
Submitted:
28 June 2023
Posted:
29 June 2023
You are already at the latest version
Abstract
Changes in epitranscriptome with N6‐methyladenine (m6A) modification could be involved in the development of multiple diseases, which might be a prevalent modification of messenger RNAs (mRNAs) in eukaryotes. The m6A modification might be performed through the action of methyltransferases, demethylases, and methylation-binding proteins. Importantly, the m6A methylation may be associated with various neurological disorders including Alzheimer's disease (AD), Parkinson's disease (PD), depression, aging related diseases, and/or aging itself. In addition, the m6A methylation might functionally regulate the eukaryotic transcriptome by influencing splicing, export, subcellular localization, translation, stability, and decay of mRNAs. Neurodegenerative diseases may possess a wide variety of phenotypes depending on the neurons that degenerate at occasions. Interestingly, increasing number of evidences have indicated that m6A modification could modulate the expression of autophagy-related genes and promote autophagy in neuronal cells. Oxidative stresses such as reactive oxygen species (ROS) could stimulate the m6A RNA methylation, which may be also related to the regulation of autophagy and/or the development of neurodegenerative diseases. Both m6A modification and autophagy could also play critical roles in regulating health condition of neurons. Therefore, a comprehensive understanding of the m6A and autophagy relationship in human diseases may benefit in developing therapeutic strategies in the future. This paper reviews advances in the understanding of the regulatory mechanisms of m6A modification in the occurrence and development of neurodegenerative diseases and/or aging, discussing the possible therapeutic procedures related to mechanisms of m6A RNA methylation and autophagy.
Keywords:
N6‐methyladenine
; reactive oxygen species
; RNA binding protein
; Alzheimer's disease
; neurodegenerative disease
1. Introduction
Neurodegenerative diseases are interlaced with the disorders of multicellular function and/or interaction in the central nervous system (CNS) [1]. The “neurodegeneration” represents a condition where neuronal cells gradually lose their potential functions, and eventually perish. Alzheimer's disease (AD) is the most common neurodegenerative disease. Neurodegenerative diseases have a huge impact on patients themselves adding to the social economic burden [2], which is regrettably expected to increase worldwide [3]. Now, progresses in resolving the mechanisms behind several neurodegenerative diseases have been considerably recognized. For example, epigenetic modifications and genetic risk factors may be implicated in their appearance, and their mixture could predict or support to identify individuals in significant risks [4]. In general, neurodegenerative diseases could result in motor dysfunctions and behavioral manifestations such as ataxia, and/or dementia [5]. Clarification of pathogenic mechanisms and new targeted drugs have been urgently needed.
More than hundred chemical alterations of RNAs have been found in all types of RNAs [6]. The most notable RNA methylation may account for more than 60 % of all RNAs alterations. Among them, the N6-methyladenosine (m6A) might be a noticeable post-transcriptional RNAs alteration in eukaryotes, which could regulate the expression of various genes [7,8]. It is noteworthy that the m6A is highly enriched in adult brain tissue [9], which might indicate to play a critical role in neurogenesis, neurodevelopment, and/or neurodegenerative disorders [10]. In addition, the profusion of m6A in the brain seems to gradually increase with age from newborns, and mounts in later life [11]. Within the structure of total RNAs, the m6A is mostly dispersed in the 3' untranslated region (UTR), coding sequence of message RNA (mRNA), and/or regions nearby the stop codon [12]. Roles of the m6A alteration in the control of gene expression may be strictly connected to various normal and/or pathological routes containing DNA damage response, cellular differentiation, and/or the occurrence of neurodegenerative diseases. In addition to the regulation of the transcriptional mRNA, m6A modification could also regulate the transcription of a variety of non-coding RNAs (ncRNAs) including microRNAs (miRNAs), circular RNAs (circRNAs), and/or long non-coding RNAs (lncRNAs) [13]. Therefore, m6A alteration could take part in the regulation of various processes in physiological cellular activities including cell development, embryonic development and/or stress responses as well as on a regulatory role of several diseases [14,15]. The motif of m6A during the development may diverge across tissues and/or developmental courses [16], in which the m6A alteration may be inclined to the coding sequence and/or 3′UTR [16]. Remarkably, even a single modification could have a vast effect on the function of RNAs. Interestingly, ncRNAs such as lncRNAs could also regulate the expression of m6A-related proteins [17].
Studies have suggested that m6A modification on mRNAs could influence the proliferation and/or differentiation of neural progenitor cells in a comprehensive understanding of RNA methylation-based diagnosis and/or therapies for neurodegenerative diseases [18,19]. In addition, many studies have also demonstrated the effects of m6A modification in the autophagy mechanisms [20], suggesting that the m6A change might play a critical role in controlling the development of neurodegenerative diseases via the control of autophagy. For example, the m6A modification could instruct direct inhibitory effects on autophagy [21]. Moreover, the m6A alteration could influence the construction of autophagosomes to dysregulate autophagy [22]. Occasionally, the m6A modification could even initiate and/or promote autophagy [23]. Interestingly, the properties of the m6A modification for autophagy may be reliant on disease situation and/or stages. Accordingly, abnormal m6A methylation could modify biological processes and/or regulate various human disorders [24]. Therefore, roles of m6A modifying enzymes might be involved in the pathological process of neurodegenerative diseases, which might be of huge importance to the development of a novel tactic for specific therapy of neurodegenerative diseases. Herein this report, we would go over the main points of the latest studies describing the impact on m6A alteration in neurodegenerative diseases via the regulation of autophagy, and discuss the role of m6A modification-autophagy relationship for the promising treatment of neurodegenerative diseases. (Figure 1)
2. m6A methylation of mRNAs in neurodegenerative diseases
Several studies have shown that m6A methylation could play an important role in the control of the neuron degeneration progression, which might mediate the incidence and/or development of multiple neuron degeneration-related disorders [25]. For example, a quantity of studies have revealed that m6A is aberrantly expressed in the brain of AD patients [26]. It may be beneficial to explore the pathogenesis of AD, which might contribute to discover new biomarkers from the perspective of m6A modification process [27]. As for the process, the m6A is a ubiquitous mRNA modification in eukaryotes, which regularly occurs through the action of methyltransferases, demethylases, and/or methylation-binding proteins. To this point, the m6A methylation of RNAs may be associated with various neurological disorders including not only AD but also Parkinson's disease (PD), Huntington's disease (HD), myasthenia gravis (MG), multiple sclerosis (MS), depression, brain injury, epilepsy, and so forth [28]. Remarkably, it has been suggested that autophagy may be involved in these neurological disorders as well as aging-related diseases [24,28]. Epigenetic modifications including m6A methylation could also play an understandable role in autophagy regulation [29]. For instance, m6A modification has been described to exactly control the expression of autophagy-related genes [30], which might modulate the cellular autophagy levels that may be involved in the development of various diseases including neurodegenerative diseases. (Figure 1) Therefore, the m6A modification is known to play a crucial role in various aging-related diseases and/or aging.
2.1. Alzheimer’s disease
The m6A abnormality is thoroughly related to AD [31], which may be one of the leading reasons of dementia and an increasing health problem worldwide. There is still no cure for AD. AD may be characterized by synaptic loss, hyper-phosphorylation of tau, extracellular plaques of amyloid-beta accumulation, and various levels of neuro-inflammation, as one of neurodegenerative diseases [32]. Several biological processes may be exaggerated by epitranscriptomic modifications, which could control the expression of specific genes via the metabolism of mRNAs. In response to modifications in m6A, the nervous system could become dysfunctional, which might play an important role in the development of AD. In general, the N6-methyladenosine levels are high in the brain [33]. Correspondingly, the expression of multiple m6A methylation regulators could change when AD occurs. In particular, m6A methylation levels are significantly elevated in the cerebral cortex and/or hippocampus of AD model mice, possibly due to the elevated expression of the m6A methyltransferase and/or the decreased expression of the m6A demethylase, which might be associated with the membrane synaptic growth [31,34]. It has been reported that the METTL3 may be downregulated at the hippocampus in human AD samples, and upregulation of METTL3 could promote autophagic p-Tau clearance and ameliorate AD both in vitro and in vivo [35]. In addition, the m6A could control protein levels of key genes involved in AD-associated pathways [33,36].
