
Submitted:
29 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Circulating extracellular microvesicles (cEVs) are characterised by presenting surface antigens of parental cells. Since their biogenesis involves the translocation of phosphatidylserine (PS) from the inner to the outer-leaflet of the plasma membrane, exposed PS has been considered as a recognition hallmark of cEVs. However, not all cEVs externalise PS. In this study we have phenotypically and quantitatively characterised cEVs by flow cytometry with special attention to the proportions of PS in chronic heart failure patients (cHF; n=119) and reference non-HF group (n=21). PS─-cEVs were predominantly found in both groups. Parental markers showed differential pattern depending on PS exposure. Endothelium-derived and connexin 43-rich cEVs were mainly PS─-cEVs and significantly increased in cHF. On the contrary, platelet-derived cEVs were mostly PS+ and were increased in non-HF. We observed similar levels of PS+- and PS─-cEVs in non-HF subjects when analysing immune cell-derived EVs, but there was a subset-specific difference in cHF patients. Indeed, those cEVs carrying CD45+, CD29+, CD11b+ and CD15+ were mainly PS+-cEVs, while those carrying CD14+, CD3+ and CD56+ were mainly PS─-cEVs. In conclusion, endothelial and red blood cells are stressed in cHF patients as detected by a high shedding of cEVs. Despite PS+-cEVs and PS─-cEVs represent two distinct cEV populations, their release and potential function as both biomarkers and shuttles for cell communication seem unrelated to PS content.
Keywords:
annexin V
; connexin 43
; endothelial cells
; extracellular vesicles
; heart failure
; leukocytes
; microvesicles
; phosphatidylserine
; platelets
; red blood cells
1. Introduction
Circulating extracellular vesicles (cEVs) are a heterogeneous population of small membrane particles involved in several physiological processes such as cell adhesion, cell-cell crosstalk, haemostasis and thrombosis, and it is demonstrated that they have a role in disease [1,2,3]. Microvesicles, with a size range between 20 and 1000 nm and originating by the blebbing of the cellular plasma membrane [4,5], are characterised for presenting in their membrane antigens from their parental cells and for carrying active molecules such as proteins, metabolites, RNA or DNA [4,5,6]. cEV release involves the loss of lipidic asymmetry of the plasma membrane exposing phospholipid phosphatidylserine (PS) in the outer membrane of the vesicle [5]. However, there are also cells that release cEVs without PS externalization [7,8,9,10,11,12].
Affecting approximately 2% of the worldwide population over 65 years of age and with an increasing incidence, heart failure (HF) accounts for a high proportion of hospitalization and cardiovascular death around the globe [13]. It is characterised by an impairment of the cardiac function that results in the inability of the cardiac muscle to reach the body’s metabolic demands. This complex clinical condition can derive from genetic or environmental insults that induce significant alterations of the heart function (decreased cardiac output, systolic or diastolic dysfunction) and structure (left ventricle dilation or hypertrophy, maladaptive remodelling, fibrosis) [14,15].
We previously investigated platelet-derived EVs (pEVs) [12] and white cell-derived EVs [16] in the circulation of chronic heart failure (cHF) patients. We also reported that patients with chronic conditions such as cHF present higher numbers of PS+-pEVs. Therefore, here we aimed to study cEVs without PS membrane exposure (PS─-cEVs) in patients with cHF and investigated their parental blood and vascular cell origin.
2. Results
2.1. Circulating Extracellular Vesicles with and Without Phosphatidylserine Membrane Exposure
The distribution of global PS─-EVs differed between non-HF and cHF groups as shown in Table 1. In the reference non-HF group, total number of EVs in the circulation were 804 (301 - 1105) / µL, with 51 % being PS+-EVs and 49 % being PS─-EVs. In the cHF group, total number of cEVs were 1,079 (426 – 4702) / µL, of those 16 % were PS+-EVs and 84% were PS─-EVs, indicating a differential exposure of PS (annexin V [AV]+ and AV─) in cEV membranes in non-HF individuals and cHF patients (P < 0.001, both) and between both groups (P = 0.022 [for PS+-EVs] and P = 0.007 [for PS─-EVs], respectively).
The relative amount of cEVs from distinct cell origin (Table 1) reveals that the majority of PS+-EVs were platelet-derived EVs (CD41a+-pEVs [integrin αIIbβ3]) accounting for 86 % in the reference non-HF group and 53% in cHF patient group (P < 0.001). Platelet also released PS─-cEVs with a 20 % in non-HF subjects and a 7 % in cHF patients (P = 0.032). In contrast, endothelial-derived EVs (CD309+-eEVs; vascular endothelial growth factor receptor 2 [VEGFR2]) without PS membrane exposure (PS─) represented a 13 % in non-HF and a 23 % in cHF (P = 0.012) while levels of CD309+/AV+-eEVs were almost null both in non-HF and cHF groups. Leukocytes are also a source of both PS─-EVs (CD45+-LEVs; leukocyte common antigen [LCA]; P = 0.586), with 6.01 % and 5.81 % in non-HF individuals and cHF patients, respectively, and PS+-CD45+-LEVs with 9.01 % of EVs in non-HF controls and 16.49 % of cEVs in patients with cHF (P = 0.016). Other identified cell-derived EVs were from erythrocytes (CD235ab+-ErEVs [glycophorin a and b]). While similar high levels of PS─-ErEVs were found in both groups (27.4 % in non-HF and 19.56 % in cHF; P = 0.245), PS+-ErEVs were mainly in cHF patients (24 %) representing only 1.33 % in non-HF subjects. Finally, an important proportion of connexin-43-carrying EVs (CX43+-EVs) were AV─ (18.06 % and 27.2 % in non-HF controls and cHF patients, respectively; P = 0.028). Small amount of CX43+/AV+-EVs was detected (0.56 % in non-HF versus 1.88 % in cHF group; P = 0.001). In summary, circulating PS─-cEVs with different cellular origin and relative releases differ between cHF and the reference non-HF groups.
2.2. Differential Patterns between Phosphatidylserine Positive and Negative Circulating Extracellular Vesicles in Non-Heart Failure Reference Group and Patients with Chronic Heart Failure
2.2.1. Endothelial-Derived Circulating Extracellular Vesicles
Differential distributions of PS+- and PS─ endothelial-derived cEVs were observed, being PS─-EVs the major proportion of eEVs both in cHF and non-HF groups (P ≤ 0.016, all), as displayed in Figure 1. Endothelium-originated PS─-eEV levels were higher in cHF patients compared to non-HF subjects, for both eEVs presenting markers of non-activated (CD309+/AV─ [VEGFR2]; P < 0.001) and activated (CD62E+/AV─ [E-selectin]; P = 0.007) endothelial cells. PS+-eEV levels were similar in both non-HF and cHF bloods, independently of the cell activation status.
2.2.2. Erythrocyte-Derived Circulating Extracellular Vesicles
Erythrocyte-derived cEVs were increased in patients with cHF compared with non-HF individuals, independently of phosphatidylserine exposure (PS─-EVs and PS+-EVs, P ≤ 0.015, both), as shown in Figure 2A. In cHF patients, significantly higher proportions of CD235ab+/AV+-EVs were observed compared to CD235ab+/AV─-EVs (P < 0.001), while no differences between them were observed in the reference non-HF group (P = 0.240).
2.2.3. Connexin-43-Carrying Circulating Extracellular Vesicles
Increased levels of PS─-EVs carrying connexin-43 (CX43+/AV─-EVs) were observed in cHF patients in comparison with non-HF subjects (P < 0.001), while CX43+-PS+-EVs were found in comparable proportions in non-HF controls and cHF patients (p=0.083). Despite both groups showed higher proportions of connexin-43-carrying PS─-EVs than PS+-EVs, only in the cHF patient group this rise was significant (P < 0.001). No differences between CX43+/AV+ and CX43+/AV─ were observed in the non-HF group (P = 0.065) (Figure 2B).
2.2.4. Platelet and Inflammatory Cell-Derived Circulating Extracellular Vesicles
Platelet-derived cEVs were mainly PS+-EVs in both non-HF subjects and patients with cHF, as previously shown [12]. In addition, while PS+-pEVs were found in lower amounts in cHF compared to non-HF controls, there were no differences in cEVs that did not expose PS, with the exception of CD31+/AV─ (Table 2). However, cEVs enriched with CD31 (PECAM-1) marker may originate either from platelets or the endothelium.
Leukocyte-derived EVs (LEVs) carrying pan-leukocyte CD45+ (LCA) antigen and CD29+ (β1 integrin), a marker of activated leucocytes, were found enriched in PS+-EVs, whereas monocyte CD14+ (LPS-receptor) epitope was found enriched in the PS─-EVs population both in non-HF individuals and cHF patients (P ≤ 0.002, all) (Figure 3).
