
Submitted:
28 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Neurodegenerative diseases (NDs) affect millions of people worldwide, and to date Alzheimer's and Parkinson's disease are the most common NDs. The risk of developing NDs increases dra-matically with age, and genetic and environmental factors can be crucial in their progression. The main feature of NDs is the abnormal presence of specific proteins that can lead to neuronal death, proteotoxic stress, and alteration of catabolic systems, which are major players in the processes of neurodegeneration and neuroinflammation. Despite research efforts to date, the therapies cur-rently in use for the treatment of NDs provide marginal therapeutic benefit, so the focus is now being direct toward the search of natural products that could be a valuable aid in combating these diseases, including the natural compound Andrographis paniculata (Ap) and especially its main constituent Andrographolide (Andro). Preclinical studies have shown that the aqueous extract of Ap is capable to modulate neuroinflammatory and neurodegenerative responses, determining a reduction in inflammatory markers and oxidative stress in various NDs. Therefore, in this review we will focus on the molecular mechanisms through which Ap and Andro are able to modulate the processes of neurodegeneration and neuroinflammation that are major causes of neuronal death and cognitive decline.
Keywords:
Andrographis paniculata
; Andrographolide
; Neurodegenerative Diseases
; Alzheimer
; Parkinson
; Brain Ischemia
; Neuroinflammation
1. Introduction
Neurodegenerative diseases (ND) have epidemiological and symptomatological specifics characteristics and different laboratory and neuroimaging tests; furthermore, the neuropathological mechanisms and treatment are also different. However, all neurodegenerative diseases can cause morbidity and reduced cognitive capacity in elderly individuals worldwide [1,2]. Alzheimer’s disease (AD) accounts for almost 80% of all dementia cases and [3,4,5] it is estimated that by 2050 the number of sufferers will reach 106.8 million, of which 16.51 million will be Europeans [6]. The incidence rate increases with age [4] and the annual risk reaches 6% in individuals over 85 years of age. Idiopathic Parkinson’s disease (PD) is the second most common neurodegenerative disease after AD. The incidence rate of Parkinson’s disease in Europe is 11-19/100 000 individuals per year [7] and the average age of onset is 60 years [8]. In addition, the aetiology of the disease in most patients is unknown, although some genes implicated in the pathogenesis of familial forms have been identified, but these only account for 5%-15% of cases. Pathologically, PD is characterised by the loss of dopaminergic neurons in the pars compacta of the substantia nigra and the accumulation of misfolded α-synuclein, which is found in intra-cytoplasmic inclusions called Lewy bodies. Dementia with Lewy bodies (DLB) and Parkinson’s disease have very overlapping features including progressive cognitive impairment, behavioural disturbances, and dyskinesia [9]. These two diseases differ, however, in the timing of onset of motor symptoms compared to cognitive symptoms. In Parkinson’s, in fact, the motor syndrome precedes the cognitive disorders [10,11]. However, the prevalence of neurodegenerative disorders is increasing overall, partly due to longer life spans but today there is no cure for any of these diseases despite ongoing research efforts [12]. In fact, neurodegenerative diseases represent an ongoing public health challenge, and the identification of effective preventive measures and disease-modifying treatments are a current necessity [13].
2. Molecular mechanisms involved in the onset of neurodegenerative diseases
Neurodegenerative diseases differ in the accumulation of specific proteins and in the different anatomical area in which the pathological neuronal variation is observed, but they share many of the pathological mechanisms that lead to neuronal dysfunction and death, such as proteotoxic stress and abnormalities in the resulting catabolic systems. Indeed, in neurodegenerative diseases, the ubiquitin-proteasome system and the autophagy/lysosomal system are found to be altered. In addition, oxidative stress, apoptotic processes, and neuroinflammation play a key role in all neurodegenerative diseases [14]. The first evidence of an ongoing inflammatory phenomenon in Alzheimer’s disease (AD) was described many years ago [15], and a large number of subsequent studies have documented the presence of an active inflammatory process also in Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS), multiple sclerosis (MS), and a growing number of other nervous system diseases. In these diseases, it appears that inflammation is not the trigger for the disease process but, as revealed by studies in animal models, an inflammatory response such that microglia and astrocytes are involved may contribute to the progression of the disease process. Therefore, the hypothesis that inhibition of the inflammatory response may be a viable strategy to reverse or slow the course of the disease appears to be well-founded. In the brain, the immune response is carried out mainly by microglia, which in turn influence neurons, astrocytes, and glial cells that perform a supporting function. Microglia normally become activated in the production of inflammatory and neurotrophic factors only under pathological conditions, with the purpose of initiating an inflammatory process with activating action on the immune system to repair tissue damage induced by, for example, an injury or microorganism. However, a sustained inflammatory state for a long time indicates persistence of the inflammatory stimulus and failure to resolve the damage. Thus, the inflammatory response that physiologically represents the initiation of a beneficial process for tissues to ensure, for example, the removal of cellular debris can, under certain conditions, turn into a risk factor. Prolonged and uncontrolled inflammation can lead to the production of neurotoxic factors capable of inducing significant tissue pathology.
Expression of genes involved in the amplification of inflammatory responses results when cells perceive evidence of infection or injury. Inflammation that occurs in response to infectious agents usually begins as a result of activation of recognition receptors such as Toll-like receptors (TLRs), which recognize certain pathogen-associated molecules that are not present in the host. The cells most involved in the innate immune response, such as macrophages and microglia, highly express these receptors. In recent years, the hypothesis that these recognition receptors may also respond to endogenous molecules such as those released during necrotic processes for example has been confirmed [16]. Purinergic receptors are also expressed on microglia and astrocytes and come into play in some pathological states [17].
As mentioned above, the brain pathological features of AD include extracellular amyloid plaques; in addition, the inflammatory response in AD induces an increase in the number, size, and motor activity of microglia as well as a morphological change whereby they change from branched (resting) to amoeboid (active). In addition, microglia surrounding plaques appears positive for activation markers and proinflammatory mediators, including MHC class II, COX-2, TNF-α, and some interleukins [18,19]. Proinflammatory cytokines, such as TNF-α, IL-1β, and IL-6, in turn can act directly on neurons to induce apoptosis [20,21], and activation of caspases and signal-dependent transcription factors, such as NF-κB and AP-1, leads to the production of numerous amplifiers (e.g., IL-1β, TNF-α, IL-6). Finally, communication between neurons and glia can amplify the production of neurotoxic factors that contribute to AD pathology; in fact, molecules with pro-inflammatory activity on neurons can increase Aβ production and consequently microglia-mediated inflammation [22].
The inflammatory reaction, characterized by a discharge of chemokines and ROS, that is triggered centrally in several acute brain diseases including in ischemic stroke leads neuroinflammation and is often followed by necrosis and apoptosis.
Immune mediators play a key proinflammatory role capable of enhancing brain cells and promoting the penetration of inflammatory cells such as macrophages, T cells, and neutrophils into the area affected by the ischemic insult. Cytokines such as IL-6 and TNF-α appear to be crucial mediators of post-ischemic inflammation Maida, 2020 [23].
3. Limitations of current therapies used for the treatment of neuroinflammation and neurodegenerative disorders in early stages of disease
There are numerous risk factors and diseases that can be considered predisposing factors for neurodegenerative diseases, including cerebrovascular disease, hyperlipidemia, smoking, diabetes, obesity, and traumatic brain injury. In contrast, protective factors are less numerous and include increased cognitive reserve, consumption of a Mediterranean diet, and regular exercise [4,24] but there are no completely effective specific therapies to date [25]. There are currently no proven pharmacological treatments that modify the course of AD, although therapies targeting aspects of both amyloid and tau protein are under active investigation. Clinical management of AD thus aims to improve patient symptoms and optimize quality of life. Acetylcholine (ACh), a widely distributed neurotransmitter in the body known to improve cognition, is reduced in patients with AD; in fact, the increase in ACh level induced by the use of acetylcholinesterase inhibitors (e.g., donepezil, rivastigmine, and galantamine) has been associated with improved cognitive ability [26,27,28]. Memantine, an N-methyl-D-aspartate (NMDA) receptor antagonist capable of acting by suppressing glutamate-mediated excitotoxicity, has been shown to reduce clinical status deterioration in patients with moderate to severe AD compared with controls [29,30], but not in patients with mild disease [31]. The combination of acetylcholinesterase inhibitors and memantine may have marginal benefit over single-drug treatment [32]. In addition, it is necessary to consider the modest benefits of these treatments and the potential side effects of each treatment option. Even in the treatment of Parkinson’s disease, currently available drug therapies do not represent a complete resolution of symptoms, and they often offer good control of motor symptoms [33,34] but do not change the evolution of the disease [35]. Drug therapies targeting the motor features of Parkinson’s disease act by potentiating dopamine signaling and involve direct implementation of the neurotransmitter Dopamine (e.g, levodopa), monoamine oxidase (MAO) enzyme inhibition (e.g., selegiline) and catechol-O-methyltransferase (COMT) inhibition (e.g., entacapone). Anticholinergics (e.g., triesiphenidyl, benzotropine) are effective in patients with a predominant tremor phenotype, and their use is preferred in the early stages of Parkinson’s disease. Deep brain stimulation (DBS) can alleviate motor fluctuations and tardive dyskinesias in patients with drug-refractory PD providing benefits for tremor, and rigidity, but gait, balance, and cognitive abilities are unlikely to improve [36]. Electrodes placed in the inner globe pallidum or subthalamic nucleus regulate abnormal neural impulses, thereby alleviating motor symptoms [37,38,39,40]. Co-treatment with DBS may reduce the dose or adverse effects of medications, but some complications such as bleeding and infection should be considered here [41,42,43]. Psychiatric manifestations of AD in some cases respond to symptomatic treatments for AD but often require more specific treatment with psychiatric medications. Depression, for example, is often treated with selective serotonin reuptake inhibitors (SSRIs) with low anticholinergic properties (e.g., citalopram, escitalopram, fluoxetine) [44]. In fact, treatment with neuroleptic drugs should be avoided because they are poorly effective and at the same time risky because of the side effects and increased risk of mortality that follow; however, when nonpharmacological treatments yield insufficient results, the use of these drugs becomes necessary [45]. In DLB and PDD, dopaminergic therapy is used to treat extrapyramidal symptoms despite the fact that this gives less symptomatic improvement than that observed in PD. Depression and anxiety in DLB and PDD can be treated with SSRIs or serotonin-norepinephrine reuptake inhibitors (SNRIs). Atypical antipsychotics may be useful for psychiatric symptoms but should be used with great caution for their adverse effects on movement and cognition. Traditional neuroleptics should be avoided because of neuroleptic hypersensitivity in patients with DLB. Disease-modifying agents are not yet clinically available [46]. Furthermore, treatment with dopamine agonists can cause side effects in the Parkinsonian patient which also include changes in the behavior probably due to overstimulation of mesolimbic dopamine receptors, in predisposed subjects and this may force the interruption of DA therapy or the substitution with a new drug [47,48].
