Submitted:
27 June 2023
Posted:
28 June 2023
You are already at the latest version
Abstract
Achieving synchronization between the central and peripheral body clocks is essential for ensuring optimal metabolic function. Meal timing is an emerging field of research that investigates the influence of eating patterns on our circadian rhythm, metabolism, and overall health. This narrative review examines the relationship between meal timing, circadian rhythm, clock genes, circadian hormones, and metabolic function. It analyzes existing literature and experimental data to explore the connection between mealtime, circadian rhythms, and metabolic processes. The available evidence highlights the importance of aligning mealtime with the body’s natural rhythms to promote metabolic health and prevent metabolic disorders. Specifically, studies show that consuming meals later in the day is associated with a elevated prevalence of metabolic disorders, while early time-restricted eating, such as having an early breakfast and an earlier dinner, improves levels of glucose in the blood and substrate oxidation. Circadian hormones, including cortisol and melatonin, interact with mealtimes and play vital roles in regulating metabolic processes. Cortisol, aligned with dawn in diurnal mammals, activates energy reserves, stimulates appetite, influences clock gene expression, and synchronizes peripheral clocks. Consuming meals during periods of elevated melatonin levels, specifically during the circadian night, has been correlated with potential implications for glucose tolerance. Understanding the mechanisms of central and peripheral clock synchronization, including genetics, interactions with chronotype, sleep duration, and hormonal changes, provides valuable insights for optimizing dietary strategies and timing. This knowledge contributes to improved overall health and well-being by aligning mealtime with the body’s natural circadian rhythm.
Keywords:
dawn
; melatonin
; clock genes
; metabolic health
; glucose control
; dinner
; cortisol
1. Introduction
Cardiometabolic disorders have a substantial impact on a large population worldwide, leading to decreased quality of life, increased healthcare utilization, and shortened life expectancy [1]. Numerous risk factors contribute to the increased susceptibility to cardiometabolic diseases, necessitating a systematic approach to comprehend these factors. The timing of fasting/feeding, mealtimes, circadian rhythm, and sleep, as well as the complex interplay between these elements, may exert influence on the circadian rhythms of various organs and cells within the body, particularly when food intake occurs at inappropriate intervals relative to the body’s circadian clock [2].
In humans and mammals, the internal circadian timing system synchronizes with the 24-hour light-dark cycle by receiving light signals that reach a central clock in the hypothalamus [3]. The circadian rhythm, an endogenous timekeeping system, is crucial in regulating various physiological processes in the human body. This biological clock is responsible for maintaining a 24-hour cycle that influences sleep, metabolism, body temperature, and hormone secretion, and the internal clock was for centuries synchronized with mealtimes at specific periods. Moreover, the circadian rhythm and meal timing mutually interact; virtually all mice models with diet-induced dysmetabolism exhibit disrupted feeding patterns, frequently marked by the absence of specific meal times and the dispersion of caloric consumption across both day and night [4].
The widespread availability of electrical lighting in recent times has increased nocturnal activities and granted humans the ability to personally select their light-dark cycles and prolong wakefulness activities well into the night [5]. This ability to modify the timing of wakefulness can lead to a misalignment between the external (environmental) and the internal circadian physiology [6]. Disharmony in circadian rhythms can compromise the functioning of these organs and impact the overall cardiometabolic systems in the body [7] and has been correlated with adverse health outcomes, including diminished energy metabolism [8], impaired glucose metabolism [9], and heightened risk of cardiovascular disease [10].
Furthermore, consuming meals during nighttime hours can disrupt sleep latency, duration, and quality [7]. Insufficient sleep duration and poor sleep quality are established risk factors for cardiometabolic diseases [7]. Additionally, emerging evidence has indicated a connection between eating patterns, mealtimes, and cardiometabolic health in humans [11]. Current evidence from animal studies [12] and human research [13] suggests that consuming food during periods typically allocated for sleep can lead to increased weight and detrimental metabolic health. Groups prone to shifting their activities to later hours, like night or rotating shift workers, or young people such as teenagers and college students, also exhibit a higher prevalence of weight gain and obesity [14]. Numerous circadian events occurring at dawn have been observed in humans, including changes in blood glucose, insulin sensitivity, and hormone levels (such as cortisol), as well as heightened activity in the autonomic nervous system [15,16], which could be linked to early morning meals.
Therefore, this narrative review provides an overview of recent research exploring the importance of synchronization of central and peripheral body clocks and meal timings and the interactions with gene expression, circadian hormones, and metabolism. It focuses on data looking at changes during the early hours of the morning (dawn) and evening (dusk) and their influence on metabolism, with a specific focus on the role of early mealtimes.
2. Search Methods
We adhered to established criteria for searching and reporting the literature for narrative reviews [17,18]. The PubMed database and Google Scholar were utilized to conduct the search using keywords such as "mealtime," "meal timing," "eating schedule," "early meal," "delayed meal," "circadian rhythm," “breakfast,” “dinner,” "circadian clock," "biological clock," "chrononutrition," "circadian misalignment," "metabolism," "metabolic rate," "energy expenditure," "obesity," "body mass index," "adiposity," "weight," "appetite," "cortisol," "melatonin," "insulin," "clock genes," and "cardiovascular health."
Inclusion criteria encompassed original articles (human and animal studies) and systematic reviews until May 31, 2023, with no age restrictions. Exclusion criteria included editorials, opinions, and articles written in languages other than English. To identify additional relevant publications, the reference lists of retrieved articles were reviewed (backward search), and recent articles citing the retrieved papers were examined (ahead search).
For studies relevant to the review’s objectives, information about study design, development, and interventions was extracted. Data collection focused on intervention type, goals, target population, study design and conduct, findings, participant demographics, intervention duration, and results. The investigators’ conclusions were also noted. Both authors independently assessed the retrieved papers for eligibility and extracted study data. Any discrepancies were resolved through discussion and agreement. During data analysis, tables were created by both authors to describe the studies and summarize the results, ensuring accuracy and comprehensiveness.
3. A Brief History of Meal Timings
Meal timing has witnessed changes in different cultures over time [19,20,21,22]. Moreover, religion may have an impact as well on mealtimes. The timing of meals during the Middle Ages was shaped by the presence of daylight. In a time without electricity, people would rise earlier to utilize natural light for a range of tasks, including meal preparation and eating [23]. Due to the absence of electricity, the option to cook dinner in the evening was not available. Peasants had their main meal around midday, although it was a considerably simpler and less extravagant event [23]. The ancient traditional Chinese Medicine, practiced for over 2500 years, suggests that the ideal time for carbohydrate-rich meals is between 0700 and 1100, with smaller meals recommended later in the day during the transition from an active phase ("yang") to a resting phase ("yin") [19,24]. This practice is rooted in the belief that consuming energy-dense meals in the evening can disrupt sleep and various bodily functions. In medieval England, dinner, the main meal of the day, was consumed around noon or 01:00 pm, while supper, a smaller meal, was closer to sunset, typically between 04:00 pm and 06:00 pm [19,25]. Religion also impacts mealtime; for example, during Islamic fasting during or outside the month of Ramadan, followers refrain from eating and drinking between dawn and sunset for a month, and are requested to eat an early pre-dawn meal called Suhur before fasting, which has a religious dimension [26]. Also, during ordinary days, Muslims were historically required to wake up every day for the dawn prayer, where they eat a meal after the prayer and start their day [26]. The advent of artificial lighting, such as oil lamps, led to a shift in dinner and other activities to later times. By the late eighteenth century, dinner had moved to around 04:00 pm or 05:00 pm, and the introduction of lunch was a response to extended fasting periods between breakfast and dinner. The Industrial Revolution also contributed to later dinner times among working-class men who split the workday with a quick noon meal [19,25]. In the USA, an examination of data derived from the National Health and Nutrition Examination Survey (NHANES) demonstrated that dinner and snacks consumed after dinner constituted the largest share of calories, providing nearly 45% of overall calorie consumption [27]. Over 40 years, from 1971-1974 to 2007-2010, the timing of breakfast and lunch generally shifted to later times, while dinner timing remained stable [27]. In the most recent NHANES analysis involving 15,341 adults from the 2009-2014 cycle, the average dinner time was 6:24 pm, with the average time of the last eating episode being 8:18 pm [47]. In fact, contrary to the commonly held belief that humans follow a strict three-meal pattern, it is currently evident that calorie intake occurs sporadically and over an extensive timeframe throughout the 24-hour cycle [28]. Hence, limiting food intake to predetermined time windows may present a readily implementable behavioral intervention with the potential to enhance outcomes in patients with metabolic syndrome.
4. Regulation and Control of the Circadian Body Clocks
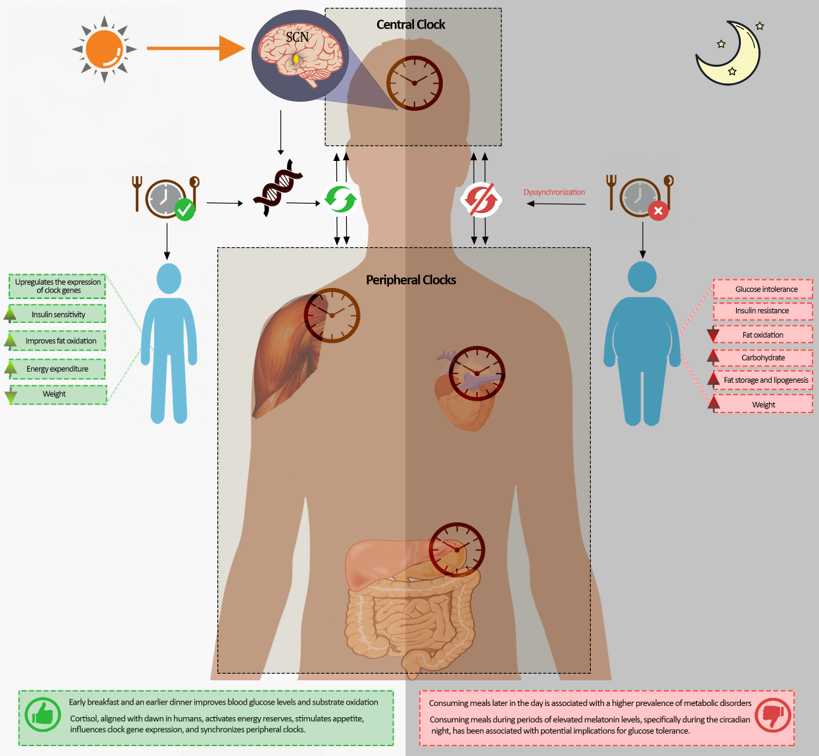
Every organ, tissue, and cell in the body functions according to a biological clock that adheres to a circadian rhythm. Biological clocks can be classified into central and peripheral clocks depending on where they are located anatomically [29]. The primary biological clock is situated within the hypothalamus, precisely within the suprachiasmatic nucleus (SCN), while peripheral clocks exist in all cells throughout the body [30]. These clocks help maintain normal tissue function by regulating the activity of tissue-specific genes [30]. The body’s internal clocks, known as circadian rhythms, rely on independent oscillators within cells to regulate gene expression; circadian clocks are intrinsic biological oscillators that exhibit a periodicity of approximately 24 hours [31], even without external cues [32]. To achieve external and internal synchrony, the body’s biological clocks must be entrained daily.
Circadian clocks are intrinsic biological oscillators that exhibit a periodicity of approximately 24 hours [31]. Found in a wide range of light-sensitive organisms, these clocks play a vital role in coordinating rhythmic activities in accordance with the natural daily cycles [33]. In order to stay synchronized with the external environment, circadian clocks rely on external signals called zeitgebers or timing cues to undergo phase resetting [31,33]. The retinohypothalamic tract primarily entrains the SCN through exposure to external light, whereas neurohormonal factors exert an influence on peripheral clocks and mealtimes [34,35]; peripheral tissues can be moderately affected by nonphotic signals, such as food intake and glucocorticoids [36,37]. The nervous and endocrine systems both regulate the peripheral clocks in vivo. One example is the secretion pattern of glucocorticoids, which is regulated by the SCN and helps synchronize the peripheral clocks [37,38]. Exposure to intense light during nighttime can disrupt the secondary clocks and rhythms that are controlled by the central clock situated in the SCN; similarly, eating meals during the night can also disrupt the peripheral clocks.
External or internal circadian system dyssynchrony has been associated with diverse cardiometabolic dysfunctions, including compromised glucose tolerance and diminished insulin sensitivity, elevated susceptibility to complications related to reduced insulin sensitivity, such as non-alcoholic fatty liver disease (NAFLD), heightened levels of proinflammatory cytokines, elevated arterial blood pressure, and decreased energy expenditure, leading to obesity [39,40,41]. Moreover, epidemiological data support the connection between circadian misalignment and elevated susceptibility to cardiometabolic disorders, diabetes, disorders affecting the cardiovascular system, and NAFLD [42,43,44,45].
Laboratory-controlled studies suggest that acute circadian rhythm misalignment increases the risk of developing cardiometabolic disorders [11]. However, more research is needed in the general population and long-term studies o investigate the potential for adaptation to long-term disruption of the circadian rhythm [46].
5. Clock Genes and Circadian Rhythms
Before discussing the impact of mealtime on cardiometabolic derangements, it is pivotal to cover the function of circadian clock genes in circadian rhythm. Clock genes play a central role in governing circadian rhythms. Their discovery and subsequent research have significantly contributed to our understanding of behavior control and human physiology [47]. The regulation of circadian rhythms is driven by an internal timing system that operates at the level of transcription, resulting in the formation of gene networks that exhibit a 24-hour oscillation [48]. In these networks, clock genes are fundamental in controlling physiological and behavioral rhythms [47]. Notably, the identification of circadian clock genes has been instrumental in understanding behavior control, and it is now evident that the circadian system exerts a significant influence on human physiology [49]. Fruit fly studies led to the discovery of the first circadian mutant gene, “period” [48], and subsequent investigations in mice identified the first circadian mutant mouse, followed by the cloning of the mammalian circadian gene, “Clock” [50,51]. The study of mammalian circadian rhythms has since witnessed remarkable progress, with the inclusion of numerous additional genes in the core clock loop (Cry1 and Cry2, mPer1, mPer2, and mPer3 and Bmal1 genes), which play a vital role in coordinating highly accurate circadian rhythms within various cellular tissues [47,52]. Moreover, clock genes may have implications beyond circadian gene expression, potentially influencing other cellular functions [53].
In mammals, the molecular mechanism that generates circadian rhythms involves a complex interplay between transcriptional activators and repressors, which creates a negative feedback loop [54]. Specifically, two proteins called BMAL1 (Brain and Muscle ARNTL-Like 1 protein) and CLOCK (Circadian Locomotor Output Cycles Kaput), which contain the PAS (Per-Arnt-Sim) domain and helix-loop-helix structure, stimulate the transcription of Cry and Per genes by binding to specific E-box motifs present in these genes [32,55]. Once the repressor proteins CRY (Cryptochrome) and PER (Period) reach a certain concentration, they form complexes with the CLOCK-BMAL1 heterodimer, reducing the transactivation potential of these transcription factors. As a result, the transcription of Per and Cry genes decreases, leading to a decrease in the accumulation of PER and CRY below the concentration required for self-repression. This allows for the initiation of a new transcription cycle for PER and CRY genes [32].
The current knowledge emphasizes the relevance of these findings for understanding the role of circadian rhythm and clock genes in various disorders and diseases and the importance of circadian alignment for better cellular functions [56]. Moreover, clock genes appear to interact with mealtimes [57]. The oscillation of daily gene expression is regulated by the interplay between circadian clocks and feeding cycles at both the transcriptional and translational levels [58]. Feeding has a rapid effect on circadian clock gene expression. For example, in fasted rats, the expressions of Per2 and Dec1 genes in the liver were induced just thirty minutes after refeeding [59]. Similarly, in mice, refeeding affected the expression of Per1, Per2, Dec1, and Rev-erbα in the liver within one hour [59]. In rodents that are active during the night, limiting feeding to the light phase of the day (providing an unlimited amount of food during the normal rest period) alters the circadian clock phase and the activation of clock-regulated genes in peripheral tissues. However, the gene activity in the central timekeeper (SCN) is not affected and remains under the control of the cycle of light and darkness [60,61].
