
Submitted:
30 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
In the case of indigenous grapevine varieties, the aim of clonal selection is twofold: to improve their technological cultivation problem while preserving the varietal character. As the sensitivity of perennial crops to climate change becomes more pronounced, clonal selection, which is already very time-consuming for perennial woody crops, may take even longer. In Badacsony, clonal selection breeding of the ‘Kéknyelű’ and ‘Juhfark’ varieties was started in the early 2000s. The problem mentioned above in cultivation technology is the poor fertility of ‘Kéknyelű’ (functionally female-flowered variety), while in the case of ‘Juhfark’ the high susceptibility to grey rot (Botrytis). Based on 11 years of data significant difference in yield between clone B.2. and the base variety was found. In this comparison the year clusters also gave different values, as in years belonging to cluster 1 the ’Kéknyelű’ yielded more than in another year. Both clones of ‘Kéknyelű’ matured with significantly lower pH compared to the base variety. ‘Kéknyelű' is renowned for the distinctive variety character of its wine, which is said to be acidic. Given the acidic nature of ‘Kéknyelű’ wine and the expected increase in must degrees and pH due to climate change, these distinctions may be beneficial in the future. Daily meteorological parameters were also recorded in the years under study, and different indexes were calculated for both of varieties for different phenological periods. The years were classified into 3 clusters and the harvest data were analyzed within each cluster. In the case of 'Kéknyelű' our results revealed significant differences in yield between vintage clusters for clone B.2. However, clone B.1 demonstrated distinct KMW and pH values. Both 'Kéknyelű’ clones have produced a substantially higher incidence of Botrytis infection in cluster 2 years than in cluster 3 years. Botrytis infection only showed statistically significant differences between year clusters for ‘Juhfark’. It is intriguing that in most years, the rotting rate was lower in both clones, especially in year clusters 1 and 3 when the overall rate of Botrytis infection was quite low. Our results highlighted the year-round sensitivity of the two varieties studied and the difficulties of clonal selection.
Keywords:
climate change
; global warming
; indigenous cultivars
; berry composition
; clonal selection
; grape breeding
1. Introduction
Since cultivated grape (Vitis vinifera L.) is a climate-sensitive crop, climate change has a major impact on the cultivation and the cultivability of individual cultivars. The main adverse effects of climate change are global warming and the increasing frequency and severity of extreme weather events, including drought, which is affecting the growing season more frequently and more severely, and is having a negative impact on the quality of white grapes in particular, due to acidity loss [1,2,3,4,5].
This is particularly the case for autochthonous varieties, where the limits of environmental tolerance are even narrower, which is why these varieties are grown only in a narrower geographical area (the growing area) - where they can be economically grown in most years. However, the great advantage of indigenous cultivars is their curiosity and, for cultural reasons, their significant marketing value [6,7,8]. However, for autochthonous varieties, it is very important to be distinguishable, as this is their speciality, which adds value to the market [9].
Much damage has been done to the nymph of the ‘Kéknyelű’ variety by the rumour that it may be identical to the Picolit variety grown in Friuli-Venezia Giulia region in Italy. This assumption was based on Goethe’s (1887) claim that the two varieties were identical [10]. Later, Németh (1967) also claimed that based on the descriptions of ‘Kéknyelű’ and Picolit are morphologically very similar, but in his opinion the identity or difference can be confirmed by a careful morphological comparison of the two cultivars planted side by side [11]. Later molecular markers were used to prove, that the two cultivars are different [12,13].
Similarly, many producers have confused the ‘Juhfark’ with the ‘Csomorika’, another ancient Hungarian vine, although the difference is morphologically visible, as the two cultivars belong to different geo-ecological groups - ‘Juhfark’ to V. vinifera L. proles orientalis Negr., ‘Csomorika’ to V. vinifera L. proles pontica Negr [10,13]. The morphological characteristics and production values of the two varieties were studied by Varga et al. [15,16]. Of course, at the molecular marker level, the two varieties are well separated [17,18,19,20,21].
The climate change and the reduced environmental tolerance of autochthonous varieties mean that their sensitivity to annual variation is even more pronounced, and therefore their selection breeding is delayed. Their low environmental tolerance is usually manifested in some specific problem, in our case the poor fertility of ‘Kéknyelű’ (a functional female-flowered variety-Figure 1.A) and the high susceptibility to Botrytis of ‘Juhfark’ (Figure 1.B) [8,22].
Figure 1.
A. The functional female flowers of ‘Kéknyelű’ causes fertilization problems. B. The dense clusters of ‘Juhfark’ also make it very susceptible to Botrytis.
Figure 1.
A. The functional female flowers of ‘Kéknyelű’ causes fertilization problems. B. The dense clusters of ‘Juhfark’ also make it very susceptible to Botrytis.

In Hungary, clonotype selection (different from polyclonal selection) and clonal selection (3 or 4-step) were the most commonly used intra-variety selection methods. Clonotype selection requires a base population already selected (at least from a phytosanitary point of view) and involves grouping grape varieties according to one or more important characteristics, such as flower type. This method is more efficient than mass selection (polyclonal selection), and was developed by Kozma in 1948 on the basis of the floral biology of Furmint and Kadarka varieties [23,24]. The method of clone selection was adapted by the Hungarians from Germany. Márton Németh developed a four-step individual method, Ottokár Luntz later reduced the number of steps to three [23,24,25].
Both methods have several advantages and disadvantages. While clonotype selection preserves much of the genetic variability in the starting material, allowing for better adaptability, the improvement of the starting population is much slower, so selection takes more time. However, for genetically degraded varieties that exist only in very small populations, this method can be used to produce the amount of reproductive material needed to “bring the variety back into production” in a relatively shorter time. Clonal selection breeding, although it is faster and allows for earlier clones, reduces the genetic variability of the variety, which reduces the environmental adaptability. From a virus-elimination point of view, clonal selection is preferable as it only requires the de-viralisation of one genotype, whereas de-viralisation of a clone type is impossible or at least very costly [24,27,28].
In practice, clonal selection is often aided by the correlation between certain morphological traits and cultivation traits [23,29]. Despite the consumer demand for Vignoles, its production is limited by the susceptibility of the bunches to rot, which is linked to the compact bunch structure. The selection objective here was to select a clone of the variety with loose clusters [28,29]. Similar considerations have been made for the selection of looser clustered types for the ‘Juhfark’. For the ‘Kéknyelű’ variety, we also looked at cluster tightness, but here we were looking for types with more fertile, and therefore slightly tighter clusters [30].
Whereas selection methods used to be aided by knowledge of the correlation of morphological traits [23] , in modern times molecular techniques can also aid clonal discrimination [33] and selection, although their application is limited compared to cross-breeding. For example, ATR-MIR spectroscopy combined with partial least squares discriminant analysis (PLS-DA) has been used to discriminate the origin and vintage year of ‘Tempranillo’ grape clones, and partial least squares (PLS) regression to predict soluble solids (SS), pH and titratable acidity (TA) [34]. To aid clonal selection of Croatian indigenous varieties, AFLP and S-SAP markers were used to assess intra-varietal genetic variability in Pejić et al. [35]. They found significant genetic variability, which makes clonal selection an effective method for improving yield and quality of existing conventional varieties. They also investigated the extent of intra-varietal genetic variability in the grapevine cultivar ‘Keshmeshi’ using 23 SSR and 7 AFLP markers, which clearly distinguished the berry skin colored clone from the others [36]. Similarly, the intraspecific genetic variability of a total of 47 clones belonging to the Pinot noir, Pinot gris, Pinot blanc, Meunier, Teroldego and Gewürztraminer varieties was assessed using SSR markers [37].
The dynamic genome of the grape is characterized by single mutations that occur only once in a clone. The frequent occurrence of mutations in different clones suggests that the mutations may be locus specific. To investigate this phenomenon, 86 Riesling clones were analyzed using ten AFLP primer combinations [38]. 38.5% of the polymorphic marker bands showed single mutations and 17% showed locus-specific mutations, confirming the observation that the grapevine genome is rather dynamic (at least in the case of the Riesling variety). This helps selection breeding, but can cause problems in cultivation due to the instability of the clones.
Clone selection of the ‘Kéknyelű’ variety was started in 2003, based on the previous clonotype selection, while clone selection of the ‘Juhfark’ variety was started in 2005 in Badacsony. Since 2011, 2 ‘Kéknyelű’ and 2 ‘Juhfark’ clones are being tested in small plot experiments. Our aim is to present the results of the breeding work so far based on the harvest results and observations.