2.2. Parkinson’s disease
Parkinson’s disease (PD) might be the second most frequent neurodegenerative disease after AD. It has been shown that multiple m6A methylated proteins are associated with PD. The m6A modifications could also regulate dopaminergic signaling. For example, the deletion of METTL14 in the substantia nigra could reduce the level of m6A mRNAs, and might impair the autonomic activity in mice [37]. FTO overexpression and m6A inhibitor cycloleucine could reduce m6A levels in PD models [38]. It has been shown that manganese exposure is a major environmental cause of PD. Remarkably, the FTO overexpression could also diminish the manganese-induced cytotoxicity, thereby could improve the symptoms of PD [39]. Accordingly, it is suggested that some inhibitors of FTO might be utilized for the treatment of PD. For example, entacapone is a catechol-O-methyltransferase inhibitor approved as an adjunctive therapy in combination with levodopa for the PD treatment, which could promote the modification of the target gene FOXO1-m6A by decreasing FTO expression [40].
2.3. Further neurodegenerative disorders and aging
Huntington's disease is an autosomal dominant neurodegenerative disorder, which is characterized by choreiform movements, cognitive deficits, and/or psychiatric symptoms [41]. These impairments may be attributed to hippocampal dysfunction and/or corticostriatal dysfunction [42]. Altered m6A RNAs methylation as a unique hallmark of HD might contribute to hippocampal memory deficits in model mice of Huntington's disease [43]. Huntington's disease is an autoimmune disease of nervous system mainly caused by the autoantibodies against acetylcholine receptor (AChR) in the postsynaptic membranes of neuromuscular junction [44]. It has been suggested that myasthenia gravis may be a complex multifactorial disease involving environmental, genetic, and/or immunological factors. Among them, the m6A regulators might play imperative roles in the pathogenesis of Myasthenia gravis [45]. Multiple sclerosis is a neuro-inflammatory disease that affects the brain, spinal cord, and optic nerves [46]. Unfortunately, there is also no permanent cure for this neurological disorder. It should be agreed at optimal treatment regimens for good quality of life (QOL) [47]. The m6A RNA modifications might be closely associated with the development of multiple sclerosis [48]. It is noteworthy that cellular senescence is an important component of the aging process. It has been suggested that aging could definitely affect m6A RNAs methylation in hippocampal regions [49]. In addition, there are aging-related differences in the modification level of the epigenome m6A [50]. Additionally, the reversibility of the m6A modification in aging may indicate its possibility to delay the aging.
3. ROS, inflammation and m6A mRNAs
The m6A modification is the most dynamic and reversible epigenetic modification of eukaryotic mRNAs [51], which could be regulated by methyltransferases and/or demethylases [52]. Methyltransferases, also called “writers”, may be composed of methyltransferase-like 3 (METTL3), methyltransferase-like 14 (METTL14), and/or wilms tumor 1-associated protein (WTAP), which could catalyze the methylation of N6-adenosine with the function of methyl connection [53]. Among them, the METTL3 has been described to play imperative roles in many pathological processes, predominantly in inflammatory and/or autoimmune responses [53]. m6A-RNAs binding proteins, also called “readers”, may generally contain HNRNPC, YTHDC1/2, IGF2BP1/2/3, and/or YTHDF1/2/3. Reader molecules might be prepared to distinguish the m6A motif, thereby accomplishing the modification of function on the m6A-RNA [54]. Additionally, the m6A modification could be detached by demethylases, also known as “erasers”, including ALKBH5 and/or FTO. (Figure 2)
For example, the half-life of Myh3 mRNA could be significantly decreased after METTL3 deletion, in which Myh3 might have several potential m6A modification sites. Downregulation of METTL3 could reverse the LPS-induced myocardial cell damages, largely by increasing Myh3 mRNA stability [55]. In addition, the deletion of METTL3 could also increase the expression of MyD88, which may prevent the activation of NF-κB signaling in the LPS treated cells [56]. Similarly, the silencing of FTO could suppress the proliferation and/or invasion of cervical cancer cells via the m6A modification of myelocytomatosis oncogene (Myc) and Zinc finger E-box binding homeobox 1 (ZEB1) [57]. The silencing of FTO could also inhibit the NLRP3 mediated IL-1β expression through the modification of the NF-κB signaling [58], which may be related to the protection of cells.
Methylation put in by the “writers” could be reversed by “erasers” [59], which might be controlled for the cellular healthy homeostasis. Therefore, dysregulations of m6A might be linked to the perturbations of cell death and/or proliferation in diverse diseases [60]. Accordingly, m6A modifications are potential to be novel diagnostic and/or therapeutic targets for several diseases including neurodegenerative disease [61]. As mentioned above, the m6A modification may be vigorously controlled by RNA methyltransferases (also known as “writers”), RNA demethylases (“erasers”) and RNA binding proteins (“readers”) [62]. Interestingly, the m6A level of RNAs may be significantly increased with the treatment of LPS [63]. In addition, METTL3 expression might be also upregulated via the increased ROS production [64]. For example, the ROS content might be elevated with the concomitant increase of m6A methylation in the liver after LPS treatment [65]. Therefore, the m6A methylation of RNAs could be modulated under oxidative stresses [65]. Furthermore, ROS could significantly increase the expression of YTHDF2, with the associated elevation of m6A methylation. Gathering these evidence has suggested that the alteration of m6A modification may be a prevalent phenomenon under oxidative stress conditions. Therefore, immunological and/or inflammatory stresses could affect the m6A level of mRNAs [66]. In general, ROS could regulate major epigenetic processes in various cells [67,68]. In particular, the change of ROS levels might contribute to the m6A RNA methylation [69]. Consequently, the m6A methylation may be related to the increased levels of ROS content.
4. m6A RNA modifications in autophagy
Epigenetic modifications such as histone modifications, DNA methylations, and/or these RNA methylations could totally control the expression of several genes involved in autophagy [70]. For example, it is noteworthy that the alteration of ULK1 mRNA expression could be post-transcriptionally altered by the m6A RNA modification, triggering a considerable inhibition of autophagy [71]. Subsequently, many investigations have revealed the effects of m6A alteration in the autophagy related mechanisms [20,72]. The m6A modification could convey inhibitory effects on autophagy, in which the METTL3 might be the key factor involved in the aberrant modification of m6A [21,73]. Sometimes it could promote autophagy instigation [23,74]. In these ways, the m6A modification might play a crucial role in regulating autophagy, which may be dependent on the disease condition. Therefore, comprehension of the m6A methylation machinery in autophagy regulation may be essential for development of therapeutic strategies [22,75].
The YTHDF2 could catch eIF4G1 transcripts with m6A methylation, and may induce the mRNA degradation, thereby promoting autophagy [76]. In addition, the m6A modification could also enhance the stability of ZFAS1 [77]. In more details for neural cells, the lnc RNA of ZFAS1 is upregulated in neural progenitor cells [77], which could regulate the expression of ATG10, and control the autophagy by preventing the PI3K/AKT pathway to stimulate the migration and/or proliferation of neural progenitor cells [77]. Similarly, the YTHDF1 could enhance the translation of ATG2A and ATG14 autophagy-related genes by binding to the m6A-modified mRNA of ATG2A and ATG14, consequently assisting autophagy [78]. The YTHDF1 could also support the translation of the other m6A-modified mRNAs [78]. Therefore, the YTHDF1-knockout may weaken the ability of RNA-binding proteins to recognize m6A, thus impeding mRNA translation, and interrupting downstream molecular functions. In general, autophagy preserves cellular homeostasis, and might support cells to react to various stresses by recycling the impaired organelles, proteins, lipids, and other cellular components. Neuron-protective autophagy could promote the survival of neurons [79]. Now, a huge number of investigations have confirmed that m6A modification could control the initiation and/or activation of autophagy by modifying the expression of ULK1, ATG5, and/or FIP200 [80]. Accordingly, m6A adjustments could control the expression of several genes related to autophagy. Elevated m6A modifications could also stimulate the formation of autophagosome as well as the lysosomal function [81,82]. Therefore, m6A modification and impaired autophagy might be related with the progress of neurodegenerative diseases, in which the m6A-autophagy partnership could play crucial roles.
