
Submitted:
28 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Microalgae are abundant components of biosphere rich in low molecular weight carbohy-drate-containing natural products (glycoconjugates). Glycoconjugates take a part in processes of photosynthesis, provide producers with important biological properties, influence other organ-isms and known by biological activities. Some of them, for example glycosylated toxins and ar-senicals, are detrimental and can be transferring via food chains into higher organisms including humans. So far, the studies on a series of particular groups of microalgal glycoconjugates were not comprehensively discussed in special reviews. In this review, a special focus was made on the isolation, structure determination, properties and approaches to the search for new bioactive metabolites. Analysis of literature data concerning structures, functions and biological activities of ribosylated arsenicals, galactosylated and sulfoquinovosylated lipids, phosphoglycolipids, gly-coside derivatives of toxins, and other groups of glycoconjugates was carried out and discussed. The discovery of a great variety of new carbohydrate-containing metabolites, important biological roles some of them, participation in regulation of microalgal blooms via viral infections as well as the detection of potent immunomodulatory activity, shown by these type metabolites could be attributed to the most interesting recent achievements. Structures, properties and biological roles of glycoconjugates from microalgae attract a great attention growing year to year.
Keywords:
microalgae
; glycoconjugates
; arsenicals
; galactolipids
; sulfoquinovosyl-containing glycolipids
; phosphoglycolipids
; steryl glycosides
; glycosylated long chain polyketides
; regulation of mi-croalgal blooms
; dietary constituents.
1. Introduction
The low molecular weight natural products such as glycero- and sphingoglycolipids, glycosylated fatty alcohols derivatives, carbohydrate-containing toxins, steroid and other glycosides, some ribose-containing arsenicals all belong to so-called glycoconjugates. Being key cellular components and participating in the both inter- and intracellular communications, metabolites of this type are ubiquitous in nature and influence important biologic properties such as fluidity and permeability of biomemranes, stimulation of apoptosis, and defense against predators. They take part in the molecular recognition processes by interactions with different target biomolecules and cells. Microalgae are producers of nutritionally important polyunsaturated fatty acids of ω-3 (n-3) series, having a wide medicinal application as well as of pigments, antioxidants, and diverse other bioregulators, which are accumulated via food chains in many marine organisms, including edible species of fish and mollusks. Glycoconjugates are a portion of a great plenty of various chemicals produced by microalgae.
Microalgae represent wide-spread eukaryotic microorganisms, populating fresh, brackish, sea waters, and bottom sediments and having economic significance growing year to year [1]. A part of microalgal species are symbionts living in host organisms, for example in corals. As a rule, microalgae are capable of performing photosynthesis, provide about a half of atmospheric oxygen and the main part of organic substances of our planet, supporting more about 60% of the total primary production in marine ecosystems [2]. Diatoms, dinoflagellates, coccolithophores, microscopic green and red microalgae, and representatives of some smaller taxa are related to different groups of microalgae, in total belonging to more than 50000 species. This provides significant biochemical diversity in them. In this review, we do not consider metabolites of cyanobacteria (cyanophytes), microorganisms, which like algae are autotrophs capable to photosynthesis with the release of oxygen, but have more similarity with bacteria than with low plants.
Glycoconjugates in microalgae form complex and difficult-to-separation mixtures of biomolecules, which are increasingly being studied by the HPLC/MS method, although it does not allow for fully characterizing structures, for example for details of stereochemistry. That is why, in this article we give structural formulae only for such carbohydrate-containing metabolites that have been obtained and purified in amounts sufficient to carry out structure analyses using NMR methods and chemical transformations. In other cases, structures of the studied molecules are described by their abbreviations.
Quite recently, we published review articles concerning low molecular weight metabolites from diatom microalgae [3] and the studies on such microalgal glycoconjugates as steryl glycosides and sphingosides [4]. The current work covering period from 1989 to 2022 is the result of an expansion and continuation of our preliminary efforts.
2. Glycosylated arsenicals
After the discovery of arsenic-containing compounds in fishes in the 1920s [5], a variety of close related natural products of this type were found in macro- and microphytes as well as in different marine animals. Arsenic specification, toxicity and metabolism of these compounds in microalgae were discussed in review of Wang et al. [6]. Typically, arsenic presents in sea water in concentration of 1–2 μg per liter and are implemented from environment into living systems in result of metabolic processes, leading to more bioavailable products. A variety of water-soluble arsenic species in marine microalgae includes not only arsenosugars, but also inorganic species such as arsenate (V) and its methylated derivatives. Chlorophytes produce glycerol- and phosphate arsenosugars, whereas glycerol-, phosphate-, and sulfate-containing arsenoribosides as well as dimethyl arsenate are more common in heterokontophytes, for example diatoms. Some arsenic-containing metabolites such as glycerol arsenosugars (1), phosphate arsenosugars (2), sulfate arsenosugars (3), and sulfonate arsenosugars (4) (Figure 1) along with glycosylated lipid-soluble arsenolipids from microalgae belong to glycoconjugates and together make up a part of relatively large class of natural compounds, so-called arsenicals. In this class, more than 70 arsenosugar compounds with established structures are known. Although their inorganic moieties can include either As (III) or As (V), the latter forms are far predominating. On the contrary with arsenosugars, arsenolipids only sometimes contain monosaccharide residues and represent lipid-soluble and frequently volatile natural products. The both arsenosugars and arsenolipids can be transferred via food chain into invertebrates and higher animals including humans. These compounds are more or less detrimental due to their toxicity and cytotoxicity.
Arsenic toxicity is a global problem. Millions of people are exposed with As-containing substances through drinking water and food. High arsenic content can induce chronic toxicity and in some cases cancer. In some individuals, acute, subacute, or chronic poisonings, characterizing by skin lesions, cardiovascular symptoms, and in some cases multi-organ failure may be developed [7].
Historically, As-containing preparations such as Salvarsan (arsphenamine) for the treatment of syphilis and arsenic trioxide for cancer were among drugs, which found wide application in medicine. The useful properties of the latter were recently rediscovered [8,9].
Arsenosugars (1–4) were found in different diatom and green microalgae such as Phaeodactylum tricornutum, Thalassiosira pseudonana, Ostreococcus tauri, Dunaliella tertiolecta, and others [10,11,12,13,14]. Generally, arsenosugars contain ribose or its derivatives, at that marine microrganisms produce arsenoribosides in higher concentrations when compared with fresh-water those. Total arsenic concentrations in algae (0.1 – 10 µg/g of dry mass) are higher in 10–100 times in comparison with terrestrial plants [15].
Lipid-soluble fractions of arsenic-containing compounds are characterized by a great diversity of metabolites. Frequently, arsenicals are analogous of common phospholipids with the change of phosphorus for arsenic. Totally, more than 100 different lipid-soluble arsenicals were isolated [16]. Very recently, Chinese scientists reported that over 300 species of naturally occurring-organoarsenicals were identified by the modern analytical methods, particularly HPLC/MS as a promising option [17].
The unicellular green alga Dunaliella tertiolecta is one of most known capable to photosynthesis marine microorganism belonging to the order Chlamydomonades. This species, like other lower plants of the genus Dunaliella, survives in hypersaline environments. Along with several previously known arsenosugars, the novel lipid-soluble arsenical phytyl 5-dimethylarsinoyl-2-O-methyl-ribofuranoside (AsSugPytol546) (5) (Figure 2) was detected in cultured microalgae D. tertiolecta as well as in extracts from oceanic phytoplankton by high resolution electrospray mass spectrometry (HR ESI MS).
In order to obtain the compound 5 in quantities sufficient for the use of NMR spectroscopy and determine its structure, this microalga was cultured in an arsenate-enriched medium. About 2 g of cultured dried cells were used for solvent partition, column chromatography on silica gel and preparative reverse-phase HPLC. As result, it was possible to obtain only a trace amount of the target compound (about 100 micrograms). Nevertheless, subsequent NMR analysis and comparison of tandem mass spectrum (MS) of the obtained arsenolipid with that of the synthesized model methyl-5-dimethylarsinoyl-2-O-methylriboside allowed establishing the structure of 5 and showed that it contains phytol as an aglycone. Glycon moiety of this arsenolipid consisted of 5-dimethylarsinoyl 2-O-methylribose, and therefore methoxy group replaces a sugar hydroxyl in this unusual compound. This structural feature, characteristic of some RNAs, was not previously found outside the RNAs world [18].
A possible pathway of the biosynthesis of AsSugPhytol546 was proposed to include the introduction of arsenic group into ribosyl moiety of S-adenosylmethionine (6→7) followed by 2-O-methylation by the corresponding methylase to yield 8. The loss of adenosyl residue at the catalysis by adenosyl nucleosidase to give ribose derivative (9), and finally interaction of the latter with phytyl diphosphate, catalyzed by riboside phytolsynthase, led to 5 (Figure 2).
In addition to the major arsenolipid 5, a new metabolite, designated as AsSugPL978 (10), accompanied with related AsSugPL958 (11) was found in the same culture. It is of particular interest that more than half of total arsenic presents in D. tertiolecta as lipid-soluble forms. Although the relative amount of total arsenolipids remained constant in different cultivation conditions, an increase in content of hydrophylic arsenosugars was indicated under various As/P regimes with rising As concentrations in media [19].
Therefore, the accumulation of arsenic species from environment into the above-mentioned microalgae stimulates the biosynthesis of various arsenic-containing glycoconjugates. Some arsenic-containing natural products of this type could occur in many popular sea foods. Further progress in the search for this type compounds was connected with using HPLC/MS after chemical derivatization [19]. It should be noted that that some oleaginous microalgae, for example Scenedesmus sp. IITRIND2 are super accumulators of arsenic. Being stressed by arsenic intake, arsenic-tolerant species modulate own cellular processes and enhance the lipid production by accumulating large lipid droplets. The biodiesel, obtained from such microalgae was comparable to plant oil methyl esters and had high cetane number and oxidative stability [20].
3. Glycolipids
Basic research of issues related to the evolution of organisms and their biochemical systems, for example, to the evolution of photosynthetic processes from cyanobacteria to microalgae, attract an attention to autotrophic microalgae and their metabolites. Glycolipids are products of mixed biogenesis, containing the both lipid and carbohydrate moieties, and represent microalgae metabolites of a particular interest and importance.
3.1. Galactolipids of microalgae
Galactolipids as a subgroup of glycolipids are major membrane constituents of microalgal plastids, which play an important role in harvesting and conversion of sunlight energy. The presence of galactolipids in thylakoids of plastids supports maximal photosynthetic efficiency as was shown by indication of specific interactions between the galactolipid head groups and photosynthetic protein complexes in plants. Exactly, it explains the preference for galactose in thylakoid lipids not only in lower, but also in many higher plants [21]. Due to their high content in thylakoid membranes and the wide distribution in lower and higher plants, galactolipids are considered to be the most abundant lipid class in the biosphere [22].
Membranes of plastids mainly are comprised of monogalactosyldiacylglycerols (MGDG, 12) and digalactosyldiacylglycerols (DGDG, 13) along with less abundant sulfoquinovosyldiacylglycerols (SQDG, 14). Galactosyl monoacyl glycerols (MGMG) 15 and 16 differ each other in position of an acyl group, attached to glycerol residue (Figure 3). Such metabolites were found in microalgae much less frequently than MGDG and DGDG. Structurally, wide-distributed in plants MGDGs are characterized by the presence of one D-galactose, β-anomerically linked to the sn-3 position of glycerol backbone. DGDGs contain an additional terminal D-α-galactose, attached to the inner β-galactose residue by 1,3-bond. SQDGs consist of β-D-6-sulfoquinovose and 1,2-diacylglycerol linked each other in the same manner as in MGDG.
Metabolites of general formulae 12 and 13 maintain chloroplast morphology and survival of plants under abiotic stresses. Lipids, making up the plastid membranes in algae are biosynthesized by either prokaryotic or eukaryotic pathways, located within the plastids and in the endoplasmic reticulum, respectively. Thylakoid glycolipidome was formed in the process of evolution from cyanobacteria via lower plants to higher plants. In microalgae, glycolipids are enriched by useful for human health polyunsaturated fatty acids (FAs) belonging to ω-3 series. Fatty acid compositions of glycolipids differ from species to species and depend on conditions of microalgae growth. It is known that glycolipids of microalgae have an important nutritional significance and often possess by antitumoral, antimicrobial and anti-inflammatory properties [23].
It was shown that galactolipids of dinoflagellates were characterized by significant structural differences, depending upon the origin of chloroplasts of these microalgae. A probable general scenario of the evolution of galactolipids in chloroplasts of algae and other plants was discussed by Sato and Awai. They suggested that chloroplasts of plants, including algae, share common origin with cyanobacteria. Biosynthesis of MGDG is realized with participation of glycosyltransferases, encoded by genes MGD-1,2,3. They catalyze the transfer of β-galactosyl residue from uridine diphospho-galactose (UDP-Gal) to diacylglycerol (DAG). Conversion of MGDG into DGDG is catalyzed by epimerases, encoded by genes DGD-1,2. One more type of enzymes, desaturases, catalyze introduction of additional double bonds in acyl substituents of DAG. In contrast with cyanobacteria, the biosynthesis of MGDG from diacylglycerols in chloroplasts of plants takes place without the formation of intermediate glucosyldiacylglycerols [24].
Carbohydrate-containing metabolites of MGDG and DGDG groups are included into photosynthesizing complexes, stabilize them and can be exported into other subcellular compartments. Evolutionary transitions comprise primary endosymbiosis from a symbiotic cyanobacterium having a primary chloroplast and secondary endosymbiosis from a symbiotic unicellular algal eukaryote with a secondary plastid [25].
Structural determination of molecular forms of these lipids must include the positioning fatty acid residues, attached to glycerol, determination of stereochemical peculiarities such as configurations of asymmetric centers in glycon and aglycon moieties and nature in acyl substituents. The procedure is performed after separation of glycolipid mixtures, isolation and purification of sufficient amount of target substances followed by NMR analyses, supporting by mass-spectroscopy and chemical transformations.
Italian scientists applied simpler, based on 1H NMR spectroscopy approach to identify and quantify glycolipid components of the corresponding fractions of three microalgal species (Thalassiosira weissflogii, Cyclotella cryptica and Nannochloropsis salina). MGDG, SQDG, and DGDG lipids were found in each the studied species. For their structure analysis, areas and positions of signals in spectra, characteristic of various fragments of these algal glycolipids, were compared with calibrated proton NMR signals of an external standard [26]. This procedure, known as the ERETIC method [27], is applicable to glycolipid mixtures without their preliminary separation and may be used for characterization of glycolipids in them.
One of the first cases of the studies on macroalgal galactolipids aimed to determination of complete structures of their molecular forms was described by Yasumoto et al. [28], who isolated the 1-O-6,9,12,15-octadecatetraenoyl-3-O-[β-D-galactopyranosyl-(1→6)-O-β-D-galactopyranosyl]-sn-glycerol (17) along with an impurity of 1-O-3,6,9,12,15-octadecapentaenoyl-3-O-[β-D-galactopyranosyl-(1→6)-O-β-D-galactopy-ranosyl]-sn-glycerol from the microalga Prymnesium parvum as hemolytically active substance (so-called Hemolysin I) (Figure 4). They used chromatographic separation of galactolipid fraction, enzymatic hydrolysis of the obtained compounds and NMR studies to solve the problem of the structure determination of 17 belonging to the group of MGMGs.
α-Galactose was found in microalgal galactolipids in results of the studies of glycolipids from the microalga Heterosigma akashiwo by the group of Kitagawa in the beginning of the eighties [29]. This microalga, whose species name “akashiwo” to be from the Japanese “red tide", a species of Raphidophyceae, which episodically is bloomi in Japanese inner sea waters. Galactolipids from the H. akashiwo were shown to consist of several molecular forms, all bearing polyunsaturated fatty acids of ω-3 (n-3) series as acyl substituents. Cultured dinoflagellates were sonicated and extracted. Extract was subjected to silica gel column chromatography to obtain a MGDG, DGDG, and SQDG fractions. Further separation of two first fractions using HPLC on a Zorbax ODS column in methanol-water mixtures gave Heterosigma-glycolipids I-IV (18–21) (Figure 4). The treatment with sodium methylate to obtain FA methyl esters allowed for determining FA compositions of these glycolipids. At that, the known glycerol galactoside was also isolated. All the obtained compounds were studied by NMR and MS methods and shown to contain in carbohydrate moieties containing β-D- or β-D- and α-D-galactose residues (in compounds 18–20 and 21, respectively).
To establish positions of two different FA residues in 18, enzymatic hydrolysis using lipase type XIII from Pseudomonas sp. was carried out. As result, monoacyl derivative (22) with a single acyl substituent at the position 2 was obtained. The compound 20 gave methyl 6,9,12,15-octadecatetraenoate after the treatment with NaOMe-MeOH. Thus, it became clear what the corresponding FA was located at the position 2 and, therefore, the total structure of 16. Similarly, the same product (22) was also obtained from Heterosigma-glycolipid II (19). Positions of acyl substituents in Heterosigma-glycolipid IV (21) were established in result of its transformation into monoacyl derivative (23) by enzymatic hydrolysis, that liberated methyl 5,8,11,14,17-eicosapentaenoate at the action of sodium methylate in MeOH [29].
