
Submitted:
28 June 2023
Posted:
29 June 2023
You are already at the latest version
Abstract
Soil salinity is a limiting factor in crop productivity. Inoculating crops with microorganisms adapted to salt stress is an alternative to increasing plant salinity tolerance. Few studies have simultaneously propagated arbuscular mycorrhizal fungi (AMF) and dark septate fungi (DSF) using different sources of native inoculum from halophyte plants, and evaluated their effectiveness. This study selected, in alfalfa plants as trap culture, the infectivity of 38 microbial consortia native from rhizospheric soil (19) or roots (19) coming from six halophyte plants, as well as their effectiveness in mitigating salinity stress. Inoculation with soil resulted in 26%-56% colonization by AMF and 12%-32% by DSF. Root inoculation produced 10%-56% and 8%-24% of colonization, respectively. There was no difference in the number of spores of AMF produced with both inoculum types. The effective consortia were selected based on low Na but high P and K shoot concentrations which is relevant for plant nutrition but more for salt stress mitigation. The microbial consortia selection is presented in a more novel and applicable context, which would allow for the production of native microbial inoculants adapted to salinity to diminish the harmful effects of salinity stress in glycophyte plants in the context of sustainable agriculture.
Keywords:
Microbial inoculant
; inoculum infectivity
; inoculum effectivity
; arbuscular mycorrhizal fungi
; dark septate fungi
; inoculum adapted to salinity
1. Introduction
Soil salinization reduces cultivable land areas and has negative effects on agricultural productivity, biodiversity, and water quality, particularly in arid and semi-arid areas. Worldwide, 932.2 million hectares have salinization problems, and most crops of economic interest are not tolerant to salinity, which decreases yields by 20% to 50% (Shrivasata and Kumar, 2015). Halophyte plants that naturally grow in saline environments contain many microorganisms in their roots that could be useful for inoculating salinity non-tolerant glycophyte plants. These microorganisms can promote plant growth, influence plant physiology and metabolism, and increase plant tolerance to salt stress.
Arbuscular mycorrhizal fungi (AMF) and dark septate fungi (DSF) are an important part of the soil microbial community. AMF establish mutualistic symbiosis with the roots of 80% of terrestrial plants (Smith and Read, 2008). These fungi are considered obligate biotrophs because they cannot complete their reproductive cycle without an alive host plant. AMF trigger several biochemical and molecular mechanisms in the plant to mitigate salt stress (Evelin et al., 2009; Ruiz-Lozano et al., 2012), including nutrient absorption, hormonal signaling, growth, and the production of regulatory osmolytes. It has been suggested that AMF act as the first selective barrier for ions that can be toxic in saline soils at high concentrations (Na and Cl). Ion discrimination could take place during fungal nutrient absorption from the soil or during transfer to the host plant (Hammer et al., 2011).
Conversely, DSF are endophytic microorganisms that colonize plant roots and are characterized by the presence of microsclerotia, a brown to black mycelium with septate and melanized hyphae in the intercellular and intracellular spaces of root tissues (Piercey et al., 2004; Knapp et al., 2015; Yuan et al., 2016). These fungi are highly abundant in extreme environments such as saline soils (Yuan et al., 2021). Moreover, DSF help the host plant to tolerate salt stress, promote plant growth, and facilitate nutrient absorption (Vergara et al., 2019). Therefore, AMF and DSF are essential components of agricultural inocula, particularly for high-salinity soils (Evelin et al., 2009).
Large-scale production of AMF and DSF inocula is necessary for applying these fungi in agriculture. Traditionally, the most commonly used techniques to isolate and produce large-scale AMF-based inoculum are aeroponics, hydroponics, inert substrate, and in vitro culture (Trejo-Aguilar and Banuelos, 2020); these methods focus on the propagation of mainly one or two species. However, inoculating plants with AMF consortia is more effective than inoculating just one or two species. Regarding DSF production, in vitro propagation and subsequent crop inoculation is the only technique so far that allows using these fungi as crop inoculants.
Selecting native microorganisms highly adapted to the local conditions, such as the salinity, is essential for the production of microbial inoculants; this has been shown for AMF inoculants (Douds et al., 2005; Bencherif et al., 2023). However, including DSF in crop inoculants is still an unexplored alternative. The present research selected native microorganisms for inoculant isolation and propagation, considering both AMF and DSF native consortia from halophyte plants using two types of crude inoculum (rhizospheric soil and roots). This procedure would offer more options for microorganisms adapted to saline conditions to assist their host plants and mitigate the damaging effects of salinity more efficiently.
In general, the selection of AMF-based inoculant production protocols focuses on infectivity criteria, considering direct indicators such as species composition, spore density, and mycorrhizal colonization. Indirect indicators such as the most probable number of propagules and soil microbial biomass can also be considered to assess the quality of a mycorrhizal inoculum (Bencherif et al., 2023). To our knowledge, no studies have considered the physiological responses to salinity mitigation in host plants that function as fungal trap culture, and posteriorly use the endorhizal fungi in salinity non-tolerant crops. These responses are related to inoculant effectiveness and may be an additional selection criterion for producing native salinity-adapted inoculants that can colonize roots and efficiently mitigate the deleterious effects of salt stress. Therefore, selecting microbial inoculants requires knowledge of the intrinsic benefits of the fungi in the context of the stress conditions under which they will be used. In addition, there is limited knowledge about the influence of the type of inoculum, i.e., obtained from the soil or the root, on the production of microbial inoculants. Most studies have isolated the inocula from the soil, which can carry small fragments of colonized roots, spores, and hyphae. Using the roots as the sole source of propagules could also provide potential inoculants for saline soils. Furthermore, DSF fungal groups have barely been included in the microbial inoculants.
The present study had the following objectives: 1) to simultaneously isolate AMF and DSF consortia from halophyte plants, 2) to compare the infectivity and effectiveness of these fungal consortia from two inoculum sources (root and soil), and 3) to analyze the mechanisms of salinity tolerance in AMF that mitigate salt toxicity in plants. The present research proposes an alternative for selecting native microbial inoculants adapted to high salinity conditions, which can be useful to mitigate the damage caused by salinity stress in sensitive plants.
2. Materials and Methods
2.1. Root and rhizospheric soil sampling of halophyte plants
The sampling site was located at the Colegio de Postgraduados in the east of the State of Mexico with geographic coordinates 19.27°N, 98.54°W, and an altitude of approximately 2,236 masl (Figure S1). This area is located in the former Lake of Texcoco and is characterized by natural salinity caused by salt deposits that rose to the surface (Valenzuela-Encina et al., 2008). Systematic sampling was conducted in February 2021, and 38 samples of rhizospheric soil and fine roots of halophyte plants dominant in the area were collected (Distichlis spicata, Cynodon dactylon, Eragrostis obtusiflora, Suaeda torreyana, Kochia scoparia y Baccharis salicifolia).
2.2. Physical and chemical analysis of rhizospheric soil
The following chemical analyses were performed on the rhizospheric samples collected: pH, electrical conductivity (EC), sodium adsorption ratio (SAR), soluble phosphorus (P), and soluble cations (Ca, Mg, K, Na). These variables were measured in the saturation paste extract. The EC was measured by conductometry, and the concentration of cations in the saturation extracts and available soil P was measured according to Mexican regulation NOM-021-RECNAT-2000. The samples were dried in the shade and sieved on a stainless steel mesh of 2 mm diameter. The saturation aqueous extract was obtained from the saturated paste by vacuum filtration. Na and K were measured by flamometry. Ca and Mg were determined by atomic absorption spectroscopy (Perkin Elmer 31000). Available phosphorus was extracted with sodium bicarbonate (0.5 M) in a 1:20 soil:sodium bicarbonate solution ratio. P was determined by UV spectrophotometry at 882 nm (Olsen, 1954).
2.3. Propagation of AMF and DSF consortia
The propagation of AMF consists of establishing trap cultures to obtain spores, fragments of colonized roots, external mycelium, and sporocarps in a short time, which are sources of inoculum. Propagation of DSFs is generally performed in in vitro cultures to obtain spores. In the present experiment, endomycorrhizal fungi (AMF and DSF) from saline soils were simultaneously propagated on alfalfa plants. Half of the trap cultures used as inoculum source soil from the rhizosphere of halophyte plants, while the other half used fine roots of these same plants.
For endorhizal fungi propagation, 19 composite samples were prepared by mixing soils with similar EC from the same halophyte plant species. Table 1 summarizes the composite samples obtained and used to inoculate alfalfa seeds (19 samples). Similarly, 19 composite samples were prepared from the root of halophyte plants that come from the same site as the soil samples. These root samples were used to inoculate alfalfa seeds.
Soil samples with different EC were used as individual soil samples for trap cultures. One hundred grams of the individual samples were inoculated on alfalfa seeds. For composite samples, 50 g of each individual sample (with similar EC and from the same halophyte plant) were mixed manually to obtain homogeneous samples. One hundred grams of each composite sample was used as inoculum to establish the trap culture.
The procedure for the trap cultures with root inoculum was similar to that for the soil. In individual samples, 2 g of roots from each halophyte plant were weighed. For composite samples, 2 g of the roots of each plant of the same species, obtained from the soil with similar EC, were mixed and homogenized. In these trap cultures, 2 g of roots in both cases (individual or composite samples) were used to inoculate alfalfa seeds.
2.4. Establishment of fungal trap cultures
From the 19 rhizospheric soil samples and 19 root samples, 38 trap cultures were prepared with three replicates (114 experimental units). Each composite soil sample or composite root sample was handled as a fungal consortium, and the same sample number was retained. Each experimental unit consisted of a 2 kg pot. For trap cultures with rhizospheric soil as inoculum, 1.6 kg of sterile silica sand was first placed at the bottom of the pot, followed by a layer of rhizospheric soil (100 g) and sterile sand, which filled the pot completely. For trap cultures with roots, 2 kg pots with 1.6 kg of sterile sand were also used, and a 2 g layer of roots was placed and covered with sterile sand. Control trap cultures were prepared in the same way but without the inoculum. Alfalfa (Medicago sativa var. Moapa) was used as a host plant (trap). Seeds were superficially disinfected with 70% (v/v) alcohol and rinsed thrice with sterile and distilled water. Excess water was removed with sterile filter paper, and 1 g of seed per pot was sown.
The trap cultures were maintained under greenhouse conditions for 11 months (average temperature 22 °C) at the Colegio de Postgraduados, Campus Montecillo, State of Mexico. The plants were irrigated every third day with a modified Hoagland nutrient solution low in phosphorus (20 µM KH2PO4), according to Millner and Kitt (1992). In the greenhouse, the pots were rotated every week in a clockwise direction.
2.5. Evaluation and selection of endorhizal fungi consortia
Plants were harvested 11 months after planting and inoculation. Endorhizal consortia (AMF and DSF) from rhizospheric soil and roots were selected based on their infectivity and effectiveness on alfalfa plants. Infectivity was based on root colonization (INVAM, 2011) and the number of spores on the substrate; both variables are normally used for AMF. Root colonization by DSF was considered an additional variable, so dual colonization results are presented. To select effective consortia, it considered the following criteria: high fresh weight production, good plant nutrition (considering foliar N and P concentrations), and adequate measures of several physiological variables related to salt stress mitigation, i.e., limited Na absorption (since this cation is toxic) and high absorption of osmoregulatory cations (K, Ca, and Mg).
2.5.1. Fungal infectivity
Alfalfa roots from the trap cultures were washed and cut into 1 cm segments. Segments were clarified with 3% KOH for 12 days until the removal of root pigmentation. Root segments were immersed in 3% HCl for 15 min and stained with 0.5% trypan blue in 50% glycerin for three days, according to Phillips and Hayman (1970). The percentage of total colonization by AMF and DSF was determined by the method described by Koske and Gemma (1989). Permanent preparations of root segments were observed under an optical microscope (40x objective). The frequency of AMF structures was quantified by considering the presence or absence of arbuscules, vesicles, or hyphae. DSF-colonized structures were established as those with microsclerotia and dark septate hyphae in the same root segments.
The frequency of AMF and DSF (%) was calculated with the following formula:
Spores from each fungal trap culture were extracted by the wet sieving technique (Gerdemann and Nicolson, 1963). Spores were observed under a stereoscopic microscope. The number of spores was reported per 20 g of dry substrate.
2.5.2. Taxonomical morphotypes identification of spores of AMF
From the selected infective and efficient fungal consortium, the analysis of morphotypes of AMF was also performed to know their taxonomical name. This was made in the original and propagated consortium samples.
2.5.3. Fungal effectiveness
Alfalfa plants were harvested, and the aerial part was separated from the root. Fungal effectiveness was assessed on the aerial part only since roots can be used as a source of inoculum for other plants after evaluating their infectivity (as mentioned previously). The aerial plant material from each pot was weighed and placed separately in paper bags for drying at 60 °C for 72 h. The material was grounded and digested with a mixture of HNO3:HClO4 (3:1) in an open system. The sample was diluted to 50 mL with water, and the following macronutrients were analyzed: N, P, K, Ca, Mg, and Na. The concentration of Ca and Mg was determined with an atomic absorption spectrometer (David, 1959), while Na and K were quantified by flamometry (Havre, 1961). P concentration was obtained by the ammonium molybdate blue method (Kitson and Mellon, 1944). The foliar percentage of N was analyzed by the Kjeldahl method (Kendall, 1998).
2.5.4. Elemental analysis in AMF spores and hyphae
To determine the possible involvement of AMF fungal structures in accumulating Na as a strategy to decrease its transport to the host plant, samples of spores and hyphae from AMF were analyzed by environmental scanning electron microscopy (ESEM) coupled to EDX. A backscattered electron detector and an X-ray detector (Bruker, Quantax 200, Germany). Elemental analysis by EDX was performed on these fungal structures with an accelerating voltage of 30 kV, a spot size of 700 (arbitrary units), and a counting rate of 1000-9000 counts per second (cps). Spores from Consortia 2, 8, and 11, pertaining to extremely saline soil, moderately saline soil, and saline soil, respectively, were chosen for this analysis.
2.6. Surface disinfection of roots, DNA extraction, and PCR amplification
It was conducted a molecular study of the original consortium, which showed the best characteristics of infectivity and effectiveness. The roots of Distichlis spicata were separated from the adhered soil, washed with 2% NaCl, and dried with sterile filter paper. Roots were processed with 70% ethanol to disinfect the surface (2 min), washed with sterile 2% NaCl (three times), disinfected with 15% H2O2 (5 min), and, finally, washed with sterile 2% NaCl (three times). The solutions obtained after the final washing of each sample were evaluated to ensure surface disinfection efficiency using agar plates. Only successfully disinfected root material was used for further analysis. DNA isolation from 20 mg of disinfected root material was performed according to the protocol (plant and fungal DNA purification kit, EURx). Isolated DNA was placed in 1.5 mL Eppendorf tubes and shipped in cold packs (Blue ice) to NOVOGEN (California, USA) for amplicon analysis (Illumina Platform-MiSeq). The region of V3-V4 16S rRNA was amplified using primers 341_F(5′- CCTAYGGGRBGCASCAG-3′) and 806_R (5′-GGACTACNNGGGTATCTAAT - 3′) and the region ITS1-5F was amplified using primers ITS5-1737_F(5′-GGAAGTAAAAGTGCTAACAAGG-3′) and ITS-2043_R (5′-GCTGCGTTCTTCATCGATGC-3′). Operational taxonomic unit (OTU) was generated using demultiplexed sequences at 97% similarity, and singletons were removed.
