
Submitted:
29 June 2023
Posted:
30 June 2023
You are already at the latest version
Abstract
Helicobacter pylori is a gastric pathogen that infects nearly half of the global population and is recognized as a group 1 carcinogen by the Word Health Organization. The global rise in antibiotic resistance has increased clinical challenges in treating H. pylori infections. Biofilm growth has been proposed to contribute to H. pylori’s chronic colonization of the host stomach, treatment failures, and the eventual development of gastric diseases. Several components of H. pylori have been identified to promote biofilm growth, and several of these may also facilitate antibiotic tolerance, including the extracellular matrix, outer membrane proteins, the coccoid morphology, modulated metabolism, efflux pumps, and virulence factors. Recent developments in therapeutic approaches targeting H. pylori biofilm have shown that synthetic compounds, such as small molecule drugs and plant-derived compounds, are effective at eradicating H. pylori biofilms. These combined topics highlight the necessity for biofilm-based research in H. pylori, to improve current H. pylori targeted therapeutic approaches and alleviate relative public health burden. In this review we discuss recent discoveries that have decoded the life cycle of H. pylori biofilms and current biofilm targeted treatment strategies.
Keywords:
Helicobacter pylori
; biofilms
; planktonic
; antibiotic resistance
; extra polymeric substance
; abiotic/biotic adhesion
; dispersion
; clinical treatment strategies
; anti-biofilm strategies
Introduction
Helicobacter pylori is a gram negative, spiral shaped, bacterial pathogen that colonizes the gastric epithelium (Cellini et al., 1994, Noach et al., 1994 and Dubois et al., 1994, Malfertheiner et al., 2023). H. pylori has been globally recognized as a high priority pathogen as it has been associated with various gastric diseases, including peptic ulcers, chronic gastritis (Labenz et al., 1994 and Blaser et al., 1994), gastric mucosa-associated tissue lymphomas (Pereira and Medeiros et al., 2014) and gastric adenocarcinomas (Asaka et al., 1994, Veldhuyzen et al., 1994, Isaacson 1994, Ma et al., 2022). Mechanisms of transmission remain unknown (Ma et al., 2022), but antibiotic therapies used to treat H. pylori infection have alarmingly been losing efficacy in regions with high infection burden (Liu et al., 2022). Antibiotic-resistant H. pylori was reported to disproportionately affect children in Asian, African, and European countries (Karbalaei et al., 2022), and in underserved communities in the US (Brown et al., 2022). One current perspective is that H. pylori in biofilms, a low growth state, may substantially promote antibiotic resistance and persistence in the host stomach (Cellini et al., 2008). Planktonic H. pylori were observed in vitro to form water-insoluble biofilms which are defined as stationary aggregates of cells encased in extra polymeric substances (EPS) (Mackay et al., 1998 and Stark et al., 1999). Although molecular mechanisms that facilitate H. pylori biofilms remain elusive in vivo, H. pylori in a form consistent with biofilms have been observed in the gastric mucosa of patients with peptic ulcers (Carron et al, 2006 and Coticchia et al., 2006). In this review we discuss recent discoveries that characterize the features of H. pylori biofilms, decode processes that regulate biofilm growth in vitro and in vivo, elucidate biofilm mechanisms that support antibiotic tolerance and current biofilm-based eradication strategies.
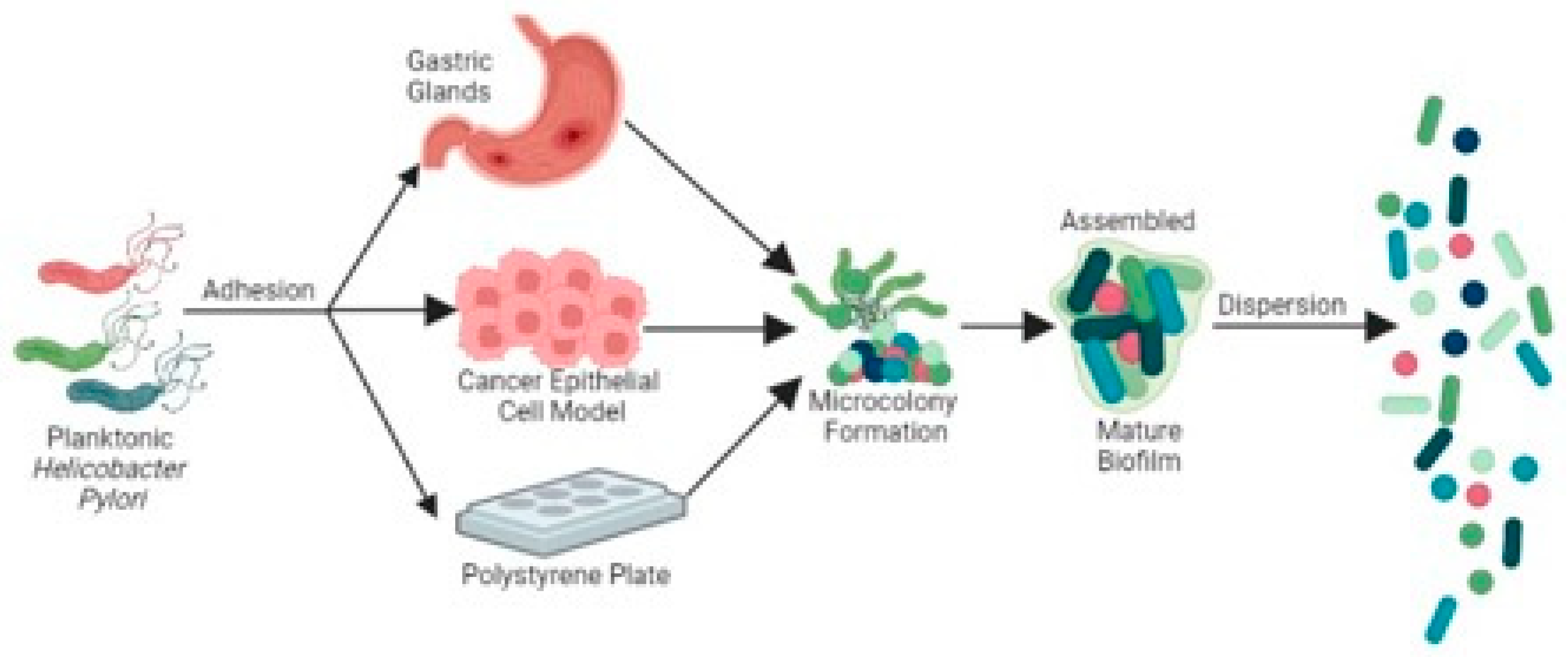
General features of H. pylori biofilms
H. pylori biofilms consist of stationary aggregates of cells encased by an extracellular matrix composed of proteins (Windham et al., 2018), extracellular DNA (Grande et al., 2011) and polysaccharides (Li et al., 2019). H. pylori biofilm formation starts from planktonic cells that adhere to either abiotic or biotic surfaces to form microcolonies with three dimensional structures (Cole et al., 2004; Hathroubi et al., 2020). Biofilm formation is initiated when planktonic H. pylori adheres to abiotic surfaces to form microcolonies (Hathroubi et al., 2018); adherence and subsequently biofilm formation were found to occur optimally in conditions lacking fetal bovine serum (Williams et al., 2008, Hathroubi et al., 2018). Aside from biofilm growth on abiotic surfaces, additional studies have also suggested that H. pylori can form a microcolony network that adhered and grew between epithelial cell junctions on human cells (Tan et al., 2009; Anderson et al., 2015) and in murine gastric glands (Sigal et al., 2015). Mature H. pylori biofilms consist of a multicellular population with a variety of cell shapes, that vary based on conditions. On abiotic surfaces, most cells adopt the coccoid morphology, with the minority displaying a rod shape (Cellini et al., 2008 and Bugli et al., 2016). As found in other bacteria, H. pylori biofilm formation exhibits a similar multiple-step process, including bacterial adherence, biofilm assembly, mature biofilm formation and dispersion. In the next sections, we dissect the features of each step in H. pylori biofilm growth.
Adherence
Adhesion is an essential process that initiates H. pylori biofilm formation and retains a role throughout the lifetime of the biofilm (Azevedoet al., 2006, Ratthawongjirakul et al., 2016, Wong et al., 2016). Prior studies have found that H. pylori can adhere to both gastric epithelial cells (Hessey et al., 1990) and abiotic surfaces (Azevedo et al., 2006). H. pylori surface adhesion and microcolony formation was first negatively associated with the concentration of supplemented fetal bovine serum (FBS); serum commonly promotes planktonic growth but inhibits surfaces adherence (Williams et al., 2008). It remains elusive which factors of serum impact H. pylori surface adhesion as FBS is an undefined medium with a non-homogeneous mix of growth factors (Zheng et al., 2006). Interestingly, H. pylori adhesion on gastric epithelial cell surface does not rely on the presence of FBS, suggesting that this bacterium may utilize specialized mechanism for surface attachment (Yonezawa et al., 2017, Senkovich et al., 2011, Lim et al., 2003, Hathroubi et al., 2020). Furthermore, studies have shown that adhesion on various surfaces directly affected the biomass of mature biofilms, a process that is independent of media components (Windham et al., 2018). H. pylori surface adhesion is also strain dependent (Wong et al., 2016), a variation that is potentially due to the heterogenicity of regulatory proteins and outer membrane proteins (OMPs) which are predicted to play a critical role in the initial adhesion step (Servetas et al., 2018, Wong et al., 2016). We will focus on discussing recent findings that have implicated flagella and OMPs as necessary components in the adherence process.
H. pylori flagella play important roles in adherence and subsequent biofilm formation. They are made up of four primary components; basal body, hook, filament, and sheath (Gu et al., 2017). Flagella are typically associated with H. pylori motility but have been recently discovered to be involved in promoting surface adhesion and maintaining biofilm architecture (Hathroubi et al., 2020). Motility itself is an essential factor required for H. pylori to initiate biofilm (Wong et al., 2016). More insight was provided for this observation by examining abiotic biofilm formation of strains that were non-motile but either retained flagella (Fla+ Mot-) due to deletion of a flagellar basal body gene motB, or lost flagella (Fla- Mot-) due to deletion of the flagellar basal body gene fliM (Hathroubi et al., 2018). More biofilm biomass was accumulated in the Fla+ mutant as compared with the Fla- strain. Flagella filaments, furthermore, were visible in the biofilm, and appeared to be forming a matrix. Fla+ Mot- strains exhibited initial attachment defects on gastric cell surfaces (Hathroubi et al., 2020). These results suggest that motility is likely involved in the attachment phase on diverse surfaces, and the presence of flagella is required for H. pylori biofilm formation.