Glycolipids are known as the both constituent of plastids and cytosolyc lipids. Taking into account that origin of plastids in microalgae is suggested to be connected with primary, secondary or tertiary symbioses, microalgal species originated by different symbiotic pathways are characterized by the difference in FA compositions.
The culture of dinoflagellate Heterocapsa circularisquama, the alga which caused mortalities of pearl oysters and other mollusks in Japanese waters contained new glycoconjugates (24–27) (Figure 5). Structures of these galactolipids including absolute configurations of asymmetric centers in sugars and in aglycon moieties were established using 1H NMR and MS methods. Presence and positions of different acyl substituents in
27 were confirmed by HR ESI MS. At the concentrations of 6.2 μg/mL or higher, these galactose-containing compounds showed cytolytic activity towards heart and gill cells of oyster [30].
A new galactolipid (28) and Hemolysin 1 (17) were isolated from a cultured marine dinoflagellate Amphidinium sp. collected off the coast-line of Spain (Figure 5). Their structures were determined by 2D-NMR spectral data and chemical degradation. Galactolipid 28 has an additional fatty acid substituent, attached to C-6 of a monosaccharide unit [31].
A MGMG (29), isolated from ethylacetate extract of cultured dinoflagellate Scrippsiella trochoidea (Figure 5), demonstrated cytotoxic activity against tumor cells P-388 and L-1210 at dose of 0.1 mg/mL [32].
Beside above-mentioned activities, immunomodulatory action is also characteristic of galactolipids from microalgae, as it was shown by the studies on this type compounds isolated from the dinoflagellate Karenia mikimotoi [33].
Galactolipids of dinoflagellates Gymnodinium mikimotoi and Gymnodinium sp. contained hemolytic mixtures of MGDG and DGDG with major ones having 18:5ω3 FA residues. These substances were separated and purified by preparative TLC on silica gel plates followed by analysis of their structures [34]. Beside common polyunsaturated C18 FAs in lipids from dinoflagellates, long chain fatty acids were also found in these microalgae [35,36]
Deblond and Chapman developed a convenient procedure for isolating galactolipids from lipid mixtures by column chromatography with a gradual increasing the polarity of the eluting systems. They obtained galactolipids from lipid extracts by elution from an UniSil silica column with acetone as eluent. The distribution of C28 and C18 fatty acids in dinoflagellates was studied, and it was demonstrated that 28:7(n-3) and 28:8(n-3) FA are obviously located in phospholipids, while 18:5(n-3) and 18:4(n-3) are constituents of galactolipids from chloroplasts. Very-long-chain fatty acids (C28) were probably biosynthesized in cytoplasm, while C18 FA galactolipids within chloroplasts [37].
A long time, the 18:5(n-3) fatty acid was considered as chemotaxonomic marker of Dinophyta, but later this fatty acid was found in other microalgal taxa such as Haptophyceae, Eustigmaphyceae [38,39], and Raphidophyceae [40,41].
Structure analysis of galactolipids was significantly facilitated by wide application of HPLC [42,43], and particularly HPLC/MS methods [44] as well as by the development of the latter approach into HPLC/ESI MS/MS analysis. This technique was repeatedly used and allowed the rapid identification of related structures in different biological objects. In 2003, Guella et al. used this approach to successfully establish positions of acyl substituents in galactolipids. Actually, fragmentation in ESI MS with collision-induced dissociation ionization of individual components from MGDG and DGDG mixtures, previously separated by high-performance HPLC, was demonstrated by tandem positive-mode ESI MS at comparison with ESI MS of sn-2 lysoglyceroglycolipids, obtained by regiospecific enzymatic hydrolysis of the corresponding diacylglycerols using Rhizopus arrhizus lipase. Positive mode MS2 of [M+H]+ ions in mass spectra of asymmetrically disubstituted galactolipids indicated the preferred loss of acyl substituent from the sn-1 position as it illustrated below (Figure 6). They have formulated the next rule for determination of positions of acyl substituents in these galactolipids: “the positional distribution of the acyl chains in galactolipids can be established knowing that, in positive-ion mode ESI MS2 measurements, the loss of the carboxylic acid linked to the sn-1 glycerol position always produces a more intense peak than that derived from the loss of the sn-2 linked acyl chain”. This rule is applicable to the both MGDGs and DGDGs [45].
Many dinoflagellates contain carotenoid pigment peridinin, the principal constituent of water-soluble light-harvesting peridinin-chlorophyll-protein complex in chloroplasts. The peridinin was first isolated over 100 years ago. Application of ESI MS to analyses of MGDGs and DGDGs in 35 peridinin-containing species from the class Dinophyceae as well as the using tandem mass-spectrometry allowed determining the positional distribution of FAs associated with different classes of galactolipids. The examined dinoflagellates were divided into 2 clusters based on the forms of MGDGs and DGDGs presented. The first cluster possessed 18:5/18:4 MGDG (sn-1/sn-2), 18:5/18:5 MGDG, 18:4/18:4 DGDG, and 18:5/18:4 DGDG as major forms, while the microalgae belonging to the second cluster contained 20:5/18:4 and 20:5/18:5 MGDG, 20:5/18:4 and 20:5/18:5 DGDG as major forms. The majority of peridinin-containing dinoflagellates have secondary plastids, presumably of red algal origin [46].
No other microalgae possess so much diversity of their galactolipids as dinoflagellates. During their evolution, some dinoflagellates lost ancestral peridinin-containing plastids several times, but after it gained new endosymbionts and restored the capability to photosynthesis as a part of tertiary endosymbiotic events [47,48]. Then, microalgae adapted biochemical pathways retained from the ancestral plastid for transcript processing in their current plastids. It was suggested that as result of the event the genera Karenia and Karlodinium possess plastids of haptophyte origin; Lepidodinium (formerly Gymnodinium) chlorophorum possess plastids of green algal, possibly prasinophyte, origin; Kryptoperidinium (formerly Peridinium) foliaceum has endosymbiont plastids of pennate diatom origin. Glycoconjugates of these species with aberrant plastids were studied by ESI MS and ESI MS/MS. L. chlorophorum and K. brevis contained 18:5/18:5 MGDG, like it was observed in several peridinin-containing dinoflagellates, along with other forms of MGDG and DGDG, previously not found these microalgae. L. chlorophorum was found to possess 18:5/16:4 MGDG and 20:5/16:4 DGDG, and in K. brevis molecular forms 18:5/14:0 MGDG and DGDG were indicated. For comparison, the corresponding compounds from the green microalgae Tetraselmis sp., the haptophyte Emiliania huxleyi and the diatom Navicula perminuta, which were considered as those having common ancestors with those of aberrant dinoflagellates, were also examined by the same methods. It was shown that MGDG and DGDG compositions of the K. foliaceum/N. perminuta pair were almost the same, whereas in the L. chlorophorum/Tetraselmis sp. and K. brevis/E. huxleyi pairs, the MGDG and DGDG compositions were similar, but not in all galactolipid components matched. In general, these data confirmed the hypothesis concerning evolution of plastids and plastid glycolipids in some microalgae as consequence of tertiary symbiosis phenomena with participation of haptophyte, green, and diatom microalgae, respectively [49].
Recent analysis of galactolipid compositions of the microalgae belonging to the genus Amphidinium, one of the largest genera of Dinophyta, showed that its metabolites could be basal to those of a group of peridinin-containing dinoflagellates. This hypothesis was proposed by Leblond et al. [50].
It is of particular interest that symbiont dinoflagellates belonging to the genus Symbiodinium (so-called zooxanthellae) are the largest class of obligatory endosymbionts in marine invertebrates, first of all, in cnidarians such as corals, sea anemones, and jellyfish. Dinoflagellates Symbiodinium spp. were found also in some species of sponges, flatworms, mollusks, foraminifera and ciliates. The symbiosis of corals with zooxanthellae is an example of mutualistic symbiosis, at that, the both partners benefit. It is known that corals having zooxanthellae calcify and increase the photosynthetic fixation of CO2 much faster than those without dinoflagellates.
Awai et al. determined the characteristic features of microalgal MGDG, DGDG and SQDG in two Symbiodinium studied strains isolated from the jellyfish Cassiopea ornata and the giant clam Tridacna crocea studied [51]. It was shown that there is a transport of FAs from hosts to symbionts, which influence total lipid structures in symbiont dinoflagellates [52]. The 13 molecular species of MDGDs and 10 such species of DGDGs from the tropical soft coral Capnella sp. were studied by high-resolution tandem mass spectrometry in zooxanthellae) As result, a unique molecular species of MGDGs (16:4/18:5) and 18:4/18:4, 18:4/20:5, and 16:2/22:6 molecular species of DGDGs and indicated [53]. The main FAs in the zooxanthellae, symbiotic of a soft coral Sinularia sp. were identified as 18:4n-3, 20:5n-3, and 22:6n-3. Polar lipids with 18:4n-3, 18:5n-3, 20:5n-3 FAs were usually considered as the most characteristic of these symbionts [54].
Four peridinin-containing, cold-adapted dinoflagellates (Gymnodinium sp., Peridinium aciculiferum, Scrippsiella hangoei, and Woloszynskia halophila), grown at 4°C, were studied for presence of chloroplastic MGDG and DGDG by the positive-ion ESI MS and ESI MS/MS. Dominant forms possessed C18 fatty acids with the exception of the 20:5/18:5 form of DGDG in Gymnodinium sp. from the Baltic Sea. Each cold-adapted dinoflagellate possessed both 18:5/18:5 and 18:5/ 18:4 DGDG, while the majority of warm-adapted dinoflagellates contained only 18:5/18:4 DGDG. The presence of the 18:1/14:0 trigalactosyldiacylglycerol (TGDG) was also established as the dominant galactolipid in Gymnodinium sp. Probably, this metabolite contains an additional D-galactopyranosyl unit, linked by α-(1→6) bond to the terminal galactose of DGDG [55].
Only a few dinoflagellates and other microcroscopic algae possess proteinaceous, carotenoid-rich eyespots. Leblond and collaborators have established that the presence of an eyespot is not associated with galactolipid compositions of such microalgae [56].
Diatoms (the phylum Bacillariophyta) along with dinoflagellates, is the largest taxa of primary producers in the oceans. In majority, they have silica cell walls, called frustules. There are two classes of diatoms, differing each other in the forms of their frustules: 1) radially symmetrical (centric) and 2) younger bilaterally symmetrical pennate diatoms. Centric diatoms are predominantly pelagic, while pennate ones are frequently benthic species.
Beginning from 1993, the structural studies on galactolipids from diatoms were started. The diatom Phaeodactylum tricornutum, which can exist as different morphotypes and change cell shape, predominately has the same classes of galactolipids as other microalgae: MGDG, DGDG and, in smaller amount, SQDG. This species contains EPA (20:5n-3) at the sn-1 position and C16:1, C16:3, and C16:4 FAs at the sn-2 position in these compounds [57].
The similar distribution was established in relation of many other studied diatom species. FA compositions of diatoms were examined in many papers, at that majority of scientists noted the high content of galactolipids in total lipid fractions. For example, Dunstan et al. [58] examined FA compositions of the genera Skeletonema, Thalassiosira, Navicula, and Haslea, totally of 14 diatom species. As a rule, the diatoms possessed larger amounts of C14:0, C16:0, C16:1, and C20:5 fatty acids when compared with dinoflagellates. The major fatty acids in the most species were identified as 14:0, 16:0, 16:1 (n-7) and 20:5(n-3). Zhukova and Aizdaicher [59] have concluded that markers of Bacillariophyceae are 16:1(n-7) over 16:0, high levels of 14:0, 20:5(n-3) FAs, at that C16 polyunsaturated FAs contain double bonds at (n-4) and (n-1) positions.
Using ultra performance liquid chromatography-electrospray ionization-quadrupole-time of flight-mass spectrometry, Yan et al. [60] examined MGDGs and DGDGs in three strain of the pennate diatom Skeletonema. The predominant species of MGDGs were identified as containing 16:3, 20:5, 16:1, and 16:3 FA. Three main DGDG were in 20:5/16:1(sn-1/sn-2), 20:5/16:2, and 16:1/16:1 molecular forms. The major molecular forms of SQDG were indicated as containing the 14:0/16:0, 14:0/16:1, and 14:0/16:3 variants. Based on the identification of FA residues in sn-2 position, it was proposed that MGDGs and DGDGs are biosynthesized within chloroplasts by prokaryotic pathway exclusively, whereas SQDGs have a mixed biogenetic origin.
Most diatoms produce a set of photosynthetic pigments, including green chlorophylls along with yellow or brown carotenoids, that provides them a green or golden-brown color. The ‘blue’ pennate diatom, Haslea ostrearia synthesizes and releases into environment the water soluble, polyphenolic, non-photosynthetic pigment marennine. At their studies on centric and pennate diatoms H. ostrearia, P. tricornutum, Skeletonema marinoi, Navicula perminuta, and Thalassiosira weissflogii Dodson et al. [61] focused on the marennine-producing pennate diatom H. ostrearia. Application of ESI MS for analysis of their MGDG and DGDG structures indicated this microalga contains primarily C18/C16 or C18/C18 forms of MGDG and DGDG on the contrary with S. marinoi, T. weissflogii, and P. tricornutum, having C20/C16 and C18/C16 molecular forms of MGDGs and DGDG.s
Raphidophyte algae (Raphidophyceae) include brown- and green-pigmented taxa. Compositions and positional distribution of FA in MGDG and DGDG were examined using ESI MS and ESI MS/MS in the positive mode. Brown-pigmented strains of the genera Chattonella, Fibrocapsa, and Heterosigma primarily possessed 20:5/18:4 MGDG and 20:5/18:4 DGDG, while the green-pigmented Gonyostomum semen had these forms along with 18:3/18:4 MGDG and DGDG, characteristic of green algae [62].
The microscopic red alga Cyanidioschyzon merolae from an Italian hot spring has extremely simple FA composition with only C16:0, C18:0, and C18:1 n-9 as major acids in their galactolipids. Interestingly, the survival of microalgae in extreme conditions of a hot spring was associated with the loss of desaturase activity in plastids and, as consequence, the presence mainly saturated FAs in galactolipids [63].
Thus, galactolipids differing from each other in the both carbohydrate fragments and FA compositions, were indicated in all the studied species of microalgae, belonging to various phyla classes. In many cases, these carbohydrate-containing metabolites contain residues of polyunsaturated FAs, often attached to sn-1 position in glycerol moiety. Being mandatory components of the photosynthetic apparatus, galactolipids serve as a reservoir of essential polyunsaturated fatty acids, necessary for human health and exhibit diverse biological activities that open up prospects for their use in medicine.
3.2. Sulfoquinovosyl-containing glycolipids
Along with galactolipids, sulfur-containing sulfoquinovosyldiacylglycerols (SQDGs) are also known as essential metabolites, localized in thylakoid membranes of plastids in many photosynthetic organisms, including microalgae. These metabolites are formed from uridine diphosphate glucose in two stages: 1) by formation of uridine diphosphate sulfoquinovose, catalysed by UDP-sulfoquinovose synthase, 2) by the subsequent reaction with diacylglycerols, catalyzed by the glycosyltransferase SQDD-synthases [64]. Chemical diversity of SQDGs is determined mainly by FAs, linked to glycerol. The distinctive feature of SQDGs consists in the presence of high polar sulfonic acid residue bonded with C-6 in quinovose residue and frequently by saturated or monounsaturated FA acyl groups.
Blooms of the chloromonad microalga Heterosigma carterae (formerly H. akashiwo) (Raphidophyceae) were observed in different areas of the World Ocean. The isolate 102R of this microalga was cultured in natural seawater, and four SQDGs (30–33) from the extract of the culture were purified and structurally identified using LC/MS/MS technique and NMR analysis (Figure 7). SQDG fractions liberated the corresponding methyl esters of FAs at the reaction with BF3 in methanol and were analyzed by GC/MS technique. Positions of acyl substituents were determined by sn-1 regioselective enzymatic cleavage of SQDGs using lipase type XI from Rhizopus arrhizus, followed by isolation and analyses of products of enzymolysis. Exact double-bond positions in acyl substituents of 30–33 were established by gas chromatography/electron impact mass spectra of nicotinate derivatives prepared from the corresponding fatty alcohols, obtained after LiAlH4 treatment of these glycolipids.
SQDGs are difficult for isolation due to their high polarity. Similar compounds, isolated from other organisms, demonstrated anti-viral activity against HIV and antitumor properties [66].
The heterotropic dinoflagellates Oxyrrhis marina and Gymnodinium dominus are efficient producers of long-chain polyenic eicosapentaenic (20:5n-3) and docosahexaenic (22:6n-3) FAs, at that the latter is a major component of O. marina. O. marina was cultured and used to isolate a new sulfoquinovosyl-containing glycolipid, the first SQDG with highly unsaturated FA residue (Figure 7). The containing residue of ω-3 docosahexaenic FA glycolipid 34 was structurally elucidated by MS and NMR analysis, including 2D NMR experiments. It demonstrated a significant nitric oxide inhibitory activity in lipopolysaccharide-induced macrophage RAW 264.7 cells [67].
