
Submitted:
29 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Bacterial biofilms can cause widespread infection. In addition to causing urinary tract infections and pulmonary infections in patients with cystic fibrosis, biofilms can help microorganisms adhere to the surfaces of various medical devices, causing biofilm-associated infections on the surfaces of biomaterials such as venous ducts, joint prostheses, mechanical heart valves, and catheters. Biofilms provide a protective barrier for bacteria and provide resistance to antimicrobial agents, which increases the morbidity and mortality of patients. This review introduces the formation process and drug resistance mechanism of biofilms in detail, and further summarizes the main characteristics of clinical persistent infection caused by biofilms and the many methods of treating biomaterial-related biofilm. This provides ideas and directions for the development of new biofilm infection strategies related to therapeutic materials.
Keywords:
Biofilm formation
; Biomaterials
; Antibiotic resistance mechanisms
; Biofilm infection
; Antibiofilm strategies
1. Introduction
A biofilm is a population structure comprising a highly structured membrane complex formed by bacteria attached to various solid surfaces and their production of extracellular polysaccharides (EPSs), matrix proteins, and extracellular DNA (eDNA) [1]. The existence of biofilms provides many biological advantages for its internal bacteria, such as high infectivity, drug resistance, and strong viability [2]. It is estimated that more than 65% of nosocomial infections, about 80% of chronic infections, and 60% of human bacterial infections are caused by biofilms [3], including chronic biofilm diseases such as cystic fibrosis and periodontitis, as well as biofilm infections related to biomedical materials, such as medical device infections, implant-related infections, and others [4].
It is worth noting that once a biofilm is formed on medical equipment, conventional methods often cannot effectively eradicate the biofilm and usually require long-term antibacterial treatment [5]. As current antimicrobial therapy is expensive and will increase the drug resistance of pathogens, there is interest in the development of new antimicrobial therapies [6]. In this review, we introduced in detail the formation process and drug resistance mechanism of biomaterial-related biofilms and summarized their characteristics in clinical infections, such as persistent infections and inflammation. Two methods to prevent and treat biofilm infection related to biomaterials, including antibacterial coatings against biofilm infection and surface modifications of biomaterials, are also discussed in order to provide ideas for the development of new anti-biofilm therapies.
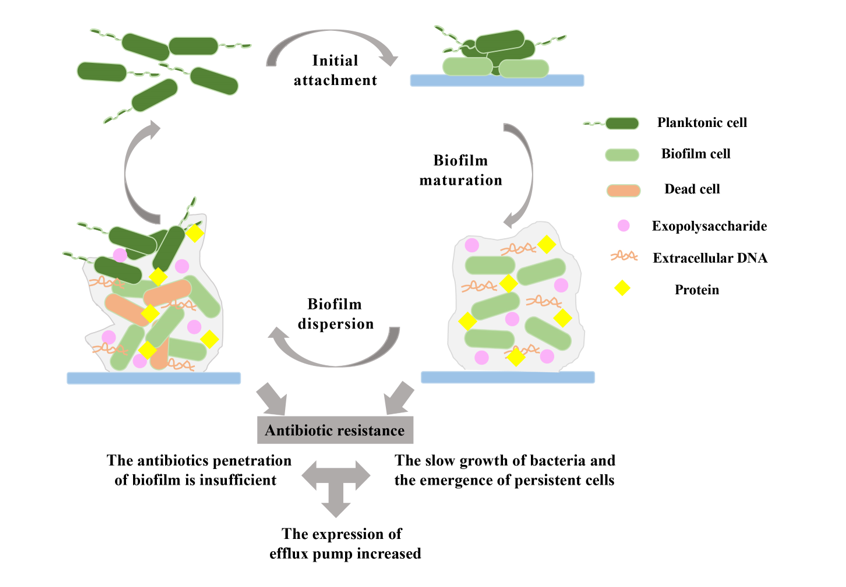
2. Biofilm formation on biomaterials
The formation of biomaterial-related biofilms mainly depends on the surface of medical devices and is a continuous, complex and multi-stage process that depends on many factors, such as the matrix, material surface medium, cell metabolism and internal characteristics, and signal molecules [7,8]. The biofilm formation includes three main steps: attachment, maturation, and dispersion [9,10] (Figure 1). In the first stage, microorganisms attach to the biomaterial surface, but the adhesion process is reversible because of the poor adhesion between the cells and the surface. Then, as the cells interact on the biomaterial surface of the material, the reversible attachment of bacteria cell becomes irreversible. Bacteria form microcolonies when their adhesion to the biomaterial surface is stable. However, when the external conditions change, such as temperature, nutrition, pH, and oxygen, the biofilm will disperse and the bacteria will return to the planktonic state [11,12].
2.1. Biofilm initial attachment
The initial attachment of a biofilm, as the name implies, refers to the attachment process of microorganisms on biomaterial surfaces [13]. However, due to the weak interaction between bacteria and the biomaterial surface, the initial adhesion stage of bacteria is not absolute, but instead a reversible process [8,14]. For example, when bacteria attach to a biomaterial surface, they are affected by the external environment, such as hydrodynamics and attraction or repulsion; or, if the composition, temperature, pressure, or pH of the biomaterial surface become unsuitable for bacteria to adhere, the bacteria will return to the planktonic state [8,15].
In this process, physical interactions such as polarity, London-van der Waals forces, electrostatic interactions, hydrophobic interactions, and protein adhesion can promote the adhesion of microorganisms to the biomaterial surface and contribute to the formation of biofilms [8,16,17]. A flagellum is a flagellate filamentous appendage that provides power for bacteria. It can attach to the surface of biomaterials to enhance the interaction between the bacteria and the attached surface and can also reduce the repulsive force between the cells and the surface, thus promoting cell attachment to the surface [18,19]. Interestingly, the reversible adhesion of bacteria can be transformed into irreversible adhesion under certain conditions. First, bacterial cells are usually negatively charged, and when the attraction between the microorganism and the implanted surface is greater than the repulsive force, the bacterial cells remain fixed and strongly adhere to the positively charged surface to form irreversible adhesion [16]. Second, bacterial adhesin plays an important role in the adhesion of microorganisms to the biomaterial surface. Fimbriae adhesin allows bacteria to adhere to each other and form an initial cell adhesion layer. For example, type 1 fimbriae is one of the most common adhesion organelles in Enterobacteriaceae, including Salmonella [20]. It can increase the initial surface adhesion of Escherichia coli and form irreversible adhesion [21]. In addition, in some bacteria, polysaccharide adhesin can regulate permanent cell adhesion, such as polysaccharide intercellular adhesin (PIA) that promotes intercellular adhesion and biofilm accumulation of Staphylococcus epidermidis [22,23]. Finally, EPSs can also promote the irreversible attachment of bacteria by interacting with surface materials and ligands [15].
2.2. Biofilm maturation
After the microorganisms adhere to the biomaterial surface and stabilize, the bacterial cells will continue to proliferate and produce intercellular adhesions to form microcolonies [1]. At this stage, as the bacterial density reaches a threshold level, the bacterial quorum sensing (QS) system is activated [16,24]. QS is a bacterial cell-to-cell communication system that relies on diffusible signal molecules. Bacteria use quorum-sensing signaling molecules to regulate their own behaviors, such as biofilm maturation, motility, and virulence factor expression [25,26]. Different types of bacteria secrete different signal molecules, such as acylated homoserine lactone (AHL) secreted by gram-negative bacteria, self-inducer peptide (AIP) secreted by gram-positive bacteria, and self-inducer-2 (AI-2) secreted by gram-negative and gram-positive bacteria [7,27]. Using Pseudomonas aeruginosa as an example, many studies have shown that Pseudomonas quinolone signal (PQS) molecules affect biofilm formation [28]. The deletion of the biosynthetic gene of a mutant PQS caused P. aeruginosa to form a defective biofilm, and it could not form the mushroom-like structure of the mature biofilm of the wild bacteria [28]. Extracellular DNA (eDNA) is essential for intercellular junction and biofilm stability [16]. The PQS also promotes the release of eDNA, which is beneficial to the maturation of P. aeruginosa biofilms [29]. In addition, QS not only regulates the entire process of biofilm formation but also promotes bacteria to secrete EPSs [30]. The secretion of EPSs helps to stabilize the biofilm structure and prevent itself from being attacked by antibacterial agents and immune cells [13]. The secretion of EPSs also contributes to the formation of three-dimensional structures of biofilm [31]. Voids and channels are formed in the three-dimensional structure of the biofilm, which are filled with water to distribute important nutrients, transmit molecular signals, remove waste from the biofilm, and act as a circulatory system [32,33]. After the formation of the first biofilm structure mediated by EPS, other bacteria of the same species and other species in the environment are also incorporated into the biofilm structure. In this manner, the biofilm structure develops from a thin layer to a mushroom-like structure [11].
2.3. Biofilm dispersion
As the biofilm matures, nutrient resources are consumed in large quantities, and toxic substances will continue to accumulate [34,35]. Therefore, to obtain more nutrients, the bacterial cells of the biofilm disperse from the biomaterial surface and migrate to other areas of the medical implant, thereby spreading the infection [8,16]. The dispersion mechanism in bacteria generally occurs in three stages: first, cells leave the microcolony; second, cells move to a new substrate; and third, cells attach to the new substrate and start a new biofilm formation process [11,36]. In fact, biofilm dispersion can be divided into active and passive modes [37]. When bacteria experience environmental stresses such as antimicrobial pressure and nutrient deficiency, they will actively dissociate from the implanted surface [38]. In the process of biofilm dispersion, some mechanisms are favorable for bacterial cell dispersion, such as the dissolution of EPSs [8]. Nutrient deprivation in biofilms stimulates small molecules of the fatty acid DSF (cis-11-methyl-2-dodecenoic acid) to trigger autophosphorylation and induce the degradation of c-di-GMP, leading to the dissolution of EPSs, which releases some of the planktonic cells [39,40]. In addition, in the process of active dispersion, the expression of genes related to bacterial motility caused by flagellum synthesis is upregulated, while the expression of genes related to bacterial attachment is downregulated, thus facilitating bacterial detachment [35].