Regarding distinct subtypes of inflammatory cell-derived EVs, cHF patients were characterised by displaying CD11b+ (macrophage-1 antigen [MAC-1], marker of activated leukocytes) and CD15+ (Sialyl Lewis X, marker of neutrophils) as well as CD3+ (T-cell co-receptor, lymphocyte marker) and CD56+ (neural cell adhesion molecule-1 [NCAM1], Natural Killer [NK] cell marker) epitopes in both PS+-EVs and PS─-EVs pools. Thus, CD11b+/AV+-LEVs and CD15+/AV+-nEVs were significantly increased compared to CD11b+/AV─-LEVs and CD15+/AV─-nEVs, respectively (P < 0.001, both). The contrary occurred with numbers of CD3+/AV+-ℓEVs and CD56+/AV+-NKc-EVs, which were reduced compared to CD3+/AV─-LEVs and CD56+/AV─-nEVs, respectively (P < 0.001, NKc-EVs), as shown in Figure S1.
Concerning non-HF reference group, no differences between PS+-EVs and PS─-EVs carrying CD11b+, CD15+, CD3+ and CD56+ surface molecules were observed in the non-HF subjects (P > 0.194, all). Indeed, both PS+-EVs and PS─-EVs had negligible content of CD3+ antigen observed in the reference non-HF group (Figure S1).
In general, subsets of specific LEVs were observed in lower levels in non-HF subjects compared to patients with cHF, independently of their PS exposure. Thus, significantly reduced levels of PS+-EVs carrying CD45+ (LEVs; P = 0.007), CD11b+ (LEVs; P < 0.001), CD14+ (mEVs; P = 0.036) and CD15+ (nEVs; P = 0.001) markers were observed in non-HF subjects in relation with patients with cHF. On the contrary, CD29+/AV+-EVs (LEVs; P < 0.001) were increased in non-HF controls compared to patients with cHF. Regarding PS─-EVs, significantly reduced levels of CD3+/AV─-EVs (ℓEVs; P = 0.003), CD45+/AV─-EVs (LEVs; P = 0.0036), CD15+/AV─-EVs (nEVs; P < 0.001) and CD56+/AV─-EVs (NKc-EVs; P = 0.005), were found in non-HF control subjects compared to cHF patients (Figure S1).
3. Discussion
No specific generic marker binding all circulating EVs has been found yet. Until now, surface phosphatidylserine content in EV membranes detected by annexin V labelling has been widely used as a universal reporter of cEVs. However, it is now well-recognised that EVs from different cell lineages can be released either with or without externalization of this phospholipid in their membranes [7,8,9,10,11,17]. PS─-EVs have been described in several distinct pathologies including multi organ dysfunction syndrome (MODS) and sepsis [18], systemic lupus erythematosus (SLE) [19,20], antiphospholipid syndrome (APS) [21,22], type 1 diabetes mellitus [23], ischaemic stroke (IS) and transient ischaemic attack (TIA) [24], familial hypercholesterolaemia [11,25], sickle cell disease [26], gastric cancer [27], carotid artery stenting [28], and ageing [29]. Literature regarding all-type cEVs in cHF is generally scarce [30,31,32,33] and there is no information on PS─-EVs. We have previously studied the role on PS+-EVs released by platelets [12] and immunity cells [16] in cHF patients. Here we have focused on the cardiovascular system-related (heart, blood, and vascular resident cells) circulating PS─-EVs from patients with chronic heart failure, unravelling that endothelial and red blood cells are stressed in the context of cHF as illustrated by high cEV shedding (Figure 4).
In agreement with other studies [21,22], we found that cHF is also characterised by displaying a higher quantity of EVs that do not expose PS in their membranes (PS─-EVs). Regarding the specific parental cell origin of cEVs, we have observed differential patterns of PS+-cEVs and PS─-cEVs in controls and in patients with cHF. Firstly, even though both groups presented higher levels of endothelium-derived cEVs that did not expose PS, cHF patients presented increased levels of PS─-cEVs compared to controls, supporting the notion of endothelial cell injury as a trigger of eEV release, in a similar fashion as in other conditions [34]. In fact, these results are also in agreement with Nozaki et al., who reported increased levels of cEVs presenting the endothelial cell marker CD144+ (VE-cadherin) but without evaluating their PS exposure in cHF patients [33]. Thus, increased levels of eEVs might reflect the endothelial dysfunction present in cHF [35].
Despite there are controversial results [7,8,9,10,11,12], we have also confirmed that platelet-derived cEVs are mainly PS+ in cHF patients, as we [12] and others [36] have previously described. It is interesting to highlight the role of CD31+-cEVs that are not exclusively platelet-derived and could also be shed by endothelial and white blood cells. The behaviour of CD31+/AV+-cEVs and CD31+/AV─-cEVs is completely different, the former following the trend of pEVs (decreased in cHF) and the later following the trend of eEVs (increased in cHF). In light of these data, the two EV subtypes could be considered as two independent populations. Indeed, other authors already have pointed out a differential pathophysiological role of EVs depending on their PS exposure profile. In a study comprising patients that had suffered an IS or TIA, CD41+/PS─, CD62P+/PS─ and CD142+/PS─ associated with an increased risk of the primary outcome (considered as fatal or non-fatal myocardial infarction and/or fatal or non-fatal recurrent ischaemic stroke), while CD41+/PS+, CD62P+/PS+ and CD142+/PS+ associated with a decreased risk [24].
The population of cEVs released from inflammatory cells showed the highest degree of complexity. Levels of specific PS─-LEVs derived from pan-leukocytes (CD45+/AV─-LEVs), lymphocytes (CD3+/AV─-ℓEVs), neutrophils (CD15+/AV─), and NK cells (CD56+/AV─-NKcEVs) were found elevated as compared to controls. Similarly to PS+-LEVs, PS─-LEVs could reflect the non-resolving inflammatory state characteristic of cHF patients [37,38]. In contrast to what we have observed with pEVs and eEVs, LEVs seem to present a variable pattern in terms of PS membrane externalization. The proportion of PS+-EVs / PS─-EVs exposure may be an index reflecting not only the cell origin but also the activation stimulus leading to cEV release.
Another class of cEVs often overlooked is those of red blood cell origin [39]. There were no differences between erythrocyte-derived PS+-EVs and PS─-EVs levels in the blood of controls, and both subtypes were significantly increased in the blood of cHF patients, being PS+-EVs the highest number of ErEVs. Interestingly, Shet et al. observed increased numbers of PS+-EVs ErEVs in sickle cell disease in comparison with controls, but when considering their PS exposure, found similar levels between PS+-EVs and PS─-EVs ErEVs [26]. On the contrary, increased levels of PS─-EVs ErEVs were observed in a study comprising patients with MODS and sepsis compared to controls, while levels of PS+-EVs ErEVs were similar [18].
To our knowledge levels of EVs presenting connexin-43 (CX43) have not been analysed before nor if this marker is co-expressed with PS. We have explored CX43, a widely distributed tissue interstitial gap junction protein as a surrogate measure of tissue and cell damage state. The majority of CX43+-cEVs were PS─-cEVs in both studied populations. High levels of PS─-CX43+-EVs in cHF patients add evidence on the myocardial stress and injury frequently observed in cHF.
Phosphatidylserine is not only a marker of apoptosis but also of other cell death mechanisms [40]. On the other hand, the biogenesis of those cEVs that do not expose PS in their membrane is still unknown. Several theories have emerged in the last years, including that PS─-EVs are true PS+-EVs with (1) very low PS exposure or (2) PS-masking (through endogenous lactadherin or β2 glycoprotein 1 [β2GP1] binding) [21,23,24]. The latter could either hamper proper PS+-EVs recognition or block PS-mediated EV clearance [21,22,24]. A previous work studying the clearance ratio of PS-exposing vesicles in immune mice (with high levels of the antiphospholipid antibody β2GP1) showed that PS─-EVs were cleared significantly slower than their PS-exposing counterparts [41]. This hypothesis was further supported by the findings of two studies in SLE and APS, where PS─-EVs were the main type of EVs found in patients but not in controls [19,22]. Similarly, it has been hypothesized that PS─-EVs could circulate in blood with a long half-life due to less uptake by liver macrophages [42]. Nevertheless, further research on the biological role of PS─-EV shedding is warranted. Increasing number of studies with patients that do not present antiphospholipid antibodies preventing EV clearance or detection have revealed high levels of PS─-EVs such as in type-1 diabetic patients and elderly individuals [23,29], while patients suffering of IS or TIA [24] and atrial fibrillation[43] exhibited greater amounts of PS+-EVs.
