Submitted:
29 June 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
Dioscorea composita is a plant native to México and Central America with a high concentration of diosgenin precursors. Currently, México is one of the two most important countries producers of this yam; however, climate change is altering the environmental conditions of its natural habits, threatening its preservation and productivity. This is why this research was focused to characterize the eco-geography of D. composita and predict its potential geographic distribution under climate change scenarios in México-Central America. A collection of 408 geo-referenced accessions was used to determine its climatic adaptation, ecological descriptors, and the current and future potential geographic distribution, which was modeled with MaxEnt model through the Kuenm R-package. For future climate scenarios, an ensemble of the GCMs HadGEM-ES and CCSM4 was used. Results showed that D. composita adapts to warm humid and very humid agro-climates and that the most contributing variables for its presence are annual and seasonal moisture availability indices, seasonal photoperiod, annual thermal range, Bio14 and Bio11. The year 2050 RCP 4.5 climate scenario would contract the potential distribution of D. composita, whilst the 2050 RCP 8.5 scenario would expand it, indicating that this species could be a good crop option under this scenario of emissions.
Keywords:
Climate change effects
; nutraceutical species
; distribution
; Ecological niche model
; Mesoamerica
Abstract.Dioscorea composita is a plant native to México and Central America with a high concentration of diosgenin precursors. Currently, México is one of the two most important countries producers of this yam; however, climate change is altering the environmental conditions of its natural habits, threatening its preservation and productivity. This is why this research was focused to characterize the eco-geography of D. composita and predict its potential geographic distribution under climate change scenarios in México-Central America. A collection of 408 geo-referenced accessions was used to determine its climatic adaptation, ecological descriptors, and the current and future potential geographic distribution, which was modeled with MaxEnt model through the Kuenm R-package. For future climate scenarios, an ensemble of the GCMs HadGEM-ES and CCSM4 was used. Results showed that D. composita adapts to warm humid and very humid agro-climates and that the most contributing variables for its presence are annual and seasonal moisture availability indices, seasonal photoperiod, annual thermal range, Bio14 and Bio11. The year 2050 RCP 4.5 climate scenario would contract the potential distribution of D. composita, whilst the 2050 RCP 8.5 scenario would expand it, indicating that this species could be a good crop option under this scenario of emissions.
1. Introduction
Dioscorea composita Hemsl is a wild species that has its origin in México and Central America [1], although some authors consider Mexico as the most probable center of origin [2,3]. Dioscorea composita has been historically recognized for the production of secondary metabolites of pharmaceutical importance, including diosgenin. However, due to advances in the chemical synthesis of these compounds, the relevance of natural diosgenin in medicine has diminished over time [3]; consequently, the industry's interest in this plant decreased, which caused that diverse aspects of this plant to remain understudied until now. Nevertheless, recently, the interest in this species has resurfaced, since it has been discovered that it may constitute a promising therapeutic agent against cancer [4,5]. In addition, D. composita is a plant with socioeconomic importance that has great nutritional and ethnobotanical value [3]. Currently, it can be considered a nutraceutical plant with use in states of southeastern México [6]. All this situation motivated the present investigation, whose objectives were to characterize the eco-geography and environmental adaptation of D. composita, as well as to predict its current and future potential distribution under climate change scenarios in its original region of occurrence (México-Central America). Climate change is causing variations in temperature and precipitation patterns, as well as in the frequency of extreme weather events [7] causing alterations in the range of species distribution, and modifying the composition and characteristics of ecosystems. México and Central America are some of the most impacted regions by climate change, due to their geographic and orographic characteristics, together with the unequal territorial distribution of natural resources [8]. Climate scenarios modeled for the middle of this century, predict increases in temperature from 1 to 3°C and a decrease in precipitation of around 10%, causing diverse environmental combinations that provoke particular climatic conditions, which alter adversely the agroclimatic conditions of these regions [9], which include the central and northern part of México [10,11]; as well as the tropical and subtropical zones [12]. Central America has experienced the ravages of this phenomenon, causing drought mainly in El Salvador, Guatemala, and Honduras in 2014 and 2015, endangering temperate and cloudy forests [8]. These changes in weather patterns impact the current distribution areas of numerous wild species in México and Central America. In the Mexican occurrence sites of D. composita, it is estimated that by the year 2050, the annual mean temperature will increase 2-3 °C and annual precipitation will decrease 10-50 mm, in relation to the average climatology 1961-2010 [13]; while, for the Central American occurrence sites, an increase of 2.5-3°C, and a decrease of 25-70 mm of precipitation are expected by the year 2050 [14]. However, the possible effects of these climatic changes on the presence and potential distribution of D. composita have not yet been assessed.
2. Materials and Methods
2.1. Occurrence data
After data curation to mainly discard duplicate and erroneous information [15] a database of D. composita occurrence sites in México-Central America was conformed considering a final list of 408 geo-referenced accessions (Table 1). University herbaria, digital herbariums, floristic inventories, and scientific articles were considered as data sources. We only considered the species occurrence sites in México-Central America because they are considered the natural populations of D. composita (this region is its center of origin), therefore, they are adequate to characterize the eco-geography of this species and the final purposes of this research [16].
2.2. Climatic data
Raster images of temperature, precipitation, evapotranspiration (ETP), and photoperiod were used, from the Agroclimatic Information System for México-Central America (SIAMEXCA) [13], which has a resolution of 30'' arc and correspond to climatic normals for the period 1961-2010. Additional parameters were derived from these images using the IDRISI Selva software [17], adding a total of 31 variables (Table 2). For climate change scenarios, we used raster images year 2050 RCPs 4.5 and 8.5 derived from an ensemble of two GCMs corresponding to the Coupled Model Inter-comparison Project Phase 5 (CMIP5): HadGEM-ES (European Network for Earth System-Met Office Hadley Center) and CCSM4 (Community Climate System Model), since they have been shown to adequately address and describe continental vegetation, including various types of vegetation [18], in addition to showing good adjustment to the climatic conditions of México and Central America [14,19,20]. Those models have been used successfully formerly to predict suitability of crops under climate change scenarios [21].
2.3. Environmental characterization of the occurrence sites
Environmental conditions of the D. composita occurrence sites were characterized by extracting values from raster images corresponding to 31 variables. This extraction was made with the system ArcMap 10.8 [22] and by using geographical coordinates for each site. With the extracted data, a data matrix was built in Microsoft Excel.
2.4. Selection of environmental variables
To obtain accurate biological information, species distribution models must be built with predictor variables that have a direct influence on the presence of the species [23]. Thus, it is important to make a good selection of variables, avoiding among other aspects, the multi-collinearity of variables, which can cause a misinterpretation of the model, due to the high level of correlation between variables [24]. Besides, severe multi-collinearity can increase the variance of regression coefficients, making them unstable [25]. Before any statistical analysis, we applied the Shapiro-Wilk test for data normality and found no normality for all variables (p<0.05). Thus, we used Spearman's r for correlation analysis. Data from the environmental data matrix were used to perform the correlation analyses; this statistical analysis was carried out with programs developed in the R software version 4.05 [26]. To determine the presence of multi-collinearity, a threshold value of correlation coefficient >0.8 [27] was established. This way, the correlated variables with a coefficient <0.8 were selected, and among the variables with collinearity, the one considered most relevant for the species presence was selected. Furthermore, with the selected variables, preliminary Maxent modeling was performed to identify the environmental variables with the greatest contribution to the distribution of D. composita. This produced a final list of selected variables, with which MaxEnt was run again to obtain an optimal model of D. composita niches distribution.