Regiochemical assignment in sufoquinovosyl diacylglycerols, like that of galactolipids, can be established using tandem mass-spectrometry, but in negative mode with collision-induced dissociation ionization. Actually, ion peaks corresponding the loss of fatty acid from sn-1 position of the studied sulfoglycolipids in these spectra are more intense in comparison with those resulted from the loss of neutral FA from sn-2 position [68].
The sulfolipid mixture with 35 as the main component was isolated by LH-20 chromatography from the methanol extract of the diatom Thalassiosira weissflogii with subsequent radial silica chromatography on Cromatotron and analyzed by NMR and LC/MS/MS. Using 35 and accompanying glycolipids as model compounds, a novel immunomodulatory compound, named β-SQDG18 (36) (Figure 7), was synthesized and proved to activate human dendritic cells (hDCs) by a TLR2/TLR4-independent mechanism. This agent stimulated the maturation of DCs and triggered immune response in vivo by the upregulation of MHC II, co-stimulatory proteins CD83, CD86, and pro-inflammatory cytokines L-12 and INF-γ. In experimental melanoma model, vaccination of C57BL/6 mice with β-SQDG18-adjuvanted hydrolyzed gelatin peptide (hgp10 peptide) induced protective response with a reduction in tumor growth and an increase in survival of animals. These studies have opened a new class of adjuvants based on a single glycolipid molecule [69].
3.3. Deacylated glycolipids
The green microalga Desmodesmus subspicatus biomass suspensions stimulated tomato (Solanum lycopersicum) seedlings via a foliar spray. A subfraction, obtained from the aqueous extract of this microalgae by chromatography on a Bio-Gel P-2 column, eluted with ultrapure water contained deacylated glycolipids 6-sulfo-α-D-quinovopyranosyl-(1→1)-glycerol (37), α-D-galactopyranosyl-(1→6)- -β-D-galactopyranosyl-(1→1)-glycerol (38), and β-D-galactopyranosyl-(1→1)-glycerol (39) along with the known zeatin (Figure 8). Tested concentrations of this subfraction induced increased hypocotyl lengths and volumes of seedlings, when compared with standard treatment (water), and provided the same effect as a commercial product derived from the brown alga Ascophyllum nodosum. At least partly, these compounds are probably responsible for plant growth biostimulatory action and have a prospect to be used in the so-called organic agriculture [70].
4. Phosphoglycolipids
The microalga Thalassiosira weissflogii (CCMP 1336) was selected to search for immunomodulatory natural products on the basis of a screening of marine extracts using human peripheral blood mononuclear cells (PBMC). Extracts of the microalga induced IL-6 cytokine production in PBMC. A novel minor phosphogalactodiacyl glycerol (40), named PGDG-1 and containing phosphate group, galactopyranose and two diacylated glycerol residues, was isolated from the methanol extract of the cultured strain of T. weissflogii by a bioassay guided fractionation using LH-20 gel chromatography followed by HPLC. Its structure was determined by NMR and HR ESI MS at comparison with those of a model compound PGDG-2 (41), obtained by the 15-step synthesis from the commercial D-galactose. PGDG-1 (40, Figure 9) exhibited immunostimulatory activity in human dendritic cells. The compound 41 showed Toll-like receptor-4 agonistic activity in human and murine dendritic cells and caused the antigen-specific T-cell activation [71].
The PGDG-1 was tested on monocyte-derived dendritic cells (moDCs) at the concentrations of 10–50 µg/mL and induced upregulation of surface markers at the lowest concentrations. When this substance was examined on human moDCs, their maturation was stimulated. The mRNA overexpression of interleukins IL-6, IL-8, and IL-12p40 was observed beginning from the concentration of 5 µg/mL. The synthetic compound (41) showed more remarkable effect on the upregulation of characteristic moDCs markers and the co-stimulatory molecules CD83 and CD86. A scheme of possible biogenetic origin of PGDG from phosphatidyldiacylglycerol (42) and galactosyldiacylglycerol (43) by reaction of transphosphatidylation was proposed and given below (Figure 10) [71].
A new platform was elaborated for selection of low molecular weight natural products with anticancer immunotherapeutic activity. This platform used immunophenotypic assay on growth factor–dependent immature DC line, derived from mouse spleen, and direct antitumor effects against 9 tumor cell lines. After it, the extracts, which induced the maturation and do not show nonspecific cytotoxicity, we studied on human DCs, generated in vitro from peripheral blood CD14+ monocytes [72]. To assess the selectivity of this screening methodology, extracts of the marine diatom Thalassiosira weissflogii containing α-sulfoquinovosyl diacylglycerols (α-SQDGs) and atypical phosphoglycolipids (PGDGs) were subjected to further investigation. Some obtained results were discussed above, and others are under progress.
5. Glycosides from microalgae
5.1. Steryl glycosides and other glycosylated derivatives from microalgae
Free sterols, important membrane constituents of eukaryotic organisms, for example microalgae, rare in prokaryotes are well studied. Sterol conjugates with carbohydrates, namely steryl glycosides, belong to one of two structural types: glucosides of sterols, predominantly of Δ5 series, and their acylated-by-fatty-acids derivatives (45, 46) (Figure 11). In the latter type, acyl group occupies the C-6 position in a sugar moiety. These glucose-containing metabolites are frequently, if not always, accompanied by other conjugated forms, such as steryl sulfates, and diverse bioactive oxidized sterol derivatives [73]. Steryl glucosides modify biological membranes and provide resistance against temperature stresses, regulate the host defense against pathogens, lipid metabolism, and development of their producers. Cholesteryl acyl glucosides (46) stimulated lymphocytes in a CD1-dependent manner and after injection into newborn animals protects them from allergy [74].
In continuation of our review [4], herein we report data concerning steryl glycosides that appeared in literature after 2018. Recently, it was shown that steryl glycosides, recovered from biodiesel tanks after the use of higher plant materials for fermentation are excellent sources of valuable plant sterols [75]. Taking into attention that the third generation of biodiesel production is based on the use of microalgae, it is of interest to examine whether precipitates forming at the fermentations of microalgal oils contain steryl glycosides also.
In the last time, modern methods of structural biology and biotechnology were used to elaborate approaches to the production of other glycosides, namely rhamnolipids by microalgae. Rhamnolipids are metabolites of some bacteria, used for bioremediation, enhancing oil recovery and as biodegradable emulsifiers, foaming agents, detergents and cleaners in other fields. Such surface-active agents as rhamnosides of 3-hydroxyacylacids may contain one or several rhamnoside residues. The green microalga Chlamydomonas reinhardtii, approved by US Food and Drug Administration as a food additive, was selected as a platform for the synthesis of 3-(hydroxydecanoyloxy)-decanoic acid HAA (47), biosynthetic precursor of biosurfactant mono-rhamnolipid (48) (Figure 12). Genes, encoding RhIA acyltransferase from the bacterium Pseudomonas aeruginosa, which catalyze the condensation of two 3-hydroxyacyl acids intermediates were used at chloroplast engineering of C. reihardtii to produce HAA lipid moiety of biosurfactants [76]. In the recent time, microalgae more and more often are considered as promising platforms for the biological synthesis of valuable metabolites not peculiar to these microorganisms themselves.
5.2. Glycosylated long chain polyketide derivatives
Studies of microalgae have rarely resulted in isolation of glycosides, in contrast to higher plants. However, unique glycosylated polyketide derivatives have been found in several microalgae species in addition to the well known steryl glucosides and cerebrosides. For example, amphidinins D and F (49, 50), belonging to the structural type of the so-called amphidinins, which are 4,5-secoanalogues of the previously known 12-membered macrolide amphidinolide Q, were obtained from a culture of the dinoflagellate Amphidinium sp. isolated from the marine acoel flatworm Amphiscolops sp. collected off Ishigaki Island, Okinawa, Japan (Figure 13). The carbon skeleton of the aglycones of these polyketides may be biosynthesized using a unique biochemical machinery for a long chain extension by one carbon atom. These compounds contain α-D-ribofuranose residues in the carbohydrate moiety and exhibit the moderate antimicrobial and antifungal properties, inhibiting the fungus Trichophyton mentagrophytes at concentrations of 16–32 μg/mL [77].
Prymnesium parvum, the golden (due to the fucoxanthin found in chloroplast) microalga belonging to the mixotrophic subphylum Haptophyta, is one of the most known species causing significant economic losses in fishery. P. parvum produces glycosylated long chain polyether toxins called prymnesins. These ichthyotoxins, due to their hemolytic action, cause massive fish kills in coastal, particularly brackish waters. They were isolated and the corresponding structures determined a long time after the toxicity of P. parvum was first detected. In fact, the studies on toxic properties of this microalga were first reported in the Netherland in 1920 [78] (cited from the recent review article [79]). Therefore, the studies on toxic P. parvum have been ongoing for 100 years. More than seventy years after this discovery, Japanese scientists isolated and determined structures of the first purified toxins of this type, prymnesins-1 and -2 [80,81], that was a result of the extremely difficult isolation and purification as well as determination of very complex structures of these toxins. The microalga was cultured and about 400 L of the culture was used to isolate 10 mg of prymnesin-1 (51) (PRM-1) and 15 mg of prymnesin-2 (52) (PRM-2) from the corresponding extract (Figure 14). However, the poor solubility of the obtained toxins made difficult the application of NMR spectroscopy for structural studies of these compounds. To improve the solubility, prymnesins were converted into N-acetates by the reaction with Ac2O in i-PrOH-water mixture (3:2). Moreover, the 13C-enriched toxins were also obtained by culturing this microalga in the medium containing Na213CO3 to facilitate using 13C NMR spectroscopy. Peracetylation, hydrogenation and dehalogenation were applied to obtain the corresponding derivatives and determine the number of free hydroxy groups, multiple bonds, and other structure peculiarities of prymnesins. COSY, DGF-COSY, and TOCSY and other NMR spectra revealed several partial structures. Their connection with each other in total structure of prymnesin-2 was established by HMBC experiments.
There were indicated also conjugated double and triple double bonds, chlorine and nitrogen atoms in aglycon moiety of these molecules. Configurations of C14 and C76−C85 asymmetric centers remained unknown. Examination of the NMR data of N-acetylprymnesins as well as analysis of their carbohydrate portions with wide application of NMR spectra, comparison with literature data, and other approaches indicated that PRM-1 contains α-D-ribofuranosyl, α-L-arabinopyranosyl, and β-D-galactofuranosyl residues at C77, C78, and C82 positions, respectively. Another toxin, PRN-1, contains only one, but a rare monosaccharide, α-L-xylofuranose, attached to C77.
Structure elucidation of polyether long-chain prymnesins-1 and -2 was the very impressive achievement in the marine bioorganic chemistry of the nineties. After partial synthesis, structures of prymnesins have been refined and reassignment of the relative configurations in the E/F ring juncture was reported [82,83]. Later, close related prymnesins began to be classified as A and B-types, respectively.
Recently, Rasmussen et al. [84] used LC/HR MS at the search for prymnesins in 10 strains of P. parvum, collected worldwide and found that only 1 of the studied strains produced the original prymnesins-1 and -2 (51, 52), isolated earlier by Japanese scientists. In addition, they reported the isolation and structure determination of prymnesin B1 (53) (Figure 14). At this, the 90% 13C enriched initial materials were used for its isolation following by extensive analysis of 2D- and 3D-NMR spectra for structure determination. Prymnesins of this type lack one 1,6-dioxadecalin unit and have a short acyclic C2 linkage instead it when compared with the original prymnesins. The nature of a sugar unit was determined using NMR and GC/MS analysis data and was based on the acid-hydrolyzed 53. The sugar in the hydrolysate of 13C-enriched sample of the toxin was identified as D-galactopyranose by GC/MS detection as an oxime-TMS derivative at the comparison with sugar standards followed by derivatization with N-methyl-bis-trifluoroacetamide and analysis of the obtained trifluoroacetyl derivative by the chiral-phase GC/MS and long-range HSQC experiment. The precise position of this sugar at C71 in core structure and the α-stereochemistry of glycoside bond were confirmed by the deshielded 13C chemical shift (δ13C = 90.3 ppm) of the anomeric carbon and 3JH-1′,H-2′ = 4 Hz as well as by a NOE correlation between H-1′ and H71. Like prymnesin-2, this novel of B-type prymnesin was extremely toxic against an RTgill-W1 cell line inhibiting these cells in the low nanomolar concentrations.
Further research by LC/HR MS method of strains of P. parvum, collected worldwide, established that only 4 examined strains contained 51 and/or 52. Other prymnesin analogues that differ each other in their backbones, chlorination, and glycosylation patterns were tentatively detected by LC/MS/MS, and C-type prymnesins were found in 5 strains studied. C-type prymnesins (a new type of these toxins) were proved to contain a C83 backbone, but their full structural analysis was not completed so far. Thus, it was indicated that evolution of prymnesins has led to a diverse family of fish-killing toxins, having a significant ecological influence on coastal brackish waters [84].
It is of interest that each of individual isolates of P. parvum contains only one type of these toxins. Geographical mapping of prymnesin strains indicated a global distribution of each type of these toxins. Prymnesins may be considered as chemotaxonomic markers of different strain clusters.
Phylogenetic analyses, based on internal transcribed spacer (ITS) sequences has shown the monophyletic origin of all the prymnesins. Probably, the appearance of new species within the P. parvum species complexes in result of their evolution is under progress nowadays [85].
Prymnesins demonstrate potent hemolytic, cytotoxic, neurotoxic, and ichthyotoxic activities at nanomolar concentrations. Structurally, these ichthyotoxins, due their biphylic structures, are detergent-like those having both polar and non-polar ends. They interact with plasmatic membranes, damage predominantly gill-breathing marine organisms and induce ion cellular leakage and lysis, followed by mortalities of fish and mollusks. Environmental factors such as salinity, pH, ion availability, and growth phase influence the prymnesin accumulation and toxicity of P. parvum strains [86].
During red tides and after it, prymnesins affect different surrounding hydrobionts, including competitors and grazers. Prymnesins can paralyze or kill organisms, which could not be caught to capture them by P. parvum due to the lack of the corresponding biological means and/or their size of victims [87]. Using own toxins, P. parvum can kill and eat the predatory heterotrophic dinoflagellate Oxyrrhis marina. At the action of toxins, cells of O. marina lost normal shape, lysed and particulate materials, originated from destroyed O. marina were ingested by cells of P. parvum [88].
Details of biosynthesis of these glycosylated toxins remains to be not established, although a cDNA library was constructed from late log-phase cultures of P. parvum with 3,415 unique tentative unigenes [TUGs]) found. Many of these TUGs, including 12 of 50 most commonly encountered transcripts, encoded novel proteins, probably involved in synthesis and secretion of prymnesins [89]. In the very recent study, the transcriptomes of 9 strains of P. parvum strains were analyzed. Numerous genes of polyketide synthases (PKS I) were found in the studied strains. Eight consensus transcripts were present in all these 9 strains examined. The detailed analysis of PKSs in P. parvum is the next step towards better understanding of the biosynthesis of prymnesins [90].
Structure diversity of prymnesins suggests that evolution of the polyketide biosynthetic machinery in microalgae has led to the variety of prymnesin backbone structures. As it was suggested by previous investigations on the biosynthesis of algal polyketides in dinoflagellates, the formation of the backbone polyketide chains in microalgal toxins is realized each time by not only by simple extension of the growing chain with two carbons of intact acetate units, but also by condensation with the cleaved acetate and glycolate units [91].
Prymnesins are more effectively biosynthesized in answer for physiological stress, for example low salinity or high irradiation. At that, toxicity and expression of selected PKS genes were increased. It confirmed PKSs to play an important role in biosynthesis of prymnesins [92].
6. Viral regulation of microalgal blooms and glycoconjugates
Recently, much attention has been attracted to issues related to the regulation of the harmful microalgae blooming as a result of viral infections. Abundance of viruses as the most common entities in the world's oceans (ten billion per liter of seawater) exceeds that of bacteria. Thirty years ago, Japanese scientists reported that the blooming of the raphidophycean microalgae Heterosigma akashiwo stopped due to the accumulation of virus. In fact, in June 1993, 3 days before the end of the red tide in the Northern part of Hiroshima Bay, Japan, they discovered viral particles in microalgae cells and suggested that the viral mortality that occurred may play a role in the decay of the red tide. Later, it has been suggested that the strain-specific HaV virus (Heterosigma akashiwo virus), clone GSNOU-30 plays the role of a selector that increases the genetic diversity of host microalgae [93,94].
A boom in the studies on marine viruses, which regulate the dynamics of microalgal blooms is continuing more than two last decades. Viruses terminate blooms by destruction of microalgae via mechanisms resemble programmed cell death in metazoans with DNA fragmentation and activation of cysteine aspartate-proteases (caspases). A particular attention was attracted to regulation of blooms caused by the microalgae participating in global geochemical cycles of carbon, calcium, and sulfur as well as harmful microalgae [94]. For example, the viral termination was established in relation of the blooming coccolithophore Emiliania huxleyi [95,96,97], major producers of calcite coccoliths, causing a transport of carbon to sediments. This alga is a producer the third part of the total marine CaCO3 production, and in addition it releases dimethyl sulfide into atmosphere enhancing cloud formation, that is the significant factor of climate regulation.