2. Materials and Methods
2.1. Experinemtal site, vineyard and growing conditions
In Badacsony, the results of small plots (20 vines) of clones of ‘Kéknyelű’ and ‘Juhfark’ from the Hungarian University of Agricultural and Life Sciences, Institute for Viticulture and Oenology, Research Station and the results of plantations of both varieties of 0.3-0.3 ha in the same area were examined. All the vineyards studied were with the same 2m x 1m vine spacing and Teleki 5C (E20) rootstock and training system (‘Juhfark’-middle-high cordon; ‘Kéknyelű’-umbrella). Bud loading was 7 buds/m2=14 buds/vines for both of the cultivars (‘Juhfark’ – 2 pieces of 3 bud spurs + 4 pieces of 2 bud spurs pro vine; ‘Kéknyelű’ – 1 piece of 12 bud cane + 1 piece of 2 bud spur pro vine).
2.2. Experimental harvest, measures
The date of the key phenological stages: budburst, beginning of flowering, end of flowering, veraison and harvest (maturity) were visually stated and recorded for both of the cultivars. The harvest date (maturity) was determined by refractometric measurement of the sugar content of 20 berries randomly sampled per cultivar.
During the experimental harvests, the following parameters were determined: yield (kg/m2), sugar content of the juice (Klosternuburger Mostwaage = KMW g / 100 g), titratable acidity of must (g/l), pH, and the degree of rot (Botrytis cinerea) infection (%) was estimated visually.
12 years (2011-2022) of meteorological data (daily minimum, maximum and mean temperature; daily precipitation) were recorded by automatic meteorological station (Luft) installed in the same site as the vineyards (Badacsony, Hungary).
2.3. Calculations, data analyses
To determine the vintage effect, the following indices were calculated for each variety at different phenological stages of the vine: growing degree days (GDD), Huglin index, hydrothermal coefficient, and the cumulative rainfall during flowering for ‘Kéknyelű’ and during maturation of the berries (from version to full maturity) for ‘Juhfark’. Years were grouped by cultivars based on these biometeorological indexes and values forming 3 clusters and principal component analysis also was performed.
Harvest data were also analyzed by the 3 groups (clusters) of years and by cultivar separately, always with a particular clone always split to the base. In this way, it was also possible to compare the vintage sensitivity of each clone.
The homogeneity of variances and the distribution of the harvest results data (normality test) were checked by Levene test and Shapiro-Wilk test respectively, and then, as these do not meet the basic conditions for analysis of variance, data were evaluated by Analysis of Variance of Aligned Rank Transformed Data (ART-ANOVA). Where the ART-ANOVA results indicated that the expected values differed at a significance level of at least 90%, the expected values were compared pairwise using “Aligned Ranked Transform Contrasts” test [39,40].
All of the results were analyzed and evaluated using the R software package [41].
3. Results
3.1. Evaluation of the meteorological data and indexes
The recorded meteorological data are detailed in Supplementary Table 1.
The date of the phenological stages of ‘Kéknyelű’ and ‘Juhfark’ are in Table 1 and Table 2 respectively.

Table 1.
Date of the key phenological stages of the ‘Kéknyelű’ variety (Badacsony, 2011-2022).
| Year | Budburst | Beginning of flowering | End of flowering | Veraison | Maturity/Harvest |
|---|---|---|---|---|---|
| 2011 | 04. 13. | 05. 30. | 06. 13. | 07. 30. | 09. 21. |
| 2012 | 04. 11. | 05. 29. | 06. 07. | 07. 27. | 09. 06. |
| 2013 | 04. 23. | 06. 07. | 06. 17. | 08. 08. | 10. 01. |
| 2014 | 04. 07. | 06. 04. | 06. 15. | 08. 05. | 09. 22. |
| 2015 | 04. 20. | 06. 04. | 06. 13. | 08. 04. | 09. 12. |
| 2017 | 04. 06. | 06. 08. | 06. 19. | 08. 01. | 09. 20. |
| 2018 | 04. 16. | 05. 21. | 05. 28. | 07. 16. | 09. 20. |
| 2019 | 04. 12. | 06. 06. | 06. 20. | 07. 29. | 10. 02. |
| 2020 | 04. 09. | 06. 04. | 06. 12. | 08. 04. | 09. 22. |
| 2021 | 04. 23. | 06. 13. | 06. 23. | 08. 06. | 09. 30. |
| 2022 | 04. 14. | 06. 02. | 06. 09. | 07. 29. | 09. 22. |
Table 2.
Date of the key phenological stages of the ‘Juhfark’ variety (Badacsony, 2011-2022).
| Year | Budburst | Beginning of flowering | End of flowering | Veraison | Maturity/Harvest |
|---|---|---|---|---|---|
| 2011 | 04. 11. | 05. 27. | 06. 08. | 07. 25. | 09. 15. |
| 2012 | 04. 04. | 05. 24. | 06. 06. | 07. 23. | 09. 10. |
| 2013 | 04. 18. | 06. 04. | 06. 14. | 07. 27. | 09. 27. |
| 2014 | 04. 04. | 05. 27. | 06. 09. | 07. 30. | 09. 16. |
| 2015 | 04. 18. | 06. 02. | 06. 06. | 07. 31. | 09. 09. |
| 2017 | 04. 04. | 06. 06. | 06. 15. | 07. 25. | 09. 13. |
| 2018 | 04. 12. | 05. 18. | 05. 25. | 07. 09. | 09. 05. |
| 2020 | 03. 30. | 05. 29. | 06. 09. | 07. 24. | 09. 10. |
| 2021 | 04. 17. | 06. 11. | 06. 18. | 07. 29. | 09. 09. |
| 2022 | 04. 09. | 05. 25. | 06. 02. | 07. 19. | 09. 01. |
The values of the calculated indexes are summarized in Table 3.
Table 3.
Values of the indexes for ‘Kéknyelű’ (A) and ‘Juhfark’ (B).
| A | |||||||||||||
| YEAR | GDD1 | GDD2 | GDD3 | GDD4 | HUG1 | HUG2 | HUG3 | HUG4 | HTC1 | HTC2 | HTC3 | HTC4 | P2 |
| 2011 | 298.53 | 165.55 | 543.61 | 651.80 | 450.89 | 217.43 | 734.16 | 877.33 | 0.20 | 0.28 | 0.82 | 0.46 | 8.60 |
| 2012 | 284.50 | 82.69 | 695.46 | 590.05 | 441.66 | 119.49 | 910.64 | 782.54 | 0.85 | 0.42 | 0.75 | 0.04 | 7.20 |
| 2013 | 297.59 | 118.33 | 754.67 | 436.35 | 448.66 | 159.35 | 995.75 | 647.92 | 1.12 | 0.39 | 0.42 | 1.28 | 8.60 |
| 2014 | 274.50 | 151.52 | 604.13 | 414.16 | 441.32 | 198.78 | 807.10 | 591.50 | 1.03 | 0.00 | 1.07 | 3.50 | 0.00 |
| 2015 | 299.63 | 125.32 | 670.72 | 488.73 | 449.31 | 162.34 | 886.58 | 665.45 | 0.98 | 0.00 | 0.32 | 0.81 | 0.00 |
| 2016 | 304.80 | 63.50 | 697.20 | 471.00 | 479.17 | 90.67 | 916.86 | 661.61 | 1.39 | 1.11 | 0.94 | 0.60 | 14.80 |
| 2017 | 350.57 | 124.98 | 592.36 | 547.79 | 516.65 | 169.15 | 787.37 | 750.64 | 0.64 | 0.11 | 0.94 | 0.99 | 2.60 |
| 2018 | 298.11 | 80.03 | 556.93 | 882.49 | 419.64 | 107.75 | 753.81 | 1216.37 | 1.30 | 0.84 | 1.26 | 1.17 | 12.60 |
| 2019 | 237.70 | 201.50 | 513.80 | 708.70 | 362.78 | 259.04 | 683.71 | 992.30 | 2.05 | 0.25 | 1.07 | 0.77 | 8.50 |
| 2020 | 274.50 | 71.70 | 637.00 | 568.80 | 463.10 | 102.80 | 857.69 | 783.14 | 0.74 | 1.58 | 1.18 | 0.62 | 24.00 |
| 2021 | 275.20 | 149.80 | 636.10 | 526.40 | 432.23 | 195.72 | 829.08 | 762.04 | 0.85 | 0.00 | 0.69 | 0.51 | 0.00 |
| 2022 | 300.50 | 80.50 | 688.10 | 613.50 | 443.73 | 106.26 | 906.52 | 830.66 | 1.28 | 2.34 | 0.57 | 0.83 | 35.20 |
| B | |||||||||||||
| YEAR | GDD1 | GDD2 | GDD3 | GDD4 | H1 | H2 | H3 | H4 | HTC1 | HTC2 | HTC3 | HTC4 | P4 |
| 2011 | 280.37 | 135.97 | 551.99 | 645.11 | 427.78 | 182.48 | 742.82 | 863.10 | 0.19 | 0.40 | 0.49 | 0.72 | 83.80 |
| 2012 | 256.88 | 108.36 | 653.20 | 693.47 | 401.71 | 159.26 | 854.12 | 922.30 | 1.06 | 0.33 | 0.72 | 0.11 | 12.80 |
| 2013 | 306.02 | 93.10 | 582.11 | 651.14 | 464.79 | 129.41 | 773.54 | 917.78 | 1.19 | 0.50 | 0.52 | 0.77 | 97.60 |
| 2014 | 239.56 | 113.87 | 608.82 | 448.57 | 387.33 | 163.94 | 811.13 | 625.89 | 1.26 | 0.02 | 1.07 | 3.12 | 289.60 |
| 2015 | 273.06 | 54.75 | 711.22 | 531.04 | 418.80 | 71.01 | 937.93 | 712.71 | 1.03 | 0.00 | 0.31 | 0.75 | 69.60 |
| 2016 | 274.60 | 75.50 | 632.30 | 484.10 | 436.59 | 107.10 | 835.80 | 668.80 | 1.19 | 1.56 | 0.97 | 0.77 | 70.20 |
| 2017 | 337.33 | 97.22 | 541.70 | 615.51 | 498.93 | 133.05 | 721.19 | 830.92 | 0.66 | 0.05 | 0.95 | 0.63 | 70.40 |
| 2018 | 298.68 | 66.44 | 513.95 | 799.28 | 420.36 | 91.89 | 690.87 | 1082.17 | 1.29 | 0.92 | 1.18 | 1.43 | 200.50 |
| 2019 | 234.60 | 187.50 | 515.90 | 643.40 | 356.53 | 238.93 | 688.70 | 870.35 | 2.22 | 0.00 | 1.07 | 0.87 | 102.50 |
| 2020 | 256.40 | 75.50 | 509.80 | 587.90 | 435.12 | 115.03 | 698.62 | 790.76 | 0.64 | 1.44 | 1.06 | 1.01 | 107.40 |
| 2021 | 255.70 | 84.70 | 624.10 | 466.80 | 411.50 | 112.88 | 810.18 | 648.69 | 1.19 | 0.32 | 0.30 | 0.90 | 79.90 |
| 2022 | 249.60 | 58.90 | 599.40 | 630.40 | 377.74 | 83.58 | 794.85 | 819.68 | 1.17 | 1.40 | 0.96 | 0.60 | 64.70 |
indexes: GDD-growing degree days; H-Huglin index; HTC-hydrothermal coefficient; P-cumulative rainfall (precipitation). Phenpohase codes: 1: from budburst to the beginning of flowering, 2 flowering; 3 from the end of flowering to veraison; from veraison to harvest (maturity).