Passive dispersion of biofilm depends on external factors, such as enzyme degradation of the biofilm matrix and shear forces [37]. The microbial community within the biofilm produces different saccharolytic enzymes that digest the polysaccharides that stabilize the biofilm structure, helping to release the microbial surfaces to new colonization areas [16,31]. For example, in alginate, Pel and Psl polysaccharides are the skeleton components of the P. aeruginosa biofilm supporting structure, which can help bacteria to absorb nutrients and communicate signals between cells [41]. Alginate lyases have been shown to be used for the cleavage and subsequent separation of EPS substrates, while the extracellular polysaccharide hydrolases PelA and PslG induce biofilm dispersion and prevent biofilm formation [16,42,43]. PelA was more effective in dispersing biofilms with Pel as the main extracellular polysaccharide, while PslG was most effective in removing biofilms formed with Psl as the main extracellular polysaccharide [44]. Previously, our research group also overexpressed PelA and PslG enzymes in P. aeruginosa engineered bacteria through synthetic biology, and lysed the engineered bacteria in active and passive ways to release two types of extracellular polysaccharide hydrolases, PelA and PslG, thus destroying cytoskeletal components Psl and Pel. The result was destruction of the P. aeruginosa biofilm [45]. In addition to enzymatic degradation of biofilm matrixes, physical means such as shear force are important factors for passive dispersion of biofilms [46]. A sudden increase in shear forces causes the cells to be immediately released from the biofilm, resulting in the dispersion of the biofilm [47]. For example, during the formation of Streptococcus mutans biofilms, a 10-fold increase in shear forces resulted in an 85% reduction in the biomass of the biofilm [48]. In addition, ultrasonic waves, laser-induced shock waves, and other technologies have been found to passively disperse biofilms [37]. Ultrasonic treatment significantly reduced E. coli and S. aureus biofilms on a stainless steel surface [49,50]. Laser-induced shock waves destroyed biofilms grown on different types of medical devices by generating plasma, and although the shock waves themselves were not harmful to the bacteria, the biofilms exposed to the shock waves were more sensitive to antibiotics than those that were not exposed [51]. To summarize, both active and passive methods can facilitate the dispersion of biofilm. However, once biofilm cells that were dispersed and settled on a new surface are stimulated by adverse conditions, the isolated microbial cells upregulate the expression of flagellar proteins to help them move quickly and return to a floating state on the surface. The bacteria then reform the biofilm in the right environment, causing continuous infection [8,52].
3. Mechanisms of antibiotic-resistant biofilms
As one of the most successful life forms of bacteria, biofilms are not only widely distributed in various environments, but they also play an important role in promoting antibiotic resistance [53]. The sensitivity test of a biofilm model in vitro showed that the survival rate of bacteria covered by a biofilm was hundreds or even thousands of times higher than that of planktonic bacteria when using antibiotics for sterilization [54,55]. Many studies have shown that in biofilms, antibiotics do not exert their antibacterial function, but instead cause the emergence of superbugs, which increases the difficulty of treating biofilm infections [56,57]. In addition, the formation of a biofilm is conducive to the emergence of inflammation, which leads to persistent infection. For example, biofilm formation plays an important role in pulmonary infection, burn infection, and medical device infection in patients with cystic fibrosis caused by P. aeruginosa [58]. Therefore, revealing the main causes of drug resistance and the mechanism of drug resistance of biofilm bacteria is of great significance for the clinical treatment of biofilm infection [59].
3.1. Prevention of antibiotic penetration by biofilms
EPSs not only maintain the structural stability of a biofilm but also serve as a barrier for antimicrobial agents such as bleaching agents and antibiotics [60]. Aminoglycoside antibiotics cannot penetrate the biofilm because the positive charge of the aminoglycoside antibiotics is neutralized by the negatively charged matrix [61]. For example, cations are chelated by a subinhibitory level of negatively charged eDNA, which limits the penetration of antibiotics by inducing the PA3552–PA3559 operon to develop resistance to cationic antimicrobial peptides and aminoglycosides [59,62]. Furthermore, the presence of enzymes in the biofilm matrix also prevent the penetration of antibiotics [63]. For example, β-lactamases accumulated in extracellular polysaccharides promote the degradation and inactivation of antibiotics, and the role of β-lactamases is mainly against β-lactam antibiotics (carbapenem, imipenem), for which drug resistance is particularly important [64,65]. It is also believed that the poor permeability of antimicrobial agents is due to the decrease in diffusion and adsorption in the biofilm matrix [6], and there is no strong interaction between most antibiotics and biofilm matrix components [66,67].
3.2. Presence of slow-growing or persistent cells in biofilms
The formation of the biofilm matrix leads to slower penetration of antimicrobial agents that develops a resistant phenotype called persistent cells, resulting in antibiotic resistance [68,69]. An experiment of a biofilm in vitro showed that the oxygen concentration on the surface of a biofilm is high, and the metabolism of surface bacteria is also high, while the oxygen concentration in the center of the biofilm can be very low, and the bacteria growth relatively slow or halted [64,70]. These slow-growing bacteria contribute to the antibiotic resistance of the biofilm [69]. For example, fluoroquinolones and tetracycline can only effectively kill metabolically active cells in the upper layer of a P. aeruginosa biofilm [71]. Persistent cells are thought to be cells that differentiate into a dormant state, and they contribute to the persistence of biofilms [63]. Because the cell metabolism of persistent cells slows or ceases, the target of the antibiotic is not active, and the antibiotic cannot work [68].
3.3. Increased expression of efflux pumps in biofilms
Increasingly more studies have shown that the presence of efflux pumps is a key factor in the development of antibiotic resistance in biofilms [72,73]. Efflux pumps, which are membrane proteins necessary to maintain bacterial homeostasis by expelling toxic substances, are present in all types of bacteria [74]. Bacteria present in the biofilms also use efflux pumps to maintain cytoplasmic concentrations of certain antibacterial compounds below critical thresholds or to expel antibiotics, and the antibiotic resistance of the biofilm is caused by the increased expression or activity of the efflux pumps [75,76]. The ATP binding box (ABC) family, the small multi-drug resistant (SMR) family, the multi-drug and toxic compound extrusion (MATE) family, the drug-resistant nodular cell division (RND) family, and the large promoter superfamily (MFS) are the five major families of efflux pumps, which play an important role in biofilm antibiotic resistance [77,78]. For example, in Acinetobacter baumannii, TetA and TetB efflux pumps of the MFS family cause resistance of its biofilm to tetracycline and minocycline, in which the Tet efflux pump uses proton exchange as an energy source to extrude tetracycline [79]. The physiological function of these pumps is to expel toxic molecules that denature cell membranes and confer resistance to bacteria that are present in the biofilms [80]. MacB is a member of ABC family, and it helps bacteria excrete macrolides and endows biofilms with antibiotic resistance [81]. In addition, P. aeruginosa MexAB-OprM and MexCD-OprJ, which belong to the RND family, play important roles in antibiotic resistance in biofilms, and MexAB-OprM pumps contribute to biofilm resistance to ofloxacin [82,83]. S. aureus responds to antibiotics by upregulating various ABC transporters, thereby increasing antibiotic resistance in biofilms. For example, GraRS may confer vancomycin resistance to a biofilm by upregulating the VraFGABC transporter [84,85]. BraRS can also activate BraDE and VraDEABC transporters and increase biofilm resistance to nisin and bacitracin [86,87].
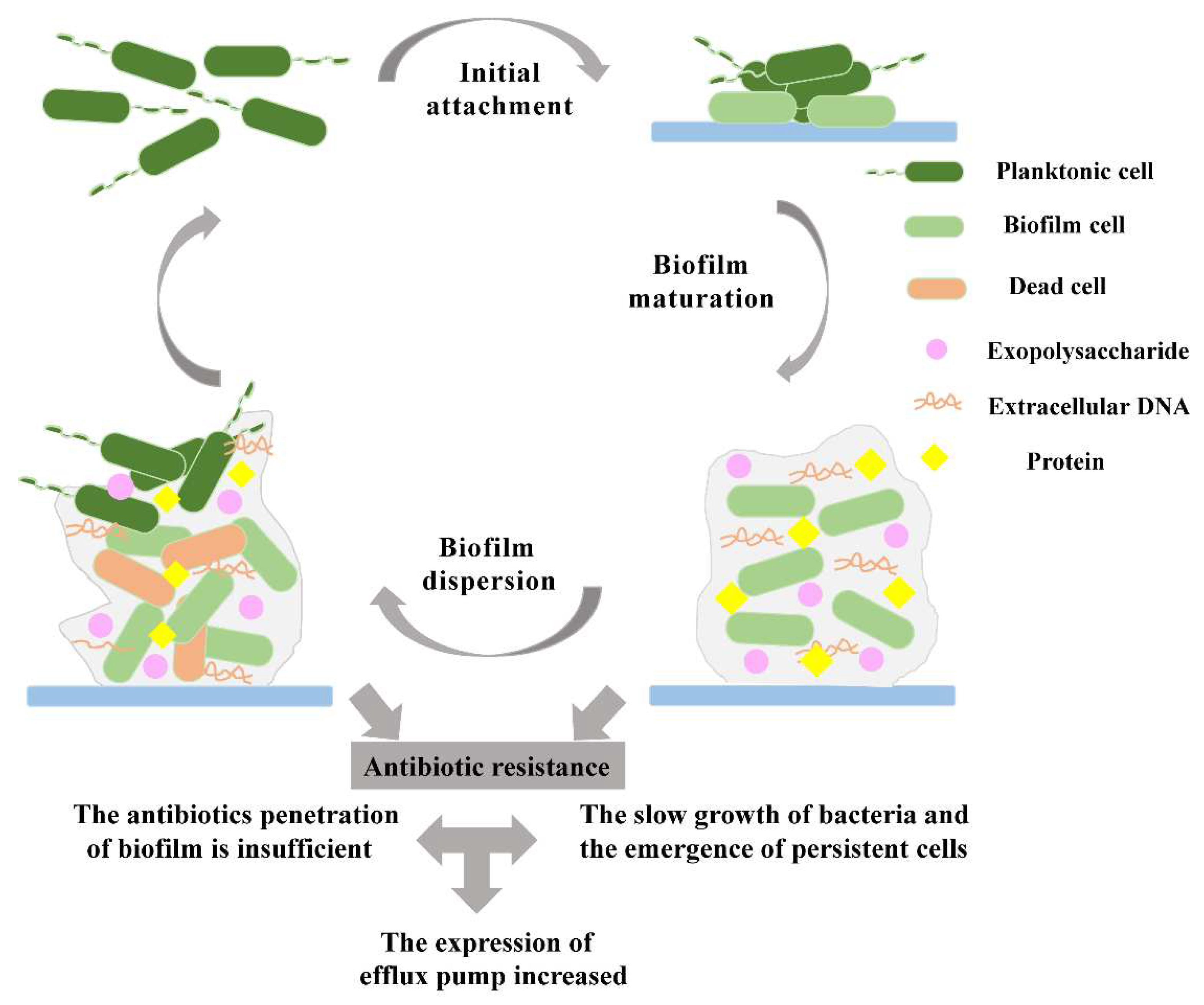
Figure 1.