By enhancing autophagy, cellular senescence and/or differentiation could be meaningfully refreshed [83]. Atypical autophagy may bring about a range of diseases containing neuron degenerative disorders, cerebral ischemic injury, innate immunity diseases, myocardial dysfunction, cardiomyopathy, nonalcoholic fatty liver disease, hypertensive nephropathy, cancers, and various inflammatory diseases [84]. Suppression or up-regulation of autophagy by m6A modification might depend on the level of m6A methylation and/or the role of downstream targets. Interestingly, defective autophagy may be related with the growth of local gram-negative bacteria in the intestine [85]. Although it has been intensely dedicated to the molecular mechanisms involved, further detailed investigations are immediately required to elucidate the exact interaction between m6A modification and autophagy under diverse pathological conditions. The relations between m6A methylation and autophagy might be a remarkable topic in cellular and molecular biology research.
5. Possible therapeutics for neurodegenerative diseases
Several patterns of neuro-degeneration may be restricted from altered pathological mechanisms. For example, AD is considered by the increase of pathologic amyloid and eventually directs to dementia, whereas PD is considered by the loss of dopaminergic neurons, and primarily directs to several extrapyramidal symptoms. As aberrant autophagy has been observed in neurodegenerative diseases, including AD, PD, HD, and/or Amyotrophic Lateral Sclerosis (ALS) [86], something to modify the autophagy might contribute to the development of treatment for these neurodegenerative diseases. In this regards, as gut microbiota can play an imperative role in the response of inflammation, autophagy, oxidative stress, and/or cellular apoptosis even for distant organs from gut [87,88], the gut microbiota might contribute to improve the pathology of neurodegenerative diseases. The comprehensive understanding of the autophagy regulation might help in interpreting their impact on human diseases and may aid in devising future therapeutic strategies.
In general, neurodegenerative diseases are mostly hard to block, and only symptomatic treatments are available. Accordingly, finding an effective treatment is very important. The recognition of m6A mRNA methylation has stretched a newfangled feature in post-transcriptional gene expression machineries [59]. Several animal experiments have also proposed the alteration roles of m6A on various neuronal functions [9,89]. However, the significance of m6A RNAs methylation in cognitive dysfunction might remain mainly unknown. As certain m6A-related regulators might serve as novel therapeutic strategies for neurodegenerative diseases, the organized assessment of m6A alterations could set an important foundation for comprehending the physical characteristics of neurodegenerative diseases including cognition. The evaluation might contribute to guiding even more effective therapeutic strategies for dementia. The link between m6A methylation and aging may also provide novel therapeutic targets with important medical implications against aging. In addition, the comprehension of interplay between m6A modifications and autophagy might support to develop future therapeutic strategies for ultimate anti-aging. In line with this, the m6A modification could have potential as a diagnostic marker and therapeutic target in the treatment of premature ageing and/or age-related diseases [90]. In particular, manipulation of the autophagy pathway may be a promising strategy to improve the development of aging related diseases. In this regards, it is meaningful that small non-coding RNAs (18–24 nucleotides) such as microRNAs (miRNAs) could regulate autophagy in a relation to age-related diseases [91]. miRNAs could also affect multiple genes, which might play diverse important roles. The miRNAs can bind to the 3′UTR of several target genes and promote the degradation of objective genes, thus regulating translation. It would be stimulating to examine the effects of these miRNAs on autophagy in the development of neurodegenerative diseases. For example, the deregulation of definite miRNA could produce a distinctive character for each disorders, and probably lead to a basic diagnosis. Adjusting conventional symptomatic treatments, it has become considerably effective for the conservative therapy in many cases, however, disease-modifying therapies are still requiring. Few have identified inhibitors absolutely targeting m6A regulatory molecules, while gene treatments with genome-editing technology for hereditary disorders have attracted attention.
Interestingly, it has been shown that the modulation of m6A levels could become a novel therapeutic target to deal with the SARS-CoV-2 infection [92]. Previous studies have found that the natural product rhein competitively binds to the FTO active site in vitro [93], which could improve virus-induced lung injury [94]. In this way, the m6A modification could play a key role in the incidence and/or development of several human diseases. However, the mechanisms are not completely clear. In addition, the exact function of each m6A factors might be different during the stages of diseases. Therefore, it might be a challenge for applying m6A technique in various disease-therapies including neurodegenerative diseases.
6. Future perspectives involved in the therapeutics
This research topic directs to explore the therapeutic effect and mechanism action as an integrative medicine for the improvement of patient healthcare of neurodegenerative diseases. The m6A, as the most common posttranscriptional alteration of eukaryotic mRNAs, has been shown as playing significant roles both in gut and/or brain [95]. Interestingly, A. muciniphila and L. plantarum in gut microbiota could affect specific m6A modifications in mice, which may emphasize epitranscriptomic modifications by commensal bacteria [95]. Studies have shown that epigenetics of m6A mRNAs could play a part in the regulation of incidence and/or the development of a variety of diseases including neurodegenerative diseases, in which gut microbiota and RNA epigenetics could construct a complicated cross regulatory network [29,96]. Interestingly, the removal of YTHDF1 could stimulate A. muciniphila colonization to increase anti-inflammatory effects as a feedback system, by promoting the expression of Foxp3 via the m6A modification [97]. Using this concept, it is possible to treat neurodegenerative diseases and/or neurodegenerative disorders with probiotics and/or fecal microbiota transplantation (FMT) [98,99]. (Figure 3) For example, the FMT has a mitigating effect on high fat diet-induced obesity, which may be due to the striking effects of FMT on the microbial composition. Interestingly, the mitigating effect with FMT could also alter intestinal lipid metabolism and/or m6A levels to accomplish the opposition to obesity [100]. Furthermore, we have already suggested that “engram theory” with the concept of improved gut microbiota might become a promising tactics for the treatment of neurodegenerative disorders [99,101]. Accordingly, non-invasive brain stimulation with the alteration of gut microbiota appears to possess much potentials. Intrinsically, emphasizing the latest research results and/or assembling the available evidence have been more imperative than ever. However, more in-depth explorations are required to develop novel therapeutic strategies based on the interaction among m6A methylation, gut microbiota and/or autophagy, in order to support further understanding and ultimately provide new methods for the prevention and/or treatment of neurodegenerative diseases. It would be also mandatory to explore the precise molecular mechanisms of these “theories” for the development of innovative treatments.
Author Contributions
Conceptualization, NS, YI, and SM; original draft preparation and editing, NS, YI, SY, KT, HS, and SM; visualization, NS, YI and SM; supervision, SM. Each author (NS, YI, SY, KT, HS, and SM) has participated sufficiently in this work of drafting the article and/or revising the article for the important rational content. Then, all authors gave final approval of the version to be submitted. Finally, all authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Competing interests statement
The authors declare that they have no competing financial interests.
Abbreviations
AChR: acetylcholine receptor; AD: Alzheimer's disease; ALS: amyotrophic lateral sclerosis; CNS: central nervous system; circRNA: circular RNA; FMT: fecal microbiota transplantation; HD: Huntington's disease; lncRNAs: long non-coding RNAs; m6A: N6-methyladenosine; METTL3: methyltransferase-like 3; METTL14: methyltransferase-like 14; miRNA: microRNA; mRNA: messenger RNA; Myc: myelocytomatosis oncogene; ncRNA: non-coding RNA; MG: myasthenia gravis; MS: multiple sclerosis; ncRNA: non-coding RNA; PD: Parkinson's disease; QOL: quality of life; UTR: untranslated region; WTAP: wilms tumor 1-associated protein; ZEB1: Zinc finger E-box binding homeobox 1
References
- Wang, M.; Pan, W.; Xu, Y.; Zhang, J.; Wan, J.; Jiang, H. Microglia-mediated neuroinflammation: a potential target for the treatment of cardiovascular diseases. J Inflamm Res. 2022, 15, 3083–3094. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martin, P.; Rodriguez-Blazquez, C.; Forjaz, M.J. Quality of life and burden in caregivers for patients with Parkinson’s disease: concepts, assessment and related factors. Expert Rev Pharmacoecon Outcomes Res. 2012, 12, 221–230. [Google Scholar] [CrossRef]
- Brodaty, H.; Breteler, M.M.; DeKosky, S.T.; Dorenlot, P.; Fratiglioni, L.; Hock, C.; Kenigsberg, P.A.; Scheltens, P.; De Strooper, B. The world of dementia beyond 2020. J Am Geriatr Soc. 2011, 59, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Do, M.D.; Tran, T.N.; Luong, A.B.; Le, L.H.G.; Van Le, T.; Le, K.T.; Van Vo, N.T.; Le, T.N.N.; Vu, H.A.; Mai, T.P. Clinical and genetic analysis of Vietnamese patients diagnosed with early-onset Parkinson's disease. Brain Behav. 2023, 13, e2950. [Google Scholar] [CrossRef] [PubMed]
- Naseri, N.; Sharma, M.; Velinov, M. Autosomal dominant neuronal ceroid lipofuscinosis: Clinical features and molecular basis. Clin Genet. 2021, 99, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Cantara, W.A.; Crain, P.F.; Rozenski, J.; McCloskey, J.A.; Harris, K.A.; Zhang, X.; Vendeix, F.A.; Fabris, D.; Agris, P.F. The RNA modification database, RNAMDB: 2011 update. Nucleic Acids Res. 2010, 39 (suppl_1), D195-D201.