Another type of molecule shown to contribute to adherence are H. pylori outer membrane proteins (OMPs), which can be on the cell surface or present in outer membrane vesicle (OMVs) along with virulence factors and eDNA (Olofsson et al., 2010 and Grande et al., 2015). OMPs play important roles in bacterial environmental adaptation and modulation of life cycle phases, including structure maintenance, substance transportation, and microbial-host interaction (Qiao et al., 2014, Egan, 2018).
H. pylori has more than 60 OMPs coding genes (Alm et al., 2000), but not all the OMP’s functions are understood (Servetas et al., 2018). Beyond inducing the pro-inflammatory responses, some OMPs were also found to promote multiple processes of H. pylori biofilm formation, one of which is to promote surface adhesion as discussed (Yonezawa et al., 2011, Tamrakar et al., 2021). H. pylori OMPs facilitate both cell-to-cell and cell-to-abiotic surface adhesion in biofilms, based on the observation of OMV localization in H. pylori biofilms via scanning electron microscopy (SEM) imaging (Yonezawa et al., 2009).
A common feature of H. pylori OMPs is anti-parallel β sheets that compose a β-barrel, highly stable pore-like structure; transmembrane domains of these proteins interact with host cell receptors (Tamrakar et al., 2021), potentially indicating these OMPs may promote bacterial cell-cell and bacterial-host connections.
One family of H. pylori OMPs is the Hom family, a group of four proteins encoded by the following genes: homA, homB, homC and homD (Alm et al., 2000). These proteins have been specifically utilized as a peptic ulcer disease marker (Tamrakar et al., 2021). Interestingly, homA and homB were found to contribute H. pylori biofilm formation as well (Servetas et al., 2018), indicating the potential association between H. pylori biofilm and relative pathology. The outer membrane protein homB (J99, jhp0870; G27 HPG27_667), was recently associated with biofilm formation (Servetas et al., 2018). This protein is interesting as it has been proposed as a biomarker of peptic ulcer disease (Oleastro et al., 2006) and gastric cancer (Talebi et al., 2011). H. pylori upregulates homB transcription via ArsRS, a two-component system, in the initial adherence and assembly phases of biofilm growth, but then levels fall back to those observed in planktonic cells after 72 hours of incubation (Servetas et al., 2018). This variation suggests the importance of HomB during the initial adhesion and later for the next biofilm assembly stages.
HomA and HomB are composed mainly of β-sheets with cysteine resides on surface loops that help to form homodimers and indicating they are potentially key to aggregation and biofilm formation (Tamrakar et al., 2021). Various studies demonstrated that HomB is negatively regulated by a two-component system, ArsRS system (Windham et al., 2018 and Servetas et al., 2018, Hathroubi et al., 2018). Other Hom family members, the homD and homC genes, are both upregulated during H. pylori biofilm formation (Hathroubi et al., 2018). Polymorphism of HomC have been linked to varied levels of biofilm formation in different H. pylori strains (Kim et al., 2016). These findings suggest Hom family OMPs are commonly involved into in the initiation steps of biofilm formation.
The outer membrane protein autotransporter is also likely play a role in regulating H. pylori biofilm formation. An uncharacterized autotransporter, paralogous to VacA, vlpC, was found to cause a defect of H. pylori biofilm formation if disrupted (Alm et al., 2000, Hathroubi et al., 2020). Specifically, this mutant was unable to form mature biofilms. vlpC has been upregulated in some biofilms, further supporting this factor is important for H. pylori biofilm formation (Hathroubi et al., 2018, Hathroubi et al., 2020)
A group of highly conserved laminin binding proteins of another OMP family, called Hop, has also been shown to be involved in H. pylori biofilm regulation as well. AlpB, a Hop family member, was implicated in biofilm formation and antibiotic resistance, since the genetic deletion of alpB caused less H. pylori biofilm formation (Senkovich et al., 2011, Yonezawa et al., 2017). Since AlpB is highly conserved among H. pylori strains, it has recently been identified and investigated as a therapeutic target to eradicate H. pylori biofilm (Xiao et al., 2022).
These findings highlight the role played aby OMPs and flagella at this stage, while also emphasizing that there is still much to be discovered.
Assembly
After surface adhesion, H. pylori starts forming microcolonies or aggregates that are recognized as the pre-mature form of biofilm (Shen et al., 2020, Hathroubi et al., 2020, Hathroubi et al., 2018). Multicellular aggregates have been observed to be formed by different strains in in vitro conditions within hours (Krzyżek et al., 2021) more complex structures as early as one day of incubation (Hathroubi et al., 2020, Hathroubi et al., 2018). H. pylori biofilms formation steps have been characterized using confocal microscopy. This work showed that H. pylori strain G27 assembles biofilms initially at the liquid air interface at 24 hours, then assembles aggregates both at the liquid air interface and under the liquid air interface as the biofilm assembled; the distribution of EPS, visualized by staining, paralleled this growth trend (Windham et al., 2018). SEM further revealed that flagella play a critical role in maintaining H. pylori biofilm structure, as discussed above (Hathroubi et al., 2020 and Hathroubi et al., 2018). Without flagella, H. pylori biofilms were slowly assembled (Ratthawongjirakul et al., 2016). Comparative genomics studies further demonstrated H. pylori biofilm assemble at rates that are similar among different strains when calculating cumulative frequency and rate of formation (Wong et al., 2016). Additionally, biofilm assembly is not significantly impacted by in vitro conditions, such as serial passaging, nutrient compositions, culturing conditions (Windham et al., 2018).
Mature Phase
The maximum biofilm mass can be observed after 3 days in vitro incubation (Hathroubi et al., 2020 and Luo et al., 2021), and can last up to 7-days in different culture conditions (Ratthawongjirakul et al., 2016, Bugli et al., 2016, Windham et al., 2018, Wong et al., 2016). Comparing biofilm growth on the surface of polystyrene plates (hydrophobic surface) that were pre-coated with poly-D-lysine (hydrophilic and positive charged) and tissues culture treated polystyrene (hydrophilic, negative charge) revealed that optimal biofilm growth is not solely dependent on surfaces being ionic; tissue culture treated and negatively charged surfaces significantly promotes biofilm growth (Windham et al. 2018). A special feature of H. pylori biofilms observed in SEM images are flagellar filaments which were discovered to promote surface cohesion and cell-to-cell connections as mentioned above, together with pili formations sustain the biofilm structural integrity on both abiotic and biotic surfaces (Hathroubi et al., 2018 and Hathroubi et al., 2020).
In the meantime, different H. pylori strains and incubation conditions can differentially impact biofilm formation kinetics. H. pylori strains with strong and poor biofilm forming abilities in tissue culture plates had consistent biomass accumulation rates during the intermediate assembly phases but had a variant cumulative biomass at the mature phase after 7 days of growth (Wong et al., 2016). In another study, H. pylori SS1 strain produced robust biofilms in relatively low FBS conditions after 3 days of growth on polystyrene plates, with most biofilm cells (~80%) being coccoid shaped (Hathroubi et al., 2018). Interestingly, H. pylori G27 strain did not rely on low-serum conditions, as biofilm formation was not impaired even at standard culture media, with 10% FBS and produced biofilms with similar morphological features as SS1 (Hathroubi et al., 2020).
In mature biofilms grown on abiotic surfaces, most cells are coccoid shaped (0.4-0.6 um long) with a minority of rod-bacillus (2-3um long) shape (Hathroubi et al., 2020). The coccoid form of H. pylori was proposed as a response to the environmental stressors, but the underlying mechanism for this morphology is not fully characterized (Kadkhodaei et al., 2020). A recent study showed that these coccoid cells maintained their membrane integrity and metabolism for up to 70 hours of incubation, which strongly suggests that they are viable dormant bacteria (El Mortaji et al., 2020). A morpho-structural analysis of H. pylori biofilms revealed that strongest biofilm producing cells show a dominance of coccoid forms unlike weak biofilm producing cells presented rod-shaped forms that were dominant in mature biofilms (Krzyżek et al., 2022).
Interestingly, H. pylori in the coccoid morphology is more tolerant to antibiotic exposure (Krzyżek et al., 2021) which aligns with H. pylori biofilms strong tolerance to antibiotics (Hathroubi et al., 2018 and Hathroubi et al., 2020). Viability staining experiments with biofilms grown on abiotic surfaces suggest that live cells and dead cells compose matured H. pylori biofilms (Hathroubi et al., 2020). Transcriptomic experiments show that biofilm cells are less metabolically active than planktonic cells due to the downregulation of multiple metabolic genes, such as atpC, atpE, and nifU (Hathroubi et al., 2018). Gastric epithelial cell lines, such as AGS, have been developed to study H. pylori biofilm formation on biotic surfaces (Hathroubi et al., 2020, Yonezawa et al., 2009). After co-incubating H. pylori and AGS cells for days, H. pylori biofilms were observed on the surfaces and between conjunctions AGS cells (Hathroubi et al., 2020, Cárdenas-Mondragón et al., 2016). Interestingly, most biofilm cells were spiral/rod shaped, a different outcome than what was observed in biofilms grown on abiotic surfaces (Hathroubi et al., 2020). Other cell lines have been employed as well, particularly mucin producing cells, like MKN-45 cells, which may present a more natural in vivo-like state similar to niches in the host. On the MKN-45 cell line, most of the biofilm cells primarily exhibited the coccoid morphology (Attaran et al., 2021), suggesting this cell line can be used as a model to study the effects of mucin on H. pylori biofilm formation. Further studies are necessary to dissect whether different incubation conditions, such as serum concentration and incubation period may modulate H. pylori biofilm features.
Figure 1.
SEM images of mature H. pylori biofilms grown on abiotic surfaces. (A) Mature H. pylori biofilms contain a mixed population of mostly cocoid shaped cells with a minority of spiral shaped cells, (B) Higher resolution image showing spiral shaped cells (white arrow) and coccoid cells aggregating via the flagella (red arrow).