2.7. Experimental design and statistical analysis
A completely randomized design with three replicates was used to evaluate the effectiveness and infectivity of fungal consortia on alfalfa plants. The number of composite mixtures represented the established fungal consortia (19), and a control treatment (uninoculated) was included. For all variables, the Shapiro-Wilk normality test was performed, and the homogeneity of variances was corroborated by Bartlett's test (α=0.05). All the variables presented normal distribution, so data transformation was not necessary. The data were analyzed with descriptive statistics, ANOVA (p ≤ 0.05), and Tukey's test for comparison of means (p≤ 0.05). All statistical analyses were performed using R statistical software. Multivariate analysis (PCA=principal component analysis) was performed with the R package Factoextra (Kassambara and Mundt, 2017).
3. Results and discussion
3.1. Physical and chemical analysis of the rhizospheric soil of each composite sample
The EC, SAR, and soluble cations analyzed in the rhizospheric soil of the study area showed high variability. The EC ranged from 0.9 to 43 dS m-1 (Table 1). Several authors have observed significant EC variability in the same geographical area; Valenzuela-Encinas et al. (2008) reported ranges from 2.3 to 200 dS m-1, Castro-Silva et al. (2013) from 71 to 159 dS m-1 and Beltrán-Hernández et al. (2007) from 22 to 150 dS m-1. The SAR ranged from 14 to 580 mmol L-1. The variability of this parameter coincides with soil samples from the former lake of Texcoco reported by Beltrán Hernández et al. (2007) and Santoyo de la Cruz et al. (2021), who reported SAR values between 103 and 1718 mmol L-1 and between 11 and 43 mmol L-1, respectively. The order of concentration of soluble cations in the soil was Na>K>Ca>Mg (Table 1). Na concentrations ranged from 12 to 656 mM, and all samples exceeded the Na concentrations considered normal for soils. However, the K and Ca concentrations were normal, and Mg concentrations were low (Table 1). The pH ranged from 7.8 to 9.6 and was classified as alkaline (Jones and Wolf, 1984). Valenzuela-Encinas et al. (2009) reported pH ranges between 7.8 and 10.1 in soils from the former lake of Texcoco. The available P concentration was between 6.41 and 17.38 mg kg-1 (Table 1). The average normal available P concentration in non-saline soils is 25 mg kg-1 soil, so all composite samples were deficient in P. Xie et al. (2022) reported the available P concentration of 12 mg kg-1 in saline soils, whereas Mahmood et al. (2013) reported 2.97 mg kg-1 and 2.76 mg kg-1 of P in two saline soils with EC of 6.59 dS m-1 and 4.21 dS m-1, respectively.
3.2. Infectivity of fungal consortia
3.2.1. Arbuscular mycorrhizal fungi
Mycorrhizal colonization
All roots of alfalfa plants inoculated with soil or root inocula showed characteristic colonization by AMF and DSF structures. AMF formed coiled hyphae, vesicles, and arbuscules on the roots. DSF structures were dark-colored septate hyphae and microsclerotia growing intracellularly in the root cortex (Figure 1). These structures are characteristic of the genera Ammopiptanthus (Hou et al., 2020) and Paraphoma (Ban et al., 2012).
When soil was used as the inoculum, the percentage of AMF colonization was between 26% and 56%. Consortia 2, 3, 4, and 6, propagated from the soil, promoted the highest root colonization (Figure 2a). This is particularly interesting because colonization above 50% is considered a desirable infectivity parameter for AMF inocula (INVAM, 2011) and because these soils had high Na and EC concentrations (Table 1). In contrast, the lowest percentage of AMF colonization was detected in plants inoculated with Consortia 13, 16, and 17, pertaining to soils with low EC and Na concentrations. There is controversy concerning the effect of salinity on mycorrhizal colonization; some studies showed that soil salinity reduces colonization (McMillen et al. 1998; Juniper 1996), but other studies did not observe this reduction (Levy et al. 1983; Hartmond et al. 1987). For example, Juniper and Abbott (2006) irrigated AMF trap cultures with 0, 150, and 300 mM NaCl and showed that Na decreased the percentage of colonization. Campanelli et al. (2013) observed that with 0 mM NaCl and 150 mM NaCl, the percentage of colonization was 69% and 47%, respectively. Laounae et al. (2019) reported that the percentage of colonization of a salinity-susceptible alfalfa variety by AMF in 0 mM NaCl and 120 mM NaCl was 31.5% and 43%, respectively; while a salinity-tolerant alfalfa variety, showed 86% and 66% of AMF colonization, respectively (Laounae et al., 2019).
In the present research, the highest percentages of colonization in alfalfa are similar to the results described under salinity conditions (Figure 2a). Recently, Bencherif et al. (2023) observed the highest colonization (86%) in soil with medium salinity (8.5 dS m-1) for a 24-month alfalfa trap culture with quarterly pruning of the aerial part of the plants. In the present study, colonization was lower (54%) in plants inoculated with soil Consortia 2, 3, 4, and 6. Soil Consortium 2 had five times higher EC, while Consortia 3, 4, and 6 had two times higher EC compared to what was observed by Bencherif et al. (2023).
Regarding the plants inoculated with the root-derived consortia as inoculum, the total percentage of colonization was between 10% and 56%. Plants inoculated with Consortia 3, 7, and 12 showed the highest percentage of colonization, while all other plants had less than 30% (Figure 2a). The average percentage of colonization differed between plants inoculated with soil and plants inoculated with roots (43% and 23%, respectively). However, Consortium 7 showed a similar percentage of colonization in alfalfa when using soil or root as inocula. Consortium 12 had a higher percentage of colonization with root inoculum than with soil inoculum. In all other plants, the percentage of colonization was higher when using soil as inoculum than with root (Figure 2a). The literature shows a limited evaluation of AMF infectivity considering different inocula sources (soil or root). Abbott et al. (1994) mentioned that root fragments have propagules (spores, hyphae, vesicles) that can colonize plant roots, but this depends on the fungal species present in the roots. For example, roots colonized with Glomus and Acaulospora may be infective, but not so when colonized by Scutellospora and Gigaspora (Abbott et al. 1994; Biermann and Linderman, 1983).
Plants inoculated with root consortia 3, 7, and 12, which had the highest percentage of colonization, consisted of Glomus sp. and Diversispora sp. (Consortium 3), Rhizophagus sp. and R. aggregatus (Consortium 7) and Septoglomus sp. (Consortium 12). These AMF genera native to saline soils were infective propagules on alfalfa. The results of the present investigation show that both inoculum sources can generate infective consortia. The results also showed that seven consortia promoted colonization greater than 50% in alfalfa roots under high salinity conditions, four consortia from the soil, and three consortia from roots as a source of inoculum. Therefore, the infectivity of the consortia supports their use as prospects for inocula. INVAM (2011) showed that commercial inocula (from non-saline sites) have root colonization between 19% and 54%, which is comparable to that obtained under salinity conditions in the present investigation.
Number of spores
The number of spores produced by the consortia using the soil inoculum ranged from 48 to 95 in 20 g of soil (Figure 3). Consortium 1 produced the highest number of spores among all consortia, despite having the highest EC value (42.5 dS m-1) and Na concentration in soil (655.9 mM). The number of spores was lower than that obtained by other authors on other plant species with lower soil EC. Using soils with natural salinity (not anthropogenic), Aliasgharzadeh et al. (2001) reported 160 spores in 20 g of soil in alfalfa plants with EC of 12.2 dS m-1, 288 spores in onion with 7.3 dS m-1, 239 spores in wheat with 12.1 dS m-1, and 230 spores in barley with 21.1 dS m-1. In other studies, the number of spores has been lower (Barrow et al., 1997), and no spores have been found in the rhizospheric soil of halophyte plants with EC higher than 45 dS m-1 (Hirrel 1981; Kim and Weber 1985). Under in vitro conditions, salinity has also been reported to decrease spore numbers. For example, Jahromi et al. (2008) found 59, 14, and 11 spores per cm-2 of Gl. intraradices (now R. irregularis) under NaCl concentrations of 0, 50, and 100 mM, respectively. The number of spores depends on the ability of AMF species to sporulate under the conditions that fungi and plants are established. Under trap culture conditions, some species sporulate readily, while others sporulate infrequently or not at all (Hayman 1970; Daft and Nicolson 1972). The number of propagules in an AMF inoculum is also related to its infectivity. However, there is no concrete data to establish the infectivity of inocula from halophyte plants based on the number of spores. Bencherif et al. (2023) observed a maximum of 650 spores in 10 g of soil in alfalfa after 24 months of trap culture with aerial part cutting and reseeding every four months. The trap culture lasted 11 months in the present investigation, and the alfalfa plants were not cut.
In plants inoculated with the root consortia, the range of spore number was between 28 and 174 per 20 g of substrate. The highest number of spores was obtained in plants with Consortia 3 and 7 (Figure 3). To the best of our knowledge, no research had previously used the roots of halophyte plants for inoculum and spore production; this study is the first to show spore production from halophyte plant roots in a trap culture.
Spore production was dependent on the composite sample and inoculum type. Spore production in Consortia 3, 7, and 12 was higher when the root was used as inoculum compared to the soil. In contrast, Consortia 1, 2, 4, 5, 6, 8, and 19 had higher spore numbers when using soil as inoculum compared to the root. However, in Consortia 10, 11, 13, 14, 15, 16, 17, and 18, the number of spores was similar with both inoculum sources.
3.2.2. Dark septate fungi
The percentage of DSF colonization from soil consortia was between 12% and 32% (Figure 2b) in the alfalfa roots. Alfalfa roots from Consortia 3, 7, 14, and 16 had the highest percentage of colonization, which differed from the other consortia. In plants inoculated with root consortia, the percentage of colonization ranged from 8% to 24%. The highest percentage of colonization was observed in Consortia 6, 7, 9, 12, and 13, which was different from the rest of the plants (Figure 2). Yuan et al. (2021) mentioned that DSF are abundant in saline environments. Our results show, for the first time, that DSF native to saline soils can colonize the roots of alfalfa plants along with AMF when using natural inocula (soil and root). AMF showed a higher percentage of colonization than DSF (Figure 2). Furthermore, microsclerotia and dark hyphae, both native to saline soils, can be infective propagules of DSF (Currah et al., 1993; Jumpponen and Trape, 1998) and colonize host plants. Colonization by DSF on alfalfa roots was higher with the soil inoculum than with the root inoculum.
3.2.2. Dark septate fungi
The percentage of DSF colonization from soil consortia was between 12% and 32% (Figure 2b) in the alfalfa roots. Alfalfa roots from Consortia 3, 7, 14, and 16 had the highest percentage of colonization, which differed from the other consortia. In plants inoculated with root consortia, the percentage of colonization ranged from 8% to 24%. The highest percentage of colonization was observed in Consortia 6, 7, 9, 12, and 13, which was different from the rest of the plants (Figure 2). Yuan et al. (2021) mentioned that DSF are abundant in saline environments. Our results show, for the first time, that DSF native to saline soils can colonize the roots of alfalfa plants along with AMF when using natural inocula (soil and root). AMF showed a higher percentage of colonization than DSF (Figure 2). Furthermore, microsclerotia and dark hyphae, both native to saline soils, can be infective propagules of DSF (Currah et al., 1993; Jumpponen and Trape, 1998) and colonize host plants. Colonization by DSF on alfalfa roots was higher with the soil inoculum than with the root inoculum.
Previous studies in vitro isolated DSF from saline environments and inoculated them on plants to evaluate their infectivity (González Mateu et al., 2020; Farias et al., 2020). For example, González Mateu et al. (2020) inoculated a native DSF consortium from saline soil into native and invasive lineages of the plant Phragmites australis and observed 61% colonization in the native lineage and 57% in the invasive lineage. Farias et al. (2020) separately inoculated the DSF Sordariomycetes sp. and Melanconiales elegans in cowpeas, both isolated from the halophyte plant Vochysia divergens. These authors observed that plants inoculated with M. elegans had a maximum colonization rate of 74%, while the colonization of Sordariomycetes sp. was 70%. The percentages of colonization by DSF discussed previously are higher than those of the present investigation, both in soil inoculum and root inoculum; this difference may be due to the low initial infectivity of the fungal propagules, to the fact that some DSF are host-specific, as mentioned by Hawksworth and Rossman (1997), or to multiple factors, such as climate, soil type, and salinity concentration, that may influence colonization.
The average percentage of AMF colonization in soil consortia was 45% and 22% for DSF; in root consortia, it was 23% for AMF colonization and 13% for DSF colonization. Fuchs and Hasewandter (2004) indicated that AMF colonization does not always predominate over DSF colonization and that the colonization pattern is host-plant dependent. Cofré et al. (2012) evaluated the percentage of AMF and DSF colonization in roots of Atriplex cordobensis in soils with different EC (2.0, 4.4, and 19.7 dS m-1). The authors observed similar colonization by AMF and DSF in the two soils with the lowest EC values, and colonization by DSF was higher than AMF in the soil with high EC. Sonjak et al. (2009) observed higher colonization by AMF than DSF in 12 halophyte plants growing naturally in saline soils. Therefore, multiple factors may influence the percentage of colonization of both types of fungi.
The present research also evaluated other variables that could be important for selecting inocula for consortia propagation that mitigate the negative effects of soil salinity. This is especially important because the efficiency of fungal isolates is not always related to the degree of mycorrhizal colonization (Corkidi et al., 2004) or the number of spores, which are the parameters commonly used to select and identify the quality of AMF inoculants. However, since DSF can be isolated and propagated in culture media, the quality of DSF inoculants would be based on the number of spores, similar to other fungal products on the market. Further research is being conducted to molecularly identify native AMF and DSF species that colonize the roots and soil of halophyte plants and those that were propagated in alfalfa trap cultures.
3.3. Mitigation of salt stress in alfalfa plants
Importantly, the effectiveness of fungal consortia on the response of alfalfa plants includes the participation of both types of fungi (AMF and DSF). Since it is impossible to separate the individual effects, the discussion will be inferred from the current knowledge of each type. In general, the results show that the response to inoculation depends on the type of inoculum (root or soil), consortium, and physical or chemical characteristics of the original soil.
3.3.1. Effectiveness of fungal consortia propagated on alfalfa
Fresh weight
Plants inoculated with Consortia 3, 7, 14, 17, and 18, pertaining to the soil, produced the highest fresh weight of the aerial part in alfalfa plants (Figure 4). The response with Consortia 3 and 7 is especially interesting because the original soils presented high EC. The increase in fresh weight was 20% for Consortium 3 and 41% for Consortium 7, compared to control (non-inoculated) plants. The dry weight of most of the other consortia was similar to that of the control plants despite the high Na concentration and high EC value of the soil in some composite samples containing these consortia.
Regarding the use of root as inoculum, plants inoculated with Consortia 15 and 19 had the highest fresh weight in the aerial part of the alfalfa plants (Figure 4). The fresh weight was different in the other inoculated plants, including the control. Alfalfa plants showed higher fresh weight when soil was used as inoculum compared to the root, except in plants with Consortia 1 and 19 (Figure 4). Previous studies showed that Na concentrations higher than 60 mM negatively affect fresh weight in alfalfa (Anand et al., 2000; Maggio et al., 2009; Bertrand et al., 2021). In the present study, Na concentration was higher in the soil of Consortia 2, 3, 7, and 17, but still produced higher fresh weight than the control plants, demonstrating their effectiveness.