6. Mealtime and cardiometabolic risk
Emerging evidence highlights the significant role of mealtime in controlling metabolic processes and its close interaction alongside the biological clock [11,64]. Chrononutrition, a new discipline, addresses the interplay between mealtime, circadian rhythm, and metabolic regulation [65]. Current research indicates that meal timing influences the circadian cycle, metabolic regulation, and body weight [66]. Consuming meals during inappropriate time periods can cause misalignment between peripheral biological clocks and the central biological clock in the SCN, increasing the potential for developing cardiometabolic disorders [7,35,64]. Studies in both nocturnal species and humans have shown comparable results, wherein consuming meals at inappropriate hours (dark hours, "inactive phase" for humans) is linked to an increased likelihood of experiencing cardiometabolic impairment [67]. In shift workers, night eating has been linked to metabolic disturbances [68,69]. Research has also shown that confining the timing of food intake to either daytime or nighttime can influence the risk of cardiometabolic disorders [28,70,71,72]. However, it is essential to acknowledge that studying the intricate interplay between mealtime, circadian rhythm, and metabolism poses challenges due to the influence of multiple interacting factors. These factors involve individual chronotype, lifestyle elements like shift work, sleep disruptions, and sleep patterns, dietary composition and portion sizes, physical activity levels, environmental factors including nighttime light exposure, age, and genetic factors can interact with these rhythms, with some evidence suggesting that high-fat meals eaten late at night may be particularly disruptive (Figure 1) [64,73,74].
In addition to caloric intake control, maintaining a consistent meal schedule is crucial as it aids in the effective management of energy balance. Consuming meals at set times from sunrise to sunset (between dawn and dusk rather than continuously throughout the day) enhances circadian rhythmicity and promotes a beneficial cycle for optimal metabolic health [19]. In experiments conducted on Wistar rats, it was observed that providing a daily serving of chocolate during the period of activity (breakfast) facilitated the adjustment of the SCN activity to the new schedule in a model of jet lag, leading to faster re-entrainment [75]. Additionally, in a model using rats simulating shift work, having a daily serving of chocolate coinciding with the start of the active phase (breakfast time) prevented disruption of the body’s internal clock by increasing the strength of the day-night activation in the SCN, specifically involving c-Fos (c-Fos is a helpful physiological marker of neural activation, which proved to be successfully utilized in the SCN) [75,76]. In contrast, chocolate consumed during dinner hindered re-synchronization in the jet lag condition and fostered disruption of circadian coordination in the shift work models [75]. Furthermore, rats consuming chocolate during breakfast showed lower weight gain, while those having chocolate during dinner exhibited increased body weight [75]. These findings emphasize the importance of meal timing in regulating circadian synchrony and metabolic function, particularly for a high-calorie and appetizing meal like chocolate.
Nonetheless, the favorable impacts of morning meal (breakfast) consumption on body weight and cardiometabolic well-being have not been consistently reported in all studies [77,78]. A consensus statement from the American Heart Association concluded that while epidemiological evidence indicates a potential adverse impact of consuming meals late in the day on cardiometabolic hazard, the scope of clinical intervention studies addressing this issue has been scant and lacks the specific focus to draw definitive conclusions or formulate recommendations [35]. The statement also emphasized that mealtime and frequency are not the sole determining factors; the length of time between meals and caloric intake are also crucial considerations [35]. In the following sections, we explore the literature that examines the effects of both early and late mealtimes. By thoroughly examining these studies, we aim to gain comprehensive insights into the implications and outcomes associated with different mealtime choices.
7. Dawn and Dusk Feeding Time: The Transition from Fasting to Feeding and Feeding to Fasting
7.1. Time of the Day and Clock Genes
To provide a comprehensive understanding of breakfast and dinner timings, it is necessary to first explore the physiological changes that take place in the early morning. By examining these early morning processes, we can gain insights into the factors that influence meal timing and their potential effect on our overall health and overall wellbeing.
For years, there has been a notion that the autoregulatory feedback cycles involved in the expression of clock genes, which are responsible for generating circadian rhythms, may be influenced differently by the timing of dawn and dusk. As a result, these cycles might play a distinct role in controlling the timing of transitions in circadian rhythms during the evening and morning [79]. At dawn, the central clock in the SCN responds to the increase in light, signaling the transition to the daytime (active) phase in humans and the daytime feeding state. This transition is central to properly regulating glucose metabolism and energy homeostasis. Clock-gene regulation exerts a substantial influence on metabolic pathways, particularly in the initial stages of the active phase (dawn), and immediately after awakening [2,80]. CLOCK-BMAL1 (muscle arnt-like protein) transcriptional regulators form heterodimers and bind to E-box (Enhancer box) enhancers in vertebrates; this binding activates the expression of additional clock genes that produce transcriptional repressors, namely the core clock genes, including PER1/2/3 (period circadian clocks 1, 2, and 3, respectively), CRY1/2 (cryptochrome circadian clocks 1 and 2, respectively), and TIM (timeless) [80]. Despite the CLOCK-BMAL1 complex being bound to its target E-box sequence during the early hours of dawn, the presence of elevated levels of CRY and PER proteins leads to the inhibition of their own transcription, resulting in repressive regulation [81]. This leads to reduced levels of CRY and PER proteins within the cell nucleus since they are not produced by sunrise. In the absence of binding to CLOCK-BMAL1, these complexes can initiate their own transcription, thereby establishing active regulation [82]. As evening approaches, CRY and PER levels increase once again due to the high expression of CG (Cytosine-Guanine) throughout the day. This allows CRY and PER proteins to enter the nucleus by forming a complex with casein kinase 1 epsilon (CK1ε), known as the PER-CRY-CK1ε complex [83]. The positive transcription of CLOCK-BMAL1 is effectively suppressed by this complex, thus initiating another cycle [81,82].
Much like the sleep-wake cycle, the daily rhythm of the feeding/fasting cycle results from a combination of homeostatic needs and temporal control. At present, human daytime feeding habits typically involve two to three meals, even for individuals who reside in an environment devoid of external time cues, following self-determined light-dark (LD) cycles, or experiencing uninterrupted illumination [84]. However, the timings of these meals may differ from one individual to another or from one culture to another, which may interact with the clock genes and affect their functions.
Studies on rats (nocturnal creatures) subjected to a 24-hour fast at various times of the day revealed that refeeding in these rats follows a daily rhythmic pattern, with the highest food consumption occurring when nearing the end of their active phase, specifically at dawn [85]. This finding highlights the influence of circadian regulation in shaping daily feeding, with the highest food consumption occurring at the end of their active phase, specifically at dawn, reflecting the influence of circadian regulation in shaping daily feeding behavior, and suggesting that even the homeostatic feeding response to fasting is under chronostatic control and indicates the importance of the connection between mealtime and the circadian clock [85].
It has been shown that when breakfast is consumed early in the morning, it activates the complex “CLOCK: BMAL1”, involves the activation of SIRT1 (part of the sirtuin family), which leads to the initiation of PERs and CRYs gene transcription [2]. Subsequently, PER and CRY proteins join together to create dimers known as C-P within the cellular cytoplasm [2]. These dimers then move back into the nucleus and inhibit the CLOCK: BMAL1 complex [86]. The inhibition of CLOCK: BMAL1 is reverted by an enzyme called casein kinase I epsilon (CKIε). The activation of SIRT1 by NAD+ results in its interaction with the CLOCK-BMAL1 complex [86].
Additionally, AMPK and SIRT1 exhibit a positive interaction. The transcription process driven by CLOCK-BMAL1 leads to the expression of several genes, including PERs, CRYs, REV-ERBα, RORα, and PGC-1α [2]. This gene activity promotes tissue-specific gene expression, such as increased insulin secretion in β-cells, enhanced GLP-1 (glucagon-like peptide-1) response in L-cells after meals, and improved glucose uptake in muscles through the activation of GLUT4 [87]. During the night, clock genes drive hepatic glucose production, promoting glycogenolysis and gluconeogenesis in the early and later parts of the resting phase [87]. Additionally, the activity of REV-ERBα, RORα, and the BMAL1 transcription factor influences the regulation of PPARα and PGC-1α in adipose tissue, specifically controlling fat breakdown during the night [88,89]. Consequently, early morning feeding, around dawn, appears to play a central role in activating the molecular clock and regulating various physiological processes throughout the day [87].
7.2. Early morning meal
Interestingly, ghrelin, a hormone that stimulates appetite and food intake, reaches its peak secretion at 08:00 in the morning [90]. Likewise, adiponectin levels experience a morning peak around 11:00 am before declining by 08:00 pm [91]. The morning surge of adiponectin activates AMPK (AMP-activated protein kinase), enhances the oxidation of fatty acids, improves insulin sensitivity, increases glucose uptake in muscles, and stimulates glycolysis [91]. This leads to a reduction in hepatic glucose production, resulting in increased glucose utilization and decreased fat accumulation during the early hours of the day [14,92]. In contrast, lower levels of adiponectin in the evening bring about metabolic changes that favor insulin-mediated anabolic processes [2]. As a result, the insulin response following a meal during the evening promotes the accumulation of fat and the process of lipogenesis through the activation of fatty acid synthesis [14,91]. Conversely, the elevation of the leptin hormone (a hormone associated with satiety) in the evening reduces fat accumulation while increasing nocturnal lipolysis [91]. Therefore, having breakfast at an earlier time (dawn meal) aligns with the transition of the circadian clock, facilitating metabolic processes like glucose uptake, insulin sensitivity, and the production of glycogen. This synchronization of mealtime with the central clock aids in maintaining the coordination of peripheral clocks in metabolic tissues such as the liver, skeletal muscle tissue, and fat tissue [2].
Also, the consumption of breakfast at dawn upregulates the expression of clock genes, such as CLOCK, BMAL1, and RORα, which are involved in regulating insulin sensitivity, glucose uptake, and energy expenditure [93]. This upregulation of clock genes also influences the secretion of hormones like GLP-1, which is elevated following meals consumed during the early part of the day in comparison to isocaloric meals consumed in the evening [94]. Thus, having an early morning breakfast is essential for maintaining the synchronization of central and peripheral clocks and promoting metabolic homeostasis. Moreover, breakfast also appears to be important to have a good night’s sleep, which in turn can contribute to earlier bedtimes and rise times and promote a healthier circadian rhythm. A recent exploratory study examined the correlation between breakfast consumption and subjective sleep quality among university students [95]. The findings indicated that students who skipped breakfast, consumed late-night snacks, or substituted meals with snacks were more prone to experiencing overall poor sleep quality [95]. A subsequent, more recent study investigated the relationship between eating habits and sleep difficulties in a large sample of children and adolescents [96]. It revealed that more frequent breakfast consumption and higher intake of fruits and vegetables were linked to fewer sleep difficulties [96].
The regulation of clock genes has been found to improve insulin sensitivity, the responsiveness of β-cells, activity of GLUT-4, glucose uptake in muscles, secretion of post-meal incretins (GLP-1, GLP-2, and GIP), its ability to stimulate insulin release mediated by incretins during the early hours of the active phase [2]. This has been supported by various studies; consequently, various studies have reported that consuming identical meals in the evening leads to a considerably greater glycemic response compared to the morning, indicating a notable disparity in how the body processes and responds to the same meals based on the timing of consumption. [94,97,98,99,100,101,102,103]. Table 1 presents a summary of randomized controlled trials that assessed the relationship between Mealtimes, Circadian Rhythm, and Metabolism [71,104,105,106,107,108,109,110,111,112,113,114,115].
7.3. Energy Expenditure and Circadian Rhythm
The energy expenditure at rest (resting energy expenditure or REE) follows a pattern of fluctuations throughout the day, with the lowest levels occurring during the resting phase [116]. In contrast, the respiratory quotient (RQ), which indicates the utilization of macronutrients, attains its highest level during the early part of the active phase [116]. Moreover, it has been suggested that the thermic effect of food, which refers to the increase in the utilization of energy after eating, is lower in the evening compared to the morning. It seems that the body’s internal clock boosts the thermogenic response to food (diet-induced thermogenesis or DIT) following meals consumed during the early part of the active phase, particularly breakfast, in comparison to the evening [117,118,119,120]. This variation in thermogenesis may be attributed to the influence of circadian rhythms [117]. Furthermore, endocrine factors in the human body may reach their peak at different times of the day due to natural oscillations [121,122,123]. For instance, during the active phase in the morning (around 7 am to 8 am), there is a peak in the hormone cortisol, which plays a role in regulating energy levels and preparing the body for activity [124,125].
In a recent study utilizing a randomized crossover design, the researchers compared early eating (between 08:00 am-06:00 pm) to isocaloric late eating (between 01:00 pm-11:00 pm) in overweight and obese adults; it was observed that late eating led to higher levels of hunger upon waking, reduced energy expenditure, and decreased serum leptin, while increasing the ghrelin-leptin ratio [126]. Another noteworthy finding was that late eating influenced the expression of genes related to adipose tissue, favoring lipid storage by downregulating genes associated with the MAPK (p38 mitogen-activated protein kinase) pathway, the TGF-β signaling pathway, modulation of insulin receptor tyrosine kinases, and autophagy. These changes ultimately promote increased adipogenesis and may contribute to an elevated risk of obesity in humans [126]. These results are in line with previous findings that demonstrate a significant impact of consuming a diet focused on the morning hours reduces hunger scores [2,93,127,128].
Analysis of gene expression in adipose tissue while abstaining from food until noon revealed changes in lipid metabolism pathways. These changes included alterations in the signaling pathway of transforming growth factor-beta (TGF-β) and the regulation of receptor tyrosine kinases, which were consistent with an increment in adipogenesis and a reduction in lipolysis [126]. Moreover, within the dietary intervention protocol that involved consuming meals later in the day, fasting until 01:00 pm resulted in a notable reduction in energy expenditure when compared to the dietary intervention protocol involving early meal consumption that included an early-timed breakfast [126]. The observed reduction in the amount of energy expended during the fasting period until noon suggests that the arrangement of omitting breakfast could potentially elevate the likelihood of developing obesity and diminish the efficiency of strategies employed for weight loss [2]. Conversely, consuming a high-energy breakfast early in the day may aid in weight loss by suppressing appetite [2]. Additionally, this provides a detailed explanation of the underlying mechanisms for the improved results in terms of weight loss observed when more energy is moved to the early part of the day in the DI [129]. Hence, the early hours of the active phase, particularly breakfast, are the ideal time for consuming food, especially for consuming carbohydrates. In contrast, it may be more suitable to reduce energy and carbohydrate intake during the evening and nighttime [130,131,132,133].
7.4. Mealtime, Insulin Sensitivity, and Glucose Response
Consuming breakfast early in the day has a positive impact on the interaction between SIRT1 and AMPK, leading to improved insulin sensitivity, translocation of GLUT-4, and uptake of glucose by muscles. This, in turn, results in better glucose and insulin responses after a meal in the morning compared to the evening. The increased expression of CLOCK and PER2 genes triggers the activation of glycogen synthase 2 (GYS2) gene transcription and promotes the synthesis of glycogen in the liver. As a result, excess glucose derived from the meal is conveyed to the liver and stored as glycogen, whilst the breakdown of glycogen (glycogenolysis) is restricted. Achieving metabolic homeostasis may depend on the temporal alignment of breakfast with the presence of light at dawn.
A study was carried out to examine the impact of prolonged fasting until noon versus eating early breakfast on the expression of clock genes and the response of glycemic, insulin, and incretin levels [134]. The study included both healthy individuals and individuals diagnosed with type 2 diabetes (T2D) [134]. Participants were randomly assigned to two single-test days: one involving consuming breakfast early in the morning at 8:00 am, followed by lunch and dinner, and the other day with solitary lunch and dinner meals, omitting breakfast [134]. Extended fasting for approximately 16 hours until noon on the no-breakfast day altered the expression of clock genes and decreased the mRNA expression of AMPK, BMAL1, PER1, and RORα before and after lunch. This fasting regimen was linked to elevated glucose levels and impaired and delayed release of insulin, while intact GLP-1 responses after lunch were reduced compared to the breakfast day. In contrast, breaking the overnight fast with a high-energy breakfast at 8:00 am on breakfast day exerted a resetting influence on the mRNA expression of these pivotal metabolic clock genes [134]. As a result, there was a notable decrease in postprandial blood sugar levels and enhanced and expedited insulin and GLP-1 responses following the midday meal (lunch) [134].