Figure 1.
SEM images of mature H. pylori biofilms grown on abiotic surfaces. (A) Mature H. pylori biofilms contain a mixed population of mostly cocoid shaped cells with a minority of spiral shaped cells, (B) Higher resolution image showing spiral shaped cells (white arrow) and coccoid cells aggregating via the flagella (red arrow).

Dispersion
Like other bacterial biofilms, H. pylori biofilms disperse after reaching optimal growth, indicated by a decrease in crystal violet staining after maximum growth has been reached (Hathroubi et al., 2018, Hathroubi et al., 2020, Krzyżek et al., 2021). Little is known about the signals that lead to H. pylori biofilm dispersal, but some evidence suggests that H. pylori utilizes a quorum sensing molecule, AI-2, as a signaling molecule to regulate biofilm generation and dispersion (Anderson et al., 2015). AI-2 was initially recognized as a chemorepellent of H. pylori sensed by chemoreceptor TlpB (Rader et al., 2011), and this molecule can be expressed by H. pylori through luxS gene in a cell density-dependent manner (Forsyth et al., 2000, Lee et al., 2006), suggesting that H. pylori can efficiently control local density through AI-2 secretion. A later study suggested that AI-2 promoted H. pylori biofilm dispersion, as genetic deletion of the luxS in H. pylori significantly promoted its biofilm formation in comparison to isogeneic WT strain through the lacunarity and fractal dimension analysis (Anderson et al., 2015). The chemotaxis system, in another aspect, was suggested to facilitate H. pylori biofilm dispersion by sensing and responding to AI-2, since chemotactic histidine kinase deficient mutant ∆cheA exhibited similar biofilm phenotype as the ∆luxS mutant (Anderson et al., 2015). Further research is required to decipher the mechanism of how H. pylori regulates biofilm maturation and dispersion.
Scheme 1.
Helicobacter pylori Biofilm Lifecycle. H. pylori adheres to both abiotic and biotic surfaces, where it forms microcolonies that susbquently assemble into mature biofilms characterized by the presence of extracellular polymeric susbtances (EPS). Dispersion allows bacteria to colonizes new niches.
Scheme 1.
Helicobacter pylori Biofilm Lifecycle. H. pylori adheres to both abiotic and biotic surfaces, where it forms microcolonies that susbquently assemble into mature biofilms characterized by the presence of extracellular polymeric susbtances (EPS). Dispersion allows bacteria to colonizes new niches.
H. pylori clinical treatment strategies and the increasing prevalence of antibiotic resistance.
The currently recommended H. pylori infection treatment are quadruple therapies that consists of proton pump inhibitor (PPI), bismuth and two additional different antibiotics, including clarithromycin, metronidazole, levofloxacin, or amoxicillin (Liu et al., 2022). However, this classical therapeutic strategy has been being less effective due to the continuing rise of antibiotic resistance. For example, in 2016 a national consensus on Chinese management of H. pylori infections reported that metronidazole, levofloxacin, and clarithromycin resistance was 40-70%, 20-50% and 20-50% respectively (Liu et al., 2018). Similarly, the elevation of antibiotic resistance was also noticed in other countries, like Indonesia, that apply the triple-therapy approach consisting of PPI and two antibiotics, (Malfertheiner et al., 2017). The prevalence of resistance to recommended antibiotics metronidazole and levofloxacin increased to 46.7% and 31.2%, respectively; resistance to non-recommended antibiotics remained low however, including amoxicillin (5.2%), tetracycline (2.6%) and clarithromycin (9.1%) (Miftahussurur et al., 2016). In 2020, a case study reported that triple therapies in Indonesia were further decreased to only 67.6% efficient (Fauzia et al., 2020).
To avoid potential therapy failure caused by antibiotic resistance, clinicians have proposed using a tailored treatment approach based on antibiotics susceptibility tests and localized resistance (Fauzia et al., 2020, Pan et al., 2020, Fallone et al., 2016, de Palma et al., 2017 and Li et al., 2022). A clinical study that analyzed the failure of H. pylori treatment revealed that patients who were not cured were often infected by diverse H. pylori strains that were resistant to different antibiotics (Mascellino et al., 2018). Clarithromycin resistance is attributed to mutations in the 23S rRNA (Redondo et al., 2018); metronidazole resistance was associated to the mutations in rdxA and frxA loci (Kim and Lee et al., 2017); levofloxacin resistance was caused by gyrA and gyrB mutations (Miftahussurur et al., 2016). These mutations are naturally occurring, but increased prevalence in the population can occur by exposing strains to sub-MIC levels of antibiotics, such as levofloxacin (Hanafi et al., 2016). Another case study indicated that the tailored treatment approach is necessary and effective in addition to traditional treatment procedures. A total of 112 isolated H. pylori strains were genotyped from the H. pylori infection prevalent regions that applied quadruple treatment. With the supplementation of the tailored treatment, the dual resistance to metronidazole and levofloxacin was observed in 20.5% of tested strains and triple resistance to metronidazole, clarithromycin and levofloxacin was observed in only ~7% of strains (Liu et al., 2022). The effectiveness of tailored therapies was also evaluated in another clinical trial study in comparison to the traditional bismuth quadruple therapy, and it was demonstrated that the tailored bismuth/quadruple therapy was more effective (Pan et al., 2020). Intriguingly, another case study examined 101 clinical H. pylori isolates from Indonesian patients with gastritis (91%), peptic ulcer disease (8.9%) and gastric cancer (1%) and discovered that 93% of the isolates formed biofilms (Fauzia et al., 2020). These studies strongly suggest that biofilm formation may play a vital role in facilitating H. pylori to acquire high antibiotic tolerance, therefore the eradication of H. pylori biofilm is likely a key process for clinical therapy. Nevertheless, there are challenges in clinal therapies: (1) planktonic susceptibility of minimal inhibitory concentration (MIC) may not be a reliable indicator of Minimal Biofilm Eradication Concentration (MBEC) with certain antibiotics (Fauzia et al., 2020; Attaran et al., 2017); (2) isolating clinical strains from infected patients are required the acquisition of gastric biopsies is an invasive procedure (Li et al., 2022). Therefore, it would be very interesting to understand if targeting biofilm formation would enhance H. pylori treatment.
Approaches utilized in H. pylori antibiotic resistance detection and prediction.
Due to the global increase in antibiotic resistance with H. pylori, efficient antibiotic susceptibility examinations would be expected for appropriate diagnostic and treatment. Currently, two major types of techniques are utilized today, either bacterial viability-based or molecular based technique.
Bacterial viability-based techniques are the standard approach to determine bacterial antibiotic susceptibility and has been utilized to track increasing antibiotic resistance (Midolo et al., 1997, Fauzia et al., 2020), by measuring bacterial viability under exposure to a certain type and amount of antibiotic. Such approaches are further divided into agar or broth dilution methods, Epsilometer test (E-test) methods (Glupczynski et al., 1991), or disk diffusion methods (Tang et al., 2020). These techniques are all capable of quantitatively determining the minimum concentration of an antibiotic that kills H. pylori (Mishra et al., 2006). Different methods have specific advantages. E-tests and disk diffusion assays are not a ‘one size fits all’ approach since the differences in susceptibility to amoxicillin, tetracycline, and furazolidone were observed between the disk diffusion method and E-test method (Tang et al., 2020). For example, the E-test method is easy to apply and time friendly (Midolo et al., 1997), while the H. pylori dilution method allows several stains to be tested simultaneously.
As various antibiotic resistance mechanisms have been characterized and the genetic elements have been identified using molecular-based approaches (Saruuljavkhlan & Yamaoka, 2021), these discoveries promote the evolution of more rapid and cost friendly molecular based methods to detect presence of responsible resistant genetic elements and susceptible elements to predict possible antibiotic resistant phenotypes (Tshibangu-Kabamba et al., 2020, Smith & Pellicano, 2019). PCR-based genetic amplification technique and Sanger sequencing approaches together are intensively developed and applied to achieve such goals; these approaches have several advantages including being easily reproducible and time efficient in comparison to traditional bacterial viability-based methods (Mégraud et al., 2007). More importantly, these techniques can be applied directly on bacteria that have not been cultured or are of low abundance including various clinical isolates from gastric tissue or gastric juice (Van Doorn et al., 2001, Schabereiter-Gurtner et al., 2004, Mitui et al., 2014, Nishizawa & Suzuki, 2014). However, this approach has limitations and is only reliable to predict certain types of antibiotics whose resistant mechanism has been specifically characterized, such as clarithromycin and tetracycline, but not for those antibiotics whose anti-mechanism is diversified, such as metronidazole and amoxicillin. To overcome such limitations, next generation sequencing (NGS) technologies have been developed as an efficient tool to detect and predict all potential antibiotic resistance mutations in a bacterial sample (Vital et al., 2022). This type of approach consists of DNA extraction from a given bacterial sample that undergoes whole genome sequencing (WGS) (Fauzia et al., 2021). There are several advantages of this approach compared to the PCR-based molecular approach. With the growing of whole microbial genome data sets, a pan-genome-based machine learning approach was recently developed to predict antimicrobial resistance activities in some bacteria, including Escherichia coli (Her & Wu, 2018). This approach uses written algorithms to predict whether a specific stain is resistant to antibiotic drugs by comparing its genome against the accessory part of the pan-genome, to yield the gene clusters that are most crucial to antimicrobial resistance activities in E. coli. A limitation of this approach is that we may not yet know all antibiotic resistance alleles. Currently, this approach has not yet applied in examining H. pylori, but it seems to be a promising one.
Mechanisms of H. pylori biofilm-promoted antibiotic resistance.