The process of biofilm formation and antibiotic resistance mechanisms. Biofilm formation is divided into three steps. (1) Initial attachment of the biofilm. (2) Biofilm maturation. (3) Biofilm dispersion. The mechanisms of antibiotic resistance in biofilms are also divided into three factors. (1) The antibiotic penetration of the biofilm is insufficient. (2) The bacteria grow slowly and persistent cells emerge in biofilms (3) The expression of efflux pumps increases.
Figure 1.
The process of biofilm formation and antibiotic resistance mechanisms. Biofilm formation is divided into three steps. (1) Initial attachment of the biofilm. (2) Biofilm maturation. (3) Biofilm dispersion. The mechanisms of antibiotic resistance in biofilms are also divided into three factors. (1) The antibiotic penetration of the biofilm is insufficient. (2) The bacteria grow slowly and persistent cells emerge in biofilms (3) The expression of efflux pumps increases.
4. Clinical characteristics of biomaterial-associated biofilms
Colonization of biomaterials by pathogens leads to the formation of biofilms, which greatly increases the potential for clinical infection [88]. This includes devices such as mechanical heart valves, prosthetic joints, endotracheal tubes, and contact lens with biofilm-related infection [89]. Once biofilms are formed on medical devices, eradicating microbes becomes extremely difficult and can be costly due to the lengthy hospital stays, surgery, and long-term antibacterial treatments that are often required [90]. The development of these biomaterials has saved millions of lives, but at the same time, the resulting biofilm infections are endangering people's health. The above-mentioned associated biofilm infections caused by biomaterials share the same clinical features [91,92]:
(1) Persistent infection (more than 7 days). For example, central venous catheter-associated biofilms and almost all indwelling central venous catheters are colonized by microorganisms that can produce biofilms. Common bacteria isolated from catheter biofilms are coagulase-negative Staphylococcus [93]. Colonization and biofilm formation on catheter surfaces by these microorganisms can occur within 24 hours of insertion and are pervasive and persistent [94]. In one study, short-term (<10 days) catheters had greater biofilm formation on the external surface; long-term catheters (>30 days) had more biofilm formation on the catheter inner lumen [31].
(2) Inflammation. An example is biofilm infection associated with mechanical heart valves. Infectious endocarditis (IE) is a disease with substantial morbidity and mortality today due to the increasing use of implantable devices, such as prosthetic heart valves [95]. Microbes initially adhere to the prosthetic valve surface via fibronectin and polysaccharides and grow in a platelet-fibrin matrix to form an intact biofilm [96]. Because biofilms formed on artificial surfaces promote inflammatory responses and hypercoagulability, the presence of foreign tissue needs to be maintained through inflammatory and thrombotic processes [97]. In addition, prosthetic joint infection (PJI), a serious complication after joint replacement, causes inflammation [98]. According to the symptoms that appear after implantation, PJIs can be divided into three categories, early infection, delayed infection, and late infection [99]. Both early and delayed infections are often the result of surgical contamination and are considered the most common cause of biomaterial-related biofilm infections. These infections are often associated with local and systemic symptoms, in addition to triggering an inflammatory response that is accompanied by increases in laboratory markers of inflammation, such as C-reactive protein, erythrocyte sedimentation rate, and white blood cell count levels[100-102]. Therefore, in addition to routine methods for detecting biofilm-associated infections, blood and tissue cultures can be used to detect PJI infections at an early stage [101,103].
(3) Failure of antibiotic treatment and recurrence of infection. It is the formation of biofilms on the surface of biomaterials that leads to the development of drug resistance in the treatment of related infections [104]. The formation of a biofilm matrix can maintain temporary dormancy of cells, which may lead to repeated infections [105]. For example, the treatment of PJI requires complex therapeutic strategies, and it is the long-term infection problems that lead to multiple surgical revisions and long-term antimicrobial therapy [98].
5. Treatment of biofilms on the surface of biomaterials
The biofilm that causes infection is progressive. Current treatments for implant-associated infections involve administration of high doses of antibiotics and surgical replacement if symptoms persist [106]. The increase in antibiotic resistance has introduced problems in the treatment of patients using medical devices [107]. Therefore, several new biomaterials have been developed to treat clinical medical device-related biofilm infection, from the addition of antimicrobials to surface modifications of the material itself, which have been used as alternative strategies for the prevention and treatment of biofilm infections (Table 1).
5.1. Antibacterial coatings
Antibacterial materials are widely used in the biomedical field, especially in the treatment of biofilm. The main pathogens of medical device-related infections are methicillin-resistant S. aureus (MRSA) and E. coli [108]. The common strategy for the treatment of this type of bacterial infection is antibiotic treatment. However, traditional antibiotic therapy will not only cause a variety of side effects but is also more likely to cause the emergence of multi-drug-resistant bacteria [109]. As a result, new methods different from antibiotic therapy have emerged to solve the problem of infection caused by medical devices, such as the use of antibacterial coatings on the surface of implants [110]. Implant coatings against biofilm-based infections can be divided into two categories [110]: (1) Active coatings, which can fight infection by releasing pre-incorporated antimicrobial agents. Coatings are prepared by soaking a porous material or coating a surface with the desired antimicrobial compound, frequently using carriers such as hydroxyapatite (HA) and polycaprolactone (PCL) [111]. (2) Passive coatings, which prevent bacterial attachment through the use of hydrophilic materials [101].
5.2. Active coatings
The most common active coatings are antibiotic or silver compound coatings, which are applied to implant surfaces as antimicrobial or anti-biofilm agents for the prevention of medical device-related infections[112-114]. For example, the antibiotic hydroxyapatite coating, which is widely used in the medical field, can be used with antibiotic solutions and modified by surface-adsorbed antibiotics, resulting in anti-biofilm effects [40]. Polymers usually have no intrinsic antibacterial properties, and polymer antibacterial materials can be prepared by using antibacterial additives to modify a polymer matrix in different ways [115]. A study incorporated antibacterial Ag nanoparticles into a hydrophobic polymer in situ in a polycaprolactone matrix and achieved long-term release of active silver. The Ag-containing composite material had strong antibacterial and anti-biofilm properties and therefore could be used in medical antibacterial equipment [116]. Some zwitterionic polymers, such as hydrogels and sulfobetaine polymers, can delay or even prevent microorganisms from attaching to the surface because the tight hydration layer around the ion surface prevents the adsorption of non-specific proteins [117,118]. However, zwitterionic polymers cannot inactivate bacterial cells and therefore can be used as carriers to achieve synergistic antibacterial effects with fungicides [118]. Impregnating ultrasmall silver nanoparticles (AgNPs) into biocompatible thermosensitive hydrogels can not only control the delivery of AgNPs at an appropriate concentration but also confer long-term storage stability and highly potent antibacterial activity [119]. AgNP hydrogels show excellent biofilm dispersion properties and are safe and non-toxic to mammalian cells in a wound environment, which provides a promising new strategy for hydrogels as an effective antibacterial platform [119]. In addition to traditional antibacterial agents, the release of other molecules (such as enzymes and antimicrobial peptides) can be used to treat infections. One study found that the acylase enzyme coating on the surface of a silicone catheter significantly inhibited the formation of a biofilm on its surface [120]. The negatively charged acylase enzymes were immobilized on the silicone catheter through alternate deposition, and the acylase enzymes on the surface reduced the biofilm by inducing the degradation of QS signals [120]. Antimicrobial peptides have been shown to effectively kill bacterial cells without toxic effects [121,122]. For example, a thin polymer multilayer film composed of chitosan and hyaluronic acid covered a catheter, and then a β-peptide coating was applied to the surface of the membrane. The film containing the peptide inhibited bacterial biofilm formation by releasing the β-peptide [123]. These β-peptide-containing films offer a new and promising localized delivery method for preventing orthopedic implant infections.
5.3. Passive coatings
Passive coatings are used to prevent bacteria from attaching. The development of coatings that prevent bacterial attachment is a promising approach to prevent biofilm formation using non-cytotoxic mechanisms [110]. The attachment of bacteria to the surface of biomaterials mainly consists of two key stages, the initial reversible attachment through physicochemical interactions, and the irreversible attachment mediated by bacterial adhesion proteins [8]. Therefore, blocking the adhesion of bacteria on the surface of biomaterials provides a good theoretical target for the development of new therapeutic methods. Strategies to reduce bacterial adhesion to biomaterial surfaces include the use of hydrophilic polymers, such as hyaluronic acid, hydrogel coatings, and heparin coatings [8]. Medical-related devices made of hydrophobic materials are more prone to biofilm infection due to the commonly observed preferable attachment of bacteria on hydrophobic surfaces [111,124]. In contrast, hydrophilic surfaces are covered by water molecules and prevent the attachment of cells and bacteria, which play a role as a physical and energy barrier [125]. Previous studies found that hyaluronic acid has notable antiadhesive properties and shows moderate activity against bacterial biofilms [126]. A recent study combined quaternized chitosan with bactericidal properties, acylase with anti-quorum sensing properties, and hyaluronic acid with anti-adhesion properties. This multifunctional coating inhibited most bacteria from initially attaching, killed the attached bacteria, and interfered with their quorum-sensing systems involved in biofilm formation [127]. In addition, chitosan can be used to prepare thermosensitive hydrogels for the treatment of bone defects. These hydrogels have attracted wide attention because they can successfully regenerate bone tissue without surgical intervention, thus reducing the invasiveness of treatment [128,129].