The results of clustering of years based on the index values detailed in Table 3. Based on this clustering 3 groups (clusters) were formed for both of the cultivars. Cluster 1 contains the years 2008, 2020 and 2022; cluster 2 the years 2011,2014, 2019 and 2021; as cluster 3 the years 2012, 2013, 2015, 2016 and 2017 for Kéknyelű. In the case of ‘Juhfark’ the clusters are the following: cluster 1: 2012, 2015, 2016, 2021 and 2022; cluster 2: 2011, 2014, 2019 and 2021 respectively (see Figure 2 A and B).
Figure 2.
Result of PCA (Principal Component Analyses) of the 3 cluster of years for ‘Kéknyelű’ (A) and ‘Juhfark’ (B) respectively.
Figure 2.
Result of PCA (Principal Component Analyses) of the 3 cluster of years for ‘Kéknyelű’ (A) and ‘Juhfark’ (B) respectively.
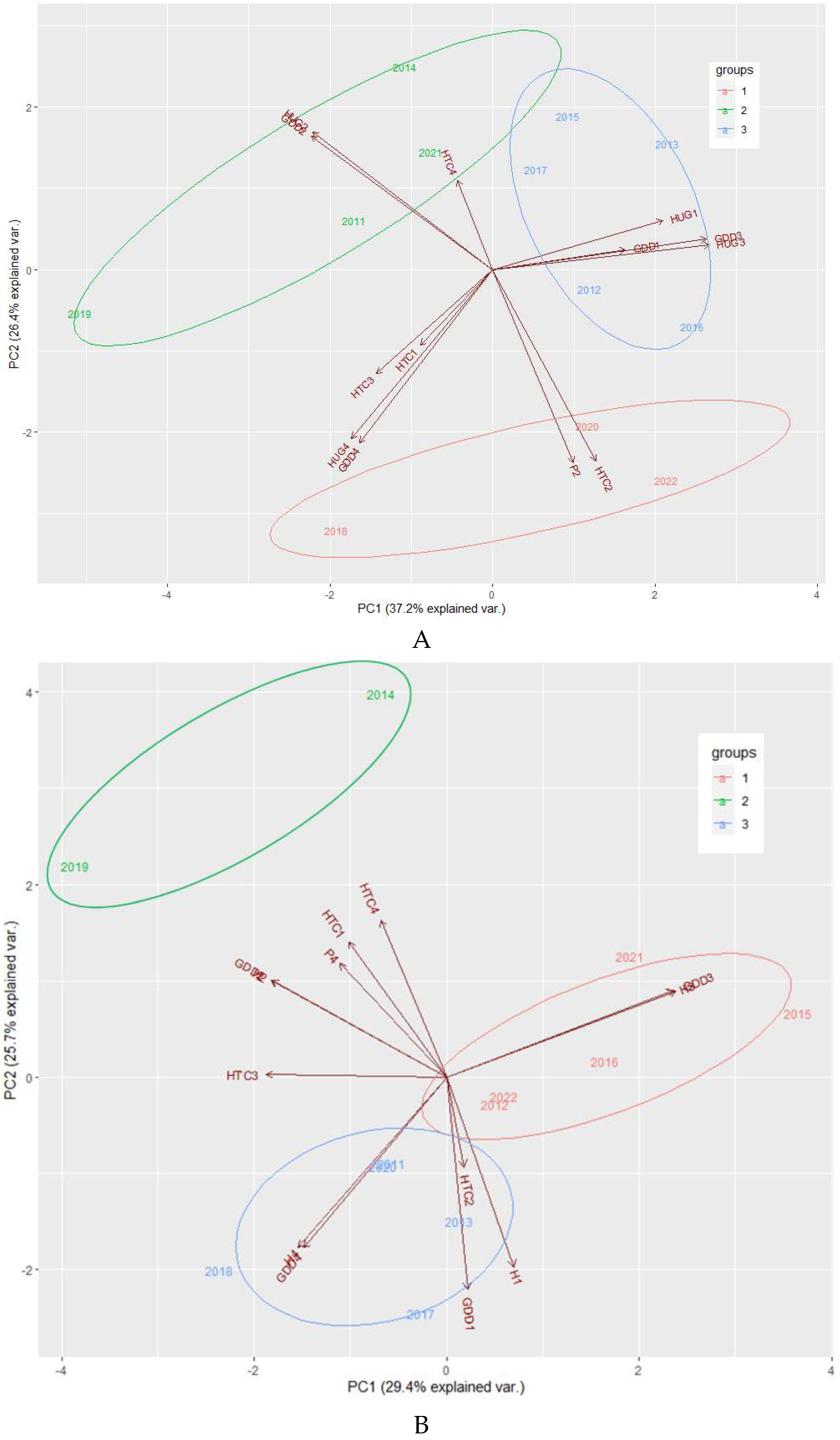
The graphical representation of the result of PCA of the 3 cluster of years is shown in Figure 2 A and B. As can be seen in the figure, the PC1 and PC2 principal components explain 37.2% and 26.4% of the variance between years for ‘Kéknyelű’ and 29.4% and 25.4% for ‘Juhfark’, respectively. For both of the cultivars the years can be clearly distinguished using these two components.
3.2. Evaluation of the harvest results of the ‘Kéknyelű’ variety
The harvest results of the ‘Kéknyelű’ variety have been evaluated on the basis of eleven years of data between 2011 and 2022. In 2016, the bird damage in the plantation was such that we were unable to harvest any appreciable amount of fruit. The harvest results are summarized in Table 4.
Table 4.