Biofilm formation may play a significant role in facilitating H. pylori antibiotic tolerance (Tshibangu-Kabamba & Yamaoka, 2021). A phenotype of tolerance manifests in that the antibiotic MIC for planktonic H. pylori does not accurately reflect the concentration needed to eradicate H. pylori biofilm cells. For example, a clinical study compared antibiotic susceptibility of H. pylori isolates between the planktonic and biofilm growth and found that H. pylori biofilms was more capable of tolerating various antibiotics relative to planktonic H. pylori, including up to 1000-fold with amoxicillin, 31.25-fold with clarithromycin, 16-fold with levofloxacin, and 8-fold with metronidazole (Fauzia et al., 2020). H. pylori biofilms have exhibited several advantages in facilitating antibiotic tolerance. Studies have proposed the correlation between high biofilm formation capacity in H. pylori and the tolerance to clarithromycin, but not however, metronidazole or levofloxacin (Krzyzek et al., 2022). While the reason for the high tolerance of H. pylori biofilms is not yet fully understood, several ideas have been proposed including that bacterial cells are protected by the biofilm structure; conjugated bacterial cells within the biofilm increased the chance of genetic exchange. Below we dissect recent mechanisms of antibiotic tolerance employed by H. pylori biofilms.
Extracellular polymeric substance matrix reduces the efficacy of antibiotics
H. pylori biofilms are encased in an extracellular polymeric substance (EPS) matrix that maintains the structural integrity of the biofilm, promotes adhesion, and facilitates cell-to-cell interactions (Li et al., 2019). Proteins, polysaccharides and eDNA were confirmed to compose the extracellular polymeric substance matrix in H. pylori biofilms (Hathroubi et al., 2018). Immunofluorescence assays with probes specific for proteins, eDNA and polysaccharides show that EPS distribution depends on cell density within the biofilm (Windham et al., 2018). Polysaccharides in the EPS can be stained with FITC-conA which targets mannose groups in polysaccharides. The green fluorescence can be used to visualize the EPS matrix in H. pylori biofilms with Confocal Laser Scanning Microscopy (CLSM) (Shen et al., 2020 and Li et al., 2019). The film tracer Sypro Ruby stain targets proteins in the EPS and can also be visualized using CLSM (Hathroubi et al., 2018 and Windham et al., 2018). EPS eDNA in H. pylori biofilms can be stained and visualized via CLSM using BOBO-3 (Hathroubi et al., 2018) and propidium iodide (Windham et al., 2018). Enzymatic assays indicate that proteins play vital role in H. pylori EPS as proteinase K treatment significantly cause dispersion of H. pylori biofilms and reduced antibiotic tolerance (Hathroubi et al., 2018 and Windham et al., 2018). While eDNA and polysaccharides also compose EPS structures, they are predicted to play minor roles compared to proteins, based on the observation that DNase I and sodium periodate treatment targeting the eDNA and polysaccharide respectively, did not cause significant H. pylori biofilm reduction (Windham et al., 2018, Hathroubi et al., 2018).
In addition to sustaining structural integrity, the EPS may reduce the efficacy of drugs from reaching the interior of the biofilm. EPS itself is minimally affected during antibiotic exposure (Li et al., 2019), supporting the idea that antibiotic treatment does not eradicate H. pylori biofilms. Removal of proteins, however, does sensitize H. pylori in biofilms to clarithromycin, although it was not demonstrated whether this is EPS or surface protein removal (Hathroubi et al 2020). Therefore, the disruption of EPS of H. pylori biofilm may be a highly significant target to effectively eradicate this bacterium (Li et al., 2019).
Coccoid Cellular morphology
Compared to spiral shape that is commonly observed in planktonic H. pylori cells, coccoid cells are more commonly found in H. pylori biofilm (Hathroubi et al., 2018, Hathroubi et al., 2020). The coccoid cellular shape was recognized to be dormant state of H. pylori that contributes to antibiotic resistance and disease induction (Reshetnyak et al., 2017, Kadkhodaei et al., 2020). H. pylori biofilms, like other bacteria, can sustain the slow growth state (Hathroubi et al., 2018), and promote antibiotic tolerance that specifically target active phase bacterium (Harris et al., 2000, Ikeda et al., 1990). Prior research has shown that significant cell wall alterations occur when H. pylori is transitioning to the coccoid morphology (Costa et al., 1999) and has been associated with biofilm growth and antibiotic tolerance (Kadkhodaei et al. 2020).
A couple of genes that modify H. pylori’s cell wall have been documented to be upregulated in H. pylori biofilms and may contribute to the coccoid form and/or antibiotic tolerance. For example, UppS, a putative undecaprenyl pyrophosphate synthase, facilitates H. pylori cell wall peptidoglycan modification (Kuo et al., 2007). Transposon inserted of uppS, resulted in a defective biofilm formation (Hathroubi et al., 2020). Some naturally occurring cell-wall related mutations may be beneficial for developing antibiotic resistance. For example, recent studies found ethoxzolamide, clinically used sulphonamide drug, can block cell wall synthesis by competitively inhibiting UppS (Modak et al., 2019); however, strains can be become resistant by acquiring mutations in the binding site of UppS (Rahman et al., 2020).
Another cell wall factor found to be important for maintaining H. pylori biofilm structure is peptidoglycan deacetylase (PdgA). The pgda gene was upregulated in H. pylori biofilms (Hathroubi et al., 2018), and was previously associated with host derived oxidative stress (Wang et al., 2010). Oxidative stress induces H. pylori biofilm formation (Zhao et al., 2021), which is consistent to a model that PdgA promotes H. pylori biofilm formation. In addition, PdgA may play an important role in maintaining H. pylori biofilm structure as the H. pylori ΔpgdA mutant is more susceptible to lysozyme exposure, an enzyme that cleaves the peptidoglycan of bacterial cell wall (Wang et al., 2012).
In addition, another gene hp0421, encoding cholesteryl-α-glucoside transferase , was also found to regulate cellular morphology in biofilms (Chou et al., 2017; Qaria et al., 2018). The hp0421 deletion caused defects in maintaining spiral morphology, an increase in susceptibility to antibiotics and promoted cellular aggregation to form pronounced biofilms faster than the wild-type controls (Qaria et al., 2018) further supporting the important role of coccoid morphology in biofilms. In conclusion, genes that have been implicated in regulating H. pylori morphologies and is synchronous with affecting biofilm phenotypes and antibiotic tolerance reveal a key topic that should be investigated to further decode H. pylori biofilms.
Downregulated Metabolism in Biofilms
Growing bacterial cells are more easily targeted by certain types of antibiotics, such as ampicillin, that is selected as an essential component of triple-therapy applications for H. pylori treatment (Marcus et al., 2012). Recently it has been revealed that H. pylori reduces its metabolic activities in the biofilm to mitigate such detrimental effects, along with the trend shifting to coccoid cellular morphology (Hathroubi et al., 2018). A recent clinical study found a positive correlation between strong biofilm former and a general decrease in metabolic rate (Wong et la., 2018). This observation is supported by another H. pylori transcriptomic study that suggests biofilm cells are less metabolically active than planktonic cells due to the downregulation of metabolic genes (Hathroubi et al., 2018). Interestingly, H. pylori is also able to upregulate specific metabolic enzymes to resist certain natural substrates, functionally as antibiotics. For example, Combretum mole extracts, an acetone-containing plant commonly consumed in South Africa to alleviate gastric illness, have bactericidal effects on H. pylori (Njume et al., 2011). To tolerate acetone exposure, acetone carboxylase gene acxA is upregulated in the H. pylori biofilm, indicating the acetone carboxylases is expressed to potentially degrade acetone during gastric colonization. Additionally, acxA deletion resulted in a significant biofilm defect (Hathroubi et al., 2020); the acxA gene is under regulation of both two-component system under the ArsRS (Loh et al., 2010) and the CrdRS (Allen et al., 2023), which are heavily involved in maintaining H. pylori biofilm and promoting gastric gland colonization (Brahmachary et al., 2008, Hathroubi et al., 2020). Both crdR and arsR regulators were found to be upregulated in biofilms (Hathroubi et al., 2018, De La Cruz et al., 2017, Servetas et al., 2016); crdR was found to be upregulated in biofilms on abiotic surfaces (Hathroubi et al., 2018 and De La Cruz et al., 2017) and upon adherence to AGS cells (De la Cruz et al., 2017). On the other hand, arsR was found to be upregulated in strain 26695 biofilms grown on abiotic surfaces and AGS (De la Cruz et al., 2017). These combined findings suggest that the acxA gene is mandatorily expressed and essential to maintain certain functions of H. pylori biofilm including protecting H. pylori in the host from acetone degradation.
Efflux pumps involved drug external transportation
Efflux pumps are commonly located on the H. pylori cell membrane and facilitate the multiple drugs external transportation (Raj et al., 2021). Efflux pumps have been strongly associated with antibiotic resistant strains and multidrug resistance in recent studies (Liu et al., 2022 and Yonezawa et al., 2019, Attaran et al., 2017) which indicates that they play a significant role in the antibiotic tolerance of H. pylori biofilms. Several efflux pumps coding genes, including Hp605 (hefA), Hp971 (hefD), Hp1327 (hefG), Hp1489, Hp1118, Hp1174 (gluP), HP0939, HP0497, and HP0471 (KefB), were found to be expressed in both planktonic and biofilm cells, suggesting that efflux pump is essential during H. pylori life cycles (Ge et al., 2018, Yonezawa et al., 2019, Cai et al., 2020). Recent studies further revealed that these efflux pump coding genes were significantly upregulated in biofilm to facilitate H. pylori antibiotics tolerance (Cai et al., 2020). HPG27_715 (a MATE-family uncharacterized efflux pump), Hp1118, gluP, HP1165 (associated with tetracycline resistance), hefA were significantly upregulated in biofilms relative to planktonic cells (Hathroubi et al., 2020, Ge et al., 2018, Attaran et al., 2017). hefA (Yonezawa et al., 2019 and Attaran et al., 2017), hefD, hefG and HP1489 were found to be particularly upregulated in biofilms from a clarithromycin resistant strain TK1402 (Yonezawa et al., 2019). gluP expression was found to be regulated by H. pylori stringent response and genetic deletions of gluP cause a biofilm defect and increased susceptibility to different types of antibiotics (Ge et al., 2018). Additionally, genetic deletions in HP0939, HP0497, and KefB also conferred with a biofilm defect (Cai et al., 2020). hefD and hefA have both recently been associated with multidrug resistance in clinical H. pylori strains isolated from Nigeria while no association with hefG was detected (Jolaiya et al., 2020). Cumulatively, these findings support the perspective that H. pylori utilizes biofilm growth to survive under antibiotic exposure and efflux pumps are a key contributor.