Due to the enormous burden of biomaterial-related infections (those associated with implants and catheters), the effectiveness of traditional antibiotic therapy is diminished, and bacterial resistance is increasing. At the same time, the transfer of antibacterial compounds onto the surface of biomaterials is also limited, and therefore, the use of antibacterial coatings has become a new approach to prevent infection. The polymer-based composite materials composed of antibacterial coatings and polymer materials are easy to manufacture and have high biocompatibility and are therefore widely used [111]. However, there are still challenges, such as the lack of long-term performance, and many antibacterial materials have only been studied in vitro without entering clinical research [110]. Therefore, follow-up studies should address these important challenges and enable these materials to make real progress in the biomedical field.
5.4. Modification of the surface of medical implants
The physicochemical properties of the material surface of implants affect the adhesion behavior of cells on the material surface and the subsequent biofilm formation process [130]. These characteristics include electrostatic interactions and van der Waals forces between the material surface and cells, surface energy and hydrophobicity of the material, morphological characteristics of the material surface, such as roughness, surface composition, and topography, and functional chemical group modifications of the material surface [131]. The surface modification of biomaterials is a potential strategy to prevent bacteria from attaching and forming biofilms on the surface of materials [132]. According to the modification process, the surface modification of polymers can be divided into physical modifications and chemical modifications.
5.5. Physical modifications
Surface-treated polymers have been widely used with biomedical materials, including optimized modified hydrogels, nanomaterials, and phage materials [133,134]. The surface modification of polymers by physics is relatively simple, economical, and effective. The surface of the polymer is modified by physical methods to change its wettability, but the modification does not change the inherent chemical properties of the polymer [135]. Common modification methods include changing the surface roughness of materials and preparing hydrophobic or superhydrophobic surfaces, which affect cell adhesion [132,133,136].
The superhydrophobic surfaces have antibacterial adhesion properties as they naturally have nano- or microscale structures in addition to the repulsion of water, which limits the access of bacteria to the superhydrophobic surface [137,138]. A femtosecond laser-induced surface structure is a large-scale nano- and microstructure formation technology, which can effectively modify the optical, electrical, mechanical, and tribological properties of materials [139]. A surface treated by a femtosecond laser shows different surface structure on scales of the order of nano- and micrometers. Jalil et al. produced superhydrophobic surfaces on gold with femtosecond laser pulses, in which the original hydrophilic Au was transformed into a superhydrophobic surface and reduced the adhesion of Escherichia coli. The physical inhibition of bacterial colonies and biofilm formation by this technique is a crucial step in reducing antibiotic resistant infections [140]. In addition, studies have designed flexible superhydrophobic surfaces decorated with copper hydroxide nanowires that lead to dual-scale roughness and superhydrophobicity. These nanowires are grown separately and transferred onto polydimethylsiloxane (PDMS) surfaces by mechanical peeling, and thus, non-planar 3D surfaces can be fabricated [141]. These surfaces have shown blood repellence, antibacterial activity, and hemocompatibility, indicating that they are suitable for medical and health care applications.
A change of roughness will change the surface energy and hydrophobicity of a material [142], which will affect cell adhesion. It is generally believed that a lower surface roughness reduces bacterial adhesion to commonly used implant materials and prevents the formation of biofilms on the biomaterial surfaces [143,144,145]. For example, etching technology is used to design micro- or nano-scale surface structures to increase the surface roughness of biomaterials [146,147]. Studies have used a simple etching method to produce multi-scale roughness on the surface of Al alloys. This method not only rapidly etched large-size substrates but also showed a high bactericidal effect on rod-shaped and coccoid-shaped bacterial cells [148]. The latest advances in surface technology and materials have led to the development of surface patterns, some of which can affect bacterial colonization and biofilm formation without added antimicrobial agents, such as sharklet micropatterns that have been shown to control the bioadhesion of pathogenic bacteria [149,150]. Studies have incorporated nano and shark micropatterns into the surface of central venous catheters. This surface modification reduced microbial colonization and biofilm formation and can be applied clinically to reduce catheter-related infection [151]. In addition, some studies have combined chemical and topographic properties to obtain a sharkskin-mimicking graphene oxide (GO)-modified chitosan membrane with antibacterial properties and cytocompatibility. GO-coated sharkskin mimicking membranes can significantly reduce the formation of bacterial biofilms while promoting cytocompatibility and therefore can be used as a convenient biomaterial in many biomedical applications, such as those requiring a biodegradable, highly biocompatible, and antibacterial implantable medical equipment coating [152].
Unlike other surface treatment methods, physical modifications are performed as part of the surface manufacturing itself. In addition, polymers modified by physical methods are strong and do not age or deteriorate, as the resulting surface roughness rarely affects the surface chemistry of the polymers [153,154].
5.6. Chemical modification
The initial adhesion of microorganisms and the formation of biofilms can be regulated by changing the chemical properties of the material surface [155]. The main methods of surface chemical modifications include covalent modification of materials, non-covalent modifications, controlled release of small molecules, and degradation of the polymerized surface [156,157].
A commonly used chemical modification technique is the self-assembled monolayer (SAM), which is a thermodynamically stable and regularly arranged monolayer formed spontaneously by a physicochemical interaction between molecules and substrate materials [158,159]. SAM technology can effectively control the types of microbial-oriented functional groups and the concentration of ligands loaded on the material surface, and then purposefully change the surface energy and surface charge density [160]. For example, chitosan inhibits bacterial growth through the interaction between positively charged amino groups and negatively charged outer membranes [161]. It was shown that chitosan hydrogel films were covalently attached to thiol-modified substrates via a thiol one-click reaction. After that, heparin-mimicking polymer chains were grafted onto the hydrogel thin film layer via surface-initiated atom transfer radical polymerization, and covalent adhesion of the hydrogel was realized [162]. This self-defense double-layer hydrogel coating can switch from a cell adhesion surface to an antibacterial adhesion surface and provides a new approach for the antibacterial protection of biomedical equipment.
There are also several functional chemical modifications designed against the formation mechanism of bacterial biofilm, such as quorum sensing inhibitors (QSIs) to inhibit the key regulatory mechanism of bacterial biofilm formation [163,164]. Surface-immobilized QSIs block bacterial communication and affect the formation of biofilms on the surface of biomaterials [165,166]. It has been found that furanone is a substance with a similar structure to N-acyl homoserine lactone (AHL), a signal molecule of gram-negative bacterial QS, which competes with AHLs for receptors, thus interfering with QS to inhibit biofilms [167]. The intergeneric quorum sensing signal molecule autoinducer-2 (AI-2) of Fusobacterium nucleatum is very important for the development of pathogenicity and biofilm formation [168]. A new brominated furanone was recently found that inhibited biofilm formation through AI-2, and thus inhibits biofilms caused by periodontopathogens, including F. nucleatum, Porphyromonasgingivalis, and Tannerella forsythia [169].
The physicochemical interaction between the material surface and cell provides a design idea for the preparation of materials that regulate microbial film formation. Through a series of complex physical, chemical, or biological mechanisms, the surface of the material has an impact on bacterial adhesion and the formation of biofilms. There are various methods for influencing these mechanisms, including the addition of an antibacterial coating on the material surface and the surface modification of biological materials. The surface properties of materials are very important to the research and development of inhibitory biofilm-forming materials, and the method of regulating biofilms through material modification provides a more comprehensive scientific reference for related research.

Table 1.
Different approaches to treat infectious biofilms on biomaterials.
| Types | Mechanisms of action | Materials | References | |
|---|---|---|---|---|
| Antibacterial coatings | Active coatings | Fight infection by releasing antimicrobial agents | Antibiotic or silver compound coatings, enzymes, antimicrobial peptides | [40,116,119,120,123] |
| Passive coatings | Prevent bacteria from attaching using hydrophilic materials | Hyaluronic acid, hydrogel coatings | [127,128,129] | |
| Modification of the surface of medical implants | Physical modification | Affect cell adhesion by changing the surface roughness and generating hydrophobic surfaces | Femtosecond laser-induced surfaces, superhydrophobic non-planar 3D surfaces, nano and shark micropattern surfaces | [140,141,148,149,150,151,152] |
| Chemical modification | Regulate the formation of biofilms by changing the chemical properties of the material surface | Chitosan hydrogel films, surface-immobilized brominated furanones | [162,169] | |
6. Conclusion
Biofilms are the most common growth state of microorganisms. For bacteria, the formation of a biofilm is a simple, multi-stage process. Bacteria must go through three steps: initial adhesion, maturation, and dispersion. Once the biofilm is formed, a biofilm community with an ordered structure and functional differentiation can provide protection for bacteria in adverse environments, thus causing infection. Particularly in clinical practice, bacterial colonization forms biofilms on various medical instruments, causing serious nosocomial infections. At present, conventional antibiotic therapy has been unable to effectively eradicate biofilms. In contrast, with the use of antibiotics, bacteria have evolved multiple resistance mechanisms, increasing the difficulty of treating biomaterial-associated biofilm infections. Therefore, it is of great practical significance to purposefully regulate the adhesion of microorganisms on material surfaces and the formation of biofilms. The use of antibacterial coatings or surface modifications of medical materials not only prevents bacterial adhesion but also directly fights biofilm infections by releasing antibacterial drugs. In addition, appropriate antibacterial therapy can save money and shorten the duration of treatment.
With the increasing number of biomaterial studies of biofilm regulation, most of which are in vitro studies, more mechanistic studies are needed in the future to overcome these shortcomings and enable these therapies to enter clinical trials. To better solve the problem of biofilm resistance, it is necessary to conduct in-depth research and optimize preventive measures, such as developing new antibacterial coatings (biodegradable, biocompatible) and optimizing medical materials. In conclusion, the research in the field of biofilms involves the intersection and integration of multidisciplinary methods such as microbiology, molecular biology, surface chemistry, and material science, which is not only of academic significance but also of extremely important application value for clinical treatment, which still needs further research.