The giant Emiliania viruses (EhVs) are surrounded by lipid envelopes, comprising of glycosphingolipids (GSLs) with lesser percentage of polar glycerolipids such as MGDG, DGDG, and SQDG. The alga is enriched by glycolipids, which made up 65% of its lipidome. Sphingolipids are a crucial factor for a successful infection, followed by the sudden crashes of E. huxleyi blooms [98]. Genome of the virus penetrates into the host and changes biosynthesis of GSLs. Viral serine palmitoyl transcriptase use as substrate C15-CoA instead of C16-CoA followed the formation of the shortened sphinganine base (C17 instead of C18) and finally after glucosylation and hydroxylation gave the glucoconjugate (54) (Figure 15) [99]. Moreover, sialo-GSLs with sialic acid instead of glucose such as in 55 are formed in the infected microalga. These glycosphingolipids induce lytic changes and cell death in E. huxleyi. Glycosphingolipids with sialic acid, namely 2-keto-3-deoxy-D-glycero-D-galacto-nononic acid as a headgroup, derived from a cluster of genes in virus genome, regulate viral infection in this alga. Being synthesized de novo in the alga during lytic infection, unusual GSLs induce the programmed cell death via metacaspase expression [100]. Generally, the bloom termination is a result of horizontal transfer of genes, encoded sphingolipid biosynthesis pathway between the virus and eukaryotic host [101].
Since viral infection involves membrane fusion between virus and host, it was postulated that specific membrane lipids may facilitate the attachment of virus. In fact, the E. huxleyi population which has a greatest content of sGSLs showed the highest rate of virus-induced mortality. A potential role of sGSLs is to regulate the sensitivity and resistance in the dynamic E. huxleyi-EhV system [102].
Recently, another large double stranded DNA virus (PpDNAV) was isolated from the site of a harmful bloom event of Prymnesium parvum in Norfolk, England. The virus was found in 5 of 15 studied tested strains of the microalga and lysed >95% of host cells in sensitive strains by 120 h post infection. Using phylogenetic clustering, it was shown the structure and probably action of this virus to be phylogenetically similar to those of other algal viruses belonging to the Megaviridae family [103].
6. Carbohydrate-containing metabolites of microalgae and deep-sea life.
A huge amount of microalgal galactolipids, the most common secondary metabolites in plants, enters the sea water and the bottom of oceans and is easily absorbed by bacteria and invertebrates that feed on suspended organic matter and detritus and live in deep sea environments. In fact, marine phytoplankton excretes photosynthate, mainly consisting of carbohydrate-containing metabolites, into sea water in the form of dissolved organic matter. 7-50% radioactivity was found in dissolved organic matter in the water after exposure of samples of lake or sea waters with added 14C-bicarbonate during 7-24 h. Production of extracellular 14C-labelled organic matter was found in different types of phytoplankton [104,105]. Dead organic matter (detritus), is accumulated in deep-water continuations of river flows, in underwater intermountain areas and on the slopes of seamounts, playing an important role as a resource and habitat for many deep sea species and increasing stability and persistence of biosystems with effects on their trophic structures and biodiversity [106].
Those geographic zones, where the supply of such substances (as well as of mineral salts or warm waters) are especially large, are oases of life in sparsely populated spaces of the ocean floor with depths of more than 200 m.
7. Conclusive remarks
Microalgae have attracted a lot of attention for the possibility of being used for the industrial production of biofuels and the obtaining components of healthy food. Carbohydrate-containing natural products from microalgae have become the focus of research on microalgae metabolites because the enhancing of attempts to expand knowledge about the structures, biological activity, biological functions and taxonimic distribution of glycolipids, as well as various glycosides, including glycosylated toxins and pollutants such as ribosylated arsenic compounds. Relevant studies are closely related to such phenomena as regulation of microalgae blooms and poisoning of humans, fish and invertebrates with toxins of blooming microalgae. Search for new natural products and studies on their useful properties, for example immunomodulatory and anticancer activities, present one of aims of the corresponding studies.
Microalgae are excellent model organisms to develop different techniques of modern omics glycobiotechnology[107], bioingeeniring and structural biology to open new frontiers in improving the corresponding strains of microalgae to obtain valuable products and discover new directions of biosynthesis and metabolism in these lower plants.
Along with bacteria, microalgae form bases of food chains, and their metabolites can be transferring as dietary constituents into organisms of higher trophic levels and to deep-water biological communities.
Author Contributions
Authors contributed equally to the work. V.A.S. writing and discussion of the review parts concerning isolation, structure determination and diversity of metabolites discussed, I.V.S. writing and discussion of the review parts concerning taxonomy and biological peculiarities of microalgae-producers and list of references. The both authors have read and agreed to the published version of the manuscript.
Funding
This review was carried out with the partially support of the Grant of the Ministry of Science and Higher Education, Russian Federation 13.1902.21.0012 (contract No 075-15-2020-796).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study were available on request from the corresponding authors and from open data bases.
Acknowledgments
Authors thank Dr. N.V. Ivanchina for the reading of manuscript and comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.A. Marine nitrogen: Phosphorus stoichiometry and the global N:P cycle. Biogeochem. 1997, 37, 237–252. [Google Scholar] [CrossRef]
- Stonik, V.; Stonik, I. Low-molecular weight metabolites from diatoms: structures, biological roles and biosynthesis. Mar. Drugs 2015, 13, 3672–3709. [Google Scholar] [CrossRef]
- Stonik, V.A.; Stonik, I.V. Sterol and sphingoid glycoconjugates from microalgae. Mar. Drugs 2018, 16, 514. [Google Scholar] [CrossRef] [PubMed]
- Sadoli, E. Investigation into the occurrence of arsenic in the organism of fish. Biochem. Z. 1928, 201, 323–331. [Google Scholar]
- Wang, Y.; Wang, S.; Xu, P.; Liu, C.; Wang, Y.; Zhang, C.; Ge, Y. Review of arsenic specification, toxicity and metabolism in microalgae. Rev. Environ. Sci. Biotechnol. 2015, 14, 427–451. [Google Scholar] [CrossRef]
- Nurchi, V.M.; Djordjevic, A.B.; Crisponi, G.; Alexander, J.; Bjørklund, G.; Aaseth, J. Arsenic toxicity: molecular targets and therapeutic agents. Biomolecules 2020, 10, 235. [Google Scholar] [CrossRef]
- Zaffiri, L.; Gardner, J.; Toledo-Pereyra, L.H. History of antibiotics. From salvarsan to cephalosporins. J. Investig. Surg. 2012, 25, 67–77. [Google Scholar] [CrossRef]
- Emadi, A.; Gore, S.D. Arsenic trioxide—an old drug rediscovered. Blood Rev. 2010, 24, 191–199. [Google Scholar] [CrossRef]
- Duncan, E.G.; Maher, W.A.; Foster, S.D.; Krikowa, F. Influence of culture regime on arsenic cycling by the marine phytoplankton Dunaliella tertiolecta and Thallassiosira pseudonana. Environ. Chem. 2013, 10, 99–101. [Google Scholar] [CrossRef]
- Foster, S.D.; Thomson, D.; Maher, W.A. Uptake and metabolism of arsenate by anexic cultures of the microalgae Dunaliella tertiolecta and Phaeodactylum tricornutum. Mar. Chem. 2008, 108, 172–183. [Google Scholar] [CrossRef]
- Zhang, S.-Y.; Sun, G.X.; Yin, X.X.; Rensing, C.; Zhu, Y.-G. Biomethylation and volatilization of arsenic by the marine microalgae Ostreococcus tauri. Chemosphere 2013, 93, 47–53. [Google Scholar] [CrossRef]
- Duncan, E.G.; Maher, W.A.; Foster, S.D.; Krikowa, F. The influence of arsenate and phosphate exposure on arsenic uptake, metabolism and species formation in the marine phytoplankton Dunaliella tertiolecta. Mar. Chem. 2013, 157, 78–85. [Google Scholar] [CrossRef]
- Duncan, E.G.; Mather, W.A.; Foster, S.D. Contribution of arsenic species in unicellular algae to cycling of arsenic in marine ecosystem. Environ. Sci. Technol. 2015, 49, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M.; Levitsky, D.O. Arsenolipids. Progress Lipid Res. 2004, 43, 403–448. [Google Scholar] [CrossRef]
- Xue, X.M.; Xiong, C.; Yoshinaga, M.; Rosen, B.; Zhu, Y.G. The enigma of environmental organoarsenicals: Insights and implications. Crit. Rev. Environ. Sci. Technol. 2022, 52, 3835–3862. [Google Scholar] [CrossRef]
- Glabonjat, R.A.; Raber, G.; Jensen, K.B.; Guttenberger, N.; Zangger, K.; Francesconi, K.A. A 2-O-methylriboside unknown outside the RNAs world contains arsenic. Angew. Chem., Int. Ed. 2017, 56, 11963–11965. [Google Scholar] [CrossRef]
- Glabonjat, R.A.; Raber, G.; Jensen, K.B.; Guttenberg, N.; Zangger, K.; Francesconi, K.A. Arsenolipid biosynthesis by the unicellular alga Dunalliella tertiolecta is influenced by As/P ratio in culture experiments. Metallomics 2018, 10, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Glabonjat, R.A.; Raber, G.; Jensen, K.B.; Ehgartner, J.; Francesconi, K.A. Quantification of arsenolipids in the certified reference material NMIJ 7405-a (Hijiki) using HPLC/mass spectrometry after chemical derivatization. Anal. Chem. 2018, 6, 10282–10287. [Google Scholar] [CrossRef]
- Arora, N.; Gulati, K.; Patel, A.; Pruthi, P.A.; Poluri, K.M.; Pruthi, V. A hybrid approach integrating arsenic detoxification with biodiesel production using oleaginous microalgae. Algal Res. 2017, 24, 29–39. [Google Scholar] [CrossRef]
- Hölzl, G.; Witt, S.; Kelly, A.A.; Heinz, E. Functional differences between galactolipids and glucolipids revealed in photosynthesis of higher plants. PNAS 2006, 103, 7512–7517. [Google Scholar] [CrossRef] [PubMed]
- Gounaris, K.; Barber, J. Monogalactosyldiacylglycerol: the most abundant polar lipid in nature. Trends Biochem. Sci. 1983, 8, 378–381. [Google Scholar] [CrossRef]
- Da Costa, E.; Silva, J.; Mendonça, S.H.; Abreu, M.H.; Domingues, M.R. Lipidomic approaches towards deciphering glycolipids from microalgae as a reservoir of bioactive lipids. Mar. Drugs 2016, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Awai, K. Diversity of biosynthetic pathways of galactolipids in the light of endosymbiotic origin of chloroplasts. Front. Plant Sci., Sec. Plant Metabolism Chemodiv. 2016, 7, 117. [Google Scholar] [CrossRef]
- Petroutsos, D.; Amiar, S.; Abida, H.; Dolch, L.J.; Bastien, O.; Rébeillé, F.; Jouhet, J.; Falconet, D.; Block, M.A.; McFadden, G.I.; Bowler, C.; Botte, C.; Maréchal, E. Evolution of galactoglycerolipid biosynthetic pathways - From cyanobacteria to primary plastids and from primary to secondary plastids. Progr. Lipid Res. 2014, 54, 68–85. [Google Scholar] [CrossRef]
- Nuzzo, G.; Gallo, C.; D’Ippolito, G.; Cutignano, A.; Sardo, A.; Fontana, A. Composition and quantification of microalgal lipids by ERITIC 1H NMR spectroscopy. Mar. Drugs 2013, 11, 3742–3753. [Google Scholar] [CrossRef]
- Akoka, S.; Barantin, L.; Trierweiler, M. Concentration measurement by proton NMR using the ERETIC method. Anal. Chem. 1999, 71, 2554–2557. [Google Scholar] [CrossRef]
- Kozaki, H.; Oshima, Y.; Yasumoto, T. lsolation and structural elucidation of Hemolysin I from the phytoflagellate Prymnesium parvum. Agric. Biol. Chem. 1982, 46, 233–236. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hayashi, K.; Kawazoe, K.; Kitagawa, I. Marine Natural Products. XXIX. Heterosigma-glycolipids I, II, III, and IV, four diacylglyceroglycolipids possessing ω3-polyunsaturated fatty acid residues, from the Raphidopycean dinoflagellate Heterosigma akashiwo. Chem. Pharm. Bull. 1992, 40, 1404–1410. [Google Scholar] [CrossRef]
- Hiraga, Y.; Shikano, T.; Widianti, T.; Ohkata, K. Three new glycolipids with cytolytic activity from cultured marine dinoflagellate Heterocapsa circularisquama. Nat. Prod. Res. 2008, 22, 649–657. [Google Scholar] [CrossRef]
- Daranas, A.H.; Fernández, J.J.; Norte, M. New monogalactosyl triacylglycerol from a cultured marine dinoflagellate Amphidinium sp. Nat. Prod. Lett. 1999, 14, 107–114. [Google Scholar] [CrossRef]
- Oshima, Y.; Yamada, S.-H.; Moriya, T.; Ohimuzu, Y. A monogalactosyl diacylglycerol from a culture marine dinoflagellate Scripsiella trocholidea. J. Nat. Prod., 1994, 57, 534–536. [Google Scholar] [CrossRef]
- Leutou, A.S.; McCall, J.R.; York, R.; Govindapur, R.R.; Bourdelais, A.J. Anti-inflammatory activity of glycolipids and a polyunsaturated fatty acid methyl ester isolated from the marine dinoflagellate Karenia mikimotoi. Mar. Drugs 2020, 18, 138. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.C.; Bodennec, G.; Gentien, P. Haemolytic glycoglycerolipids from Gymnodinium species. Phytochemistry 1998, 47, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Monsour, M.P.; Volkmam, J.K.; Jackson, A.E.; Blackburn, S.I. Very-long-chain (C28) highly unsaturated fatty acids in marine dinoflagellates. Phytochemistry 1999, 50, 541–548. [Google Scholar] [CrossRef]
- Volkman, J.K.; Barrett, S.M.; Blackburn, S.I.; Mansour, M.P.; Sikes, E.L.; Gelin, F. Microalgal biomarkers: a review of recent research developments. Org. Geochem. 1998, 29, 1168–1179. [Google Scholar] [CrossRef]
- Leblond, J.D.; Chapman, P.J. Lipid class distribution of highly unsaturated long chain fatty acids in marine dinoflagellates. J. Phycol. 2000, 36, 1103–1198. [Google Scholar] [CrossRef]
- Renaud, S.M.; Parry, D.L.; Thinh, L.V.; Kuo, C.; Padovan, A.; Sammy, N. Effect of light intensity on the proximate biochemical and fatty acid composition of Isochrysis sp. and Nannochloropsis oculata for use in tropical aquaculture. J. Appl. Phycol. 1991, 3, 43–53. [Google Scholar] [CrossRef]
- Okyama, H.; Morita, N.; Kogame, K. Occurrence of octadecapentaenoic acid in lipids of a cold stenotermic alga prymnesiophyte strain B. J. Phycol. 1992, 25, 465–472. [Google Scholar] [CrossRef]
- Nichols, P.D.; Volkman, J.K.; Hallegraeff, G.M.; Blackburn, S.I. Sterols and fatty acids of the red tide flagellates Heterosigma akashiwo and Chattonella antiqua (Raphidophyceae). Phytochemistry 1987, 26, 2537–2541. [Google Scholar] [CrossRef]
- Mostaert, A.S.; Karsten, U.; Hara, Y.; Watanabe, M.M. Pigments and fatty acids of marine raphidophytes: a chemotaxonomic re-evaluation. Phycol. Res. 1998, 46, 215–220. [Google Scholar] [CrossRef]
- Lynch, D.V.; Gundersen, R.E.; Thompson, G.A. Jr. Separation of galactolipids molecular species by high-performance liquid chromatography. Plant Physiol. 1983, 72, 903–905. [Google Scholar] [CrossRef]
- Yongmanitchai, W.; Ward, O.P. Positional distribution of fatty acids, and molecular species of polar lipids, in the diatom Phaeodactylum tricornutum. Microbiology, 1993, 139, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.-H.; Hong, J.; Choe, J.C.; Kim, Y.-H. Analysis of fatty acyl groups of diacyl galactolipid molecular species by HPLC/ESI-MS with in-source fragmentation. Bull. Korean Chem. Soc. 2003, 24, 1163–1168. [Google Scholar] [CrossRef]