4. The potential role of Andrographis paniculata in the treatment of neuroinflammation and neurodegenerative diseases
Despite research efforts, treatments currently used for neurodegenerative diseases as already seen provide marginal symptomatic benefit. In recent years, research has focused on products of natural origin and thus phytocompounds that could be a valuable aid in combating various diseases [49,50,51,52]. Among the most studied medicinal plants, Andrographis paniculata (Ap) and especially its main constituent, Andrographolide (Andro), a diterpene lactone, have attracted particular interest. Ap is a plant of Asian origin already known since ancient times under the name “King of Bitters”. Ap is known for its anti-inflammatory, antibacterial, antiviral, antineoplastic, hepatoprotective, hypolipidemic, and hypoglycemic properties [53,54]. More specifically, in preclinical studies in rats, the aqueous extract of Ap was able to result in a reduction in the expression of inflammatory markers such as TNF-α, IL-1β, IL-6 and oxidative stress makers such as ROS and Thiobarbituric Acid Reactive Substances (TBARS) with an enhancement of SOD (Superoxide Dismutase), CAT (Catalase) and GSH (Glutathione) activity [55]. In addition, through pharmacokinetic analyses conducted on Wistar rats treated with Ap and Andro extract, it was seen that the compound is able to easily cross the blood-brain barrier and generate effects at the neuronal level due to its high distribution capacity in the brain [56]. Especially in the hippocampus Ap is able to reduce the activity of LPS-induced cholinesterases, thus showing anti-inflammatory activity and improving mnemonic processes, as confirmed by spatial learning tests [55]. The role of Andro has also been studied in the context of neuroinflammation. A study performed in vivo, by LPS-induced neuroinflammation, showed that treatment with Andro reduced cortical levels of certain chemokines such as CCL2, CCL5. Furthermore, the same authors observed the anti-inflammatory effect of Andro in vitro by inducing neuroinflammation of astrocytes with both LPS and IL-1β. In the LPS-induced neuroinflammation model, Andro treatment was able to inhibit LPS-induced TNF- α release through inhibition of NFkB and JNK, additionally increasing Nrf2 levels and HO-1 expression through the p38/MAPK and ERK-dependent signaling pathway [57]. While in the IL-1β-induced neuroinflammation model, the compound had been able to reduce the release of CCL5, phosphorylation of NFkB p65 and IkBα, and glial fibrillary acidic protein (GFAP) [58]. To investigate the role of Andro in LPS-induced neuroinflammation and memory, Das et al. evaluated the expression of TLR4 and its endogenous ligand High Mobility Group Box 1 (HMGB1) in primary mixed glial cells (PMLCs) in adult prefrontal cortices. Andro treatment was successful in reducing TLR4 expression and LPS-induced p-NFκB-p65 activation. In microglia, on the other hand, a reduction in CD-68 expression levels and an increase in arginase-1 levels were observed. While at the cortical level Andro was able to reduce the expression of GFAP, exerting a neuroprotective action in astrocytes through inhibition of the release of pro-inflammatory molecules such as iNOS, COX-2, nitrates, NLRP3, and caspase-1. Finally, in the context of cognitive memory processes Andro had been able to antagonize overexpression of PKC and phosphorylation of cAMP responsive element binding protein (CREB), which normally result up-regulated in cognitive deficits. The same authors also observed that at the level of the prefrontal cortex Andro had reduced levels of β-amyloid, APP, ptau, BACE-1 (β-secretase-1), activation of the pro-apoptotic caspase-3 and bax genes and increased levels of the anti-apoptotic gene Bcl-2, improving neuronal viability. At the hippocampal level, on the other hand, an increase in Post Synaptic Density Protein 95 (PSD-95) and synapsin was observed, resulting in increased neuronal plasticity, reducing the expression levels of inflammatory cytokines such as TNF-α and Macrophage Inflammatory Protein-1 (MIP-1), and an increasing those of TGF-β and IL-10 [59]. In another study conducted by Adedayo et al, the effect of Andro in counteracting the amnesic effect of scopolamine, similar to that found in AD, was evaluated. Rats treated with the aqueous extract of Ap, showed an improvement in cognitive function, as assessed by the Y-Labyrinth Test and the Morris Water Maze (MWM) Test, and through other analyses performed, significant reductions in AChE and Buttiril-Che, of some Monoamine Oxidases (MAO), purinergic enzymes such as ATPdase, ADPdase, while the levels of 5-Nucletidase and Adenosine Deaminase at the hippocampal level were found to be increased [60]. Because neuroinflammation also plays a key role in diseases such as depression and schizophrenia [61,62], some studies have wanted to investigate the possible role of Andro in these diseases. In an in vivo model of schizophrenia induced by the administration of phencyclidine (PCP), Andro reduced the levels of IL-1β and TNF-α, p-p65, p-IκBα, p-p38 and p-ERK1/2 in the prefrontal cortex, improved oxidative stress instead through the activation of antioxidant enzymes such as SOD, CAT and GSH-Px and increased the levels of NRF-2, HO-1 and NQO-1. Cognitive improvement was also confirmed by behavioral tests that showed increased locomotor activity [63]. Andro was also found to be effective in the treatment of stress-induced depression. In fact, mice subjected to chronic unpredictable mild stress (CUMS) and treated with Andro showed reduced levels of NO, iNOS, IL-1β, IL-6, TNF-α, COX-2, p-p65, p-IκBα, and NLR family pyrin domain containing 3 (NLRP3) in the prefrontal cortex, compared with untreated mice. In addition, Andro exerted a pro-autophagic action through increased Beclin-1 expression and decreased p-mTOR [64]. In a recent study, the effects of Andro in the treatment of neurological disorders induced by heavy metal exposure were evaluated. Specifically, the effects of Andro on aluminum intoxication in Drosophila were evaluated. Andro treatment on flies resulted in improved survival, improved locomotor performance, and improved learning and memory through reduced AChE and MAO activity and increased catalase activity [65]. In 2005, Tobar et al., also observed the protective action of Andro in the treatment of Multiple Sclerosis, an autoimmune neurodegenerative disease in which immune defenses and particularly lymphocytes, attack components of the nervous system. Indeed, in dendritic cells (DCs) pulsed with hen’s egg ovalbumin (OVA) Andro counteracted the generation of peptide-MHC complexes required for T-cell activation by inhibiting the up-regulation of maturation markers I-Ab, CD40 and CD86 in LPS-treated dendritic cells. Andro treatment was also tested in vivo on C57Bl/6 mice with experimental autoimmune encephalomyelitis (induced with the peptide MOG35-55) by significantly reducing the incidence of the disease as also demonstrated by reduced production of IFN and IL-2 [66]. Therefore, based on the data reported in the literature, we can state that both Ap and Andro are effective in treating the inflammatory and degenerative effects that occur during various disease states in the nervous system [54,55,67,68]. Here we will examine these effects on diseases involving the nervous system focusing on those most prevalent in the world population.

Table 1.