2.5. Eco-geography of D. composita: Determination of climatic adaptation and ecological descriptors
The climatic adaptation of D. composita was determined by identifying the occurrence of the species in different agro-climatic regions, for this, the map of agro-climatic regions for México and Central America was used [16]. In this way, a list of the agro-climatic conditions in which this species is present was derived.
For the determination of the ecological descriptors, the extracted data matrix was used, considering only information from 20 environmental variables, which were selected after the correlation analysis to discriminate variables highly correlated.
2.6. Ecological niche modeling
The MaxEnt algorithm was used for ecological niche modeling (ENM) of D. composita in the current climate scenarios as well as in the climate change scenarios addressed in this research. MaxEnt model was implemented with the assistance of the Kuenm R-package [26,28], which automates and optimizes the ENM process. The preliminary models were created using Kuenm's Kuenm_ceval function, which utilizes occurrence data and environmental predictors, and their efficiency was evaluated through the cal_eval function that determines their statistical significance. To ensure accuracy, ten replicates were performed via cross-validation, using logistic outputs for current and future climatic scenarios. Final model assessments included partial ROC calculations and omission rates (based on E = 5%) using an independent dataset. The functions kuenm_mod and kuenm_feval were used to finalize the models and perform evaluations, respectively [28]. The quality of the model was assessed with the AUC value from the ROC curve, and the Akaike Information Criterion (AIC), corrected for small sample sizes (AICc) [28]. To identify any potential model transfer extrapolation risks, mobility-oriented parity (MOP) analyses were conducted using the kuenm_mmop function [29].
For this research, models were tested using a sequential order of the FC (L, LQ, H, LQH, LQHP, LQHPT) and regularization multiplier (RM) values of 0.1 to 10 with 0.1 increases, a maximum omission rate of 5%, and run 25 k-fold replicates of each configuration; 500 iterations were used [30]. Two models were generated for each parameter setting, one based on the complete set of occurrences and the other based only on the training data.
3. Results
3.1. Statistical analysis and environmental variables selection
Based on Spearman's correlation analysis, 20 environmental variables were selected and used to characterize the eco-geography of D. composita. However, for the purpose of the species distribution modeling, the preliminary ENM with MaxEnt revealed that seven variables are the ones that contribute the most to the eco-geographical distribution of D. composita, which were used for the final ENM processes with Kuenm R-package [31].
3.2. Eco-geography of D. composita
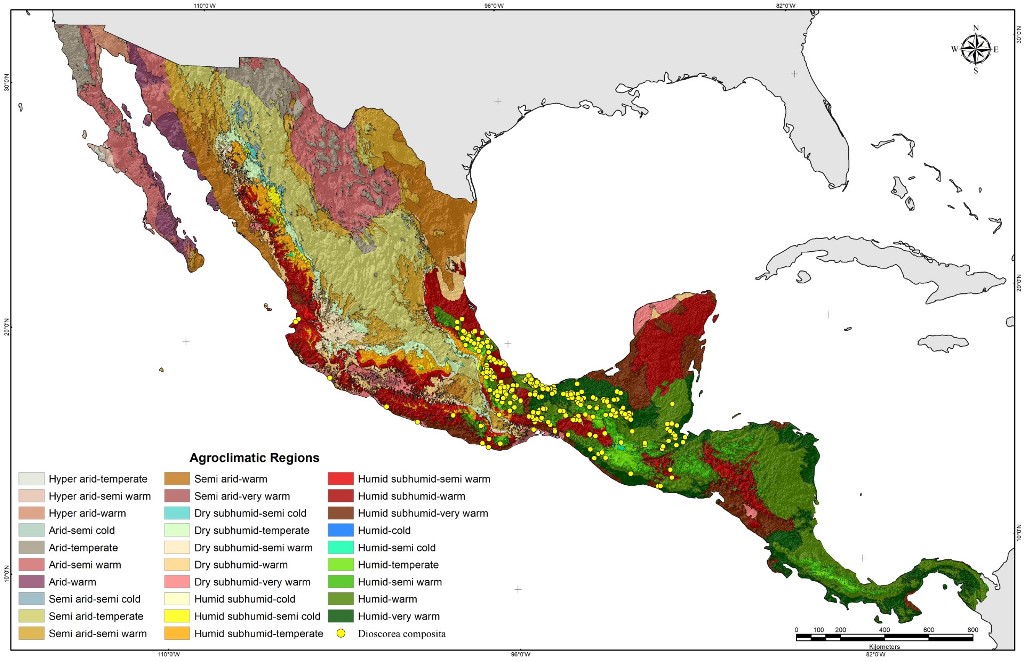
Figure 1 shows the current geographical distribution of D. composita in the agro-climatic regions of México and Central America. The occurrence sites for this species are distributed mainly from the north of Veracruz in México to some areas of Guatemala, Belize, and El Salvador. In this area, most of the D. composita accessions are located in the agro-climates humid warm (209 accessions, 51.2%), and very humid warm (83, 20.3%), remarking the preference of this species for humid warm climates.
The map of Figure 1 also shows presence sites in other regions of México with different environmental conditions, proving the ability of D. composita to colonize new habitats and adapt to other environmental conditions. Thus, D. composita is currently distributed in 10 agro-climates that go from temperate to very warm and from semi-arid to very humid environments (Table 3).
Climatic characterization of each accession site is described in the Supplemental File. Based on such data, in Table 4, the ecological descriptors of D. composita are shown, which include the minimum, maximum and optimal range for each parameter. We decided not to include the average value but the optimal intervals, which correspond with the highest frequency of the species occurrence sites. Since this climatic characterization derives from its center of origin, the most original habitats of D. composita were taken into account to do it, hence, this climatic characterization should be considered as a valid reference of its eco-geography. These results may be worthy to perform the species agro-ecological zonation and other applications, now that Latin American governments are more involved in the inventory of native plant genetic resources [32].
Four important climatic aspects for D. composita presence may be distinguished in Table 4 data: moisture availability, temperature conditions, thermal range, and photoperiod, most of them important as annually as well as seasonal parameters. The species is present in Mesoamerica region, from 5 to 3001 m above sea level, although its optimal elevation range is 5-1125 m.
It is interesting to notice that some of the ecological descriptors included in Table 4, are not commonly reported, such as the moisture available index (MAI), in this table expressed by the annual and seasonal moisture indices. We obtained that D. composita distributes in the range of 0.42 to 3.75, with an optimal interval of 1.1 to 2.9, which is in correspondence with the optimal interval of annual precipitation of 1940 to 3839 mm (Table 4). The thermal oscillation also influences the distribution and development of tubers, the optimal range determined for D. composita is from 10.1 to 12.9 °C, whereas photoperiod was selected as an important seasonal variable in the period spring-summer (May-October) as well as in autumn-winter (November-April), with optimal ranges of 12.9-13.15 h and 11.27-11.5 h, respectively. The optimal range for the annual mean temperature (23-28°C) proves that D. composita is a species whose natural habitat is the warm thermal zones (Table 4).
3.3. Modeling the potential distribution of Dioscorea composita
Kuenm R-package enabled to obtain 341 MaxEnt models (Figure 2), all of them significant, however, only one model positively met the Akaike criterion (AIC=0) and a maximum omission rate of 5%. The Kuenm R-package implemented with MaxEnt determined that 2.0 was the optimum regularization multiplier. The model finally selected to depict the distribution of D. composita was judged excellent since the AUC of the ROC curve accounted for 0.946.
The Jackknife test revealed that the variables with the greatest contribution to the distribution niche model of D. composita are AMAI (38.4%), NAPH (20.7%), and ATO (17.1%) (Table 5).