- Guella, G.; Frasssantio, R.; Mancini, I. A new solution for an old problem: the regiochemical distribution of the acyl chains in galactolipids can be established by electrospray ionization tandem mass spectrometry. Rapid Comm. Mass Spectrom. 2003, 17, 982–1984. [Google Scholar] [CrossRef]
- Gray, C.G.; Lasiter, A.D.; Li, C.; Leblond, J.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. I. Peridinin-containing taxa. Eur. J. Phycol. 2009, 44, 191–197. [Google Scholar] [CrossRef]
- Yoon, H.S.; Hackett, J.D.; Van Dolah, F.M.; Nosenko, T.; Lidie, K.L.; Bhattacharya, D. Tertiary endosymbiosis driven genome evolution in dinoflagellate algae. Mol. Biol. Evol. 2005, 22, 1299–1308. [Google Scholar] [CrossRef]
- Dorrell, R.G.; Howe, C.J. Integration of plastids with their hosts: Lessons learned from dinoflagellates. Proc. Natl Acad. Sci. USA 2015, 112, 10247–10254. [Google Scholar] [CrossRef]
- Leblond, J.D.; Andrew, D.; Lasiter, A.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. II. Lepidodinium chlorophorum, Karenia brevis, and Kryptoperidinium foliaceum, three dinoflagellates with aberrant plastids. Eur. J. Phycol. 2009, 44, 199–205. [Google Scholar] [CrossRef]
- Leblond, J.D.; Elkins, F.C.; Graeff, J.E.; Sabir, K. Galactolipids of the genus Amphidinium (Dinophyceae): an hypothesis that they are basal to those of other peridinin-containing dinoflagellates. Eur. J. Phycol. 2022, 58, 190–198. [Google Scholar] [CrossRef]
- Awai, K.; Matsuoka, R.; Shioi, Y. Lipid and fatty acid compositions of Symbiodinium strains. In Proceedings of the 12th international coral reef symposium. 2012, 9–12. [Google Scholar]
- Imbs, A.B.; Yakovleva, I.M.; Dautova, T.N.; Bui, L.H.; Jones, P. Diversity of fatty acid composition of symbiotic dinoflagellates in corals: evidence for the transfer of host PUFAs to the symbionts. Phytochemistry 2014, 101, 76–82. [Google Scholar] [CrossRef]
- Imbs, A.B.; Rybina, V.G.; Kharlamenko, V.I.; Dang, L.P.T.; Nguyen, N.T.; Pham, K.M.; Pham, L.Q. Polyunsaturated molecular species of galactolipids: Markers of zooxanthellae in a symbiotic association of the soft coral Capnella sp. (Anthozoa: Alcyonacea). Russ. J. Mar Biol. 2015, 41, 461–467. [Google Scholar] [CrossRef]
- Imbs, A.B.; Yakovleva, I.M.; Pham, L.Q. Distribution of lipids and fatty acids in the zooxanthellae and host of the soft coral Sinularia sp. Fish. Sci. 2010, 76, 375–380. [Google Scholar] [CrossRef]
- Gray, C.G.; Lasiter, A.D.; Leblond, J.D. Mono- and digalactosyldiacylglycerol composition of dinoflagellates. III. Four cold-adapted, peridinin-containing taxa and the presence of trigalactosyldiacylglycerol as an additional glycolipid. Eur. J. Phycol. 2009, 44, 439–445. [Google Scholar] [CrossRef]
- Graeff, J.E.; Elkins, L.C.; Leblond, J.D.; Graeff, J.E.; Elkins, L.C.; Leblond, J.D. Plastid-associated galactolipid composition in eyespot-containing dinoflagellates: a review. Algae 2021, 36, 73–90. [Google Scholar] [CrossRef]
- Yongmanitchai, W.; Ward, O.P. Positional distribution of fatty acids, and molecular species of polar lipids, in the diatom Phaeodactylum tricornutum. J. Gen. Microbiol. 1993, 139, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, G.A.; Volkman, J.K.; Barrett, S.M.; Leroi, J.M.; Jeffrey, S.W. Essential polyunsaturated fatty acids from 14 species of diatom (Bacillariophyceae). Phytochemistry 1994, 35, 155–161. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Aizdaicher, N.A. Fatty acid composition of 15 species of marine microalgae. Phytochemistry 1995, 39, 351–356. [Google Scholar] [CrossRef]
- Yan, X.; Chen, D.; Xu, J.; Zhou, C. Profiles of photosynthetic glycerolipids in three strains of Skeletonema determined by UPLC-Q-TOF-MS. J. Appl. Phycol. 2011, 23, 271–282. [Google Scholar] [CrossRef]
- Dodson, J.; Dahmen, J.L.; Mouget, J.L.; Leblond, J.D. Mono-and digalactosyldiacylglycerol composition of the marennine-producing diatom, Haslea ostrearia: Comparison to a selection of pennate and centric diatoms. Phycol. Res. 2013, 61, 199–207. [Google Scholar] [CrossRef]
- Roche, S.A.; Leblond, J.D. Mono- and digalactosyldiacylglycerols compositions of rhaphidophytes (Raphidophyceae): a modern interpretation using positive-ion/ mass-spectrometry/mass-spectrometry. Journal of phycology 2011, 47, 106–111. [Google Scholar] [CrossRef]
- Sato, N.; Moriyama, T. Genomic and biochemical analysis of lipid biosynthesis in the unicellular rhodophyte Cyanidioschyzon merolae: lack of a plastidic desaturation pathway results in the coupled pathway of galactolipid synthesis. Eukaryotic cell 2007, 6, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Benning, C. Biosynthesis and functions of the sulfolipid sulfoquinosyl diacylglycerol. Annual Rev. Plant Physiol. Plant Molecul. Biol. 1998, 49, 53–75. [Google Scholar] [CrossRef]
- Keusgen, M.; Curtis, J.M.; Thibault, P.; Walter, J.A.; Windust, A.; Ayer, S.W. Sulfoquinovosyl diacylglycerols from the alga Heterosigma carterae. Lipids 1997, 32, 1102–1112. [Google Scholar] [CrossRef]
- Gustafson, K.R.; Cardellina, J.H., II; Fuller, R.W.; Weislow, O.S.; Kiser, R.F.; Snader, K.M.; Patterson, G.M.L.; Boyd, M.R. AIDS-Antiviral Sulfolipids from Cyanobacteria (Blue-Green Algae). J. Natl. Cancer Inst. 1989, 81, 1254–1258. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.Y.; Yang, A.R.; Park, J.; Moon, S.J.; Jeong, E.J.; Rho, J.-R. Characterization of a new trioxilin and a sulfoquinovosyl diacylglycerol with anti-inflammatory properties from the dinoflagellate Oxyrrhis marina. Mar. Drugs, 2017, 15, 57. [Google Scholar] [CrossRef]
- Zianni, R.; Bianco, G.; Lelario, F.; Losito, I.; Palmisano, F.; Cataldi, T.R. Fatty acid neutral losses observed in tandem mass spectrometry with collision-induced dissociation allows regiochemical assignment of sulfoquinovosyl-diacylglycerols. J. Mass Spectr. 2013, 48, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Cutignano, A.; Pagano, D.; Gallo, C.; Barra, G.; Nuzzo, G.; Sansone, C.; Ianora, A.; Urbanek, K.; Fenoglio, D.; Ferrera, F.; Bernardi, C.; Parodi, A.; Pasqually, G.; Leonardi, A.; Filaci, G.; De Palma, R.; Fontana, A. A new marine-derived sulfoglycolipid triggers dendritic cell activation and immune adjuvant response. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Mazepa, E.; Malburg, B.V.; Mógor, G.; de Oliveira, A.C.; Amatussi, J.O.; Corrêa, D.O.; Lemos, J.S.; Diogo, R.B.; Ducatti, D.R.; Duarte, M.E.R.; Mogor, A.F.; Noseda, M.D. Plant growth biostimulant activity of the green microalga Desmodesmus subspicatus. Algal Res. 2021, 59, 102434. [Google Scholar] [CrossRef]
- Manzo, E.; Gallo, C.; Sartorius, R.; Nuzzo, G.; Sardo, A.; De Berardinis, P.; Fontana, A.; Cutignano, A. Immunostimulatory phosphatidylmonogalactosyldiacylglycerols (PGDG) from the Marine Diatom Thalassiosira weissflogii: Inspiration for a Novel Synthetic Toll-Like Receptor 4 Agonist. Mar. Drugs, 2019, 17, 103. [Google Scholar] [CrossRef] [PubMed]
- Gallo, C.; Barra, G.; Saponaro, M.; Manzo, E.; Fioretto, L.; Zico, M.; Nuzzo, G.; d’Ippolito, G.; De Palma, R.; Fontana, A. A new bioassay platform design for the discovery of small molecules with anticancer immunotherapeutic activity. Mar. drugs, 2020, 18, 604. [Google Scholar] [CrossRef]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev., 2001, 70, 673–715. [Google Scholar] [CrossRef]
- Shimamura, M. Structure, metabolism and biological functions of steryl glycosides in mammals. Archiv. Immunolog. Therap. 2012, 60, 351–359. [Google Scholar] [CrossRef]
- Carlucci, R.; Jäger, S.N.; Labadie, G.R. Steryl glucosides recovered from biodiesel tank deposits are an excellent source of phytosterols. Industrial Crops and Prod. 2022, 187, 115307. [Google Scholar] [CrossRef]
- Mario-Vynvyals, B.; Artigues, M.; Wostrikoff, K.; Monte, E.; Bruto-Puig, G. Chloroplast engineering of the green microalgae Chlamydomonas reinhardtii for the production of HAA the lipid moiety of rhamnolipid biosurfactants. New biotechn. 2023, 76, 1–12. [Google Scholar]
- Kubota, T.; Iwai, T.; Sakai, K.; Gonoi, T.; Kobayashi, J. Amphidinins C−F, Amphidinolide Q Analogues from marine dinoflagellate Amphidinium sp. Org. Lett. 2014, 16, 5624–5627. [Google Scholar] [CrossRef]
- Liebert, F.; Deerns, W.M. Onderzoek naar de oorzak van een Vischsterfte in den Polder Workumer Nieuwland, nabij Workum. Verhandungen en Rapporten uitgegeven door Rijkinstituten voor Visscherijonderzoek, 1920, 1, 81–93. [Google Scholar]
- Wagstaff, B.A.; Hems, E.S.; Rejzek, M.; Pratscher, J.; Brooks, E.; Kuhaudomlarp, S.K.; O’Neill, E.C.; Matthew, I.; Donaldson, M.I.; Lane, S.; Currie, J.; Hindes, A.M.; Malin, G.; Murrell, J.C.; Field, R.A. Insights into toxic Prymnesium parvum blooms: the role of sugars and algal viruses. Biochem. Soc. Transactions, 2018, 46, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, T.; Satake, M.; Yasumoto, T. Prymnesin-2: a potent ichthyotoxic and hemolytic glycoside isolated from the red tide alga Prymnesium parvum. J. Am. Chem. Soc. 1996, 118, 479–480. [Google Scholar] [CrossRef]
- Igarashi, T.; Satake, M.; Yasumoto, T. Structures and partial stereochemical assignments for prymnesin-1 and prymnesin-2: potent hemolytic and ichthyotoxic glycosides isolated from the red tide alga Prymnesium parvum. J. Am. Chem. Soc. 1999, 121, 37–8499. [Google Scholar] [CrossRef]
- Sasaki, M.; Shida, T.; Tachibana, K. Synthesis and stereochemical confirmation of the HI/JK ring system of prymnesins, potent hemolytic and ichthyotoxic glycoside toxins isolated from the red tide alga. Tetrahedron Lett. 2001, 42, 5725–5728. [Google Scholar] [CrossRef]
- Sasaki, M.; Ebine, M.; Takagi, H.; Takakura, H.; Shida, T.; Satake, M.; Oshima, Y.; Igarashi, T.; Yasumotao, T. Synthesis of the CDE/FG ring models of prymnesins: reassignment of the relative configuration of the E/F ring juncture. Org. Lett. 2004, 6, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.A.; Meier, S.; Andersen, N.G.; Blossom, H.E.; Duus, J.Q.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Chemodiversity of Ladder-Frame Prymnesin Polyethers in Prymnesium parvum. J. Nat. Prod. 2016, 79, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Binzer, S.B.; Svenssen, D.K.; Daugbjerg, N.; Alves-de-Souza, C.; Pinto, E.; Hansen, P.J.; Varga, E. A-, B-and C-type prymnesins are clade specific compounds and markers in Prymnesium parvum. Harmful Algae, 2019, 81, 10–17. [Google Scholar] [CrossRef]
- Manning, S.R.; La Claire II, J.W. Prymnesins: Toxic Metabolites of the Golden Alga, Prymnesium parvum Carter (Haptophyta). Mar. Drugs 2010, 8, 678–704. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U. Phagotrophy by a plastidic haptophyte, Prymnesium patelliferum. Aquat. Microb. Ecol. 1998, 14, 155–160. [Google Scholar] [CrossRef]
- Tillmann, U. Kill and eat your predator: A winning strategy of the planktonic flagellate Prymnesium parvum. Aquat. Microb. Ecol. 2003, 32, 73–84. [Google Scholar] [CrossRef]
- La Claire, J.W., II. Analysis of expressed sequence tags from the harmful alga, Prymnesium parvum (Prymnesiophyceae, Haptophyta). Mar. Biotech. 2006, 8, 534–546. [Google Scholar] [CrossRef]
- Anestis, K.; Kohli, G.; Wohlrab, S.S.; Varga, E.; Larsen, T.O.; Hansen, P.J.; Jon, U. Polyketide synthase genes and molecular trade-offs in the ichthyotoxic species Prymnesium parvum. Sci. Total Environ., 2021, 795, 148878. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Satake, M.; Wright, J.L. Polyketide biosynthesis in dinoflagellates: what makes it different? Nat. Prod. Rep. 2014, 31, 1101–1137. [Google Scholar] [CrossRef]
- Freitag, M.; Beszteri, S.; Vogel, H.; John, U. Effects of physiological shock treatments on toxicity and polyketide synthase gene expression in Prymnesium parvum (Prymnesiophyceae). Eur. J. Phycol., 2011, 46, 193–201. [Google Scholar] [CrossRef]
- Nagasaki, K.; Masashi, A.; Itakura, S.; Imai, I.; Ishida, Y. Viral mortality in the final stage of Heterosigma akashiwo (Raphidophyceae) red tide. J. Plankt. Res. 1994, 16, 1595–1599. [Google Scholar] [CrossRef]
- Nagasaki, K.; Yamaguchi, M. Isolation of a virus infectious to the harmful bloom causing microalga Heterosigma akashiwo (Raphidophyceae). Aquat. Microb. Ecol. 1997, 13, 135–140. [Google Scholar] [CrossRef]
- Fuhrman, J.A. Marine viruses and their biogeochemical and ecological effects. Nature 1999, 399, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, D.C.; Oke, J.; Malin, G.; Wilson, W.H. Coccolithovirus (Phycodnaviridae): Characterization of a new large dsDNA algal virus that infects Emiliana huxleyi. Arch. Virol. 2002, 147, 1685–1698 doiorg103390/v9030052. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; Tarran, G.A.; Schroeder, D.; Cox, M.; Oke, J.; Malin, G. Isolation of viruses responsible for the demise of an Emiliania huxleyi bloom in the English Channel. J. Mar. Biol. Assoc. UK, 2002, 82, 369–377. [Google Scholar] [CrossRef]
- Vardi, A.; Haramaty, L.; Van Mooy, B.A.S.; Fredricks, H.F.; Kimmance, S.; Larsen, A.; Bidle, K.D. Host-virus dynamics and subcellular controls of cell fate in a natural coccolithophore population. Proc. Natl. Acad. Sci. U S A, 2012, 109, 19327–19332. [Google Scholar] [CrossRef] [PubMed]
- Vardi, A.; Van Mooy, B.A.S.; Fredricks, H.F.; Popendorf, K.J.; Ossolinski, J.E.; Haramaty, L.; Bidle, K.D. Viral glycosphingolipids induce lytic infection and cell death in marine phytoplankton. Science 2009, 326, 861–865. [Google Scholar] [CrossRef]
- Pagarete, A.; Allen, M.J.; Wilson, W.H.; Kimmance, S.A.; de Vargas, C. Host–virus shift of the sphingolipid pathway along an Emiliania huxleyi bloom: survival of the fattest. Environ. Micobiol. 2009, 11, 2840–2848. [Google Scholar] [CrossRef]
- Wilson, W.H.; Schroeder, D.C.; Allen, M.J.; Holden, M.T.G.; Parkhill, J.; Barrell, B.G.; Ghazal, P. Complete genome sequence and lytic phase transcription profile of a Coccolithovirus. Science 2005, 309, 1090–1092. [Google Scholar] [CrossRef]
- Fulton, J.M.; Fredricks, H.F.; Bidle, K.D.; Bidle, K.D.; Vardi, A.; Kendrick, B.J.; DiTullio, G.R.; Van Mooy, B.A.S. Novel molecular determinants of viral susceptibility and resistance in the lipidome of Emiliania huxleyi. Environ. Microbiol. 2014, 16, 1137–1149. [Google Scholar] [CrossRef]
- Wagstaff, B.A.; Vladu, I.C.; Barclay, J.E.; Schroeder, D.C.; Malin, G.; Field, R.A. Isolation and Characterization of a Double Stranded DNA Megavirus Infecting the Toxin-Producing Haptophyte Prymnesium parvum. Viruses 2017, 9, 40. [Google Scholar] [CrossRef]
- Fogg, G.T.; Nalewajko, C.; Watt, W. Extracellular products of phytoplankton photosynthesis. Proc. Roy Soc London B 1965, 162, 517–534. [Google Scholar] [CrossRef]
- Fenchel, T. Marine phytoplankton food chains. Ann. Rev. Ecol. Syst. 1988, 19, 19–38. [Google Scholar] [CrossRef]
- Moore, J.C.; Berlow, E.L.; Coleman, D.C.; de Ruiter, P.C.; Dong, Q.; Hastings, A.; Johnson, N.C.; McCann, K.S.; Melville, K.; Morin, P.J.; Nadelhoffer, K.; Rosemond, A.D.; Post, D.M.; Sabo, J.L.; Scow, K.M.; Vanni, M.J.; Wall, D.H. Detritus, trophic dynamics and biodiversity. Ecol. letters 2004, 7, 584–600. [Google Scholar] [CrossRef]
- Sirohi, R.; Joun, J.; Choi, H.I.; Gaur, V.K.; Sim, S.J. Algal glycobiotechnology: omics approaches for strain improvement. Microbial Cell Fact 2021, 20, 163. [Google Scholar] [CrossRef]
Figure 1.