Harvest results of the ‘Kéknyelű’ variety (Badacsony, 2011-2022).
| Clone | Yield kg/m2 |
Sugar content of the juice KMW |
Titratable acid contant of the juice g/l |
pH | Botrytis infection % |
Yield kg/m2 |
|---|---|---|---|---|---|---|
| B.1. | 2011 | 0.88 | 20.20 | 7.25 | 3.08 | 10.00 |
| B.2. | 2011 | 1.26 | 17.40 | 7.09 | 3.07 | 10.00 |
| Base | 2011 | 0.99 | 18.60 | 6.59 | 3.38 | 10.00 |
| B.1. | 2012 | 0.95 | 18.30 | 6.52 | 3.55 | 0.00 |
| B.2. | 2012 | 1.10 | 17.50 | 6.36 | 3.67 | 0.00 |
| Base | 2012 | 1.08 | 18.50 | 4.64 | 3.47 | 0.00 |
| B.1. | 2013 | 1.39 | 18.90 | 11.15 | 3.26 | 0.00 |
| B.2. | 2013 | 1.61 | 17.70 | 10.36 | 3.31 | 0.00 |
| Base | 2013 | 1.21 | 21.20 | 9.60 | 3.56 | 0.00 |
| B.1. | 2014 | 1.01 | 18.10 | 16.60 | 3.19 | 30.00 |
| B.2. | 2014 | 0.91 | 18.00 | 17.91 | 3.25 | 30.00 |
| Base | 2014 | 0.72 | 19.50 | 15.57 | 3.24 | 30.00 |
| B.1. | 2015 | 1.07 | 18.40 | 7.82 | 3.39 | 0.00 |
| B.2. | 2015 | 1.20 | 18.00 | 6.83 | 3.40 | 0.00 |
| Base | 2015 | 0.97 | 18.20 | 6.98 | 3.51 | 0.00 |
| B.1. | 2017 | 1.15 | 18.10 | 6.71 | 3.25 | 0.00 |
| B.2. | 2017 | 1.33 | 17.70 | 5.72 | 3.38 | 0.00 |
| Base | 2017 | 0.92 | 18.20 | 6.74 | 3.36 | 0.00 |
| B.1. | 2018 | 1.23 | 17.70 | 8.26 | 3.43 | 3.00 |
| B.2. | 2018 | 1.90 | 18.20 | 6.96 | 3.39 | 0.00 |
| Base | 2018 | 1.76 | 17.70 | 6.85 | 3.55 | 5.00 |
| B.1. | 2019 | 1.39 | 18.70 | 8.70 | 3.28 | 0.00 |
| B.2. | 2019 | 1.46 | 18.60 | 3.29 | 8.00 | 0.00 |
| Base | 2019 | 1.27 | 18.70 | 7.56 | 3.44 | 5.00 |
| B.1. | 2020 | 1.26 | 17.70 | 8.68 | 3.14 | 0.00 |
| B.2. | 2020 | 1.31 | 18.20 | 7.60 | 3.43 | 0.00 |
| Base | 2020 | 1.03 | 18.80 | 8.57 | 3.55 | 0.00 |
| B.1. | 2021 | 1.28 | 18.90 | 8.62 | 3.29 | 0.00 |
| B.2. | 2021 | 1.24 | 20.20 | 7.40 | 3.25 | 0.00 |
| Base | 2021 | 1.04 | 20.20 | 9.38 | 3.37 | 0.00 |
| B.1. | 2022 | 2.79 | 16.50 | 7.40 | 3.19 | 0.00 |
| B.2. | 2022 | 2.88 | 16.30 | 6.10 | 3.41 | 0.00 |
| Base | 2022 | 2.03 | 17.00 | 6.20 | 3.58 | 0.00 |
Harvest data show that the ’Kéknyelű’ has yielded an average of 1.32 kg/m2 over eleven years, which allows the variety to be grown economically.
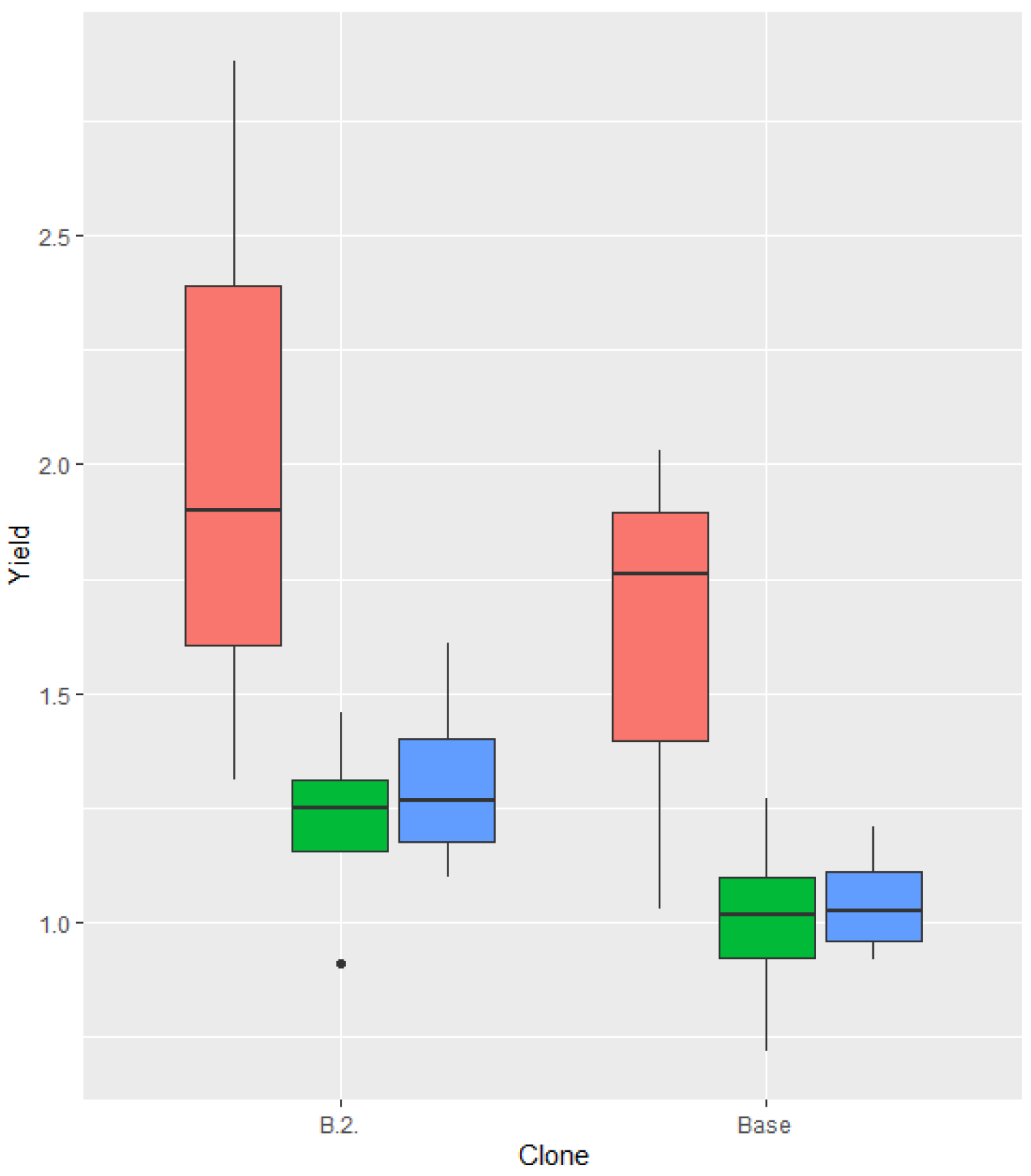
There was significant difference in yield between the clone B.2. and the base variety. In this comparison the year clusters also gave different values, as in years belonging to cluster 1 the ’Kéknyelű’ yielded more than in another years (Figure 3). These differences were were significalt at 90% confidence level.
Figure 3.
Yield results of the ‘Kéknyelű’ clone B.2. and base variety (Badacsony, 2011-2022; data in kg/m2).
Figure 3.
Yield results of the ‘Kéknyelű’ clone B.2. and base variety (Badacsony, 2011-2022; data in kg/m2).