Effects of Ap and Andro on various animal and cellular models and on various tissues.
| Effects on proinflammatory molecules and on oxidative stress mediators expression | Reduction of TNF-α, IL-1β, IL-6, ROS and TBARS expression; reduction of Chemokine ligand 5 (CCL5) release, phosphorylation of NFkB p65 and IkBa, as well as GFAP (Glial fibrillary acidic protein) induced by IL-1b; LPS-induced reduction of TLR4 expression and p-NFκB-p65 activation; reduction of the levels of inflammatory cytokines such as TNF-α and MIP-1 (Macrophage Inflammatory Protein-1); Increase in anti-inflammatory cytokines such as TGF-β and IL-10; reduction of pro-inflammatory molecules expression such as iNOS, COX-2, nitrates, NLRP3, caspase-1; improvement of SOD (Superoxide Dismutase), CAT (Catalase) and GSH (Glutathione) activity. |
|---|---|
| Effects on the hippocampus | Reduction of Cholinesterase activity induced by Lipopolysaccharide showing an anti-inflammatory activity and improving memory; increased expression of PSD-95 (Post Synaptic Density Protein 95) and synapsin which are involved in synaptic plasticity; increase in purinergic enzymes such as ATPdase, ADPdase. 5-Nucletidase and Adenosine Deaminase. |
| Effects in models of neuroinflammation | Reduction of cortical levels of chemokines such as CCL2, CCL5; |
| Effects on astrocytes | Increased Nrf2 levels and HO-1 expression through the p38 MAPK and ERK dependent pathway; anti-inflammatory effect in vitro; reduction of GFAP expression. |
| Effects on microglia | Reduction of CD-68 expression; increased levels of arginase-1. |
| Effects on prefrontal cortex | Reduction in β-amyloid, APP, ptau, BACE-1 (β-secretase-1) levels; activation of caspase-3 and bax; increased levels of the anti-apoptotic gene Bcl-2. |
| Effects on the activity of neurotransmitters | Reduction of AChE and Buttyryl-Che, and of Monoamine Oxidases (MAO). |
| Effects on schizophrenia | Reduction of IL-1β and TNF-α, p-p65, p-IκBα, p-p38 and p-ERK1/2 levels in the prefrontal cortex; activation of antioxidant enzymes such as SOD, CAT and GSH-Px; increased levels of NRF-2, HO-1 and NQO-1; increased locomotor activity. |
| Effects on stress-induced depression | Reduction of NO, iNOS, IL-1β, IL-6, TNF-α, COX-2, p-p65, p-IκBα levels and NLRP3 inflammasome assembly in the prefrontal cortex; pro-autophagic action through an increase in Beclin-1 expression and a reduction in p-mTOR. |
| Effects in aluminum poisoning | Improved survival, locomotor performance, learning and memory through a reduction of AChE and MAO activity and increased catalase activity. |
| Effects on Multiple Sclerosis | Prevents the generation of peptide-MHC complexes required for T cell activation; inhibition of up-regulation of maturation markers I-Ab, CD40 and CD86 in LPS-treated dendritic cells. |
| Effects on experimental autoimmune encephalomyelitis | Significant reduction in the incidence of the disease as demonstrated by the reduced production of IFN and IL-2. |
4.1. Alzheimer Disease
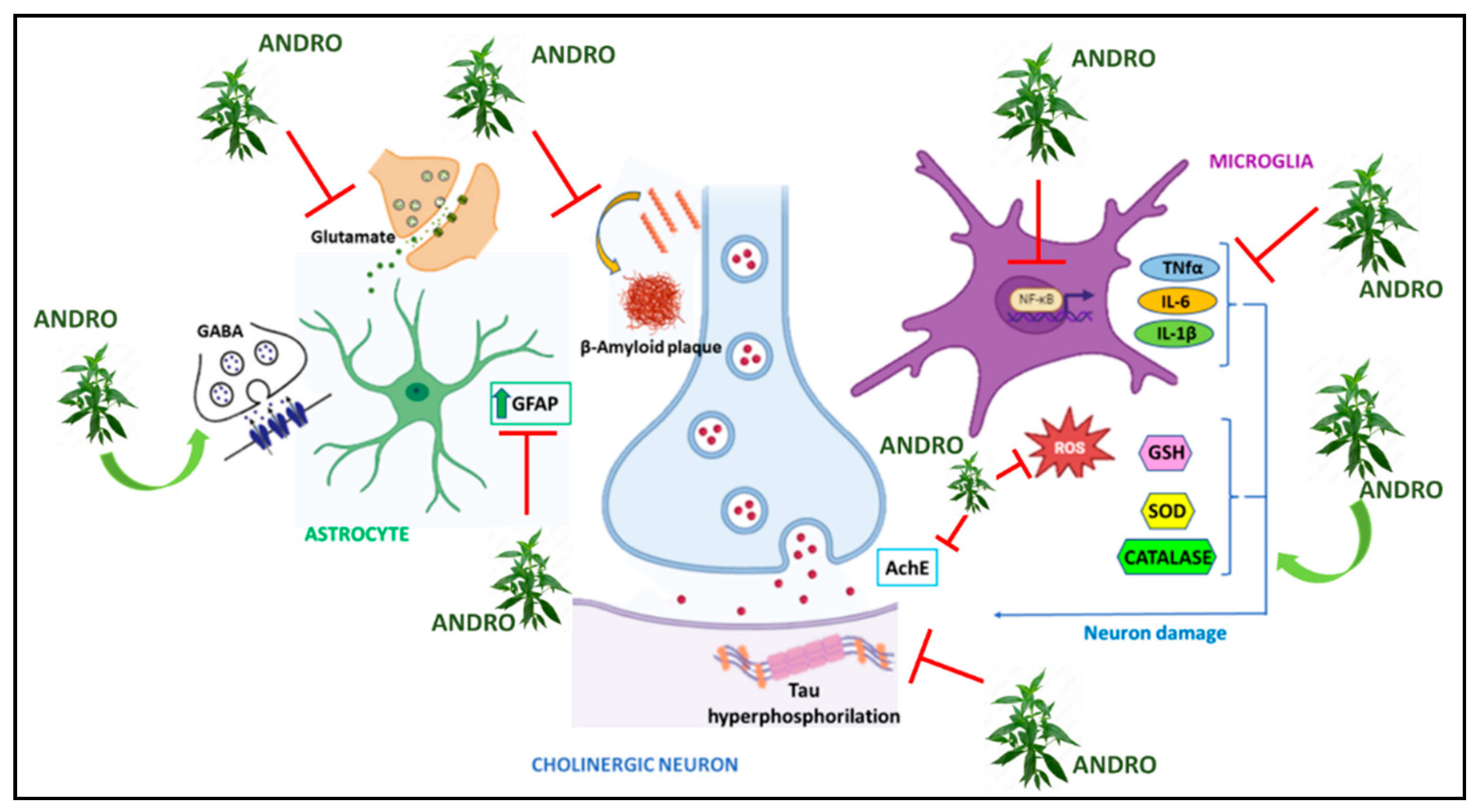
In vitro studies showed that andrographolide and some of its derivatives were able to activate α-secretase, which is involved in the inhibition of Aβ formation, and to inhibit β-secretase, which is involved in Aβ formation, without the protective treatment causing toxicity [69]. On hippocampal neuronal cells of HT-22 mice, treatment with Andro increased the levels of nuclear factor erythroid-derived 2-like 2 / Kelch-like ECH-associated protein 1 (Nrf2/Keap), Antioxidant Response Element (ARE) gene, and HO-1 enzyme. Since these factors are major players in anti-inflammatory and antioxidant responses, again Andro was able to induce a cytoprotective response in the brain [70]. On mouse microglia BV-2 (microglial cells derived from C57/BL6 murine), on the other hand, Andro reduced the expression of Aβ improving neuronal viability and, through the NF-kB-mediated signaling pathway, reduced the levels of TNF-α, COX-2, PGE2, i-NOS, NO, and cytokines such as IL-1β, IL-6, protecting neurons from damage produced by inflammation [71].In addition to Andro, its analogs have also been used on these same cells, achieving complementary results. Indeed, the analogs have shown the ability to inhibit LPS-induced NO production but also iNOS expression, TNF-α and IL-6 production [72]. One enzyme known to be involved in learning and memory processes and especially in tau protein phosphorylation and increased β-amyloid production is Glycogen Synthase Kinase-3β (GSK3β) [73]. Andro treatment on primary hippocampal neuronal cultures induced inhibition of GSK3β enzyme and reduction of its active form [74]. The same treatment conducted on HEK293 cells (Human embryonic kidney cells) showed comparable results. Furthermore, because GSK3β is involved in the Wnt/β-catenin signaling pathway known for its role in neurogenesis, the possible involvement of Andro on this pathway was also investigated [75]. Andro treatment on hippocampal cells showed an induction of Wnt gene transcription by restoring its proper activity [74]. Several studies on the effects of Ap have also been conducted in mouse models. In fact, in the literature, we find several transgenic mouse models used to reproduce Alzheimer’s disease [76,77]. In one study in which Andro was tested on 2-month-old mice that had mutations on genes encoding for APP and presenilin (PS1), known proteins involved in AD, increased expression of the Wnt/β-catenin signaling pathway was observed [78]. In another experiment in which mice were used instead with ages 7-12 months, Andro was able to alter the maturation of amyloid plaques in the cortex and hippocampus; in fact, in the early stages of the disease, the number of plaques was found to be reduced. Furthermore, in the same study, Andro was also observed to exert its effects on tau protein phosphorylation, leading to a significant increase in post-synaptic proteins including Shank, GluN2B, GluA2, PSD-95, and an increase in the inactive form of GSK-3β. While behavioral tests found that Andro-treated mice showed improved learning and lower latency values and improved spatial memory performance [79]. In addition to transgenic mouse models, some work has also been conducted on Octodon degus mice, which are capable of naturally reproducing the neurodegeneration and neurological signs of Alzheimer’s with advancing age [77]. Rivera et al., performed several behavioral tests on these 12- and 56-month-old rodents to assess their degree of spatial learning and memory after Andro administration. The results obtained showed neuroprotective effects that resulted in recovery of memory and learning, enhancement of excitatory postsynaptic field potential (fEPSP), protection of certain proteins such as synaptophysin (SYP), increased vesicular glutamate transporter 1 (vGluT1) and NMDA receptor subunit GluN2A. Their results also showed a decrease in phosphorylated tau protein and Aβ aggregate maturation in aged mice [80]. In contrast, on these same animals, Lindsay et al. reported the neuroprotective effects of Andro through a reduction in Aβ, GFAP, IL-6, COX-2, and oxidative stress markers such as 4-HNE (4-Hydroxynonenal) and N-Tyr (N-Tyrosin) in the brain [81]. It is well known that impaired glucose metabolism at the neuronal level may be related to several neurodegenerative diseases, including AD [82,83,84,85,86,87]. Cognitive deficits and symptoms comparable to AD are also reproducible following intracerebroventricular administration of streptozotocin (SZT). Rats that underwent this treatment and treated with Andro show improved spatial memory in the Morris Water Maze (Morris Water) and the Elevated Plus Maze test compared with the untreated group of animals. Andro also reduced levels of neuroinflammatory markers such as TNF-α, IL-1B and IL-16, reduced levels of the neurotransmitter glutamate (GLU), and increased levels of GABA. Because STZ administration induces increased levels of AChE and p-tau, the group of animals that had received Andro treatment showed a significant reduction in the expression levels of AChE and p-tau, as well as a reduction in oxidative stress through attenuation of MDA (Malondialdehyde) and nitrite, and increased levels of GSH, SOD and catalase compared with the group treated with STZ alone [88]. Comparable results were also obtained on diabetic rats given oral administrations of STZ and subsequently treated with Andro [89]. In another study on rat primary hippocampal neurons, treatment with Andro resulted in increased glucose uptake through increased GLUT translocation and increased ATP production resulting in promotion of AMPK-dependent glycolysis [90].