- Li, Y.; Xiao, J.; Bai, J.; Tian, Y.; Qu, Y.; Chen, X.; Wang, Q.; Li, X.; Zhang, Y.; Xu, J. Molecular characterization and clinical relevance of m6A regulators across 33 cancer types. J.Mol Cancer. 2019, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, Y.; Wuermanbieke, S.; Hu, S.; Huang, G. N6-methyladenosine (m6A) methyltransferase METTL3-mediated LINC00680 accelerates osteoarthritis through m6A/SIRT1 manner. Cell Death Discov. 2022, 8, 240. [Google Scholar] [CrossRef]
- Weng, Y.-L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H. Epitranscriptomic m6A regulation of axon regeneration in the adult mammalian nervous system. Neuron. 2018, 97, 313–325. [Google Scholar] [CrossRef]
- Widagdo, J.; Anggono, V. The m6A-epitranscriptomic signature in neurobiology: from neurodevelopment to brain plasticity. J Neurochem. 2018, 147, 137–152. [Google Scholar] [CrossRef]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: transcriptome plasticity in brain development and function. Nat Rev Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Ding, X.; Wang, Y.; Hu, X.; Sun, T.; Wei, M.; Wang, X.; Wu, H. The relationship between the network of non-coding RNAs-molecular targets and N6-methyladenosine modification in colorectal cancer. Front Cell Dev Biol. 2021, 9, 772542. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chen, C.; Ji, X.; Liu, J.; Zhou, Q.; Wang, G.; Yuan, W.; Kan, Q.; Sun, Z. The interplay between m6A RNA methylation and noncoding RNA in cancer. J Hematol Oncol. 2019, 12, 1–15. [Google Scholar] [CrossRef]
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat Rev Mol Cell Biol. 2017, 18, 31–42. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, X.; Huang, T.; Zhao, X.; Chen, W.; Gu, N.; Zhang, R. Dynamic landscape and evolution of m6A methylation in human. Nucleic Acids Res. 2020, 48, 6251–6264. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Dong, D.; Yu, P.; Chen, T.; Gao, R.; Wei, J.; Mo, Z.; Zhou, H.; Yang, Q.; Yue, C. Prognostic model based on m6A -associated lncRNAs in esophageal cancer. Front Endocrinol (Lausanne). 2022, 13, 947708. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, B.; Sun, H.; Xu, X.; Wang, Y. Epigenetic regulations in neural stem cells and neurological diseases. Stem Cells Int. 2018, 2018. 6087143.
- Li, J.; Yang, X.; Qi, Z.; Sang, Y.; Liu, Y.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. The role of mRNA m6A methylation in the nervous system. Cell Biosci. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Chen, K.; Zhang, Q.; Li, G.; Zhuo, L.; Zhai, B.; Sui, X.; Hu, X.; Xie, T. The emerging molecular mechanism of m6A modulators in tumorigenesis and cancer progression. Biomed Pharmacother. 2020, 127, 110098. [Google Scholar] [CrossRef]
- Song, H.; Feng, X.; Zhang, H.; Luo, Y.; Huang, J.; Lin, M.; Jin, J.; Ding, X.; Wu, S.; Huang, H. METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy. 2019, 15, 1419–1437. [Google Scholar] [CrossRef]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m6A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy. 2020, 16, 1221–1235. [Google Scholar] [CrossRef]
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L. The mechanism of m6A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by β-elemene. Cell Death Dis. 2020, 11, 969. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Luo, R.; Zhang, W.; He, S.; Wang, B.; Liang, H.; Song, Y.; Ke, W.; Shi, Y.; Feng, X. m6A hypomethylation of DNMT3B regulated by ALKBH5 promotes intervertebral disc degeneration via E4F1 deficiency. Clin Transl Med. 2022, 12, e765. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhan, Y.; Zong, Y. METTL3-mediated LINC00657 promotes osteogenic differentiation of mesenchymal stem cells via miR-144-3p/BMPR1B axis. Cell Tissue Res. 2022, 388, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Zhou, X.; Li, Y.-M.; Yang, T.; Zhang, S.-J.; Wang, Y.; Jia, S.-H.; Peng, D.-T. N6-methyladenine-modified DNA was decreased in Alzheimer’s disease patients. World J Clin Cases. 2022, 10, 448. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Fischer, C.; Maralani, P.J.; Black, S.E.; Moody, A.R.; Khademi, A. Alzheimer’s and vascular disease classification using regional texture biomarkers in FLAIR MRI. Neuroimage Clin. 2023, 38, 103385. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Shi, M.; Li, M.; Zeng, J.; He, J. Roles of m6A modification in neurological diseases. Zhong nan da xue xue bao. Yi xue ban = Journal of Central South University. Medical sciences. 2022, 47, 109–115. [Google Scholar]
- Tang, F.; Chen, L.; Gao, H.; Xiao, D.; Li, X. m6A: An emerging role in programmed cell death. Front Cell Dev Biol. 2022, 10, 80. [Google Scholar] [CrossRef]
- Han, L.; Lei, G.; Chen, Z.; Zhang, Y.; Huang, C.; Chen, W. IGF2BP2 regulates MALAT1 by serving as an N6-methyladenosine reader to promote NSCLC proliferation. Front Mol Biosci. 2022, 8, 1328. [Google Scholar] [CrossRef]
- Han, M.; Liu, Z.; Xu, Y.; Liu, X.; Wang, D.; Li, F.; Wang, Y.; Bi, J. Abnormality of m6A mRNA methylation is involved in Alzheimer’s disease. J.Front Neurosci. 2020, 14, 98. [Google Scholar] [CrossRef]
- Pinheiro, L.; Faustino, C. Therapeutic strategies targeting amyloid-β in Alzheimer’s disease. Curr Alzheimer Res. 2019, 16, 418–452. [Google Scholar] [CrossRef]
- Shafik, A.M.; Zhang, F.; Guo, Z.; Dai, Q.; Pajdzik, K.; Li, Y.; Kang, Y.; Yao, B.; Wu, H.; He, C. N6-methyladenosine dynamics in neurodevelopment and aging, and its potential role in Alzheimer’s disease. Genome Biol. 2021, 22, 1–19. [Google Scholar] [CrossRef]
- Shao, N.; Ye, T.; Xuan, W.; Zhang, M.; Chen, Q.; Liu, J.; Zhou, P.; Song, H.; Cai, B. The effects of N 6-methyladenosine RNA methylation on the nervous system. Mol Cell Biochem. 2023, 1–13. [Google Scholar] [CrossRef]
- Tang, Z.; Cao, J.; Yao, J.; Fan, X.; Zhao, J.; Zhao, M.; Duan, Q.; Han, B.; Duan, S. KDM1A-mediated upregulation of METTL3 ameliorates Alzheimer's disease via enhancing autophagic clearance of p-Tau through m6A-dependent regulation of STUB1. Free Radic Biol Med. 2023, 195, 343–358. [Google Scholar] [CrossRef]
- Hao, X.; Li, Y.; Huang, G.; Zeng, Y. Role of the N6-methyladenosine regulatory factor in reducing the risk of cardiovascular disease: subtype diagnosis following aerobic exercise-assisted weight loss. Am J Transl Res. 2022, 14, 5363. [Google Scholar] [PubMed]