Furthermore, results from the Jackknife analysis also indicated that when testing the three cases "with only variable", "without variable", and "with all variables", the variable with the highest gain was ATO (annual mean thermal oscillation), with a regularized training gain > 1.1 (Figure 3). Other important environmental variables were NAPH (November-April mean photoperiod), AMAI (annual mean moisture availability index), MOMAI (May-October mean moisture availability index), NAMAI (November-April mean moisture availability index), BIO07 (Temperature Annual Range) BIO14 (precipitation of the driest month), and BIO11 (mean temperature of the coldest quarter), all of them with regularized training gains > 1.0 (Figure 3).
Areas with current environmental aptitude for D. composita were generated as a binomial map (Figure 4). We used the method of Balance training omission, predicted area, and threshold value (BTOPATV) to delimit suitable areas for D. composita. Thus, Figure 3 shows the current and potential distribution of D. composita. The total potential distribution area for D. composita accounted 692,123 km2, and is located in Durango, Nayarit, Jalisco, Colima, Michoacán, Guanajuato, San Luis Potosí, Tamaulipas, Quintana Roo, Yucatán, Campeche, Tabasco, Veracruz, Guerrero, Oaxaca, and Chiapas, in México, and in some areas of Guatemala, Belize, El Salvador, Honduras, and Nicaragua.
3.4. Impact of climate change on the potential distribution of D. composita
To generate potential distribution maps under climate change scenarios (2050 RCP 4.5 and RCP 8.5), we also generated binary maps by using the Balance Training Omission, Predicted Area, and Threshold Value (BTOPATV) method. Since an appropriate selection of a thresholding method enables to avoid an important bias in the final distribution map [33], several aspects must be taken into account, such as minimizing the omission rate of the map as possible but without neglecting to seek a realistic map [16], this is why we decided to use BTOPATV.
Figure 5 shows the environmental suitability areas for D. composita in the year 2050 under two different climate paths (RCP 4.5 and RCP 8.5). Both contraction and expansion areas were produced for the distribution of D. composita.
Table 6 indicates that when considering both contraction and expansion areas, a negative balance is predicted under RCP 4.5, suggesting that climate change would have an adverse impact on the environmental suitability for D. composita. The contraction in the environmental suitability area in México would affect the states of Durango, Michoacán, San Luis Potosí, Tamaulipas, Oaxaca, Quintana Roo, Yucatán, Campeche, Veracruz, and Chiapas, whilst other affected areas would be located in Guatemala, Belize, El Salvador, and Honduras. Opposite, with RCP 8.5, a positive balance is predicted, which includes an increase in the environmental suitability area for D. composita. The expansion of the environmental suitability would imply territory of México in the states of Durango, Jalisco, Michoacán, Tamaulipas, Yucatán, and Campeche, as well as territory of Guatemala, El Salvador, Honduras, and Nicaragua.
4. Discussion
D. composita occurrence sites are concentrated in a region of humid warm climates that goes from the north of Veracruz, México, to the north of Guatemala. Since México-Central America is the most likely center of origin for D. composita [1], this geographic area could be suggested as a possible center of origin of this species, although, this hypothetical assertion must be proven with phylogenetically targeted genomic, morphological, and Earth system data [34]. In addition, as expected, niche modeling with MaxEnt showed this region with the highest environmental suitability for D. composita, which appends another argument to support such a hypothetical suggestion, since it is known that the centers of origin of the species meet the best environmental conditions for their growth [35].
Even when D. composita mostly distributes in humid warm climates, the presence of occurrence sites in other environmental conditions evidences the species’ capabilities to adapt to an ampler climatic scope. This provides D. composita the possibility of better adaptation to novel climates that global warming is bringing on [36,37,38].
The former results match with previous reports that state that the species of the genus Dioscorea are distributed in warm climates as well as in temperate zones [39], even at elevations above 2,200 m [40]. Likewise, our results agree with previous findings that Dioscorea species also are adapted to both tropical and subtropical zones [41,42].
According to annual mean precipitation range characterization for 18 species of Dioscorea genus with a presence in India [40], they distribute in environments with annual precipitation ranging from 2165 to 3778 mm, reinforcing the wetland plants’ character. In our research, we obtained a range of 737 to 4,874 annual millimeters; the value 737 mm corresponds with an accession located on the borderline of Mexican states Guerrero and Oaxaca, near the Pacific coast. In fact, 12 of 27 occurrence sites located near the Pacific coastline, register annual precipitation lower than 1,500 mm, indicating that D. composita has strived to adapt to novel drier environments. These accessions could be of interest for a possible breeding program focused on giving rise to varieties better capable to cope with climate change drought episodes.
Photoperiod also was determined to be an important variable for D. composita distribution. Short days have been reported to favor Dioscorea species tubers, whereas a photoperiod of more than 12 hours has been shown to promote the growth of leaf area, long stems, and vigorous vines [43,44].
Results evidenced that annual thermal oscillation (ATO) and annual thermal range (ATR) contributed to explaining D. composita presence and distribution; the optimal intervals determined for D. composita are 10.1-12.9 °C and 16.9-20.2 °C, respectively (Table 4). These values are indicative of a species not typically from thermo-regulated climates, as might be expected due to its presence in mostly humid and warm climates, but rather of a species that tolerate the occurrence of extreme temperatures. According to the Köppen-García Climate Classification System [45], these environments correspond to extreme thermal climates (ATO = 7 – 14 °C).
Growth of the Dioscorea species requires temperatures in the range of 25 to 30 °C to exhibit normal development; we obtained a very similar range (23-28 °C, Table 4) as an optimal interval in this parameter [41]. This author also reports that the growth of Dioscorea species is restricted by temperatures below 20 °C; such conditions are present in the occurrence sites located in the original region of distribution of D. composita (yellow color area in map of Figure 3). However, for the rest of the distribution sites, temperatures keep below 20 °C many days during the year (Supplemental File), denoting D. composita is being subjected to adaptation in other environments.
Warm temperatures favor foliage growth, but they also favor high respiration rates, which retard tuber growth and alter the production of diosgenin [46]. However, the optimal range determined in this research for the mean maximum temperature in the hot season (May-October) was 29-35 °C, which supposes certain comfort status of D. composita even under temperatures considered extreme (>32 °C) for many plant species [47,48].
Niche model with an AUC value greater than 0.7 can make good estimations [49]. Classify the accuracy of the models according to their AUC value in five categories: 0.50-0.60 insufficient model; 0.60-0.70 poor model; 0.70-0.80 average acceptable model; 0.80-0.90 good model, and 0.90-1.00 excellent model [50,51]. According to this classification, the obtained models for all climatic scenarios studied are excellent to describe the potential distribution of D. composita and constitute an adequate tool to derive the eco-geographic characterization of the territories where this species is distributed [15].
The results of the Jackknife test revealed that the environmental variables that most determine the presence and distribution of D. composita are AMAI, NAPH, ATO, MOMAI, NAMAI, Bio14, and Bio11 (Table 5). These results partially match with previous reports y [37] which mention that the most important variables in the growth and development of the species, especially in the production of diosgenin, are precipitation and solar radiation. For Solanum tuberosum with which the species of the genus Dioscorea are compared for the production of tubers, the most important environmental variables are annual precipitation and average soil temperature, in such a way that at higher soil temperature and lower soil moisture, the distribution of the species that produce tubers is limited [52].