The improved metabolic effects linked to consuming breakfast may be attributed to food serving as a regulator of the peripheral clocks, potentially aligning and coordinating their functions.
7.5. Early Breakfast or No Early Breakfast
The definition proposed by Timlin and Pereira [135] is widely recognized as the academic standard. According to their definition, breakfast is the initial meal of the day, ingested either before or at the onset of daily activities such as duties, travel, or work. This meal should be eaten within two hours of waking up, preferably no later than 10:00 am in the morning, and should provide an energy intake ranging from 20% to 35% of the individual’s total daily energy requirements. However, it is crucial to acknowledge that this definition does not specify the duration of the overnight fast, which is relevant in the present discussion. However, there is inconsistency in the interpretation of breakfast across various research studies, and there is no unanimous agreement regarding the positive impact of consuming breakfast on weight and cardiometabolic health, as reported in various studies.
A recent meta-analysis comprising 12 randomized controlled trials sourced from affluent countries, aimed to evaluate the influence of consistent breakfast consumption on body weight and energy consumption in adults [136]. The results of this analysis revealed a slight disparity in weight, with a preference towards individuals who skipped breakfast; mean follow-up of 7 weeks; range 2-16 weeks). However, the results across the trials were inconsistent, and individuals assigned to the breakfast group exhibited elevated levels of daily energy intake (approximately 260 extra calories/day) in comparison to individuals who did not consume breakfast (mean difference of 259.79 kcal/day). Moreover, overall, the studies included in the analysis generally exhibited low methodological quality, leading the authors to caution against definitive interpretations of their findings [136].
In the majority of breakfast-related studies that compared the impacts of having or skipping breakfast, there was a lack of consideration for the length of the overnight fasting period [137]. This factor, which could significantly influence metabolic outcomes, was an important variable that should have been taken into account. To illustrate, let us consider two individuals who both had their last meals at 11:00 pm the previous night. One of them had breakfast at 05:00 am the following morning, while the other consumed breakfast at 11:00 am. The discrepancy in the duration of their overnight fasting periods could potentially be the reason behind notable metabolic differences [138]. Conversely, if individuals had similar durations of overnight fasting, regardless of the timing of their meals, it is possible that they would exhibit similar metabolic profiles.
Another more recent comprehensive evaluation and synthesis of observational studies indicated that skipping breakfast in real-world settings may contribute to weight gain and the onset of excess weight and obesity [139]. However, these findings should be interpreted cautiously due to the limited number of studies and the heterogeneity observed, which may introduce publication bias and a small study effect. The methodological differences among the included studies and the lack of information on breakfast composition, quantity, and quality limit the precision and robustness of the results.
These inconsistencies highlight the need for large, high-quality, randomized controlled trials to comprehensively evaluate the influence of breakfast consumption on body weight and the risk of cardiometabolic conditions. A new consensus statement by the American Heart Association acknowledged that data from population-based studies indicate a potentially harmful effect of late mealtime on cardiometabolic risk. However, has been a limitation in conducting intervention studies that can establish a causal relationship and lack the sufficient focus to draw definitive conclusions and make specific recommendations [35]. Furthermore, the consensus statement emphasized that mealtime and frequency alone are not the sole factors contributing to these outcomes; the duration between meals and the caloric intake in each meal are also important considerations [35].
Studies investigating the impact of Ramadan daytime fasting, where individuals consume breakfast in the early morning (30 minutes before dawn) and late evening meals (at sunset), have shown reductions in weight and cardiometabolic risk [7,140]. It is important to note that observational studies have limitations in that they can only establish associations between behaviors and diseases, without being able to ascertain causality or directionality.
A recent small, randomized crossover trial of healthy adult participants suggested that disruption of circadian rhythms induced by delaying mealtimes by 4 hours relative to circadian alignment shifts nutrient metabolism, leading to elevated carbohydrate oxidation and decreased fat oxidation [141]. Despite these metabolic changes, the 24-hour energy expenditure remained consistent between the two conditions. In another controlled randomized experimental crossover study conducted in free-living conditions, eight young and healthy lean volunteers followed two-week and eight-week eating schedules [142]. The schedules, a daytime schedule (08:00 am-07:00 pm) and a delayed schedule (12:00 pm-11:00 pm), were counterbalanced with a 2-week washout period in between [142]. The results showed that the daytime schedule led to weight loss, improved energy metabolism, reduced insulin resistance, and favorable changes in glucose and lipid profiles compared to the delayed schedule [142]. These findings suggest that adopting a daytime eating schedule can be an effective and feasible behavioral modification to promote weight loss and metabolic improvements in real-world conditions.
These inconsistencies highlight the need for large, high-quality, randomized controlled trials to comprehensively assess the impact of breakfast consumption on weight and cardiometabolic risk.
7.6. Skipping Breakfast and Genetics
In addition to the above, a group of scholars developed a Genome-Wide Association Study (GWAS) on breakfast skipping using the UK Biobank dataset, which consisted of approximately 200,000 participants [129]. The results were then replicated in other European populations, including Twin UK and CHARGE [143]. The study identified six genetic variants associated with breakfast skipping. These variants were found to be involved in caffeine metabolism, carbohydrate metabolism, and the control of the biological rhythm. The findings from this large-scale study, involving 200,000 participants, provided evidence suggesting a causal association between genetically determined breakfast skipping and obesity.
Mendelian randomization analysis provided evidence suggesting a causal relationship between skipping breakfast and obesity [144]. However, it is important to interpret these results with caution, considering the limitations associated with Mendelian randomization. One such limitation is that DNA alone does not encompass all the necessary information to fully determine the phenotype, among other potential limitations [143].
7.7. Evening and Late Night Meals
At dusk, the central clock in the SCN responds to the decrease in light, signaling the transition from the daytime feeding state to the nocturnal fasting state. In simple terms, eating late at night and going to bed soon after a late-night dinner can cause a prolonged elevation in blood glucose levels after eating, especially if there is an absence of physical activity during sleep. This can make it harder for blood sugar levels to return to normal when breakfast is eaten without enough time in between. While it may be helpful for most people to eat breakfast, those who regularly eat dinner late at night and go to bed soon after should consider their late-night dinner habits and overall lifestyle when deciding whether to eat breakfast to improve their heart and metabolic health [145].
In theory, having dinner late at night, especially right before going to bed, can cause a long-lasting increase in blood sugar levels after eating. This is because of various influences, like the lack of physical activity during sleep [146]. A cohort study was the first to provide evidence of the connection between consuming dinner late at night and blood sugar control in individuals with type 2 diabetes [147]. It found that having dinner after 8 pm was independently associated with an increase in HbA1c [147]. In several short-term trials, both healthy individuals and those with type 2 diabetes showed higher levels of blood glucose and insulin after eating meals at night [148]. These studies suggest that disrupting the body’s natural circadian rhythm can worsen the normal decrease in glucose tolerance that occurs at night. Moreover, findings derived from the KNHANES study indicated a correlation between eating late at night and a higher prevalence of metabolic syndrome (odds ratio [OR] 1.25; 95% confidence interval [CI]: 1.04–1.49) [149]. Additionally, late-night eating was linked to a reduction in the level of high-density cholesterol (OR 1.18; 95% CI: 1.01–1.38).
Consuming meals late in the day can potentially disrupt the synchronization between the central and peripheral clocks, potentially contributing to the emergence of metabolic disorders [35]. At the same time, new data suggest that consuming an early dinner could be advantageous in maintaining a modest postprandial glycemic response and stable blood glucose levels during the nighttime [109,150]. A randomized controlled trial study examined the effects of early time-restricted eating on blood glucose levels and postprandial lipid metabolism in healthy adults. The results showed that eating dinner at 06:00 pm, as opposed to 09:00 pm, led to improved 24-hour blood glucose levels and better substrate oxidation after breakfast on the following day [109]. Another randomized crossover trial explored the effects of excessive consumption of a late dinner on nocturnal metabolism in young, healthy volunteers (n=20) [151]. The study found that eating a late dinner (at 10:00 pm vs. 06:00 pm) had higher glucose levels, delayed triglyceride peak, reduced free fatty acids, and dietary fatty acid oxidation; it also increased plasma cortisol levels [151]. These metabolic changes were most significant in individuals who typically went to bed earlier, as determined by actigraphy monitoring. These findings suggest that late dinners may contribute to nocturnal glucose intolerance and hinder fat metabolism, potentially promoting obesity if this pattern persists chronically.
However, some previous studies on time-restricted eating (TRE) may have been influenced by the inherent calorie restriction, making it difficult to separate the effects of TRE from calorie restriction [152]. A population-based study utilizing a larger population of the NHANES attempted to address this limitation by investigating the connection between fasting and cardiometabolic markers without the confounding factor of overlapping calorie restriction [152]. Data analysis revealed that delayed timing of the first meal was correlated with elevated levels of CRP, HbA1c %, insulin, glucose, total cholesterol, and LDL cholesterol. It was also associated with lower HDL cholesterol levels [152]. These findings suggest that initiating energy consumption earlier in the day can benefit cardiometabolic endpoints.
8. The Interaction between Mealtime and Circadian Hormones
The classical rodent circadian system simulation proposed by “Pittendrigh and Daan (1976)” has also been suggested to be applicable to humans [153]. According to this model and its expansion by “Illnerova and Vanecek (1982)” [154], the circadian timing system is composed of two distinct rhythmic components; one oscillator is attuned to dusk, regulating evening movement pattern and the initiation of production of melatonin in rodents that are active during the night. The second oscillator is aligned to dawn, controlling morning locomotor activity and the cessation of melatonin secretion.
In a human study, the circadian rhythms of wakefulness, internal body temperature, and hormone release were observed to display clear patterns throughout the 24-hour cycle [155]. These patterns indicated distinct diurnal (daytime) and nocturnal (nighttime) states, with noticeable abrupt transitions resembling a biological "dawn" and "dusk" [155].
When considering humans, the complex circadian pacemaker components synchronized with dusk and dawn can be seen as responsible for regulating the transitions in hormonal secretions, such as melatonin secretion and cortisol secretion, during the evening and morning. These components also help modify the timing of these shifts in accordance with seasonal variations in daylight duration. Certain hormones, like melatonin and glucocorticoids, are tightly regulated by the SCN clock and exert a significant influence on the timing of secondary clocks that express their corresponding receptors [156]. Although peripheral clocks do not directly receive light cues, they can still uphold a daily rhythm under a light-dark cycle, even without a functioning master clock [157].
9. Cortisol
The circadian pattern of corticosteroids in diurnal and nocturnal mammals follows opposite patterns in accordance with the cycle of light and darkness, specifically aligning with dawn and dusk, respectively. However, it functions in both groups to anticipate the beginning of the daily period of wakefulness and activity, aiding in the activation of energy reserves and the stimulation of appetite [158,159]. Glucocorticoids play a crucial role as essential timing signals for numerous peripheral oscillators, facilitating their appropriate synchronization and adjustment to the light-dark (LD) cycle [37,160,161]. Furthermore, it has been suggested that they contribute to preventing sudden shifts in the timing of peripheral clocks within the circadian rhythm. This is particularly important when peripheral clocks become disconnected as a result of consecutive days of fasting and subsequent refeeding cycles [162]. Research has shown that individuals who fast from dawn to dusk exhibit two peaks (acrophases) of cortisol during dawn and dusk, compared to those who do not fast, and individuals with a single peak (acrophase) [163]. These findings suggest that when meals are timed immediately before and after a fasting period that spans from dawn to dusk, the fasting-induced biphasic cortisol circadian rhythm synchronizes the peripheral clocks with the central clock, ensuring their phase alignment, thereby preventing phase shifts between the central and peripheral clocks.
Extensive research has focused on investigating the role of glucocorticoids, which have been linked to the central clock situated in the SCN [164,165]. The secretion of adrenocorticotropin hormone (ATCH) from the anterior pituitary gland is regulated by the SCN [166]. Consequently, this endocrine hormone plays a pivotal role in modulating the release of glucocorticoid hormones from the adrenal glands, which subsequently coordinate the functioning of peripheral tissue circadian clocks.
Studies have provided evidence showing that the regulation of clock gene expression and dietary rhythmicity involves the influence of glucocorticoid receptors [37]. Therefore, considering this relationship, it is plausible to suggest a potential connection between clock genes, the hypothalamic-pituitary-adrenal (HPA) axis, and metabolism [167].
Glucocorticoids, through the glucocorticoid receptor (GR), have been observed to exert a broad impact on gene expression [64,168]. Upon activation, the inactive complexed state of cytoplasmic GR experiences structural modifications and moves into the nucleus after dimerization. In the nucleus, the binding of GR to glucocorticoid response elements (GREs) facilitates the transcription of genes targeted by glucocorticoids [169]. These GREs are responsible for regulating the expression of several genes, including core clock genes such as Per1, Per2, Npas2, and Rev-erbß [170,171]. Both in laboratory settings (in vitro) and living organisms (in vivo), glucocorticoids have shown the capability to alter the circadian rhythms of peripheral clocks [37]. The synthetic glucocorticoid analog dexamethasone, for example, has been observed to stimulate the expression of clock genes and genes influenced by the circadian clock in rat fibroblasts [37]. Additionally, depending on the timing of administration to mice, dexamethasone was observed to either postpone or accelerate the timing of clock gene expression in the liver, kidney, and heart [37]. More recently, there is evidence suggesting that glucocorticoids can influence the human adipose tissue’s biological rhythms as well [172].
The association between meal timing and the diurnal fluctuations of cortisol levels has been documented, highlighting the possibility of both adrenal and extra-adrenal regulatory influences [173]. Glucocorticoids levels are affected by meals and mealtime; it is suggested that when meals are timed immediately before and after dawn and dusk, the dual-phase cortisol circadian rhythm during periods of fasting aligns the timing of peripheral clocks with the central clock, promoting synchronization and preventing disruptions or transitions between the central and peripheral clocks [174].
Nevertheless, it is necessary to conduct randomized-controlled clinical trials that employ a comprehensive multi-omics analysis, encompassing circadian gene expression profiling and metabolomics, and proteomics, to investigate the impact of meals around dawn and dusk in individuals suffering from chronic metabolic conditions and metabolic syndrome.
10. Melatonin
Melatonin levels follow a circadian pattern, reaching their highest point during sleep, decreasing towards the early morning hours, and remaining low until nighttime [175]. At night, melatonin levels begin to rise again in preparation for sleep. Apart from its function in regulating the sleep-wake cycle, melatonin also possesses antioxidant and anti-inflammatory properties and plays a role in controlling glucose and lipid metabolism and the pathophysiology of cardiovascular diseases [176,177]. The reduced levels of melatonin and its major metabolite, 6-sulphatoxymelatonin, have been reported in various cardiovascular diseases, including myocardial infarcts, coronary heart disease, congestive heart failure, and nocturnal hypertension [178,179]. Furthermore, melatonin deficiency caused by factors such as shift work, aging, and exposure to illuminated environments at night can result in glucose intolerance, insulin resistance, metabolic circadian disorganization, and sleep disturbance, all of which pose a threat to health conditions [180].
Meals’ timing may also interact with melatonin to affect circadian rhythm and metabolism. Consuming a meal during nighttime when melatonin levels are elevated, particularly during night shifts, has been suggested as a potential mechanism for an elevated risk of heart disease and diabetes. A study conducted with 40 overweight/obese women of European ancestry who were habitual late eaters revealed that taking melatonin (5 mg) had a negative effect on glucose tolerance [181]. Specifically, the participants who had dinner within 2.5 hours of their usual bedtime and had high levels of natural melatonin experienced a decrease in glucose tolerance. This suggests that when meal timing coincides with elevated melatonin levels, it impairs glucose tolerance. It is important to note that melatonin levels typically rise about 30 minutes before bedtime [182]. Another study utilized a mobile phone application with time-stamped pictures to track participants’ food intake over seven consecutive days, while also evaluating their body composition and the timing of melatonin release in a laboratory setting [14]. The findings revealed that individuals with higher body fat, known as non-lean individuals, consumed most of their calories approximately 1.1 hours closer to the onset of melatonin release, which signifies the start of the biological night, compared to individuals with lower body fat, known as lean individuals [14]. These results provide additional evidence that the timing of meal intake throughout the circadian evening and/or night, independent of more conventional risk factors such as the quantity or content of consumed food and activity level, plays a vital role in determining body composition [14].