Author Contributions
J.L. conceptualized the article and critically revised the work. P.L., R.Y., and J.C. performed the literature search and wrote the manuscript. P.L. and R.Y. prepared the figure and table. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (32070103, 31700031), by the Qinchuang Yuan “Scientist + Engineer” Team Construction Project of Shaanxi Province (2023KXJ-019), by the Regional Development Talent Project of the “Special Support Plan” of Shaanxi Province, a grant from the Outstanding Young Talent Support Plan of the Higher Education Institutions of Shaanxi Province, and by the Youth Innovation Team of Shaanxi Universities (2022).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: a common cause of persistent infections. Science. 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Huang, M.; Melander, C.; Kjellerup, B.V. Dispersal and inhibition of biofilms associated with infections. J Appl Microbiol. 2020, 128, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Assefa, M.; Amare, A. Biofilm-associated multi-drug resistance in hospital-acquired infections: A review. Infect Drug Resist. 2022, 15, 5061–5068. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.M.; Tran, H.; Booth, M.A.; Fox, K.E.; Nguyen, T.H.; Tran, N.; Tran, P.A. Nanomaterials for treating bacterial biofilms on implantable medical devices. Nanomaterials (Basel). 2020, 10, 2253. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, Q.; Zhang, D.; Yan, W. Mechanisms and control strategies of antibiotic resistance in pathological biofilms. Journal of Microbiology and Biotechnology. 2021, 31, 1–7. [Google Scholar] [CrossRef]
- Mishra, S.; Gupta, A.; Upadhye, V.; Singh, S.C.; Sinha, R.P.; Häder, D.P. Therapeutic strategies against biofilm infections. Life (Basel). 2023, 13, 172. [Google Scholar] [CrossRef]
- Liu, X.; Yao, H.; Zhao, X.; Ge, C. Biofilm formation and control of foodborne pathogenic bacteria. Molecules. 2023, 28, 2432. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon. 2018, 4, e01067. [Google Scholar] [CrossRef]
- Wang, D.; Fletcher, G.C.; On, S.L.W.; Palmer, J.S.; Gagic, D.; Flint, S.H. Biofilm formation, sodium hypochlorite susceptibility and genetic diversity of Vibrio parahaemolyticus. Int J Food Microbiol. 2023, 385, 110011. [Google Scholar] [CrossRef]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The biofilm life cycle: expanding the conceptual model of biofilm formation. Nat Rev Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Öztürk, F.Y.; Darcan, C.; Kariptaş, E. The determination, monitoring, molecular mechanisms and formation of biofilm in E. coli. Braz J Microbiol. 2023, 54, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Cheng, J.; Wang, J.; Li, P.; Lin, J. Treatment of Pseudomonas aeruginosa infectious biofilms: Challenges and strategies. Front Microbiol. 2022, 13, 955286. [Google Scholar] [CrossRef] [PubMed]
- De Silva, L.; Heo, G.J. Biofilm formation of pathogenic bacteria isolated from aquatic animals. Arch Microbiol. 2022, 205, 36. [Google Scholar] [CrossRef] [PubMed]
- Abebe, G.M. The role of bacterial biofilm in antibiotic resistance and food contamination. Int J Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef]
- Gupta, P.; Sarkar, S.; Das, B.; Bhattacharjee, S.; Tribedi, P. Biofilm, pathogenesis and prevention--a journey to break the wall: a review. Arch Microbiol. 2016, 198, 1–15. [Google Scholar] [CrossRef]
- Stoica, P.; Chifiriuc, M.C.; Rapa, M.; Lazăr, V. Overview of biofilm-related problems in medical devices. Biofilms and Implantable Medical Devices 2017, 3–23. [Google Scholar]
- Li, P.; Zong, W.; Zhang, Z.; Lv, W.; Ji, X.; Zhu, D.; Du, X.; Wang, S. Effects and molecular mechanism of flagellar gene flgK on the motility, adhesion/invasion, and desiccation resistance of Cronobacter sakazakii. Food Res Int. 2023, 164, 112418. [Google Scholar] [CrossRef]
- Nedeljković, M.; Sastre, D.E.; Sundberg, E.J. Bacterial flagellar filament: A supramolecular multifunctional nanostructure. Int J Mol Sci. 2021, 22, 7521. [Google Scholar] [CrossRef]
- Kolenda, R.; Ugorski, M.; Grzymajlo, K. Everything you always wanted to know about salmonella type 1 fimbriae, but were afraid to ask. Front Microbiol. 2019, 10, 1017. [Google Scholar] [CrossRef]
- Puri, D.; Fang, X.; Allison, K.R. Evidence of a possible multicellular life cycle in Escherichia coli. iScience. 2023, 26, 105795. [Google Scholar] [CrossRef] [PubMed]
- Azara, E.; Longheu, C.M.; Attene, S.; Sanna, S.; Sale, M.; Addis, M.F.; Tola, S. Comparative profiling of agr locus, virulence, and biofilm-production genes of human and ovine non-aureus staphylococci. BMC Vet Res. 2022, 18, 212. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear beta-1,6-linked glucosaminoglycan: purification and structural analysis. J Bacteriol. 1996, 178, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Federle, M.J.; Bassler, B.L. Interspecies communication in bacteria. J Clin Invest. 2003, 112, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zou, H.; Li, J.; Song, T.; Lv, W.; Wang, W.; Wang, Z.; Tao, S. Impact of quorum sensing signaling molecules in gram-negative bacteria on host cells: current understanding and future perspectives. Gut Microbes. 2022, 14, 2039048. [Google Scholar] [CrossRef]
- Lin, J.; Cheng, J.; Wang, Y.; Shen, X. The Pseudomonas Quinolone Signal (PQS): Not just for quorum sensing anymore. Front Cell Infect Microbiol. 2018, 8, 230. [Google Scholar] [CrossRef]
- Wang, Y.; Bian, Z.; Wang, Y. Biofilm formation and inhibition mediated by bacterial quorum sensing. Appl Microbiol Biotechnol. 2022, 106, 6365–6381. [Google Scholar] [CrossRef]
- Cooke, A.C.; Florez, C.; Dunshee, E.B.; Lieber, A.D.; Terry, M.L.; Light, C.J.; Schertzer, J.W. Pseudomonas Quinolone Signal-induced outer membrane vesicles enhance biofilm dipersion in Pseudomonas aeruginosa. mSphere 2020, 5, e01109-01120. [Google Scholar] [CrossRef]
- Allesen-Holm, M.; Barken, K.B.; Yang, L.; Klausen, M.; Webb, J.S.; Kjelleberg, S.; Molin, S.; Givskov, M.; Tolker-Nielsen, T. A characterization of DNA release in Pseudomonas aeruginosa cultures and biofilms. Mol Microbiol. 2006, 59, 1114–1128. [Google Scholar] [CrossRef]
- Fu, H.; Wang, J.; Liu, Q.; Ding, L.; Ren, H. The role of immobilized quorum sensing strain in promoting biofilm formation of Moving Bed Biofilm Reactor during long-term stable operation. Environ Res. 2022, 215, 114159. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J Chin Med Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: an emerging link to disease pathogenesis. Annu Rev Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Cullen, M.C.; Rickard, A.H.; Zeef, L.A.; Davies, D.G.; Gilbert, P. Characterization of nutrient-induced dispersion in Pseudomonas aeruginosa PAO1 biofilm. J Bacteriol. 2004, 186, 7312–7326. [Google Scholar] [CrossRef]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb Perspect Med. 2013, 3, a010306. [Google Scholar] [CrossRef]
- Shen, D.; Langenheder, S.; Jürgens, K. Dispersal modifies the diversity and composition of active bacterial communities in response to a salinity disturbance. Front Microbiol. 2018, 9, 2188. [Google Scholar] [CrossRef]
- Wille, J.; Coenye, T. Biofilm dispersion: The key to biofilm eradication or opening Pandora's box? Biofilm. 2020, 2, 100027. [Google Scholar] [CrossRef]
- Fleming, D.; Rumbaugh, K.P. Approaches to dispersing medical biofilms. Microorganisms. 2017, 5, 15. [Google Scholar] [CrossRef]
- Oppenheimer-Shaanan, Y.; Steinberg, N.; Kolodkin-Gal, I. Small molecules are natural triggers for the disassembly of biofilms. Trends Microbiol. 2013, 21, 594–601. [Google Scholar] [CrossRef]
- Veerachamy, S.; Yarlagadda, T.; Manivasagam, G.; Yarlagadda, P.K. Bacterial adherence and biofilm formation on medical implants: a review. Proc Inst Mech Eng H. 2014, 228, 1083–1099. [Google Scholar] [CrossRef]
- Ma, L.Z.; Wang, D.; Liu, Y.; Zhang, Z.; Wozniak, D.J. Regulation of biofilm exopolysaccharide biosynthesis and degradation in Pseudomonas aeruginosa. Annu Rev Microbiol. 2022, 76, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. Polysaccharases for microbial exopolysaccharides. Carbohydrate Polymers. 1999, 319–328. [Google Scholar] [CrossRef]
- Kalia, M.; Resch, M.D.; Cherny, K.E.; Sauer, K. The alginate and motility regulator AmrZ is essential for the regulation of the dispersion response by Pseudomonas aeruginosa biofilms. mSphere. 2022, 7, e0050522. [Google Scholar] [CrossRef] [PubMed]
- Colvin, K.M.; Irie, Y.; Tart, C.S.; Urbano, R.; Whitney, J.C.; Ryder, C.; Howell, P.L.; Wozniak, D.J.; Parsek, M.R. The Pel and Psl polysaccharides provide Pseudomonas aeruginosa structural redundancy within the biofilm matrix. Environ Microbiol. 2012, 14, 1913–1928. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.T.; Niu, Y.T.; Zhang, H.; Li, P.X.; Zhang, N.M.; Cheng, J.L.; LIN, J.S. An engineered bacterium for the targeted delivery of proteins to destroy Pseudomonas aeruginosa biofilms. Acta Microbiologica Sinica 2021, 61, 2726–2748. [Google Scholar]
- Tsagkari, E.; Connelly, S.; Liu, Z.; McBride, A.; Sloan, W.T. The role of shear dynamics in biofilm formation. NPJ Biofilms Microbiomes. 2022, 8, 33. [Google Scholar] [CrossRef]