Foliar concentration of N and P
The N content in the foliage of plants inoculated with soil ranged from 1.2% to 4.6%. Plants inoculated with Consortium 11 had the highest percentage of N. This result is relevant because the N content is considered normal despite the high Na concentration (181 mM) in the soil mixture that generated this consortium (Table 1). In contrast, the N content in the control plants was 1.08% (Figure 5a). According to Mattson (1980), foliar N content between 3% and 7% is normal, but values between 0.5% and 1.5% are low. Consortia 1, 4, and 6, from soil with high Na concentration, also influenced normal foliar N percentages. Some research has shown that soil Na concentration higher than 50 mM negatively affects N absorption (Evelin et al., 2012; Campanelli et al., 2013; Porcel et al., 2016). Consortia 1, 4, 6, and 11 mitigated this negative effect on N absorption in alfalfa plants. When root was used as inoculum, the foliar N content had a narrower range (1.17% - 3.74%) than when soil was used as inoculum. Again, Consortia 4 and 6, as well as 16, influenced the highest foliar N percentage (3.68%, 3.08%, and 3.74%, respectively). Therefore, these consortia are efficient in N absorption in alfalfa plants.
Soil salinity is known to decrease P availability (Grattan and Grieve 1998) and inhibit its absorption by plants (Martinez et al. 1996; Martinez and Läuchli 1994). However, AMF consortia isolated from saline soils can increase foliar P concentration in sorghum when used as a trap culture (Chandra et al., 2022). DSFs offer similar benefits to those known in AMF.
For example, they enhance P absorption in their host plants (Vergara et al., 2019) and participate in mitigating the effects of salt stress (Castro-Farias et al., 2020; González-Mateu et al., 2020). In agreement with this, the present research most plants inoculated with consortia from both soil and root increased the foliar concentration of P. The foliar concentration of P was higher in plants inoculated with Consortia 1, 3, and 10 (from soil). Also, P concentrations differed from the rest of the inoculated plants and the control (Figure 5b). This result is relevant because the soils that generated Consortia 1 and 3 had high Na concentrations but increased P absorption in alfalfa plants. Plants inoculated with
Consortia 12 and 14 (soil inoculum) showed foliar P concentration similar to control plants. These foliar P concentrations are considered lower than the normal range (0.5 - 5 g P kg-1 PS), according to Malhotra et al., (2018). The highest P concentration was observed in Consortia 8, 10, 13, and 14 when roots were used as the inoculum source. Interestingly, Consortium 10 of both inoculum sources promoted the highest foliar P concentration.
AMF and DSF participated in the nutritional status of alfalfa plants inoculated with consortia from two inoculum types: soil and root. It can be hypothesized that each fungal type participates in the alfalfa mineral nutrition through different mechanisms. Della Monica et al. (2015) observed a close relationship between AMF and DSF with P availability and absorption in plants. While DSF increase the P reserve in the rhizosphere, AMF transfer P to the host plant. Co-colonization of plants by AMF and DSF shows a synergistic effect. Future research should address this hypothesis to understand plant nutrition in saline soils, considering the dual participation of these beneficial endophytic fungi.
Foliar concentration of Na and protective osmolytes
The foliar Na concentration of plants inoculated with soil consortia ranged from 168 to 453 mmol kg-1 PS (Figure 6a). In control alfalfa plants without soil addition (and therefore, without an external Na source), the foliar concentration was less than 50 mmol kg-1.
The normal Na concentration in plants is less than 87 mmol kg-1. Some research has shown that inoculation with AMF decreases Na absorption in host plants. However, these findings are difficult to interpret because such investigations were conducted in greenhouse conditions using non-saline soil (substrate), and salt stress was induced with a Na source. For example, under greenhouse conditions, Campanelli et al. (2013) observed lower leaf Na concentration (1,447 mmol kg-1 PS) in alfalfa plants inoculated with Gl. viscosum than in control plants (1,600 mmol kg-1 PS); these plants had vermiculite and peat as substrate and were irrigated with a 150 mM NaCl saline solution. Conversely, Ben-Laouane et al. (2020) sowed alfalfa seeds in soil with low EC (0.75 dS m-1) and irrigated the plants with 120 mM NaCl saline solution; these authors reported lower leaf Na concentration (870 mmol kg-1 PS) in plants with the AMF consortium (Glomus sp., Sclerocystis sp., and Acaulospora sp.) compared to control plants (1,520 mmol kg-1 PS). Both studies report higher concentrations than those observed in the present investigation in saline soils with native microorganisms from halophilic plants.
Plants inoculated with Consortia 1, 4, 9, 12, and 15 (Figure 6a) had the lowest leaf Na concentration. Interestingly, soils from Consortia 1 and 4 had high Na concentrations (655.89 mM and 472.45 mM) and high EC (42.5 dS m-1 and 17.50 dS m-1), equivalent to three times more Na on average than the experiments conducted by Campanelli et al. (2013) and Ben-Laouane et al. (2020). However, these authors quantified eight times higher foliar Na concentration than the present study. The results strengthen the idea that salinity-adapted consortia can control Na absorption in inoculated plants, as observed by other authors (Latef et al., 2011; Gaber et al., 2020). Ion discrimination could occur during the fungal absorption of nutrients from the soil or transfer to the host plant (Hammer et al., 2011). Estrada et al. (2013) observed different behavior between AMF from saline soils and from non-saline soils; in this work, corn plants were irrigated with 0.1 M NaCl previously inoculated with R. irregularis from non-saline soil or Claroideoglomus etunicatum from saline soil. These authors reported that plants inoculated with R. irregularis showed a foliar Na concentration of 521 mmol kg-1 PS; plants inoculated with Claroideoglomus etunicatum (adapted to salinity) had 260 mmol kg-1 PS of foliar Na; and non-inoculated plants had 869 mmol kg-1 PS of foliar Na. AMF decreased Na absorption in both inoculated plants, but absorption was more efficient in the saline soil isolate. These results enable the selection of native halophilic consortia that significantly affect salinity mitigation, as measured from the reduced foliar absorption of Na. Regarding inocula obtained from the roots, inoculated plants had similar leaf Na concentrations to non-inoculated plants, and all plants showed normal Na concentrations.
Stress tolerance is a prerequisite for a successful symbiotic relationship between DSF and host plants that decreases salt stress. Therefore, fungi adapted to saline conditions will confer tolerance compared to non-adapted fungi (Gaber et al., 2020). The DSF Curvularia sp. isolated from the halophyte plant Suaeda salsa established a beneficial symbiosis with white poplar (Populus tomentosa), increasing its response to salt stress through higher antioxidant activity (Pan et al., 2018). In the present investigation, native DSF associated with halophyte plants could confer tolerance to abiotic stress in alfalfa, but separating their effects from those of AMF is impossible. Therefore, specific studies are necessary to test the independent efficiency of these endophytic fungi.
Protective osmolytes (K, Ca, and Mg) are key in mitigating plant salt stress because they can act as osmotic adjusters. In general, Na ions compete with K ions for binding sites that are essential for several cellular functions (Ruiz-Lozano et al., 2012).
Research show that inoculation with AMF favors K absorption over Na absorption in saline or sodic soils. Zuccarini and Okurowska (2008) observed higher leaf K concentration (1,069 mmol kg-1 PS) in plants inoculated with a mycorrhizal consortium composed of Gl. mosseae (now Funneliformis mosseae), R. irregularis, and F. coronatum than in control plants (820 mmol kg-1) when irrigated with 50 mM NaCl; however, no differences were found with 250 mM NaCl irrigation. In another study, the authors also observed no differences with 50 mM NaCl irrigation in control plants and those inoculated with R. irregularis in Ocimum basilicum (Zuccarini and Okurowska, 2008). Therefore, the results may differ depending on the fungi and the Na:K ion ratio. In the present study, leaf K concentration increased significantly in alfalfa plants inoculated with the soil consortia except for 15, which had a similar concentration as non-inoculated plants (310 mmol kg-1 PS). Plants inoculated with Consortia 1, 8, 17, 18, and 19 from soil had the highest leaf K concentration (1016, 1086, 1028, 1028, 888, and 984 mmol kg-1 PS, respectively). The results show that although the soil from Consortia 1 and 8 had more Na than K, the plants absorbed more K than Na (Figure 6b).
The present data suggest that AMF and the native DSF communities in the consortia participated in the significant K increase in alfalfa plants. Yun et al. (2018) reported that Zea mays plants irrigated with 200 mM NaCl and inoculated with Piriformospora indica had higher foliar K concentration than their non-inoculated counterparts. Similar results were reported by Song et al. (2015) in Hordeum vulgare inoculated with Epichloe sp. and irrigated with 200 mM NaCl: inoculated plants had higher leaf K concentration (512 mmol kg-1 PS) than the control (385 mmol kg-1 PS). In contrast, Ghabooli (2014) observed no differences in leaf K concentration between control plants of H. vulgare and those inoculated with P. indica, both with 200 mM irrigation. Therefore, the type of fungus may have a relevant effect on the absorption of K, an osmoregulatory element.
Regarding the plants inoculated with root as inoculum, all consortia except 1 and 3 increased the foliar concentration of K. Consortia 2, 4, 13, and 14 were the most efficient (Figure 6b). Importantly, all plants with the root consortia as inoculum only received nutrient solution as an external source of K.
According to LaHaye and Epstein (1969), soil Na concentration greater than 50 mM affects Ca absorption in plants. However, AMF can favor Ca absorption in their host plants under saline conditions. For example, Evelin et al. (2012) reported differences in leaf Ca concentration between Trigonella foenum-graecum plants inoculated (115 mmol kg-1 ) and non-inoculated (95 mmol kg-1 PS) with Gl. intraradices and irrigated with 200 mM NaCl. In the present research, the original soil of all the consortia, except 16, 18, and 19, overcame the saline condition with a concentration higher than 200 mM. However, in all consortia (from both soil and root), the foliar Ca concentration in alfalfa plants was significantly higher than that of the control plants, which was 270 mmol kg-1 PS (Figure 6c). Marschner (1995) indicated that the optimum range of foliar Ca concentration in plants is between 25-1200 mmol kg-1 PS. In particular, Consortia 16 and 19 from soil were the ones that increased the foliar Ca concentration the most (998 and 1040.67 mmol kg-1 PS, respectively). However, plants inoculated with Consortia 1, 2, and 3, with very high Na concentration in the soil of origin, had 2.7 times higher Ca concentration than control plants.
Regarding the plants inoculated with the root consortia, without Na from the soil, Consortium 8 substantially increased the foliar Ca concentration (1102 mmol kg-1 PS) compared to control plants (270 mmol kg-1 PS). It is well-documented that under non-saline conditions, AMF can increase Ca absorption. For example, tomato plants inoculated with F. mosseae had foliar Ca concentration of 412 kg-1 PS, and non-inoculated plants had foliar Ca concentrations of 375 mmol kg-1 PS (Karagiannidis et al., 2002); and olive trees mycorrhized with R. irregularis and not mycorrhized had foliar Ca concentration of 45 mmol kg-1 PS and 37 mmol kg-1 PS, respectively (Tekaya et al., 2017).
It is known that DSF increase leaf Ca concentration under non-saline conditions. In this sense, cucumber plants inoculated with Penicillium sp., and P. glomerata (both reported as DSF) presented higher foliar Ca concentration (10 mmol kg-1 PS) than non-inoculated plants (7 mmol kg-1 PS) (Waqas et al., 2012). Under salinity conditions, DSF can also increase leaf Ca concentration. For example, in H. vulgare inoculated with P. indica and irrigated with 300 mM NaCl, the foliar Ca concentration was 299 mmol kg-1 PS, and in non-inoculated plants, it was 155 mmol kg-1 PS (Ghabooli, 2014). In Lolium arundinaceum plants irrigated with 250 mM NaCl, inoculated and not inoculated with DSF, the leaf concentration was 64 52 mmol kg-1 PS and 52 mmol kg-1 PS, respectively (Yin et al., 2014).
In the present research, the native consortia of the two inoculum types increased the leaf Mg concentration in alfalfa (Figure 6d), although the concentration is considered normal in inoculated and non-inoculated alfalfa plants (61 mmol kg-1 PS to 329 mmol kg-1 PS) according to Marschner et al. (1989). The range of leaf Mg concentration of alfalfa plants inoculated with the soil-derived fungal consortia was between 113 mmol kg-1 PS to 280 mmol kg-1 PS, whereas control plants had a leaf Mg concentration of 50 mmol kg-1 PS (Figure 6d). Consortia 1, 3, 4, 6, 7, 7, 8, 13, 14, 15, 18, and 19 had the highest leaf Mg concentration, although the Mg concentration in soil was lower than the concentration normally found in soil solution (800 mM). The range of leaf Mg concentration in the plants inoculated with the root consortia was between 157 mmol kg-1 PS and 368 mmol kg-1 PS, with Consortia 8 and 9 promoting the highest leaf Mg concentration compared to the control plants (Figure 6d). Research has shown that AMF can increase leaf Mg concentration under saline (Giri and Mukerji, 2004) and non-saline conditions (Zare-Maivan et al., 2017), although this is not always the case. Giri and Mukerji (2004) inoculated Sesbania aegyptiaca and S. grandiflora plants with Gl. macrocarpum isolated from saline soil (15 dS m-1 EC and 150 mM NaCl) and reported 67 mmol kg-1 PS in non-mycorrhizal plants and 134 mmol kg-1 PS in mycorrhizal plants. Evelin et al. (2012) found no differences in leaf Mg concentration between inoculated and non-inoculated plants in T. foenum-graecum plants inoculated with Gl. intraradices and irrigated with 200 mM NaCl.
DSF also improve leaf Mg concentration, but this has only been shown in non-saline conditions. For example, in tomato plants inoculated individually with DSF (A101, A103, and A105), the average leaf Mg concentration was 3 mmol kg-1 PS, while in non-inoculated plants, the concentration of Mg was 2.5 mmol kg-1 PS (Vergara et al., 2017). In contrast, rice crops showed a significant difference in leaf Mg concentration between plants inoculated with DSF (A101 and A103) and non-inoculated plants (Vergara et al., 2019). Further research should describe the individual and combined functions of AMF and DSF on nutrient transfer, moreover on the functionality of AMF and DSF under saline conditions should be integrated. Several studies with AMF have ignored the natural participation of DSF, and sometimes they even were reported as contaminants (Porras-Alfaro and Bayman, 2011).
3.4. Scanning electron microscopy (SEM) and elemental analysis in AMF fungal structures
Figure 7 shows micrographs obtained by ESEM and the elemental composition of spores and external mycelium of AMF in some trap cultures. The Na concentration in spores was low (0.03% - 0.29%), and Na was not detected in the external hyphae despite its abundance in the soil of provenance, for example, Consortia 2 and 8. Using PIXE analysis, Hammer et al. (2011) also observed low Na content in spores and hyphae from two natural saline sites with different Na concentrations (Site 1: 150 mM and Site 2: 200 mM). The result obtained in the present research is relevant because it clarifies that the lower amount of Na in alfalfa tissues is not due to the participation of these fungal structures in Na accumulation but that other mitigation mechanisms might be participating.