Therefore, in countries where dinner is served early, such as Sweden and Germany, the chances of food intake aligning with elevated melatonin levels are low [129]. However, in Spain, where dinner is usually around 10 pm, melatonin levels at dinner time are approximately three times higher, especially among young individuals who have higher natural melatonin levels than older individuals [129]; this situation increases the likelihood of metabolic changes related to glucose [183]. In a randomized crossover study involving a Spanish population, researchers investigated the impact of late eating and elevated melatonin levels on glucose control, particularly in individuals carrying the G allele in the MTNR1B gene associated with type 2 diabetes [184]. The study found that late dinner timing resulted in significantly higher melatonin levels and impaired glucose tolerance, with lower insulin response and higher glucose levels. These effects were more pronounced in individuals carrying the G allele, suggesting that the combination of high melatonin and carbohydrate intake during late eating can lead to insulin secretion defects and impaired glucose control [184].
Therefore, it is advisable to have dinner early, around dusk, and refrain from consuming meals, particularly those with high glycemic content, in close proximity to exogenous melatonin intake or during nighttime when endogenous melatonin levels are typically elevated.
11. Meal timing, Circadian Rhythm, and Gut Microbiota
There is a growing acknowledgment of the crucial role played by the gut microbiome in human health. Additionally, substantial variations in microbiome composition have been observed among different global lifestyles, and it is probable that the modern lifestyle has an impact on the gut microbiome [185]. In recent years, the connection between circadian rhythms and the composition of the gut microbiota (GM) has gained attention [186]. It has been observed that over 50% of the overall microbial composition exhibits rhythmic fluctuations throughout the day [186]. Circadian disruption leads to various changes in gut function, such as increased gut permeability, which can cause dysfunction of the gut barrier and modify the composition of the gut microbiome [187,188]. Furthermore, research has revealed that the GM plays a role in synchronizing the host’s circadian biological clock through various signaling mechanisms [189].
The complex relationship between the circadian rhythm and the GM involves multiple communication pathways, forming a complex two-way system. Within this interplay, diet (composition and timing) plays a crucial role [186]. Therefore, any disruption or alteration in the circadian rhythm can significantly impact the rhythmicity of the gut microbiota, thereby leading to harmful effects on the host’s overall health [186]. Meal timing can have a major effect on the gut microbiota, both in animals and humans. The gut microbiota of mammals exhibits daily fluctuations regulated by the rhythmic patterns of food intake. The timing of food intake influences the daily fluctuations in the composition of the microbiota, and it is observed that the rhythmicity of the microbiota is a dynamic process that can be disrupted or restored in response to changes in feeding behaviors. Consequently, the timing of feeding serves as a connection between the circadian patterns of host behavior and the diurnal variations in the composition and role of the microbiota [190,191]. Furthermore, circadian misalignment in humans can modify the composition of the microbiota, potentially leading to an increase in proinflammatory taxa and a decrease in functional pathways mediated by the microbiota [192]. One of the affected pathways is the biosynthesis of tryptophan, which is crucial for serotonin production [192].
TRE has been revealed to reintroduce members of the Ruminococcaceae family, specifically within the Oscillibacter genus. These members are believed to confer resistance to the metabolic effects of obesity [190,191]. A higher abundance of Firmicutes species in the gut microbiome has been associated with increased adiposity, suggesting their potential involvement in obesity development [193]. However, research evaluating the microbiome in both normal mice and mice under TRE conditions indicates that the abundance of Firmicutes species is more closely linked to the food and feeding pattern rather than obesity or dysmetabolism per se [193]. One prevalent explanation for the documented advantages of TRE is that it emulates innate eating patterns aligned with circadian rhythms, resembling the dietary habits of humans prior to the advent of artificial lighting and the availability of high-energy foods around the clock [4,194,195].
During a study, male Wistar rats were subjected to a 16-hour light and 8-hour dark cycle. They were divided into four groups, each following specific dietary patterns that mimicked breakfast, lunch, dinner, and late-night eating [196]. Remarkably, engaging in late-night eating habits, such as skipping dinner for a night eating (BLN) or skipping breakfast and having a night eating (LDN), led to significant changes in the composition and functions of the gut microbiota [196]. These alterations in the gut microbiota are believed to be involved in the emergence of metabolic disorders.
A small cross-sectional study involving human adults with metabolic syndrome examined the associations between sleep quality, night eating behavior, and gut microbiome composition [197]. The study’s findings indicated potential links between the composition of the gut microbiome and disturbances in circadian rhythms caused by sleep disturbances or late-night eating; nevertheless, it was difficult to determine if the changes in microbiota are related to late eating, sleep disruption, or both. Additional evidence supporting the significance of meal timing in shaping the make-up of the gut microbiota was furnished by the findings of the initial investigation that examined the gut microbiota composition based on chronotype [198]. Carasso and colleagues discovered that individuals classified as evening chronotypes exhibited a greater abundance of Lachnospira [198]. This increased abundance has recently been linked to the consumption of greater quantities of energy in the afternoon and evening, which is a typical characteristic of individuals classified as evening chronotypes [195]. Another small study revealed that individuals with late-night eating habits had a higher abundance of Erysipelotrichales, a member of the Firmicutes phylum [199]. This bacterial group has previously been associated with metabolic disorders and obesity.
In summary, further research is needed to understand better the potential bidirectional relationship between meal timing and the gut microbiome and its implications for overall health. Future research needs to explore the relationship between mealtime, meal frequency and consistency, circadian rhythms, and GM; this area of research remains relatively unexplored thus far. Future studies should explore the specific mechanisms by which meal timing influences the rhythmicity and composition of the gut microbiota while controlling for other potential confounders like meal composition and sleep disruption. Also, investigations should aim to elucidate the bidirectional relationship between meal timing and the gut microbiota, including the potential role of specific bacterial taxa and functional pathways. Understanding these connections will provide valuable insights into the interplay between meal timing, circadian rhythms, and metabolic disorders.
12. Concluding Remarks and Future Directions
The role of diet in maintaining good health is crucial, and aligning food consumption with an individual’s internal circadian clock has been shown to support metabolic well-being. Innovative dietary approaches like Time-Restricted Eating (TRE) have the potential to enhance circadian alignment, leading to a reduction in various metabolic risks [200,201,202]. However, despite the advantages of intermittent fasting and TRE for health, certain limitations exist. There is currently no consensus on the optimal timing of eating and fasting for achieving the best health outcomes, resulting in variations in meal timing across different studies. Moreover, one notable methodological limitation is that TRE inherently results in moderate calorie restriction, which makes it challenging to differentiate the effects of TRF from those of calorie restriction [28,203,204]. Meal timing significantly influences human physiology, and when there is a mismatch between feeding/fasting patterns and the endogenous circadian system, it can impact an individual’s health. The concept of meal timing presents a promising and innovative dietary strategy that holds importance for cardiometabolic health.
The synchronization between the central and peripheral clocks is key for metabolic regulation. Disruptions in this synchronization, such as consuming meals late in the day, have been connected to an elevated susceptibility to metabolic disorders. On the other hand, early time-restricted eating, such as having dinner earlier in the evening, has been shown to positively influence blood glucose levels and substrate oxidation. Aligning meal timing with the natural circadian rhythm may yield favorable effects on metabolic health.
Late-night meals and going to bed soon after can disrupt the natural transition from the daytime feeding state to the nocturnal fasting state. This can result in prolonged increases in blood sugar levels. Regularly eating dinner late at night and going to bed soon after should prompt individuals to reevaluate their late-night eating habits and overall lifestyle choices. Considering the impact on heart and metabolic health, it is essential to be mindful of the timing of our meals.
The precise mechanisms that underlie these associations are not yet fully comprehended. One limitation of large observational studies is the absence of direct measurements of the circadian system or circadian phase, which refers to the timing of the internal clock. Most of the previously described studies did not directly assess the internal circadian system, such as dim light melatonin onset or body temperature rhythms, thus making it challenging to evaluate the extent of circadian rhythm disruption.
Eating at inappropriate times can disrupt the alignment of circadian rhythms in different bodily tissues. Metabolites derived from food consumption also serve as time signals for peripheral clocks. Nevertheless, it is crucial to acknowledge that other factors, such as an unhealthy diet, low sleep quality or duration, and reduced physical activity, can also contribute to poor health. These noncircadian mechanisms may further connect these factors to negative health outcomes. Gaining a better understanding of the underlying mechanisms is essential in order to determine whether and how to intervene in relation to the behaviors above.
Circadian hormones, particularly cortisol, and melatonin, play pivotal roles in the regulation of metabolic processes. Cortisol, which follows a circadian pattern aligned with dawn in diurnal mammals, assists in the activation of energy reserves, appetite stimulation, and peripheral clock functioning. Glucocorticoids, including cortisol, influence the expression of clock genes and the synchronization of peripheral clocks with the central clock. Melatonin, responsible for regulating the sleep-wake cycle, also possesses antioxidant and anti-inflammatory properties. Meal timing that coincides with elevated melatonin levels, especially during the circadian evening and night, may impair glucose tolerance. Being mindful of melatonin release and avoiding meals close to bedtime or during elevated melatonin levels can help maintain healthy glucose control.
To advance our understanding of circadian health and cardiometabolic well-being, Additional studies are required to establish the optimal meal timing and dietary patterns. This research should involve direct measurements of the circadian system and account for noncircadian factors that affect cardiometabolic risk, such as diet quality, sleep quality and duration, and physical activity levels. Furthermore, well-designed RCTs with adequate sample sizes and appropriate follow-up periods are necessary to understand the meal timing and dietary patterns that enhance circadian and cardiometabolic health. Additionally, research should explore the relationship between mealtime, circadian rhythms, and gut microbiota.
Author Contributions
Conceptualization, ASB; writing—original draft preparation, ASB and ARP; writing—review and editing, ASB and ARP; supervision, ASB. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Strategic Technologies Program of the National Plan for Sciences and Technology and Innovation in the Kingdom of Saudi Arabia (MED511-02-08). The study sponsors played no role in the study design, the collection, analysis, or interpretation of data, writing the manuscript, or the decision to submit the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ndisang, J.F.; Rastogi, S. Cardiometabolic diseases and related complications: Current status and future perspective. Biomed Res Int 2013, 2013, 467682. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Rosenblum, R.C.; Wainstein, J.; Twito, O. Influence of Fasting until Noon (Extended Postabsorptive State) on Clock Gene mRNA Expression and Regulation of Body Weight and Glucose Metabolism. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Panda, S. Daily Eating Patterns and Their Impact on Health and Disease. Trends Endocrinol Metab 2016, 27, 69–83. [Google Scholar] [CrossRef]
- Wright, K.P., Jr.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr Biol 2013, 23, 1554–1558. [Google Scholar] [CrossRef]
- de la Iglesia, H.O.; Fernandez-Duque, E.; Golombek, D.A.; Lanza, N.; Duffy, J.F.; Czeisler, C.A.; et al. Access to Electric Light Is Associated with Shorter Sleep Duration in a Traditionally Hunter-Gatherer Community. J Biol Rhythms 2015, 30, 342–350. [Google Scholar] [CrossRef]
- Almeneessier, A.S.; Pandi Perumal, S.R.; BaHammam, A.S. Intermittent fasting, insufficient sleep, and circadian rhythm: Interaction and impact on the cardiometabolic system. Current Sleep Medicine Reports 2018, 4, 179–195. [Google Scholar] [CrossRef]
- McHill, A.W.; Melanson, E.L.; Higgins, J.; Connick, E.; Moehlman, T.M.; Stothard, E.R.; et al. Impact of circadian misalignment on energy metabolism during simulated nightshift work. Proc Natl Acad Sci USA 2014, 111, 17302–17307. [Google Scholar] [CrossRef]
- Morris, C.J.; Yang, J.N.; Garcia, J.I.; Myers, S.; Bozzi, I.; Wang, W.; et al. Endogenous circadian system and circadian misalignment impact glucose tolerance via separate mechanisms in humans. Proc Natl Acad Sci USA 2015, 112, E2225–E2234. [Google Scholar] [CrossRef]
- Morris, C.J.; Purvis, T.E.; Hu, K.; Scheer, F.A. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc Natl Acad Sci USA 2016, 113, E1402–E1411. [Google Scholar] [CrossRef]
- BaHammam, A.S.; Almeneessier, A.S. Recent Evidence on the Impact of Ramadan Diurnal Intermittent Fasting, Mealtime, and Circadian Rhythm on Cardiometabolic Risk: A Review. Front Nutr 2020, 7, 28. [Google Scholar] [CrossRef]
- Salgado-Delgado, R.; Angeles-Castellanos, M.; Saderi, N.; Buijs, R.M.; Escobar, C. Food intake during the normal activity phase prevents obesity and circadian desynchrony in a rat model of night work. Endocrinology 2010, 151, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Baron, K.G.; Reid, K.J.; Horn, L.V.; Zee, P.C. Contribution of evening macronutrient intake to total caloric intake and body mass index. Appetite 2013, 60, 246–251. [Google Scholar] [CrossRef] [PubMed]
- McHill, A.W.; Phillips, A.J.; Czeisler, C.A.; Keating, L.; Yee, K.; Barger, L.K.; et al. Later circadian timing of food intake is associated with increased body fat. Am J Clin Nutr 2017, 106, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Ushijima, K.; Shimba, S.; Fujimura, A. Daily Fasting Blood Glucose Rhythm in Male Mice: A Role of the Circadian Clock in the Liver. Endocrinology 2016, 157, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Oster, H.; Damerow, S.; Kiessling, S.; Jakubcakova, V.; Abraham, D.; Tian, J.; et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab 2006, 4, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Paré, G.; Kitsiou, S. Methods for Literature Reviews. In Handbook of eHealth Evaluation: An Evidence-based Approach; Lau, F., Kuziemsky, C., Eds.; University of Victoria: Victoria, British Columbia, 2017; pp. 157–179. [Google Scholar]
- Templier, M.; Paré, G. Transparency in literature reviews: An assessment of reporting practices across review types and genres in top IS journals. European Journal of Information Systems 2018, 27, 503–550. [Google Scholar] [CrossRef]
- Dashti, H.S.; Scheer, F.; Saxena, R.; Garaulet, M. Timing of Food Intake: Identifying Contributing Factors to Design Effective Interventions. Adv Nutr 2019, 10, 606–620. [Google Scholar] [CrossRef]
- Fjellström, C. Mealtime and meal patterns from a cultural perspective. Scandinavian Journal of Nutrition 2004, 48, 161–164. [Google Scholar] [CrossRef]
- Ochs, E.; Shohet, M. The cultural structuring of mealtime socialization. New Dir Child Adolesc Dev 2006, 35–49. [Google Scholar] [CrossRef]
- Cinotto, S. Everyone would be around the table: American family mealtimes in historical perspective, 1850–1960. New Directions for Child and Adolescent Development 2006, 111, 17–33. [Google Scholar] [CrossRef]
- Winterman, D. Breakfast, lunch and dinner: Have we always eaten them? BBC News Magazine [Internet]. 2012. Available online: https://www.bbc.com/news/magazine-20243692 (accessed on 19 June 2023).
- Eating with the Chinese body clock: Queiscence: Acupuncture & Apothecary. 2016. Available online: https://chinesemedicinemelbourne.com.au/eating-with-the-chinese-body-clock/ (accessed on 17 June 2023).
- McMillan, S. What Time is Dinner? History Magazine [Internet]. 2001. Available online: https://www.history-magazine.com/dinner2.html (accessed on 17 June 2023).