- Choi, Y.C.; Morgenroth, E. Monitoring biofilm detachment under dynamic changes in shear stress using laser-based particle size analysis and mass fractionation. Water Sci Technol. 2003, 47, 69–76. [Google Scholar] [CrossRef]
- Hwang, G.; Klein, M.I.; Koo, H. Analysis of the mechanical stability and surface detachment of mature Streptococcus mutans biofilms by applying a range of external shear forces. Biofouling. 2014, 30, 1079–1091. [Google Scholar] [CrossRef]
- Oulahal-Lagsir, N.; Martial-Gros, A.; Bonneau, M.; Blum, L.J. "Escherichia coli-milk" biofilm removal from stainless steel surfaces: synergism between ultrasonic waves and enzymes. Biofouling. 2003, 19, 159–168. [Google Scholar]
- Oulahal, N.; Martial-Gros, A.; Bonneau, M.; Blum, L.J.J.B. Combined effect of chelating agents and ultrasound on biofilm removal from stainless steel surfaces. Application to Escherichia coli milk and Staphylococcus aureus milk biofilms. 2004, 1, 65–73. [Google Scholar]
- Nigri, G.R.; Tsai, S.; Kossodo, S.; Waterman, P.; Fungaloi, P.; Hooper, D.C.; Doukas, A.G.; LaMuraglia, G.M. Laser-induced shock waves enhance sterilization of infected vascular prosthetic grafts. Lasers Surg Med. 2001, 29, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal infections: mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu Rev Med. 2013, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yang, Q.; Zhang, D.; Yan, W. Mechanisms and control strategies of antibiotic resistance in pathological biofilms. J Microbiol Biotechnol. 2021, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J Clin Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Michael, *!!! REPLACE !!!*; Waturangi, D.E. Antibiofilm activity from endophyte bacteria, Vibrio cholerae strains, and actinomycetes isolates in liquid and solid culture. BMC Microbiol. 2023, 23, 83. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, K.; Dawadi, B.R.; Shrestha, L.B. Role of biofilm in bacterial infection and antimicrobial resistance. JNMA J Nepal Med Assoc. 2022, 60, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Carbone, D.; Parrino, B.; Pecoraro, C.; Giovannetti, E.; Cirrincione, G.; Diana, P. Therapeutic strategies to counteract antibiotic resistance in MRSA biofilm-associated infections. ChemMedChem. 2021, 16, 65–80. [Google Scholar] [CrossRef]
- Thomsen, K.; Høiby, N.; Jensen, P.; Ciofu, O.; Moser, C. Immune response to biofilm growing pulmonary Pseudomonas aeruginosa infection. Biomedicines. 2022, 10, 2064. [Google Scholar] [CrossRef]
- Güneş, B.; Akçelik, N. The role of eDNA in biofilm structure of Enterococcus faecalis and investigation of the efficiency of enzyme and antibiotic application in biofilm eradication. Mikrobiyol Bul. 2022, 56, 606–619. [Google Scholar]
- Sheng, Y.; Chen, Z.; Wu, W.; Lu, Y. Engineered organic nanoparticles to combat biofilms. Drug Discov Today. 2023, 28, 103455. [Google Scholar] [CrossRef]
- Tian, C.; Yuan, M.; Tao, Q.; Xu, T.; Liu, J.; Huang, Z.; Wu, Q.; Pan, Y.; Zhao, Y.; Zhang, Z. Discovery of novel resistance mechanisms of Vibrio parahaemolyticus biofilm against aminoglycoside antibiotics. Antibiotics (Basel). 2023, 12, 638. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. Apmis. 2017, 125, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents. 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Mohamed, H.M.A.; Alnasser, S.M.; Abd-Elhafeez, H.H.; Alotaibi, M.; Batiha, G.E.; Younis, W. detection of β-Lactamase resistance and biofilm genes in Pseudomonas species isolated from chickens. Microorganisms. 2022, 10, 1975. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Moreau, A.; Khodaparast, S.; Perazzo, A.; Feng, J.; Fei, C.; Mao, S.; Mukherjee, S.; Košmrlj, A.; Wingreen, N.S.; Bassler, B.L.; Stone, H.A. Bacterial biofilm material properties enable removal and transfer by capillary peeling. Adv Mater. 2018, 30, e1804153. [Google Scholar] [CrossRef]
- Spoering, A.L.; Lewis, K. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef]
- Mah, T.F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int J Med Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu Rev Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Pamp, S.J.; Gjermansen, M.; Johansen, H.K.; Tolker-Nielsen, T. Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol Microbiol. 2008, 68, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, A.; Bakht, P.; Saini, M.; Pandey, S.; Pathania, R. Role of bacterial efflux pumps in antibiotic resistance, virulence, and strategies to discover novel efflux pump inhibitors. Microbiology (Reading). 2023, 169, 001333. [Google Scholar] [CrossRef] [PubMed]
- Seukep, A.J.; Mbuntcha, H.G.; Kuete, V.; Chu, Y.; Fan, E.; Guo, M.Q. What approaches to thwart bacterial efflux pumps-mediated resistance? Antibiotics (Basel). 2022, 11, 1287. [Google Scholar] [CrossRef] [PubMed]
- Başaran, S.N.; Öksüz, L. The role of efflux pumps ın antıbıotıc resıstance of gram negatıve rods. Arch Microbiol. 2023, 205, 192. [Google Scholar] [CrossRef] [PubMed]
- Mudde, S.E.; Schildkraut, J.A.; Ammerman, N.C.; de Vogel, C.P.; de Steenwinkel, J.E.M.; van Ingen, J.; Bax, H.I. Unraveling antibiotic resistance mechanisms in Mycobacterium abscessus: the potential role of efflux pumps. J Glob Antimicrob Resist. 2022, 31, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, V.; Kashyap, A.; Raja, R.; Newar, K.; Rai, D.; Dixit, N.M.; Mehra, S. The extent of antimicrobial resistance due to efflux pump regulation. ACS Infect Dis. 2022, 8, 2374–2388. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; Hossain, M.J.; Koirala, N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Grossman, T.H. Tetracycline antibiotics and resistance. Cold Spring Harb Perspect Med. 2016, 6, a025387. [Google Scholar] [CrossRef]
- Venkatesan, N.; Perumal, G.; Doble, M. Bacterial resistance in biofilm-associated bacteria. Future Microbiol. 2015, 10, 1743–1750. [Google Scholar] [CrossRef]
- Shi, K.; Cao, M.; Li, C.; Huang, J.; Zheng, S.; Wang, G. Efflux proteins MacAB confer resistance to arsenite and penicillin/macrolide-type antibiotics in Agrobacterium tumefaciens 5A. World J Microbiol Biotechnol. 2019, 35, 115. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, L.; Haghshenas, M.R.; Mirzaei, B.; Khalili, Y.; Goli, H.R. Role of MexAB-OprM efflux pump in the emergence of multidrug-resistant clinical isolates of Pseudomonas aeruginosa in Mazandaran province of Iran. Mol Biol Rep. 2023, 50, 2603–2609. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Espada, R.; Shahrour, H.; Pitts, B.; Stewart, P.S.; Sánchez-Gómez, S.; Martínez-de-Tejada, G. A permeability-increasing drug synergizes with bacterial efflux pump inhibitors and restores susceptibility to antibiotics in multi-drug resistant Pseudomonas aeruginosa strains. Sci Rep. 2019, 9, 3452. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Bayer, A.S.; Mishra, N.N.; Meehl, M.; Ledala, N.; Yeaman, M.R.; Xiong, Y.Q.; Cheung, A.L. The Staphylococcus aureus two-component regulatory system, GraRS, senses and confers resistance to selected cationic antimicrobial peptides. Infect Immun. 2012, 80, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Costa, S.K.; Wierzbicki, R.M.; Rigby, W.F.C.; Cheung, A.L. The extracellular loop of the membrane permease VraG interacts with GraS to sense cationic antimicrobial peptides in Staphylococcus aureus. PLoS Pathog. 2021, 17, e1009338. [Google Scholar] [CrossRef] [PubMed]
- Hiron, A.; Falord, M.; Valle, J.; Débarbouillé, M.; Msadek, T. Bacitracin and nisin resistance in Staphylococcus aureus: a novel pathway involving the BraS/BraR two-component system (SA2417/SA2418) and both the BraD/BraE and VraD/VraE ABC transporters. Mol Microbiol. 2011, 81, 602–622. [Google Scholar] [CrossRef] [PubMed]
- Arii, K.; Kawada-Matsuo, M.; Oogai, Y.; Noguchi, K.; Komatsuzawa, H. Single mutations in BraRS confer high resistance against nisin A in Staphylococcus aureus. Microbiologyopen. 2019, 8, e791. [Google Scholar] [CrossRef]
- Babushkina, I.V.; Bondarenko, A.S.; Ulyanov, V.Y.; Mamonova, I.A. Biofilm formation by gram-negative bacteria during implant-associated infection. Bull Exp Biol Med. 2020, 169, 365–368. [Google Scholar] [CrossRef]
- Barros, J.; Monteiro, F.J.; Ferraz, M.P. Bioengineering approaches to fight against orthopedic biomaterials related-infections. Int J Mol Sci. 2022, 23, 11658. [Google Scholar] [CrossRef]
- Chouirfa, H.; Bouloussa, H.; Migonney, V.; Falentin-Daudré, C. Review of titanium surface modification techniques and coatings for antibacterial applications. Acta Biomater. 2019, 83, 37–54. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Holá, V.; Imbert, C.; Kirketerp-Møller, K.; Lebeaux, D.; Oliver, A.; Ullmann, A.J.; Williams, C. ESCMID guideline for the diagnosis and treatment of biofilm infections 2014. Clin Microbiol Infect. 2015, 21 Suppl 1, S1–25. [Google Scholar] [CrossRef]