The EDAX analysis showed that spores had low K concentration (0.13% to 0.15%), while hyphae presented K concentration between 0.12% and 0.28%. Other investigations analyzed the elemental composition of AMF spores and hyphae in monoxenic cultures. For example, Olsson et al. (2008) cultured Gl. intraradices spores in KH2PO4 and performed PIXE analysis; these authors observed that the concentration of K decreased as the dose of this element in the culture medium increased (0.89% in spores grown with 2500 mM KH2PO4 and 0.28% with lower concentration (250 mM). Hammer et al. (2011) reported K concentration in spores and hyphae from two natural saline sites, Site 1 (300 mM of soil K) and Site 2 (400 mM of soil K). The K concentration in Site 1 spores was 0.064%, while in hyphae, it was 0.15%. The K concentration in spores from Site 2 was 0.89%.
In the present work, the spores of Consortia 2 and 8 from soil had 2.57% and 2.59% of Ca, respectively. The Ca content was 6 and 7 times higher than the Na content in Consortia 2 and 8, respectively. Spores of Consortium 11, from root inocula, was 1.30%. The external hyphae of Consortia 2 and 8 had 1.69% and 2.15% Ca, while the K content was 9 and 17 times higher than the Ca content in Consortia 2 and 8. Hammer et al. (2011) reported the Ca content in spores and hyphae from two naturally saline sites; the saline soil of Site 1 had 28 mM Ca, while that of Site 2 had 280 mM. The Ca content in the spores and hyphae of Site 1 was 0.96% and 0.87%, respectively, while the spores of Site 2 had 12% Ca. The latter results are higher than those obtained in the present investigation. In spores from Consortia 2 and 8 (from soil inoculum), 0.96% of Mg was observed, while in Consortium 11 (from root inoculum), it was 0.43%. Hammer et al. (2011) reported spores from Site 1 Mg accumulation of 0.05% and Site 2 of 0.96%.
Little is known about the osmotic mechanisms by which AMF tolerates salinity conditions, as well as how other ions, such as Ca, K, and Mg, participate in the process that mitigates Na toxicity. In plants under salt stress, Ca absorption prevents Na toxicity, improves cell wall stability, and enhances nutrient absorption (Yildirim et al. 2009). The high percentage of Ca in AMF spores and hyphae possibly acts as an osmotic equivalent in fungal tissues, demonstrating the ion-selective uptake capacity of AMF.
Fe, Cu, and Si were also detected in the spores and hyphae of the consortia. Si content in spores and hyphae of AMF consortia is especially interesting because spores of Consortia 2 and 8, pertaining to the soil, had 0.88% Si, while in spores of Consortium 11, pertaining to the root, had 0.17% Si. The external hyphae of Consortia 2 and 8 had 0.09% Si and 0.75% Si, respectively. The presence of Si in the spores and hyphae of Consortium 11 is relevant because the inoculum source, i.e., the root, produced the structures containing this element, suggesting that Si constitutes the structures of these AMF from saline soils with lacustrine origin. The study of Hammer et al. (2011) was the only one that reported Si content in spores and hyphae from two natural saline sites. Saline Site 1 had 0.71 mM Si, and Si content in Site 2 was not determined. Spores from Site 1 had 0.32% Si, and hyphae had 0.16%, while spores from Site 2 had 1%. Si addition has been proposed as an alternative to increase plant resilience to salinity because it modulates physiological and biochemical processes altered by salt stress. However, the molecular mechanisms are still being studied (Dhiman et al., 2021). Under salinity conditions, AMF enhance Si absorption in plants and induce Na precipitation in cell walls of root cells (Saqib et al. 2008), thereby reducing Na concentration in shoots (Ashraf et al. 1986). The mechanisms whereby AMF participate in Si absorption are still unclear, so further study is needed to understand their function and the involvement of DSFs in this process.
3.5. Principal component analysis (PCA)
The first three components of the PCA explained 53% of the total accumulated variation (Figure 8). PC1 accumulated 25.3% and revealed the most significant variables in the study: the foliar concentration of N, P, and K; the percentage of AMF colonization; the number of spores; and the soil concentration of P, K, and Na. PC2 explained 15.4% of the variance and was influenced by the foliar concentration of Na, Ca, and Mg and DSF colonization. PC3 accumulated 13.0% of the variation, the related variables were fresh weight, soil Ca concentration, soil Mg concentration, and EC. The analysis also showed that the response of the control plants was separated from the effect of the inoculated plants. Plants inoculated with Consortia 1 and 2 from both soil and root were grouped in Component 1. These results indicated that the microbial communities composing these consortia were closely related to K and P nutrition, which were important in PC1. Several studies have shown that AMF improve P absorption in plants (Nagy et al., 2009). The present investigation also shows that the efficiency of Consortia 1, 3, and 10 of some AMF native to saline soils promote P absorption in alfalfa plants when using two types of inoculum. Therefore, P absorption by AMF should be consistently evaluated as a measure of their effectiveness in saline soils.
PCA showed no relationship between AMF and DSF colonization. However, studies should be conducted to better understand the functioning of these endorhizal fungi on their host plants under salt-stress conditions. This will promote their beneficial use in saline soils.
3.6. Selection of fungal inoculants
Based on the previous results, Consortium 1 from soil was selected for further use as an inoculant. Soil from this consortium had the highest EC and Na concentration (Table 1). Under these conditions, which negatively affect fungal infectivity, Consortium 1 had the highest spore production (96 in 20 g of the substrate), and the percentage of AMF colonization was within the range (43%) proposed by INVAM (2011) for inoculant selection. Consortium 1 had a DSF colonization percentage of 19%.
Effectiveness is also useful for selecting AMF inoculants. Abbott et al. (1992) postulated that inoculant effectiveness is based on improving plant P absorption. However, effectiveness must also be based on the absorption of other important nutrients under particular conditions, such as high salinity. Therefore, the selection of inoculants from saline soils should be based on physiological processes (nutrient absorption) that highlight the intrinsic abilities of AMF and generate benefits for the plant. Soil Consortium 1 was selected for its effectiveness in nutrient absorption under saline conditions. Plants inoculated with this consortium had the lowest foliar Na concentration (180 mmol kg-1), despite having the highest Na concentration in the original soil (656 mM), i.e., the soil had four times more Na than the leaf tissue. These results show that the AMF present in Consortium 1 effectively decreased Na translocation to tissues. K absorption is another outstanding variable under saline conditions. Soil Consortium 1 also effectively increased foliar K concentration despite the high Na concentration in the soil. P absorption is negatively affected under saline conditions. Although the soil P concentration of Consortium 1 was below the normal soil concentration, the AMF present in Consortium 1 favored P absorption.
3.7. Identification of AMF spore morphospecies in the selected consortium
Soil Consortium 1 morphospecies before trap culture were F. mosseae (67%), C. claroideum (13%), R. irregularis (12%), and Diversispora sp. (8%). Soil Consortium 1 morphospecies after trap culture were F. mosseae (47%), C. claroideum (21%), R. irregularis (21%), and Diversispora sp. (11 %). However, only the presence of R. irregularis was confirmed by sequencing analysis. A more directed molecular analysis should be performed for the detection of the other AMF observed in this consortium. Figure 9 shows these fungal morphospecies. Wang et al. (2004) mentioned that the AMF commonly observed in saline soils are Glomus sp. Aliasgharzadeh et al. (2001) observed that the predominant AMF species in saline soils with EC of 162 dS m-1 were Gl. intraradices (now R. irregularis), Gl. versiforme and Gl. etunicatum (now C. etunicatum). Bencherif et al. (2023) observed 12 AMF species after establishing a trap culture with alfalfa by using soil as inoculum at three EC levels. Dominika sp. was the dominant mycorrhizal species in the highly saline soil with EC of 9.9 dS m-1, whereas R. irregularis was the dominant mycorrhizal species in the medium saline soil with 8.5 dS m-1 and saline soil with 4.1 dS m-1.
3.8. Endophytic microbial community of the selected consortium
When crude soil and root inocula are used to propagate AMF and DSF consortia, the endophytic bacterial community present in the inocula cannot be excluded. This research focused on the endorhizal microorganisms such as AMF and DSF. To complement our study, it was analyzed the endophytic microbial community present in the roots of Distichlis spicata; which is the dominant halophyte where the Consortium 1 originated. This was selected based on its infectivity and effectiveness. The microbial composition present in the soil of Consortium 1 was also analyzed. Analysis of 16S sequences revealed the presence of 9 phyla: Proteobacteria (37%), Actinobacteria (16%), Bacteroidota (1%), Myxococcota (6%), Bdellovibrionota (1%), Patrescibacteria (1%), Fibrobacteria (0.9%), Acidobacteria (1%), and Chloroflexi (6%) (Figure 10a). The endophytic bacterial community consists of 22 genera, and 11 were the most abundant in the phyla: Ramlibacter (2%), Steroidobacter (2%), and Acidibacter (2%). In Actinobacteria, the most abundant genera were Kibdelosporangium (7%) and Lechevalieria (4%). Zhang et al. (2019) reported Proteobacteria dominance (14%) in superficially disinfected roots of D. spicata and other halophyte plants growing in desert saline soils. Shi et al. (2015) examined the diversity of endophytic bacteria associated with two halophyte plants: Salicornia europaea and Sueada aralocaspica; they found a predominance of the phyla Proteobacteria (41% and 99%, respectively). These authors did not show the chemical data of the soil where S. europaea and S. aralocaspica roots were collected, but they mentioned that these plants are indicators of saline soils.
In the present research, the analysis of ITS sequences revealed the predominance of 2 phyla: Ascomycota (59%) and Glomeromycota (8%), as well as 33% of unidentified phyla (Figure 10b). At the genus level, the genus Fusarium (29%) and Preussia (5%) were the most abundant in the phyla Ascomycota, while Rhizophagus (4%) was abundant in Glomeromycota. Currently, all DSF described belong to the phyla Ascomycota (Berthelot et al., 2019). Sequencing results corroborated the presence of DSF in D. spicata roots. In addition, the genera Fusarium and Preussia have been classified as DSF, and some species have even been reported as plant growth promoters (Porras-Alfaro and Bayman, 2011; Mapperson et al., 2014). Although the molecular analysis identified the genus Rhizophagus, it did not detect the other three morphotypes mentioned previously. Therefore, more specific molecular analyses for AMF should be performed to identify the morphospecies observed in Consortium 1. Few reports have reported the composition of the endophytic fungal community of D. spicata. Redman et al. (2022) informed that DSF of the genera Phoma sp. and Fusarium sp. were the most dominant in roots of D. spicata in soils with EC of 0.18 dS m-1. Other studies also agree that the dominant phyla in some halophyte plants (Inula crithmoides) is Ascomycota, and the genus Fusarium (50%) is the most predominant (Macía-Vicente et al., 2012). Of note, Fusarium is a very diverse fungal group that includes pathogenic or saprotrophic species. Fusarium did not cause damage in alfalfa roots in the fungal trap culture; however, its effect should be analyzed before its wide use in other plants. These results expand the knowledge of the microbial components that may be related to the quality of the endorhizal fungal inoculum produced.
4. Conclusions
Alfalfa was useful as a trap culture for the simultaneous propagation of native AMF and DSF consortia from halophyte plants growing in saline/sodic saline soils with high EC and high Na concentrations. This is the first study addressing the selection of native microorganisms adapted to high-salinity soils, using soil and roots of halophyte plants as sources of microbial inoculum. Likewise, this study evaluated two important fungal groups that favor host plant tolerance to saline soils based on infectivity and effectiveness. In this context, the results provide further information on variables that are useful for selecting native inocula from saline soils that participate in the mitigation of salinity in plants and that may have the potential for the sustainable management of agroecosystems. Soil salinity imposes ionic toxicity on plants and a deficiency of nutrients such as N, P, K, Ca, and Mg. In sustainable agriculture, solutions to salinity problems should include the application of biological processes such as inoculation with endorhizal fungi. Some of the consortia tested could be of interest for mass propagation and application in glycophytic crops. In particular, Consortium 1 improved the absorption of P and K; and decreased the foliar concentration of Na in alfalfa; it also produced the highest number of spores. Consortium 3 had the highest percentage of colonization of both AMF and DSF and significantly increased leaf Mg concentration. Traditionally, fungal type, mycorrhizal colonization, and spore number determine the infectivity of fungal inoculants used for commercial and research purposes. However, results obtained in this research show that under salt stress conditions, the foliar concentration of K, Ca, Mg (protective osmolytes that mitigate salinity damage), N, and P (essential nutrients) can be used as indicators for the selection of inoculants that effectively increase plant tolerance under high salinity conditions. These results offer a novel alternative for producing microbial inoculants adapted to salinity that are effective for protecting plants in saline soils. The alternative provided by this study should be scaled up to increase crop yields in saline soils. Future research should analyze other salinity tolerance mechanisms by which these endorhizal fungi protect their plants from the effects of salinity, a problem predicted to increase under the current climate change scenario.
References
- Abbott, L. K., and Gazey, C. An ecological view of the formation of VA mycorrhizas. Plant Soil 1994, 159, 69–78. [CrossRef]
- Abbott, L. K., Robson, A. D., & Gazey, C. Selection of Inoculant Vesicular-arbuscular Mycorrhizal Fungi. In Methods in microbiology 1992, (Vol. 24, pp. 1-21). Academic Press. [CrossRef]
- Aliasgharzadeh, N., Rastin, S. N., Towfighi, H., & Alizadeh, A. Occurrence of arbuscular mycorrhizal fungi in saline soils of the Tabriz Plain of Iran in relation to some physical and chemical properties of soil. Mycorrhiza 2001, 11, 119-122. [CrossRef]
- Anand, A., Baig, M. J., & Mandal, P. K. Response of alfalfa genotypes to saline water irrigation. Biologia Plantarum 2000, 43(3), 455-457. [CrossRef]
- Ashraf, M., McNeilly, T., & Bradshaw, A. D. The potential for evolution of salt (NaCl) tolerance in seven grass species. New Phytologist 1986, 103(2), 299-309. [CrossRef]
- Ban, Y., Tang, M., Chen, H., Xu, Z., Zhang, H., & Yang, Y. The response of dark septate endophytes (DSE) to heavy metals in pure culture. PLoS one, 2012, 7(10), e47968. [CrossRef]
- Barrow, J. R., Havstad, K. M., & McCaslin, B. D. Fungal root endophytes in fourwing saltbush, Atriplex canescens, on arid rangelands of southwestern USA. Arid Land Research and Management 1997, 11(2), 177-185. [CrossRef]
- Beltrán-Hernández, R. I., Luna-Guido, M. L., & Dendooven, L. (2007). Emission of carbon dioxide and dynamics of inorganic N in a gradient of alkaline saline soils of the former lake Texcoco. Applied soil ecology 2007, 35(2), 390-403. [CrossRef]
- Bencherif, K., Laruelle, F., Tisserant, B., Dalpé, Y., & Lounés-Hadj Sahraoui, A. Engineering approach for production of arbuscular mycorrhizal inoculum adapted to saline soil management. Stresses 2023, 3(2), 404-423. [CrossRef]
- Ben Laouane, R., Baslam, M., Ait-El-Mokhtar, M., Anli, M., Boutasknit, A., Ait-Rahou, Y., & Meddich, A. Potential of native arbuscular mycorrhizal fungi, rhizobia, and/or green compost as alfalfa (Medicago sativa) enhancers under salinity. Microorganisms 2020, 8(11), 1695. [CrossRef]
- Berthelot, C., Chalot, M., Leyval, C., & Blaudez, D. From darkness to light: emergence of the mysterious dark septate endophytes in plant growth promotion and stress alleviation. Endophytes for a growing world 2019, 143-164. [CrossRef]
- Bertram, N. A., Alfonso, C., Grande, S., Chiacchiera, S., Ohanian, A., Bonvillani, J., & Angeletti, F. R. Efecto de la concentración salina y el régimen hídrico sobre la germinación, emergencia y establecimiento de alfalfa (Medicago sativa L.). RIA. Revista de investigaciones agropecuarias 2021, 47(2), 267-272.