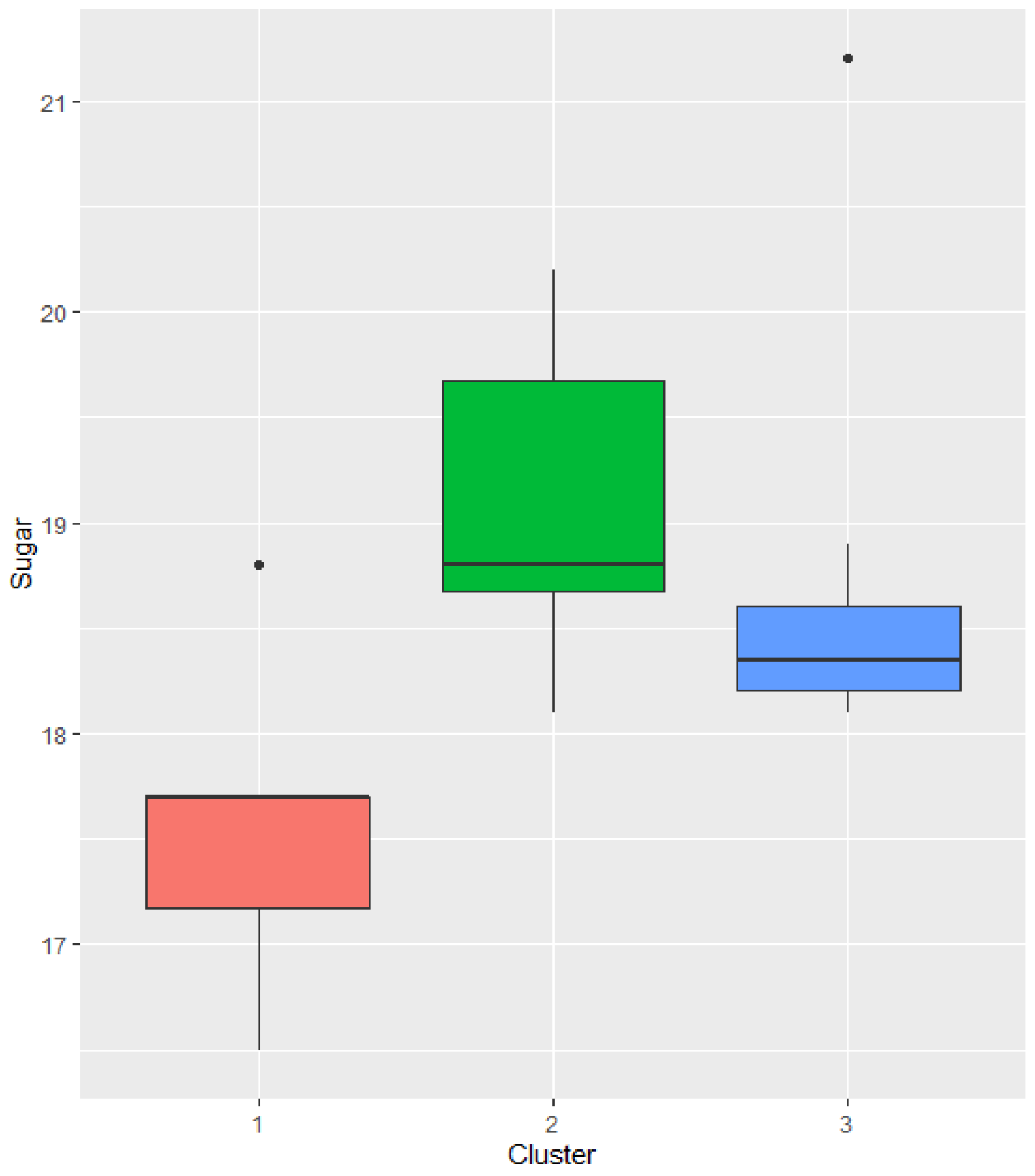
The average sugar content of the ‘Kéknyelű’ grape juice in the examined years was 18.36 KMW. Examining the Klosternuburger Mostwaage results, we found no significant differences between the single clones and the base variety, but the ‘Kéknyelű’ clone B.1 had higher KMW for years in cluster 2 than in cluster 1 in 99% confidence level.
Figure 4.
Sugar content of the juice of the ‘Kéknyelű’ clone B.1. and base variety in different year clusters (Badacsony, 2011-2022; data in Klosternuburger Mostwaage-KMW).
Figure 4.
Sugar content of the juice of the ‘Kéknyelű’ clone B.1. and base variety in different year clusters (Badacsony, 2011-2022; data in Klosternuburger Mostwaage-KMW).
The average titratable acid content of the grape juice was 8.24 g/l over the examined years, no significant differences was detected between clones or year clusters.
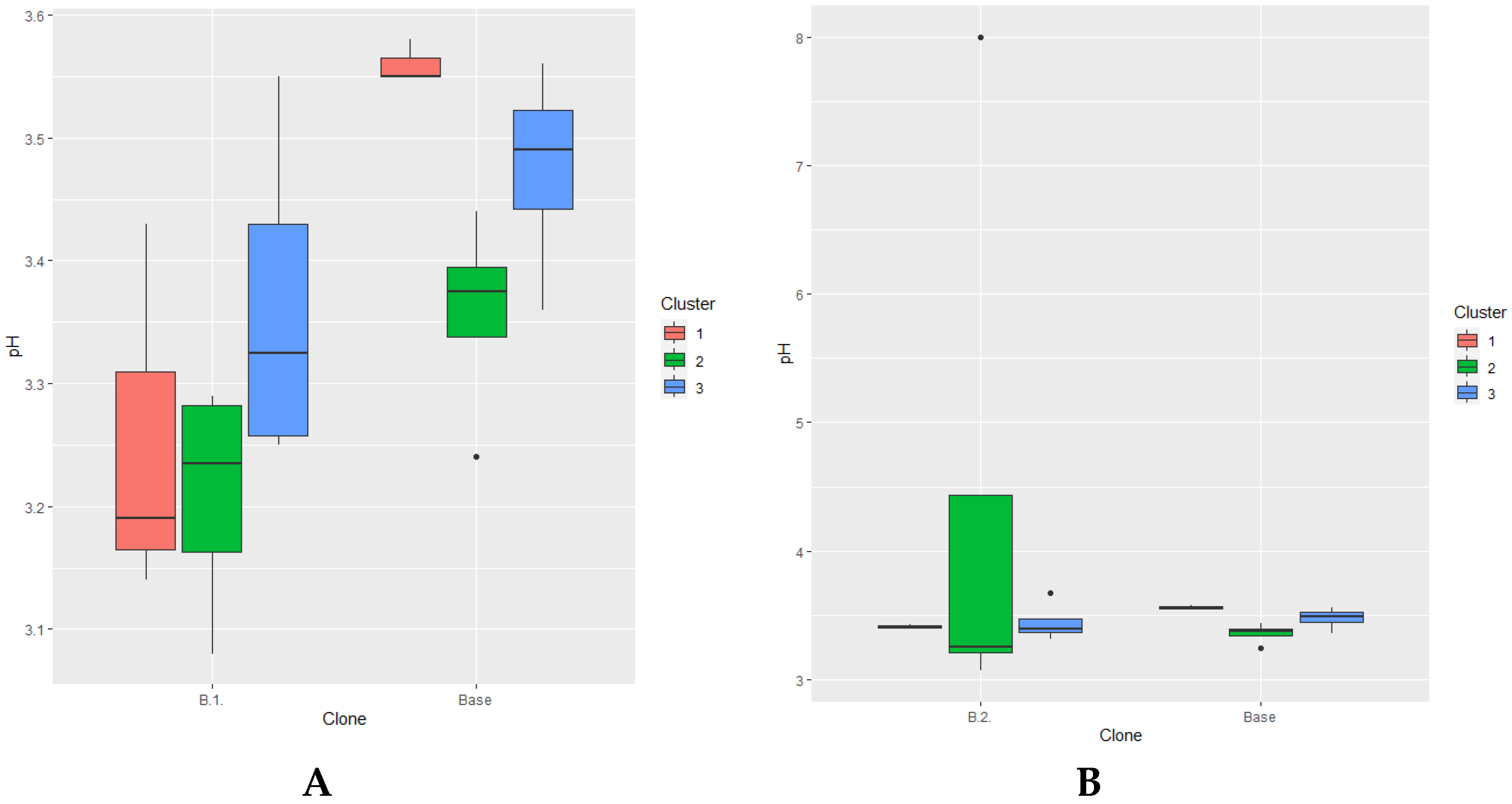
The pH of the grape juice was 3.5. Both clones of ‘Kéknyelű’ matured with significantly lower pH at the 99% and 95% level (Figure 5 A and B). Only clone B.1. showed a significant difference between year types (95% level) indicating the vintage sensitivity of the clone on the basis of pH.
Figure 5.
pH values in different year clusters compared to the ‘Kéknyelű’ base cultivar by B.1.(A) and B.2.(B) clones respectively.
Figure 5.
pH values in different year clusters compared to the ‘Kéknyelű’ base cultivar by B.1.(A) and B.2.(B) clones respectively.
The average percentages of rotting of ‘Kéknyelű’ is 4.03, which is very low. There were no significant differences between clones and the base variety, but there were significant differences (at 95% level) in Botrytis infection among the different clusters of years.
The difference between the individual years was significant at the 90% level or higher for all harvest parameters.
3.3. Evaluation of the results of the harvest of the variety ‘Juhfark’
The harvest performance of the ‘Juhfark’ variety was evaluated based on eleven years of data between 2011 and 2022. In 2016 the plantation suffered bird damage to such an extent that we were unable to harvest a qualifiable crop (in 2019 only the base and in 2022 only the B.1. clone and the base vineyard could be harvested respectively). Harvest results are summarized in Table 5.
Table 5.