Figure 1.
Graphic representation of molecular mechanisms underlying Alzheimer’s Disease on which Andrographis paniculata acts.
Figure 1.
Graphic representation of molecular mechanisms underlying Alzheimer’s Disease on which Andrographis paniculata acts.
4.2. Parkinson’s disease
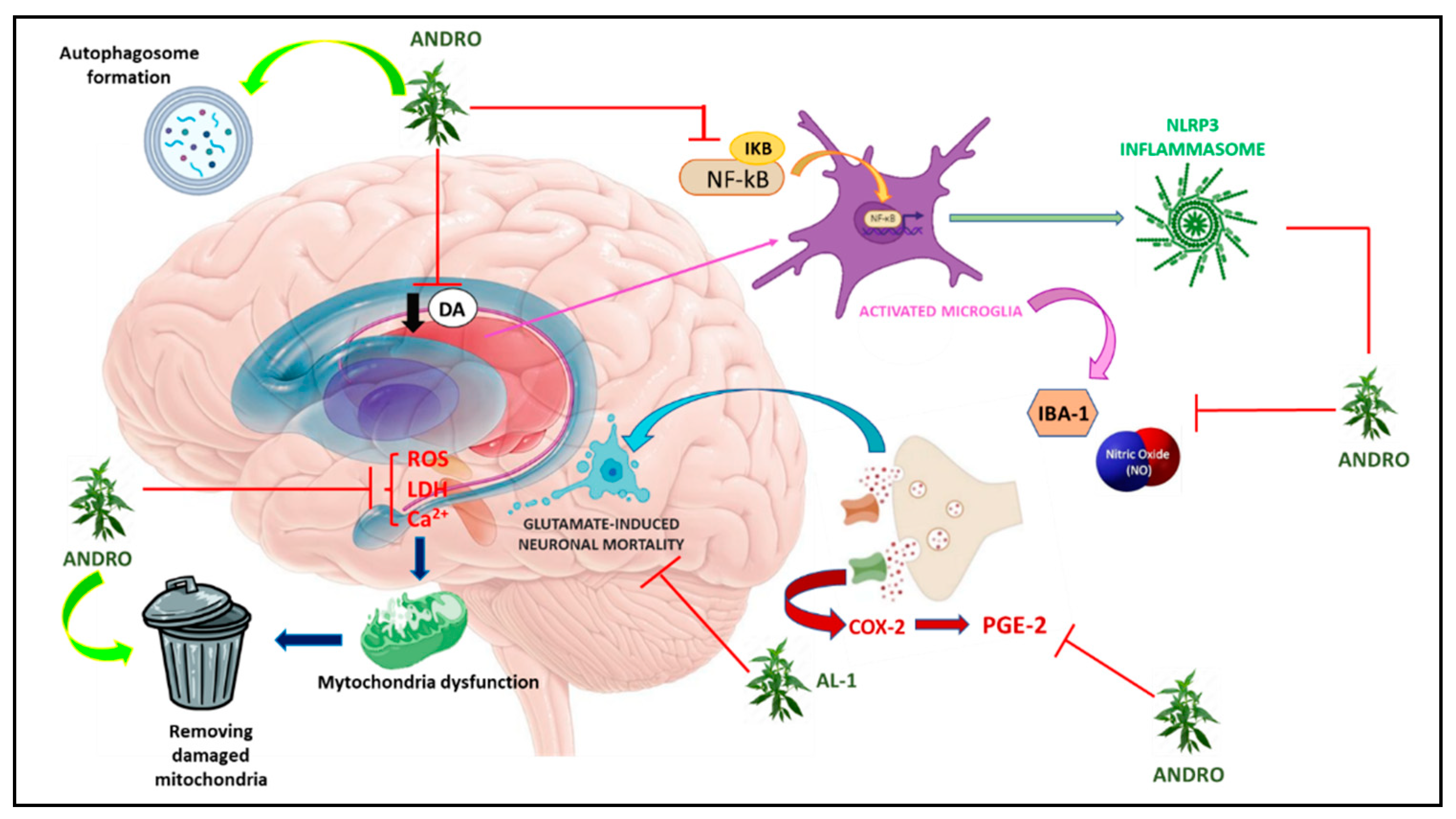
As already widely described, inflammation and apoptosis are important factors in various neurodegenerative diseases and Parkinson’s is one of them [91,92]. To study the effects of Andro in this disease, rat midbrain glia cultures were pre- and post-treated concomitantly with LPS-induced dopaminergic neurodegeneration. The results obtained showed that Andro was able to attenuate LPS-induced dopaminergic neurodegeneration by reducing the activation of microglia and inflammatory factors such as ROS, TNF-α, NO and PGE2. In addition, pretreatment with Andro on BV2 microglia cells reduced the expression of COX-2 and iNOS. However, in this work, the neuroprotection of Andro on neurodegeneration induced by 1-methyl-4-phenyl-pyridine (MPP), a metabolite of MPTP used to induce Parkinson’s, was also tested, although pretreatment with Andro failed to reduce neuronal damage [93]. In another study conducted instead on mice subjected to intraperitoneal administration of MPTP, treatment with Andro produced positive results in all behavioral tests performed including the catalepsy test, grip strength (grip strenght) and rota rod, leading to improved motor conditions [64]. In vitro, it was seen that LPS- and MPP-induced microglia activation was reduced following Andro administration resulting in decreased NLRP3 inflammasome activation, and these data were confirmed by inhibition of the microglial expression marker Iba-1 both in vitro and in vivo on mouse brain [94,95]. Andro has also been shown to ameliorate mitochondrial dysfunction through inhibition of ROS formation and maintenance of mitochondrial membrane potential (in vitro) by also promoting autophagosome formation and elimination of damaged mitochondria [95]. The neuroprotective effects of Andro on MPP-induced damage were also studied on neuroblastoma cells (SH-SY5Y). In these cells, andrographolide-lipoic acid (AL-1) conjugate protected against MPP-induced damage leading to increased cell viability and inhibited phosphorylation of NF-κB p65 and IκBα. While in MPTP-treated mice, AL-1 protected against the loss of TH-positive dopaminergic neurons in the substantia nigra pars compacta, attenuated dopamine loss in the striatum, and improved motor functions as demonstrated by behavioral tests [96]. However, in a later study performed by Ketterman et al, in SH-SY5H cells, administration of Andro did not demonstrate protective effects against oxidative stress [94]. Dysregulation of GLU levels is involved in several processes that may result in neuronal damage [97,98,99]. In this regard Yang et al., in 2014, studied the neuroprotective effect of Ap extract on HT22 neuronal cells in the mouse hippocampus following glutamate-induced damage. The results showed that Ap can significantly reduce GLU-induced neuronal mortality as well as reduce cytosolic lactate dehydrogenase (LDH) levels, reduce Ca2+ influx, and reduce intracellular ROS production induced by GLU in a dose-dependent manner. In addition, the same authors also observed a significant decrease in the phosphorylation of MAPK, p38, ERK and JNK, while Western blotting analysis revealed a restoration of the levels of anti-apoptotic proteins such as Bcl-2, Bid, Bax and a reduction in apoptosis-inducing factor (AIF) after Andro treatment [70].