Main types of arsenosugar derivatives.
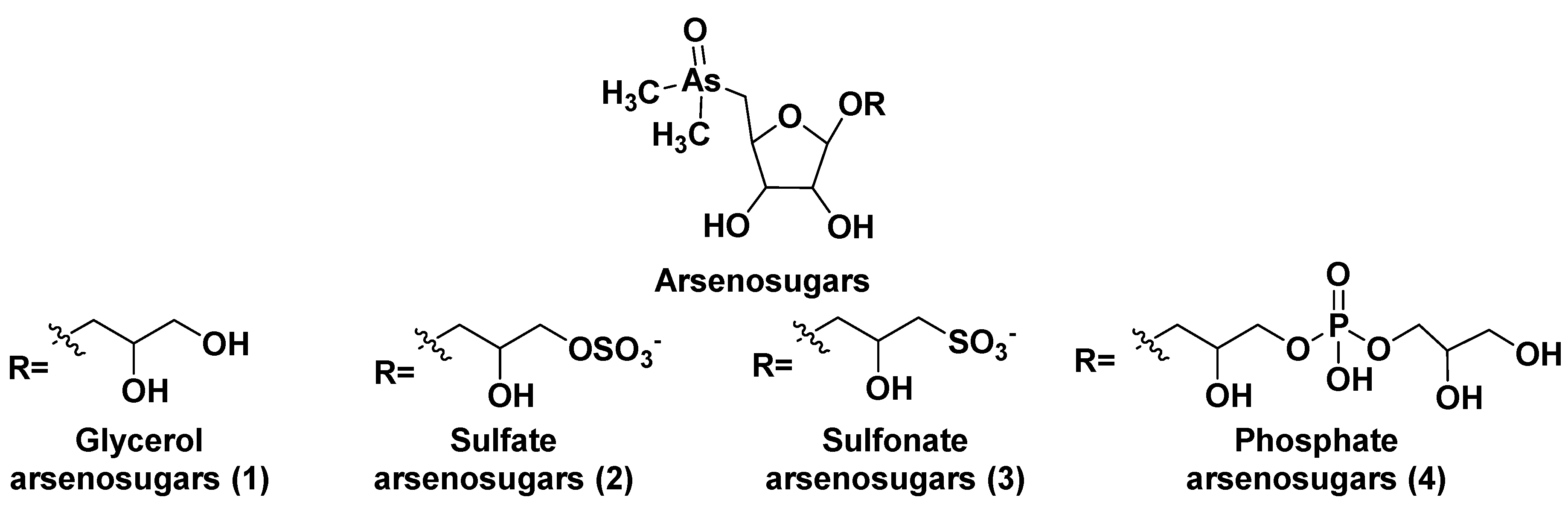
Figure 2.
Biosynthesis of 5 and structures of other arsenolipids found in D. tertiolecta.
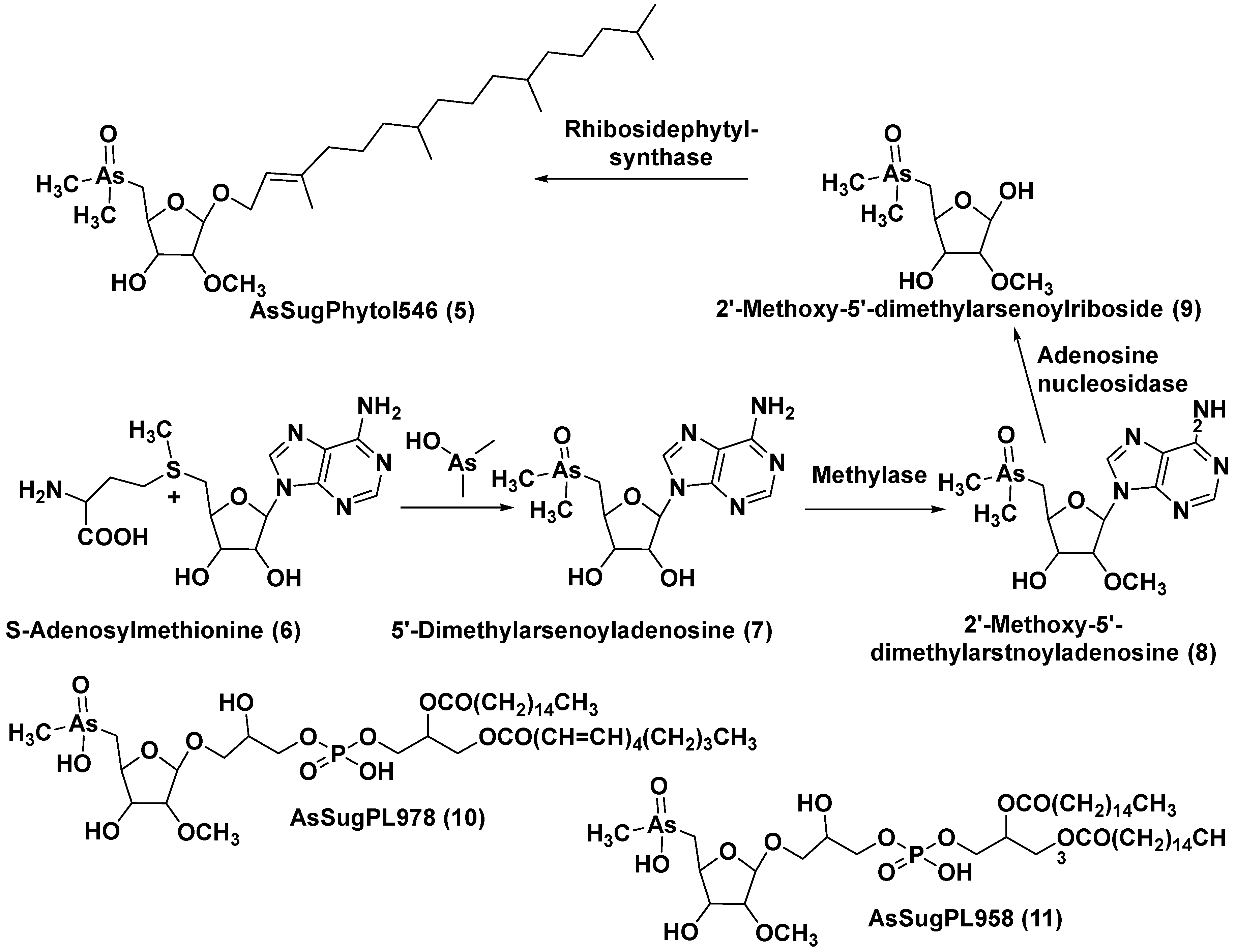
Figure 3.
Main structural types of galactolipids.
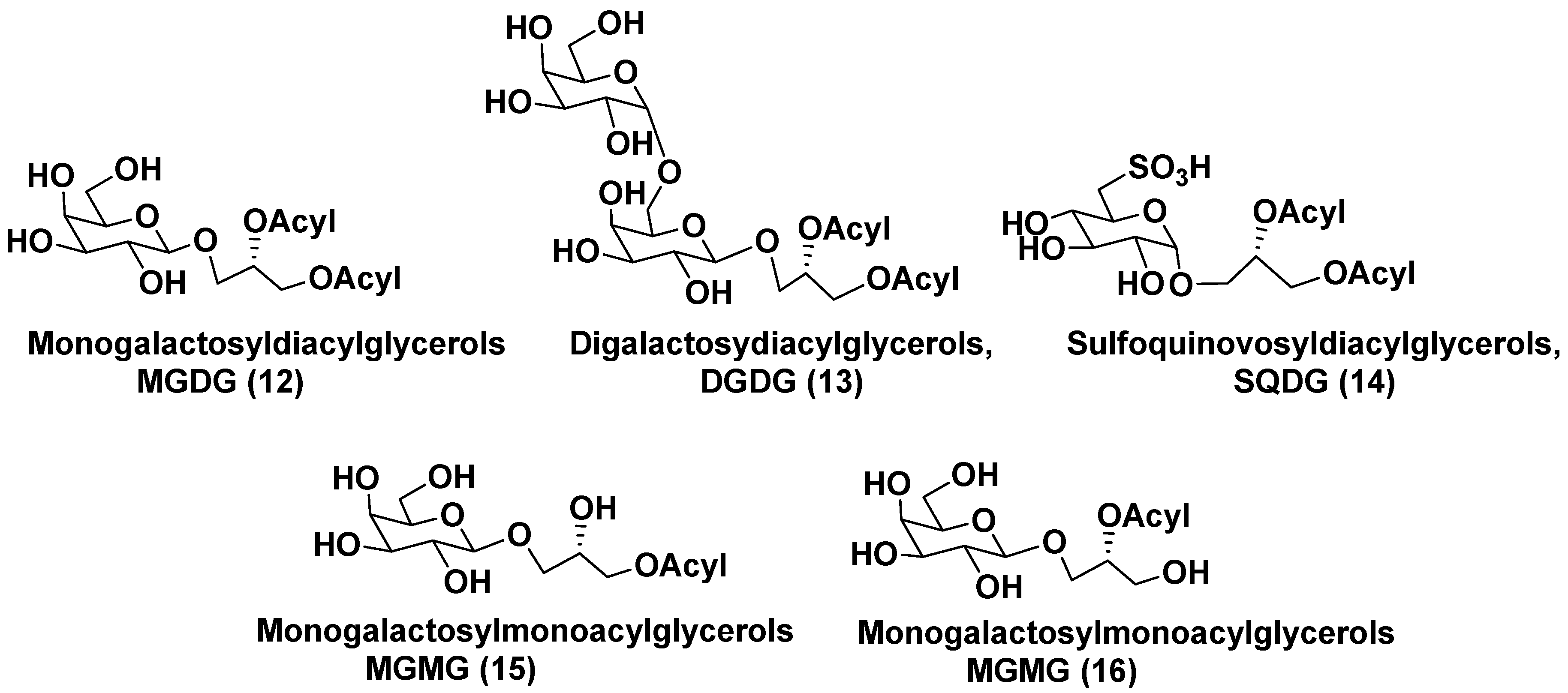
Figure 4.
Structures and enzymatic conversions of some galactolipids.
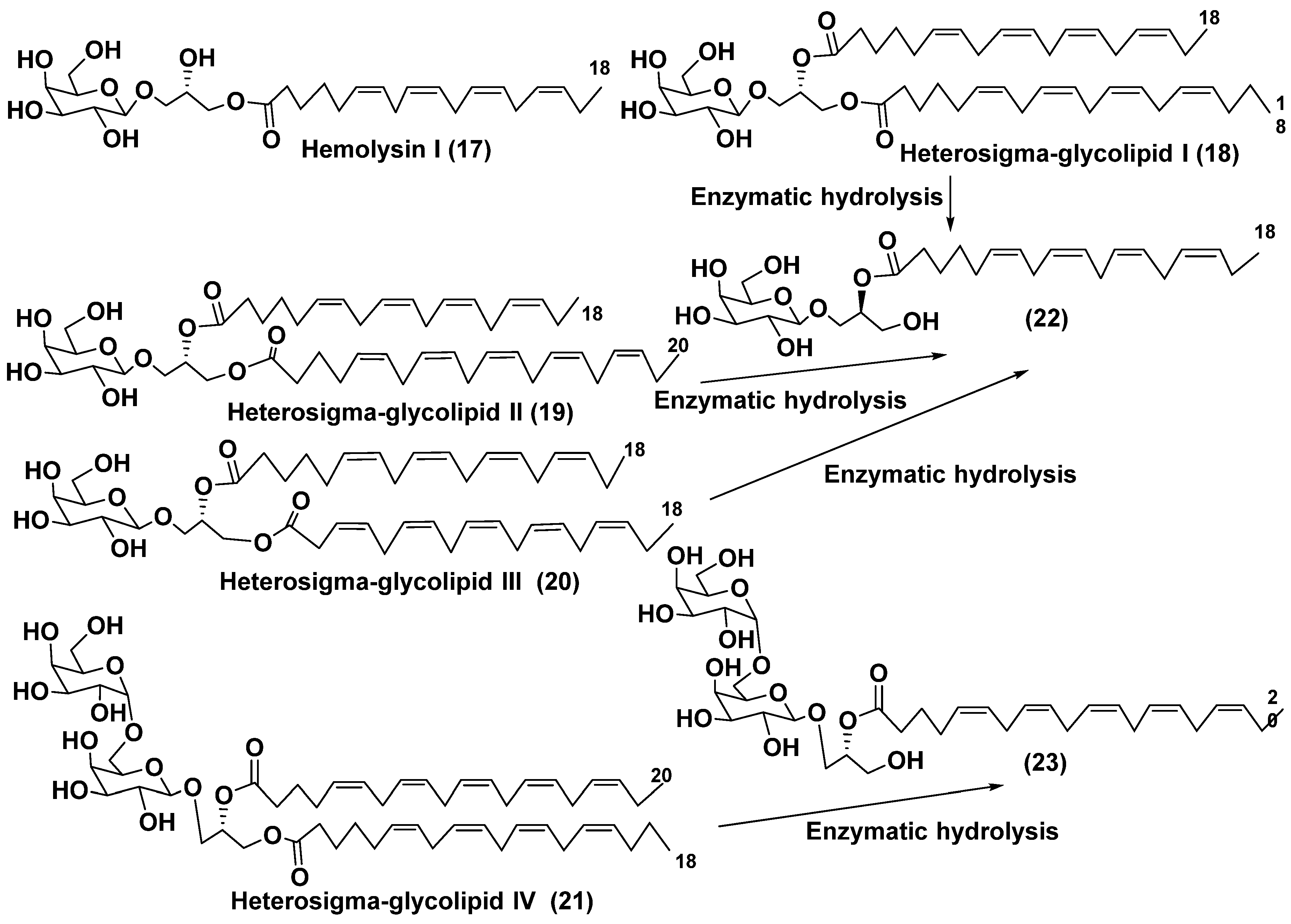
Figure 5.
Structures of other galactolipids from microalgae.
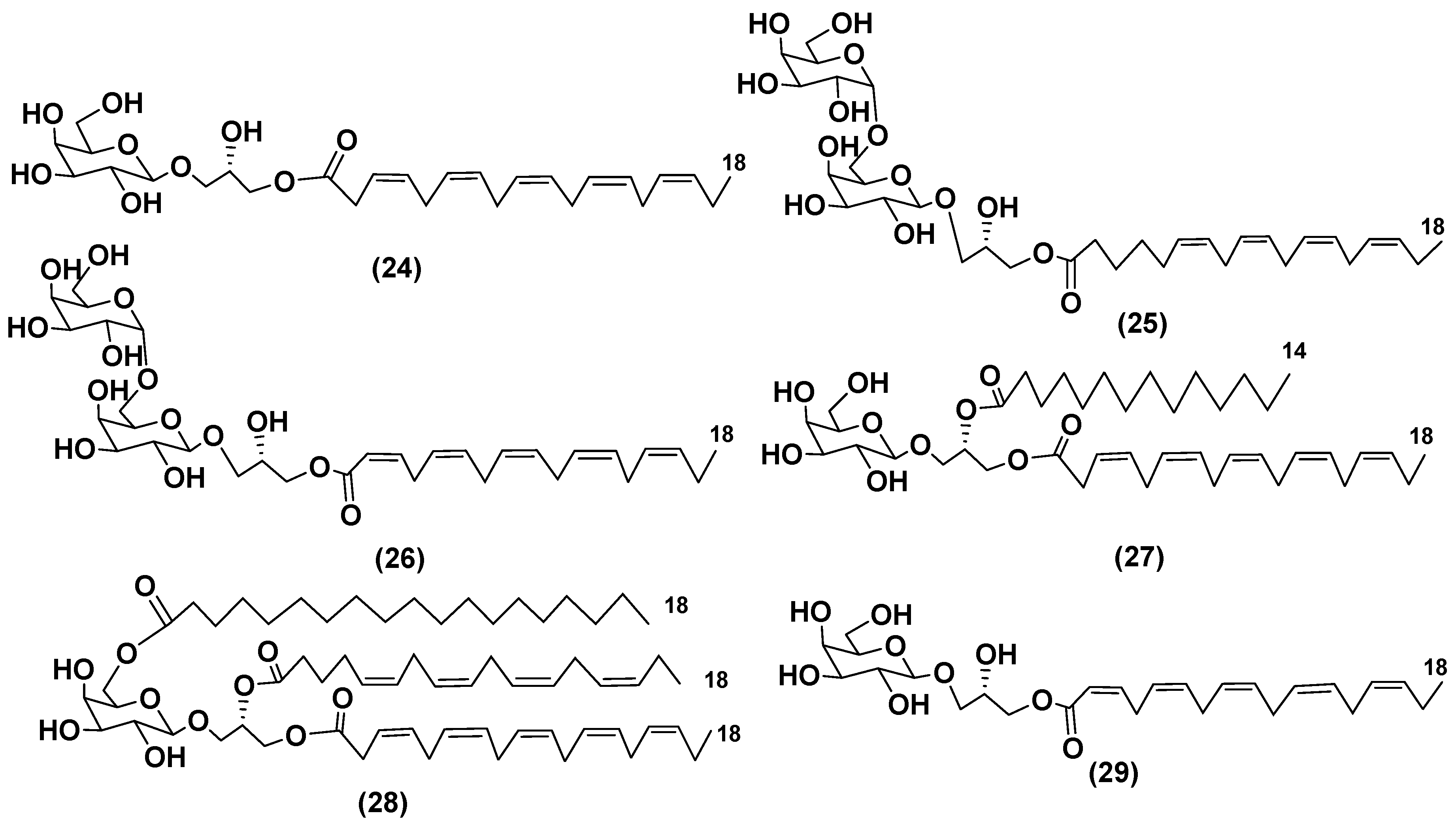
Figure 6.