Furthermore, the pathophysiological role exerted by PS─-EVs remains also unclear. The presence of PS in the EV membranes might likely discriminate two populations with different roles, with PS+-EVs suggested being more procoagulant and PS─-EVs reflecting a proinflammatory state [8,29]. The potential procoagulant proprieties of PS+-EVs are widely accepted and have been already demonstrated in several studies [44,45]. In heart failure condition, Kou et al. determined that the exposure of PS in both blood cells and cEVs is related to the procoagulant state of these patients [31,46]. As of PS─-EV inflammatory role, high levels of PS─-EVs have been detected in inflammatory diseases such as SLE or APS [19,22]. Further, cHF patients present a chronic and unresolved inflammatory state, which could also be reflected in the high proportion of PS─-EV release. In addition, increased levels of inflammation are observed in elderly individuals in comparison with younger subjects [47], who were already shown to display higher proportions of PS─-EVs [29].
Although the study of EV population without PS exposure is still on its infancy, it is evident that their release and function is unrelated to those EVs bearing PS. Our clinical model of study involving a chronic heart disease as heart failure, which is a condition with a large systemic affectation and cellular stresses, compared with non-HF control subjects, has evidenced the presence in blood of different types of circulating EVs. While immune cells secrete variety of cEVs depending on cell type and red blood cells both PS─ and PS+-EVs, those released by platelets are mainly PS+-EVs and those released by endothelial cells and connexin 43+-rich cells are mainly PS─-EVs. These differences should be taken into consideration in future studies addressing the role of cEVs in cHF. In conclusion, our findings shed new light on the link between cellular stresses, endothelial dysfunction, pro-oxidant and pro-inflammatory environment, and organ damage in the context of chronic heart failure, as uncovered by a high degree of distinct cell activation processes leading to extracellular vesicle shedding in the circulation.
4. Materials and Methods
4.1. Healthy and Diseased Blood Donors
Guideline-directed medically treated chronic heart failure patients from the outpatient HF unit of our hospital were enrolled for this study, including patients with both preserved left ventricular ejection fraction (LVEF) (≥ 50%) and reduced LVEF (< 40%). As a reference to cHF patient group, a non-heart failure control group (for circulating extracellular microvesicle values) was matched by age (65.7 ± 8.6 years [non-HF] and 67.0 ± 11.8 years [cHF]) and sex (24% of women [non-HF] and 32% [cHF]). Individuals with HF with mild reduction in ejection fraction (40-50% of LVEF), previous inflammatory disorders, infections, sepsis or cancer, as well as pregnant women, were excluded from the study. Baseline demographic and pharmacological data of all individuals (cHF patients and reference non-HF subjects) recruited for the study are listed in Table 3 and Table S1, respectively.
The ethics committee for clinical research of our institution, Hospital de la Santa Creu i Sant Pau in Barcelona (Spain), approved the study protocol (Ref. 16/44), which was conducted according the Declaration of Helsinki. Before recruitment, a written informed consent was obtained from all participants.
4.2. Blood Sampling and Circulating Extracellular Vesicle Isolation
Venous blood was withdrawn from the cubital vein without tourniquet using a 20-gauge needle after 10-14 hours (h) of fasting into 3.8% sodium citrate tubes (Vacutainer, Becton Dickinson [BD]). After the extraction and within the first 2 h, samples were identically processed as follows: firstly, blood was centrifuged at 1,560 ×g for 20 min at 20ºC (Eppendorf 5810R GLOOB04932 centrifuge, A-4-81 rotor, Eppendorf) to avoid in vitro platelet activation. Next, platelet-poor plasma (PPP) was carefully aspirated, leaving about a 1 mm undisturbed layer on top of cells. Then, a second centrifugation to obtain platelet-free plasma (PFP) by ensuring a complete removal of cells was pursued at 1,500×g for 10 min at 20ºC (Eppendorf 5415R centrifuge, FA45-24-11 rotor, Eppendorf). Finally, until flow cytometry studies PFP aliquots were stored at -80ºC.
A two-step high-speed centrifugation was used to isolate cEVs from PFP, as previously described [16,48,49]. In brief, thawed PFP was centrifuged at 1,500 ×g for 10 min at 20ºC (5417R centrifuge, FA45-24-11 rotor, Eppendorf). Then, a volume of 250 µL of PFP from the upper part of the vial was carefully transferred to a new tube. This tube was then centrifuged at 20,000 ×g for 30 min at 20ºC to pellet the cEVs. The supernatant was discarded and the cEV-enriched pelleted was washed with citrate-phosphate buffered saline (PBS) solution (citrate-PBS; 1.4mmol/L phosphate, 154mmol/L NaCl, 10.9mmol/L trisodium citrate, pH 7.4) before an identical centrifugation was pursued. Finally, the remaining pellets were resuspended in a final volume of 100 µL citrate-PBS. Citrate-PBS was filtered through 0.22 μm pores before each analysis on a daily basis. The procedure for flow cytometric analysis of circulating EVs followed the MIFlowCyt-EV and MIFlowCyt checklists, as previously reported [12].
4.3. Flow Cytometric Analysis of Circulating Extracellular Vesicles
Three-label flow cytometric analysis was pursued as previously described [16,48,49]. In short, washed cEV suspensions were diluted in PBS containing 2.5mmol of CaCl2 (annexin binding buffer [ABB], BD Biosciences, CA, San Jose). Next, combinations of AV conjugated with CF-Blue (to detect PS) and two monoclonal antibodies (mAb) labelled with phycoerythrin (PE) or fluorescein isothiocyanate (FITC), were added and incubated for 20 min at 20ºC in the dark (Table S2). Samples were then diluted with ABB and immediately analysed on a FACSCantoII™ (BD, Franklin Lakes, NJ, USA) flow cytometer.
Each sample was acquired for 1 min at low flow rate. Forward scatter (FSC), side scatter (SSC), and fluorescence data were obtained with the settings in the logarithmic scale. Gate limits were established following previously described criteria [16,48,49]. Megamix-Plus FSC beads (BioCytex, Marseille, France) were used to set the upper threshold for FSC. These beads are a mix of beads with diameters of 0.1 µm, 0.3 µm, 0.5 µm and 0.9 µm, and according to their signal, the lower detection limit was set as a threshold above the electronic background noise of the flow cytometer for FSC and the second logarithm for SSC. EVs within the established gate limits (> 0.1 to 1 µm) were identified and quantified based on their reactivity to cell-specific mAb and binding to AV. To identify positive marked events, thresholds of fluorescence were also set based on samples incubated with the same final concentration of isotype-matched control mAb after titration experiments. AV binding level was corrected for auto-fluorescence using fluorescence signals obtained with EVs in a calcium-free buffer (PBS). Additional controls to correct for PE-, FITC- and CFBlue-fluorescence were also performed (unstained and single-stained controls), as well as serial dilutions to ensure proper event detection and swarming prevention. Finally, to confirm the presence of EVs in the cEVs suspension, 5% saponin-treated controls were investigated. Buffers were prepared daily and filtered through 0.22 µm pore-size filters under vacuum to reduce background noise.
BD FACSDiva™ Software (version 6.1.3, Becton Dickinson, Franklin Lakes, NJ, USA) was used to analyse data. Nieuwland’s formula [50] was used to calculate cEV concentration (number of EVs per µL of platelet-free plasma). Considering sample’s volume, flow cytometer’s flow rate and number of fluorescence-positive events (N), the formula is as follows: EVs/µL = N x (Vf/Va) x (Vt/FR) x (1/Vi), where Vf(µL)=final volume of washed EVs suspension, Va(µL)=volume of washed EVs suspension used for each labelling analysis, Vt(µL)=volume of EVs suspension before fluorescence-activated cell sorting analysis, FR(µL/min)=flow rate of the cytometer at low mode (the average volume of cEV suspension analysed in 1 min), 1 is the microliter unit of volume, and Vi(µL)=original volume of plasma used for cEVs isolation.
To calculate which cEVs did not have PS in their membrane, the number of AV+ events (PS+) was subtracted from the total number of fluorescence-positive gated events (which included PS+ and PS─ events), obtaining the number of PS─-EVs. Once obtained, this value was used to calculate EVs concentration by using the formula stated above [50].
4.4. Statistical Analysis
Statistical analyses were pursued using SPSS Statistical Analysis System (version 26.0, IBM Corp. Armonk, NY). Shapiro-Wilk test was utilised to assess variables’ normality. Descriptive analyses for quantitative variables were expressed using mean ± standard deviation or median [interquartile range 0.25-0.75], whereas for qualitative variables the number of cases and percentages were utilised. Chi-squared analysis was used to compare the frequencies of qualitative variables between groups. Non-parametric tests were used to contrast median values of quantitative variables and a P < 0.05 was considered statistically significant. Sample size was determined using the GRANMO sample size calculator (version 7.12, April 2012). To detect mean differences in the number of cEVs, a total of 126 individuals (healthy and diseased subjects) would be needed to complete the study (α risk=0.05, beta risk=0.2, two-sided test).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Distribution of leukocyte-derived circulating extracellular vesicles in a group of chronic heart failure patients and a reference non-heart failure group; Table S1: Clinical characteristics of the studied reference non-heart failure group; and Table S2: Cell surface molecules for extracellular microvesicle identification and characterization.