- Wi, Y.M.; Patel, R. Understanding biofilms and novel approaches to the diagnosis, prevention, and treatment of medical devicea-Associated infections. Infect Dis Clin North Am. 2018, 32, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, D.; Fernández-Hidalgo, N.; Chauhan, A.; Lee, S.; Ghigo, J.M.; Almirante, B.; Beloin, C. Management of infections related to totally implantable venous-access ports: Challenges and perspectives. Lancet Infect Dis. 2014, 14, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Gominet, M.; Compain, F.; Beloin, C.; Lebeaux, D. Central venous catheters and biofilms: where do we stand in 2017? Apmis. 2017, 125, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Mangner, N.; Woitek, F.; Haussig, S.; Schlotter, F.; Stachel, G.; Höllriegel, R.; Wilde, J.; Lindner, A.; Holzhey, D.; Leontyev, S.; Mohr, F.W.; Schuler, G.; Linke, A. Incidence, predictors, and outcome of patients developing infective endocarditis following transfemoral transcatheter aortic valve replacement. J Am Coll Cardiol. 2016, 67, 2907–2908. [Google Scholar] [CrossRef] [PubMed]
- Elgharably, H.; Hussain, S.T.; Shrestha, N.K.; Blackstone, E.H.; Pettersson, G.B. Current hypotheses in cardiac surgery: biofilm in infective endocarditis. Semin Thorac Cardiovasc Surg. 2016, 28, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Nappi, F.; Iervolino, A.; Singh, S.S.A. The new challenge for heart endocarditis: from conventional prosthesis to new devices and platforms for the treatment of structural heart disease. Biomed Res Int. 2021, 2021, 7302165. [Google Scholar] [CrossRef] [PubMed]
- Izakovicova, P.; Borens, O.; Trampuz, A. Periprosthetic joint infection: current concepts and outlook. EFORT Open Rev. 2019, 4, 482–494. [Google Scholar] [CrossRef]
- Kamihata, S.; Ando, W.; Nakahara, I.; Enami, H.; Takashima, K.; Uemura, K.; Hamada, H.; Sugano, N. Optimizing vancomycin release from novel carbon fiber-reinforced polymer implants with small holes: periprosthetic joint infection treatment. J Artif Organs. (Online ahead of print). 2023, 1–8. [Google Scholar] [CrossRef]
- Belgiovine, C.; Pellegrino, L.; Bulgarelli, A.; Lauta, F.C.; Di Claudio, A.; Ciceri, R.; Cancellara, A.; Calcaterra, F.; Mavilio, D.; Grappiolo, G.; Chiappetta, K.; Loppini, M.; Rusconi, R. Interaction of bacteria, immune cells, and surface topography in periprosthetic joint infections. Int J Mol Sci. 2023, 24, 9028. [Google Scholar] [CrossRef]
- Gbejuade, H.O.; Lovering, A.M.; Webb, J.C. The role of microbial biofilms in prosthetic joint infections. Acta Orthop. 2015, 86, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R. Diagnosis and management of prosthetic joint infection: clinical practice guidelines by the Infectious Diseases Society of America. Clin Infect Dis. 2013, 56, e1–e25. [Google Scholar] [CrossRef] [PubMed]
- Jacqueline, C.; Caillon, J. Impact of bacterial biofilm on the treatment of prosthetic joint infections. J Antimicrob Chemother. 2014, 69 Suppl 1, i37–40. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: an emerging battleground in microbial communities. Antimicrob Resist Infect Control. 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Pham, D.T.N.; Tabassum, N.; Oloketuyi, S.F.; Kim, Y.M. Treatment strategies targeting persister cell formation in bacterial pathogens. Crit Rev Microbiol. 2020, 46, 665–688. [Google Scholar] [CrossRef] [PubMed]
- Sousa, V.; Mardas, N.; Spratt, D.; Hassan, I.A.; Walters, N.J.; Beltrán, V.; Donos, N. The effect of microcosm biofilm decontamination on surface topography, chemistry, and biocompatibility dynamics of implant titanium surfaces. Int J Mol Sci. 2022, 23, 10033. [Google Scholar] [CrossRef]
- Folliero, V.; Franci, G.; Dell'Annunziata, F.; Giugliano, R.; Foglia, F.; Sperlongano, R.; De Filippis, A.; Finamore, E.; Galdiero, M. Evaluation of antibiotic resistance and biofilm production among clinical strain isolated from medical devices. Int J Microbiol. 2021, 2021, 9033278. [Google Scholar] [CrossRef] [PubMed]
- Olivares, E.; Badel-Berchoux, S.; Provot, C.; Prévost, G.; Bernardi, T.; Jehl, F. Clinical impact of antibiotics for the treatment of Pseudomonas aeruginosa biofilm infections. Front Microbiol. 2019, 10, 2894. [Google Scholar] [CrossRef]
- Srinivasan, R.; Santhakumari, S.; Poonguzhali, P.; Geetha, M.; Dyavaiah, M.; Xiangmin, L. Bacterial biofilm inhibition: A focused rview on recent therapeutic strategies for combating the biofilm mediated infections. Front Microbiol. 2021, 12, 676458. [Google Scholar] [CrossRef]
- Cloutier, M.; Mantovani, D.; Rosei, F. Antibacterial coatings: Challenges, perspectives, and opportunities. Trends Biotechnol. 2015, 33, 637–652. [Google Scholar] [CrossRef]
- Olmo, J.A.-D.; Ruiz-Rubio, L.; Pérez-Alvarez, L.; Sáez-Martínez, V.; Vilas-Vilela, J.L. Antibacterial Coatings for Improving the Performance of Biomaterials. Coatings. 2020, 10. [Google Scholar] [CrossRef]
- Bruna, T.; Maldonado-Bravo, F.; Jara, P.; Caro, N. Silver nanoparticles and their antibacterial applications. Int J Mol Sci. 2021, 22, 7202. [Google Scholar] [CrossRef] [PubMed]
- Romanò, C.L.; Scarponi, S.; Gallazzi, E.; Romanò, D.; Drago, L. Antibacterial coating of implants in orthopaedics and trauma: a classification proposal in an evolving panorama. J Orthop Surg Res. 2015, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Kuehl, R.; Brunetto, P.S.; Woischnig, A.K.; Varisco, M.; Rajacic, Z.; Vosbeck, J.; Terracciano, L.; Fromm, K.M.; Khanna, N. Preventing implant-associated infections by silver coating. Antimicrob Agents Chemother. 2016, 60, 2467–2475. [Google Scholar] [CrossRef]
- Haktaniyan, M.; Bradley, M. Polymers showing intrinsic antimicrobial activity. Chem Soc Rev. 2022, 51, 8584–8611. [Google Scholar] [CrossRef]
- Tran, P.A.; Hocking, D.M.; O'Connor, A.J. In situ formation of antimicrobial silver nanoparticles and the impregnation of hydrophobic polycaprolactone matrix for antimicrobial medical device applications. Mater Sci Eng C Mater Biol Appl. 2015, 47, 63–69. [Google Scholar] [CrossRef]
- Leng, C.; Hung, H.C.; Sun, S.; Wang, D.; Li, Y.; Jiang, S.; Chen, Z. Probing the surface hydration of nonfouling zwitterionic and PEG materials in contact with proteins. ACS Appl Mater Interfaces. 2015, 7, 16881–16888. [Google Scholar] [CrossRef]
- Liu, C.; Faria, A.F.; Ma, J.; Elimelech, M. Mitigation of biofilm development on thin-film composite membranes functionalized with zwitterionic polymers and silver nanoparticles. Environ Sci Technol. 2017, 51, 182–191. [Google Scholar] [CrossRef]
- Haidari, H.; Kopecki, Z.; Bright, R.; Cowin, A.J.; Garg, S.; Goswami, N.; Vasilev, K. Ultrasmall AgNP-impregnated biocompatible hydrogel with highly effective biofilm elimination properties. ACS Appl Mater Interfaces. 2020, 12, 41011–41025. [Google Scholar] [CrossRef]
- Ivanova, K.; Fernandes, M.M.; Mendoza, E.; Tzanov, T. Enzyme multilayer coatings inhibit Pseudomonas aeruginosa biofilm formation on urinary catheters. Appl Microbiol Biotechnol. 2015, 99, 4373–4385. [Google Scholar] [CrossRef]
- Costa, B.; Martínez-de-Tejada, G.; Gomes, P.A.C.; MC, L.M.; Costa, F. Antimicrobial peptides in the battle against orthopedic implant-related infections: A review. Pharmaceutics. 2021, 13, 1918. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Costa, F.; Monteiro, C.; Duarte, F.; Martins, M.C.L.; Gomes, P. Antimicrobial coatings prepared from Dhvar-5-click-grafted chitosan powders. Acta Biomater. 2019, 84, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Lopez, A.L.; Lee, M.R.; Ortiz, B.J.; Gastfriend, B.D.; Whitehead, R.; Lynn, D.M.; Palecek, S.P. Preventing S. aureus biofilm formation on titanium surfaces by the release of antimicrobial beta-peptides from polyelectrolyte multilayers. Acta Biomater. 2019, 93, 50–62. [Google Scholar] [CrossRef]
- Guo, S.; Zhu, X.; Loh, X.J. Controlling cell adhesion using layer-by-layer approaches for biomedical applications. Mater Sci Eng C Mater Biol Appl. 2017, 70, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, D.; van den Boogaert, I.; Miller, J.; Presswell, R.; Jouhara, H. Hydrophilic and hydrophobic materials and their applications. Energy Sources, Part A: Recovery, Utilization, and Environmental Effects 2018, 40, 2686–2725. [Google Scholar] [CrossRef]
- Drago, L.; Cappelletti, L.; De Vecchi, E.; Pignataro, L.; Torretta, S.; Mattina, R. Antiadhesive and antibiofilm activity of hyaluronic acid against bacteria responsible for respiratory tract infections. APMIS. 2014, 122, 1013–1019. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, C.; Zhang, H.; Wu, Y.; Lin, Y.; Cheng, J.; Lu, K.; Li, L.; Zhang, Y.; Chen, H.; Yu, Q. Three lines of defense: A multifunctional coating with anti-adhesion, bacteria-killing and anti-quorum sensing properties for preventing biofilm formation of Pseudomonas aeruginosa. Acta Biomater. 2022, 151, 254–263. [Google Scholar] [CrossRef]
- Saravanan, S.; Vimalraj, S.; Thanikaivelan, P.; Banudevi, S.; Manivasagam, G. A review on injectable chitosan/beta glycerophosphate hydrogels for bone tissue regeneration. Int J Biol Macromol. 2019, 121, 38–54. [Google Scholar] [CrossRef]
- Robert, B.; Chenthamara, D.; Subramaniam, S. Fabrication and biomedical applications of Arabinoxylan, Pectin, Chitosan, soy protein, and silk fibroin hydrogels via laccase - Ferulic acid redox chemistry. International Journal of Biological Macromolecules. 2022, 201, 539–556. [Google Scholar] [CrossRef]
- Nemani, S.K.; Annavarapu, R.K.; Mohammadian, B.; Raiyan, A.; Sojoudi, H.J.A.M.I. Surface modification: Surface modification of polymers: methods and applications. Adv. Mater. Interfaces 2018, 5, 1870121. [Google Scholar] [CrossRef]