Most studies about the effects of climate change on the environmental suitability for species, report more contraction than expansion areas for their potential distribution [53]. However, this type of climate change effects should not be considered a generalization, since the new environmental conditions brought about by climate change can represent comparative advantages for diverse species, mostly causing the expansion of their potential distribution areas [54]; also, some species could remain in their current distribution areas without being significantly affected by climate change. In this way, species of origin and adaptation to temperate environments would see their potential distribution reduced by the year 2050, as it is the case of Solanum tuberosum [52]; while species of tropical origin and adaptation to warm environments would benefit from the expansion of their potential distribution areas; an example of this statement is Dioscorea alata, which is predicted to have a significant increase in production and potential distribution area by 2040s decade [55].
Environmental factors are the main drivers of changes in the distribution of Dioscorea species [56]. Based on the values in Table 6, which describe the dynamics of environmental suitability for D. composita under different climate change scenarios, it is concluded that scenario RCP 4.5 would have a negative impact on the species, with a decrease in its potential distribution area. Loss of biodiversity, as a consequence of climate change, has been observed in similar environments in other species such as passion fruit, which, being a climbing species, will also be affected in scenario RCP 4.5, decreasing its potential area due to increased temperature and decreased precipitation (and hence, reduced water availability), and new physico-chemical soil characteristics [57].
Based on the results, the year 2050 RCP 8.5 climatology would have a positive effect on the environmental suitability of D. composita. However, studies on the effects of climate change indicate mainly impacts on areas of environmental suitability, resulting in contraction areas being greater than expansion areas [53]. Notwithstanding the foregoing, the response of each plant species may differ, with some decreasing their distribution, others changing or expanding it, and perhaps some not being affected at all [54]. While climate change has the potential to negatively affect the distribution and biodiversity of plant species, specific effects may vary depending on the species and the scenario considered. In the case of D. composita, the RCP 8.5 scenario suggests a positive effect on its potential distribution area. Furthermore, the variation in the impacts of climate change on population growth rates is mainly due to differences in the climatic response of the species populations [58].
5. Conclusions
Since results of this research showed that D. composita is mainly distributed in a compact zone of southern México-Central America with warm and humid climates. However, some D. composita populations were found to adapt to sub-humid and semi-arid conditions, and to semi-warm and temperate environments, which could contribute to this plant better cope with environmental stress that climate change is imposing through drought and heat episodes.
Modeling of environmental suitability for current conditions allowed for the identification of a region that stretches from northern Veracruz, México, to northern Guatemala in Central America, which could be the center of origin of D. composita. The parameters that most influence its distribution are the annual and seasonal moisture availability indices, the November-April mean photoperiod, the annual mean thermal oscillation, the precipitation of the driest month, and the mean temperature of the coldest quarter.
According to climate change scenarios for the year 2050, the research indicates a decrease in environmental suitability for D. composita under the RCP 4.5 scenario and an increase under the RCP 8.5 scenario, thus, this species could be a good crop option under this scenario of emissions.
The data generated in this research can contribute to a better understanding of the plant-environment interactions for D. composita, thus, easing the determination of its potentiality as a crop under the current and future climatologies, as well as design strategies for its natural populations in México and Central America. Such possible actions will undoubtedly be very useful now that D. composita is back in the public interest for its nutraceutical properties.
References
- Couto, R.S.; Martins, A.C.; Bolson, M.; Lopez, R.C.; Smidt, E.C.; and Braga, J.M.A. Time calibrated tree of Dioscorea (Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene. Botanical Journal of the Linnean Society, 2018, 188, 144–160. [Google Scholar] [CrossRef]
- Martin, F.W.; Ortiz, S. New chromosome numbers in some Dioscorea species. Cytologia 1966, 31, 105–107. [Google Scholar] [CrossRef]
- Álvarez, Q.V. , Caso, B.L., Aliphat, F.M. Galmiche TÁ. Plantas medicinales con propiedades frías y calientes en la cultura Zoque de Ayapa. Tabasco, México. Boletín Latinoamericano y del Caribe de plantas medicinales y aromáticas. 2017, 16, 428–454. [Google Scholar]
- Zhang, S.; Fan, M.; Ye, G.; Zhang, H.; Xie, J. Biorefinery of Dioscorea composita Hemsl with ferric chloride for saponins conversion to diosgenin and recycling the waste to biomethane. Industrial Crops and Products. 2019, 135, 122–129. [Google Scholar] [CrossRef]
- Wei, M.; Bai, Y.; Ao, M.; Jin, W.; Yu, P.; Zhu, M.; Yu, L. Novel method utilizing microbial treatment for cleaner production of diosgenin from Dioscorea zingiberensis CH Wright (DZW). Bioresource technology. 2013, 146, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Román-Cortés, N. R. , García-Mateos, M. D. R., Castillo-González, A. M., Sahagún-Castellanos, J., & Jiménez-Arellanes, M. A. Características nutricionales y nutracéuticas de hortalizas de uso ancestral en México. Revista fitotecnia mexicana. 2018, 41, 245–253. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen (Para adaptarlo a las condiciones de la República Mexicana). 5ª. Edición Instituto de Geografía UNAM-Enriqueta García. Ciudad de México. 2004, pp.90. http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/view/83/82/251-1.
- Bolaños, G.B. Biopolíticas del cambio climático para Centroamérica. Trace (México, DF). 2018a, 74, 135–158. [Google Scholar] [CrossRef]
- Ruiz-Corral, J.A.; Medina-García, G.; Flores-López, H.E. , Ramírez-Díaz, J.L., De la Cruz-Larios, L.; Villalpando-Ibarra, J.F.; Ruiz-Álvarez, O. Impacto del cambio climático sobre la estación de crecimiento en el estado de Jalisco, México. Revista Mexicana Ciencias Agrícolas . 2016; 13, 2627–2638. Available online: http://www.scielo.org.mx/scielo.php?pid=S200709342016000902627&script=sci_arttext.
- Lobato, S.R.; Altamirano, C.M.A. Detección de la tendencia local del cambio de la temperatura en México. Tecnología y Ciencias del Agua. 2017, 8, 101–116. [Google Scholar] [CrossRef]
- Rodríguez, M.V.M; Medina, G.G.; Díaz, P.G.; Ruiz, C.J.A.; Estrada, A.J.E.; Mauricio, R.J.E. ¿Por qué México es un país altamente vulnerable al cambio climático? Revista Mexicana Ciencias Agrícolas. 2021, 25, 45–57. [Google Scholar] [CrossRef]
- Mora, C.; Caldwell, I.R.; Caldwell, J.M.; Fisher, M.R.; Brandon, M.G.; Running, S.W. Suitable days for plant growth disappear under projected climate change: potential human and biotic vulnerability. PloS one Biology. 2015, 587. [Google Scholar] [CrossRef]
- Ruiz-Corral, J.A.; Medina-García, G.; García-Romero, G. Sistema de información agroclimática para México-Centroamérica. Revista Mexicana Cienciencas. Agrícolas. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Barrera-Sánchez, C.F.; Ruiz-Corral, J.A.; Zarazúa, V.P.; Lépiz, I.R.; González, E.D.R. Cambio climático y distribución potencial de frijol lima en Mesoamérica y Aridoamérica. Revista Mexicana de Ciencias Agrícolas. 2020, 11, 1361–1375. [Google Scholar] [CrossRef]
- Kong, F.; Tang, L.; He, H.; Yang, F.; Tao, J.; Wang, W. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environmental Science and Pollution Research. 2021, 28, 34655–34663. [Google Scholar] [CrossRef]
- Sánchez-González, J.D.J.; Ruiz, C.J.A.; García, G.M.; Ojeda, G.R.; Larios, L.D.L.C.; Holland, J.B.; García, R.G.E. Ecogeography of teosinte . PLoS One. 2018, 13, e0192676. [Google Scholar] [CrossRef]
- Eastman, J.R. Idrisi Selva Manual. Clark Labs, Clark University. Worcester, MA, USA. 2012, 322 p.