Harvest results of the ‘Juhfark’ variety (Badacsony, 2011-2021).
| Clone | Yield kg/m2 |
Sugar content of the juice KMW |
Titratable acid contant of the juice g/l |
pH | Botrytis infection % |
Yield kg/m2 |
|---|---|---|---|---|---|---|
| B.1. | 2011 | 0.28 | 20.80 | 7.47 | 3.13 | 50.00 |
| B.2. | 2011 | 0.64 | 20.20 | 6.86 | 2.96 | 35.00 |
| Base | 2011 | 0.87 | 20.70 | 8.55 | 2.96 | 30.00 |
| B.1. | 2012 | 1.38 | 17.40 | 8.31 | 3.59 | 0.00 |
| B.2. | 2012 | 1.34 | 17.40 | 7.21 | 3.42 | 0.00 |
| Base | 2012 | 1.06 | 20.10 | 7.16 | 3.55 | 0.00 |
| B.1. | 2013 | 1.35 | 20.70 | 12.07 | 3.37 | 3.00 |
| B.2. | 2013 | 1.56 | 19.90 | 10.82 | 3.33 | 2.00 |
| Base | 2013 | 1.08 | 19.80 | 11.50 | 3.21 | 5.00 |
| B.1. | 2014 | 0.73 | 17.40 | 19.84 | 3.28 | 60.00 |
| B.2. | 2014 | 0.21 | 16.50 | 17.52 | 3.18 | 80.00 |
| Base | 2014 | 0.21 | 15.10 | 16.41 | 3.20 | 85.00 |
| B.1. | 2015 | 1.01 | 19.40 | 8.62 | 3.52 | 3.00 |
| B.2. | 2015 | 1.21 | 18.20 | 8.04 | 3.47 | 5.00 |
| Base | 2015 | 1.46 | 17.80 | 9.91 | 3.50 | 10.00 |
| B.1. | 2017 | 1.31 | 17.70 | 7.71 | 3.36 | 5.00 |
| B.2. | 2017 | 1.28 | 18.40 | 7.45 | 3.33 | 5.00 |
| Base | 2017 | 1.57 | 17.50 | 9.44 | 3.28 | 5.00 |
| B.1. | 2018 | 2.26 | 18.00 | 7.29 | 3.46 | 7.00 |
| B.2. | 2018 | 1.99 | 17.40 | 8.48 | 3.47 | 5.00 |
| Base | 2018 | 2.02 | 17.90 | 9.94 | 3.35 | 10.00 |
| B.1. | 2019 | nd. | nd. | nd. | nd. | nd. |
| B.2. | 2019 | nd. | nd. | nd. | nd. | nd. |
| Base | 2019 | 1.43 | 20.80 | 12.55 | 3.42 | 40.00 |
| B.1. | 2020 | 1.17 | 20.40 | 11.88 | 3.41 | 20.00 |
| B.2. | 2020 | 1.06 | 19.90 | 11.08 | 3.34 | 25.00 |
| Base | 2020 | 1.55 | 17.20 | 12.40 | 3.29 | 20.00 |
| B.1. | 2021 | 1.12 | 19.60 | 14.20 | 3.14 | 20.00 |
| B.2. | 2021 | 1.09 | 19.00 | 12.40 | 3.18 | 20.00 |
| Base | 2021 | 1.48 | 15.10 | 18.80 | 3.01 | 20.00 |
| B.1. | 2022 | 0.96 | 17.70 | 7.47 | 3.37 | 10.00 |
| B.2. | 2022 | nd. | nd. | nd. | nd. | nd. |
| Base | 2022 | 2.25 | 17.10 | 10.54 | 3.14 | 10.00 |
nd. = no data (Crop haven’t been harvested because of the hard bird damage.
The ‘Juhfark’ was harvested with a mean yield of 1.23 kg/m2 over ten years the differences were not statistically significant.
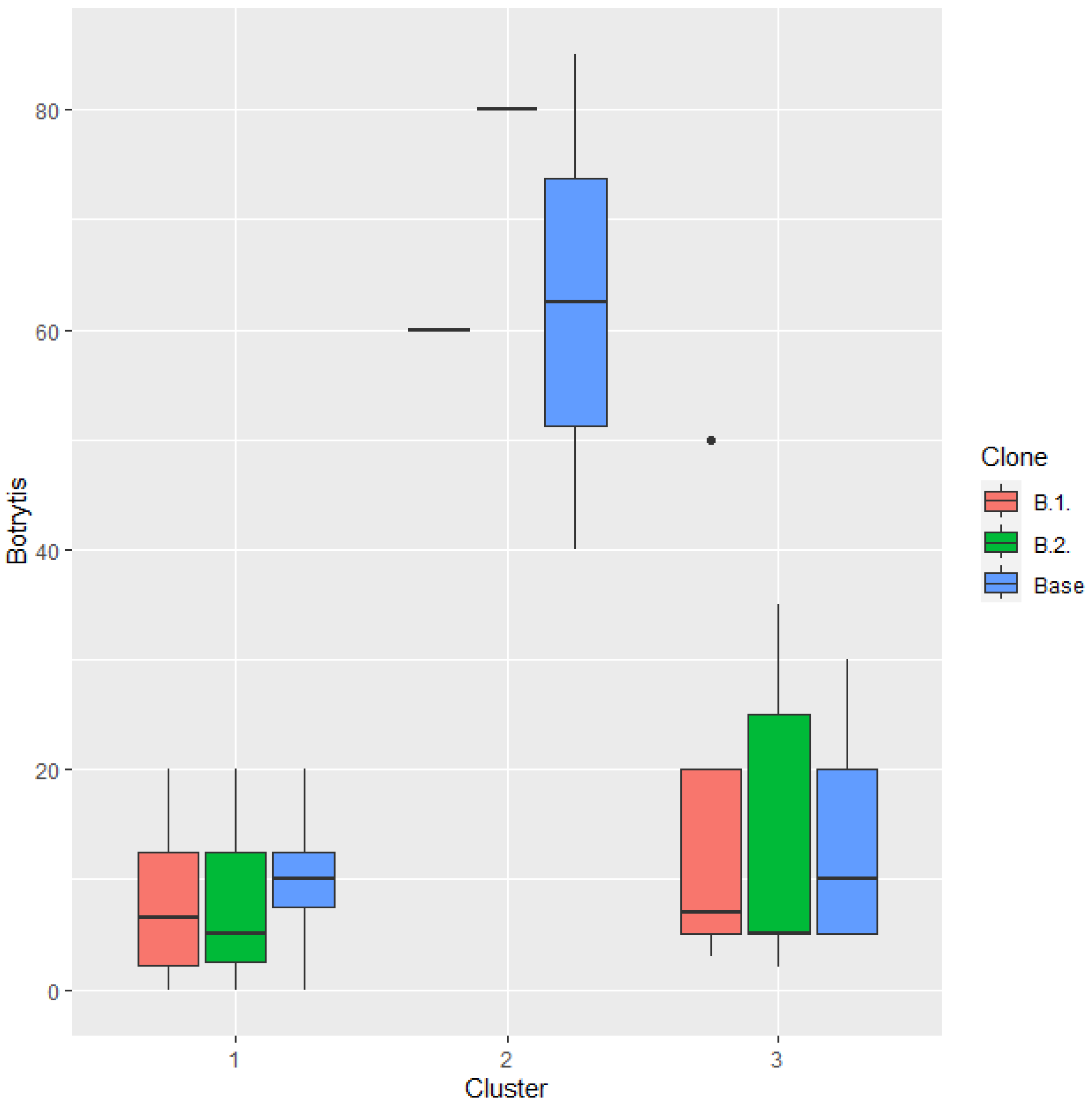
The average sugar content of the ‘Juhfark’ grape juice in the examined years was 18.5 KMW. Titratable acid content of the grape juice was 10.73 g/l, which is quite high, but this is characteristic for this variety. The average pH value of the must was 3.31, and the rate of Botrytis infection was 19.67%, which is high, also characteristic for this variety. Significant differences were only detected between year clusters in Botrytis infection. It is interesting, that in most of the years, the rate of rotting was lower in both of the clones, in specially year cluster, when the overall rotting was quite low (year cluster 1 and 3). The average rate of infection was lower in clone B.1. compared to the base variety in every year clusters (Figure 6), however this difference was not statistically significant.
Figure 6.
Rate of Botrytis infection (%) in different year clusters compared to the ‘Juhfark’ base cultivar by B.1. and B.2. clones.
Figure 6.
Rate of Botrytis infection (%) in different year clusters compared to the ‘Juhfark’ base cultivar by B.1. and B.2. clones.
4. Discussion
In our work, the meteorological data measured over the years were evaluated as the value of different biometeorological indices (GDD, Huglin index, etc.) and average or sum of meteorological parameters calculated at key phenophases of the two grapevine cultivars under study.