Author Contributions
Conceptualization, T.P., S. M., and L.B.; methodology, R.S., A.V-F., T.P., and L.B.; formal analysis, R.S., A.V-F., T.P., and L.B.; investigation, R.S., A.V-F., T.P., and L.B.; resources, T.P. and L.B.; data curation, R.S. and T.P.; writing— original draft preparation R.S., A.V-F., and L.B., writing—review, and editing, R.S., A.V-F., T.P, S. M. and L.B.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Society of Cardiology (SEC2016 Research Grant) to L.B.; Spanish Ministry of Economy and Competitiveness of Science and Agencia Estatal de Investigación (AEI/10.13039/501100011033-[PID2019-107160RB-I00]) to L.B.; Institute of Health Carlos III (ISCIII, FIS [PI19/01687] to T.P. and CIBERCV [CB16/11/00411] to L.B.); and Secretariat of University and Research of the Department of Business and Knowledge (Government of Catalonia [2017 SGR 1480]), cofounded by European Regional Development Fund (ERDF): "A way of making Europe"). We thank Fundación Jesus Serra and Fundación de Investigación Cardiovascular (Barcelona, Spain) for their continuous support.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Hospital de la Santa Creu i Sant Pau in Barcelona, Spain (protocol reference: 16/44).
Informed Consent Statement
A written informed consent to participate was obtained from all subjects involved in the study before their recruitment.
Data Availability Statement
Any data used for this manuscript can be obtained from the authors upon reasonable request.
Acknowledgments
We thank heart failure patients and non-heart failure subjects for their willingness to participate in this study, as well as physicians that helped in the recruitment.
Conflicts of Interest
L.B. declares to have acted as SAB member of Sanofi, Ionis and Novo Nordisk, to have a Research Grant of AstraZeneca, to have received speaker fees from Sanofi and to have founded the Spin-offs Glycardial Diagnostics SL and Ivastatin Therapeutics S (all unrelated to this work). T.P. declares to be co-founder of the Spin-offs Glycardial Diagnostics SL and Ivastatin Therapeutics S (all unrelated to this work). Remaining authors have no competing interests.
References
- Badimon, L.; Padro, T.; Arderiu, G.; Vilahur, G.; Borrell-Pages, M.; Suades, R. Extracellular vesicles in atherothrombosis: From biomarkers and precision medicine to therapeutic targets. Immunol Rev 2022, 312, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Suades, R.; Greco, M.F.; Padro, T.; Badimon, L. Extracellular Vesicles as Drivers of Immunoinflammation in Atherothrombosis. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Suades, R.; Padro, T.; Badimon, L. The Role of Blood-Borne Microparticles in Inflammation and Hemostasis. Semin Thromb Hemost 2015, 41, 590–606. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol 2019, 21, 9–17. [Google Scholar] [CrossRef]
- van Niel, G.; D'Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Badimon, L.; Suades, R.; Fuentes, E.; Palomo, I.; Padro, T. Role of Platelet-Derived Microvesicles As Crosstalk Mediators in Atherothrombosis and Future Pharmacology Targets: A Link between Inflammation, Atherosclerosis, and Thrombosis. Front Pharmacol 2016, 7, 293. [Google Scholar] [CrossRef]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: determination of their morphology, size, phenotype and concentration. J Thromb Haemost 2014, 12, 614–627. [Google Scholar] [CrossRef]
- Connor, D.E.; Exner, T.; Ma, D.D.; Joseph, J.E. The majority of circulating platelet-derived microparticles fail to bind annexin V, lack phospholipid-dependent procoagulant activity and demonstrate greater expression of glycoprotein Ib. Thromb Haemost 2010, 103, 1044–1052. [Google Scholar] [CrossRef]
- Matsumura, S.; Minamisawa, T.; Suga, K.; Kishita, H.; Akagi, T.; Ichiki, T.; Ichikawa, Y.; Shiba, K. Subtypes of tumour cell-derived small extracellular vesicles having differently externalized phosphatidylserine. J Extracell Vesicles 2019, 8, 1579541. [Google Scholar] [CrossRef]
- Perez-Pujol, S.; Marker, P.H.; Key, N.S. Platelet microparticles are heterogeneous and highly dependent on the activation mechanism: studies using a new digital flow cytometer. Cytometry A 2007, 71, 38–45. [Google Scholar] [CrossRef]
- Suades, R.; Padro, T.; Alonso, R.; Mata, P.; Badimon, L. Lipid-lowering therapy with statins reduces microparticle shedding from endothelium, platelets and inflammatory cells. Thromb Haemost 2013, 110, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Vilella-Figuerola, A.; Cordero, A.; Mirabet, S.; Munoz-Garcia, N.; Suades, R.; Padro, T.; Badimon, L. Platelet-Released Extracellular Vesicle Characteristics Differ in Chronic and in Acute Heart Disease. Thromb Haemost 2023. [Google Scholar] [CrossRef] [PubMed]
- Groenewegen, A.; Rutten, F.H. Sodium-glucose co-transporter 2 inhibitors and acute heart failure. Eur J Heart Fail 2020, 22, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.D.; Conte, J.V. The pathophysiology of heart failure. Cardiovasc Pathol 2012, 21, 365–371. [Google Scholar] [CrossRef]
- Tanai, E.; Frantz, S. Pathophysiology of Heart Failure. Compr Physiol 2015, 6, 187–214. [Google Scholar] [CrossRef]
- Vilella-Figuerola, A.; Padro, T.; Roig, E.; Mirabet, S.; Badimon, L. New factors in heart failure pathophysiology: Immunity cells release of extracellular vesicles. Front Cardiovasc Med 2022, 9, 939625. [Google Scholar] [CrossRef]
- Boulanger, C.M.; Loyer, X.; Rautou, P.E.; Amabile, N. Extracellular vesicles in coronary artery disease. Nat Rev Cardiol 2017, 14, 259–272. [Google Scholar] [CrossRef]
- Joop, K.; Berckmans, R.J.; Nieuwland, R.; Berkhout, J.; Romijn, F.P.; Hack, C.E.; Sturk, A. Microparticles from patients with multiple organ dysfunction syndrome and sepsis support coagulation through multiple mechanisms. Thromb Haemost 2001, 85, 810–820. [Google Scholar] [CrossRef]
- Mobarrez, F.; Vikerfors, A.; Gustafsson, J.T.; Gunnarsson, I.; Zickert, A.; Larsson, A.; Pisetsky, D.S.; Wallen, H.; Svenungsson, E. Microparticles in the blood of patients with systemic lupus erythematosus (SLE): phenotypic characterization and clinical associations. Sci Rep 2016, 6, 36025. [Google Scholar] [CrossRef]
- Nielsen, C.T.; Ostergaard, O.; Johnsen, C.; Jacobsen, S.; Heegaard, N.H. Distinct features of circulating microparticles and their relationship to clinical manifestations in systemic lupus erythematosus. Arthritis Rheum 2011, 63, 3067–3077. [Google Scholar] [CrossRef]
- Mobarrez, F.; Gunnarsson, I.; Svenungsson, E. Altered beta(2) -glycoprotein I expression on microparticles in the presence of antiphospholipid antibodies. J Thromb Haemost 2017, 15, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Vikerfors, A.; Mobarrez, F.; Bremme, K.; Holmstrom, M.; Agren, A.; Eelde, A.; Bruzelius, M.; Antovic, A.; Wallen, H.; Svenungsson, E. Studies of microparticles in patients with the antiphospholipid syndrome (APS). Lupus 2012, 21, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Bergen, K.; Mobarrez, F.; Jorneskog, G.; Wallen, H.; Tehrani, S. Phosphatidylserine expressing microvesicles in relation to microvascular complications in type 1 diabetes. Thromb Res 2018, 172, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, A.; Mobarrez, F.; Rooth, E.; Thalin, C.; von Arbin, M.; Henriksson, P.; Gigante, B.; Laska, A.C.; Wallen, H. Prognostic Value of Circulating Microvesicle Subpopulations in Ischemic Stroke and TIA. Transl Stroke Res 2020, 11, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Suades, R.; Padro, T.; Crespo, J.; Sionis, A.; Alonso, R.; Mata, P.; Badimon, L. Liquid Biopsy of Extracellular Microvesicles Predicts Future Major Ischemic Events in Genetically Characterized Familial Hypercholesterolemia Patients. Arterioscler Thromb Vasc Biol 2019, 39, 1172–1181. [Google Scholar] [CrossRef]