- Bellouin, N.; Boucher, O.; Haywood, J.; Johnson, C.; Jones, A.; Rae, J.; Woodward, S. Improved representation of aerosols for HadGEM2. Hadley Centre Technical Note 73, Met Office Hadley Centre. 2007. Available online: https://www.google.com/search?q=Improved+representation+of+aerosols+for+HadGEM2.+Hadley+Centre+Technical+Note+73%2C+Met+Office+Hadley+Centre.+Exeter&rlz=1C1ALOY_esMX969MX973&oq=Improved+representation+of+aerosols+for+HadGEM2.+Hadley+Centre+Technical+Note+73%2C+Met++Office+Hadley+Centre.+Exeter&aqs=chrome..69i57.1981j0j15&sourceid=chrome&ie=UTF-8.
- 19. Instituto de Ecología (INE). México cuarta comunicación nacional ante la convención marco de las Naciones Unidas sobre cambio climático. Secretaría de Medio Ambiente y Recursos Naturales, (SEMARNAT). D. F México. 2009, 274 p. Available online: https://www.gob.mx/cms/uploads/attachment/file/666958/cuarta_Comunicacion.pdf.
- Conde, C.; Estrada, F.; Martinez, B.; Sánchez, O.; Gay, C. Regional climate change scenarios for México. Atmósfera. 2011, 24, 125–140. Available online: http://www.scielo.org.mx/pdf/atm/v24n1/v24n1a9.pdf.
- Zuza, E.J.; Maseyk, K.; Bhagwat, S.A.; De Sousa, K.; Emmott, A.; Rawes, W.; Araya, Y.N. . Climate suitability predictions for the cultivation of macadamia (Macadamia integrifolia) in Malawi using climate change scenarios. PloS one. 2021, 16, e0257007. [Google Scholar] [CrossRef] [PubMed]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop: Release 10. Environmental Systems Research Institute. Redlands, CA, USA. 2010, 15 p. Available at. http://earthobservations.org/about_geo.shtml.
- Newbold, T. Applications and limitations of museum data for conservation and ecology, with particular attention to species distribution models. Progress in Physical Geography. 2010, 34, 3–22. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L. , Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecological Modelling. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- del Valle, M.J.; Guerra, B.W. La multicolinealidad en modelos de regresión lineal múltiple. Revista Ciencias Técnicas Agropecuarias. 2012, 21(4), 80–83. Available online: http://scielo.sld.cu/pdf/rcta/v21n4/rcta13412.pdf.
- Core, T.R. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Austria. 2021. Available at. https://www.R-project.org/.
- Jayasinghe, S.L.; Kumar, L. Modeling the climate suitability of tea [Camellia sinensis (L.) O. Kuntze] in Sri Lanka in response to current and future climate change scenarios. Agricultural and Forest Meteorology. 2019, 272, 102–117. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Osorio, O.L.; Jiménez, G.D. An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecological Informatics. 2019, 53, 100983. [Google Scholar] [CrossRef]
- Peterson, A.T. , Cobos, M.E., Jiménez, G.D. Major challenges for correlational ecological niche model projections to future climate conditions. Annals of the New York Academy of Sciences. 2018, 1429, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; DeWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecological Modelling. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Flores, T.M.; Ortiz, E. ; Villaseñor. J.L. Modelos de nicho ecológico como herramienta para estimar la distribución de comunidades vegetales. Revista Mexicana de Biodiversidad 90. 2220. [Google Scholar]
- Goettsch, B.; Urquiza, H.T.; Koleff, P. Acevedo, G.F. Extinction risk of Mesoamerican crop wild relatives. Plants, People, Planet. 2021, 3, 775–795. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The Effects of Sampling Bias and Model Complexity on the Predictive Performance of MaxEnt Species Distribution Models. PLoS one. 2013, 8, e55158. [Google Scholar] [CrossRef]
- Bowles, A. M. , Williamson, C. J., Williams, T. A., Lenton, T. M., & Donoghue, P. C. The origin and early evolution of plants. Trends in Plant Science. 2022, 28, 312–329. [Google Scholar] [CrossRef]
- Purseglove, J.W. Tropical crops: Monocotyledons. Longman Scientific and Technical. New York, NY, USA; 1985.
- Villers, R.L.; Trejo, V.I. El cambio climático y la vegetación en México. México: Una Visión Hacia El Siglo XXI El Cambio Climático En México, Universidad Nacional Autónoma de México, México. D.F. 2000, 57-66. Available at. https://www.uv.mx/personal/tcarmona/files/2010/08/Villers-y-Trejo-.pdf.
- Shen, L.; Xu, J.; Luo, L.; Hu, H.; Meng, X.; Li, X.; Chen, S. Predicting the potential global distribution of diosgenin-contained Dioscorea species. Chinese medicine. 2018, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Kipling, R.P.; Topp, C.F.; Bannink, A.; Bartley, D.J.; Blanco, P.I.; Cortignani. R.; Eory, V. To what extent is climate change adaptation a novel challenge for agricultural modellers? Environmental Modelling & Software. 2019, 120, 104492. [Google Scholar]
- Téllez, V.O.; Ramírez, R.R.O. Las dioscóreas (Dioscoreaceae) del Estado de Morelos, México. Anales del Instituto de Biología serie Botánica. 1992, 63-001. Available at. https://www.redalyc.org/pdf/400/40063104.pdf.
- Saikia, B.; Rawat, J.S; Tag, H.; Das, A.K. An investigation on the taxonomy and ecology of Dioscorea in Arunachal Pradesh, India. Journal of Frontline Research. 2011, 01, 44–53. Available online: https://d1wqtxts1xzle7.cloudfront.net/31930407/Dioscorea_paper_B_saikia-with-cover-page-v2.pdf?
- Rodríguez, W. Botánica, domesticación y fisiología del cultivo de ñame (Dioscorea alata). Agronomía Mesoamericana. 2000, 11, 133–152. Available online: https://www.redalyc.org/pdf/437/ 43711221.pdf. [CrossRef]
- González, V.M.E. El ñame (Dioscorea spp.). Características, usos y valor medicinal. Aspectos de importancia en el desarrollo de su cultivo. Cultivos Tropicales. 2012, 33, 05–15. Available online: http://scielo.sld.cu/scielo.php?pid=S0258-59362012000400001&script=sci_arttext&tlng=en.
- Coursey, D.G. Yams: Dioscorea spp. In: J. Smartt & N. Simmonds, Evolution of Crop Plants. Wiley 2nd Edition. London. 1976, pp.70-74. Available at. https://agris.fao.org/agrissearch/search.do?recordID=US201303063121.
- Puga, B.E. Manejo agronómico del cultivo de ñame. Guía técnica. 2 ed. MIDA. Panamá; 1995.
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen (Para adaptarlo a las condiciones de la República Mexicana). 5ª. Edición Instituto de Geografía UNAM-Enriqueta García. Ciudad de México, México. 2004, 90 p. http://www.publicaciones.igg.unam.mx/index.php/ig/catalog/view/83/82/251-1.