- Teng, Y.; Liu, Z.; Chen, X.; Liu, Y.; Geng, F.; Le, W.; Jiang, H.; Yang, L. Conditional deficiency of m6A methyltransferase Mettl14 in substantia nigra alters dopaminergic neuron function. J Cell Mol Med. 2021, 25, 8567–8572. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, C.; Guo, M.; Zheng, X.; Ali, S.; Huang, H.; Zhang, L.; Wang, S.; Huang, Y.; Qie, S. Down-regulation of m6A mRNA methylation is involved in dopaminergic neuronal death. ACS Chem Neurosci. 2019, 10, 2355–2363. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Wang, S.; Li, J.; Wen, Y.; Cui, R.; Zhang, K.; Liu, Y.; Yang, X.; Zhang, L.; Xu, B. Protective role of mRNA demethylase FTO on axon guidance molecules of nigro-striatal projection system in manganese-induced parkinsonism. J Hazard Mater. 2022, 426, 128099. [Google Scholar] [CrossRef]
- Ondo, K.; Isono, M.; Nakano, M.; Hashiba, S.; Fukami, T.; Nakajima, M. The N6-methyladenosine modification posttranscriptionally regulates hepatic UGT2B7 expression. Biochem Pharmacol. 2021, 189, 114402. [Google Scholar] [CrossRef]
- Foroud, T.; Siemers, E.; Kleindorfer, D.; Bill, D.J.; Hodes, M.; Norton, J.A.; Conneally, P.M.; Christian, J.C. Cognitive scores in carriers of Huntington's disease gene compared to noncarriers. Ann Neurol. 1995, 37, 657–664. [Google Scholar] [CrossRef]
- Harris, K.L.; Armstrong, M.; Swain, R.; Erzinclioglu, S.; Das, T.; Burgess, N.; Barker, R.A.; Mason, S.L. Huntington's disease patients display progressive deficits in hippocampal-dependent cognition during a task of spatial memory. Cortex. 2019, 119, 417–427. [Google Scholar] [CrossRef]
- Pupak, A.; Singh, A.; Sancho-Balsells, A.; Alcalá-Vida, R.; Espina, M.; Giralt, A.; Martí, E.; Ørom, U.A.V.; Ginés, S.; Brito, V. Altered m6A RNA methylation contributes to hippocampal memory deficits in Huntington’s disease mice. Cell Mol Life Sci. 2022, 79, 416. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.M.; Van Den Broeck, J.; Vrolix, K.; Janssen, S.P.; Lemmens, M.A.; Van Der Esch, E.; Duimel, H.; Frederik, P.; Molenaar, P.C.; Martínez-Martínez, P. Antibody effector mechanisms in myasthenia gravis—pathogenesis at the neuromuscular junction. Autoimmunity. 2010, 43 (5-6), 353-370.
- Li, S.; Liu, H.; Ruan, Z.; Guo, R.; Sun, C.; Tang, Y.; Huang, X.; Gao, T.; Hao, S.; Li, H. Landscape analysis of m6A modification regulators related biological functions and immune characteristics in myasthenia gravis. J Transl Med. 2023, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Dalmau, J. Diagnostic and treatment challenges, a new section in N2. Neurol Neuroimmunol Neuroinflamm. 2018, 5, e511. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.-B.; Lei, S.-F.; Qian, Q.-Y.; Guo, Y.-F.; Zhang, Y.-H.; Zhang, H. Integrative analysis revealed potential causal genetic and epigenetic factors for multiple sclerosis. J Neurol. 2019, 266, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Castro-Hernández, R.; Berulava, T.; Metelova, M.; Epple, R.; Peña Centeno, T.; Richter, J.; Kaurani, L.; Pradhan, R.; Sakib, M.S.; Burkhardt, S. Conserved reduction of m6A RNA modifications during aging and neurodegeneration is linked to changes in synaptic transcripts. Proc Natl Acad Sci U S A. 2023, 120, e2204933120. [Google Scholar] [CrossRef]
- Su, X.; Shen, Y.; Jin, Y.; Kim, I.-m.; Weintraub, N.L.; Tang, Y. Aging-associated differences in epitranscriptomic m6A regulation in response to acute cardiac ischemia/reperfusion injury in female mice. Front Pharmacol. 2021, 12, 654316. [Google Scholar] [CrossRef]
- Chen, Y.-s.; Ouyang, X.-p.; Yu, X.-h.; Novák, P.; Zhou, L.; He, P.-p.; Yin, K. N6-Adenosine methylation (m6A) RNA modification: an emerging role in cardiovascular diseases. J Cardiovasc Transl Res. 2021, 14, 857–872. [Google Scholar] [CrossRef]
- Shi, H.; Wei, J.; He, C. Where, when, and how: context-dependent functions of RNA methylation writers, readers, and erasers. Mol Cell. 2019, 74, 640–650. [Google Scholar] [CrossRef]
- Luo, J.; Xu, T.; Sun, K. N6-methyladenosine RNA modification in inflammation: roles, mechanisms, and applications. Front Cell Dev Biol. 2021, 9, 670711. [Google Scholar] [CrossRef]
- Mapperley, C.; van de Lagemaat, L.N.; Lawson, H.; Tavosanis, A.; Paris, J.; Campos, J.; Wotherspoon, D.; Durko, J.; Sarapuu, A.; Choe, J. The mRNA m6A reader YTHDF2 suppresses proinflammatory pathways and sustains hematopoietic stem cell function. J Exp Med. 2020, 218, e20200829. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Wu, J.; Li, H.; Luo, C.; Ji, G.; Guan, X.; Liu, J.; Wang, M. METTL3 achieves lipopolysaccharide-induced myocardial injury via m6A -dependent stabilization of Myh3 mRNA. Biochim Biophys Acta Mol Cell Res. 2023, 1870, 119503. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Li, Q.; Meng, R.; Yi, B.; Xu, Q. METTL 3 regulates alternative splicing of MyD88 upon the lipopolysaccharide-induced inflammatory response in human dental pulp cells. J Cell Mol Med. 2018, 22, 2558–2568. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Jin, C.; Wang, Y.; Yu, J.; Wang, R.; Tian, X. FTO promotes the progression of cervical cancer by regulating the N6-methyladenosine modification of ZEB1 and Myc. Mol Carcinog. 2023. [Google Scholar] [CrossRef]
- Luo, J.; Wang, F.; Sun, F.; Yue, T.; Zhou, Q.; Yang, C.; Rong, S.; Yang, P.; Xiong, F.; Yu, Q. Targeted inhibition of FTO demethylase protects mice against LPS-induced septic shock by suppressing NLRP3 inflammasome. Front Immunol. 2021, 12, 663295. [Google Scholar] [CrossRef]
- Yue, Y.; Liu, J.; He, C. RNA N6-methyladenosine methylation in post-transcriptional gene expression regulation. Genes Dev. 2015, 29, 1343–1355. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA modifications in gene expression regulation. Cell. 2017, 169, 1187–1200. [Google Scholar] [CrossRef]
- Deng, Y.; Zhu, H.; Xiao, L.; Liu, C.; Liu, Y.-L.; Gao, W. Identification of the function and mechanism of m6A reader IGF2BP2 in Alzheimer’s disease. Aging (Albany NY). 2021, 13, 24086. [Google Scholar] [CrossRef]
- Yang, Y.; Hsu, P.J.; Chen, Y.-S.; Yang, Y.-G. Dynamic transcriptomic m6A decoration: writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, Y.; Ma, J.; Yu, Y.; Wang, Q.; Gao, P.; Wang, L.; Xu, Z.; Wei, X.; Jing, M. m6A mRNA Methylation Was Associated With Gene Expression and Lipid Metabolism in Liver of Broilers Under Lipopolysaccharide Stimulation. Front Genet. 2022, 13. 818357.