Figure 2.
Graphic representation of molecular mechanisms underlying Parkinson’s disease on which Andrographis paniculata acts.
Figure 2.
Graphic representation of molecular mechanisms underlying Parkinson’s disease on which Andrographis paniculata acts.
4.3. Brain Ischemia-reperfusion injury
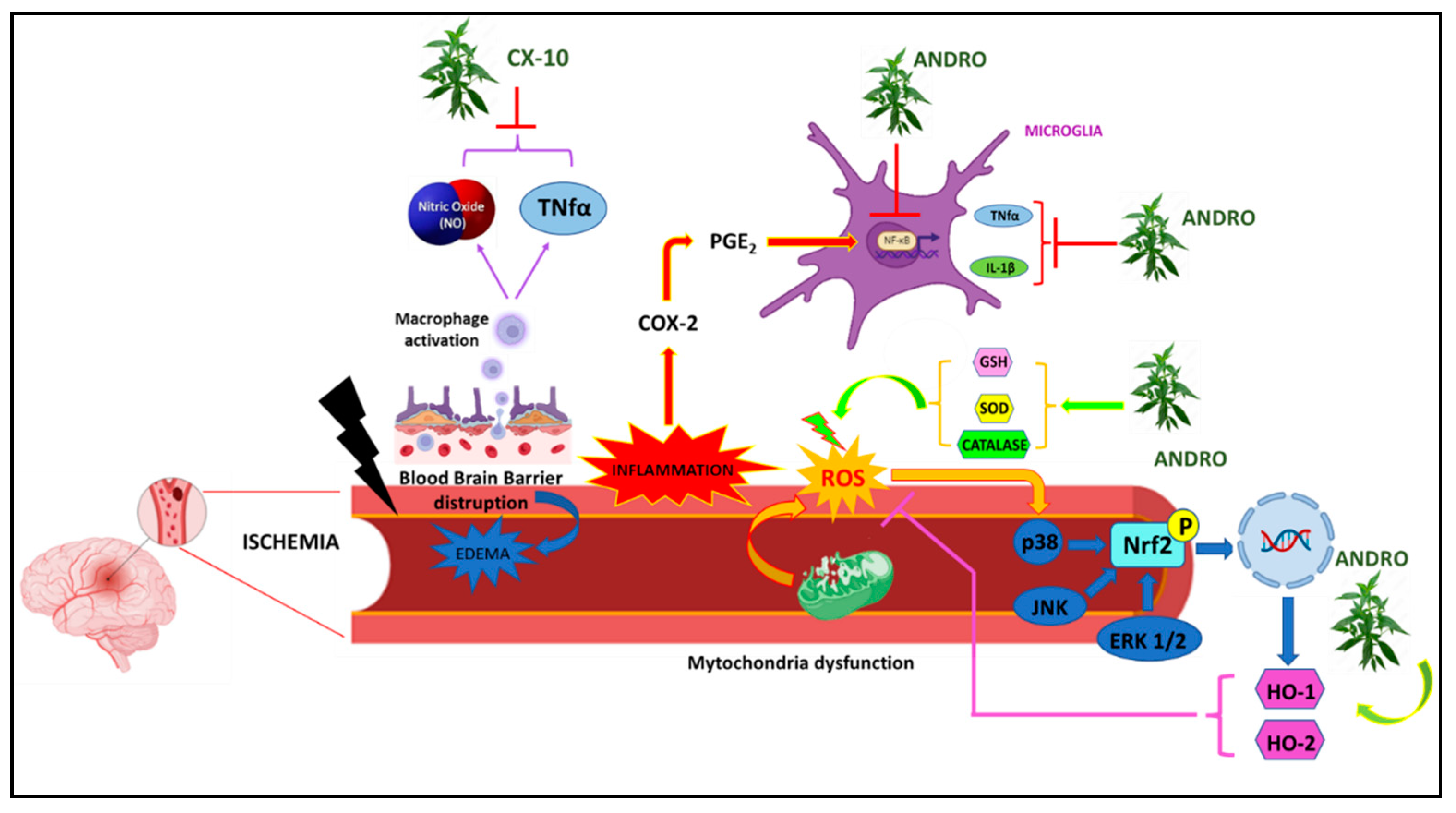
Cerebral ischemia is a pathological condition characterized by reduced blood supply that can affect different areas of the brain and induce major neuronal damages [100]. In a study conducted on mouse brain endothelial cells, no positive effects were found following the administration of Andro, as the compound not only did not improve the viability of these cells but induced a marked reduction in the number of neuronal cells as a result of increased LDH release and increased apoptosis, blocking cell growth in the G0/G1 phase [101]. However, from many other studies instead, especially in recent years, the protective action of Andro and its derivatives on ischemic damage seems to emerge and be confirmed. More specifically, in a rat mouse model with permanent middle cerebral artery occlusion (pMCAO), Andro reduced infarct volume and microglia activation in peri-infarct areas by going against increased levels of inflammatory markers such as IL-1β, TNF-α, PGE2 and the transcription factor NF-kB [102]. In another study, however, rats undergoing ischemic brain injury/reperfusion (CI/R) treated with Andro showed decreased cerebral infarction, reduced superoxide anion and nitrotyrosine, reduced expression of gp91phox/NOX2, IL-1β and Hypoxia Inducible Factor (HIF), as well as reduced p65NFkB, thus confirming the protective effects of Andro in counteracting an ischemic attack [103]. Mice with CI/R-induced oxidative brain damage and treated with Andro also reported reduced magnitude of infarction and subsequent neurological deficits, accompanied by reduced free radical production, nitrotyrosine, CD11b, NOX2 and iNOS formation [104].
Another andrographolide derivative, triacetylandrographolide (CX-10), has been used to test possible neuroprotective effects against cerebral ischemia. On the macrophage cell line RAW264.7, both Andro and CX-10 counteracted NO and TNF-α production. While in BALB rats, CX-10 reduced LPS-induced TNF-α production. In Sprague-Dawley rats with middle cerebral artery occlusion (MCAO), reduced infarct size as well as improved motor performance were observed. At the level of brain tissues, CX-10 reduced the levels of TNF-α and IL-1β and increased the activity of antioxidant enzymes such as SOD, CAT, and GSH-P. Western blot analysis also showed the positive effects of Andro in reducing the expression levels of TLR4, NF-κB, TNF-α, iNOS proteins by increasing the expression of Nuclear factor erythroid 2-related factor 2 (Nrf2) and HO-1 [103]. In mouse Cerebral Endothelial Cells (CECs), treated with 10μM Andro, an increase in HO-1 expression mediated by phosphorylation of ERK1/2, p38 MAPK and JNK1/2, and an increase in Heme oxygenase 2 (HO-2) protein, via the Nrf2 signalling pathway, was observed. In this way, Andro was able to protect neuronal cells from cell death induced by oxygen-glucose deprivation (OGD), thus improving cell viability. While in rats subjected to MCAO, Andro’s antioxidant effect in counteracting free radical production was observed, and a reduction in cerebral oedema and infarct volume was observed in brain tissue analyses [105].
Figure 3.
Graphic representation of molecular mechanisms underlying Ischemia on which Andrographis paniculata acts.
Figure 3.
Graphic representation of molecular mechanisms underlying Ischemia on which Andrographis paniculata acts.
5. Conclusions
In conclusion, Ap and Andro exert neuroprotective effects in different NDs. The emerging informations of the Ap and Andro properties exerted by cytoprotective, anti-inflammatory, and antioxidant effects seems to explain many mechanisms at the basis of the neuroprotective action. In addition, since they are phytochemical compound, they could also be considered as adjuvant to conventional drugs therapies, and a greater understanding of the neuroprotective molecular mechanisms of Ap and Andro could be crucial for identifying new therapeutic targets in NDs.