Anti-biofilm strategies
Since chronic infection with H. pylori causes various gastric diseases, approaches are being developed to efficiently eradicate this bacterium. Here, we summarize several approaches based on the anti-biofilm treatments including synthetic compounds, natural compounds, and small molecule drugs.
Antimicrobial peptides
Antimicrobial peptides (AMPs) are promising alternatives to antibiotics for combating biofilm infections. One of the advantages of using AMPs is that these molecules are also less likely to induce resistance in bacteria than antibiotics because they target multiple components within the bacterial cell. These small peptides can penetrate the extracellular matrix that surrounds biofilm cells and thus target the bacteria directly.
Another antimicrobial peptide was also recently investigated, Cbf-K16, Cathelicidin-like peptide which showed a good antimicrobial activity against clarithromycin- and amoxicillin- resistant H. pylori in vitro and in vivo (Jiang et al., 2019). In mouse gastritis model Cbf-K16 demonstrated a 3.9-log10 reduction of bacterial counts in stomach tissues compared to untreated mice group (Jing et al., 2019). Interestingly, treatments with Cbf-K16 significantly downregulated the expression levels of the adhesion-associated genes alpA and alpB mRNA, both factors play a role in H. pylori adhesion and biofilms as mentioned above (Senkovich et al., 2011, Yonezawa et al., 2017, Jiang et al., 2019).
The antimicrobial peptide MSI-78A, also known as Pexiganan, is a 22-amino acid peptide Magainin-2 analogue was reported to have antibacterial activity in solution (Zhang et al., 2015, Parreira et al., 2019). When surface grafted MSI_78A still demonstrated activity with a high bacterial eradication rate (>90% after 2h) thus not able to proliferate and establish biofilms (Parreira et al., 2019).
Several synthetic peptides were also applied and have been shown to promote biofilm dispersion in H. pylori, individually or synergistically with host antimicrobial peptides (Windham et al., 2018). For instance, when H. pylori biofilms were treated with synthetic peptides IDR-1018 and DJK-5, it became more susceptible to the host derived anti-microbial peptides (Windham et al., 2018). In addition, DJK-5 is a synthetic short D-enantiomeric peptide designed to be resistant bacterial proteases (de la Fuente-Núñezet al., 2015) and IDR-1018 was designed by altering bactenecin from bovine neutrophils (Mansour et al., 2015). Both DJK-5 and IDR-1018 are capable of degrading a second messenger nucleotide, a stringent response molecule, called (p)ppGpp (de la Fuente-Núñezet al., 2015 and Mansour et al., 2015). Prior in vitro studies from several H. pylori strains (J99, 26695 and G27) suggested that H. pylori utilizes a stringent response at low pH or with poor nutrients to produce significant amounts of ppGpp (Wells et al, 2006). H. pylori contains an enzyme called SpoT, a (p)ppGpp synthase and hydrolase, whose genetic deletion causes a defective biofilm phenotype and an increased susceptibility to antibiotics (Ge et al., 2018). DJK-5 and IDR-1018 were tested on H. pylori biofilms and were observed to not affect viability of planktonic bacterial viability; biofilm assembly, however, was inhibited only by DJK-5 (dose dependent). In contrast, IDR-1018 reduced mature H. pylori biofilms without affecting the bacterial viability within the biofilm matrix (Windham et al., 2018). These findings suggest that synthetic cationic peptides specifically target H. pylori in the form of biofilms and that H. pylori utilizes mechanisms in biofilms homologous to other bacterial species affected by the same peptides (Windham et al., 2018).
Extracts from Natural Resources
Extractions from natural resources such as plants and other bacteria are commonly applied to treat various microbial infections, including H. pylori. Some extractions have been found to be particularly effective in eradicating H. pylori by specifically targeting biofilm stability.
Probiotics can inhibit bacterial biofilms and thus play an auxiliary role in bacterial antibiotic therapy. As documented, the effects of different probiotic strains may play a varied role in restricting certain bacterial biofilms, including H. pylori biofilm. Probiotic Lactobacillus fermentum UCO-979C was previously found to play a role in inhibiting H. pylori biofilm formation (Salas-Jara et al., 2016). Furthermore, another microbial study found that Lactobacillus plantarum LN66 cell-free supernatant (CFS) can weaken H. pylori biofilm formation, an effect monitored by SEM and confocal laser scanning microscopy (CLSM) (Ji & Yang, 2021). Probiotics combined with other antibiotics was found to increase treatment efficacy for levofloxacin as LN66 CFS facilitate this antibiotic function to inhibit EPS secretion (Salas-Jara et al., 2016, Jin and Yang 2021). Another intriguing finding is armeniaspirols, which is a novel class of natural products isolated from Streptomyces armeniacus previously identified as antibacterial agents against Gram-positive pathogens (Dufour et al., 2012). Armeniaspirol A (ARM1) exhibited potent antibacterial activity against H. pylori as well by inhibiting H. pylori biofilm formation in a dose-dependent manner. In a mouse model to study multidrug-resistant H. pylori, dual therapy with ARM1 and omeprazole showed efficient killing efficacy, comparable to the standard triple therapy, and induced negligible toxicity against normal tissues (Jia et al., 2022). Moreover, at acidic pH 2.5, ARM1 exhibited a much more potent anti-H. pylori activity than metronidazole (Jia et al., 2022). All these advantages promote the possibility of ARM1 being used in a clinical application.
Plants are another major resource that organic products are extracted from to treat bacterial infections. A variety of materials have been found to efficiently restrict H. pylori infection. For example, Antractylodes lancea volatile oils were recently found to inhibit H. pylori biofilm formation. This oil complex also exhibits a robust ability to reduce H. pylori virulence factor CagA translocation into host cells, a finding observed in a cell culture infection model (Yu et al., 2019). Additional screenings were applied to search for natural molecules to target H. pylori biofilm stability. Phytochemicals from Acorus calamus, Colocasia esculenta Vitex trifolia, Azadirachta indica A. Juss exhibited a significant effect on inhibiting H. pylori biofilm formation as well (Marina et al., 2022, Prasad et al., 2019). Among screening tests, Acorus calamus exhibited the highest H. pylori anti-biofilm activity via a dose-dependent pattern (Prasad et al., 2019). Phytochemicals from the neem tree (Azadirachta indica A. Juss) were also previously shown to have bactericidal properties and several other Neem tree phytochemicals (nimbolide, azadirachtin, gedunin) and were tested for toxicity towards H. pylori but only Nimbolide was found to kill both planktonic and biofilm H. pylori without having hemolytic activity; Nimbolide was effective towards the nine strains of H. pylori tested in a time and dose dependent manner under various stressful growth conditions and metabolic activities (Marina et al., 2022). Dihydroatanshinone I, a natural herbal compound, is another agent that clearly inhibits H. pylori biofilm in both in vitro and in vivo studies when combined with omeprazole as a dual therapy, even more efficiently compared to the standard triple therapy approach that includes metronidazole; more interestingly, this compound exhibited negligible toxicity against normal tissues, indicating the potential in its clinical application (Luo et al., 2021). Extracts from hibiscus flowers (Hibiscus rosa sinensis L. flower) also showed properties of inhibiting biofilms and bactericidal effects on drug resistant H. pylori strains (Trung et al., 2020). Alginate lyases, a compound found naturally in brown algae that degrades the EPS was found to enhance the efficacy of clarithromycin when both components are synergistically used to treat biofilms (Bugli et al., 2016). These recent findings present promising possibilities of discovering compounds in nature that are effective at killing H. pylori even in biofilm forms.
Small Molecule Drug and Nanodrugs
Various small molecule-based compounds that facilitate traditionally applied antibiotics, have been found to be effective at treating bacterial infections. These compounds include both organic and inorganic monomers or polymers that target bacterial essential enzymes, pathways, or structure. For example, carvacrol and thymol were found to inhibit H. pylori biofilms by inhibiting an enzyme required for biofilm growth, carbonic anhydrase (Grande et al., 2021). Lipid polymer nanoparticles can eradicate H. pylori biofilm by enhancing the encapsulation of a given antibiotic, such as clarithromycin, to reduce biofilm viability and structural integrity more efficiently via bypassing the mucus layer and the EPS of the H. pylori biofilm (Li et al., 2019). A following study further found that the function of N-acylhomoserine lactonase silver nanoparticles (aka nanodrugs) in inhibiting H. pylori quorum sensing system, potentially combats H. pylori biofilm formation (Gopalakrishnan et al., 2020). Additionally, synthesized silver ultra-nano clusters (SUNCs) in another study were found to inhibit H. pylori biofilm formation when synergized with other antibiotics, like metronidazole (Grande et al., 2020; Huang et al., 2022). Nanodrugs are slightly negative charged/ hydrophilic oral drugs made of berberine derivatives and rhamnolipids (RHL) that penetrate the mucus layer and effectively clear H. pylori biofilms in vitro and in vivo (Shen et al., 2020, Li et al., 2019); RHL is a biosurfactant composed of di and mono-rhamnose sugars attached to fatty acids produced by Pseudomonas aeruginosa (Li et al., 2019) and berberine is a quaternary ammonium alkaloid isolate from Coptis chinensis that is proposed to enhance the efficacy of triple therapy for H. pylori infections (Shen et al., 2020). Nanoparticles modified with mannose were specifically found to be effective towards multi-drug resistant H. pylori and their biofilms (Arif et al., 2022). All these studies show that the combination of nanodrugs with antibiotics efficiently disrupts H. pylori biofilm and provides a feasible strategy to eradicate H. pylori infection.
Conclusion and Perspective
In conclusion, the scientific community has made considerable strides in unraveling the intricate nature of the gastric chronic pathogen, H. pylori, and its biofilm formation mechanisms. Notably, studies employing clinically isolated strains have played a crucial role in advancing our understanding and have paved the way for the development of promising biofilm-based approaches for eradicating H. pylori.
Nevertheless, there is still a need for more targeted research to comprehensively evaluate the pharmacological effects of the newly proposed treatments on both the host and the effectiveness of H. pylori eradication. These investigations should extend beyond in vitro experiments and encompass comprehensive animal models and rigorous clinical trials. By conducting such studies, we can obtain a more accurate assessment of the therapeutic potential of these proposed treatments and their impact on both the host and the pathogen.