- Qi, L.; Wang, Y.; Hu, H.; Li, P.; Hu, H.; Li, Y.; Wang, K.; Zhao, Y.; Feng, M.; Lyu, H. m6A methyltransferase METTL3 participated in sympathetic neural remodeling post-MI via the TRAF6/NF-κB pathway and ROS production. J Mol Cell Cardiol. 2022, 170, 87–99. [Google Scholar] [CrossRef]
- Zhong, X.; Yu, J.; Frazier, K.; Weng, X.; Li, Y.; Cham, C.M.; Dolan, K.; Zhu, X.; Hubert, N.; Tao, Y. Circadian clock regulation of hepatic lipid metabolism by modulation of m6A mRNA methylation. Cell Rep. 2018, 25, 1816–1828. [Google Scholar] [CrossRef]
- Han, Y.-C.; Xie, H.-Z.; Lu, B.; Xiang, R.-L.; Zhang, H.-P.; Li, J.-Y.; Zhang, S.-Y. Lipopolysaccharide alters the m6A epitranscriptomic tagging of RNAs in cardiac tissue. Front Mol Biosci. 2021, 8, 670160. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, Y.; Chen, Z.; Yue, Y.; Huang, H.; Wu, B.; Liu, Y.; Zhou, D.-X.; Zhao, Y. ROS-stimulated protein lysine acetylation is required for crown root development in rice. J Adv Res. 2023, 48, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ni, X. ROS-mediated DNA methylation pattern alterations in carcinogenesis. Curr Drug Targets. 2015, 16, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhuo, R.; Tao, S.; Liang, Y.; Liu, C.; Liu, Q.; Wang, T.; Zhong, X. m6A RNA Methylation Mediates NOD1/NF-kB Signaling Activation in the Liver of Piglets Challenged with Lipopolysaccharide. Antioxidants (Basel). 2022, 11, 1954. [Google Scholar] [CrossRef]
- Shu, F.; Xiao, H.; Li, Q.-N.; Ren, X.-S.; Liu, Z.-G.; Hu, B.-W.; Wang, H.-S.; Wang, H.; Jiang, G.-M. Epigenetic and post-translational modifications in autophagy: biological functions and therapeutic targets. Signal Transduct Target Ther. 2023, 8, 32. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, X.; Miao, Y.; Liang, P.; Zhu, K.; She, Y.; Wu, Y.; Liu, D.-A.; Huang, J.; Ren, J. , m6A RNA modification controls autophagy through upregulating ULK1 protein abundance. Cell Res. 2018, 28, 955–957. [Google Scholar] [CrossRef]
- Liu, J.; Shao, Y.; Li, D.; Li, C. N6-methyladenosine helps Apostichopus japonicus resist Vibrio splendidus infection by targeting coelomocyte autophagy via the AjULK-AjYTHDF/AjEEF-1α axis. Commun Biol. 2023, 6, 547. [Google Scholar] [CrossRef]
- Yu, T.; Qi, X.; Zhang, L.; Ning, W.; Gao, D.; Xu, T.; Ma, Y.; Knott, J.G.; Sathanawongs, A.; Cao, Z. Dynamic reprogramming and function of RNA N6-methyladenosine modification during porcine early embryonic development. Zygote. 2021, 29, 417–426. [Google Scholar] [CrossRef]
- Fang, Z.M.; Zhang, S.M.; Luo, H.; Jiang, D.S.; Huo, B.; Zhong, X.; Feng, X.; Cheng, W.; Chen, Y.; Feng, G. Methyltransferase-like 3 suppresses phenotypic switching of vascular smooth muscle cells by activating autophagosome formation. Cell Prolif. 2023, 56, e13386. [Google Scholar] [CrossRef]
- Liang, J.; Sun, J.; Zhang, W.; Wang, X.; Xu, Y.; Peng, Y.; Zhang, L.; Xiong, W.; Liu, Y.; Liu, H. Novel Insights into The Roles of N6-methyladenosine (m6A) Modification and Autophagy in Human Diseases. Int J Biol Sci. 2023, 19, 705. [Google Scholar] [CrossRef]
- Wang, F.; Liao, Y.; Zhang, M.; Zhu, Y.; Wang, W.; Cai, H.; Liang, J.; Song, F.; Hou, C.; Huang, S. N6-methyladenosine demethyltransferase FTO-mediated autophagy in malignant development of oral squamous cell carcinoma. Oncogene. 2021, 40, 3885–3898. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, S.; Zhang, Y.; Zhao, M. Rat bone marrow mesenchymal stem cells (BMSCs) inhibit liver fibrosis by activating GSK3β and inhibiting the Wnt3a/β-catenin pathway. Infect Agent Cancer. 2022, 17, 1–10. [Google Scholar] [CrossRef]
- Huang, P.; Liu, M.; Zhang, J.; Zhong, X.; Zhong, C. The Potential Role of m6A in the Regulation of TBI-Induced BGA Dysfunction. Antioxidants (Basel). 2022, 11, 1521. [Google Scholar] [CrossRef]
- Yu, H.; Zhong, H.; Sun, J.; Li, N.; Chen, J.; Shen, B.; Huang, P.; Shen, X.; Huang, S.; Zhong, Y. Molecular signaling from microglia impacts macroglia autophagy and neurons survival in glaucoma. iScience. 2023, 26. 106839.
- Chen, S.-j.; Bao, L.; Keefer, K.; Shanmughapriya, S.; Chen, L.; Lee, J.; Wang, J.; Zhang, X.-Q.; Hirschler-Laszkiewicz, I. ; Merali, S Transient receptor potential ion channel TRPM2 promotes AML proliferation and survival through modulation of mitochondrial function, ROS, and autophagy. Cell Death Dis. 2020, 11, 247. [Google Scholar] [CrossRef]
- Hao, W.; Dian, M.; Zhou, Y.; Zhong, Q.; Pang, W.; Li, Z.; Zhao, Y.; Ma, J.; Lin, X.; Luo, R. Autophagy induction promoted by m6A reader YTHDF3 through translation upregulation of FOXO3 mRNA. Nat Commun. 2022, 13, 5845. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Dian, M.; Wang, J.; Sun, Y.; Xiao, D. Epitranscriptomic turbo for autophagy boost: m6A reader YTHDF3. Autophagy. 2022, 19, 1882–1884. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wang, J.; Wang, Q.; Liu, P.; Zhao, H. , Interaction between N6-methyladenosine and autophagy in the regulation of bone and tissue degeneration. Front Bioeng Biotechnol. 2022, 10, 978283. [Google Scholar] [CrossRef] [PubMed]
- Almasi, S.; Crawford Parks, T.E.; Ravel-Chapuis, A.; MacKenzie, A.; Côté, J.; Cowan, K.N.; Jasmin, B.J. Differential regulation of autophagy by STAU1 in alveolar rhabdomyosarcoma and non-transformed skeletal muscle cells. Cell Oncol (Dordr). 2021, 44, 851–870. [Google Scholar] [CrossRef]