Author Contributions
Conceptualization, Vincenzo Mollace (VM) and Giovambattista De Sarro (GDS); writing—original draft preparation, Francesca Bosco (BF), Lorenza Guarnieri (LG), and Stefano Ruga (SR); writing—review and editing, BF, LG, Antonio Leo (AL), Rita Citraro (RC) and Juan Hancke (JH); Images realization, FB; participation in drafting the article, Jessica Maiuolo (JM), Francesca Oppedisano (FO), Roberta Macrì (RM), Federica Scarano (FS), Saverio Nucera (SN), Irene Bava (IB), Ernesto Palma (EP), and Carolina Muscoli (CM). All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest
References
- Prusiner, S.B. Biology and genetics of prions causing neurodegeneration. Annu Rev Genet 2013, 47, 601–623. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.G.; Lue, L.F.; Serrano, G.; Adler, C.H.; Caviness, J.N.; Sue, L.I.; Beach, T.G. Altered Expression Patterns of Inflammation-Associated and Trophic Molecules in Substantia Nigra and Striatum Brain Samples from Parkinson’s Disease, Incidental Lewy Body Disease and Normal Control Cases. Front Neurosci 2015, 9, 507. [Google Scholar] [CrossRef] [PubMed]
- Reitz, C.; Rogaeva, E.; Foroud, T.; Farrer, L.A. Genetics and genomics of late-onset Alzheimer’s disease and its endophenotypes. Int J Alzheimers Dis 2011, 2011, 284728. [Google Scholar] [CrossRef] [PubMed]
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer disease. Cold Spring Harb Perspect Med 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Ortiz, A.L.; Acosta-Castillo, I.; Prince, M.J. Epidemiology of dementias and Alzheimer’s disease. Arch Med Res 2012, 43, 600–608. [Google Scholar] [CrossRef]
- Niu, H.; Álvarez-Álvarez, I.; Guillén-Grima, F.; Aguinaga-Ontoso, I. Prevalence and incidence of Alzheimer’s disease in Europe: A meta-analysis. Neurologia 2017, 32, 523–532. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur J Neurol 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Moisan, F.; Kab, S.; Mohamed, F.; Canonico, M.; Le Guern, M.; Quintin, C.; Carcaillon, L.; Nicolau, J.; Duport, N.; Singh-Manoux, A. , et al. Parkinson disease male-to-female ratios increase with age: French nationwide study and meta-analysis. J Neurol Neurosurg Psychiatry 2016, 87, 952–957. [Google Scholar] [CrossRef]
- Lippa, C.F.; Knopman, D.S. Dementia: many roads, but not built in a day. Neurology 2007, 69, 2193–2194. [Google Scholar] [CrossRef]
- Thenganatt, M.A.; Jankovic, J. Parkinson disease subtypes. JAMA Neurol 2014, 71, 499–504. [Google Scholar] [CrossRef]
- Alves, G.; Larsen, J.P.; Emre, M.; Wentzel-Larsen, T.; Aarsland, D. Changes in motor subtype and risk for incident dementia in Parkinson’s disease. Mov Disord 2006, 21, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Heemels, M.T. Neurodegenerative diseases. Nature 2016, 539, 179. [Google Scholar] [CrossRef] [PubMed]
- Logroscino, G.; Urso, D.; Savica, R. Descriptive Epidemiology of Neurodegenerative Diseases: What Are the Critical Questions? Neuroepidemiology 2022, 56, 309–318. [Google Scholar] [CrossRef]
- Dugger, B.N.; Dickson, D.W. Pathology of Neurodegenerative Diseases. Cold Spring Harb Perspect Biol 2017, 9. [Google Scholar] [CrossRef]
- Akiyama, H. [Alzheimer’s disease and the immune system response]. Nihon Rinsho 1994, 52, 2990–2994. [Google Scholar] [PubMed]
- Balistreri, C.R.; Colonna-Romano, G.; Lio, D.; Candore, G.; Caruso, C. TLR4 polymorphisms and ageing: implications for the pathophysiology of age-related diseases. J Clin Immunol 2009, 29, 406–415. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Ceruti, S.; Bramanti, P.; Abbracchio, M.P. Purinergic signalling in inflammation of the central nervous system. Trends Neurosci 2009, 32, 79–87. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L. , et al. Inflammation and Alzheimer’s disease. Neurobiol Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Cartier, L.; Hartley, O.; Dubois-Dauphin, M.; Krause, K.H. Chemokine receptors in the central nervous system: role in brain inflammation and neurodegenerative diseases. Brain Res Brain Res Rev 2005, 48, 16–42. [Google Scholar] [CrossRef]
- McCoy, M.K.; Tansey, M.G. TNF signaling inhibition in the CNS: implications for normal brain function and neurodegenerative disease. J Neuroinflammation 2008, 5, 45. [Google Scholar] [CrossRef]
- Simi, A.; Tsakiri, N.; Wang, P.; Rothwell, N.J. Interleukin-1 and inflammatory neurodegeneration. Biochem Soc Trans 2007, 35, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Maida, C.D.; Norrito, R.L.; Daidone, M.; Tuttolomondo, A.; Pinto, A. Neuroinflammatory Mechanisms in Ischemic Stroke: Focus on Cardioembolic Stroke, Background, and Therapeutic Approaches. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Petersson, S.D.; Philippou, E. Mediterranean Diet, Cognitive Function, and Dementia: A Systematic Review of the Evidence. Adv Nutr 2016, 7, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Durães, F.; Pinto, M.; Sousa, E. Old Drugs as New Treatments for Neurodegenerative Diseases. Pharmaceuticals (Basel) 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Birks, J.S.; Harvey, R. Donepezil for dementia due to Alzheimer’s disease. Cochrane Database Syst Rev 2003, CD001190. [Google Scholar] [CrossRef]
- Olin, J.; Schneider, L. Galantamine for Alzheimer’s disease. Cochrane Database Syst Rev 2001, CD001747. [Google Scholar] [CrossRef]
- Birks, J.S.; Chong, L.Y.; Grimley Evans, J. Rivastigmine for Alzheimer’s disease. Cochrane Database Syst Rev 2015, 9, CD001191. [Google Scholar] [CrossRef]
- Howard, R.; McShane, R.; Lindesay, J.; Ritchie, C.; Baldwin, A.; Barber, R.; Burns, A.; Dening, T.; Findlay, D.; Holmes, C. , et al. Donepezil and memantine for moderate-to-severe Alzheimer’s disease. N Engl J Med 2012, 366, 893–903. [Google Scholar] [CrossRef]
- Reisberg, B.; Doody, R.; Stöffler, A.; Schmitt, F.; Ferris, S.; Möbius, H.J.; Group, M.S. Memantine in moderate-to-severe Alzheimer’s disease. N Engl J Med 2003, 348, 1333–1341. [Google Scholar] [CrossRef]
- McShane, R.; Areosa Sastre, A.; Minakaran, N. Memantine for dementia. Cochrane Database Syst Rev 2006, CD003154. [Google Scholar] [CrossRef] [PubMed]
- Farrimond, L.E.; Roberts, E.; McShane, R. Memantine and cholinesterase inhibitor combination therapy for Alzheimer’s disease: a systematic review. BMJ Open 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Nemade, D.; Subramanian, T.; Shivkumar, V. An Update on Medical and Surgical Treatments of Parkinson’s Disease. Aging Dis 2021, 12, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Bosco, D.; Plastino, M.; Bosco, F.; Fava, A.; Rotondo, A. Daily motor performance after switching levodopa to melevodopa: an open-label on advanced Parkinson’s disease with “delayed-on” and/or”wearing-off”. Minerva Med 2011, 102, 125–132. [Google Scholar]
- Rascol, O.; Payoux, P.; Ory, F.; Ferreira, J.J.; Brefel-Courbon, C.; Montastruc, J.L. Limitations of current Parkinson’s disease therapy. Ann Neurol 2003, 53 Suppl 3, S3-12; discussion S12-15. 3. [CrossRef]
- Fasano, A.; Daniele, A.; Albanese, A. Treatment of motor and non-motor features of Parkinson’s disease with deep brain stimulation. Lancet Neurol 2012, 11, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Benabid, A.L.; Pollak, P.; Louveau, A.; Henry, S.; de Rougemont, J. Combined (thalamotomy and stimulation) stereotactic surgery of the VIM thalamic nucleus for bilateral Parkinson disease. Appl Neurophysiol 1987, 50, 344–346. [Google Scholar] [CrossRef]
- Siegfried, J.; Lippitz, B. Bilateral chronic electrostimulation of ventroposterolateral pallidum: a new therapeutic approach for alleviating all parkinsonian symptoms. Neurosurgery 1994, 35, 1126–1129. [Google Scholar] [CrossRef]