Furthermore, it is important to explore the long-term effects of these novel approaches to ensure their safety and efficacy in real-world scenarios. Additionally, investigating potential resistance mechanisms that H. pylori may employ in response to biofilm-targeting therapies would be instrumental in designing more robust treatment strategies.
In conclusion, while significant progress has been made in understanding H. pylori biofilm formation and developing potential eradication approaches, further research is necessary to evaluate the pharmacological effects, efficacy, and safety of these treatments in animal models and clinical trials. By addressing these research gaps, we can bring us closer to achieving more effective and personalized strategies for combating H. pylori infection and its associated complications.
Author Contributions
Yasmine Elshenawi and Dr. Hu contributed equally to writing the original manuscript, conceiving content, and searching for literature references. Dr. Hathroubi coordinated to write the manuscript, provide feedback, and help draft literature review sections. Dr. Ottemann edited the manuscript. All authors have read and approved the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cellini, L., Allocati, N., Angelucci, D., Iezzi, T., Campli, E. D., Marzio, L., & Dainelli, B. Coccoid Helicobacter pylori not culturable in vitro reverts in mice. Microbiology and immunology 1994, 38(11), 843–850.
- A Noach, L.; Rolf, T.M.; Tytgat, G.N. Electron microscopic study of association between Helicobacter pylori and gastric and duodenal mucosa. J. Clin. Pathol. 1994, 47, 699–704. [Google Scholar] [CrossRef]
- Dubois, A.; Fiala, N.; Heman-Ackah, L.M.; Drazek, E.; Tarnawski, A.; Fishbein, W.N.; Perez-Perez, G.I.; Blaser, M.J. Natural gastric infection with Helicobacter pylori in monkeys: A model for spiral bacteria infection in humans. Gastroenterology 1994, 106, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P., Camargo, M. C., El-Omar, E., Liou, J. M., Peek, R., Schulz, C., ... & Suerbaum, S. Helicobacter pylori infection. Nature Reviews Disease Primers 2023, 9(1), 19.
- Labenz, J.; Borsch, G. Evidence for the essential role of Helicobacter pylori in gastric ulcer disease. Gut 1994, 35, 19–22. [Google Scholar] [CrossRef]
- Pereira, M.-I. Role ofHelicobacter pyloriin gastric mucosa-associated lymphoid tissue lymphomas. World J. Gastroenterol. 2014, 20, 684–98. [Google Scholar] [CrossRef]
- Blaser, M.J. Helicobacter pyloriPhenotypes Associated with Peptic Ulceration. Scand. J. Gastroenterol. 1994, 29, 1–5. [Google Scholar] [CrossRef]
- Asaka, M.; Kimura, T.; Kato, M.; Kudo, M.; Miki, K.; Ogoshi, K.; Kato, T.; Tatsuta, M.; Graham, D.Y. Possible role ofHelicobacter pylori infection in early gastric cancer development. Cancer 1994, 73, 2691–2694. [Google Scholar] [CrossRef]
- Veldhuyzen van Zanten SJ, Sherman PM. Helicobacter pylori infection as a cause of gastritis, duodenal ulcer, gastric cancer and nonulcer dyspepsia: a systematic overview. CMAJ. PMID: 8287340; PMCID: PMC1486230. 1994, 150(2), 177–185.
- Isaacson, P.G. Gastric Lymphoma and Helicobacter pylori. New Engl. J. Med. 1994, 330, 1310–1311. [Google Scholar] [CrossRef]
- Ma, J.; Yu, M.; Shao, Q.; Yu, X.; Zhang, C.; Zhao, J.; Yuan, L.; Qi, Y.; Hu, R.; Wei, P.; et al. Both family-basedHelicobacter pyloriinfection control and management strategy and screen-and-treat strategy are cost-effective for gastric cancer prevention. Helicobacter 2022, 27, e12911. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Yang, F.; Chi, W.; Ding, L.; Liu, T.; Zhu, F.; Ji, D.; Zhou, J.; Fang, Y.; et al. Antimicrobial resistance patterns and genetic elements associated with the antibiotic resistance of Helicobacter pylori strains from Shanghai. Gut Pathog. 2022, 14, 1–23. [Google Scholar] [CrossRef]
- Karbalaei, M.; Keikha, M.; Abadi, A.T.B. Prevalence of Primary Multidrug-resistant Helicobacter pylori in Children: A Systematic Review and Meta-analysis. Arch. Med Res. 2022, 53, 634–640. [Google Scholar] [CrossRef]
- Brown, H.; Cantrell, S.; Tang, H.; Epplein, M.; Garman, K.S. Racial Differences in Helicobacter pylori Prevalence in the US: A Systematic Review. Gastro Hep Adv. 2022, 1, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Cellini, L.; Grande, R.; Di Campli, E.; Traini, T.; Di Giulio, M.; Lannutti, S.N.; Lattanzio, R. Dynamic colonizationof Helicobacter pyloriin human gastric mucosa. Scand. J. Gastroenterol. 2008, 43, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Mackay, W.; Gribbon, L.; Barer, M.; Reid, D. Biofilms in drinking water systems: a possible reservoir for Helicobacter pylori. J. Appl. Microbiol. 1998, 85, 52S–59S. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.M.; Gerwig, G.J.; Pitman, R.S.; Potts, L.F.; Williams, N.A.; Greenman, J.; Weinzweig, I.P.; Hirst, T.R.; Millar, M.R. Biofilm formation byHelicobacter pylori. Lett. Appl. Microbiol. 1999, 28, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Carron, M.A.; Tran, V.R.; Sugawa, C.; Coticchia, J.M. Identification of Helicobacter pylori Biofilms in Human Gastric Mucosa. J. Gastrointest. Surg. 2006, 10, 712–717. [Google Scholar] [CrossRef]
- Coticchia, J.M.; Sugawa, C.; Tran, V.R.; Gurrola, J.; Kowalski, E.; Carron, M.A. Presence and Density of Helicobacter pylori Biofilms in Human Gastric Mucosa in Patients With Peptic Ulcer Disease. J. Gastrointest. Surg. 2006, 10, 883–889. [Google Scholar] [CrossRef]
- Windham, I.H.; Servetas, S.L.; Whitmire, J.M.; Pletzer, D.; Hancock, R.E.W.; Merrell, D.S. Helicobacter pylori Biofilm Formation Is Differentially Affected by Common Culture Conditions, and Proteins Play a Central Role in the Biofilm Matrix. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed]
- Grande, R.; Di Giulio, M.; Bessa, L.; Di Campli, E.; Baffoni, M.; Guarnieri, S.; Cellini, L. Extracellular DNA in Helicobacter pylori biofilm: a backstairs rumour. J. Appl. Microbiol. 2010, 110, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Chen, X.; Shen, Y.; Li, H.; Zou, Y.; Yuan, G.; Hu, P.; Hu, H. Mucus penetration enhanced lipid polymer nanoparticles improve the eradication rate of Helicobacter pylori biofilm. J. Control. Release 2019, 300, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.P.; Harwood, J.; Lee, R.; She, R.; Guiney, D.G. Characterization of Monospecies Biofilm Formation by Helicobacter pylori. J. Bacteriol. 2004, 186, 3124–3132. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Hu, S.; Ottemann, K.M. Genetic requirements and transcriptomics of Helicobacter pylori biofilm formation on abiotic and biotic surfaces. npj Biofilms Microbiomes 2020, 6, 1–14. [Google Scholar] [CrossRef]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Involves a Multigene Stress-Biased Response, Including a Structural Role for Flagella. mBio 2018, 9, e01973–18. [Google Scholar] [CrossRef]
- Williams, J.C.; McInnis, K.A.; Testerman, T.L. Adherence of Helicobacter pylori to Abiotic Surfaces Is Influenced by Serum. Appl. Environ. Microbiol. 2008, 74, 1255–1258. [Google Scholar] [CrossRef]
- Tan, S.; Tompkins, L.S.; Amieva, M.R. Helicobacter pylori Usurps Cell Polarity to Turn the Cell Surface into a Replicative Niche. PLOS Pathog. 2009, 5, e1000407. [Google Scholar] [CrossRef]
- Anderson, J.K.; Huang, J.Y.; Wreden, C.; Sweeney, E.G.; Goers, J.; Remington, S.J.; Guillemin, K. Chemorepulsion from the Quorum Signal Autoinducer-2 Promotes Helicobacter pylori Biofilm Dispersal. mBio 2015, 6, e00379–15. [Google Scholar] [CrossRef]
- Sigal, M.; Rothenberg, M.E.; Logan, C.Y.; Lee, J.Y.; Honaker, R.W.; Cooper, R.L.; Passarelli, B.; Camorlinga, M.; Bouley, D.M.; Alvarez, G.; et al. Helicobacter pylori Activates and Expands Lgr5+ Stem Cells Through Direct Colonization of the Gastric Glands. Gastroenterology 2015, 148, 1392–1404. [Google Scholar] [CrossRef]
- Cellini, L., Grande, R., Di Campli, E., Di Bartolomeo, S., Di Giulio, M., Traini, T., & Trubiani, O. Characterization of an Helicobacter pylori environmental strain. Journal of applied microbiology 2008, 105(3), 761–769.
- Bugli, F., Palmieri, V., Torelli, R., Papi, M., De Spirito, M., Cacaci, M., ... & Sanguinetti, M. In vitro effect of clarithromycin and alginate lyase against Helicobacter pylori biofilm. Biotechnology Progress 2016, 32(6), 1584–1591.
- Azevedo, N.F.; Pinto, A.R.; Reis, N.M.; Vieira, M.J.; Keevil, C.W. Shear Stress, Temperature, and Inoculation Concentration Influence the Adhesion of Water-Stressed Helicobacter pylori to Stainless Steel 304 and Polypropylene. Appl. Environ. Microbiol. 2006, 72, 2936–41. [Google Scholar] [CrossRef] [PubMed]
- Ratthawongjirakul, P.; Thongkerd, V.; Chaicumpa, W. The impacts of a fliD mutation on the biofilm formation of Helicobacter pylori. Asian Pac. J. Trop. Biomed. 2016, 6, 1008–1014. [Google Scholar] [CrossRef]
- Hessey, S.J.; Spencer, J.; I Wyatt, J.; Sobala, G.; Rathbone, B.J.; Axon, A.T.; Dixon, M.F. Bacterial adhesion and disease activity in Helicobacter associated chronic gastritis. Gut 1990, 31, 134–138. [Google Scholar] [CrossRef]
- Zheng, X., Baker, H., Hancock, W. S., Fawaz, F., McCaman, M., & Pungor Jr, E. Proteomic analysis for the assessment of different lots of fetal bovine serum as a raw material for cell culture. Part IV. Application of proteomics to the manufacture of biological drugs. Biotechnology progress 2006, 22(5), 1294–1300.