- Singh, S.B.; Lin, H.C. Autophagy counters LPS-mediated suppression of lysozyme. Innate Immun. 2017, 23, 537–545. [Google Scholar] [CrossRef]
- Sánchez-Vidaña, D.I.; Li, J.; Abokyi, S.; Chan, J.N.-M.; Ngai, S.P.-C.; Lau, B.W.-M. In vitro methods in autophagy research: Applications in neurodegenerative diseases and mood disorders. Front Mol Neurosci. 2023, 16, 130. [Google Scholar] [CrossRef]
- Nagase, N.; Ikeda, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Efficacy of probiotics on the modulation of gut microbiota in the treatment of diabetic nephropathy. World J Diabetes. 2022, 13, 150. [Google Scholar] [CrossRef]
- Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Tsuji, A.; Matsuda, S. Promising role of D-amino acids in irritable bowel syndrome. World J Gastroenterol. 2022, 28, 4471. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jang, S. RNA m6A methyltransferase Mettl3 regulates spatial neural patterning in Xenopus laevis. Mol Cell Biol. 2021, 41, e00104–e00121. [Google Scholar] [CrossRef]
- Wu, Z.; Shi, Y.; Lu, M.; Song, M.; Yu, Z.; Wang, J.; Wang, S.; Ren, J.; Yang, Y.-G.; Liu, G.-H. METTL3 counteracts premature aging via m6A-dependent stabilization of MIS12 mRNA. Nucleic Acids Res. 2020, 48, 11083–11096. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Huang, T.; Liu, Y.; Fu, J.; Wei, X.; Liu, D.; Ma, W.; Gu, H.; Yuan, Z. Nuclear factor IC disrupts cellular homeostasis between autophagy and apoptosis via miR-200b-Ambra1 in neural tube defects. Cell Death Dis. 2021, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Zannella, C.; Rinaldi, L.; Boccia, G.; Chianese, A.; Sasso, F.C.; De Caro, F.; Franci, G.; Galdiero, M. Regulation of m6A methylation as a new therapeutic option against COVID-19. Pharmaceuticals (Basel). 2021, 14, 1135. [Google Scholar] [CrossRef]
- Chen, B.; Ye, F.; Yu, L.; Jia, G.; Huang, X.; Zhang, X.; Peng, S.; Chen, K.; Wang, M.; Gong, S.; et al. Development of cell-active N6-methyladenosine RNA demethylase FTO inhibitor. J Am Chem Soc. 2012, 134, 17963–17971. [Google Scholar] [CrossRef]
- Shen, C.; Zhang, Z.; Xie, T.; Ji, J.; Xu, J.; Lin, L.; Yan, J.; Kang, A.; Dai, Q.; Dong, Y. Rhein suppresses lung inflammatory injury induced by human respiratory syncytial virus through inhibiting NLRP3 inflammasome activation via NF-κB pathway in mice. Front Pharmacol. 2020, 10, 1600. [Google Scholar] [CrossRef]
- Jabs, S.; Biton, A.; Bécavin, C.; Nahori, M.-A.; Ghozlane, A.; Pagliuso, A.; Spanò, G.; Guérineau, V.; Touboul, D. ; Giai Gianetto, Q Impact of the gut microbiota on the m6A epitranscriptome of mouse cecum and liver. Nat Commun. 2020, 11, 1344. [Google Scholar] [CrossRef]
- Koh, Y.-C.; Ho, C.-T.; Pan, M.-H. , Recent advances in cancer chemoprevention with phytochemicals. J Food Drug Anal. 2020, 28, 14–37. [Google Scholar] [CrossRef] [PubMed]
- Medina-Larqué, A.-S.; Rodríguez-Daza, M.-C.; Roquim, M.; Dudonné, S.; Pilon, G.; Levy, É.; Marette, A.; Roy, D.; Jacques, H.; Desjardins, Y. Cranberry polyphenols and agave agavins impact gut immune response and microbiota composition while improving gut barrier function, inflammation, and glucose metabolism in mice fed an obesogenic diet. Front Immunol. 2022, 13, 871080. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, A.; Yoshikawa, S.; Ikeda, Y.; Taniguchi, K.; Sawamura, H.; Morikawa, S.; Nakashima, M.; Asai, T.; Matsuda, S. Tactics with prebiotics for the treatment of metabolic dysfunction-associated fatty liver disease via the improvement of mitophagy. Int J Mol Sci. 2023, 24, 5465. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. A New Concept of Associations between Gut Microbiota, Immunity and Central Nervous System for the Innovative Treatment of Neurodegenerative Disorders. Metabolites. 2022, 12, 1052. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Fu, J.; Xiao, X.; Wang, F.; Jin, M.; Fang, W.; Wang, Y.; Zong, X. Faecal microbiota transplantation-mediated jejunal microbiota changes halt high-fat diet-induced obesity in mice via retarding intestinal fat absorption. Microb Biotechnol. 2022, 15, 337–352. [Google Scholar] [CrossRef]
- Sawamura, H.; Taniguchi, K.; Ikeda, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. , Gut microbiota could modulate the effects of neuro-immune responses and memory traces via the gut-brain-immune axis in schizophrenia. Explor. Neuroprot. Ther. 2022, 2, 74–86. [Google Scholar] [CrossRef]
Figure 1.
Illustration of the relation of m6A RNAs methylation to various neurodegenerative diseases. Roles of m6A RNAs methylation with autophagy have been proposed in several neurodegenerative diseases including Alzheimer’s disease, Parkinson’s disease, Huntington's disease, myasthenia gravis, multiple sclerosis as well as in aging-related diseases and aging. Consequently, m6A RNAs methylation could be diagnostic and/or therapeutic tools for these diseases.
Figure 1.
Illustration of the relation of m6A RNAs methylation to various neurodegenerative diseases. Roles of m6A RNAs methylation with autophagy have been proposed in several neurodegenerative diseases including Alzheimer’s disease, Parkinson’s disease, Huntington's disease, myasthenia gravis, multiple sclerosis as well as in aging-related diseases and aging. Consequently, m6A RNAs methylation could be diagnostic and/or therapeutic tools for these diseases.
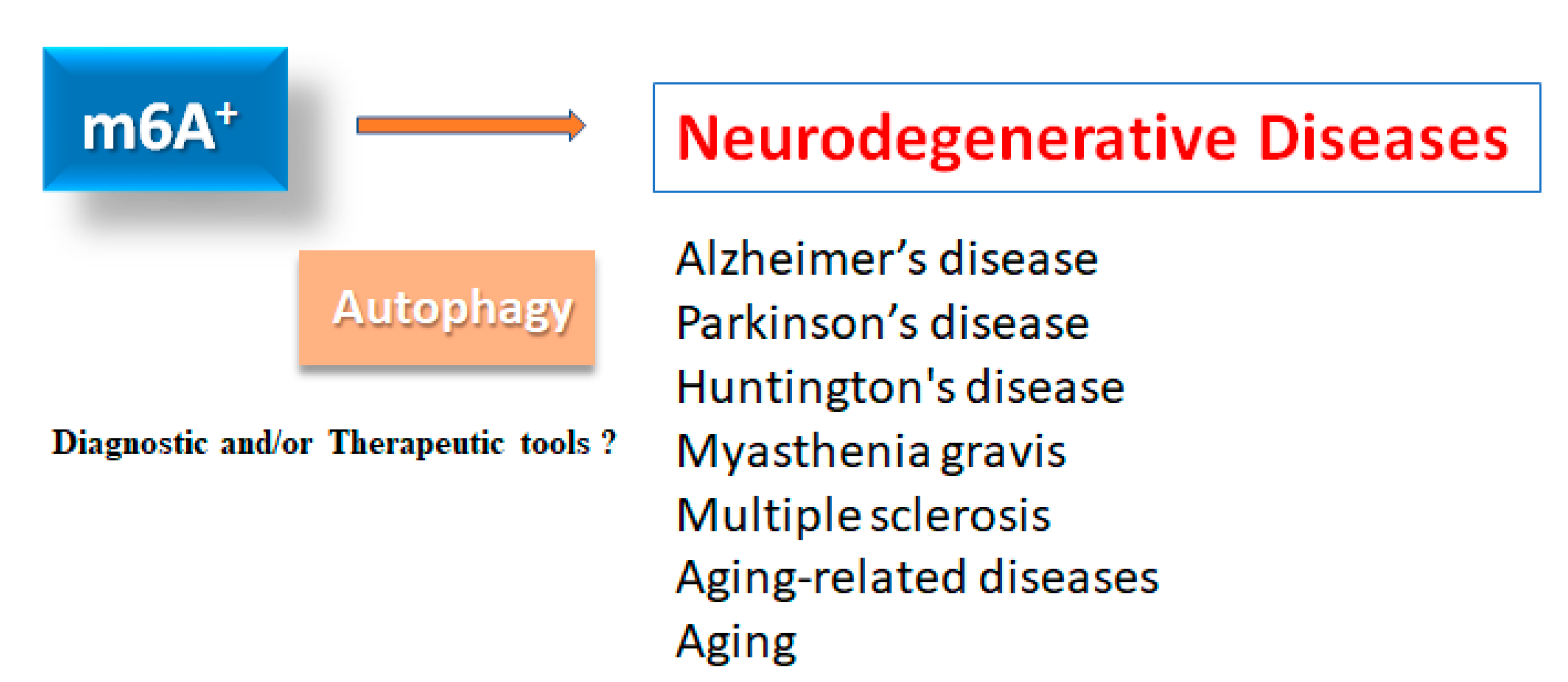
Figure 2.