- Follett, K.A.; Weaver, F.M.; Stern, M.; Hur, K.; Harris, C.L.; Luo, P.; Marks, W.J.; Rothlind, J.; Sagher, O.; Moy, C. , et al. Pallidal versus subthalamic deep-brain stimulation for Parkinson’s disease. N Engl J Med 2010, 362, 2077–2091. [Google Scholar] [CrossRef]
- Odekerken, V.J.; Boel, J.A.; Schmand, B.A.; de Haan, R.J.; Figee, M.; van den Munckhof, P.; Schuurman, P.R.; de Bie, R.M.; group, N.s. GPi vs STN deep brain stimulation for Parkinson disease: Three-year follow-up. Neurology 2016, 86, 755–761. [Google Scholar] [CrossRef]
- Lyons, K.E.; Pahwa, R. Deep brain stimulation in Parkinson’s disease. Curr Neurol Neurosci Rep 2004, 4, 290–295. [Google Scholar] [CrossRef]
- Guridi, J.; Rodriguez-Oroz, M.C.; Alegre, M.; Obeso, J.A. Hardware complications in deep brain stimulation: electrode impedance and loss of clinical benefit. Parkinsonism Relat Disord 2012, 18, 765–769. [Google Scholar] [CrossRef]
- Pouratian, N.; Thakkar, S.; Kim, W.; Bronstein, J.M. Deep brain stimulation for the treatment of Parkinson’s disease: efficacy and safety. Degener Neurol Neuromuscul Dis 2012, 2012, 107–117. [Google Scholar] [CrossRef]
- Seitz, D.P.; Adunuri, N.; Gill, S.S.; Gruneir, A.; Herrmann, N.; Rochon, P. Antidepressants for agitation and psychosis in dementia. Cochrane Database Syst Rev 2011, CD008191. [Google Scholar] [CrossRef]
- Sink, K.M.; Holden, K.F.; Yaffe, K. Pharmacological treatment of neuropsychiatric symptoms of dementia: a review of the evidence. JAMA 2005, 293, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Paolini Paoletti, F.; Gaetani, L.; Parnetti, L. The Challenge of Disease-Modifying Therapies in Parkinson’s Disease: Role of CSF Biomarkers. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, A.; Bosco, D.; Plastino, M.; Consoli, A.; Bosco, F. Clozapine for medication-related pathological gambling in Parkinson disease. Mov Disord 2010, 25, 1994–1995. [Google Scholar] [CrossRef]
- Bosco, D.; Plastino, M.; Colica, C.; Bosco, F.; Arianna, S.; Vecchio, A.; Galati, F.; Cristiano, D.; Consoli, A.; Consoli, D. Opioid antagonist naltrexone for the treatment of pathological gambling in Parkinson disease. Clin Neuropharmacol 2012, 35, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics (Review). Mol Med Rep 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Quispe, C.; Hossain, R.; Islam, M.T.; Al-Harrasi, A.; Al-Rawahi, A.; Martorell, M.; Mamurova, A.; Seilkhan, A.; Altybaeva, N. , et al. Neuropharmacological Effects of Quercetin: A Literature-Based Review. Front Pharmacol 2021, 12, 665031. [Google Scholar] [CrossRef]
- Zhang, R.; Miao, Q.W.; Zhu, C.X.; Zhao, Y.; Liu, L.; Yang, J.; An, L. Sulforaphane ameliorates neurobehavioral deficits and protects the brain from amyloid β deposits and peroxidation in mice with Alzheimer-like lesions. Am J Alzheimers Dis Other Demen 2015, 30, 183–191. [Google Scholar] [CrossRef]
- Maiuolo, J.; Bosco, F.; Guarnieri, L.; Nucera, S.; Ruga, S.; Oppedisano, F.; Tucci, L.; Muscoli, C.; Palma, E.; Giuffrè, A.M. , et al. Protective Role of an Extract Waste Product from. Plants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Dai, Y.; Chen, S.R.; Chai, L.; Zhao, J.; Wang, Y. Overview of pharmacological activities of. Crit Rev Food Sci Nutr 2019, 59, S17–S29. [Google Scholar] [CrossRef] [PubMed]
- Hossain, R.; Quispe, C.; Herrera-Bravo, J.; Beltrán, J.F.; Islam, M.T.; Shaheen, S.; Cruz-Martins, N.; Martorell, M.; Kumar, M.; Sharifi-Rad, J. , et al. Neurobiological Promises of the Bitter Diterpene Lactone Andrographolide. Oxid Med Cell Longev 2022, 2022, 3079577. [Google Scholar] [CrossRef]
- Sani, D.; Khatab, N.I.O.; Kirby, B.P.; Yong, A.; Hasan, S.; Basri, H.; Stanslas, J. A standardised. J Adv Res 2019, 16, 87–97. [Google Scholar] [CrossRef]
- Bera, R.; Ahmed, S.K.; Sarkar, L.; Sen, T.; Karmakar, S. Pharmacokinetic analysis and tissue distribution of andrographolide in rat by a validated LC-MS/MS method. Pharm Biol 2014, 52, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.Y.; Tan, M.G.; Banks, W.A.; Wong, W.S.; Wong, P.T.; Lai, M.K. Andrographolide attenuates LPS-stimulated up-regulation of C-C and C-X-C motif chemokines in rodent cortex and primary astrocytes. J Neuroinflammation 2016, 13, 34. [Google Scholar] [CrossRef]
- Wong, S.Y.; Chan, S.J.; Wong, W.S.; Wong, P.T.; Lai, M.K. Andrographolide attenuates interleukin-1β-stimulated upregulation of chemokine CCL5 and glial fibrillary acidic protein in astrocytes. Neuroreport 2014, 25, 881–886. [Google Scholar] [CrossRef]
- Das, S.; Mishra, K.P.; Ganju, L.; Singh, S.B. Andrographolide - A promising therapeutic agent, negatively regulates glial cell derived neurodegeneration of prefrontal cortex, hippocampus and working memory impairment. J Neuroimmunol 2017, 313, 161–175. [Google Scholar] [CrossRef]
- Adedayo, B.C.; Jesubowale, O.S.; Adebayo, A.A.; Oboh, G. Effect of Andrographis paniculata leaves extract on neurobehavioral and biochemical indices in scopolamine-induced amnesic rats. J Food Biochem 2021, 45, e13280. [Google Scholar] [CrossRef]
- Vallée, A. Neuroinflammation in Schizophrenia: The Key Role of the WNT/β-Catenin Pathway. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Hurley, L.L.; Tizabi, Y. Neuroinflammation, neurodegeneration, and depression. Neurotox Res 2013, 23, 131–144. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.; Dai, Z.; Sui, Y. Andrographolide improves PCP-induced schizophrenia-like behaviors through blocking interaction between NRF2 and KEAP1. J Pharmacol Sci 2021, 147, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Liu, J.; Yuan, X.; Liu, W.; Guo, W. Andrographolide triggers autophagy-mediated inflammation inhibition and attenuates chronic unpredictable mild stress (CUMS)-induced depressive-like behavior in mice. Toxicol Appl Pharmacol 2019, 379, 114688. [Google Scholar] [CrossRef] [PubMed]
- Adedayo, B.C.; Ogunsuyi, O.B.; Akinniyi, S.T.; Oboh, G. Effect of. Drug Chem Toxicol 2022, 45, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Iruretagoyena, M.I.; Tobar, J.A.; González, P.A.; Sepúlveda, S.E.; Figueroa, C.A.; Burgos, R.A.; Hancke, J.L.; Kalergis, A.M. Andrographolide interferes with T cell activation and reduces experimental autoimmune encephalomyelitis in the mouse. J Pharmacol Exp Ther 2005, 312, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, Y.; Zhao, Y.; Zhang, Y.; Liu, Y.; Ma, F.; Wang, X.; Fu, J. Andrographolide ameliorates neuroinflammation in APP/PS1 transgenic mice. Int Immunopharmacol 2021, 96, 107808. [Google Scholar] [CrossRef]
- Lu, J.; Ma, Y.; Wu, J.; Huang, H.; Wang, X.; Chen, Z.; Chen, J.; He, H.; Huang, C. A review for the neuroprotective effects of andrographolide in the central nervous system. Biomed Pharmacother 2019, 117, 109078. [Google Scholar] [CrossRef]
- Dey, A.; Chen, R.; Li, F.; Maitra, S.; Hernandez, J.F.; Zhou, G.C.; Vincent, B. Synthesis and Characterization of Andrographolide Derivatives as Regulators of βAPP Processing in Human Cells. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Seo, J.Y.; Pyo, E.; An, J.P.; Kim, J.; Sung, S.H.; Oh, W.K. Andrographolide Activates Keap1/Nrf2/ARE/HO-1 Pathway in HT22 Cells and Suppresses Microglial Activation by A. Mediators Inflamm 2017, 2017, 5906189. [Google Scholar] [CrossRef]
- Yang, S.L.; Kuo, F.H.; Chen, P.N.; Hsieh, Y.H.; Yu, N.Y.; Yang, W.E.; Hsieh, M.J.; Yang, S.F. Andrographolide suppresses the migratory ability of human glioblastoma multiforme cells by targeting ERK1/2-mediated matrix metalloproteinase-2 expression. Oncotarget 2017, 8, 105860–105872. [Google Scholar] [CrossRef]