- Onwueme, IC. Sett weight effects on time of tuber formation, and on tuber yield characteristics, in water yam (Dioscorea alata L.). Journal Agricultural Science. 1978, 91, 317–319. [Google Scholar] [CrossRef]
- Ruiz-Corral, J.A.; Medina, G.G.; González, A.I.J.; Flores, L.H.E.; Ramírez, O.G.; Ortiz, T.C.; Byerly, M.K.F.; Martínez, P.A.R.A. Requerimientos agroecológicos de cultivos. Libro Técnico Núm. 3. 2ª Ed. INIFAP-Prometeo Editores. Guadalajara, Jalisco, México. 2013, pp.564 Available at. https://www.researchgate.net/publication/343047223_REQUERIMIENTOS_AGROECOLOGICOS_DE_CULTIVOS_2da_Edicion.
- FAO. ECOCROP Database. Food and Agriculture Organization of the United Nations. Rome, Italy.2019.
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecological Modelling. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Reynoso, S.R.; Pérez, H.M.J.; López, B.W.; Hernández, R.J.; Muñoz, F.H.J.; Cob, U.J.V.; Reynoso, S.M.D. El nicho ecológico como herramienta para predecir áreas potenciales de dos especies. Revista Mexicana de Ciencias Forestales. 2018, 9, 47–68. [Google Scholar] [CrossRef]
- Navarro, G.M.A.; Jove, C.C.A.; Ignacio, A.J.M. Modelamiento de nichos ecológicos de flora amenazada para escenarios de cambio climático en el departamento de Tacna-Perú. Colombia Forestal. 2020, 23, 51–67. [Google Scholar] [CrossRef]
- Khalil., T.; Asad, S.A.; Khubaib, N.; Baig, A.; Atif, S.; Umar, M.; Baig, S. Climate change and potential distribution of potato (Solanum tuberosum) crop cultivation in Pakistan using Maxent. AIMS Agriculture and Food. 2021, 6, 663–676. [Google Scholar] [CrossRef]
- Sork, V.; Davis, F.; Westfall. ; R.; Flints, A.; Ikegami, M.; Wang, H.; Grivet, D.D. Gene movements and genetic association with regional gradients in California valley oak (Quercus lobata Née) in the face of climate change. Molecular Ecology. 2010, 19, 3806–3823. [Google Scholar] [CrossRef]
- Ovando, H.N.; Tun, J.; Mendoza, G.G.; Parra, T.V. Efecto del cambio climático en la distribución de especies clave en la vegetación de duna costera en la península de Yucatán. México. Revista Mexicana de Biodiversidad 90 2019. [Google Scholar]
- Srivastava, A.K.; Gaiser, T.; Paeth, H.; Ewert, F. The impact of climate change on Yam (Dioscorea alata) yield in the savanna zone of West Africa. Agriculture, Ecosystems & Environment. 2012, 153, 57–64. [Google Scholar]
- Du, Z.X.; Wu, J. Meng, X.X.; Li, J.H.; Huang, L.F. Predicting the global potential distribution of four endangered Panax species in middle-and low-latitude regions of China by the geographic information system for global medicinal plants (GMPGIS). Molecules. 2017, 22, 10–1630. [Google Scholar] [CrossRef]
- Munar, A.M.; Carlosama, A.R.; España, J.L.M. Potenciales áreas cultivables de pasifloras en una región tropical considerando escenarios de cambio climático. Revista de Investigación Agraria y Ambiental. 2022, 13, 109–129. [Google Scholar] [CrossRef]
- Louthan, A.M.; Morris, W. Climate change impacts on population growth across a species’ range differ due to nonlinear responses of populations to climate and variation in rates of climate change. PloS one. 2021, 16, e0247290. [Google Scholar] [CrossRef]
Figure 1.
Current distribution of D. composita in agroclimatic regions of México and Central America.
Figure 1.
Current distribution of D. composita in agroclimatic regions of México and Central America.
Figure 2.
Models obtained and evaluated by Kuenm R-Package for D. composita.

Figure 3.
Results of Jackknife test of the relative importance of predictor environmental variables in MaxEnt model for D. composita in México and Central America.
Figure 3.
Results of Jackknife test of the relative importance of predictor environmental variables in MaxEnt model for D. composita in México and Central America.
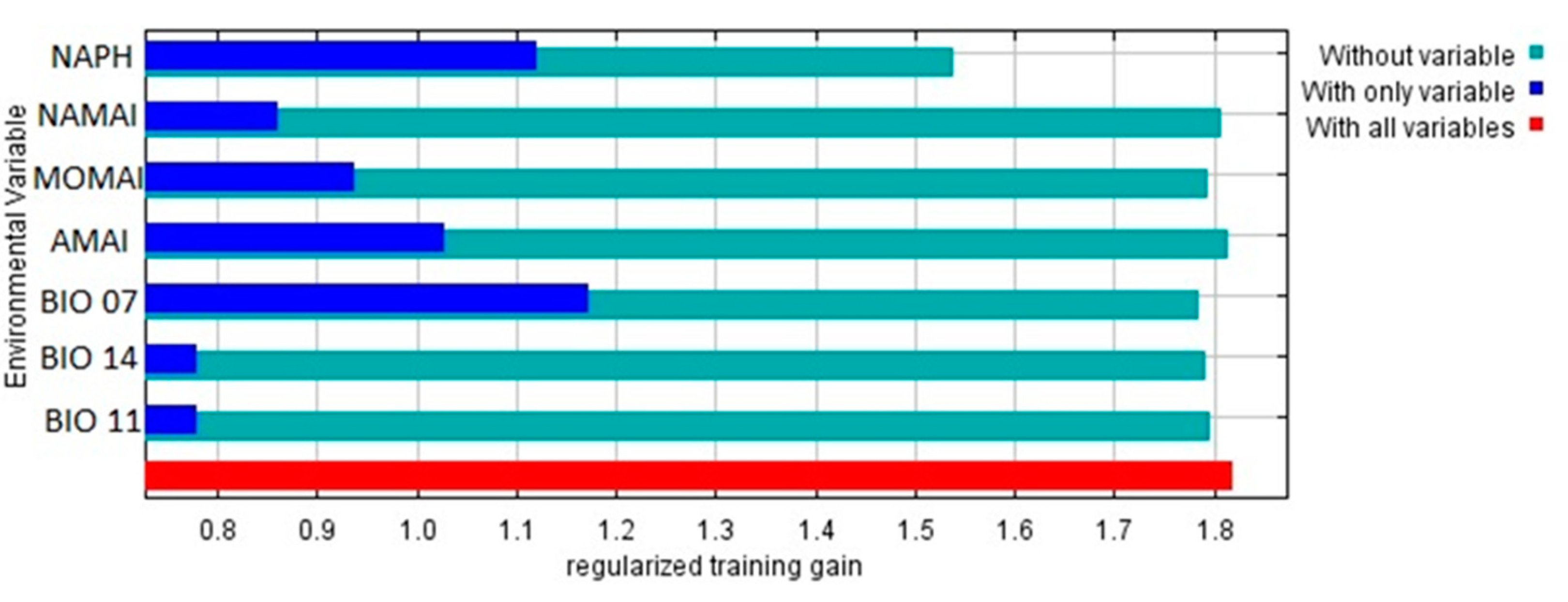
Figure 4.
Areas with environmental suitability for D. composta in México and Central America.