- BaHammam, A.S.; Alghannam, A.F.; Aljaloud, K.S.; Aljuraiban, G.S.; AlMarzooqi, M.A.; Dobia, A.M.; et al. Joint consensus statement of the Saudi Public Health Authority on the recommended amount of physical activity, sedentary behavior, and sleep duration for healthy Saudis: Background, methodology, and discussion. Ann Thorac Med 2021, 16, 225–238. [Google Scholar] [CrossRef]
- Kant, A.K.; Graubard, B.I. 40-year trends in meal and snack eating behaviors of American adults. J Acad Nutr Diet 2015, 115, 50–63. [Google Scholar] [CrossRef]
- Gill, S.; Panda, S. A Smartphone App Reveals Erratic Diurnal Eating Patterns in Humans that Can Be Modulated for Health Benefits. Cell Metab 2015, 22, 789–798. [Google Scholar] [CrossRef]
- Rumanova, V.S.; Okuliarova, M.; Foppen, E.; Kalsbeek, A.; Zeman, M. Exposure to dim light at night alters daily rhythms of glucose and lipid metabolism in rats. Front Physiol 2022, 13, 973461. [Google Scholar] [CrossRef]
- Van Laake, L.W.; Luscher, T.F.; Young, M.E. The circadian clock in cardiovascular regulation and disease: Lessons from the Nobel Prize in Physiology or Medicine 2017. Eur Heart J 2017. [Google Scholar] [CrossRef]
- Manella, G.; Bolshette, N.; Golik, M.; Asher, G. Input integration by the circadian clock exhibits nonadditivity and fold-change detection. Proc Natl Acad Sci USA 2022, 119, e2209933119. [Google Scholar] [CrossRef]
- Ripperger, J.A.; Schibler, U. Rhythmic CLOCK-BMAL1 binding to multiple E-box motifs drives circadian Dbp transcription and chromatin transitions. Nat Genet 2006, 38, 369–374. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat Rev Mol Cell Biol 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Zelinski, E.L.; Deibel, S.H.; McDonald, R.J. The trouble with circadian clock dysfunction: Multiple deleterious effects on the brain and body. Neurosci Biobehav Rev 2014, 40, 80–101. [Google Scholar] [CrossRef]
- St-Onge, M.P.; Ard, J.; Baskin, M.L.; Chiuve, S.E.; Johnson, H.M.; Kris-Etherton, P.; et al. Meal Timing and Frequency: Implications for Cardiovascular Disease Prevention: A Scientific Statement From the American Heart Association. Circulation 2017, 135, e96–e121. [Google Scholar] [CrossRef] [PubMed]
- Birnie, M.T.; Claydon, M.D.B.; Troy, O.; Flynn, B.P.; Yoshimura, M.; Kershaw, Y.M.; et al. Circadian regulation of hippocampal function is disrupted with corticosteroid treatment. Proc Natl Acad Sci USA 2023, 120, e2211996120. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Brown, S.; Marcacci, L.; Tronche, F.; Kellendonk, C.; Reichardt, H.; et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Kon, N.; Iizuka, K.; Fukada, Y.; Sakurai, T.; Hirano, A. Singularity response reveals entrainment properties in mammalian circadian clock. Nat Commun 2023, 14, 2819. [Google Scholar] [CrossRef]
- Reutrakul, S.; Knutson, K.L. Consequences of circadian disruption on cardiometabolic health. Sleep Med Clin 2015, 10, 455–468. [Google Scholar] [CrossRef]
- Saran, A.R.; Dave, S.; Zarrinpar, A. Circadian Rhythms in the Pathogenesis and Treatment of Fatty Liver Disease. Gastroenterology 2020, 158, 1948–1966.e1. [Google Scholar] [CrossRef]
- Alasmari, A.A.; Al-Khalifah, A.S.; BaHammam, A.S.; Alodah, A.S.; Almnaizel, A.T.; Alshiban, N.M.S.; et al. Ramadan Fasting Model Exerts Hepatoprotective, Anti-obesity, and Anti-Hyperlipidemic Effects in an Experimentally-induced Nonalcoholic Fatty Liver in Rats. Saudi Journal of Gastroenterology 2023, in press. [CrossRef]
- Lajoie, P.; Aronson, K.J.; Day, A.; Tranmer, J. A cross-sectional study of shift work, sleep quality and cardiometabolic risk in female hospital employees. BMJ Open 2015, 5, e007327. [Google Scholar] [CrossRef]
- Nagata, C.; Tamura, T.; Wada, K.; Konishi, K.; Goto, Y.; Nagao, Y.; et al. Sleep duration, nightshift work, and the timing of meals and urinary levels of 8-isoprostane and 6-sulfatoxymelatonin in Japanese women. Chronobiol Int 2017, 34, 1187–1196. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, L.; Zhang, Y.; Zhang, B.; He, Y.; Xie, S.; et al. Meta-analysis on night shift work and risk of metabolic syndrome. Obesity reviews: An official journal of the International Association for the Study of Obesity 2014, 15, 709–720. [Google Scholar] [CrossRef]
- Perez-Diaz-Del-Campo, N.; Castelnuovo, G.; Caviglia, G.P.; Armandi, A.; Rosso, C.; Bugianesi, E. Role of Circadian Clock on the Pathogenesis and Lifestyle Management in Non-Alcoholic Fatty Liver Disease. Nutrients 2022, 14, 5053. [Google Scholar] [CrossRef] [PubMed]
- Arble, D.M.; Bass, J.; Behn, C.D.; Butler, M.P.; Challet, E.; Czeisler, C.; et al. Impact of Sleep and Circadian Disruption on Energy Balance and Diabetes: A Summary of Workshop Discussions. Sleep 2015, 38, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Medicine 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Konopka, R.J.; Benzer, S. Clock mutants of Drosophila melanogaster. Proc Natl Acad Sci USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef]
- Ruben, M.D.; Wu, G.; Smith, D.F.; Schmidt, R.E.; Francey, L.J.; Lee, Y.Y.; et al. A database of tissue-specific rhythmically expressed human genes has potential applications in circadian medicine. Sci Transl Med 2018, 10. [Google Scholar] [CrossRef]
- Vitaterna, M.H.; King, D.P.; Chang, A.M.; Kornhauser, J.M.; Lowrey, P.L.; McDonald, J.D.; et al. Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science 1994, 264, 719–725. [Google Scholar] [CrossRef]
- King, D.P.; Zhao, Y.; Sangoram, A.M.; Wilsbacher, L.D.; Tanaka, M.; Antoch, M.P.; et al. Positional cloning of the mouse circadian clock gene. Cell 1997, 89, 641–653. [Google Scholar] [CrossRef]
- Bolsius, Y.G.; Zurbriggen, M.D.; Kim, J.K.; Kas, M.J.; Meerlo, P.; Aton, S.J.; et al. The role of clock genes in sleep, stress and memory. Biochem Pharmacol 2021, 191, 114493. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat Rev Genet 2017, 18, 164–179. [Google Scholar] [CrossRef]
- Lowrey, P.L.; Takahashi, J.S. Genetics of the mammalian circadian system: Photic entrainment, circadian pacemaker mechanisms, and posttranslational regulation. Annu Rev Genet 2000, 34, 533–562. [Google Scholar] [CrossRef]
- Marri, D.; Filipovic, D.; Kana, O.; Tischkau, S.; Bhattacharya, S. Prediction of mammalian tissue-specific CLOCK–BMAL1 binding to E-box DNA motifs. Sci Rep 2023, 13, 7742. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Silvestre-Roig, C.; Ortega-Gomez, A.; Lemnitzer, P.; Poelman, H.; Schumski, A.; et al. Chrono-pharmacological Targeting of the CCL2-CCR2 Axis Ameliorates Atherosclerosis. Cell Metab 2018, 28, 175–182.e5. [Google Scholar] [CrossRef]
- Chaudhari, A.; Gupta, R.; Makwana, K.; Kondratov, R. Circadian clocks, diets and aging. Nutr Healthy Aging 2017, 4, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Vollmers, C.; Gill, S.; DiTacchio, L.; Pulivarthy, S.R.; Le, H.D.; Panda, S. Time of feeding and the intrinsic circadian clock drive rhythms in hepatic gene expression. Proc Natl Acad Sci USA 2009, 106, 21453–21458. [Google Scholar] [CrossRef] [PubMed]
- Tahara, Y.; Otsuka, M.; Fuse, Y.; Hirao, A.; Shibata, S. Refeeding after fasting elicits insulin-dependent regulation of Per2 and Rev-erbalpha with shifts in the liver clock. J Biol Rhythms 2011, 26, 230–240. [Google Scholar] [CrossRef]
- Damiola, F.; Le Minh, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev 2000, 14, 2950–2961. [Google Scholar] [CrossRef]
- Stokkan, K.A.; Yamazaki, S.; Tei, H.; Sakaki, Y.; Menaker, M. Entrainment of the circadian clock in the liver by feeding. Science 2001, 291, 490–493. [Google Scholar] [CrossRef]
- Lanfumey, L.; Mongeau, R.; Hamon, M. Biological rhythms and melatonin in mood disorders and their treatments. Pharmacol Ther 2013, 138, 176–184. [Google Scholar] [CrossRef]
- Sunderram, J.; Sofou, S.; Kamisoglu, K.; Karantza, V.; Androulakis, I.P. Time-restricted feeding and the realignment of biological rhythms: Translational opportunities and challenges. J Transl Med 2014, 12, 79. [Google Scholar] [CrossRef]
- Kessler, K.; Pivovarova-Ramich, O. Meal Timing, Aging, and Metabolic Health. Int J Mol Sci 2019, 20, 1911. [Google Scholar] [CrossRef]
- Pot, G.K. Chrono-nutrition—An emerging, modifiable risk factor for chronic disease? Nutrition Bulletin 2021, 46, 114–119. [Google Scholar] [CrossRef]
- Allison, K.C.; Goel, N. Timing of eating in adults across the weight spectrum: Metabolic factors and potential circadian mechanisms. Physiology & Behavior 2018, 192, 158–166. [Google Scholar] [CrossRef]
- Billingsley, H.E. The effect of time of eating on cardiometabolic risk in primary and secondary prevention of cardiovascular disease. Diabetes Metab Res Rev 2023, e3633. [Google Scholar] [CrossRef] [PubMed]
- Knutsson, A.; Karlsson, B.; Ornkloo, K.; Landstrom, U.; Lennernas, M.; Eriksson, K. Postprandial responses of glucose, insulin and triglycerides: Influence of the timing of meal intake during night work. Nutr Health 2002, 16, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.B.; Coates, A.M.; Davidson, Z.E.; Bonham, M.P. Dietary Patterns under the Influence of Rotational Shift Work Schedules: A Systematic Review and Meta-Analysis. Adv Nutr 2023, 14, 295–316. [Google Scholar] [CrossRef]
- Kwak, J.; Jang, K.A.; Kim, H.R.; Kang, M.S.; Lee, K.W.; Shin, D. Identifying the Associations of Nightly Fasting Duration and Meal Timing with Type 2 Diabetes Mellitus Using Data from the 2016-2020 Korea National Health and Nutrition Survey. Nutrients 2023, 15, 1385. [Google Scholar] [CrossRef]
- Jamshed, H.; Beyl, R.A.; Della Manna, D.L.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves 24-Hour Glucose Levels and Affects Markers of the Circadian Clock, Aging, and Autophagy in Humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef]
- Sutton, E.F.; Beyl, R.; Early, K.S.; Cefalu, W.T.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves Insulin Sensitivity, Blood Pressure, and Oxidative Stress Even without Weight Loss in Men with Prediabetes. Cell Metab 2018, 27, 1212–1221.e3. [Google Scholar] [CrossRef]
- Molina-Montes, E.; Rodriguez-Barranco, M.; Ching-Lopez, A.; Artacho, R.; Huerta, J.M.; Amiano, P.; et al. Circadian clock gene variants and their link with chronotype, chrononutrition, sleeping patterns and obesity in the European prospective investigation into cancer and nutrition (EPIC) study. Clin Nutr 2022, 41, 1977–1990. [Google Scholar] [CrossRef]
- Jones, S.E.; Lane, J.M.; Wood, A.R.; van Hees, V.T.; Tyrrell, J.; Beaumont, R.N.; et al. Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms. Nature Communications 2019, 10, 343. [Google Scholar] [CrossRef]
- Escobar, C.; Espitia-Bautista, E.; Guzmán-Ruiz, M.A.; Guerrero- Vargas, N.N.; Hernández-Navarrete, M.Á.; Ángeles-Castellanos, M.; et al. Chocolate for breakfast prevents circadian desynchrony in experimental models of jet-lag and shift-work. Sci Rep 2020, 10, 6243. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yamaguchi, Y.; Suzuki, T.; Doi, M.; Okamura, H. Effect of Daily Light on c-Fos Expression in the Suprachiasmatic Nucleus under Jet Lag Conditions. Acta Histochem Cytochem 2018, 51, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Ruddick-Collins, L.C.; Morgan, P.J.; Johnstone, A.M. Mealtime: A circadian disruptor and determinant of energy balance? J Neuroendocrinol 2020, 32, e12886. [Google Scholar] [CrossRef] [PubMed]
- Chawla, S.; Beretoulis, S.; Deere, A.; Radenkovic, D. The Window Matters: A Systematic Review of Time Restricted Eating Strategies in Relation to Cortisol and Melatonin Secretion. Nutrients 2021, 13, 2525. [Google Scholar] [CrossRef]
- Daan, S.; Albrecht, U.; van der Horst, G.T.; Illnerova, H.; Roenneberg, T.; Wehr, T.A.; et al. Assembling a clock for all seasons: Are there M and E oscillators in the genes? J Biol Rhythms 2001, 16, 105–116. [Google Scholar] [CrossRef]
- Sanford, A.B.A.; da Cunha, L.S.; Machado, C.B.; de Pinho Pessoa, F.M.C.; Silva, A.; Ribeiro, R.M.; et al. Circadian Rhythm Dysregulation and Leukemia Development: The Role of Clock Genes as Promising Biomarkers. Int J Mol Sci 2022, 23, 8212. [Google Scholar] [CrossRef]
- Sinturel, F.; Gos, P.; Petrenko, V.; Hagedorn, C.; Kreppel, F.; Storch, K.F.; et al. Circadian hepatocyte clocks keep synchrony in the absence of a master pacemaker in the suprachiasmatic nucleus or other extrahepatic clocks. Genes Dev 2021, 35, 329–334. [Google Scholar] [CrossRef]
- Crespo, M.; Leiva, M.; Sabio, G. Circadian Clock and Liver Cancer. Cancers 2021, 13, 3631. [Google Scholar] [CrossRef]
- Cox, K.H.; Takahashi, J.S. Circadian clock genes and the transcriptional architecture of the clock mechanism. J Mol Endocrinol 2019, 63, R93–R102. [Google Scholar] [CrossRef]
- Aschoff, J.; von Goetz, C.; Wildgruber, C.; Wever, R.A. Meal timing in humans during isolation without time cues. J Biol Rhythms 1986, 1, 151–162. [Google Scholar] [CrossRef]
- Rivera-Estrada, D.; Aguilar-Roblero, R.; Alva-Sanchez, C.; Villanueva, I. The homeostatic feeding response to fasting is under chronostatic control. Chronobiol Int 2018, 35, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Rosensweig, C.; Green, C.B. Periodicity, repression, and the molecular architecture of the mammalian circadian clock. Eur J Neurosci 2020, 51, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Hira, T.; Trakooncharoenvit, A.; Taguchi, H.; Hara, H. Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity. Int J Mol Sci 2021, 22, 6623. [Google Scholar] [CrossRef]
- Froy, O.; Garaulet, M. The Circadian Clock in White and Brown Adipose Tissue: Mechanistic, Endocrine, and Clinical Aspects. Endocr Rev 2018, 39, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Stenvers, D.J.; Jongejan, A.; Atiqi, S.; Vreijling, J.P.; Limonard, E.J.; Endert, E.; et al. Diurnal rhythms in the white adipose tissue transcriptome are disturbed in obese individuals with type 2 diabetes compared with lean control individuals. Diabetologia 2019, 62, 704–716. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Neurobiology of food anticipatory circadian rhythms. Physiology & Behavior 2011, 104, 535–545. [Google Scholar] [CrossRef]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int J Mol Sci 2019, 20, 1190. [Google Scholar] [CrossRef]