- Shet, A.S.; Aras, O.; Gupta, K.; Hass, M.J.; Rausch, D.J.; Saba, N.; Koopmeiners, L.; Key, N.S.; Hebbel, R.P. Sickle blood contains tissue factor-positive microparticles derived from endothelial cells and monocytes. Blood 2003, 102, 2678–2683. [Google Scholar] [CrossRef]
- Yang, C.; Ma, R.; Jiang, T.; Cao, M.; Zhao, L.; Bi, Y.; Kou, J.; Shi, J.; Zou, X. Contributions of phosphatidylserine-positive platelets and leukocytes and microparticles to hypercoagulable state in gastric cancer patients. Tumour Biol 2016, 37, 7881–7891. [Google Scholar] [CrossRef]
- Zhao, L.; Wu, X.; Si, Y.; Yao, Z.; Dong, Z.; Novakovic, V.A.; Guo, L.; Tong, D.; Chen, H.; Bi, Y.; et al. Increased blood cell phosphatidylserine exposure and circulating microparticles contribute to procoagulant activity after carotid artery stenting. J Neurosurg 2017, 127, 1041–1054. [Google Scholar] [CrossRef]
- Arauna, D.; Chiva-Blanch, G.; Padro, T.; Fuentes, E.; Palomo, I.; Badimon, L. Frail older adults show a distinct plasma microvesicle profile suggesting a prothrombotic and proinflammatory phenotype. J Cell Physiol 2021, 236, 2099–2108. [Google Scholar] [CrossRef]
- Berezin, A.E. Microparticles in Chronic Heart Failure. Adv Clin Chem 2017, 81, 1–41. [Google Scholar] [CrossRef]
- Kou, Y.; Zou, L.; Liu, R.; Zhao, X.; Wang, Y.; Zhang, C.; Dong, Z.; Kou, J.; Bi, Y.; Fu, L.; et al. Intravascular cells and circulating microparticles induce procoagulant activity via phosphatidylserine exposure in heart failure. J Thromb Thrombolysis 2019, 48, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Matan, D.; Mobarrez, F.; Lofstrom, U.; Corbascio, M.; Ekstrom, M.; Hage, C.; Lynga, P.; Persson, B.; Eriksson, M.; Linde, C.; et al. Extracellular vesicles in heart failure - A study in patients with heart failure with preserved ejection fraction or heart failure with reduced ejection fraction characteristics undergoing elective coronary artery bypass grafting. Front Cardiovasc Med 2022, 9, 952974. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, T.; Sugiyama, S.; Sugamura, K.; Ohba, K.; Matsuzawa, Y.; Konishi, M.; Matsubara, J.; Akiyama, E.; Sumida, H.; Matsui, K.; et al. Prognostic value of endothelial microparticles in patients with heart failure. Eur J Heart Fail 2010, 12, 1223–1228. [Google Scholar] [CrossRef]
- Jimenez, J.J.; Jy, W.; Mauro, L.M.; Horstman, L.L.; Ahn, Y.S. Elevated endothelial microparticles in thrombotic thrombocytopenic purpura: findings from brain and renal microvascular cell culture and patients with active disease. Br J Haematol 2001, 112, 81–90. [Google Scholar] [CrossRef]
- Zuchi, C.; Tritto, I.; Carluccio, E.; Mattei, C.; Cattadori, G.; Ambrosio, G. Role of endothelial dysfunction in heart failure. Heart Fail Rev 2020, 25, 21–30. [Google Scholar] [CrossRef]
- Brisson, A.R.; Tan, S.; Linares, R.; Gounou, C.; Arraud, N. Extracellular vesicles from activated platelets: a semiquantitative cryo-electron microscopy and immuno-gold labeling study. Platelets 2017, 28, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Biasucci, L.M.; La Rosa, G.; Pedicino, D.; D'Aiello, A.; Galli, M.; Liuzzo, G. Where Does Inflammation Fit? Curr Cardiol Rep 2017, 19, 84. [Google Scholar] [CrossRef]
- Shirazi, L.F.; Bissett, J.; Romeo, F.; Mehta, J.L. Role of Inflammation in Heart Failure. Curr Atheroscler Rep 2017, 19, 27. [Google Scholar] [CrossRef]
- Suades, R.; Padro, T.; Vilahur, G.; Martin-Yuste, V.; Sabate, M.; Sans-Rosello, J.; Sionis, A.; Badimon, L. Growing thrombi release increased levels of CD235a(+) microparticles and decreased levels of activated platelet-derived microparticles. Validation in ST-elevation myocardial infarction patients. J Thromb Haemost 2015, 13, 1776–1786. [Google Scholar] [CrossRef]
- Shlomovitz, I.; Speir, M.; Gerlic, M. Flipping the dogma - phosphatidylserine in non-apoptotic cell death. Cell Commun Signal 2019, 17, 139. [Google Scholar] [CrossRef]
- Dombroski, D.; Balasubramanian, K.; Schroit, A.J. Phosphatidylserine expression on cell surfaces promotes antibody-dependent aggregation and thrombosis in beta2-glycoprotein I-immune mice. J Autoimmun 2000, 14, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Takahashi, Y.; Ogata, K.; Kitamura, S.; Nakagawa, N.; Yamamoto, A.; Ishihama, Y.; Takakura, Y. Phosphatidylserine-deficient small extracellular vesicle is a major somatic cell-derived sEV subpopulation in blood. iScience 2021, 24, 102839. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bi, Y.; Yu, M.; Li, T.; Tong, D.; Yang, X.; Zhang, C.; Guo, L.; Wang, C.; Kou, Y.; et al. Phosphatidylserine-exposing blood cells and microparticles induce procoagulant activity in non-valvular atrial fibrillation. Int J Cardiol 2018, 258, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Sinauridze, E.I.; Kireev, D.A.; Popenko, N.Y.; Pichugin, A.V.; Panteleev, M.A.; Krymskaya, O.V.; Ataullakhanov, F.I. Platelet microparticle membranes have 50- to 100-fold higher specific procoagulant activity than activated platelets. Thromb Haemost 2007, 97, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Zara, M.; Guidetti, G.F.; Camera, M.; Canobbio, I.; Amadio, P.; Torti, M.; Tremoli, E.; Barbieri, S.S. Biology and Role of Extracellular Vesicles (EVs) in the Pathogenesis of Thrombosis. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- De Lorenzo, F.; Saba, N.; Kakkar, V.V. Blood coagulation in patients with chronic heart failure: evidence for hypercoagulable state and potential for pharmacological intervention. Drugs 2003, 63, 565–576. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Exosomal vesicles enhance immunosuppression in chronic inflammation: Impact in cellular senescence and the aging process. Cell Signal 2020, 75, 109771. [Google Scholar] [CrossRef]
- Suades, R.; Padro, T.; Alonso, R.; Lopez-Miranda, J.; Mata, P.; Badimon, L. Circulating CD45+/CD3+ lymphocyte-derived microparticles map lipid-rich atherosclerotic plaques in familial hypercholesterolaemia patients. Thromb Haemost 2014, 111, 111–121. [Google Scholar] [CrossRef]
- Suades, R.; Padro, T.; Alonso, R.; Mata, P.; Badimon, L. High levels of TSP1+/CD142+ platelet-derived microparticles characterise young patients with high cardiovascular risk and subclinical atherosclerosis. Thromb Haemost 2015, 114, 1310–1321. [Google Scholar] [CrossRef]
- Nieuwland, R.; Berckmans, R.J.; McGregor, S.; Boing, A.N.; Romijn, F.P.; Westendorp, R.G.; Hack, C.E.; Sturk, A. Cellular origin and procoagulant properties of microparticles in meningococcal sepsis. Blood 2000, 95, 930–935. [Google Scholar] [CrossRef]
Figure 1.
Distribution of circulating extracellular vesicles of endothelial origin in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) from (a) non- (CD309+) and (b) activated (CD62E+) endothelial cells in non-heart failure control subjects (n=21) and patients with chronic heart failure (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P < 0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular vesicles; eEVs, endothelial-derived EVs; HF, heart failure; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─, EVs that do not expose phosphatidylserine.
Figure 1.
Distribution of circulating extracellular vesicles of endothelial origin in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) from (a) non- (CD309+) and (b) activated (CD62E+) endothelial cells in non-heart failure control subjects (n=21) and patients with chronic heart failure (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P < 0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular vesicles; eEVs, endothelial-derived EVs; HF, heart failure; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─, EVs that do not expose phosphatidylserine.