As both varieties are considered local, their cultivation is limited to a small geographical area in the Balaton highlands. Since autochthonous varieties are characterized by their year-round sensitivity, the selected clones have to be evaluated over many years. Our results show that, for most harvest parameters, differences between clones are not even apparent after 11 years of testing.
In the case of ‘Kéknyelű’ our results showed that both of the clones yielded higher than the base variety, the difference was significant only for clone B.2. This is important because low yields due to poor fertility are the main problem in ‘Kéknyelű’ [8,12,22]. Among the quality parameters, in the context of global warming, the significantly lower pH of both clones should be highlighted. Strong relationships between total acidity, titratable acidity and pH was shown in several times [42,43], the lower pH is associated with higher acidity. ‘Kéknyelű’ is famous about the special variety character of its wine, which is told to have pronounced acidity. Given the acidic character of ‘Kéknyelű’ wine and the increasing must degrees and increasing pH expected due to climate change these differences could be positive in the long term [44,45,46,47,48].
No statistically verifiable difference in harvest parameters was observed for ‘Juhfark’, suggesting that further studies will be needed due to climate change and the high vintage sensitivity of the variety [49,50,51].
An interesting result is that the two varieties responded differently to different weather conditions in different seasons. In the case of ‘Kéknyelű’, our results showed significant differences in yield in clone B.2. between vintage clusters. However, clone B.1. showed differences in KMW values and pH. Both of the ‘Kéknyelű clones have given significantly higher rate of Botrytis infection in the years belonging to cluster 2, than in cluster 3.
In the ‘Kéknyelű’ clone B.1., in years belonging to the cluster1 the pH was significantly lower, and the rate of rotting was significantly higher than in years belonging tocluster 3. The pH value depends to a large extent on the acid composition of the grape juice, with a lower pH usually associated with a higher malic acid content. In this case, the relationship between the lower pH and higher rotting is essentially could be explained by the fact that the Botrytis fungus uses sugars as a food source originates from the conversion of organic acids, mainly malic acid. It was reported, that during grape berry ripening, the conversion of acids into sugars indulgence the development of Botrytis cinerea [52,53]. Sugars, malic acid, potassium, and sodium content of berry exudates promote mycelial growth of pathogen [54].
In the case of the ‘Juhfark’, the vintage sensitivity is related to the problem of the variety, the sensitivity to bunch rot. Both of the clones showed significantly different sensitivity to Botrytis infection in different year clusters, in years of cluster 2 gave higher values (worst results) than the years in another year clusters.
5. Conclusions
In addition to improving the breeding problem in clonal selection breeding of indigenous varieties, it is very important to maintain the varietal character of the cultivar, as this is what makes these varieties unique [51]. Both the ‘Kéknyelű’ and the ‘Juhfark’ varieties have an acidic character, retaining their acidity even in drought conditions, and this is reflected in the varietal character of their wines [8,32]. Importantly, we have managed to maintain relatively high titratable acidity values for both varieties in the clones tested.
Based on eleven years of data, only small differences were found between the tested varieties and their clones, most of which were not statistically verifiable. As breeding of woody plants (such as grapes) takes a very long time, the potential effects of climate change have to be taken into account already during a selection cycle and clones have to be selected accordingly.
6. Patents
The clones of the ‘Kéknyelű’ and ‘Juhfark’ varieties tested in this manuscript have been entered for state registration in Hungary.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table 1: Detailed daily meteorological data.
Author Contributions
Conceptualization, methodology, formal analysis, visualization, project administration writing—original draft preparation, J.G.; validation, formal analysis, data curation: K.Sz.Gy and N.Cs. writing—review and editing: D.T., O.R., O.K.; supervision and funding acquisition: Sz.B. and Ny.S.D.Á.
Funding
This research was funded by the National Research, Development and Innovation Office, grant number NKFIH-831-10/2019
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- van Leeuwen, C.; Darriet, P. The Impact of Climate Change on Viticulture and Wine Quality. Journal of Wine Economics 2016, 11, 150–167. [Google Scholar] [CrossRef]
- van Leeuwen, C.; Destrac-Irvine, A. Modified Grape Composition under Climate Change Conditions Requires Adaptations in the Vineyard. Oeno One 2017, 51, 147–154. [Google Scholar] [CrossRef]
- Ollat, N.; van Leeuwen, C.; de Cortazar-Atauri, I.G.; Touzard, J.M. The Challenging Issue of Climate Change for Sustainable Grape and Wine Production. Oeno One 2017, 51, 59–60. [Google Scholar] [CrossRef]
- Németh, K. Klímaváltozás Hatása a Borszőlő Biológiai Jellemzőire, Termésmennyiségére, Minőségére. ÉRTÉKÁLLÓ ARANYKORONA 2012, 12, 6–7. [Google Scholar]
- Németh, K. Száraz Termőhelyen. Kertesz Szolesz 2015, 20–21. [Google Scholar]
- Tkhakushinov, E.K.; Hachemizova, E.A. Development Problems of the Regional Agro-Industrial Complex Management System; 2018; Volume 7.
- Ostroukhova, E.; Levchenko, S.; Vasylyk, I.; Volynkin, V.; Lutkova, N.; Boyko, V. COMPARISON OF THE PHENOLIC COMPLEX OF CRIMEAN AUTOCHTHONOUS AND CLASSIC WHITE-BERRY GRAPE CULTIVARS. [CrossRef]
- Májer, J.; Győrffyné Jahnke, G. Autochton Szőlőfajták Optimális Termesztéstechnológiáját Megalapozó Kísérletek Eredményei Badacsonyban. Borászati Füzetek 2005, 17, 4–9. [Google Scholar]
- Somogyi, N.; Németh, K. A Kárpát-Medence Kincsei Határon Innen És Túl. MEZŐHÍR: ORSZÁGOS AGRÁRINFORMÁCIÓS SZAKLAP 2018, 22, 44–46. [Google Scholar]
- Goethe, H. Handbuch Der Ampelographie; Verlag Parely: Berlin, 1887. [Google Scholar]
- Németh, M. Ampelográfiai Album, I. (Termesztett Borszőlőfajtáink 1.); Mezőgazdasági Kiadó: Budapest, 1967. [Google Scholar]
- Jahnke, G.; Korbuly, J.; Májer, J.; Györffyné Molnár, J. Discrimination of the Grapevine Cultivars “Picolit” and “Kéknyelu” with Molecular Markers. Sci Hortic 2007, 114. [Google Scholar] [CrossRef]
- Györffyné Jahnke, G.; Májer, J.; Korbuly, J. Distinguishing the Grapevine Cultivars “Picolit” and “Kéknyelü” with Isozymes and Microsatellite Markers. Acta Hortic 2009, 827, 159–162. [Google Scholar] [CrossRef]
- Bakonyi, L.; Bényei, F.; Fazekas, I.; Hajdu, E.; Korbuly, J.; Lőrincz, A.; Marcinkó, F.; Pernesz, Gy.; Romenda, R.; Zanathy, G. Borszőlőfajták, Csemegeszőlő-Fajták És Alanyok; Bényei, F., Lőrincz, A., Eds.; Mezőgazda Kiadó: Budapest, 2005; ISBN 9789632865362. [Google Scholar]
- Varga, Zs.; Bényei, F.; Lőrincz, A.; Ulz, A. Yields and Quality of Former Vitis-Cultivars at Tokaj-Hegyalja. Borászati Füzetek 2006, 16, 1–8. [Google Scholar]
- Varga, Z.; Ferenczy, A.; Bényei, F.; Zanathy, G. EXAMINATION OF THE RELATIONS OF ORIGIN OF OLD GRAPE VINE CULTIVARS WITH CLUSTER ANALYSIS IN TOKAJ-HEGYALJA. Acta Hortic 2009, 169–176. [Google Scholar] [CrossRef]
- Halász, G.; Veres, A.; Kozma, P.; Kiss, E.; Balogh, A.; Galli, Z.; Szőke, A.; Hoffmann, S.; Heszky, L. Microsatellite Fingerprinting of Grapevine (Vitis Vinifera L.) Varieties of the Carpathian Basin; 2005; Volume 44.