- Charlot, A.; Hutt, F.; Sabatier, E.; Zoll, J. Beneficial Effects of Early Time-Restricted Feeding on Metabolic Diseases: Importance of Aligning Food Habits with the Circadian Clock. Nutrients 2021, 13, 1405. [Google Scholar] [CrossRef]
- Ruddick-Collins, L.C.; Morgan, P.J.; Fyfe, C.L.; Filipe, J.A.N.; Horgan, G.W.; Westerterp, K.R.; et al. Timing of daily calorie loading affects appetite and hunger responses without changes in energy metabolism in healthy subjects with obesity. Cell Metab 2022, 34, 1472–1485.e6. [Google Scholar] [CrossRef]
- Biancolin, A.D.; Martchenko, A.; Mitova, E.; Gurges, P.; Michalchyshyn, E.; Chalmers, J.A.; et al. The core clock gene, Bmal1, and its downstream target, the SNARE regulatory protein secretagogin, are necessary for circadian secretion of glucagon-like peptide-1. Mol Metab 2020, 31, 124–137. [Google Scholar] [CrossRef]
- Faris, M.E.; Vitiello, M.V.; Abdelrahim, D.N.; Cheikh Ismail, L.; Jahrami, H.A.; Khaleel, S.; et al. Eating habits are associated with subjective sleep quality outcomes among university students: Findings of a cross-sectional study. Sleep Breath 2022, 26, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qu, D.; Liang, K.; Bao, R.; Chen, S. Eating habits matter for sleep difficulties in children and adolescents: A cross-sectional study. Frontiers in Pediatrics 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Dyar, K.A.; Ciciliot, S.; Wright, L.E.; Bienso, R.S.; Tagliazucchi, G.M.; Patel, V.R.; et al. Muscle insulin sensitivity and glucose metabolism are controlled by the intrinsic muscle clock. Mol Metab 2014, 3, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Gil-Lozano, M.; Mingomataj, E.L.; Wu, W.K.; Ridout, S.A.; Brubaker, P.L. Circadian secretion of the intestinal hormone GLP-1 by the rodent L cell. Diabetes 2014, 63, 3674–3685. [Google Scholar] [CrossRef]
- Kuang, J.; Hou, X.; Zhang, J.; Chen, Y.; Su, Z. Identification of insulin as a novel retinoic acid receptor-related orphan receptor alpha target gene. FEBS Lett 2014, 588, 1071–1079. [Google Scholar] [CrossRef]
- Pinho, A.V.; Bensellam, M.; Wauters, E.; Rees, M.; Giry-Laterriere, M.; Mawson, A.; et al. Pancreas-Specific Sirt1-Deficiency in Mice Compromises Beta-Cell Function without Development of Hyperglycemia. PLoS ONE. 2015, 10, e0128012. [Google Scholar] [CrossRef]
- Saad, A.; Dalla Man, C.; Nandy, D.K.; Levine, J.A.; Bharucha, A.E.; Rizza, R.A.; et al. Diurnal pattern to insulin secretion and insulin action in healthy individuals. Diabetes 2012, 61, 2691–2700. [Google Scholar] [CrossRef]
- Yoshino, J.; Imai, S. A clock ticks in pancreatic beta cells. Cell Metab 2010, 12, 107–108. [Google Scholar] [CrossRef]
- Wehrens, S.M.T.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; et al. Meal Timing Regulates the Human Circadian System. Current Biology: CB 2017, 27, 1768–1775.e3. [Google Scholar] [CrossRef]
- Bandin, C.; Scheer, F.A.; Luque, A.J.; Avila-Gandia, V.; Zamora, S.; Madrid, J.A.; et al. Meal timing affects glucose tolerance, substrate oxidation and circadian-related variables: A randomized, crossover trial. Int J Obes (Lond) 2015, 39, 828–833. [Google Scholar] [CrossRef]
- Bo, S.; Fadda, M.; Castiglione, A.; Ciccone, G.; De Francesco, A.; Fedele, D.; et al. Is the timing of caloric intake associated with variation in diet-induced thermogenesis and in the metabolic pattern? A randomized cross-over study. Int J Obes (Lond) 2015, 39, 1689–1695. [Google Scholar] [CrossRef]
- Collado, M.C.; Engen, P.A.; Bandin, C.; Cabrera-Rubio, R.; Voigt, R.M.; Green, S.J.; et al. Timing of food intake impacts daily rhythms of human salivary microbiota: A randomized, crossover study. FASEB J 2018, 32, 2060–2072. [Google Scholar] [CrossRef] [PubMed]
- Manoogian, E.N.C.; Zadourian, A.; Lo, H.C.; Gutierrez, N.R.; Shoghi, A.; Rosander, A.; et al. Feasibility of time-restricted eating and impacts on cardiometabolic health in 24-h shift workers: The Healthy Heroes randomized control trial. Cell Metab 2022, 34, 1442–1456.e7. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Purvis, T.E.; Mistretta, J.; Scheer, F.A. Effects of the Internal Circadian System and Circadian Misalignment on Glucose Tolerance in Chronic Shift Workers. J Clin Endocrinol Metab 2016, 101, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Tajiri, E.; Hatamoto, Y.; Ando, T.; Shimoda, S.; Yoshimura, E. Eating Dinner Early Improves 24-h Blood Glucose Levels and Boosts Lipid Metabolism after Breakfast the Next Day: A Randomized Cross-Over Trial. Nutrients 2021, 13, 2424. [Google Scholar] [CrossRef] [PubMed]
- Pizinger, T.; Kovtun, K.; RoyChoudhury, A.; Laferrere, B.; Shechter, A.; St-Onge, M.P. Pilot study of sleep and meal timing effects, independent of sleep duration and food intake, on insulin sensitivity in healthy individuals. Sleep Health 2018, 4, 33–39. [Google Scholar] [CrossRef]
- Qian, J.; Dalla Man, C.; Morris, C.J.; Cobelli, C.; Scheer, F. Differential effects of the circadian system and circadian misalignment on insulin sensitivity and insulin secretion in humans. Diabetes Obes Metab 2018, 20, 2481–2485. [Google Scholar] [CrossRef]
- Sharma, A.; Laurenti, M.C.; Dalla Man, C.; Varghese, R.T.; Cobelli, C.; Rizza, R.A.; et al. Glucose metabolism during rotational shift-work in healthcare workers. Diabetologia 2017, 60, 1483–1490. [Google Scholar] [CrossRef]
- Xie, Z.; Sun, Y.; Ye, Y.; Hu, D.; Zhang, H.; He, Z.; et al. Randomized controlled trial for time-restricted eating in healthy volunteers without obesity. Nature Communications 2022, 13, 1003. [Google Scholar] [CrossRef]
- Hutchison, A.T.; Regmi, P.; Manoogian, E.N.C.; Fleischer, J.G.; Wittert, G.A.; Panda, S.; et al. Time-Restricted Feeding Improves Glucose Tolerance in Men at Risk for Type 2 Diabetes: A Randomized Crossover Trial. Obesity (Silver Spring) 2019, 27, 724–732. [Google Scholar] [CrossRef]
- Jones, R.; Pabla, P.; Mallinson, J.; Nixon, A.; Taylor, T.; Bennett, A.; et al. Two weeks of early time-restricted feeding (eTRF) improves skeletal muscle insulin and anabolic sensitivity in healthy men. Am J Clin Nutr 2020, 112, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Zitting, K.M.; Vujovic, N.; Yuan, R.K.; Isherwood, C.M.; Medina, J.E.; Wang, W.; et al. Human Resting Energy Expenditure Varies with Circadian Phase. Curr Biol 2018, 28, 3685–3690.e3. [Google Scholar] [CrossRef]
- Morris, C.J.; Garcia, J.I.; Myers, S.; Yang, J.N.; Trienekens, N.; Scheer, F.A. The Human Circadian System Has a Dominating Role in Causing the Morning/Evening Difference in Diet-Induced Thermogenesis. Obesity (Silver Spring) 2015, 23, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Bideyan, L.; Nagari, R.; Tontonoz, P. Hepatic transcriptional responses to fasting and feeding. Genes Dev 2021, 35, 635–657. [Google Scholar] [CrossRef] [PubMed]
- Romon, M.; Edme, J.L.; Boulenguez, C.; Lescroart, J.L.; Frimat, P. Circadian variation of diet-induced thermogenesis. Am J Clin Nutr 1993, 57, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, E.; Simonneaux, V.; Challet, E. Reciprocal Interactions between Circadian Clocks, Food Intake, and Energy Metabolism. Biology (Basel) 2023, 12, 539. [Google Scholar] [CrossRef]
- Carroll, T.; Raff, H.; Findling, J.W. Late-night salivary cortisol measurement in the diagnosis of Cushing’s syndrome. Nat Clin Pract Endocrinol Metab 2008, 4, 344–350. [Google Scholar] [CrossRef]
- Espelund, U.; Hansen, T.K.; Hojlund, K.; Beck-Nielsen, H.; Clausen, J.T.; Hansen, B.S.; et al. Fasting unmasks a strong inverse association between ghrelin and cortisol in serum: Studies in obese and normal-weight subjects. J Clin Endocrinol Metab 2005, 90, 741–746. [Google Scholar] [CrossRef]
- Gavrila, A.; Peng, C.K.; Chan, J.L.; Mietus, J.E.; Goldberger, A.L.; Mantzoros, C.S. Diurnal and ultradian dynamics of serum adiponectin in healthy men: Comparison with leptin, circulating soluble leptin receptor, and cortisol patterns. J Clin Endocrinol Metab 2003, 88, 2838–2843. [Google Scholar] [CrossRef]
- Azizi, M.; Rahmani-Nia, F.; Mohebbi, H. Cortisol Responses and Energy Expenditure atDifferent Times of Day in Obese Vs. Lean Men. World Journal of Sport Sciences 2012, 6, 314–320. [Google Scholar] [CrossRef]
- Brillon, D.J.; Zheng, B.; Campbell, R.G.; Matthews, D.E. Effect of cortisol on energy expenditure and amino acid metabolism in humans. Am J Physiol. 1995, 268 (3 Pt 1), E501–E513. [Google Scholar] [CrossRef]
- Vujovic, N.; Piron, M.J.; Qian, J.; Chellappa, S.L.; Nedeltcheva, A.; Barr, D.; et al. Late isocaloric eating increases hunger, decreases energy expenditure, and modifies metabolic pathways in adults with overweight and obesity. Cell Metab 2022, 34, 1486–1498.e7. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Barnea, M.; Wainstein, J.; Froy, O. Effects of caloric intake timing on insulin resistance and hyperandrogenism in lean women with polycystic ovary syndrome. Clin Sci (Lond) 2013, 125, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Jakubowicz, D.; Landau, Z.; Tsameret, S.; Wainstein, J.; Raz, I.; Ahren, B.; et al. Reduction in Glycated Hemoglobin and Daily Insulin Dose Alongside Circadian Clock Upregulation in Patients With Type 2 Diabetes Consuming a Three-Meal Diet: A Randomized Clinical Trial. Diabetes Care 2019, 42, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Minguez, J.; Gomez-Abellan, P.; Garaulet, M. Timing of Breakfast, Lunch, and Dinner. Effects on Obesity and Metabolic Risk. Nutrients 2019, 11, 2624. [Google Scholar] [CrossRef] [PubMed]
- Manoogian, E.N.C.; Chow, L.S.; Taub, P.R.; Laferrere, B.; Panda, S. Time-restricted Eating for the Prevention and Management of Metabolic Diseases. Endocr Rev. 2022, 43, 405–436. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hutchison, A.T.; Heilbronn, L.K. Carbohydrate intake and circadian synchronicity in the regulation of glucose homeostasis. Curr Opin Clin Nutr Metab Care 2021, 24, 342–348. [Google Scholar] [CrossRef]
- Clayton, D.J.; Mode, W.J.A.; Slater, T. Optimising intermittent fasting: Evaluating the behavioural and metabolic effects of extended morning and evening fasting. Nutr Bull 2020, 45, 444–455. [Google Scholar] [CrossRef]
- Kelly, K.P.; Ellacott, K.L.J.; Chen, H.; McGuinness, O.P.; Johnson, C.H. Time-optimized feeding is beneficial without enforced fasting. Open Biol 2021, 11, 210183. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Wainstein, J.; Landau, Z.; Raz, I.; Ahren, B.; Chapnik, N.; et al. Influences of Breakfast on Clock Gene Expression and Postprandial Glycemia in Healthy Individuals and Individuals With Diabetes: A Randomized Clinical Trial. Diabetes Care 2017, 40, 1573–1579. [Google Scholar] [CrossRef]
- Timlin, M.T.; Pereira, M.A. Breakfast frequency and quality in the etiology of adult obesity and chronic diseases. Nutr Rev 2007, 65 (6 Pt 1), 268–281. [Google Scholar] [CrossRef]
- Sievert, K.; Hussain, S.M.; Page, M.J.; Wang, Y.; Hughes, H.J.; Malek, M.; et al. Effect of breakfast on weight and energy intake: Systematic review and meta-analysis of randomised controlled trials. BMJ 2019, 364, l42. [Google Scholar] [CrossRef] [PubMed]
- Zilberter, T.; Zilberter, E.Y. Breakfast: To skip or not to skip? Front Public Health 2014, 2, 59. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.J.; Kaur, B.; Quek, R.Y.C. Chrononutrition in the management of diabetes. Nutr Diabetes 2020, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Wicherski, J.; Schlesinger, S.; Fischer, F. Association between Breakfast Skipping and Body Weight-A Systematic Review and Meta-Analysis of Observational Longitudinal Studies. Nutrients 2021, 13, 272. [Google Scholar] [CrossRef]
- Fernando, H.A.; Zibellini, J.; Harris, R.A.; Seimon, R.V.; Sainsbury, A. Effect of Ramadan Fasting on Weight and Body Composition in Healthy Non-Athlete Adults: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 478. [Google Scholar] [CrossRef]
- Carabuena, T.J.; Boege, H.L.; Bhatti, M.Z.; Whyte, K.J.; Cheng, B.; St-Onge, M.P. Delaying mealtimes reduces fat oxidation: A randomized, crossover, controlled feeding study. Obesity (Silver Spring) 2022, 30, 2386–2395. [Google Scholar] [CrossRef]
- Allison, K.C.; Hopkins, C.M.; Ruggieri, M.; Spaeth, A.M.; Ahima, R.S.; Zhang, Z.; et al. Prolonged, Controlled Daytime versus Delayed Eating Impacts Weight and Metabolism. Curr Biol 2021, 31, 650–657.e3. [Google Scholar] [CrossRef]
- Dashti, H.S.; Merino, J.; Lane, J.M.; Song, Y.; Smith, C.E.; Tanaka, T.; et al. Genome-wide association study of breakfast skipping links clock regulation with food timing. Am J Clin Nutr 2019, 110, 473–484. [Google Scholar] [CrossRef]
- LEWONTINRC. The analysis of variance and the analysis of causes. International Journal of Epidemiology 2006, 35, 520–525. [Google Scholar] [CrossRef]
- Nakajima, K.; Suwa, K. Association of hyperglycemia in a general Japanese population with late-night-dinner eating alone, but not breakfast skipping alone. J Diabetes Metab Disord 2015, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Nakamura, K.; Ogata, H.; Miyashita, A.; Nagasaka, S.; Omi, N.; et al. Acute effect of late evening meal on diurnal variation of blood glucose and energy metabolism. Obesity research & clinical practice 2011, 5, e169–e266. [Google Scholar] [CrossRef]
- Sakai, R.; Hashimoto, Y.; Ushigome, E.; Miki, A.; Okamura, T.; Matsugasumi, M.; et al. Late-night-dinner is associated with poor glycemic control in people with type 2 diabetes: The KAMOGAWA-DM cohort study. Endocr J 2018, 65, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Leung, G.K.W.; Huggins, C.E.; Ware, R.S.; Bonham, M.P. Time of day difference in postprandial glucose and insulin responses: Systematic review and meta-analysis of acute postprandial studies. Chronobiol Int 2020, 37, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Ha, K.; Song, Y. Associations of Meal Timing and Frequency with Obesity and Metabolic Syndrome among Korean Adults. Nutrients 2019, 11, 2437. [Google Scholar] [CrossRef]