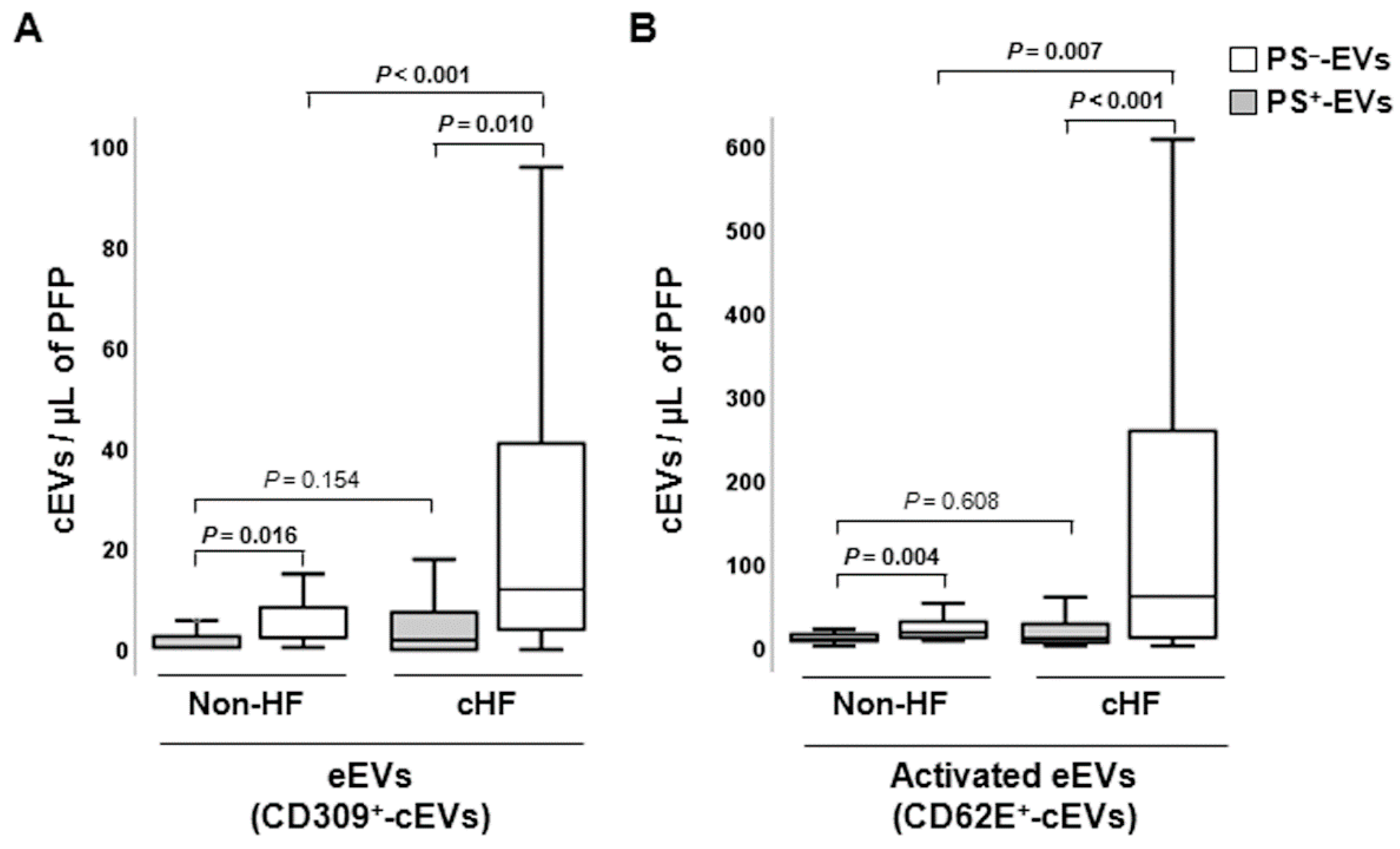
Figure 2.
Distribution of red blood cell and connexin 43-rich circulating extracellular vesicles in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) from (a) erythrocytes (CD235ab+) and (b) carrying connexin-43 (CX43+) in non-heart failure control subjects (n=21) and patients with chronic heart failure (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P < 0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular microvesicles; ErEVs, erythrocyte-derived EVs; HF, heart failure; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─-, EVs that do not expose phosphatidylserine.
Figure 2.
Distribution of red blood cell and connexin 43-rich circulating extracellular vesicles in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) from (a) erythrocytes (CD235ab+) and (b) carrying connexin-43 (CX43+) in non-heart failure control subjects (n=21) and patients with chronic heart failure (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P < 0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular microvesicles; ErEVs, erythrocyte-derived EVs; HF, heart failure; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─-, EVs that do not expose phosphatidylserine.
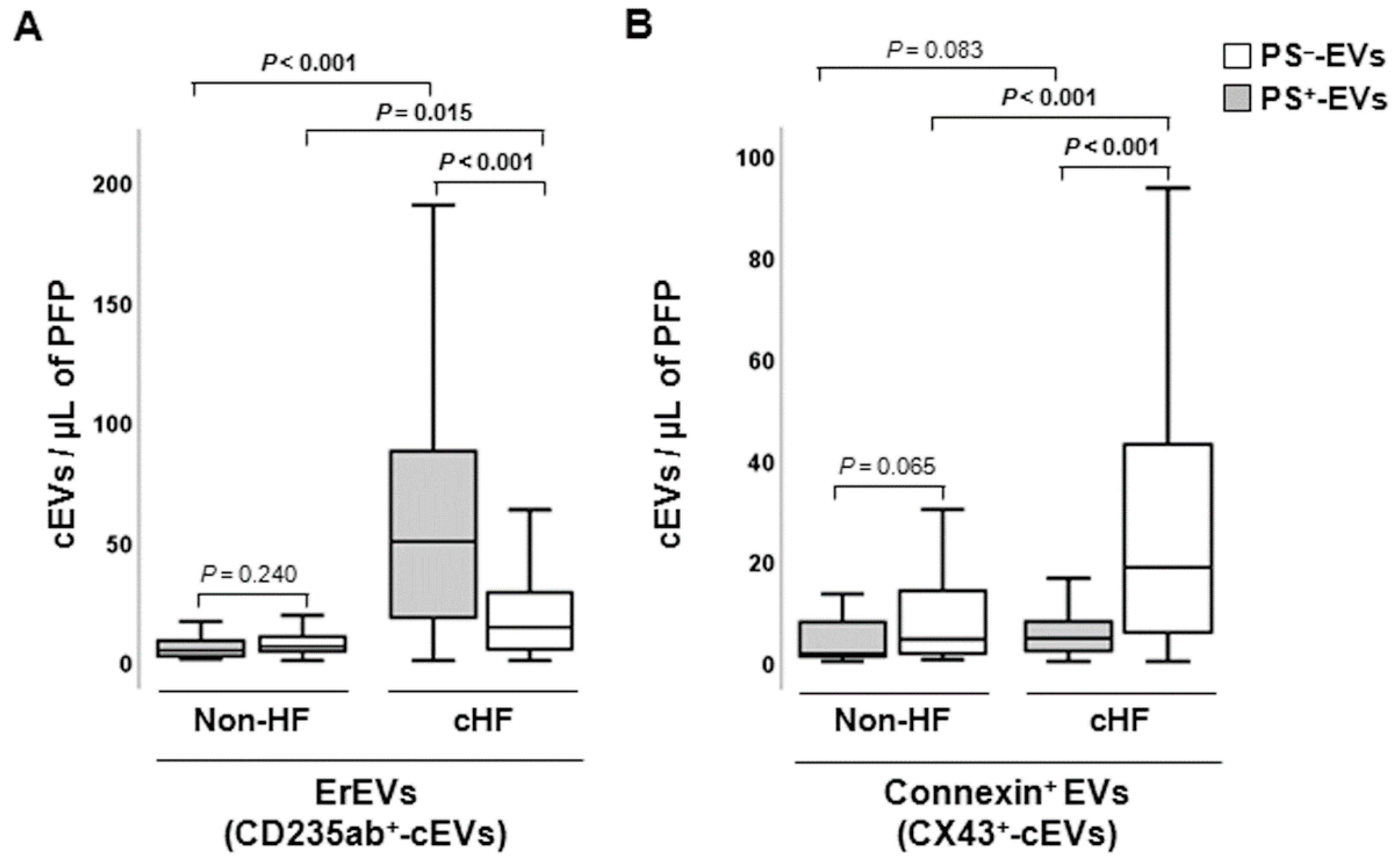
Figure 3.
Distribution of leukocyte-derived circulating extracellular vesicles in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) of (a) leukocyte-derived (CD45+), (b) activated leukocyte-derived (CD29+) and (c) monocyte-derived (CD14+) circulating extracellular vesicles in non-heart failure control subjects (n=21) and chronic heart failure patients (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P <0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular microvesicles; HF, heart failure; LEVs, leukocyte-derived EVs; mEVs, monocyte-derived EVs; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─-, EVs that do not expose phosphatidylserine.