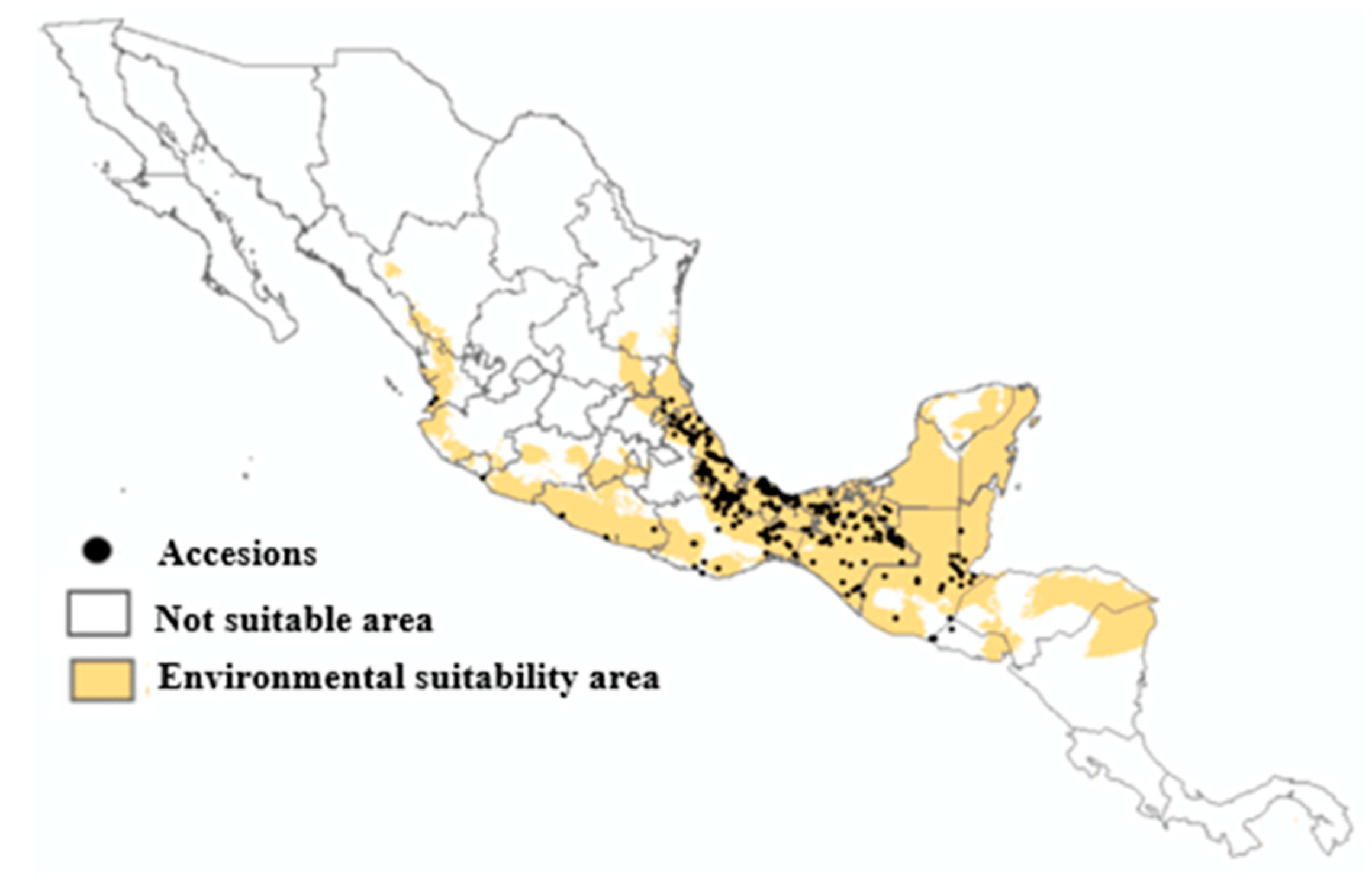
Figure 5.
Current and potential distribution of D. composita under three climatic scenarios: a) 1961-2010; b) year 2050 rcp 4.5; c) year 2050 rcp 8.5. Areas of contraction and expansion of environmental suitability are shown in orange and green colors, respectively.
Figure 5.
Current and potential distribution of D. composita under three climatic scenarios: a) 1961-2010; b) year 2050 rcp 4.5; c) year 2050 rcp 8.5. Areas of contraction and expansion of environmental suitability are shown in orange and green colors, respectively.
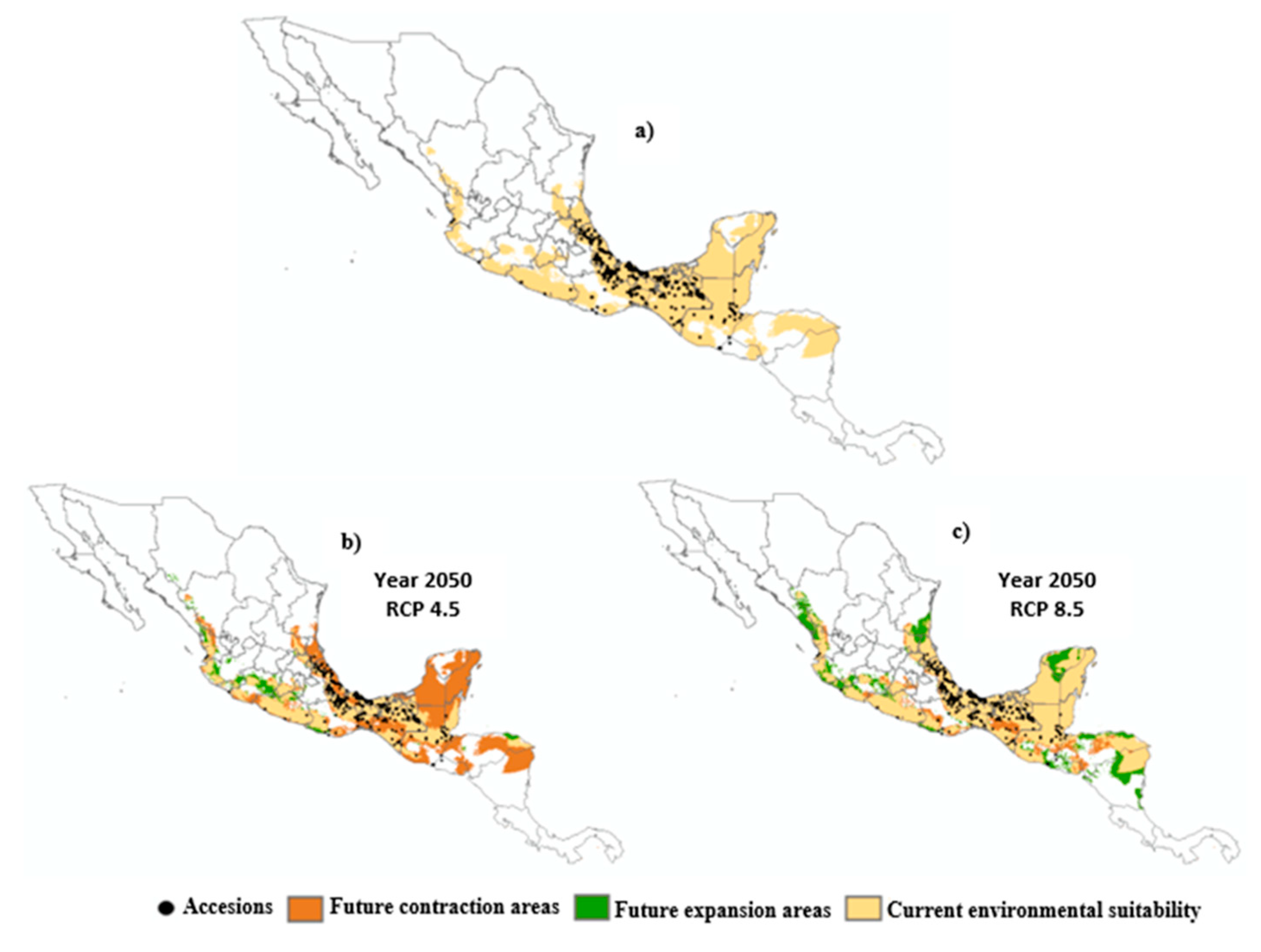

Table 1.