- Biermann, B., & Linderman, R. G. Use of vesicular-arbuscular mycorrhizal roots, intra-radical vesicles and extra-radical vesicles as inoculum. New Phytologist 1983, 95(1), 97-105.
- Campanelli, A., Ruta, C., De Mastro, G., y Morone-Fortunato, I. The role of arbuscular mycorrhizal fungi in alleviating salt stress in Medicago sativa L. var. icon. Symbiosis 2013, 59(2), 65-76. [CrossRef]
- Castro Silva, C. Análisis del cambio en las comunidades microbianas en un suelo salino-alcalino del exlago de Texcoco en presencia de antraceno 2013, Tesis doctoral. Cinvestav, Unidad Zacatenco.
- Chandra, P., Singh, A., Prajapat, K., Rai, A. K., & Yadav, R. K. Native arbuscular mycorrhizal fungi improve growth, biomass yield, and phosphorus nutrition of sorghum in saline and sodic soils of the semi–arid region. Environmental and Experimental Botany 2022, 201, 104982. [CrossRef]
- Cofré, M. N., Becerra, A. G., Nouhra, E. R., & Soteras, M. F. (2012). Arbuscular mycorrhizae and dark-septate endophytes on Atriplex cordobensis in saline sites from Argentina. Journal of Agricultural Technology 2012, 8(7), 2201-2214.
- Corkidi, L., Allen, E. B., Merhaut, D., Allen, M. F., Downer, J., Bohn, J., & Evans, M. (2004). Assessing the infectivity of commercial mycorrhizal inoculants in plant nursery conditions. Journal of Environmental Horticulture 2004, 22(3), 149-154. [CrossRef]
- Currah, R.S., Tsuneda, A., and Murakami, S. Morphology and ecology of Phialocephala fortinii in roots of Rhododendron brachycarpum. Canadian Journal Botany 1993, 71: 1639–1644. [CrossRef]
- Daft, M. J., & Nicolson, T. H. (1972). Effect of Endogone mycorrhiza on plant growth. IV. Quantitative relationships between the growth of the host and the development of the endophyte in tomato and maize. New Phytologist 1972, 287-295.
- David, D. J. Determination of calcium in plant material by atomic-absorption spectrophotometry. Analyst 1959, 84(1002), 536-545. [CrossRef]
- Della Mónica, I. F., Saparrat, M. C., Godeas, A. M., & Scervino, J. M. The co-existence between DSE and AMF symbionts affects plant P pools through P mineralization and solubilization processes. Fungal Ecology 2015, 17, 10-17. [CrossRef]
- Dhiman, P., Rajora, N., Bhardwaj, S., Sudhakaran, S. S., Kumar, A., Raturi, G., ... & Deshmukh, R. Fascinating role of silicon to combat salinity stress in plants: An updated overview. Plant Physiology and Biochemistry 2021, 162, 110-123. [CrossRef]
- Douds Jr, D. D., Nagahashi, G., Pfeffer, P. E., Kayser, W. M., & Reider, C. On-farm production and utilization of arbuscular mycorrhizal fungus inoculum. Canadian Journal of Plant Science 2005, 85(1), 15-21.
- Estrada, B., Aroca, R., Maathuis, F. J., Barea, J. M., & RUIZ-LOZANO, J. M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant, cell & environment 2013, 36(10), 1771-1782. [CrossRef]
- Evelin, H., Giri, B., & Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenumgraecum. Mycorrhiza 2012, 22, 203-217. [CrossRef]
- Evelin, H., Kapoor, R., y Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: a review. Annals of botany 2009, 104(7), 1263-1280. [CrossRef]
- Farias, G. C., Nunes, K. G., Soares, M. A., de Siqueira, K. A., Lima, W. C., Neves, A. L. R., y de Lacerda, C. F. Dark septate endophytic fungi mitigate the effects of salt stress on cowpea plants. Brazilian Journal of Microbiology 2020, 51(1), 243-253. [CrossRef]
- Fuchs, B., & Haselwandter, K. Red list plants: colonization by arbuscular mycorrhizal fungi and dark septate endophytes. Mycorrhiza 2004, 14(4), 277-281. [CrossRef]
- Gaber, D. A., Berthelot, C., Camehl, I., Kovacs, G. M., Blaudez, D., y Franken, P. Salt stress tolerance of dark septate endophytes is independent of melanin accumulation. Frontiers in microbiology 2020, 3158. [CrossRef]
- Gerdemann, J. W., & Nicolson, T. H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological society 1963, 46(2), 235-244. [CrossRef]
- Ghabooli., M. Effect of Piriformospora indica inoculation on some physiological traits of barley (Hordeum vulgare) under salt stress. Chemistry of Natural Compounds 2014, 50:1082–1087. [CrossRef]
- Giri, B., & Mukerji, K. G. Mycorrhizal inoculant alleviates salt stress in Sesbania aegyptiaca and Sesbania grandiflora under field conditions: evidence for reduced sodium and improved magnesium uptake. Mycorrhiza 2004, 14, 307-312. [CrossRef]
- Gonzalez Mateu, M., Baldwin, A. H., Maul, J. E., & Yarwood, S. A. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis. The ISME journal 2020, 14(8), 1943-1954. [CrossRef]
- Grattan, S. R., & Grieve, C. M. Salinity mineral nutrient relations in horticultural crops. Scientia horticulturae 1998, 78(1-4), 127-157. [CrossRef]
- Hammer, E. C., Nasr, H., Pallon, J., Olsson, P. A., & Wallander, H. Elemental composition of arbuscular mycorrhizal fungi at high salinity. Mycorrhiza 2011, 21, 117-129. [CrossRef]
- Hartmond, U., Schaesberg, N. V., Graham, J. H., & Syvertsen, J. P. (1987). Salinity and flooding stress effects on mycorrhizal and non-mycorrhizal citrus rootstock seedlings. Plant and Soil 1987, 104, 37-43. [CrossRef]
- Havre, G. N. The flame photometric determination of sodium, potassium and calcium in plant extracts with special reference to interference effects. Analytica Chimica Acta 1961, 25(6), 557-566. [CrossRef]
- Hawksworth, D. L., & Rossman, A. Y. Where are all the undescribed fungi? Phytopathology 1997, 87(9), 888-891. [CrossRef]
- Hayman, D. S. Endogone spore numbers in soil and vesicular-arbuscular mycorrhiza in wheat as influenced by season and soil treatment. Transactions of the British mycological Society 1970, 54(1), 53-IN12. [CrossRef]
- Hirrel, M.C. The effect of sodium and chloride salts on the germination of Gigaspora margarita. Mycologia 1981, 73:610-617. [CrossRef]
- Hou, L., Yu, J., Zhao, L., & He, X. (2020). Dark septate endophytes improve the growth and the tolerance of Medicago sativa and Ammopiptanthus mongolicus under cadmium stress. Frontiers in microbiology 2020, 10, 3061. [CrossRef]
- INVAM. Results of infectivity (MIP) assays of commercial Inoculants. Available at: http://invam.caf.wvu.edu/otherinfo/commercial/commercial_MIPs.pdf. Accessed 29 December 2011 2011.
- Jahromi, F., Aroca, R., Porcel, R., & Ruiz-Lozano, J. M. Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microbial Ecology 2008, 55, 45-53. [CrossRef]
- Jumpponen, A. R. I., & Trappe, J. M. Dark septate endophytes: a review of facultative biotrophic root-colonizing fungi. The New Phytologist 1998, 140(2), 295-310. [CrossRef]
- Juniper, S. The effect of sodium chloride on some vesicular–arbuscular mycorrhizal fungi. Ph.D. thesis 1996, The University of Western Australia.
- Juniper, S., y Abbott, L. K. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza 2006, 16(5), 371-379. [CrossRef]
- Karagiannidis, N., Bletsos, F., & Stavropoulos, N. Effect of Verticillium wilt (Verticillium dahliae Kleb.) and mycorrhiza (Glomus mosseae) on root colonization, growth and nutrient uptake in tomato and eggplant seedlings. Scientia horticulturae 2002, 94(1-2), 145-156. [CrossRef]
- Karley, A. J., & White, P. J. Moving cationic minerals to edible tissues: potassium, magnesium, calcium. Current opinion in plant biology 2009, 12(3), 291-298. [CrossRef]
- Kassambara, A., y Mundt, F. (2017). Package ‘factoextra’. Extract and visualize the results of multivariate data analyses 2017, 76(2).
- Kendall, C. Tracing nitrogen sources and cycling in catchments. In Isotope tracers in catchment hydrology 1998, (pp. 519-576). Elsevier. [CrossRef]
- Kim, C.K., & Weber D.J. Distribution of VA mycorrhiza on halophytes on inland salt playas. Plant Soil 1985, 83:207-214. [CrossRef]
- Kitson, R. E., & Mellon, M. G. Colorimetric determination of germanium as molybdigermanic acid. Industrial & Engineering Chemistry Analytical Edition 1944, 16(2), 128-130. [CrossRef]
- Knapp, D. G., Kovács, G. M., Zajta, E., Groenewald, J. Z., & Crous, P. W. (2015). Dark septate endophytic Pleosporalean genera from semiarid areas. Persoonia-Molecular Phylogeny and Evolution of Fungi 2015, 35(1), 87-100. [CrossRef]
- Koske, R. E., y Gemma, J. N. A modified procedure for staining roots to detect VA mycorrhizas. Mycological research 1989, 92(4), 486. [CrossRef]
- LaHaye, P., & Epstein, E. Salt toleration by plants: enhancement with calcium. Science 1969, 166(3903), 395-396. [CrossRef]
- Laouane, B., Meddich, A., Bechtaoui, N., Oufdou, K., & Wahbi, S. Effects of arbuscular mycorrhizal fungi and rhizobia symbiosis on the tolerance of Medicago sativa to salt stress. Gesunde Pflanzen 2019, 71(2), 135-146. [CrossRef]
- Latef, A. A. H. A., & Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Scientia Horticulturae 2011, 127(3), 228-233. [CrossRef]
- Levy, Y., Dodd, J., & Krikun, J. Effect of irrigation, water salinity and rootstock on the vertical distribution of vesicular-arbuscular mycorrhiza in citrus roots. New Phytologist 1983, 95(3), 397-403. [CrossRef]
- Maciá-Vicente, J. G., Ferraro, V., Burruano, S., & Lopez-Llorca, L. V. Fungal assemblages associated with roots of halophytic and non-halophytic plant species vary differentially along a salinity gradient. Microbial ecology 2012, 64, 668-679. [CrossRef]
- Maggio, A., Chiaranda, F. Q., Cefariello, R., y Fagnano, M. Responses to ozone pollution of alfalfa exposed to increasing salinity levels. Environmental Pollution 2009, 157(5), 1445-1452. [CrossRef]
- Mahmood, I. A., Arshad, A., Aslam, M., Shahzad, A., Sultan, T., & Hussain, F. Phosphorus availability in different salt-affected soils as influenced by crop residue incorporation. International Journal of Agriculture and Biology 2013, 15(3).
- Malhotra, H., Sharma, S., & Pandey, R. Phosphorus nutrition: plant growth in response to deficiency and excess. Plant nutrients and abiotic stress tolerance 2018, 171-190. [CrossRef]
- Mapperson, R. R., Kotiw, M., Davis, R. A., & Dearnaley, J. D. The diversity and antimicrobial activity of Preussia sp. endophytes isolated from Australian dry rainforests. Current microbiology 2014, 68, 30-37. [CrossRef]
- Marschner, H., & Cakmak, I. High light intensity enhances chlorosis and necrosis in leaves of zinc, potassium, and magnesium deficient bean (Phaseolus vulgaris) plants. Journal of Plant physiology 1989, 134(3), 308-315. [CrossRef]
- Martínez, C. M., Marcos, M. F., & Rodriguez, E. A. Factors influencing phosphorus adsorption in mine soils in Galicia, Spain. Science of the total environment 1996, 180(2), 137-145. [CrossRef]
- Martinez, V., & Läuchli, A. Salt-induced inhibition of phosphate uptake in plants of cotton (Gossypium hirsutum L.). New Phytologist 1994, 126(4), 609-614.
- Mattson Jr, W. J. Herbivory in relation to plant nitrogen content. Annual review of ecology and systematics 1980, 11(1), 119-161.
- McMillen, B. G., Juniper, S., & Abbott, L. K. Inhibition of hyphal growth of a vesicular-arbuscular mycorrhizal fungus in soil containing sodium chloride limits the spread of infection from spores. Soil Biology and Biochemistry 1998, 30(13), 1639-1646. [CrossRef]
- Millner, P. D., y Kitt, D. G. The Beltsville method for soilless production of vesicular-arbuscular mycorrhizal fungi. Mycorrhiza 1992, 2(1), 9-15. [CrossRef]
- Nagy, R., Drissner, D., Amrhein, N., Jakobsen, I., & Bucher, M. (2009). Mycorrhizal phosphate uptake pathway in tomato is phosphorus-repressible and transcriptionally regulated. New Phytologist 2009, 181(4), 950-959. [CrossRef]
- Olsen, S. R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate (No. 939). US Department of Agriculture 1954.
- Olsson, P. A., Hammer, E. C., Pallon, J., Van Aarle, I. M., & Wallander, H. (2011). Elemental composition in vesicles of an arbuscular mycorrhizal fungus, as revealed by PIXE analysis. Fungal biology 2011, 115(7), 643-648. [CrossRef]
- Pan, X., Qin, Y., & Yuan, Z. Potential of a halophyte-associated endophytic fungus for sustaining Chinese white poplar growth under salinity. Symbiosis 2018, 76, 109-116. [CrossRef]
- Phillips, J. M., y Hayman, D. S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British mycological Society 1970, 55(1), 158-IN18. [CrossRef]
- Piercey, M. M., Graham, S. W., & Currah, R. S. Patterns of genetic variation in Phialocephala fortinii across a broad latitudinal transect in Canada. Mycological Research 2004, 108(8), 955-964. [CrossRef]
- Porcel, R., Aroca, R., Azcon, R., & Ruiz-Lozano, J. M. Regulation of cation transporter genes by the arbuscular mycorrhizal symbiosis in rice plants subjected to salinity suggests improved salt tolerance due to reduced Na+ root-to-shoot distribution. Mycorrhiza 2016, 26, 673-684. [CrossRef]
- Porras-Alfaro, A., & Bayman, P. Hidden fungi, emergent properties: endophytes and microbiomes. Annual review of phytopathology 2011, 49, 291-315. [CrossRef]
- Redman, R. S., Anderson, J. A., Biaggi, T. M., Malmberg, K. E., Rienstra, M. N., Weaver, J. L., & Rodriguez, R. J. Symbiotic modulation as a driver of niche expansion of coastal plants in the San Juan Archipelago of Washington State. Frontiers in Microbiology 2022, 13. [CrossRef]
- Rodrı́guez-Navarro, A. (2000). Potassium transport in fungi and plants. Biochimica et Biophysica Acta (BBA)-Reviews on Biomembranes 2000, 1469(1), 1-30. [CrossRef]
- Ruiz-Lozano, J. M., Porcel, R., Azcon, C., & Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: new challenges in physiological and molecular studies. Journal of Experimental Botany 2012, 63(11), 4033-4044. [CrossRef]
- Santoyo de la Cruz, M. F., Flores-Magdaleno, H., Khalil-Gardezi, A., Mancilla-Villa, Ó. R., & Rubiños-Panta, J. E. Composición iónica y comparación de índices de salinidad de suelo agrícola de Texcoco, México. Nova scientia 2021, 13(27).