- Yonezawa, H.; Osaki, T.; Fukutomi, T.; Hanawa, T.; Kurata, S.; Zaman, C.; Hojo, F.; Kamiya, S. Diversification of the AlpB Outer Membrane Protein of Helicobacter pylori Affects Biofilm Formation and Cellular Adhesion. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef]
- Senkovich, O.A.; Yin, J.; Ekshyyan, V.; Conant, C.; Traylor, J.; Adegboyega, P.; McGee, D.J.; Rhoads, R.E.; Slepenkov, S.; Testerman, T.L. Helicobacter pylori AlpA and AlpB Bind Host Laminin and Influence Gastric Inflammation in Gerbils. Infect. Immun. 2011, 79, 3106–3116. [Google Scholar] [CrossRef]
- Lim, J.W.; Kim, H.; Kim, K.H. Cell adhesion-related gene expression by Helicobacter pylori in gastric epithelial AGS cells. Int. J. Biochem. Cell Biol. 2003, 35, 1284–1296. [Google Scholar] [CrossRef]
- Wong, E.H.J.; Ng, C.G.; Chua, E.G.; Tay, A.C.Y.; Peters, F.; Marshall, B.J.; Ho, B.; Goh, K.L.; Vadivelu, J.; Loke, M.F. Comparative Genomics Revealed Multiple Helicobacter pylori Genes Associated with Biofilm Formation In Vitro. PLOS ONE 2016, 11, e0166835. [Google Scholar] [CrossRef]
- Servetas, S.L.; Kim, A.; Su, H.; Cha, J.; Merrell, D.S. Comparative analysis of the Hom family of outer membrane proteins in isolates from two geographically distinct regions: The United States and South Korea. Helicobacter 2018, 23, e12461. [Google Scholar] [CrossRef]
- Oleastro, M.; Monteiro, L.; Lehours, P.; Mégraud, F.; Ménard, A. Identification of Markers for Helicobacter pylori Strains Isolated from Children with Peptic Ulcer Disease by Suppressive Subtractive Hybridization. Infect. Immun. 2006, 74, 4064–4074. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, A.; Vallström, A.; Petzold, K.; Tegtmeyer, N.; Schleucher, J.; Carlsson, S.; Haas, R.; Backert, S.; Wai, S.N.; Gröbner, G.; et al. Biochemical and functional characterization of Helicobacter pylori vesicles. Mol. Microbiol. 2010, 77, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Grande, R.; Di Marcantonio, M.C.; Robuffo, I.; Pompilio, A.; Celia, C.; Di Marzio, L.; Paolino, D.; Codagnone, M.; Muraro, R.; Stoodley, P.; et al. Helicobacter pylori ATCC 43629/NCTC 11639 Outer Membrane Vesicles (OMVs) from Biofilm and Planktonic Phase Associated with Extracellular DNA (eDNA). Front. Microbiol. 2015, 6, 1369. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Kurata, S.; Fukuda, M.; Kawakami, H.; Ochiai, K.; Hanawa, T.; Kamiya, S. Outer Membrane Vesicles of Helicobacter pylori TK1402 are Involved in Biofilm Formation. BMC Microbiol. 2009, 9, 197–197. [Google Scholar] [CrossRef] [PubMed]
- Servetas, S. L. et al. ArsRS-dependent regulation of homB contributes to Helicobacter pylori biofilm formation. Front. Microbiol. 2018, 2(9), 1497.
- Oleastro, M.; Monteiro, L.; Lehours, P.; Mégraud, F.; Ménard, A. Identification of Markers for Helicobacter pylori Strains Isolated from Children with Peptic Ulcer Disease by Suppressive Subtractive Hybridization. Infect. Immun. 2006, 74, 4064–4074. [Google Scholar] [CrossRef]
- Abadi, A.T.B.; Rafiei, A.; Ajami, A.; Hosseini, V.; Taghvaei, T.; Jones, K.R.; Merrell, D.S. Helicobacter pylori homB, but Not cagA, Is Associated with Gastric Cancer in Iran. J. Clin. Microbiol. 2011, 49, 3191–3197. [Google Scholar] [CrossRef]
- Shen, Y.; Zou, Y.; Chen, X.; Li, P.; Rao, Y.; Yang, X.; Sun, Y.; Hu, H. Antibacterial self-assembled nanodrugs composed of berberine derivatives and rhamnolipids against Helicobacter pylori. J. Control. Release 2020, 328, 575–586. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Siavoshi, F.; Noghabi, K.A. Mucoid and coccoid Helicobacter pylori with fast growth and antibiotic resistance. Helicobacter 2019, 25, e12678. [Google Scholar] [CrossRef]
- El Mortaji, Lamya, Alejandro Tejada-Arranz, Aline Rifflet, Ivo G. Boneca, Gérard Pehau-Arnaudet, J. Pablo Radicella, Stéphanie Marsin, and Hilde De Reuse. "A peptide of a type I toxin− antitoxin system induces Helicobacter pylori morphological transformation from spiral shape to coccoids". Proceedings of the National Academy of Sciences 2020, 117(49), 31398–31409.
- Krzyżek, P.; Migdał, P.; Grande, R.; Gościniak, G. Biofilm Formation of Helicobacter pylori in Both Static and Microfluidic Conditions Is Associated With Resistance to Clarithromycin. Front. Cell. Infect. Microbiol. 2022, 12, 868905. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Paluch, E.; Karwańska, M.; Wieliczko, A.; Gościniak, G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 2695. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Mondragón, M.G.; Ares, M.A.; Panunzi, L.G.; Pacheco, S.; Camorlinga-Ponce, M.; Girón, J.A.; Torres, J.; De la Cruz, M.A. Transcriptional Profiling of Type II Toxin–Antitoxin Genes of Helicobacter pylori under Different Environmental Conditions: Identification of HP0967–HP0968 System. Front. Microbiol. 2016, 7, 1872. [Google Scholar] [CrossRef] [PubMed]
- Attaran, B.; Falsafi, T.; Kabiri, M. Biofilm Formation Capability of Clinical Helicobacter pylori Isolates on MKN-45 Cells. Jentashapir J. Cell. Mol. Biol. 2021, 12. [Google Scholar] [CrossRef]
- Rader, B.A.; Wreden, C.; Hicks, K.G.; Sweeney, E.G.; Ottemann, K.M.; Guillemin, K. Helicobacter pylori perceives the quorum-sensing molecule AI-2 as a chemorepellent via the chemoreceptor TlpB. Microbiology 2011, 157, 2445–2455. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, M.H.; Cover, T.L. Intercellular Communication in Helicobacter pylori : luxS Is Essential for the Production of an Extracellular Signaling Molecule. Infect. Immun. 2000, 68, 3193–3199. [Google Scholar] [CrossRef]
- Lee, W.-K.; Ogura, K.; Loh, J.T.; Cover, T.L.; Berg, D.E. Quantitative Effect of luxS Gene Inactivation on the Fitness of Helicobacter pylori. Appl. Environ. Microbiol. 2006, 72, 6615–6622. [Google Scholar] [CrossRef]
- Qiao, S.; Luo, Q.; Zhao, Y.; Zhang, X.C.; Huang, Y. Structural basis for lipopolysaccharide insertion in the bacterial outer membrane. Nature 2014, 511, 108–111. [Google Scholar] [CrossRef]
- Egan, A.J.F. Bacterial outer membrane constriction. Mol. Microbiol. 2018, 107, 676–687. [Google Scholar] [CrossRef]
- Alm, R.A.; Bina, J.; Andrews, B.M.; Doig, P.; Hancock, R.E.W.; Trust, T.J. Comparative Genomics of Helicobacter pylori : Analysis of the Outer Membrane Protein Families. Infect. Immun. 2000, 68, 4155–4168. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Woo, T.; Kurata, S.; Zaman, C.; Hojo, F.; Hanawa, T.; Kato, S.; Kamiya, S. Analysis of outer membrane vesicle protein involved in biofilm formation of Helicobacter pylori. Anaerobe 2011, 17, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Tamrakar, A.; Singh, R.; Kumar, A.; Makde, R.D.; Ashish; Kodgire, P. Biophysical characterization of the homodimers of HomA and HomB, outer membrane proteins of Helicobacter pylori. Sci. Rep. 2021, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Servetas, S.L.; Kang, J.; Kim, J.; Jang, S.; Choi, Y.H.; Su, H.; Jeon, Y.-E.; Hong, Y.A.; Yoo, Y.-J.; et al. Helicobacter pylori outer membrane protein, HomC, shows geographic dependent polymorphism that is influenced by the Bab family. J. Microbiol. 2016, 54, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Shang, K.; Zhang, L.; Li, W.; Wang, X. A rapid anti-Helicobacter pylori biofilm drug screening biosensor based on AlpB outer membrane protein and colloidal gold/nanoporous gold framework. Biosens. Bioelectron. 2022, 215, 114599. [Google Scholar] [CrossRef]
- Liu, W.Z.; Xie, Y.; Lu, H.; Cheng, H.; Zeng, Z.R.; Zhou, L.Y.; Chen, Y.; Bin Wang, J.; Du, Y.Q.; Lu, N.H.; et al. Fifth Chinese National Consensus Report on the management ofHelicobacter pyloriinfection. Helicobacter 2018, 23, e12475. [Google Scholar] [CrossRef]
- Miftahussurur, M.; Syam, A.F.; Nusi, I.A.; Makmun, D.; Waskito, L.A.; Zein, L.H.; Akil, F.; Uwan, W.B.; Simanjuntak, D.; Wibawa, I.D.N.; et al. Surveillance of Helicobacter pylori Antibiotic Susceptibility in Indonesia: Different Resistance Types among Regions and with Novel Genetic Mutations. PLOS ONE 2016, 11, e0166199. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management ofHelicobacter pyloriinfection—the Maastricht V/Florence Consensus Report. European Helicobacter and Microbiota Study Group and Consensus panel Gut 2017, 66, 6–30. [Google Scholar] [CrossRef]