- Lencsés, A.K.; Szőke, A.; Kozma, P.; Halász, G.; Katuláné Debreceni, D.; Veres, A.; Györffyné Jahnke, G.; Kiss, E. Mikroszatellit Markerek Alkalmazása a Magyarországi Szőlő Génforrások Megőrzésére. Kertgazdasag - Horticulture 2010, 42, 58–67. [Google Scholar]
- Jahnke, G.; Májer, J.; Lakatos, A.; Molnár, J.G.; Deák, E.; Stefanovits-Bányai, E.; Varga, P. Isoenzyme and Microsatellite Analysis of Vitis Vinifera L. Varieties from the Hungarian Grape Germplasm. Sci Hortic 2009, 120. [Google Scholar] [CrossRef]
- Jahnke, G.; Májer, J.; Varga, Z.; Deák, E.; Varga, P. An Acid Phosphatase Isoenzyme Pattern Is Characteristic for the Pontican Cultivars. Cereal Res Commun 2009, 37. [Google Scholar] [CrossRef]
- Bodor, P.; Varga, Zs.; Deák, T.; Pedryc, A.; Bisztray, Gy.D. Old Hungarian Grapevine Cultivars and Their Relations Characterized with Microsatellite Markers. Int J Hortic Sci 2008, 14. [Google Scholar] [CrossRef]
- Györffyné Jahnke, G.; Májer, J. Results of the Experiments for the Improvement of the Fertilisation of the Functional Female Flowered Grapevine Cultivar “Kéknyelű”. Acta Hortic 2003, 603, 767–773. [Google Scholar] [CrossRef]
- Kozma, P. A Szőlő Nemesítése. (Breeding of the Vine); Mezőgazdasági Kiadó: Budapest, 1951. [Google Scholar]
- Hajdu, E. Grapevine Breeding in Hungary. Grapevine Breeding Programs for the Wine Industry 2015, 103–134. [Google Scholar] [CrossRef]
- Németh, M. A Szőlő Klónszelekciós Nemesítéséről. (About the Clonal Selection of the Vine). Agrártudomány 1958, 43–49. [Google Scholar]
- Lunz, O. A Klónszelekcióü Hazai Helyzete És Eredményei (National Situataion and Results of Clonal Selection). Szőlőtermesztés és Borászat 1990, 12, 2–7. [Google Scholar]
- Hajdu, E.; Korać, N.; Cindrić, P.; Medić, M. Genetical Variations in Vine (Mutation) The Importance of Clonal Selection of Grapevine and the Role of Selected Clones in Production of Healthy Propagating Stocks. Int J Hortic Sci 2011, 17, 15–24. [Google Scholar] [CrossRef]
- Hajdu, E. A Kertészeti Növények Nemesítése a Szőlő Példáján. (The Horticultural Plant Breeding on the Example of Grape. Gradus 2016, 3, 378–383. [Google Scholar]
- Szűgyi-Reiczigel, Z.; Ladányi, M.; Bisztray, G.D.; Varga, Z.; Bodor-Pesti, P. Morphological Traits Evaluated with Random Forest Method Explains Natural Classification of Grapevine (Vitis Vinifera L.) Cultivars. Plants 2022, 11, 3428. [Google Scholar] [CrossRef] [PubMed]
- Cousins, P.; Garris, A. Quality Improvement in “Vignoles” through Clonal Selection. Acta Hortic 2014, 1046, 287–290. [Google Scholar] [CrossRef]
- Stover, E.; Aradhya, M.; Dangl, J.; Prins, B.; Cousins, P. Grape Genetic Resources and Research at the Davis California National Clonal Germplasm Repository. Acta Hortic 2009, 827, 193–196. [Google Scholar] [CrossRef]
- Győrffyné Jahnke, G.; Knolmájerné Szigeti, G.; Németh, C.; Nagy, Z.A.; Májer, J. Kéknyelű És Juhfark Fajták Klónszelekciója Badacsonyban. Agrofórum Extra 2018, 46, 26–28. [Google Scholar]
- Jahnke, G.; Májer, J.; Varga, P.; SzÖke, B. Analysis of Clones of Pinots Grown in Hungary by SSR Markers. Sci Hortic 2011, 129, 32–37. [Google Scholar] [CrossRef]
- Lemos, A.M.; Machado, N.; Egea-Cortines, M.; Barros, A.I. ATR-MIR Spectroscopy as a Tool to Assist ‘Tempranillo’ Clonal Selection Process: Geographical Origin and Year of Harvest Discrimination and Oenological Parameters Prediction. Food Chem 2020, 325, 126938. [Google Scholar] [CrossRef]
- Pejić, I.; Šimon, S.; Preiner, D.; Žulj Mihaljević, M.; Maletić, E.; Zdunić, G.; Petric, I. v.; Anhalt, U.; Forneck, A.; Ruehl, E. Estimate of Intravarietal Genetic Variation as a Prerequisite for Successful Clonal Selection in Grapevine. Acta Hortic 2015, 1082, 105–112. [Google Scholar] [CrossRef]
- Doulati Baneh, H. Molecular Markers to Detect Genetic Variation in Superior Clones of Grapevine (Vitis Vinifera L. ’Keshmeshi’). Acta Hortic 2015, 1082, 183–188. [Google Scholar] [CrossRef]
- Vezzulli, S.; Leonardelli, L.; Malossini, U.; Stefanini, M.; Velasco, R.; Moser, C. Assessing the Genetic Variability of Grape Clones. Acta Hortic 2014, 1046, 357–362. [Google Scholar] [CrossRef]
- Anhalt, U.C.M.; Crespo Martínez, S.; Rühl, E.; Forneck, A. An AFLP-Marker Study of the Vitis Vinifera L. Cultivar “White Riesling” Comprising 86 Clones to Investigate the Stability of Clones. Acta Hortic 2014, 1046, 681–684. [Google Scholar] [CrossRef]
- Wobbrock, J.O.; Findlater, L.; Gergle, D.; Higgins, J.J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only ANOVA Procedures. Conference on Human Factors in Computing Systems - Proceedings 2011, 143–146. [Google Scholar] [CrossRef]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. UIST 2021 - Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology 2021, 754–768. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.r-project.org/.
- Bonada, M.; Sadras, V.O. Review: Critical Appraisal of Methods to Investigate the Effect of Temperature on Grapevine Berry Composition. Aust J Grape Wine Res 2015, 21, 1–17. [Google Scholar] [CrossRef]
- Boulton, R. The Relationships between Total Acidity, Titratable Acidity and PH in Grape Tissue. VITIS - Journal of Grapevine Research 1980, 19, 113–113. [Google Scholar] [CrossRef]
- Neethling, E.; Petitjean, T.; Quénol, H.; Barbeau, G. Assessing Local Climate Vulnerability and Winegrowers’ Adaptive Processes in the Context of Climate Change. Mitig Adapt Strateg Glob Chang 2017, 22, 777–803. [Google Scholar] [CrossRef]
- Neethling, E.; Barbeau, G.; Coulon-Leroy, C.; Quénol, H. Spatial Complexity and Temporal Dynamics in Viticulture: A Review of Climate-Driven Scales. Agric For Meteorol 2019, 276–277. [Google Scholar] [CrossRef]
- Szenteleki, K.; Horváth, L.; Ladányi, M. Climate Risk and Climate Analogies in Hungarian Viticulture. International Conference on Future Environment and Energy 2012, 28, 250–254. [Google Scholar]
- Szenteleki, K.; Ladányi, M.; Gaál, M.; Zanathy, G.; Bisztray, G.Y. Climatic Risk Factors of Central Hungarian Grape Growing Regions. Appl Ecol Environ Res 2011, 10, 87–105. [Google Scholar] [CrossRef]
- Navrátilová, M.; Beranová, M.; Severová, L.; Šrédl, K.; Svoboda, R.; Abrhám, J. The Impact of Climate Change on the Sugar Content of Grapes and the Sustainability of Their Production in the Czech Republic. Sustainability (Switzerland) 2021, 13, 1–18. [Google Scholar] [CrossRef]
- Carámbula, C.; Moreno, M.T.; Riera, D.; Cretazzo, E.; Tomás, M.; A 1; Medrano, H. Selección Clonal de Las Principales Variedades Autóctonas de Baleares.
- Roby, J.P.; Leeuwen, C. van About the Need of Maintaining Simultaneously Mass and Clonal Selection for Conservation of Genetic Diversity in Both International and Autochtoneous Varieties. In Proceedings of the 2nd International Symposium. Exploitation of autochthonous and more common vine varieties. Genetic pedigree and phenotyping, tolerance and stress, diseases to control, rootstocks. OENOVITI INTERNATIONAL network; Geisenheim, Germany, November 3 2014; pp. 93–97. [Google Scholar]
- Roby, J.-P.; Leeuwen, C. van; Gonçalves, E.; Graça, A.; Martins, A. The Preservation of Genetic Resources of the Vine Requires Cohabitation between Institutional Clonal Selection, Mass Selection and Private Clonal Selection. BIO Web Conf 2014, 3, 01018. [Google Scholar] [CrossRef]
- Prusky, D. PATHOGEN QUIESCENCE IN POSTHARVEST DISEASES. Annual review of Phytopathology 2003, 34, 413–434. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and Necrotrophic Lifestyle Choice During Postharvest Disease Development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Padgett, M.; Morrison, J.C. Changes in Grape Berry Exudates during Fruit Development and Their Effect on Mycelial Growth of Botrytis Cinerea. Journal of the American Society for Horticultural Science 1990, 115, 269–273. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