- Shi, Y. W., Lou, K., Li, C., Wang, L., Zhao, Z. Y., Zhao, S., & Tian, C. Y. Illumina-based analysis of bacterial diversity related to halophytes Salicornia europaea and Sueada aralocaspica. Journal of Microbiology 2015, 53, 678-685. [CrossRef]
- Shrivastava, P., y Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi journal of biological Sciences 2015, 22(2), 123-131. [CrossRef]
- Smith, S. E., y Read, D. J. Mycorrhizal symbiosis. 2008, (Academic Press Ltd: Cambridge, UK).
- Song, M., Chai, Q., Li, X., Yao, X., Li, C., Christensen, M.J, & Nan, Z. An asexual Epichloë endophyte modifies the nutrient stoichiometry of wild barley (Hordeum brevisubulatum) under salt stress. Plant Soil 2015, 387:153–165. [CrossRef]
- Sonjak, S., Udovič, M., Wraber, T., Likar, M., & Regvar, M. Diversity of halophytes and identification of arbuscular mycorrhizal fungi colonising their roots in an abandoned and sustained part of Sečovlje salterns. Soil Biology and Biochemistry 2009, 41(9), 1847-1856. [CrossRef]
- Saqib, M., Zörb, C., & Schubert, S. Silicon-mediated improvement in the salt resistance of wheat (Triticum aestivum) results from increased sodium exclusion and resistance to oxidative stress. Functional Plant Biology 2008, 35(7), 633-639. [CrossRef]
- Tekaya, M., Mechri, B., Mbarki, N., Cheheb, H., Hammami, M., & Attia, F. Arbuscular mycorrhizal fungus Rhizophagus irregularis influences key physiological parameters of olive trees (Olea europaea L.) and mineral nutrient profile. Photosynthetica 2017, 55(2), 308-316. [CrossRef]
- Trejo-Aguilar, D., & Banuelos, J. Isolation and culture of arbuscular mycorrhizal fungi from field samples. Arbuscular Mycorrhizal Fungi: Methods and Protocols 2020, 1-18. [CrossRef]
- Valenzuela-Encinas, C., Neria-González, I., Alcántara-Hernández, R. J., Enríquez-Aragón, J. A., Estrada-Alvarado, I., Hernández-Rodríguez, C., & Marsch, R. Phylogenetic analysis of the archaeal community in an alkaline-saline soil of the former lake Texcoco (Mexico). Extremophiles 2008, 12, 247-254. [CrossRef]
- Vergara, C., Araujo, K. E. C., Sperandio, M. V. L., Santos, L. A., Urquiaga, S., y Zilli, J. É. Dark septate endophytic fungi increase the activity of proton pumps, efficiency of 15N recovery from ammonium sulphate, N content, and micronutrient levels in rice plants. Brazilian Journal of Microbiology 2019, 50(3), 825-838. [CrossRef]
- Vergara, C., Araujo, K. E., Urquiaga, S., Schultz, N., Balieiro, F. D. C., Medeiros, P. S., & Zilli, J. E. Dark septate endophytic fungi help tomato to acquire nutrients from ground plant material. Frontiers in Microbiology 2017, 8, 2437. [CrossRef]
- Wang, F. Y., Liu, R. J., Lin, X. G., & Zhou, J. M. Arbuscular mycorrhizal status of wild plants in saline-alkaline soils of the Yellow River Delta. Mycorrhiza 2004, 14, 133-137. [CrossRef]
- Waqas, M., Khan, A.L., Kamran M, Hamayun, M., Kang S-M, Kim Y-H, Lee I-J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules 2012, 17:10754–10773. [CrossRef]
- Xie, W., Yang, J., Gao, S., Yao, R., & Wang, X. The effect and influence mechanism of soil salinity on phosphorus availability in coastal salt-affected soils. Water 2022, 14(18), 2804. [CrossRef]
- Yin, L., Ren, A., Wei, M., Wu, L., Zhou, Y., Li X., & Gao, Y. Neotyphodium coenophialum-infected tall fescue and its potential application in the phytoremediation of saline soils. International Journal Phytoremediation 2014, 16:235–246. [CrossRef]
- Yuan, Z., Druzhinina, I. S., Gibbons, J. G., Zhong, Z., Van de Peer, Y., Rodriguez, R. J., & Martin, F. M. Divergence of a genomic island leads to the evolution of melanization in a halophyte root fungus. The ISME journal 2021, 15(12), 3468-3479. [CrossRef]
- Yuan, Z., Druzhinina, I. S., Labbé, J., Redman, R., Qin, Y., Rodriguez, R., ... & Lin, F. Specialized microbiome of a halophyte and its role in helping non-host plants to withstand salinity. Scientific reports 2016, 6(1), 32467. [CrossRef]
- Yun P, Xu L, Wang S-S, Shabala L, Shabala S, Zhang W. Y. Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoots. Plant Growth Regul 2018, 86:323–331. [CrossRef]
- Zare-Maivan, H., Khanpour-Ardestani, N., & Ghanati, F. Influence of mycorrhizal fungi on growth, chlorophyll content, and potassium and magnesium uptake in maize. Journal of plant Nutrition 2017, 40(14), 2026-2032. [CrossRef]
- Zhang, Q., Acuña, J. J., Inostroza, N. G., Mora, M. L., Radic, S., Sadowsky, M. J., & Jorquera, M. A. Endophytic bacterial communities associated with roots and leaves of plants growing in Chilean extreme environments. Scientific reports 2019, 9(1), 4950. [CrossRef]
- Zuccarini, P., & Okurowska, P. Effects of mycorrhizal colonization and fertilization on growth and photosynthesis of sweet basil under salt stress. Journal of Plant Nutrition 2008, 31(3), 497-513. [CrossRef]
Figure 1.
Endorhizal root colonization in alfalfa as host plant in trap cultures. Alfalfa inoculated with soil: a) Consortium 1, b) Consortium 3, c) Consortium 6; or roots: d) Consortium 3, e) Consortium 1 and f) Consortium 7. Structures of arbuscular mycorrhizal fungi= arbuscules (A), vesicles (V), and hyphae (H). Structures of dark septate fungi= microsclerotium (ME) and dark septate hyphae (HS).
Figure 1.
Endorhizal root colonization in alfalfa as host plant in trap cultures. Alfalfa inoculated with soil: a) Consortium 1, b) Consortium 3, c) Consortium 6; or roots: d) Consortium 3, e) Consortium 1 and f) Consortium 7. Structures of arbuscular mycorrhizal fungi= arbuscules (A), vesicles (V), and hyphae (H). Structures of dark septate fungi= microsclerotium (ME) and dark septate hyphae (HS).

Figure 2.
Colonization by arbuscular mycorrhizal fungi (a) and dark septate fungi (b). Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard deviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
Figure 2.
Colonization by arbuscular mycorrhizal fungi (a) and dark septate fungi (b). Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard deviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
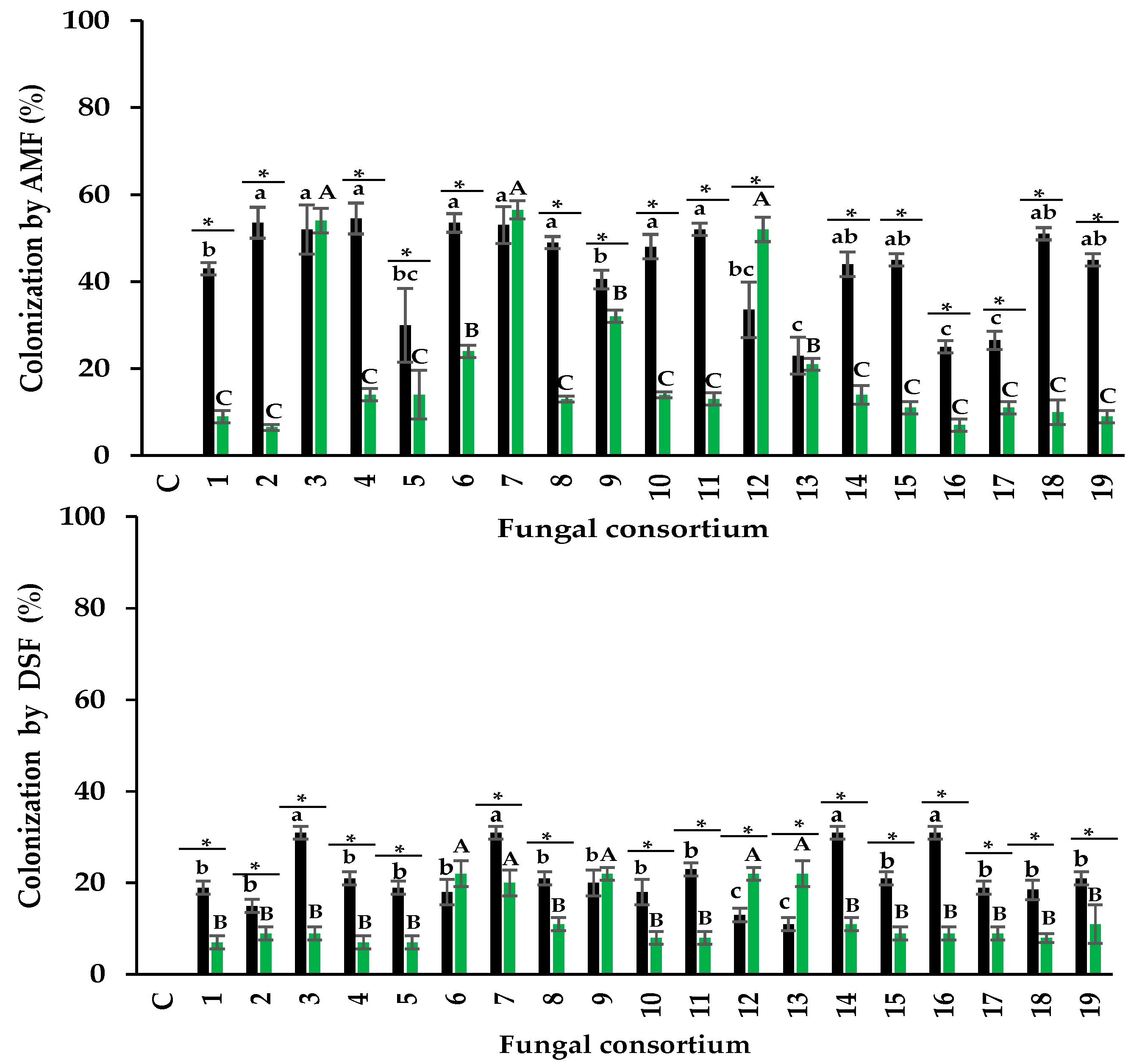
Figure 3.
Number of spores in alfalfa trap cultures with two types of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
Figure 3.
Number of spores in alfalfa trap cultures with two types of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
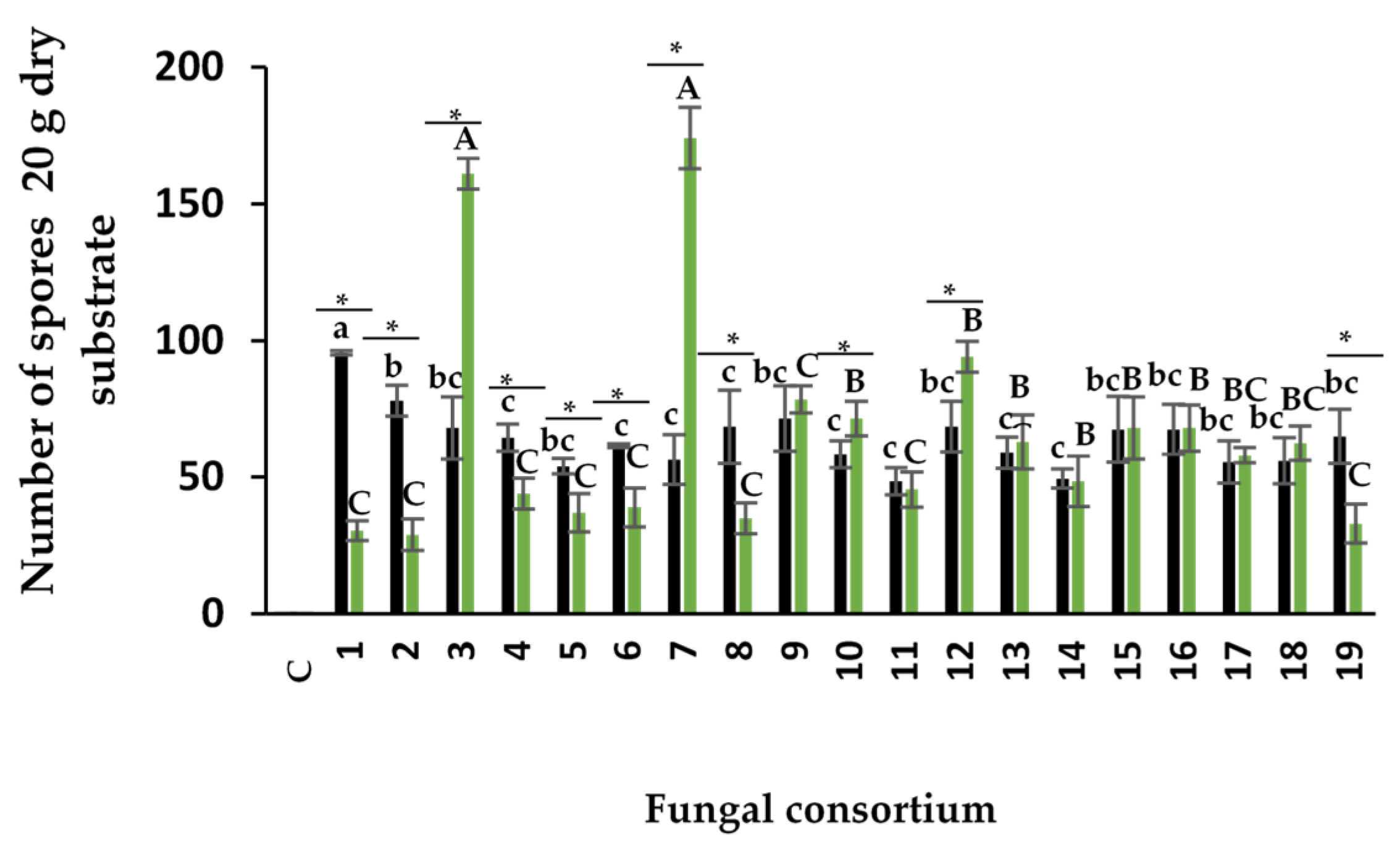
Figure 4.
Fresh weight of the aerial part in alfalfa culture with two types of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
Figure 4.