Figure 3.
Distribution of leukocyte-derived circulating extracellular vesicles in reference non-heart failure and chronic heart failure groups. Box and whisker plots show numbers of cEVs per microliter of platelet-free plasma (cEVs/µL of PFP) of (a) leukocyte-derived (CD45+), (b) activated leukocyte-derived (CD29+) and (c) monocyte-derived (CD14+) circulating extracellular vesicles in non-heart failure control subjects (n=21) and chronic heart failure patients (n=119). Lines within boxes represent median values, the upper and lower boxes represent the 25th and 75th percentiles, respectively, and the upper and lower boxes outside the boxes represent the 10th and 90th percentiles, respectively. A P <0.05 was considered significant (U-Mann Whitney test). P-values in bold correspond to significant differences. CD indicates cluster of differentiation; cHF, chronic heart failure; cEVs, circulating extracellular microvesicles; HF, heart failure; LEVs, leukocyte-derived EVs; mEVs, monocyte-derived EVs; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─-, EVs that do not expose phosphatidylserine.
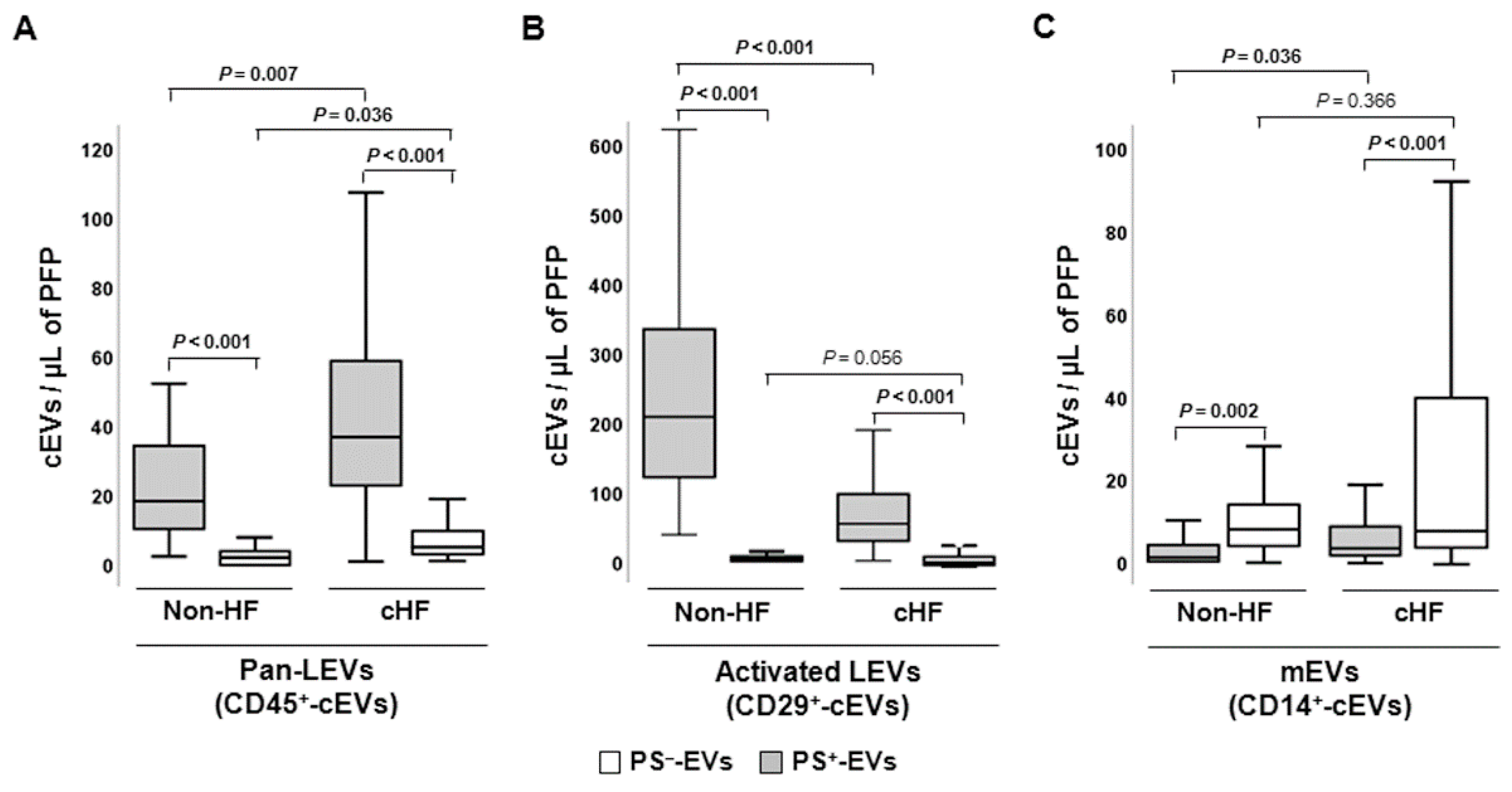
Figure 4.
Schematic summary showing main study results of the circulating extracellular vesicle characterization focusing on phosphatidylserine membrane exposure in chronic heart failure. CD indicates cluster of differentiation; cEVs, circulating extracellular vesicles; cHF, chronic heart failure; CX43, connexin 43; eEVs, endothelial cell-derived EVs; ErEVs, erythrocyte-derived EVs; EVs, extracellular microvesicles; ℓEVs, lymphocyte-derived EVs; LEVs, platelet-derived EVs; nEVs, neutrophil-derived EVs; NKcEVs, NK cell-derived EVs; mEVs, monocyte-derived EVs; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─, EVs that do not expose phosphatidylserine.
Figure 4.
Schematic summary showing main study results of the circulating extracellular vesicle characterization focusing on phosphatidylserine membrane exposure in chronic heart failure. CD indicates cluster of differentiation; cEVs, circulating extracellular vesicles; cHF, chronic heart failure; CX43, connexin 43; eEVs, endothelial cell-derived EVs; ErEVs, erythrocyte-derived EVs; EVs, extracellular microvesicles; ℓEVs, lymphocyte-derived EVs; LEVs, platelet-derived EVs; nEVs, neutrophil-derived EVs; NKcEVs, NK cell-derived EVs; mEVs, monocyte-derived EVs; PFP, platelet-free plasma; PS+, EVs exposing phosphatidylserine; and PS─, EVs that do not expose phosphatidylserine.
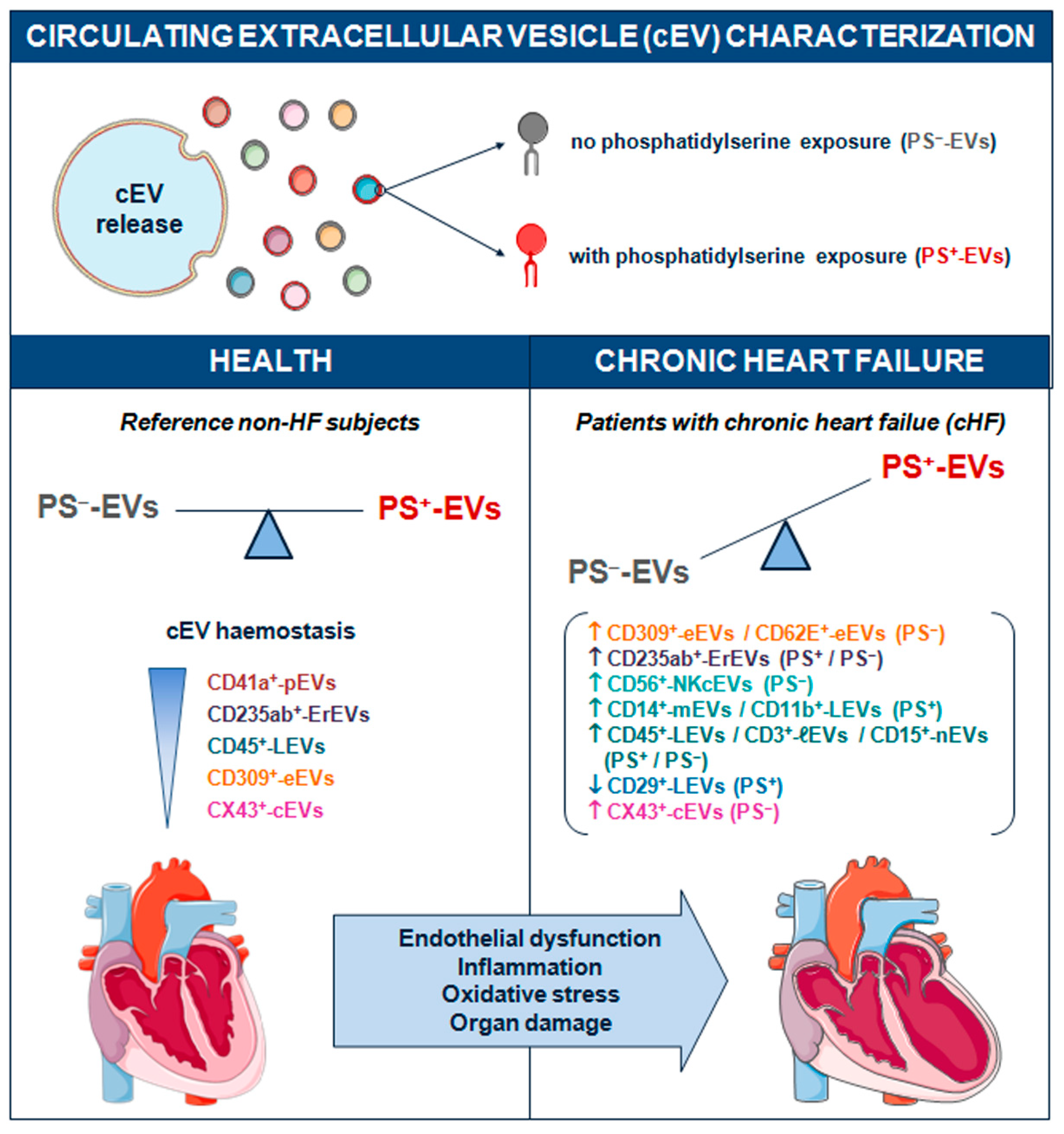

Table 1.