Schematic representation of the players involved in the m6A RNAs methylation. The m6A modification is regulated by methyltransferases “writers” and demethylases “erasers”. The m6A-RNAs binding proteins are called “readers”. Example molecules are also shown for each players. Genetic factors and/or inflammation with ROS may affect the function of these players. The arrowhead means stimulation, and hammerhead represents inhibition. Note that some critical pathways have been omitted for clarity.
Figure 2.
Schematic representation of the players involved in the m6A RNAs methylation. The m6A modification is regulated by methyltransferases “writers” and demethylases “erasers”. The m6A-RNAs binding proteins are called “readers”. Example molecules are also shown for each players. Genetic factors and/or inflammation with ROS may affect the function of these players. The arrowhead means stimulation, and hammerhead represents inhibition. Note that some critical pathways have been omitted for clarity.
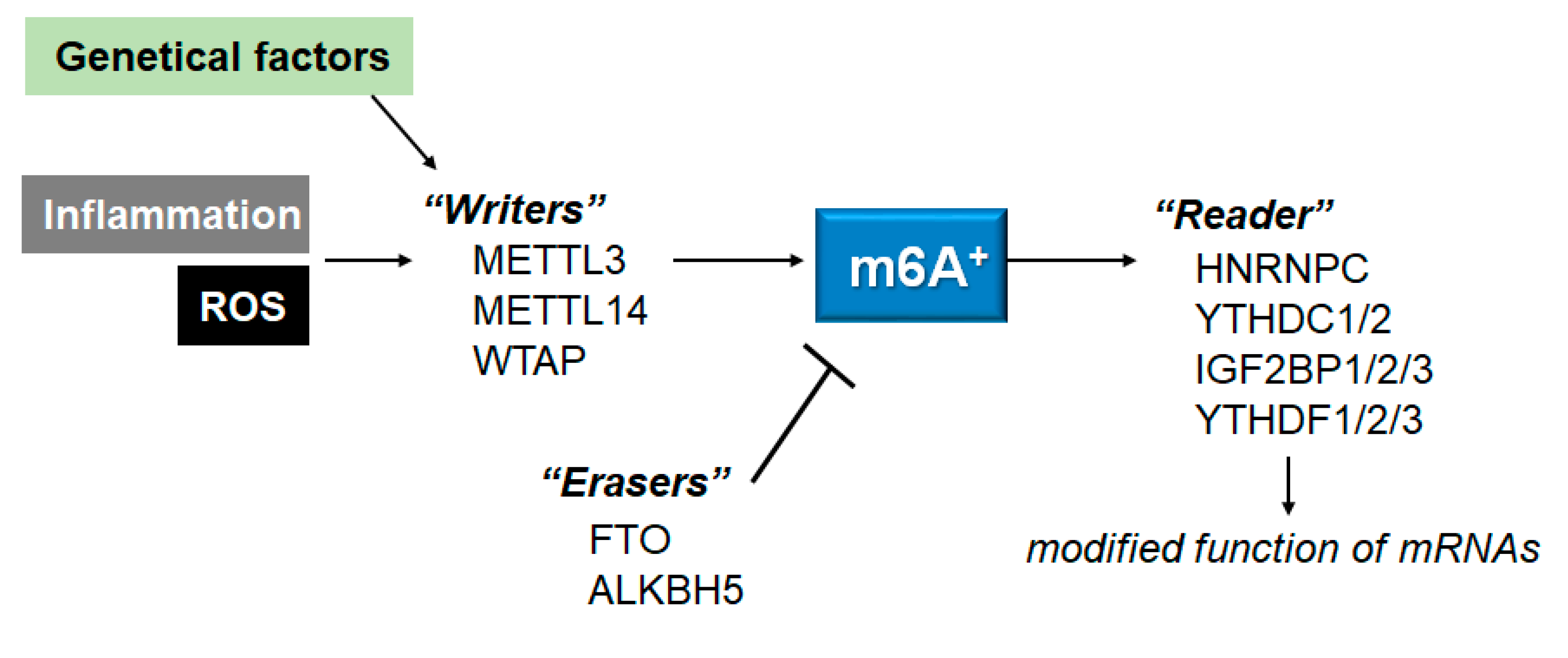
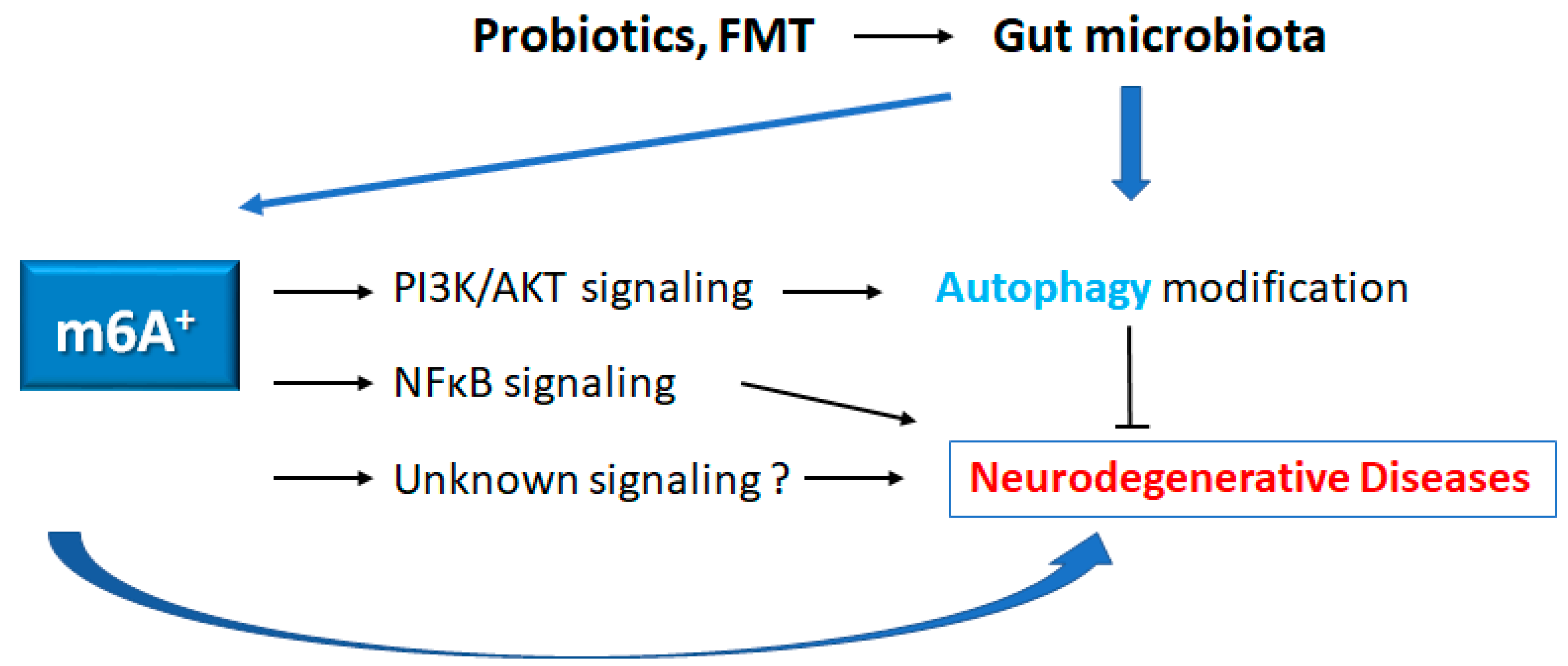
Figure 3.
Schematic representation of the possible inhibitory tactics against the pathogenesis of neurodegenerative diseases. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) could contribute to the alteration of gut microbial community for the alteration of autophagy and/or m6A RNAs methylation, which might be beneficial for the treatment of neurodegenerative diseases. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several important activities such as cytokine-induction, ROS production, and/or inflammatory reactions have been omitted for clarity.
Figure 3.
Schematic representation of the possible inhibitory tactics against the pathogenesis of neurodegenerative diseases. Some kinds of probiotics and/or fecal microbiota transplantation (FMT) could contribute to the alteration of gut microbial community for the alteration of autophagy and/or m6A RNAs methylation, which might be beneficial for the treatment of neurodegenerative diseases. Arrowhead indicates stimulation whereas hammerhead shows inhibition. Note that several important activities such as cytokine-induction, ROS production, and/or inflammatory reactions have been omitted for clarity.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