Fresh weight of the aerial part in alfalfa culture with two types of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
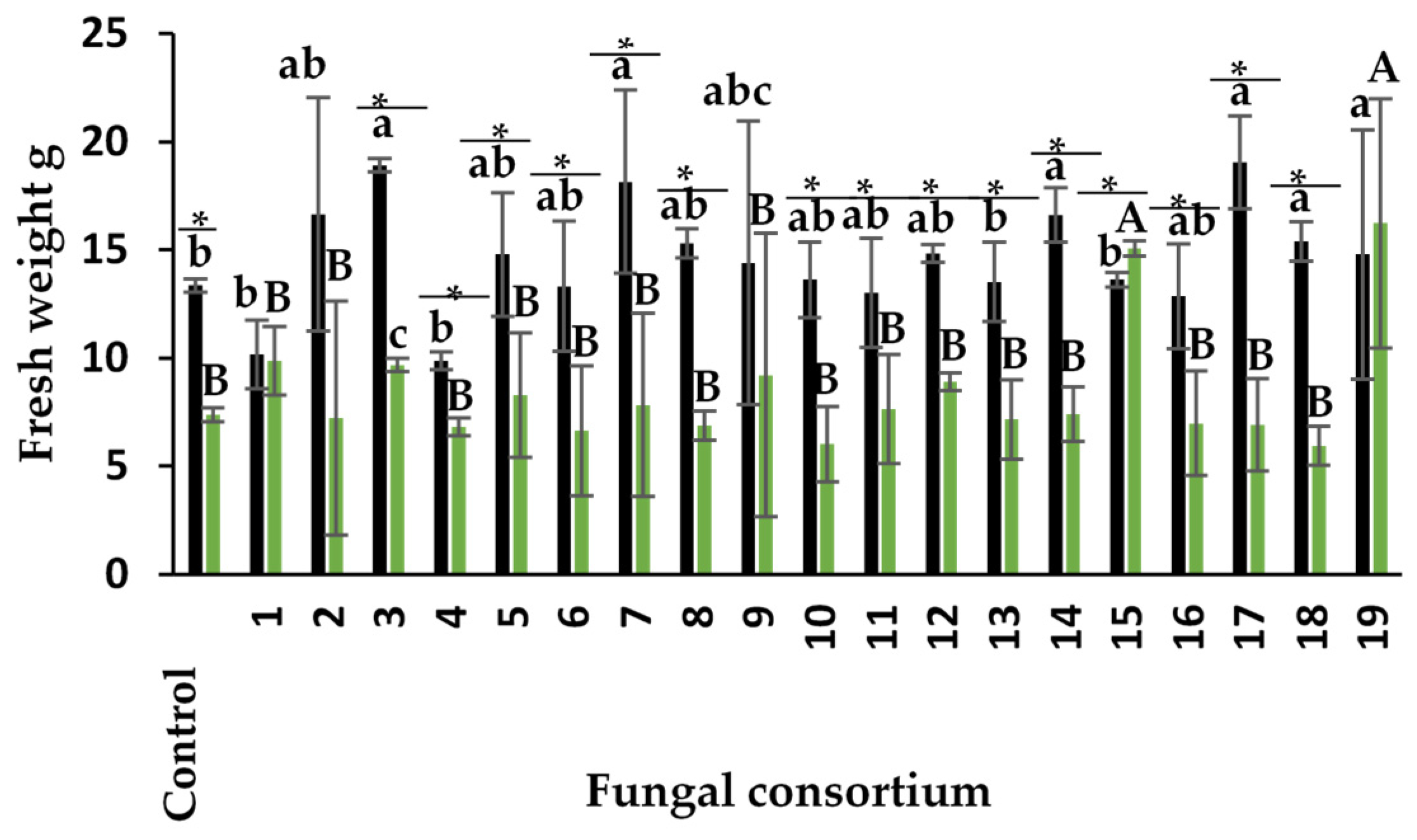
Figure 5.
Influence of inoculation of fungal consortia on foliar nutrient concentration. a) foliar nutrient concentration of nitrogen and b) phosphorus in alfalfa trap culture with two sources of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
Figure 5.
Influence of inoculation of fungal consortia on foliar nutrient concentration. a) foliar nutrient concentration of nitrogen and b) phosphorus in alfalfa trap culture with two sources of inoculum. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
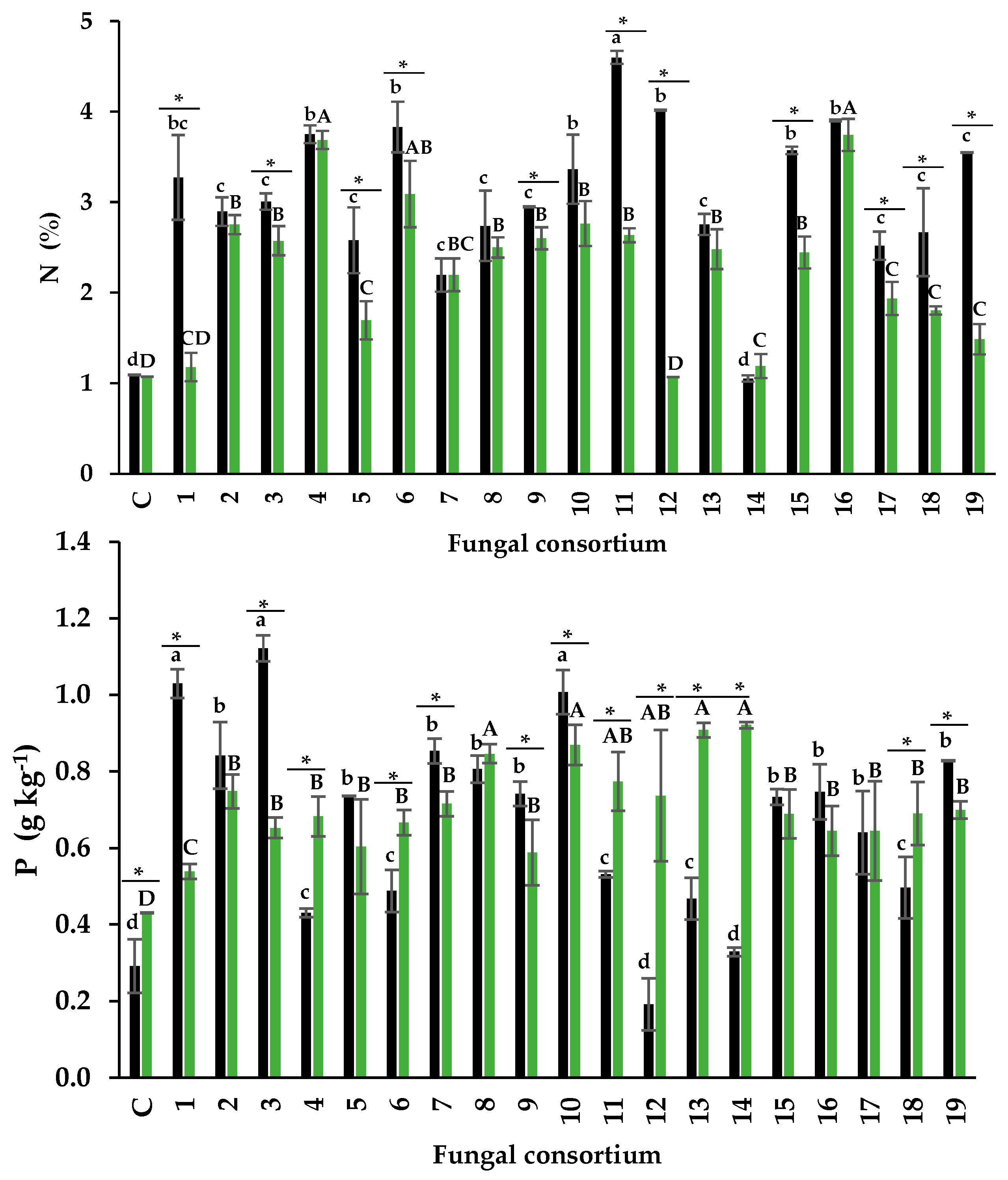
Figure 6.
Foliar concentration of Na and osmoprotective elements in alfalfa as fungal trap culture to propagate endophytic fungi. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
Figure 6.
Foliar concentration of Na and osmoprotective elements in alfalfa as fungal trap culture to propagate endophytic fungi. Black bars= inoculum soil and green bars= root inoculum. Values correspond to average ± standard desviation, n= 3. Different lowercase letters show differences in colonization when comparing among fungal consortia using soil as inoculum (Tukey, α=0.05). Different capital letters show differences in colonization when comparing among fungal consortia using roots as inoculum (Tukey, α=0.05). *Show significant difference in the percentage of colonization of each fungal consortium when comparing between soil and root inoculum (Tukey, α=0.05).
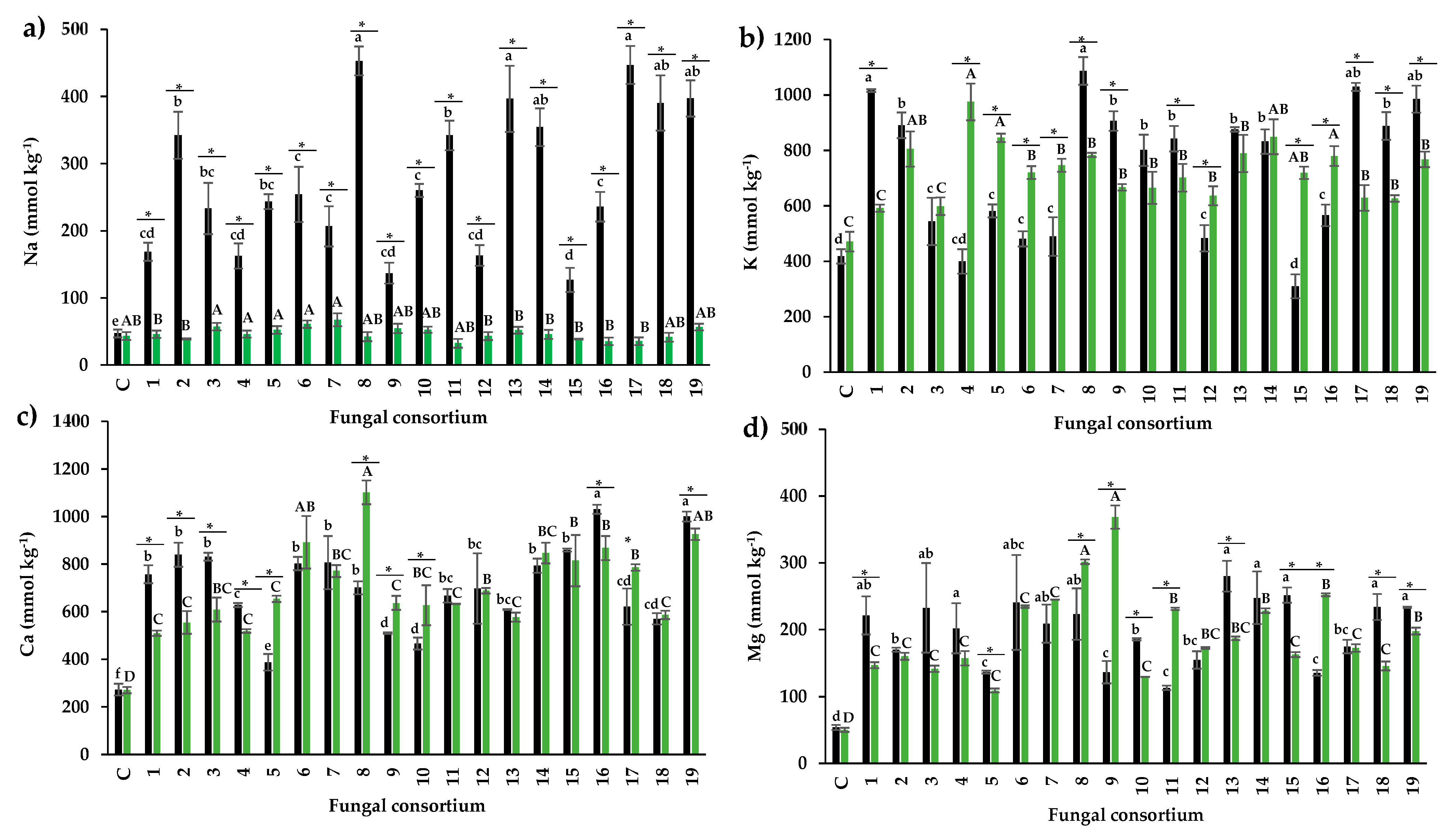
Figure 7.
Micrographs acquired with an environmental electron microscopy of spores and hyphae from the alfalfa trap culture and the elemental content obtained by EDX. a) Consortium 2, b) Consortium 8, both from soil used as inoculum in trap cultures.
Figure 7.
Micrographs acquired with an environmental electron microscopy of spores and hyphae from the alfalfa trap culture and the elemental content obtained by EDX. a) Consortium 2, b) Consortium 8, both from soil used as inoculum in trap cultures.
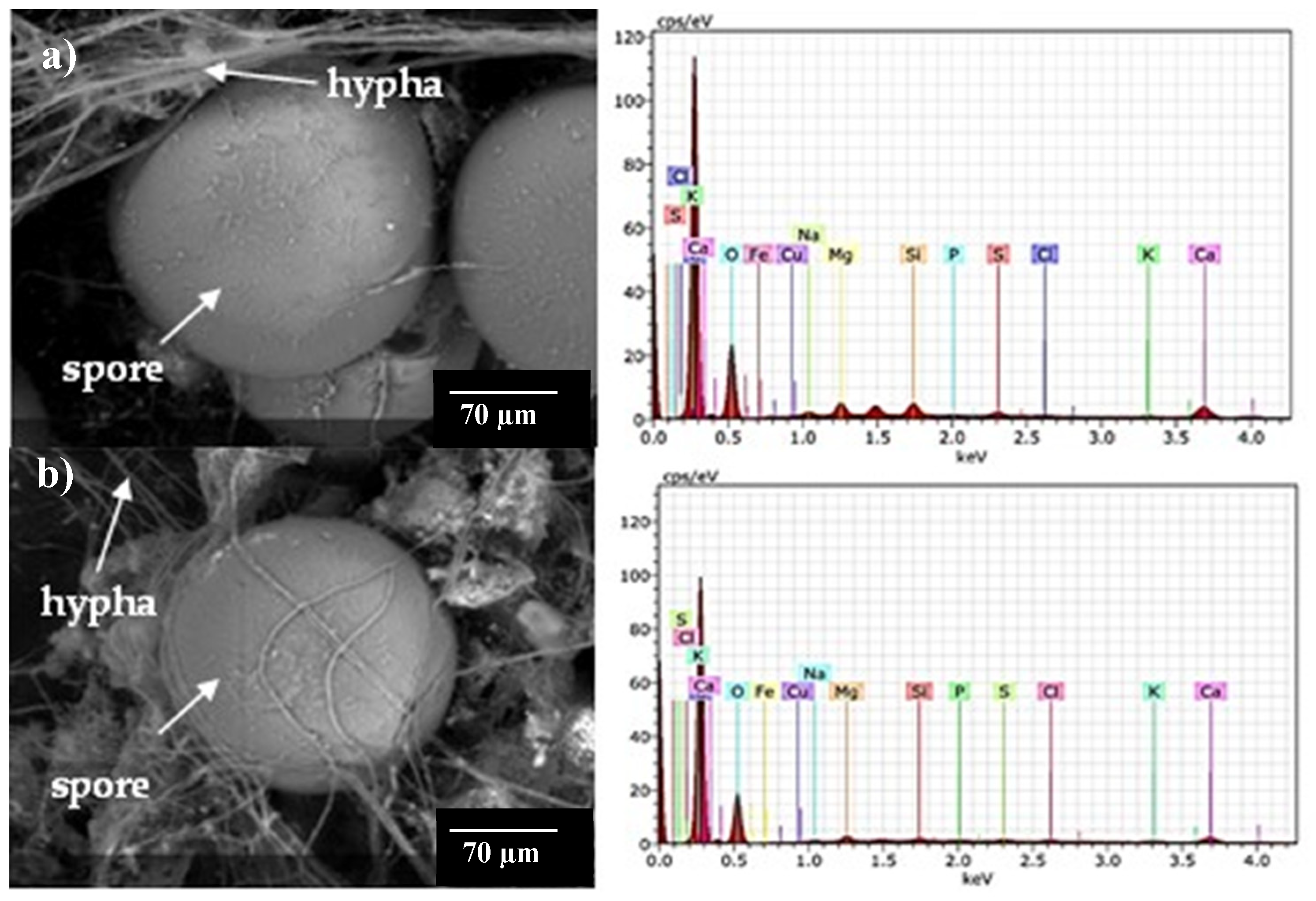
Figure 8.