Fragmentation of galactolipids at ESI MS.
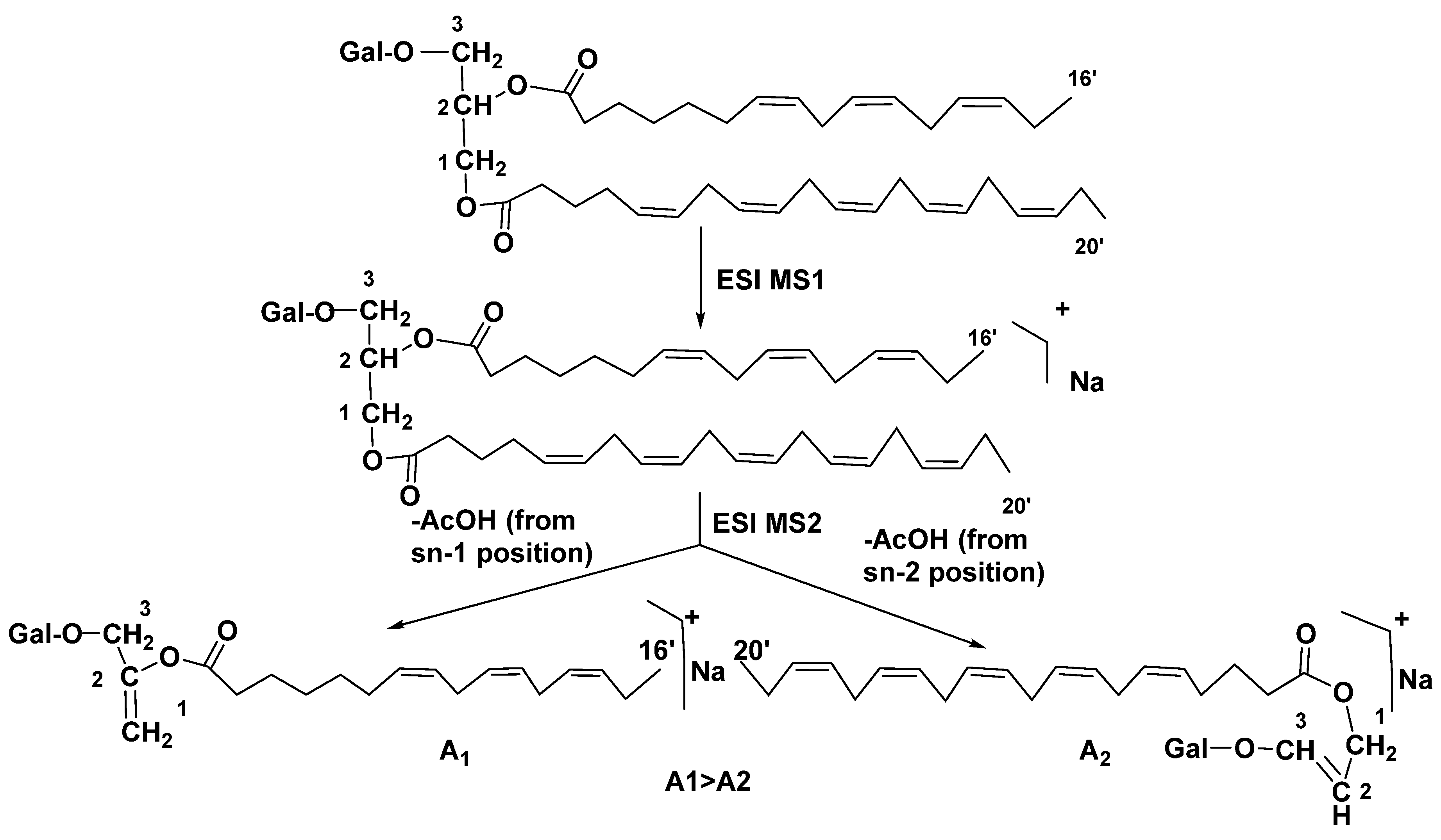
Figure 7.
Structures of sulfoquinovosyl diglycerides 30–36.
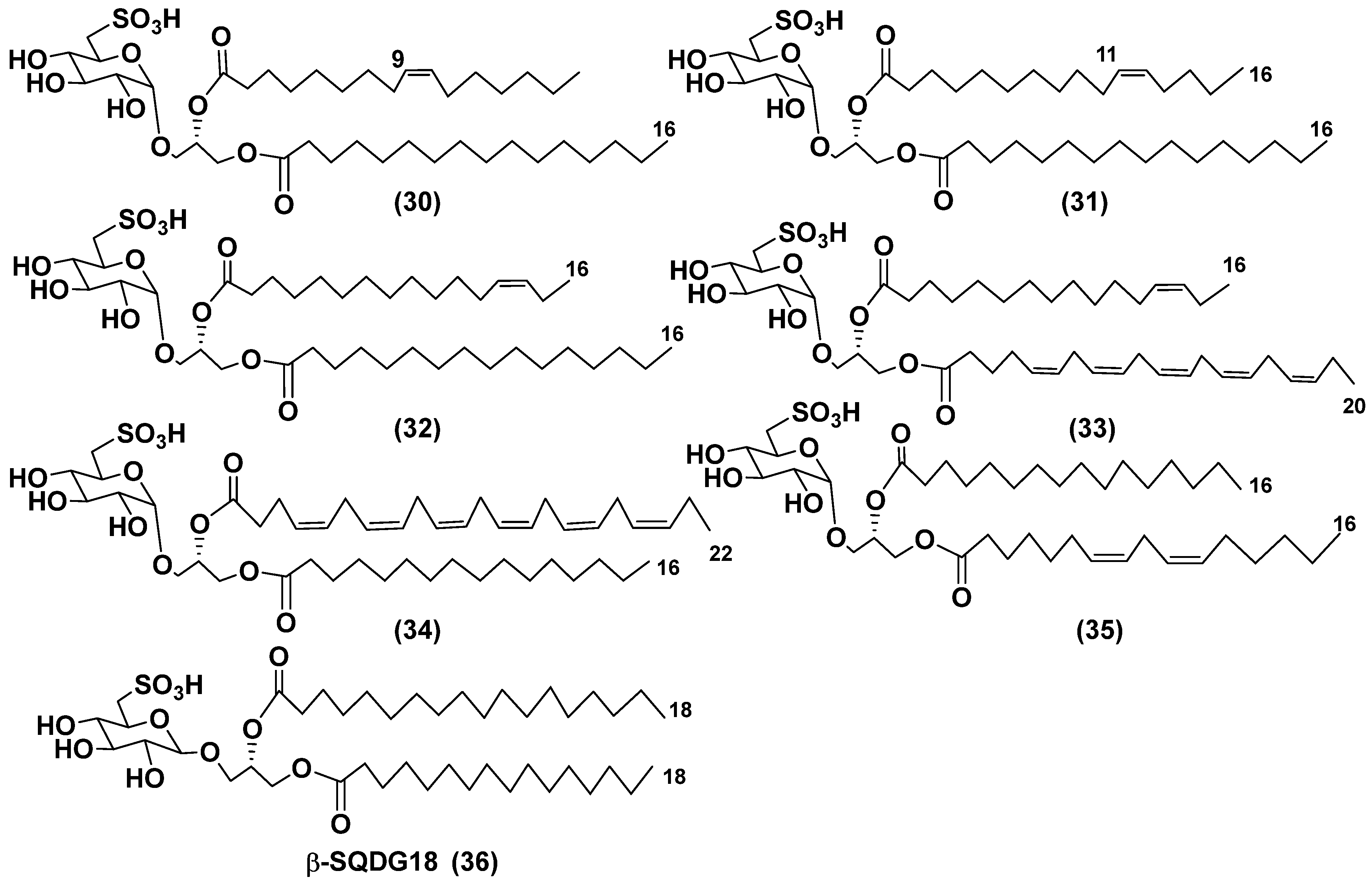
Figure 8.
New plant growth stimulators.
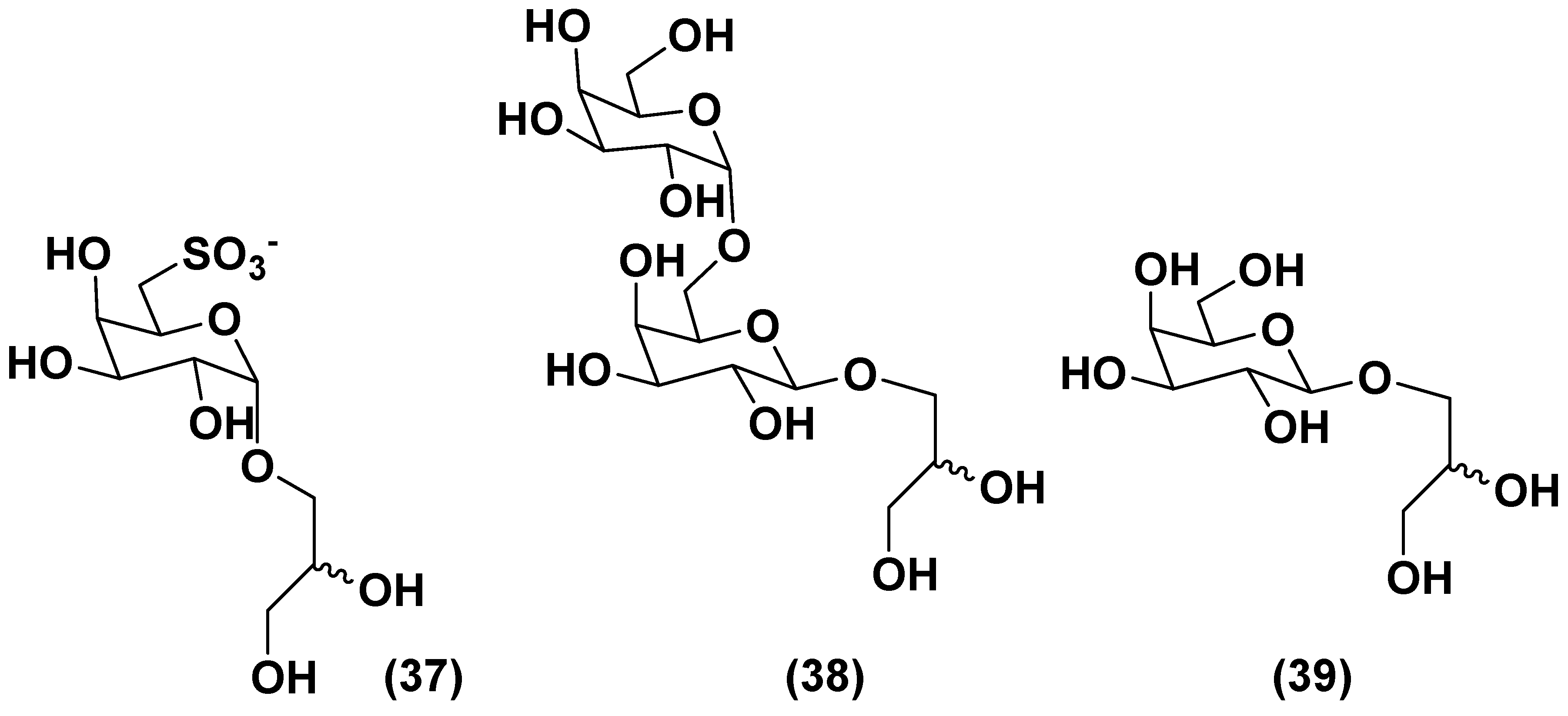
Figure 9.
Phosphogalactodiacyl glycerols.
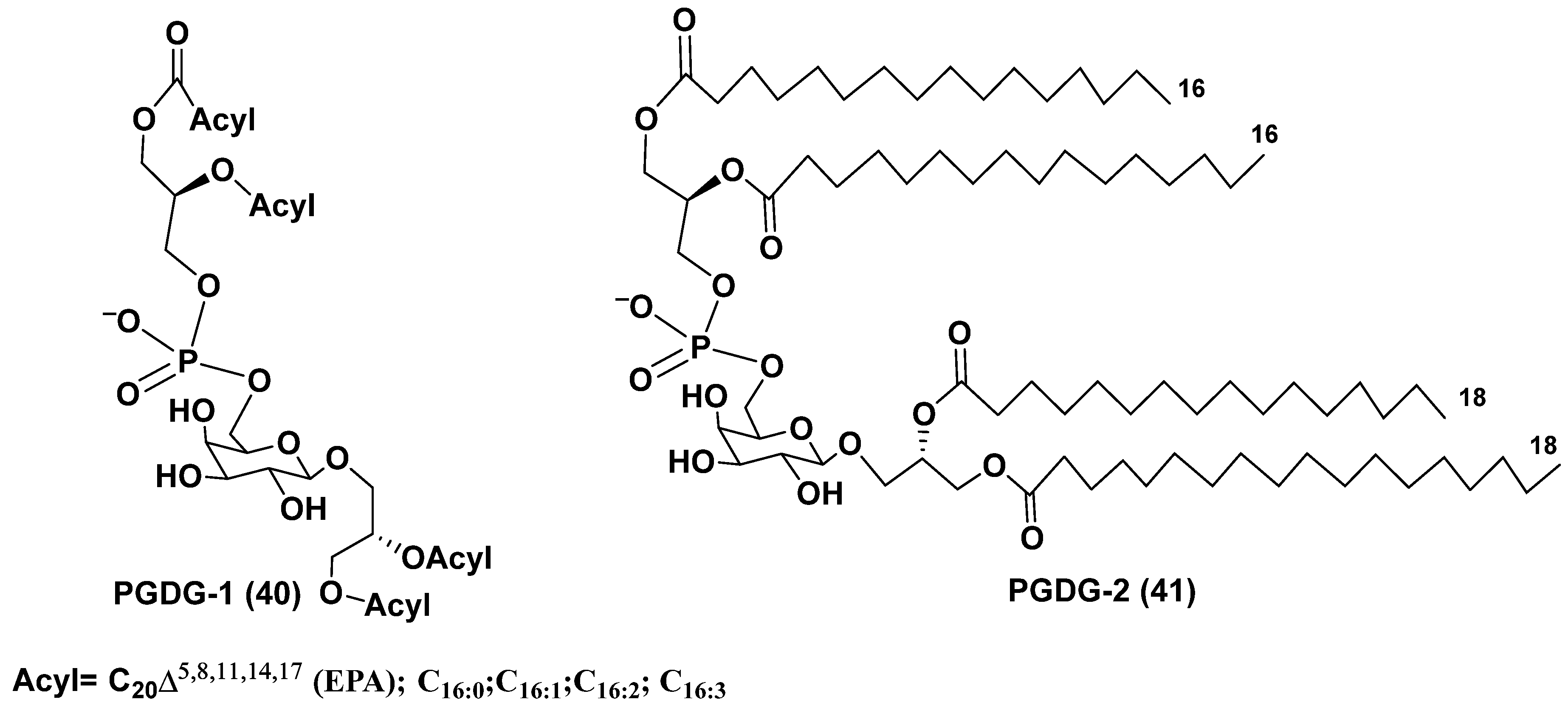
Figure 10.
Biogenesis of phosphogalactodiacyl glycerols.