Absolute numbers and relative amounts of annexin V-positive and negative circulating extracellular vesicles in reference non-heart failure subjects and chronic heart failure patients.
Table 1.
Absolute numbers and relative amounts of annexin V-positive and negative circulating extracellular vesicles in reference non-heart failure subjects and chronic heart failure patients.
| cEVs | cHF (n = 119) | Non-HF (n = 21) | P-value | |
|---|---|---|---|---|
| AV+-EVs + AV─-EVs | 1079.12 [425.93 - 4702.2] | 803.72 [300.6 - 1104.67] | 0.135 | |
| AV+-EVs | ||||
| Total | n | 159.6 [110.8 - 252.37] | 309.2 [121.14 - 453.31] | 0.022 |
| % | 15.76 [3.76 - 32.47] | 51.48 [28.18 - 78.19] | <0.001 | |
| CD309+ | n | 1.9 [0-7.52] | 0 [0 - 2.90] | 0.154 |
| % | 0.61 [0 - 2.16] | 0 [0 - 0.45] | 0.094 | |
| CD41a+ | n | 124 [76.1 - 180.5] | 406 [110 - 646] | 0.001 |
| % | 53.4 [44.03 - 62.52] | 85.61 [77.11 - 96.76] | <0.001 | |
| CD45+ | n | 36 [21.45 - 58] | 18 [9 - 38] | 0.007 |
| % | 16.49 [11.37 - 24.59] | 9.01 [2.24 - 18.77] | 0.016 | |
| CD235ab+ | n | 49.7 [17.5 - 88.35] | 4.3 [1.75 - 8.98] | <0.001 |
| % | 24.17 [13.91 - 33.10] | 1.33 [0.54 - 2.49] | <0.001 | |
| CX43+ | n | 4.6 [2 - 7.98] | 1.6 [0.8 - 8.0] | 0.083 |
| % | 1.88 [1.05 - 2.98] | 0.56 [0.17 - 1.65] | 0.001 | |
| AV─-EVs | ||||
| Total | n | 787.8 [262.2 - 4576.7] | 253.86 [99.57 - 703.16] | 0.007 |
| % | 84.25 [67.53 - 96.24] | 48.52 [21.80 - 71.82] | <0.001 | |
| CD309+ | n | 12 [4 - 43.32] | 2 [1 - 8.44] | <0.001 |
| % | 23.08 [10.4 - 41.24] | 12.82 [2.03 - 19.05] | 0.012 | |
| CD41a+ | n | 4.16 [2.0 - 9.5] | 5 [0.5 - 14.17] | 0.773 |
| % | 6.76 [1.15 - 13.89] | 20.41 [0 - 38.9] | 0.032 | |
| CD45+ | n | 4 [1.92 - 9.12] | 2.2 [0 - 4.0] | 0.036 |
| % | 5.81 [1.74 - 12.51] | 6.01 [0 - 14.28] | 0.586 | |
| CD235ab+ | n | 14 [4.63 - 28.5] | 6.0 [4.0 - 11.0] | 0.015 |
| % | 19.56 [9.1 - 36.49] | 27.4 [11.11 - 47.62] | 0.245 | |
| CX43+ | n | 18.60 [5.4 - 43.2] | 4.4 [1.6 - 14.0] | 0.001 |
| % | 27.2 [15.23 - 42.81] | 18.06 [9.09 - 30.43] | 0.028 | |
Data are expressed as median [interquartile range] of absolute values and relative expression. AV+/- indicates annexin V+/-; CD, cluster of differentiation; cEVs, circulating extracellular vesicles; cHF, chronic heart failure; EVs, extracellular vesicles; and HF, heart failure.
Table 2.
Absolute numbers of distinct subsets of platelet-derived annexin V-positive and negative circulating extracellular vesicles in reference non-heart failure subjects and chronic heart failure patients.
Table 2.
Absolute numbers of distinct subsets of platelet-derived annexin V-positive and negative circulating extracellular vesicles in reference non-heart failure subjects and chronic heart failure patients.
| pEVs | cHF patients | Non-HF subjects | P-value |
|---|---|---|---|
| AV+-pEVs | |||
| CD31+ | 28.5 [16 - 58.9] | 166 [53 - 283.78] | < 0.001 |
| CD41a+ | 124 [80 - 178.6] | 406 [126 - 642] | 0.001 |
| CD31+/CD41a+ | 28.25 [12 - 58.9] | 122 [40 - 220] | 0.001 |
| CD62P+ | 30 [14 - 51.3] | 58 [31 - 92] | 0.010 |
| AV─-pEVs | |||
| CD31+ | 17.1 [6 - 44] | 4 [2 - 10] | 0.002 |
| CD41a+ | 4.16 [2 - 9.5] | 5 [1 - 12.78] | 0.773 |
| CD31+/CD41a+ | 0 [0 - 0] | 0 [0 - 0] | 0.123 |
| CD62P+ | 2 [0 - 6] | 0 [0 - 4] | 0.085 |
Data are expressed as median [interquartile range] of absolute values. AV+/- indicates annexin V+/-; CD, cluster of differentiation; cHF, chronic heart failure; HF, heart failure; and pEVs, platelet-derived extracellular vesicles.
Table 3.
Clinical characteristics of the studied chronic heart failure patient population.
| Parameters | Patients with cHF (n=119) |
|---|---|
| Demographic characteristics | |
| Male, n (%) | 81 (68) |
| Female, n (%) | 38 (32) |
| Age, years | 67 ± 11.8 |
| Clinical data | |
| Systolic blood pressure, mmHg | 120.4 ± 19.1 |
| Diastolic blood pressure, mmHg | 73.9 ± 11.1 |
| Left ventricular ejection fraction, % | 45.59 ± 18.98 |
| Comorbidities | |
| Smokers, n (%) | 13 (10.9) |
| Hypertension, n (%) | 82 (68.9) |
| Pulmonary hypertension, n (%) | 49 (41.1) |
| Dyslipidaemia, n (%) | 64 (53.7) |
| Chronic kidney disease, n (%) | 46 (38.6) |
| Diabetes mellitus, n (%) | 53 (44.5) |
| Atrial fibrillation, n (%) | 50 (42) |
| Background medication | |
| Angiotensin-converting-enzyme inhibitors, n (%) | 48 (40.3) |
| Angiotensin II receptor blockers, n (%) | 35 (29.4) |
| Angiotensin receptor neprilysin inhibitors, n (%) | 17 (14.2) |
| Beta-blockers, n (%) | 100 (84) |
| Aldosterone antagonists, n (%) | 66 (55.4) |
| Diureticsa, n (%) | 103 (86.5) |
| Ivabradine, n (%) | 14 (11.7) |
| Statins, n (%) | 77 (64.7) |
| Insulin, n (%) | 16 (13.4) |
| Anti-diabetic drugs, n (%) | 40 (33.6) |
| Anticoagulants, n (%) | 61 (51.2) |
| Antiplatelet agents, n (%) | 46 (38.6) |
| Anti-arrhythmic drugs, n (%) | 26 (21.8) |
Data are expressed either by mean ± standard deviation or number of cases (percentage). a Diuretics include furosemide, hydrochlorothiazide, torasemide, and indapamide. cHF indicates chronic heart failure.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