Principal component analysis of data from infectivity and effectivity responses in alfalfa plants used as fungal trap culture inoculated with soil or roots from halophytes established in saline soils. Variables related to fungi (blue arrows), plants (green arrows) and original soil variables (gray arrows). AMF=colonization by arbuscular mycorrhizal fungi or by dark septate fungi (DSF); Concentration of elements in soil= chemical symbols or foliar alfalfa concentration= chemical symbols with f, and EC=Electrical conductivity. The numbers refer to consortium from soil (black color) or from roots (green color).
Figure 8.
Principal component analysis of data from infectivity and effectivity responses in alfalfa plants used as fungal trap culture inoculated with soil or roots from halophytes established in saline soils. Variables related to fungi (blue arrows), plants (green arrows) and original soil variables (gray arrows). AMF=colonization by arbuscular mycorrhizal fungi or by dark septate fungi (DSF); Concentration of elements in soil= chemical symbols or foliar alfalfa concentration= chemical symbols with f, and EC=Electrical conductivity. The numbers refer to consortium from soil (black color) or from roots (green color).
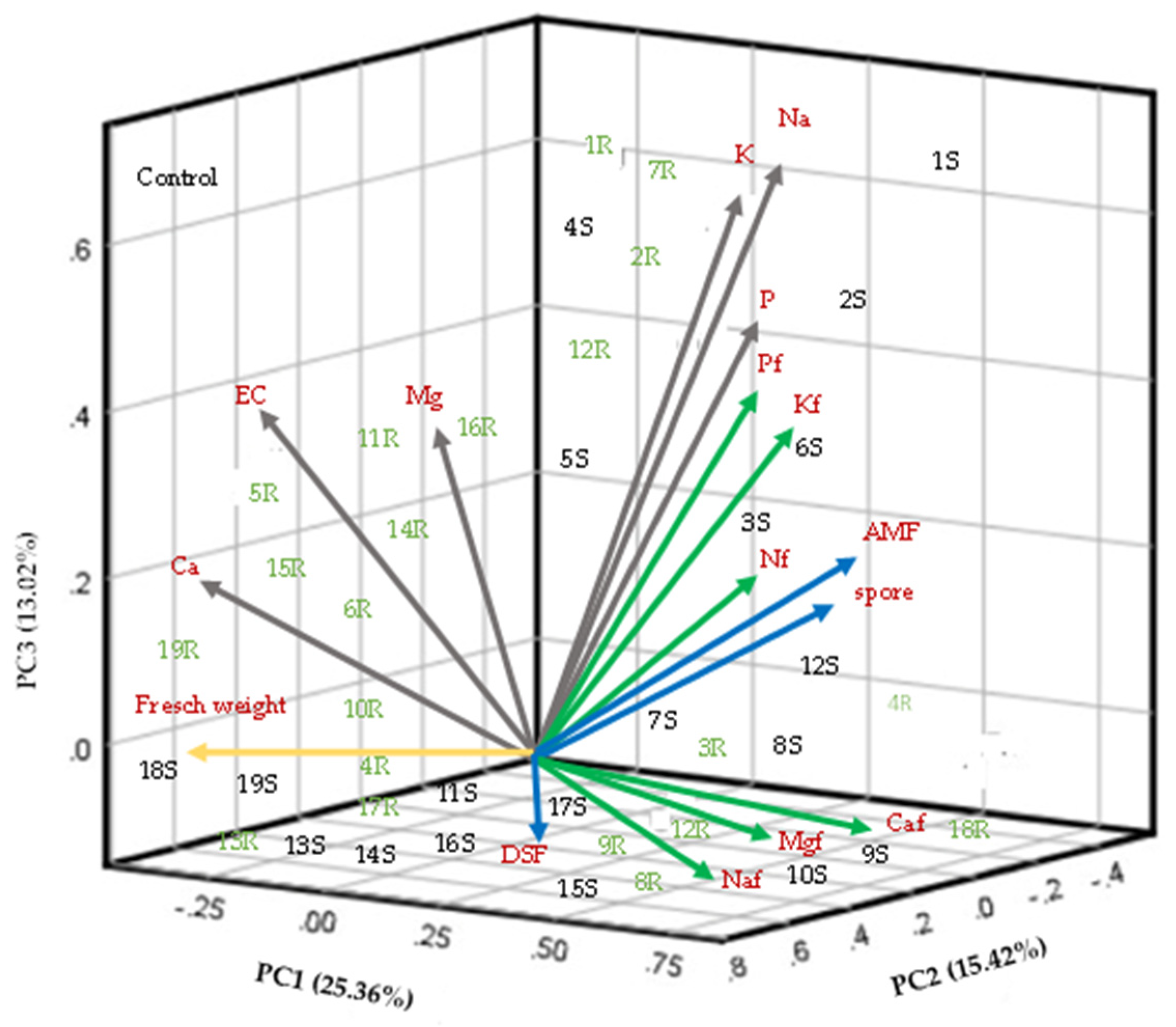
Figure 9.
Arbuscular mycorrhizal fungi collected after and before soil Consortium 1 trap culture in alfalfa. a) Funneliformis mosseae, b) Rhizophagus aggregatus, c) Claroideoglomus claroideum and d) Diversispora sp.
Figure 9.
Arbuscular mycorrhizal fungi collected after and before soil Consortium 1 trap culture in alfalfa. a) Funneliformis mosseae, b) Rhizophagus aggregatus, c) Claroideoglomus claroideum and d) Diversispora sp.
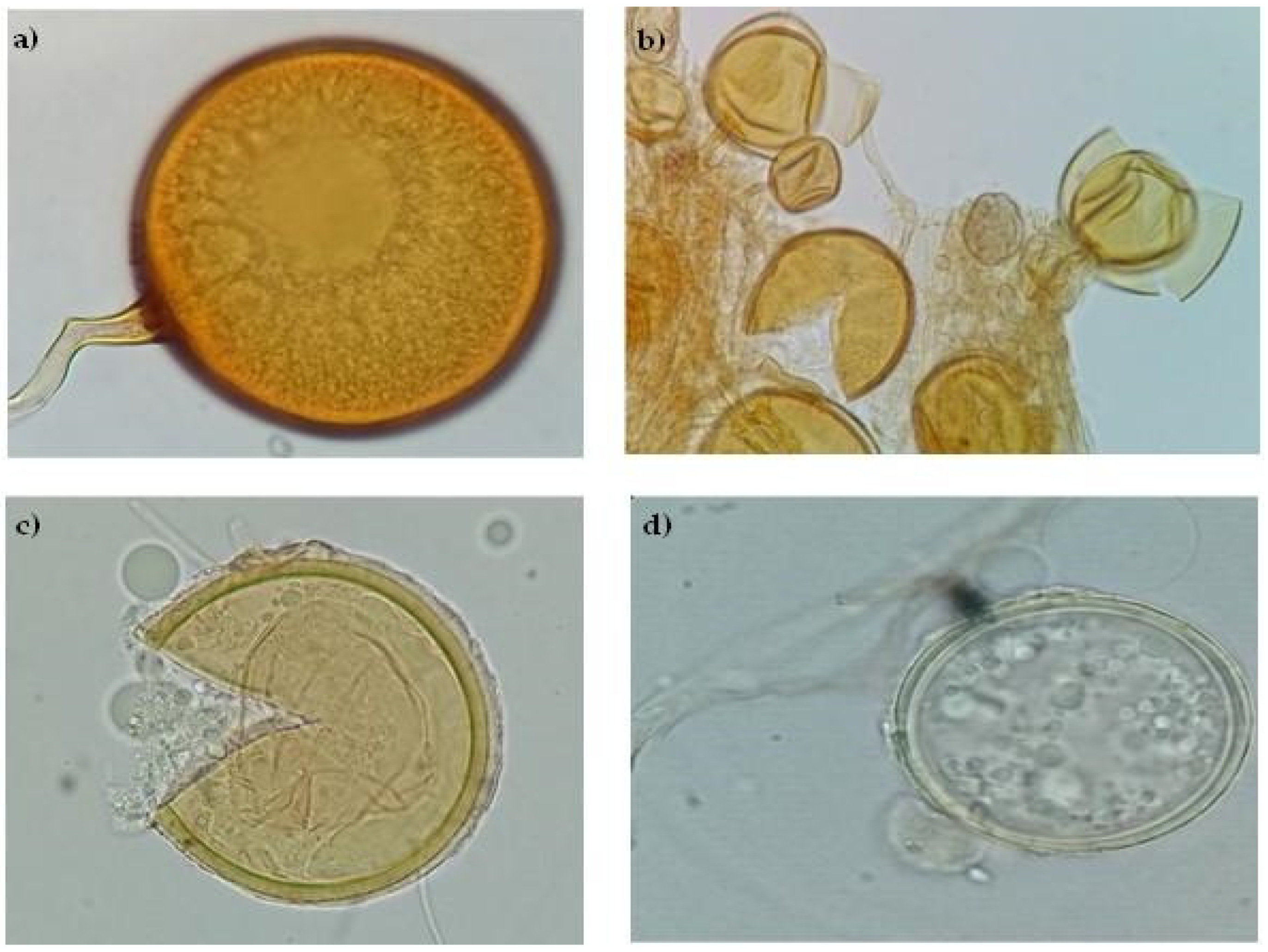
Figure 10.
Analysis of the taxonomic annotation for endophytic microorganisms found in Distichlis spicata , where Consortium 1, originated. a) bacteria and b) fungi.
Figure 10.
Analysis of the taxonomic annotation for endophytic microorganisms found in Distichlis spicata , where Consortium 1, originated. a) bacteria and b) fungi.


Table 1.
Cations and phosphorus concentration, electrical conductivity (EC), pH, and Na adsorption ratio (SAR) in soil compound samples from halophyte species in saline soils used for endorhizal fungal trap cultures.
Table 1.
Cations and phosphorus concentration, electrical conductivity (EC), pH, and Na adsorption ratio (SAR) in soil compound samples from halophyte species in saline soils used for endorhizal fungal trap cultures.
| Consortium | # of individual soil sample † | Cations concentration (mM)†† |
EC (dS m-1) †† | pH†† | SAR (mM)†† | Soil type | PO4-2 Olsen (mg kg-1) †† |
Vegetal species | ||||
| Na | K | Ca | Mg | S | SS | |||||||
| 1 | 5 | 655.9 | 60.2 | 2.0 | 0.7 | 42.5 | 7.8 | 580.4 | + | 10.91 | Distichlis spicata | |
| 2 | 8, 10 | 585.3 | 70.8 | 5.1 | 1.4 | 36.3 | 8.1 | 325.1 | + | 15.46 | Distichlis spicata | |
| 3 | 19 | 322.9 | 39.3 | 3.3 | 0.8 | 20.3 | 8.1 | 225.1 | + | 12.19 | Kochia scoparia | |
| 4 | 29 | 272.5 | 15.2 | 3.0 | 0.5 | 17.5 | 9.2 | 206.4 | + | 15.12 | Eragrostis obtusiflora | |
| 5 | 13 | 222.0 | 18.3 | 3.3 | 0.8 | 15.4 | 8.3 | 154.0 | + | 13.86 | Distichlis spicata | |
| 6 | 2, 18 | 212.0 | 24.7 | 3.3 | 2.4 | 14.8 | 8.5 | 213.6 | + | 15.69 | Suaeda torreyana | |
| 7 | 6 | 94.9 | 12.6 | 2.3 | 1.5 | 8.5 | 8.1 | 69.2 | + | 13.28 | Distichlis spicata | |
| 8 | 9, 11 | 131.2 | 18.6 | 4.4 | 3.1 | 9.1 | 8.4 | 67.2 | + | 16.49 | Baccharis salicifolia | |
| 9 | 20, 23 | 52.4 | 16.8 | 8.2 | 2.1 | 8.7 | 8.6 | 23.0 | + | 12.91 | Distichlis spicata | |
| 10 | 7 | 80.8 | 9.1 | 3.9 | 1.6 | 6.0 | 9.4 | 49.0 | + | 17.38 | Kochia scoparia | |
| 11 | 12, 14, 16 | 181.6 | 12.3 | 5.7 | 3.2 | 6.4 | 9.6 | 86.5 | + | 15.00 | Eragrostis obtusiflora | |
| 12 | 32, 37 | 212.0 | 12.6 | 3.1 | 1.7 | 6.6 | 8.2 | 135.2 | + | 6.41 | Eragrostis obtusiflora | |
| 13 | 17 | 50.5 | 9.1 | 3.9 | 1.0 | 3.8 | 8.5 | 32.5 | + | 8.13 | Cynodon dactylon | |
| 14 | 4 | 52.5 | 8.1 | 4.9 | 1.4 | 3.1 | 8.7 | 29.6 | + | 9.96 | Baccharis salicifolia | |
| 15 | 3 | 131.1 | 8.1 | 6.0 | 1.4 | 2.6 | 8.7 | 68.3 | + | 16.15 | Kochia scoparia | |
| 16 | 21, 22 | 38.3 | 7.3 | 3.9 | 3.0 | 2.2 | 8.4 | 20.6 | + | 10.76 | Distichlis spicata | |
| 17 | 1, 27, 28, 30, 31, 33 | 141.3 | 3.4 | 3.1 | 1.2 | 3.8 | 7.9 | 96.8 | + | 7.18 | Eragrostis obtusiflora | |
| 18 | 15, 34, 35, 36, 38 | 31.3 | 6.3 | 6.3 | 3.3 | 2.0 | 8.0 | 14.3 | + | 8.79 | Baccharis salicifolia | |
| 19 | 24, 25 | 31.3 | 6.0 | 5.2 | 1.6 | 0.9 | 8.4 | 17.0 | + | 12.14 | Kochia scoparia | |
| Normal in the soil solution | 0.2††† | 20†††† | 0.1‡ | 8‡‡ | ||||||||
† Individual soil samples (38) from the rhizosphere halophytes established in saline (S) or sodic saline (SS) soil. †† Values correspond to the final sample used in the fungal trap cultures (consortium). ††† Data accoring to Rodríguez-Navarro (2000), †††† Borax (2002), ‡ Wood et al. (2005), ‡‡ Karley and White (2009).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