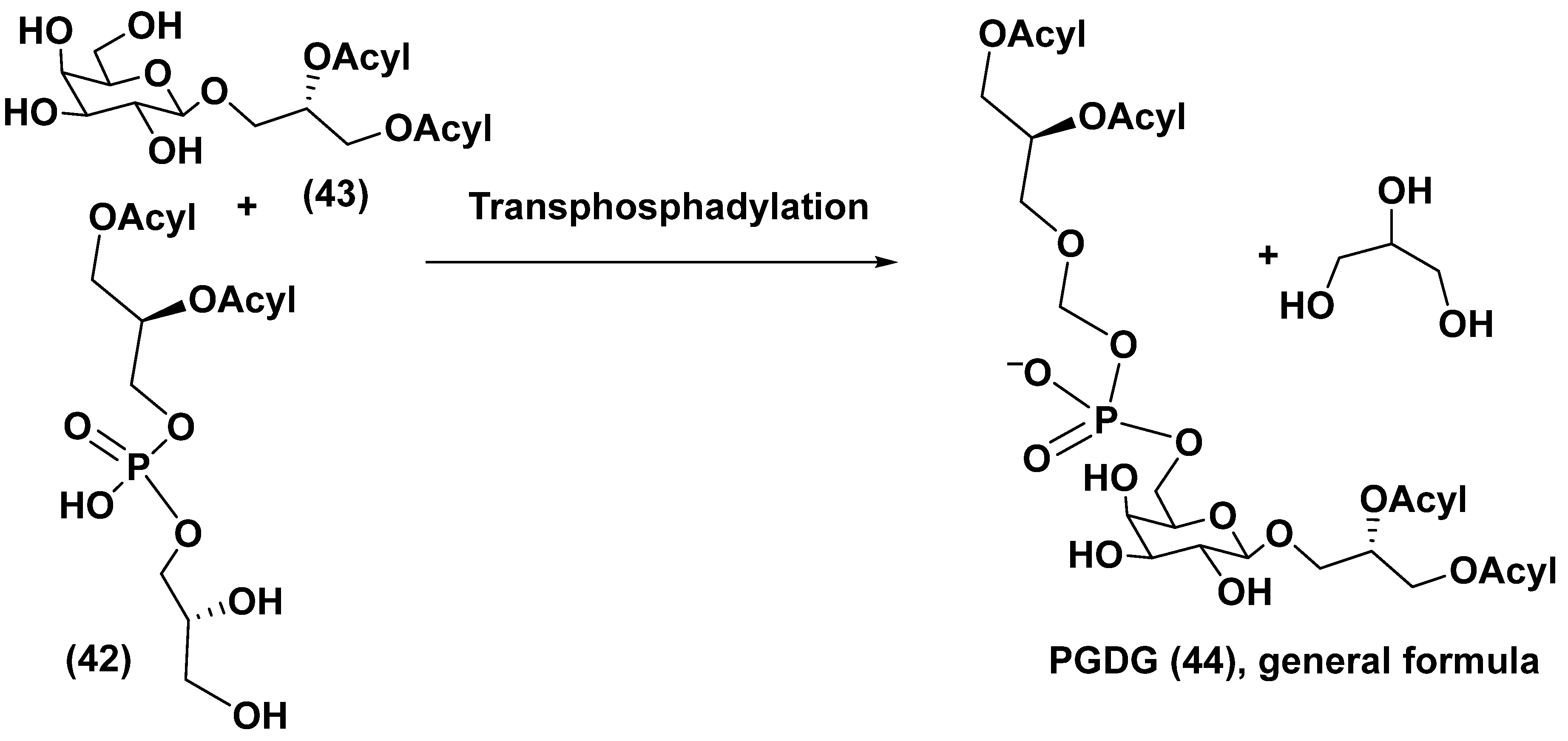
Figure 11.
General formulae of steryl glucosides.
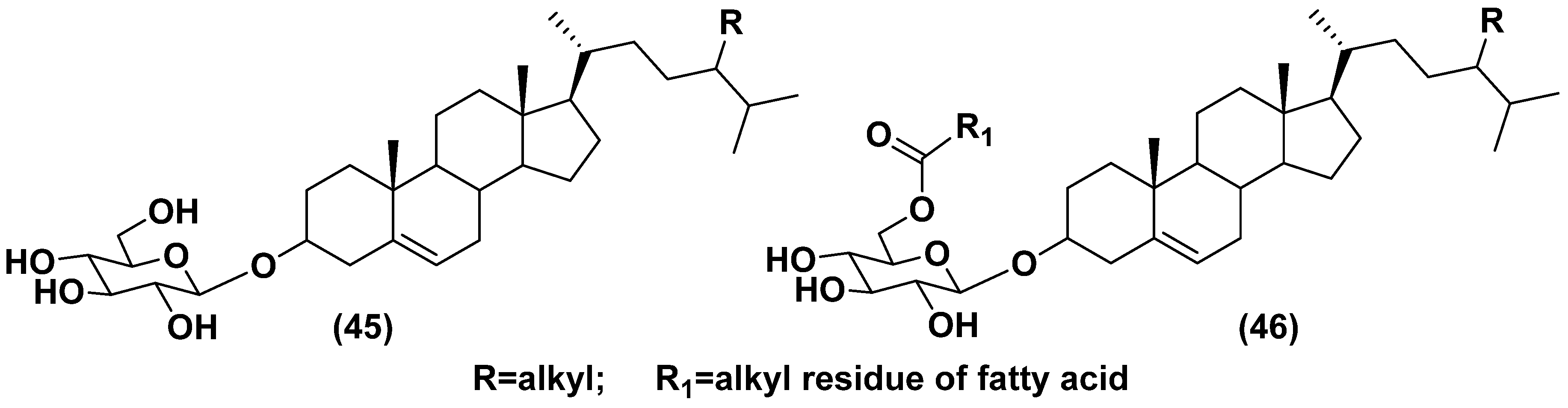
Figure 12.
Biogenesis of monorhamnolids.
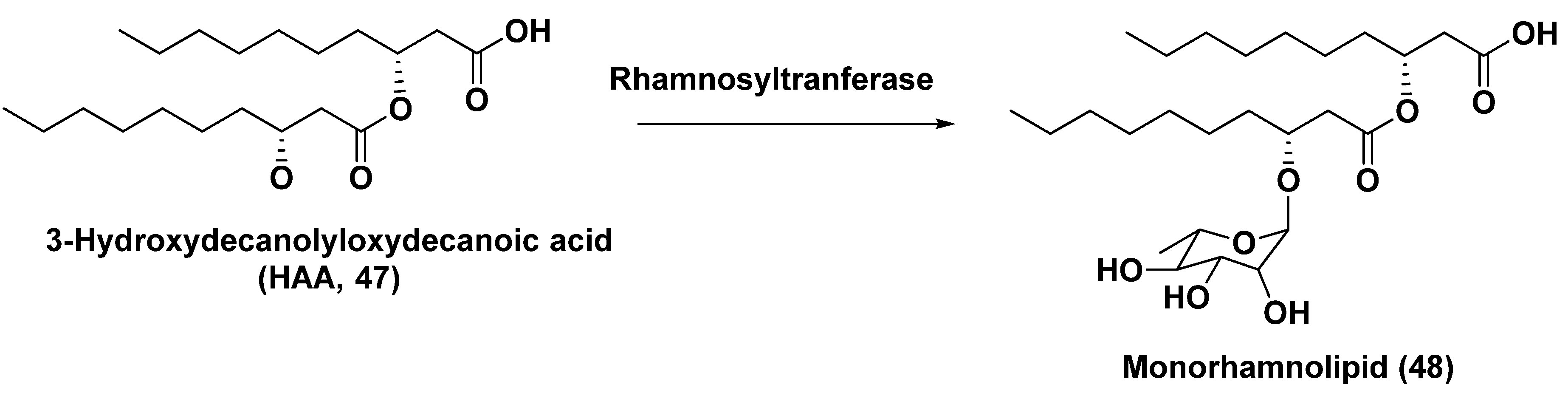
Figure 13.
Amphidinins D (49) and F (50).
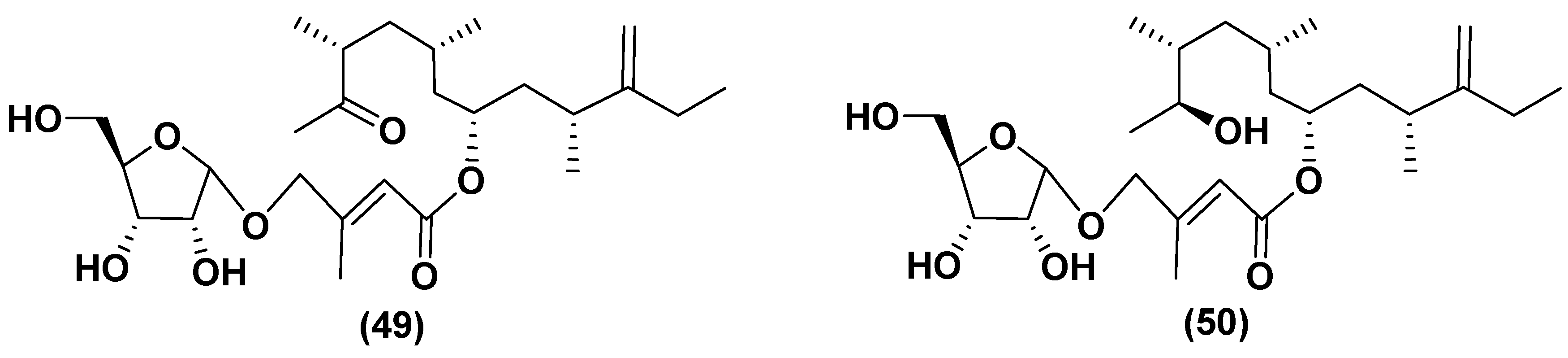
Figure 14.
Prymnesins.
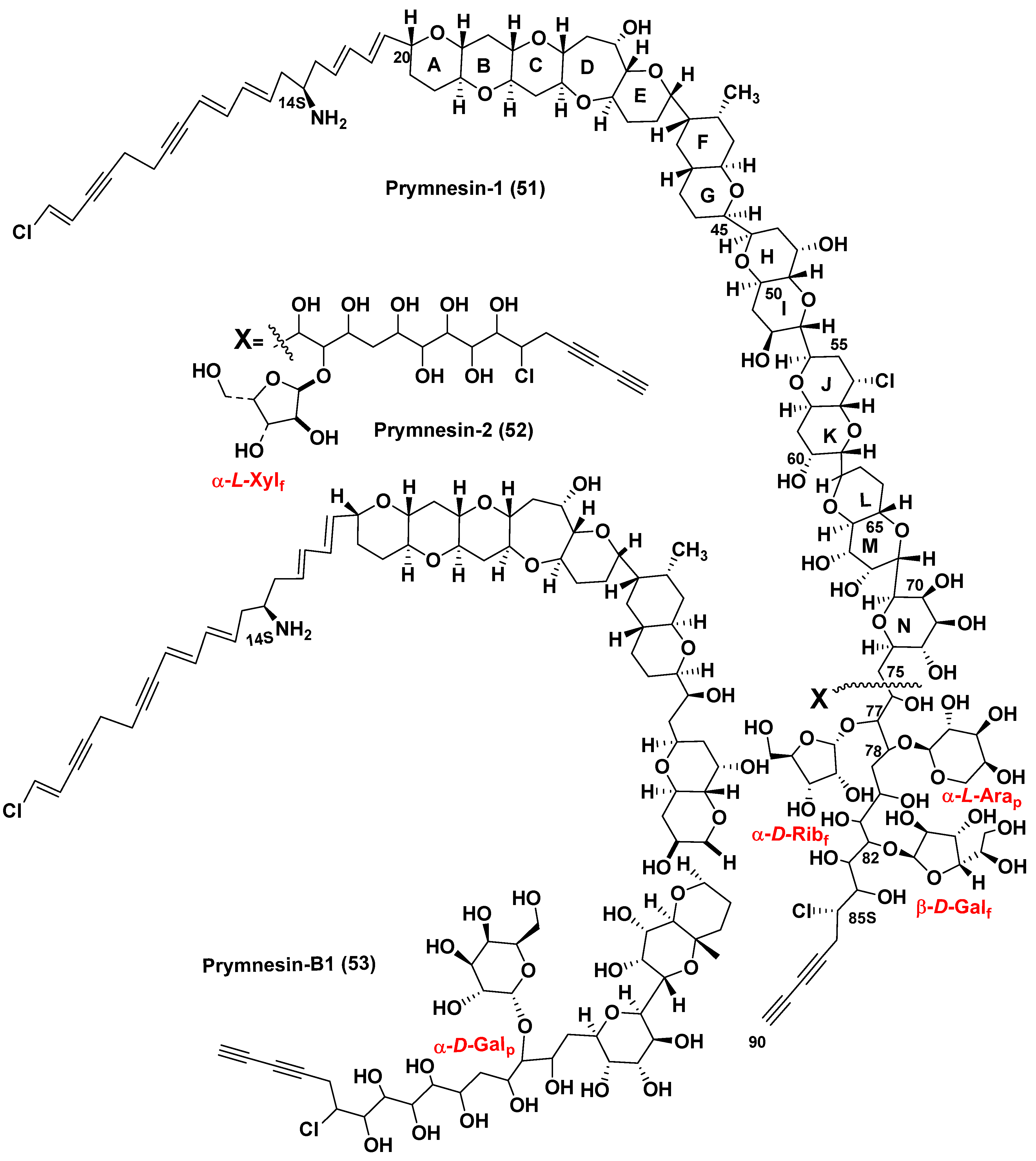
Figure 15.
Unusual glycosphingolipids from virus-infected E. huxleyi.
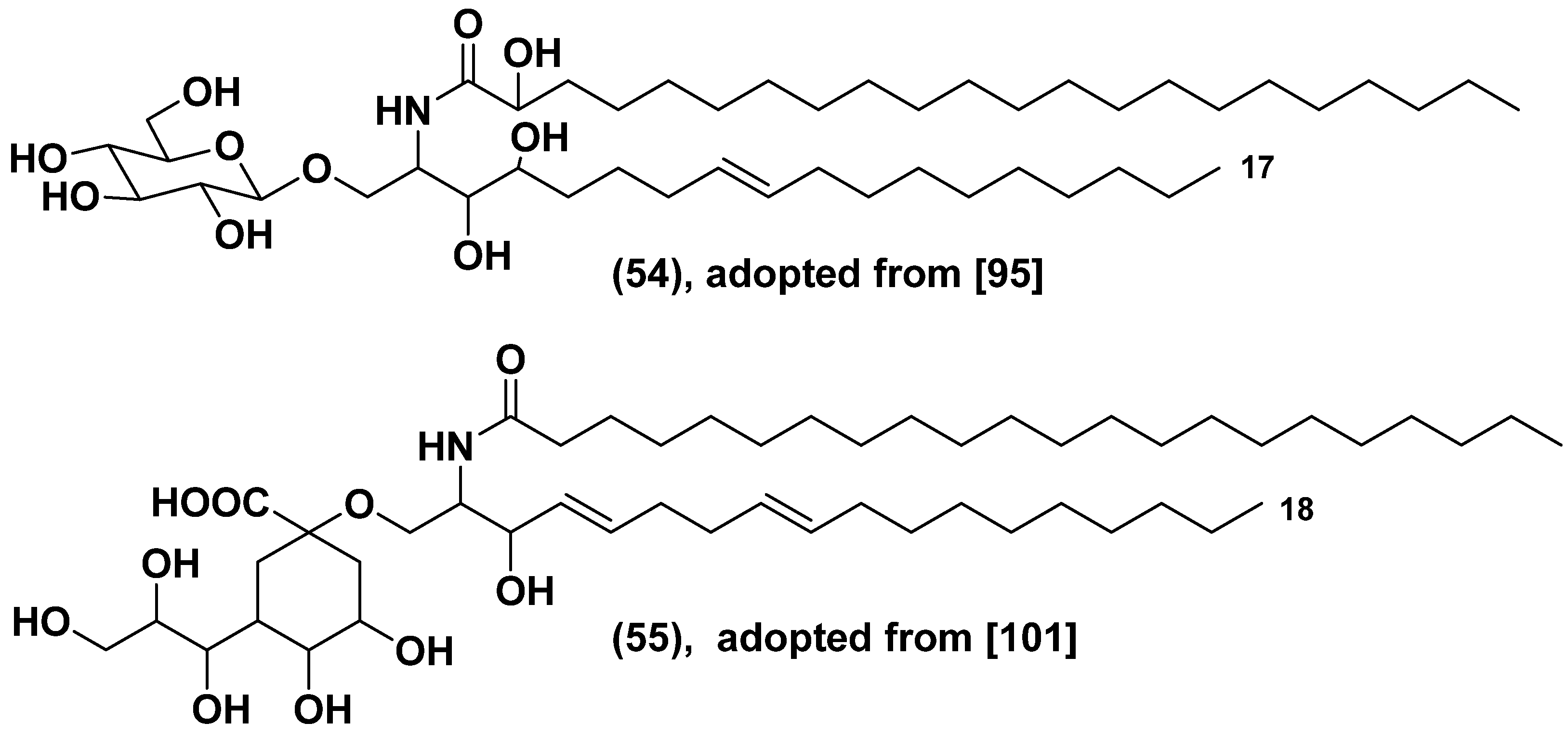
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