Sources of occurrence data for D. composita in México-Central America.
| Institution | Accessions | |
|---|---|---|
| Universidad Nacional Autónoma de México (Instituto de Biología) | 179 | |
| Instituto Nacional de Ecología y Cambio Climático | 131 | |
| Instituto Nacional de Estadística, Geografía e Informática (Dep. Botánica) | 3 | |
| Universidad Autónoma de Veracruz (Centro de Investigaciones Tropicales) | 1 | |
| Universidad Autónoma de Veracruz (Instituto de Investigaciones Biológicas) | 10 | |
| Colegio de la Frontera Sur (Unidad Tapachula) | 5 | |
| Universidad Autónoma de Chiapas | 5 | |
| Universidad Autónoma de Puebla | 4 | |
| Universidad Juárez Autónoma de Tabasco | 5 | |
| Universidad Autónoma Benito Juárez de Oaxaca | 4 | |
| Universidad Autónoma de Guerrero | 8 | |
| JSTOR Plant Science and the Global Plants Initiative | 3 | |
| Trópicos.org | 1 | |
| Red de Herbarios del Noroeste de México. | 16 | |
| Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Herbario Digital) | 7 | |
| Global Biodiversity Information Facility (GBIF) | 26 | |
| Total | 408 | |
Table 2.
Environmental variables considered in this research.
| Variable | Description | Temporal scale |
|---|---|---|
| BIO01 | (Annual mean temperature) | Annual |
| BIO05 | (Maximum temperature of the warmest month) | Month |
| BIO06 | (Minimum temperature of the coldest month) | Month |
| BIO07 | Temperature annual range | Annual |
| BIO08 | (Mean temperature of the wettest quarter) | Quarter |
| BIO09 | (Mean temperature of the driest quarter) | Quarter |
| BIO10 | (Mean temperature of the warmest quarter) | Quarter |
| BIO11 | (Mean temperature of the coldest quarter) | Quarter |
| BIO12 | Annual precipitation | Annual |
| BIO13 | Precipitation of the wettest month | Month |
| BIO14 | Precipitation of the driest month | Month |
| BIO16 | Precipitation of the wettest quarter | Quarter |
| BIO17 | Precipitation of the driest quarter | Quarter |
| BIO18 | Precipitation of the warmest quarter | Quarter |
| BIO19 | Precipitation of the coldest quarter | Quarter |
| N-AMT | November-April mean temperature | Seasonal |
| M-OMT | May-October mean temperature | Seasonal |
| M-OXT | May-October maximum temperature | Seasonal |
| N-AXT | November-April maximum temperature | Seasonal |
| AXT | Annual maximum temperature | Annual |
| M-OIT | May-October minimum temperature | Seasonal |
| N-AIT | November-April minimum temperature | Seasonal |
| AIT | Annual minimum temperature | Annual |
| ATO | Annual thermal oscillation | Annual |
| M-OP | May-October precipitation | Seasonal |
| N-AP | November-April Precipitation | Seasonal |
| M-OPH | May-October photoperiod | Seasonal |
| N-APH | November-April photoperiod | Seasonal |
| AMAI | Annual moisture availability index | Annual |
| M-OMI | May-October moisture availability index | Seasonal |
| N-AMI | November-April moisture availability index | Seasonal |
Table 3.
Intervals of annual moisture availability index and annual mean temperature for 10 agro-climatic regions of México-Central America where D. composita is currently distributed.
Table 3.
Intervals of annual moisture availability index and annual mean temperature for 10 agro-climatic regions of México-Central America where D. composita is currently distributed.
| Agro-climatic region | Annual moisture availability index* | Annual mean temperature (°C)* |
|---|---|---|
| Semi-arid very warm | 0.2 to 0.5 | > 26 |
| Dry sub-humid very warm | 0.5 to 0.65 | > 26 |
| Dry sub-humid temperate | 0.5 to 0.65 | 12 to 18 |
| Humid sub-humid semi-warm | 0.65 to 1.0 | 18 to 22 |
| Humid sub-humid warm | 0.65 to 1.0 | 22 to 26 |
| Humid sub-humid very warm | 0.65 to 1.0 | > 26 |
| Humid temperate | > 1.0 | 12 to 18 |
| Humid semi-warm | > 1.0 | 18 to 22 |
| Humid warm | > 1.0 | 22 to 26 |
| Humid very warm | > 1.0 | > 26 |
Table 4.
Ecological descriptors of D. composita, based on 1961-2010 average values.
| Variable | Minimum | Maximum | Optimum |
|---|---|---|---|
| Annual mean moisture index | 0.49 | 4.95 | 1.128-2.932 |
| May-October mean moisture index | 0.92 | 10.52 | 2.02-4.78 |
| November-April mean moisture index | 0.026 | 3.82 | 0.62-2.62 |
| Annual mean precipitation | 737 | 4874 | 1940-3839 |
| May-October mean precipitation (mm) | 648 | 3658 | 1524-2880 |
| November-April mean precipitation (mm) | 26 | 1657 | 416-1059 |
| Annual mean temperature | 12.1 | 29 | 23-28 |
| Annual mean maximum temperature | 17 | 35 | 28-33.5 |
| Annual mean minimum temperature | 7.2 | 23 | 18-22.5 |
| May-October mean temperature | 11 | 31.9 | 24-29 |
| May-October mean maximum temperature | 15.5 | 39.4 | 29-35 |
| May-October mean minimum temperature | 6.51 | 24.5 | 19-23 |
| November-April mean temperature | 11.6 | 30.3 | 22-26.9 |
| November-April mean maximum temperature | 17.8 | 35.7 | 26.5-32 |
| November-April mean minimum temperature | 5.5 | 24.9 | 17.5-21.8 |
| May-October mean photoperiod | 12.30 | 13.59 | 12.90-11.15 |
| November-April mean photoperiod | 10.44 | 11.70 | 11.27-11.50 |
| Annual mean thermal oscillation | 8.22 | 21.10 | 10.1-12.9 |
| Annual range of temperature | 11.73 | 22.4 | 16.87-20.15 |
| Elevation | 5 | 3001 | 5-1125 |
Table 5.
Contribution of seven environmental variables to depict the presence and distribution of D. composita.
Table 5.
Contribution of seven environmental variables to depict the presence and distribution of D. composita.
| Environmental variables | Contribution (%) | Permutation importance (%) | |
|---|---|---|---|
| Annual moisture availability index | 38.4 | 3.5 | |
| November-April photoperiod | 20.7 | 26.8 | |
| Annual thermal oscillation | 17.1 | 9.5 | |
| May-October moisture index | 9.1 | 42.7 | |
| November-April moisture availability index | 8.4 | 6.2 | |
| Precipitation of the driest month | 3.7 | 2.9 | |
| Mean temperature of the coldest quarter | 2.5 | 8.3 | |
Table 6.
Current and predicted environmental suitability by the year 2050 for Dioscorea composita in México-Central America.
Table 6.
Current and predicted environmental suitability by the year 2050 for Dioscorea composita in México-Central America.
| Area (km2) | |||
|---|---|---|---|
| 1961-2010 | 2050 RCP4.5 | 2050 RCP8.5 | |
| Environmental suitability | 692,123 | 365,680 | 763,589 |
| Suitability expansion | 46,826 | 154,530 | |
| Suitability contraction | 373,269 | 83,064 | |
| Suitability without change | 318,854 | 609,059 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