- Kajiyama, S.; Imai, S.; Hashimoto, Y.; Yamane, C.; Miyawaki, T.; Matsumoto, S.; et al. Divided consumption of late-night-dinner improves glucose excursions in young healthy women: A randomized cross-over clinical trial. Diabetes Res Clin Pract 2018, 136, 78–84. [Google Scholar] [CrossRef]
- Gu, C.; Brereton, N.; Schweitzer, A.; Cotter, M.; Duan, D.; Borsheim, E.; et al. Metabolic Effects of Late Dinner in Healthy Volunteers-A Randomized Crossover Clinical Trial. J Clin Endocrinol Metab 2020, 105, 2789–2802. [Google Scholar] [CrossRef]
- Wirth, M.D.; Zhao, L.; Turner-McGrievy, G.M.; Ortaglia, A. Associations between Fasting Duration, Timing of First and Last Meal, and Cardiometabolic Endpoints in the National Health and Nutrition Examination Survey. Nutrients 2021, 13, 2686. [Google Scholar] [CrossRef]
- Pittendrigh, C.S.; Daan, S. A functional analysis of circadian pacemakers in nocturnal rodents. Journal of comparative physiology 1976, 106, 333–355. [Google Scholar] [CrossRef]
- Illnerová, H.; Vaněček, J. Two-oscillator structure of the pacemaker controlling the circadian rhythm of N-acetyltransferase in the rat pineal gland. Journal of Comparative Physiology 1982, 145, 539–548. [Google Scholar] [CrossRef]
- Wehr, T.A.; Aeschbach, D.; Duncan, W.C., Jr. Evidence for a biological dawn and dusk in the human circadian timing system. J Physiol 2001, 535 Pt 3, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Buijs, R.M.; Wortel, J.; Van Heerikhuize, J.J.; Feenstra, M.G.; Ter Horst, G.J.; Romijn, H.J.; et al. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur J Neurosci 1999, 11, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Gnocchi, D.; Bruscalupi, G. Circadian Rhythms and Hormonal Homeostasis: Pathophysiological Implications. Biology (Basel) 2017, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.C.; Wood, S.A.; Kershaw, Y.M.; Bate, E.; Lightman, S.L. Diurnal variation in the responsiveness of the hypothalamic-pituitary-adrenal axis of the male rat to noise stress. J Neuroendocrinol 2006, 18, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Quabbe, H.J.; Gregor, M.; Bumke-Vogt, C.; Hardel, C. Pattern of plasma cortisol during the 24-hour sleep/wake cycle in the rhesus monkey. Endocrinology 1982, 110, 1641–1646. [Google Scholar] [CrossRef]
- Koyanagi, S.; Okazawa, S.; Kuramoto, Y.; Ushijima, K.; Shimeno, H.; Soeda, S.; et al. Chronic treatment with prednisolone represses the circadian oscillation of clock gene expression in mouse peripheral tissues. Mol Endocrinol 2006, 20, 573–583. [Google Scholar] [CrossRef]
- Pezuk, P.; Mohawk, J.A.; Wang, L.A.; Menaker, M. Glucocorticoids as entraining signals for peripheral circadian oscillators. Endocrinology 2012, 153, 4775–4783. [Google Scholar] [CrossRef]
- Le Minh, N.; Damiola, F.; Tronche, F.; Schutz, G.; Schibler, U. Glucocorticoid hormones inhibit food-induced phase-shifting of peripheral circadian oscillators. EMBO J 2001, 20, 7128–7136. [Google Scholar] [CrossRef]
- Bogdan, A.; Bouchareb, B.; Touitou, Y. Ramadan fasting alters endocrine and neuroendocrine circadian patterns. Meal-time as a synchronizer in humans? Life Sci 2001, 68, 1607–1615. [Google Scholar] [CrossRef]
- Salazar, A.; von Hagen, J. Circadian Oscillations in Skin and Their Interconnection with the Cycle of Life. Int J Mol Sci 2023, 24, 5635. [Google Scholar] [CrossRef]
- Palomino-Segura, M.; Hidalgo, A. Circadian immune circuits. J Exp Med 2021, 218. [Google Scholar] [CrossRef]
- Curtis, A.M.; Bellet, M.M.; Sassone-Corsi, P.; O’Neill, L.A. Circadian clock proteins and immunity. Immunity. 2014, 40, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, I.; Dumbell, R.; Oster, H. Circadian Clocks and the Interaction between Stress Axis and Adipose Function. Int J Endocrinol 2015, 2015, 693204. [Google Scholar] [CrossRef] [PubMed]
- Nader, N.; Chrousos, G.P.; Kino, T. Interactions of the circadian CLOCK system and the HPA axis. Trends Endocrinol Metab 2010, 21, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; et al. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol Cell Endocrinol 2013, 380, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Segall, L.A.; Milet, A.; Tronche, F.; Amir, S. Brain glucocorticoid receptors are necessary for the rhythmic expression of the clock protein, PERIOD2, in the central extended amygdala in mice. Neurosci Lett 2009, 457, 58–60. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nakahata, Y.; Tanaka, M.; Yoshida, M.; Soma, H.; Shinohara, K.; et al. Acute physical stress elevates mouse period1 mRNA expression in mouse peripheral tissues via a glucocorticoid-responsive element. J Biol Chem 2005, 280, 42036–42043. [Google Scholar] [CrossRef]
- Gomez-Abellan, P.; Diez-Noguera, A.; Madrid, J.A.; Lujan, J.A.; Ordovas, J.M.; Garaulet, M. Glucocorticoids affect 24 h clock genes expression in human adipose tissue explant cultures. PLoS ONE. 2012, 7, e50435. [Google Scholar] [CrossRef]
- Stimson, R.H.; Mohd-Shukri, N.A.; Bolton, J.L.; Andrew, R.; Reynolds, R.M.; Walker, B.R. The postprandial rise in plasma cortisol in men is mediated by macronutrient-specific stimulation of adrenal and extra-adrenal cortisol production. J Clin Endocrinol Metab 2014, 99, 160–168. [Google Scholar] [CrossRef]
- Mindikoglu, A.L.; Park, J.; Opekun, A.R.; Abdulsada, M.M.; Wilhelm, Z.R.; Jalal, P.K.; et al. Dawn-to-dusk dry fasting induces anti-atherosclerotic, anti-inflammatory, and anti-tumorigenic proteome in peripheral blood mononuclear cells in subjects with metabolic syndrome. Metabol Open 2022, 16, 100214. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; et al. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr Neuropharmacol 2017, 15, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; et al. Melatonin and Human Cardiovascular Disease. J Cardiovasc Pharmacol Ther 2017, 22, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Brown, G.M.; Spence, D.W.; Bharti, V.K.; Kaur, C.; et al. Melatonin antioxidative defense: Therapeutical implications for aging and neurodegenerative processes. Neurotox Res 2013, 23, 267–300. [Google Scholar] [CrossRef]
- Baker, J.; Kimpinski, K. Role of melatonin in blood pressure regulation: An adjunct anti-hypertensive agent. Clin Exp Pharmacol Physiol 2018, 45, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Imenshahidi, M.; Karimi, G.; Hosseinzadeh, H. Effects of melatonin on cardiovascular risk factors and metabolic syndrome: A comprehensive review. Naunyn Schmiedebergs Arch Pharmacol 2020, 393, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.; Afeche, S.C.; Tan, D.X.; Reiter, R.J. Melatonin, energy metabolism, and obesity: A review. J Pineal Res 2014, 56, 371–381. [Google Scholar] [CrossRef]
- Rubio-Sastre, P.; Scheer, F.A.; Gomez-Abellan, P.; Madrid, J.A.; Garaulet, M. Acute melatonin administration in humans impairs glucose tolerance in both the morning and evening. Sleep 2014, 37, 1715–1719. [Google Scholar] [CrossRef]
- Lopez-Minguez, J.; Saxena, R.; Bandin, C.; Scheer, F.A.; Garaulet, M. Late dinner impairs glucose tolerance in MTNR1B risk allele carriers: A randomized, cross-over study. Clin Nutr 2018, 37, 1133–1140. [Google Scholar] [CrossRef]
- Gabel, V.; Reichert, C.F.; Maire, M.; Schmidt, C.; Schlangen, L.J.M.; Kolodyazhniy, V.; et al. Differential impact in young and older individuals of blue-enriched white light on circadian physiology and alertness during sustained wakefulness. Sci Rep 2017, 7, 7620. [Google Scholar] [CrossRef]
- Garaulet, M.; Lopez-Minguez, J.; Dashti, H.S.; Vetter, C.; Hernandez-Martinez, A.M.; Perez-Ayala, M.; et al. Interplay of Dinner Timing and MTNR1B Type 2 Diabetes Risk Variant on Glucose Tolerance and Insulin Secretion: A Randomized Crossover Trial. Diabetes Care 2022, 45, 512–519. [Google Scholar] [CrossRef]
- Carter, M.M.; Olm, M.R.; Merrill, B.D.; Dahan, D.; Tripathi, S.; Spencer, S.P.; et al. Ultra-deep sequencing of Hadza hunter-gatherers recovers vanishing gut microbes. Cell 2023. [Google Scholar] [CrossRef] [PubMed]
- Lotti, S.; Dinu, M.; Colombini, B.; Amedei, A.; Sofi, F. Circadian rhythms, gut microbiota, and diet: Possible implications for health. Nutr Metab Cardiovasc Dis 2023. [Google Scholar] [CrossRef] [PubMed]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front Microbiol 2018, 9, 737. [Google Scholar] [CrossRef]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc Natl Acad Sci USA 2015, 112, 10479–10484. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liu, Y.; Wu, Z.; Wang, J.; Zhang, X. Effects of Diet and Exercise on Circadian Rhythm: Role of Gut Microbiota in Immune and Metabolic Systems. Nutrients 2023, 15, 2743. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab 2014, 20, 1006–1017. [Google Scholar] [CrossRef]
- Chellappa, S.L.; Engen, P.A.; Naqib, A.; Qian, J.; Vujovic, N.; Rahman, N.; et al. Proof-of-principle demonstration of endogenous circadian system and circadian misalignment effects on human oral microbiota. FASEB J 2022, 36, e22043. [Google Scholar] [CrossRef]
- Zeb, F.; Osaili, T.; Obaid, R.S.; Naja, F.; Radwan, H.; Cheikh Ismail, L.; et al. Gut Microbiota and Time-Restricted Feeding/Eating: A Targeted Biomarker and Approach in Precision Nutrition. Nutrients 2023, 15, 259. [Google Scholar] [CrossRef]
- Fonken, L.K.; Workman, J.L.; Walton, J.C.; Weil, Z.M.; Morris, J.S.; Haim, A.; et al. Light at night increases body mass by shifting the time of food intake. Proc Natl Acad Sci USA 2010, 107, 18664–18669. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Thompson, S.V.; Holscher, H.D. Complex interactions of circadian rhythms, eating behaviors, and the gastrointestinal microbiota and their potential impact on health. Nutr Rev 2017, 75, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Wu, L.; Jiang, J.; Yang, T.; Wang, Z.; Ma, L.; et al. Late-Night Eating-Induced Physiological Dysregulation and Circadian Misalignment Are Accompanied by Microbial Dysbiosis. Mol Nutr Food Res 2019, 63, e1900867. [Google Scholar] [CrossRef] [PubMed]
- Bellikci-Koyu, E.; Sarer-Yurekli, B.P.; Akyon, Y.; Ozgen, A.G.; Brinkmann, A.; Nitsche, A.; et al. Associations of sleep quality and night eating behaviour with gut microbiome composition in adults with metabolic syndrome. Proceedings of the Nutrition Society 2021, 80, E58. [Google Scholar] [CrossRef]
- Carasso, S.; Fishman, B.; Lask, L.S.; Shochat, T.; Geva-Zatorsky, N.; Tauber, E. Metagenomic analysis reveals the signature of gut microbiota associated with human chronotypes. FASEB J 2021, 35, e22011. [Google Scholar] [CrossRef]
- Mohammadi, Z.; Bishehsari, F.; Masoudi, S.; Hekmatdoost, A.; Stewart, D.A.; Eghtesad, S.; et al. Association between Sleeping Patterns and Mealtime with Gut Microbiome: A Pilot Study. Arch Iran Med 2022, 25, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Ansu Baidoo, V.; Knutson, K.L. Associations between circadian disruption and cardiometabolic disease risk: A review. Obesity (Silver Spring) 2023, 31, 615–624. [Google Scholar] [CrossRef]
- Ravussin, E.; Beyl, R.A.; Poggiogalle, E.; Hsia, D.S.; Peterson, C.M. Early Time-Restricted Feeding Reduces Appetite and Increases Fat Oxidation But Does Not Affect Energy Expenditure in Humans. Obesity (Silver Spring) 2019, 27, 1244–1254. [Google Scholar] [CrossRef]
- Wilkinson, M.J.; Manoogian, E.N.C.; Zadourian, A.; Lo, H.; Fakhouri, S.; Shoghi, A.; et al. Ten-Hour Time-Restricted Eating Reduces Weight, Blood Pressure, and Atherogenic Lipids in Patients with Metabolic Syndrome. Cell Metab 2020, 31, 92–104.e5. [Google Scholar] [CrossRef]
- O’Connor, S.G.; Boyd, P.; Bailey, C.P.; Shams-White, M.M.; Agurs-Collins, T.; Hall, K.; et al. Perspective: Time-Restricted Eating Compared with Caloric Restriction: Potential Facilitators and Barriers of Long-Term Weight Loss Maintenance. Adv Nutr. 2021, 12, 325–333. [Google Scholar] [CrossRef]
- Welton, S.; Minty, R.; O’Driscoll, T.; Willms, H.; Poirier, D.; Madden, S.; et al. Intermittent fasting and weight loss: Systematic review. Can Fam Physician 2020, 66, 117–125. [Google Scholar]
- Lowe, D.A.; Wu, N.; Rohdin-Bibby, L.; Moore, A.H.; Kelly, N.; En Liu, Y.; et al. Effects of Time-Restricted Eating on Weight Loss and Other Metabolic Parameters in Women and Men With Overweight and Obesity:The TREAT Randomized Clinical Trial. JAMA Intern Med. 2020, 180, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
It illustrates the interaction between chrononutrition and other factors. Chrononutrition, an emerging field of study, focuses on the complex relationship between mealtime, the circadian clock, and metabolism. However, assessing the impact of chrononutrition faces challenges due to the influence of numerous interacting factors. These factors encompass individual chronotype, lifestyle elements such as shift work, sleep disruptions and sleep patterns, dietary composition and portion sizes, levels of physical activity, environmental factors including exposure to nighttime light, age, and genetic factors.
Figure 1.
It illustrates the interaction between chrononutrition and other factors. Chrononutrition, an emerging field of study, focuses on the complex relationship between mealtime, the circadian clock, and metabolism. However, assessing the impact of chrononutrition faces challenges due to the influence of numerous interacting factors. These factors encompass individual chronotype, lifestyle elements such as shift work, sleep disruptions and sleep patterns, dietary composition and portion sizes, levels of physical activity, environmental factors including exposure to nighttime light, age, and genetic factors.


Table 1.
A Summary of Randomized Controlled Trials Investigating the Relationship between Mealtimes, Circadian Rhythm, and Metabolism. Abbreviations: eTRF and mTRF, Early and midday time-restricted feeding; RMR: resting metabolic rate; T wrist: Wrist temperature; EE and LE: early and late eating.
Table 1.