- Al-Gunaid, T.A.; Krupa, I.; Ouederni, M.; Krishnamoorthy, S.K.; Popelka, A. Enhancement of adhesion characteristics of low-density polyethylene using atmospheric plasma iitiated-grafting of polyethylene glycol. Polymers (Basel). 2021, 13, 1309. [Google Scholar] [CrossRef] [PubMed]
- Sterzenbach, T.; Helbig, R.; Hannig, C.; Hannig, M. Bioadhesion in the oral cavity and approaches for biofilm management by surface modifications. Clin Oral Investig. 2020, 24, 4237–4260. [Google Scholar] [CrossRef] [PubMed]
- Nemani, S.K.; Annavarapu, R.K.; Mohammadian, B.; Raiyan, A.; Heil, J.; Haque, M.A.; Abdelaal, A.; Sojoudi, H. Surface Modification of Polymers: Methods and Applications. Advanced Materials Interfaces. 2018, 5, 1870121. [Google Scholar] [CrossRef]
- Klodzinska, S.N.; Wan, F.; Jumaa, H.; Sternberg, C.; Rades, T.; Nielsen, H.M. Utilizing nanoparticles for improving anti-biofilm effects of azithromycin: A head-to-head comparison of modified hyaluronic acid nanogels and coated poly (lactic-co-glycolic acid) nanoparticles. J Colloid Interface Sci. 2019, 555, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.; Kumari, R.M.; Verma, V.; Nimesh, S. Recent development in therapeutic strategies targeting Pseudomonas aeruginosa biofilms – A review. Materials Today: Proceedings 2021, 46, 2359–2373. [Google Scholar] [CrossRef]
- Encinas, N.; Pantoja, M.; Abenojar, J.; Martínez, M.A. Control of Wettability of Polymers by Surface Roughness Modification. Journal of Adhesion Science and Technology. 2010, 24, 1869–1883. [Google Scholar] [CrossRef]
- Epperlein, N.; Menzel, F.; Schwibbert, K.; Koter, R.; Bonse, J.; Sameith, J.; Krüger, J.; Toepel, J. Influence of femtosecond laser produced nanostructures on biofilm growth on steel. Applied Surface Science. 2017, 418, 420–424. [Google Scholar] [CrossRef]
- Gillett, A.; Waugh, D.; Lawrence, J.; Swainson, M.; Dixon, R. Laser surface modification for the prevention of biofouling by infection causing Escherichia Coli. Journal of Laser Applications. 2016, 28, 022503. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, Q.; Long, M.; Han, R.; Cao, K.; Zhang, S.; Feng, D.; Jia, T.; Sun, Z.; Qiu, J.; Xu, H. Femtosecond laser-induced periodic structures: mechanisms, techniques, and applications. Opto-Electronic Science 2022, 1, 220005. [Google Scholar] [CrossRef]
- Jalil, S.A.; Akram, M.; Bhat, J.A.; Hayes, J.J.; Singh, S.C.; ElKabbash, M.; Guo, C. Creating superhydrophobic and antibacterial surfaces on gold by femtosecond laser pulses. Appl Surf Sci. 2020, 506, 144952. [Google Scholar] [CrossRef]
- Tripathy, A.; Kumar, A.; Sreedharan, S.; Muralidharan, G.; Pramanik, A.; Nandi, D.; Sen, P. Fabrication of low-cost flexible superhydrophobic antibacterial surface with dual-scale roughness. ACS Biomater Sci Eng. 2018, 4, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, Y.; Cao, Y.; Li, G.; Liao, Y. Influence of surface roughness on contact angle hysteresis and spreading work. Colloid and Polymer Science. 2020, 298, 1107–1112. [Google Scholar] [CrossRef]
- Wu, S.; Altenried, S.; Zogg, A.; Zuber, F.; Maniura-Weber, K.; Ren, Q. Role of the surface nanoscale roughness of stainless steel on bacterial adhesion and microcolony formation. ACS Omega. 2018, 3, 6456–6464. [Google Scholar] [CrossRef] [PubMed]
- Kozmos, M.; Virant, P.; Rojko, F.; Abram, A.; Rudolf, R.; Raspor, P.; Zore, A.; Bohinc, K. Bacterial adhesion of Streptococcus mutans to dental material surfaces. Molecules. 2021, 26, 1152. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Gao, Y.; Jin, T.; Luo, X.; Zeng, Q.; Shang, Z. Effects of surface roughness and texture on the bacterial adhesion on the bearing surface of bio-ceramic joint implants: An in vitro study. Ceramics International. 2020, 46, 6550–6559. [Google Scholar] [CrossRef]
- Matos, G.R.M. Surface roughness of dental implant and osseointegration. J Maxillofac Oral Surg. 2021, 20, 1–4. [Google Scholar] [CrossRef]
- Nazarov, D.V.; Smirnov, V.M.; Zemtsova, E.G.; Yudintceva, N.M.; Shevtsov, M.A.; Valiev, R.Z. Enhanced osseointegrative properties of ultra-fine-grained titanium implants modified by chemical etching and atomic layer deposition. ACS Biomater Sci Eng. 2018, 4, 3268–3281. [Google Scholar] [CrossRef]
- Hasan, J.; Jain, S.; Padmarajan, R.; Purighalla, S.; Sambandamurthy, V.K.; Chatterjee, K. Multi-scale surface topography to minimize adherence and viability of nosocomial drug-resistant bacteria. Mater Des. 2018, 140, 332–344. [Google Scholar] [CrossRef]
- May, R.M.; Hoffman, M.G.; Sogo, M.J.; Parker, A.E.; O'Toole, G.A.; Brennan, A.B.; Reddy, S.T. Micro-patterned surfaces reduce bacterial colonization and biofilm formation in vitro: Potential for enhancing endotracheal tube designs. Clin Transl Med. 2014, 3, 1–9. [Google Scholar] [CrossRef]
- Xu, B.; Wei, Q.; Mettetal, M.R.; Han, J.; Rau, L.; Tie, J.; May, R.M.; Pathe, E.T.; Reddy, S.T.; Sullivan, L.; Parker, A.E.; Maul, D.H.; Brennan, A.B.; Mann, E.E. Surface micropattern reduces colonization and medical device-associated infections. J Med Microbiol. 2017, 66, 1692–1698. [Google Scholar] [CrossRef]
- May, R.M.; Magin, C.M.; Mann, E.E.; Drinker, M.C.; Fraser, J.C.; Siedlecki, C.A.; Brennan, A.B.; Reddy, S.T. An engineered micropattern to reduce bacterial colonization, platelet adhesion and fibrin sheath formation for improved biocompatibility of central venous catheters. Clin Transl Med. 2015, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Rostami, S.; Puza, F.; Ucak, M.; Ozgur, E.; Gul, O.; Ercan, U.K.; Garipcan, B. Bifunctional sharkskin mimicked chitosan/graphene oxide membranes: Reduced biofilm formation and improved cytocompatibility. Applied Surface Science. 2021, 544, 148828. [Google Scholar] [CrossRef]
- Wardhono, E.Y.; Pinem, M.P.; Susilo, S.; Siom, B.J.; Sudrajad, A.; Pramono, A.; Meliana, Y.; Guénin, E. Modification of physio-mechanical properties of chitosan-based films via physical treatment approach. Polymers (Basel). 2022, 14, 5216. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Seenivasan, M.K.; Inbarajan, A. A literature review on biofilm formation on silicone and poymethyl methacrylate used for maxillofacial prostheses. Cureus. 2021, 13, e20029. [Google Scholar] [CrossRef] [PubMed]
- Desrousseaux, C.; Sautou, V.; Descamps, S.; Traoré, O. Modification of the surfaces of medical devices to prevent microbial adhesion and biofilm formation. J Hosp Infect. 2013, 85, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Renner, L.D.; Weibel, D.B. Physicochemical regulation of biofilm formation. MRS Bull. 2011, 36, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.Z.; Zhao, X.X.; Liao, Y.H. Effect of material surface characteristics on biofilm formation and its application. Microbiol China 2018, 45, 155–165. [Google Scholar]
- Freitas, S.C.; Correa-Uribe, A.; Martins, M.C.L.; Pelaez-Vargas, A. Self-assembled monolayers for dental implants. Int J Dent. 2018, 2018, 4395460. [Google Scholar] [CrossRef]
- Somasundaram, S. Silane coatings of metallic biomaterials for biomedical implants: A preliminary review. J Biomed Mater Res B Appl Biomater. 2018, 106, 2901–2918. [Google Scholar] [CrossRef]
- Nicosia, C.; Huskens, J. Reactive self-assembled monolayers: from surface functionalization to gradient formation. Mater Horiz. 2014, 1, 32–45. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Kim, K.D.; Chun, S.C. Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes. 2020, 8. [Google Scholar] [CrossRef]
- He M, W.Q.; Zhao, W. A self-defensive bilayer hydrogel coating with bacteria triggered switching from cell adhesion to antibacterial adhesion. Polymer Chemistry. 2017, 8, 5344–5353. [Google Scholar] [CrossRef]
- Tan, W.-S.; Law, J.W.-F.; Law, L.N.-S.; Letchumanan, V.; Chan, K.-G. Insights into quorum sensing (QS): QS-regulated biofilm and inhibitors. Progress In Microbes & Molecular Biology 2020, 3, a0000141. [Google Scholar]
- Sionov, R.V.; Steinberg, D. Targeting the holy triangle of quorum sensing, biofilm formation, and antibiotic resistance in pathogenic bacteria. Microorganisms. 2022, 10, 1239. [Google Scholar] [CrossRef]
- Vashistha, A.; Sharma, N.; Nanaji, Y.; Kumar, D.; Singh, G.; Barnwal, R.P.; Yadav, A.K. Quorum sensing inhibitors as Therapeutics: Bacterial biofilm inhibition. Bioorganic Chemistry. 2023, 136, 106551. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, Y.; Ge, Y.; Zhu, X.; Pan, J. Regulatory mechanisms and promising applications of quorum sensing-inhibiting agents in control of bacterial biofilm Formation. Front Microbiol. 2020, 11, 589640. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S.; Hachicha, M.; Boukraa, M.; Soulère, L.; Efrit, M.L.; Queneau, Y. Heterocyclic chemistry applied to the design of N-Acyl homoserine lactone analogues as bacterial quorum sensing signals mimics. Molecules. 2021, 26, 5135. [Google Scholar] [CrossRef]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat Rev Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Park, J.S.; Ryu, E.J.; Li, L.; Choi, B.K.; Kim, B.M. New bicyclic brominated furanones as potent autoinducer-2 quorum-sensing inhibitors against bacterial biofilm formation. Eur J Med Chem. 2017, 137, 76–87. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