- Xu, Y.; Tang, D.; Wang, J.; Wei, H.; Gao, J. Neuroprotection of Andrographolide Against Microglia-Mediated Inflammatory Injury and Oxidative Damage in PC12 Neurons. Neurochem Res 2019, 44, 2619–2630. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.; Killick, R.; Lovestone, S. The GSK3 hypothesis of Alzheimer’s disease. J Neurochem 2008, 104, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Rojas, C.; Schüller, A.; Lindsay, C.B.; Ureta, R.C.; Mejías-Reyes, C.; Hancke, J.; Melo, F.; Inestrosa, N.C. Andrographolide activates the canonical Wnt signalling pathway by a mechanism that implicates the non-ATP competitive inhibition of GSK-3β: autoregulation of GSK-3β in vivo. Biochem J 2015, 466, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, S.B.; Valenzuela-Bezanilla, D.; Mardones, M.D.; Varela-Nallar, L. Role of Wnt Signaling in Adult Hippocampal Neurogenesis in Health and Disease. Front Cell Dev Biol 2020, 8, 860. [Google Scholar] [CrossRef]
- Esquerda-Canals, G.; Montoliu-Gaya, L.; Güell-Bosch, J.; Villegas, S. Mouse Models of Alzheimer’s Disease. J Alzheimers Dis 2017, 57, 1171–1183. [Google Scholar] [CrossRef]
- Castro-Fuentes, R.; Socas-Pérez, R. Octodon degus: a strong attractor for Alzheimer research. Basic Clin Neurosci 2013, 4, 91–96. [Google Scholar] [PubMed]
- Varela-Nallar, L.; Arredondo, S.B.; Tapia-Rojas, C.; Hancke, J.; Inestrosa, N.C. Andrographolide Stimulates Neurogenesis in the Adult Hippocampus. Neural Plast 2015, 2015, 935403. [Google Scholar] [CrossRef]
- Serrano, F.G.; Tapia-Rojas, C.; Carvajal, F.J.; Hancke, J.; Cerpa, W.; Inestrosa, N.C. Andrographolide reduces cognitive impairment in young and mature AβPPswe/PS-1 mice. Mol Neurodegener 2014, 9, 61. [Google Scholar] [CrossRef]
- Rivera, D.S.; Lindsay, C.; Codocedo, J.F.; Morel, I.; Pinto, C.; Cisternas, P.; Bozinovic, F.; Inestrosa, N.C. Andrographolide recovers cognitive impairment in a natural model of Alzheimer’s disease (Octodon degus). Neurobiol Aging 2016, 46, 204–220. [Google Scholar] [CrossRef]
- Lindsay, C.B.; Zolezzi, J.M.; Rivera, D.S.; Cisternas, P.; Bozinovic, F.; Inestrosa, N.C. Andrographolide Reduces Neuroinflammation and Oxidative Stress in Aged Octodon degus. Mol Neurobiol 2020, 57, 1131–1145. [Google Scholar] [CrossRef]
- Fava, A.; Colica, C.; Plastino, M.; Messina, D.; Cristiano, D.; Opipari, C.; Vaccaro, A.; Gorgone, G.; Bosco, F.; Fratto, A. , et al. Cognitive impairment is correlated with insulin resistance degree: the “PA-NICO-study”. Metab Brain Dis 2017, 32, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Liang, J.; Zhou, B. Glucose Metabolic Dysfunction in Neurodegenerative Diseases-New Mechanistic Insights and the Potential of Hypoxia as a Prospective Therapy Targeting Metabolic Reprogramming. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Hu, Y.; Wang, B.; Wang, S.; Zhang, X. Metabolic Dysregulation Contributes to the Progression of Alzheimer’s Disease. Front Neurosci 2020, 14, 530219. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, P.; Sabater, L.; Mathieu, E.; Faller, P.; Hureau, C. Why the Ala-His-His Peptide Is an Appropriate Scaffold to Remove and Redox Silence Copper Ions from the Alzheimer’s-Related Aβ Peptide. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Cunnane, S.C.; Sieber, C.C.; Swerdlow, R.H.; Cruz-Jentoft, A.J. Mild cognitive impairment: when nutrition helps brain energy rescue-a report from the EuGMS 2020 Congress. Eur Geriatr Med 2021, 12, 1285–1292. [Google Scholar] [CrossRef]
- Bosco, D.; Plastino, M.; Bosco, F.; Consoli, A.; Labate, A.; Pirritano, D.; Consoli, D.; Fava, A. Bell’s palsy: a manifestation of prediabetes? Acta Neurol Scand 2011, 123, 68–72. [Google Scholar] [CrossRef]
- Patel, R.; Kaur, K.; Singh, S. Protective effect of andrographolide against STZ induced Alzheimer’s disease in experimental rats: possible neuromodulation and Aβ. Inflammopharmacology 2021, 29, 1157–1168. [Google Scholar] [CrossRef]
- Thakur, A.K.; Rai, G.; Chatterjee, S.S.; Kumar, V. Beneficial effects of an Andrographis paniculata extract and andrographolide on cognitive functions in streptozotocin-induced diabetic rats. Pharm Biol 2016, 54, 1528–1538. [Google Scholar] [CrossRef]
- Gherardelli, C.; Cisternas, P.; Gutiérrez, J.; Martinez, M.; Inestrosa, N.C. Andrographolide restores glucose uptake in rat hippocampal neurons. J Neurochem 2021, 157, 1222–1233. [Google Scholar] [CrossRef]
- Stoker, T.B.; Greenland, J.C. Parkinson’s Disease: Pathogenesis and Clinical Aspects. 2018.
- Pajares, M.; I Rojo, A.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9. [Google Scholar] [CrossRef]
- Wang, T.; Liu, B.; Zhang, W.; Wilson, B.; Hong, J.S. Andrographolide reduces inflammation-mediated dopaminergic neurodegeneration in mesencephalic neuron-glia cultures by inhibiting microglial activation. J Pharmacol Exp Ther 2004, 308, 975–983. [Google Scholar] [CrossRef]
- Ketterman, A.J.; Wongtrakul, J.; Saisawang, C. Phytochemical andrographolide modulates NF-κB and JNK in human neuroblastoma SH-SY5Y cells, a cell model for Parkinson’s disease. Heliyon 2020, 6, e04121. [Google Scholar] [CrossRef]
- Ahmed, S.; Kwatra, M.; Ranjan Panda, S.; Murty, U.S.N.; Naidu, V.G.M. Andrographolide suppresses NLRP3 inflammasome activation in microglia through induction of parkin-mediated mitophagy in in-vitro and in-vivo models of Parkinson disease. Brain Behav Immun 2021, 91, 142–158. [Google Scholar] [CrossRef]
- Zhang, Z.; Lai, D.; Wang, L.; Yu, P.; Zhu, L.; Guo, B.; Xu, L.; Zhou, L.; Sun, Y.; Lee, S.M. , et al. Neuroprotective effects of the andrographolide analogue AL-1 in the MPP⁺/MPTP-induced Parkinson’s disease model in vitro and in mice. Pharmacol Biochem Behav 2014, 122, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Hyun, J.; Park, J.; Jung, S.; Oh, Y.; Kim, Y.; Ryu, S.H.; Kim, S.H.; Jeong, E.I.; Jo, D.G. , et al. Aberrant role of pyruvate kinase M2 in the regulation of gamma-secretase and memory deficits in Alzheimer’s disease. Cell Rep 2021, 37, 110102. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Varma, V.R.; Varma, S.; Casanova, R.; Dammer, E.; Pletnikova, O.; Chia, C.W.; Egan, J.M.; Ferrucci, L.; Troncoso, J. , et al. Evidence for brain glucose dysregulation in Alzheimer’s disease. Alzheimers Dement 2018, 14, 318–329. [Google Scholar] [CrossRef]
- Kumar, V.; Kim, S.H.; Bishayee, K. Dysfunctional Glucose Metabolism in Alzheimer’s Disease Onset and Potential Pharmacological Interventions. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Pluta, R. Cerebral Ischemia. 2021.
- Yen, T.L.; Hsu, W.H.; Huang, S.K.; Lu, W.J.; Chang, C.C.; Lien, L.M.; Hsiao, G.; Sheu, J.R.; Lin, K.H. A novel bioactivity of andrographolide from Andrographis paniculata on cerebral ischemia/reperfusion-induced brain injury through induction of cerebral endothelial cell apoptosis. Pharm Biol 2013, 51, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.J.; Wong, W.S.; Wong, P.T.; Bian, J.S. Neuroprotective effects of andrographolide in a rat model of permanent cerebral ischaemia. Br J Pharmacol 2010, 161, 668–679. [Google Scholar] [CrossRef]
- Yang, M.Y.; Yu, Q.L.; Huang, Y.S.; Yang, G. Neuroprotective effects of andrographolide derivative CX-10 in transient focal ischemia in rat: Involvement of Nrf2/AE and TLR/NF-κB signaling. Pharmacol Res 2019, 144, 227–234. [Google Scholar] [CrossRef]
- Chern, C.M.; Liou, K.T.; Wang, Y.H.; Liao, J.F.; Yen, J.C.; Shen, Y.C. Andrographolide inhibits PI3K/AKT-dependent NOX2 and iNOS expression protecting mice against hypoxia/ischemia-induced oxidative brain injury. Planta Med 2011, 77, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Yen, T.L.; Chen, R.J.; Jayakumar, T.; Lu, W.J.; Hsieh, C.Y.; Hsu, M.J.; Yang, C.H.; Chang, C.C.; Lin, Y.K.; Lin, K.H. , et al. Andrographolide stimulates p38 mitogen-activated protein kinase-nuclear factor erythroid-2-related factor 2-heme oxygenase 1 signaling in primary cerebral endothelial cells for definite protection against ischemic stroke in rats. Transl Res 2016, 170, 57–72. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