A Summary of Randomized Controlled Trials Investigating the Relationship between Mealtimes, Circadian Rhythm, and Metabolism. Abbreviations: eTRF and mTRF, Early and midday time-restricted feeding; RMR: resting metabolic rate; T wrist: Wrist temperature; EE and LE: early and late eating.
| Authors (year) | Study design | Main results | Comments |
|---|---|---|---|
| Nakamura K. et al. (2021) [109] | A Randomized Cross-Over Trial: To assess the effect of the early evening meal on 24-h blood glucose levels and postprandial lipid metabolism in healthy adults. Twelve participants (2 males and 10 females) participated in a 3-day randomized cross-over study: eating a late dinner (at 21:00) or an early dinner (at 18:00), monitoring blood glucose levels by continuous blood glucose measuring device, metabolic measurements by indirect calorimetry method on the morning of day 3. | Significant differences between the two groups were observed in mean 24-h blood glucose levels on day 2 (p = 0.03). There was a significant decrease in the postprandial respiratory quotient 30 min and 60 min after breakfast on day 3 in the early dinner group compared with the late dinner group (p < 0.05). | Even a small difference 3 h, eating dinner early (at 18:00) has a positive effect on blood glucose level fluctuation and substrate oxidation compared with eating dinner late (at 21:00). No of the subjects is very small studied acute effects. |
| Xie Z. et al. (2022) [113] | Randomized controlled trial: To compare the effects of the two TRF regimens (early and midday) in healthy individuals without obesity. Ninety participants were randomized to eTRF (n=30), mTRF (n=30), or control groups (n=30). Eighty-two participants completed the entire five-week trial and were analyzed (28 in eTRF, 26 in mTRF, 28 in control groups). The primary outcome was the change in insulin resistance. | eTRF was more effective than mTRF at improving insulin sensitivity; eTRF, but not mTRF, improved fasting glucose, reduced total body mass and adiposity, ameliorated inflammation, and increased gut microbial diversity. |
Demonstrates a positive effect of meal timings (from early to midday) on metabolism during TRF regimen, although the study had multiple level biases (participant selection meal plan etc.). |
| Bo S. et al. (2015) [105] | A randomized cross-over study: To assess food-induced thermogenesis in the morning and evening. Twenty subjects were enrolled and randomly received the same standard meal in the morning and, 7 days after, in the evening, or vice versa. A 30-min basal, a further 60-min and 120-min calorimetry was performed after the beginning of the meal. Blood samples were drawn every 30-min for 180-min. General linear models, adjusted for period and carry-over, were used to evaluate the ‘morning effect’, that is, the difference of morning delta (after-meal minus fasting values) minus evening delta (after-meal minus fasting values) of the variables. | Fasting resting metabolic rate (RMR) did not change from morning to evening; after-meal RMR values were significantly higher after the morning meal (1916; 95% confidence interval (CI) = 1792, 2041 vs. 1756; 1648, 1863 kcal; P<0.001). RMR was significantly increased after the morning meal (90.5; 95% CI = 40.4, 140.6 kcal; P<0.001), whereas differences in areas-under-the curve for glucose (−1800; − 2564, − 1036 mg /dlx h, P<0.001), log-insulin (−0.19; − 0.30, − 0.07 μUml /h; P = 0.001) and fatty free acid concentrations (−16.1; − 30.0, − 2.09 mmol/ l xh; P = 0.024) were significantly lower. Delayed and larger increases in glucose and insulin concentrations were found after the evening meals. | The same meal consumed in the evening determined a lower RMR, and increased glycemic/insulinemic responses, suggesting circadian variations in the energy expenditure and metabolic pattern of healthy individuals. Conditions were experimental in terms of food selection and interval. |
| Bandin C. et al. (2015 [104] | A randomized, cross-over trial: Thirty-two women completed two randomized, cross-over protocols: one protocol (P1) includes an assessment of resting energy expenditure (indirect-calorimetry) and glucose tolerance (mixed-meal test) (n = 10), and the other (P2) includes circadian-related measurements based on profiles in salivary cortisol and Wrist temp. (T wrist) (n = 22). In each protocol, participants were provided with standardized meals during the two meal intervention weeks and were studied under two lunch-eating conditions: Early Eating (EE; lunch at 01:00 pm) and Late Eating (LE; lunch at 04:30 pm). | LE, as compared with EE, resulted in decreased pre-meal resting-energy expenditure (P = 0.048), a lower pre-meal protein-corrected respiratory quotient (CRQ) and a changed post-meal profile of CRQ (P = 0.019). These changes reflected a significantly lower pre-meal utilization of carbohydrates in LE versus EE (P = 0.006). LE also increased glucose area under the curve above baseline by 46%, demonstrating decreased glucose tolerance (P = 0.002). Changes in the daily profile of cortisol and T wrist were also found with LE blunting the cortisol profile, with lower morning and afternoon values, and suppressing the postprandial. T wrist peak (P<0.05). |
Eating late is associated with decreased resting-energy expenditure, decreased fasting carbohydrate oxidation, decreased glucose tolerance, blunted daily profile in free cortisol concentrations, and decreased thermal effect of food on T wrist. These results may be implicated in the differential effects of meal timing on metabolic health. |
| Manoogian ENC. et al. (2022) [107] | A randomized control trial: It included 137 firefighters who work 24-h shifts (23-59 years old, 9% female), 12 weeks of 10-h time-restricted eating (TRE) was feasible, with TRE participants decreasing their eating window (baseline, mean 02:13 pm, 95% CI 13.78-14.47 h; intervention, 11:13 am, 95% CI 10:73-11:54 h, p = 3.29E-17) | Compared to the standard of care (SOC) arm, TRE significantly decreased VLDL particle size. In participants with elevated cardiometabolic risks at baseline, there were significant reductions in TRE compared to SOC in glycated hemoglobin A1C and diastolic blood pressure | 10-h time-restricted eating is feasible for shift workers on a 24-h schedule. TRE improved very low-density lipoprotein size and quality of life measures. TRE decreased HbA1c and blood pressure for participants with cardiometabolic risk. A consistent 10-h eating window (TRE) had no adverse effects. However, it had a skewed gender ratio and heterogenous baseline health factors. |
| Qian J. et al. (2018) [111] | Randomized, cross-over design trial: To determine the separate and relative impacts of the circadian system, behavioural/environmental cycles, and their interaction (i.e., circadian misalignment) on insulin sensitivity and β-cell function, the minimal oral model was used to quantitatively assess the major determinants of glucose control in 14 healthy adults using a randomized, cross-over design with two 8-day laboratory protocols. Both protocols involved 3 baseline inpatient days with habitual sleep/wake cycles, followed by 4 inpatient days with the same nocturnal bedtime (circadian alignment) or with 12-hour inverted behavioral/environmental cycles (circadian misalignment) | Data showed that the circadian phase and circadian misalignment affect glucose tolerance through different mechanisms. While the circadian system reduces glucose tolerance in the biological evening compared to the biological morning mainly by decreasing both dynamic and static β-cell responsivity, circadian misalignment reduced glucose tolerance mainly by lowering insulin sensitivity, not by affecting β-cell function. | Results revealed that the endogenous circadian system and circadian misalignment, after controlling for behavioral cycle influences, have independent and differential impacts on insulin sensitivity and β-cell function in healthy adults. |
| Collado MC. et al. (2018) [106] | A randomized, cross-over study: It recruited 10 healthy normal-weight young women to test the effect of the timing of food intake on the human microbiota in the saliva and fecal samples to determine whether eating late alters the daily rhythms of human salivary microbiota; the researchers interrogated salivary microbiota in samples obtained at 4 specific time points over 24 h, to achieve a better understanding of the relationship between food timing and metabolic alterations in humans | A significant diurnal rhythm in salivary diversity and relative bacterial abundance (i.e., TM7 and Fusobacteria) across both early and late eating conditions. Meal timing affected diurnal rhythms in a diversity of salivary microbiota toward an inverted rhythm between the eating conditions, and eating late increased the number of putative pro-inflammatory taxa, showing a diurnal rhythm in the saliva | The impact of the timing of food intake on human salivary microbiota. Eating the main meal late inverts the daily rhythm of salivary microbiota diversity, which may have a deleterious effect on the metabolism of the host. |
| Pizinger T. et al. (2018) [110] | Randomized control trial: It aimed to test the independent and interactive effects of sleep and mealtimes on insulin sensitivity (Si) in overweight adults, Six participants were enrolled (4 men, 2 women; (1 did not complete). Participants underwent a 4-phase randomized cross-over inpatient study differing in sleep times: normal (Ns: mid-night-08:00 am) or late (Ls: 03:30 am -11:30 am); and in meal times: normal (Nm: 1, 5, 11, and 12.5 hours after awakening) or late (Lm: 4.5, 8.5, 14.5, and 16 hours after awakening). An insulin-modified frequently sampled intravenous glucose tolerance test at scheduled breakfast time, and a meal tolerance test, at scheduled lunch time, were performed to assess Si after 3 days in each condition. | Meal time influenced concentrations of glucose (P = 0.012) and insulin (P = 0.069) during the overnight hours Average cortisol concentrations between 2200 and 0700 h tended to be affected by mealtime. Melatonin concentrations from the overnight sampling period showed no effect on mealtime. |
Mealtimes may be relevant for health. |
| Morris C.J et al. (2016) [108] | A randomized cross-over study: The study aimed to test the hypothesis that the endogenous circadian system and circadian misalignment separately affect glucose tolerance in shift workers, both independently from behavioral cycle effects; 9 healthy. The intervention included simulated night work comprised of 12-hour inverted behavioral and environmental cycles (circadian misalignment) or simulated day work (circadian alignment). Postprandial glucose and insulin responses to identical meals given at 8:00 am and 8:00 pm in both protocols | Circadian misalignment increased postprandial glucose by 5.6%, independent of behavioral and circadian effects (P =.0042). | Internal circadian time affects glucose tolerance in shift workers. Separately, circadian misalignment reduces glucose tolerance in shift workers, providing a mechanism to help explain the increased diabetes risk in shift workers. |
| Sharma A. et al. (2017) [112] | Randomized control trial: It aimed to determine the effect of rotational shift work on glucose metabolism. 12 healthy nurses performing rotational shift work using a randomized cross-over study design, underwent an isotope-labeled mixed meal test during a simulated day shift and a simulated night shift, enabling simultaneous measurement of glucose flux and beta cell function using the oral minimal model. | The postprandial glycaemic excursion was higher during the night shift (381±33 vs. 580±48 mmol/l per 5 h, p<0.01). The time to peak insulin, C-peptide, and nadir glucagon suppression in response to meal ingestion was also delayed during the night shift. While insulin action did not differ between study days, the beta cell responsivity to glucose (59±5 vs. 44±4 × 10-9 min-1; p<0.001) and disposition index were decreased during the night shift. | Impaired beta cell function during the night shift may result from normal circadian variation, the effect of rotational shift-work or a combination of both. As a consequence, higher postprandial glucose concentrations are observed during the night shift. |
| Vujovic N. et al. (2022) [126] | A randomized, controlled, cross-over trial with 18 subjects to determine the effects of late versus early eating while rigorously controlling for nutrient intake, physical activity, sleep, and light exposure. The parameters measured were subjective (hunger) and objective (hormones related to metabolism) | Late eating increased hunger (p < 0.0001) and altered appetite-regulating hormones, increasing waketime and 24-h ghrelin leptin ratio (p < 0.0001 and p = 0.006, respectively). Furthermore, late eating decreased waketime energy expenditure (p = 0.002) and 24-h core body temperature (p = 0.019) | These findings show converging mechanisms by which late eating may result in positive energy balance and increased obesity risk. |
| Jamshed H et al. (2019) [71] | This study employed a 4-day randomized crossover design to investigate the impact of time-restricted feeding (TRF) on gene expression, circulating hormones, and diurnal patterns in cardiometabolic risk factors. Eleven overweight adults participated in the study, following two different eating schedules: early TRF (eTRF) from 8 am to 2 pm and a control schedule from 8 am to 8 pm. Continuous glucose monitoring was conducted, and blood samples were collected to assess various factors. | Compared to the control schedule, eTRF led to a significant decrease in mean 24-hour glucose levels by 4 ± 1 mg/dl (p = 0.0003) and glycemic excursions by 12 ± 3 mg/dl (p = 0.001). In the morning, before breakfast, eTRF increased ketones, cholesterol, and the expression of the stress response and aging gene SIRT1, as well as the autophagy gene LC3A (all p < 0.04). In the evening, eTRF showed a tendency to increase brain-derived neurotropic factor (BNDF; p = 0.10) and significantly increased the expression of MTOR (p = 0.007), a key protein involved in nutrient sensing and cell growth. Moreover, eTRF resulted in altered diurnal patterns in cortisol levels and the expression of several circadian clock genes (p < 0.05). | The findings of this study indicate that eTRF improves 24-hour glucose levels, modifies lipid metabolism and circadian clock gene expression, and may enhance autophagy while potentially exerting anti-aging effects in humans. |
| Lowe DA. et a. (2020) [205] | This 12-week randomized clinical trial aimed to investigate the impact of 16:8-hour time-restricted eating (TRE) on weight loss and metabolic risk markers. Participants (n=116) were divided into two groups: the consistent meal timing (CMT) group, instructed to consume three structured meals per day, and the TRE group, instructed to eat ad libitum from 12:00 pm until 8:00 pm and observe a complete caloric intake abstention from 8:00 pm until 12:00 pm the following day. The study was conducted using a custom mobile study application, with in-person testing for a subset of 50 participants located near San Francisco, California. | The TRE group showed a significant decrease in weight (-0.94 kg; 95% CI, -1.68 to -0.20; P = 0.01). However, there was no significant change in the CMT group (-0.68 kg; 95% CI, -1.41 to 0.05, P = 0.07) or between the two groups (-0.26 kg; 95% CI, -1.30 to 0.78; P = 0.63). In the in-person cohort (n = 25 TRE, n = 25 CMT), the TRE group exhibited a significant within-group decrease in weight (-1.70 kg; 95% CI, -2.56 to -0.83; P < 0.001). Additionally, the two groups had a significant difference in the appendicular lean mass index (-0.16 kg/m2; 95% CI, -0.27 to -0.05; P = 0.005). However, no significant changes were observed in any of the other secondary outcomes within or between the groups. Estimated energy intake did not differ significantly between the groups. | These findings suggest that the effectiveness of time-restricted feeding for weight loss may depend on factors such as meal timing. Specifically, skipping the early morning meal may not lead to weight loss benefits. |
| Hutchison AT, et a. (2019) [114] | This study employed a randomized controlled trial design to assess the impact of 9-hour time-restricted feeding (TRF) on glucose tolerance in men at risk for type 2 diabetes. Fifteen male participants with an average age of 55 ± 3 years and a BMI of 33.9 ± 0.8 kg/m2 were enrolled in the study. The participants wore a continuous glucose monitor for 7 days during the baseline assessment and two 7-day TRF conditions. They were randomly assigned to either early TRF (TRFe) from 8 am to 5 pm or delayed TRF (TRFd) from 12 pm to 9 pm, with a 2-week washout phase between the two conditions. Glucose, insulin, triglycerides, nonesterified fatty acids, and gastrointestinal hormone levels were measured and analyzed based on incremental areas under the curve following a standard meal at specific time points. | The results demonstrated that both TRFe and TRFd improved glucose tolerance, as evidenced by a reduction in glucose incremental area under the curve (P = 0.001) and fasting triglycerides (P = 0.003) on day 7 compared to day 0. However, no significant interactions between mealtime and TRF existed for any of the variables examined. TRF did not significantly affect fasting and postprandial insulin, nonesterified fatty acids, or gastrointestinal hormone levels. As measured by continuous glucose monitoring, mean fasting glucose was lower in TRFe (P = 0.02) but not in TRFd (P = 0.17) compared to baseline, with no significant difference observed between the two TRF conditions. | This study’s findings suggest that early and delayed time-restricted feeding (TRFe and TRFd) improve glycemic responses to a test meal in men at risk for type 2 diabetes. Although only TRFe resulted in a lower mean fasting glucose level, reflecting better results with early feeding time. |
| Jones R, et al. (2020) [115] | This study employed a randomized controlled trial to investigate the chronic effects of early time-restricted feeding (eTRF) compared to an energy-matched control on insulin and anabolic sensitivity in healthy males. Sixteen participants with an average age of 23 ± 1 years and a BMI of 24.0 ± 0.6 kg·m-2 were assigned to two groups: eTRF (n = 8) and control/caloric restriction (CON:CR; n = 8). The eTRF group followed the eTRF diet for 2 weeks, while the CON:CR group underwent a calorie-matched control diet after the eTRF intervention. During eTRF, daily energy intake was restricted to the period between 08:00 and 16:00, which extended the overnight fasting period by approximately 5 hours. Metabolic responses were assessed before and after the interventions, following a 12-hour overnight fast, using a carbohydrate/protein drink. | The results showed that eTRF improved whole-body insulin sensitivity compared to CON:CR, with a between-group difference of 1.89 (95% CI: 0.18, 3.60; P = 0.03; η2p = 0.29). eTRF also enhanced skeletal muscle uptake of glucose (between-group difference: 4266 μmol·min-1·kg-1·180 min; 95% CI: 261, 8270; P = 0.04; η2p = 0.31) and branched-chain amino acids (BCAAs) (between-group difference: 266 nmol·min-1·kg-1·180 min; 95% CI: 77, 455; P = 0.01; η2p = 0.44). The eTRF group experienced a reduction in energy intake (approximately 400 kcal·d-1) and weight loss (-1.04 ± 0.25 kg; P = 0.01), which was comparable to the weight loss observed in the CON:CR group (-1.24 ± 0.35 kg; P = 0.01). | Under free-living conditions, eTRF demonstrated improvements in whole-body insulin sensitivity and increased skeletal muscle glucose and BCAA uptake. These metabolic benefits were observed independent of weight loss and reflect chronic adaptations rather than the immediate effects of the last overnight fast, suggesting that eTRF can have positive effects on metabolic health. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