- Miftahussurur, M.; Shrestha, P.K.; Subsomwong, P.; Sharma, R.P.; Yamaoka, Y. Emerging Helicobacter pylori levofloxacin resistance and novel genetic mutation in Nepal. BMC Microbiol. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Fauzia, K.A.; Miftahussurur, M.; Syam, A.F.; Waskito, L.A.; Doohan, D.; Rezkitha, Y.A.A.; Matsumoto, T.; Tuan, V.P.; Akada, J.; Yonezawa, H.; et al. Biofilm Formation and Antibiotic Resistance Phenotype of Helicobacter pylori Clinical Isolates. Toxins 2020, 12, 473. [Google Scholar] [CrossRef]
- Pan, J.; Shi, Z.; Lin, D.; Yang, N.; Meng, F.; Lin, L.; Jin, Z.; Zhou, Q.; Wu, J.; Zhang, J.; et al. Is tailored therapy based on antibiotic susceptibility effective ? a multicenter, open-label, randomized trial. Front. Med. 2020, 14, 43–50. [Google Scholar] [CrossRef]
- Fallone, C.A.; Chiba, N.; van Zanten, S.V.; Fischbach, L.; Gisbert, J.P.; Hunt, R.H.; Jones, N.L.; Render, C.; Leontiadis, G.I.; Moayyedi, P.; et al. The Toronto Consensus for the Treatment of Helicobacter pylori Infection in Adults. Gastroenterology 2016, 151, 51–69. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.Z.; Mendiondo, N.; Wonaga, A.; Viola, L.; Ibarra, D.; Campitelli, E.; Salim, N.; Corti, R.; Goldman, C.; Catalano, M. Occurrence of Mutations in the Antimicrobial Target Genes Related to Levofloxacin, Clarithromycin, and Amoxicillin Resistance inHelicobacter pyloriIsolates from Buenos Aires City. Microb. Drug Resist. 2017, 23, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, Y.; Song, X.; Tang, X.; Hu, R.; Marshall, B.J.; Tang, H.; Benghezal, M. Need for standardization and harmonization of Helicobacter pylori antimicrobial susceptibility testing. Helicobacter 2022, 27, e12873. [Google Scholar] [CrossRef]
- Mascellino, M.T.; Oliva, A.; De Angelis, M.; Pontone, S.; Porowska, B. Helicobacter pylori infection: antibiotic resistance and eradication rate in patients with gastritis showing previous treatment failures. new Microbiol. 2018, 41, 306–309. [Google Scholar]
- Redondo, J.J.; Keller, P.M.; Zbinden, R.; Wagner, K. A novel RT-PCR for the detection of Helicobacter pylori and identification of clarithromycin resistance mediated by mutations in the 23S rRNA gene. Diagn. Microbiol. Infect. Dis. 2018, 90, 1–6. [Google Scholar] [CrossRef]
- Lee, Sun Min, Nayoung Kim, Yong Hwan Kwon, Ryoung Hee Nam, Jung Mogg Kim, Jong Youn Park, Yeon Suk Lee, and Dong Ho Lee. "rdxA, frxA, and efflux pump in metronidazole-resistant Helicobacter pylori: their relation to clinical outcomes". Journal of Gastroenterology and Hepatology 2018, 33(3), 681–688. [CrossRef]
- Hanafi, A.; Lee, W.C.; Loke, M.F.; Teh, X.; Shaari, A.; Dinarvand, M.; Lehours, P.; Mégraud, F.; Leow, A.H.R.; Vadivelu, J.; et al. Molecular and Proteomic Analysis of Levofloxacin and Metronidazole Resistant Helicobacter pylori. Front. Microbiol. 2016, 7, 2015. [Google Scholar] [CrossRef]
- Attaran, Bahareh, and Tahereh Falsafi. "Identification of factors associated with biofilm formation ability in the clinical isolates of Helicobacter pylori". Iranian Journal of Biotechnology 2017, 15(1), 58. [CrossRef]
- Attaran, Bahareh, Tahereh Falsafi, and Nassim Ghorbanmehr. "Effect of biofilm formation by clinical isolates of Helicobacter pylori on the efflux-mediated resistance to commonly used antibiotics". World Journal of Gastroenterology 2017, 23(7), 1163. [CrossRef]
- Midolo, P.D.; Bell, J.M.; Lambert, J.R.; Turnidge, J.D.; Grayson, M.L. Antimicrobial resistance testing of Helicobacter pylori: A comparison of etest and disk diffusion methods. Pathology 1997, 29, 411–414. [Google Scholar] [CrossRef]
- Glupczynski, Y. O. U. R. I., Max Labbe, Willy Hansen, Françoise Crokaert, and Eugène Yourassowsky. "Evaluation of the E test for quantitative antimicrobial susceptibility testing of Helicobacter pylori". Journal of Clinical Microbiology 1991, 29(9), 2072–2075. [CrossRef]
- Tang, X.; Shen, Y.; Hu, R.; Yang, T.; Benghezal, M.; Li, H.; Tang, H. Re-assessment of the disk diffusion technique for routine antimicrobial susceptibility testing for Helicobacter pylori. Helicobacter 2020, 25, e12703. [Google Scholar] [CrossRef]
- Mishra, K.K.; Srivastava, S.; Garg, A.; Ayyagari, A. Antibiotic Susceptibility of Helicobacter pylori Clinical Isolates: Comparative Evaluation of Disk-Diffusion and E-Test Methods. Curr. Microbiol. 2006, 53, 329–334. [Google Scholar] [CrossRef]
- Fauzia, Kartika Afrida, Hafeza Aftab, Muhammad Miftahussurur, Langgeng Agung Waskito, Vo Phuoc Tuan, Takashi Matsumoto, Michiyuki Yurugi et al. "Genetic Determinants of Biofilm Formation and Antibiotic Resistance of Helicobacter Pylori using Whole Genome Sequencing." (2021). [CrossRef]
- Saruuljavkhlan, B.; Yamaoka, Y. Benefits of a Molecular-Based Method for the Detection of Clarithromycin-Resistant Helicobacter pylori. Gut Liver 2021, 15, 487–489. [Google Scholar] [CrossRef]
- Tshibangu-Kabamba, E.; de Jesus Ngoma-Kisoko, P.; Tuan, V.P.; Matsumoto, T.; Akada, J.; Kido, Y.; Tshimpi-Wola, A.; Tshiamala-Kashala, P.; Ahuka-Mundeke, S.; Ngoyi, D.M.; et al. Next-Generation Sequencing of the Whole Bacterial Genome for Tracking Molecular Insight into the Broad-Spectrum Antimicrobial Resistance of Helicobacter pylori Clinical Isolates from the Democratic Republic of Congo. Microorganisms 2020, 8, 887. [Google Scholar] [CrossRef]
- Smith, S.; Fowora, M.; Pellicano, R. Infections with Helicobacter pylori and challenges encountered in Africa. World J. Gastroenterol. 2019, 25, 3183–3195. [Google Scholar] [CrossRef]
- Mégraud, F.; Lehours, P. Helicobacter pylori Detection and Antimicrobial Susceptibility Testing. Clin. Microbiol. Rev. 2007, 20, 280–322. [Google Scholar] [CrossRef]
- van Doorn, L.-J.; Glupczynski, Y.; Kusters, J.G.; Mégraud, F.; Midolo, P.; Maggi-Solcà, N.; Queiroz, D.M.M.; Nouhan, N.; Stet, E.; Quint, W.G.V. Accurate Prediction of Macrolide Resistance in Helicobacter pylori by a PCR Line Probe Assay for Detection of Mutations in the 23S rRNA Gene: Multicenter Validation Study. Antimicrob. Agents Chemother. 2001, 45, 1500–1504. [Google Scholar] [CrossRef]
- Schabereiter-Gurtner, C.; Hirschl, A.M.; Dragosics, B.; Hufnagl, P.; Puz, S.; Kovách, Z.; Rotter, M.; Makristathis, A. Novel Real-Time PCR Assay for Detection of Helicobacter pylori Infection and Simultaneous Clarithromycin Susceptibility Testing of Stool and Biopsy Specimens. J. Clin. Microbiol. 2004, 42, 4512–4518. [Google Scholar] [CrossRef]
- Mitui, M.; Patel, A.; Leos, N.K.; Doern, C.D.; Park, J.Y. Novel Helicobacter pylori Sequencing Test Identifies High Rate of Clarithromycin Resistance. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 6–9. [Google Scholar] [CrossRef]
- Nishizawa, T.; Suzuki, H. Mechanisms of Helicobacter pylori antibiotic resistance and molecular testing. Front. Mol. Biosci. 2014, 1, 19–19. [Google Scholar] [CrossRef] [PubMed]
- Vital, J.S.; Tanoeiro, L.; Lopes-Oliveira, R.; Vale, F.F. Biomarker Characterization and Prediction of Virulence and Antibiotic Resistance from Helicobacter pylori Next Generation Sequencing Data. Biomolecules 2022, 12, 691. [Google Scholar] [CrossRef] [PubMed]
- Her, H.-L.; Wu, Y.-W. A pan-genome-based machine learning approach for predicting antimicrobial resistance activities of the Escherichia coli strains. Bioinformatics 2018, 34, i89–i95. [Google Scholar] [CrossRef] [PubMed]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter pylori infection and antibiotic resistance — from biology to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613–629. [Google Scholar] [CrossRef]
- Li, P.; Chen, X.; Shen, Y.; Li, H.; Zou, Y.; Yuan, G.; Hu, P.; Hu, H. Mucus penetration enhanced lipid polymer nanoparticles improve the eradication rate of Helicobacter pylori biofilm. J. Control. Release 2019, 300, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Reshetnyak, V.I.; Reshetnyak, T.M. Significance of dormant forms ofHelicobacter pyloriin ulcerogenesis. World J. Gastroenterol. 2017, 23, 4867–4878. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.G.; Hazell, S.L.; Netting, A.G. Use of digoxigenin-labelled ampicillin in the identification of penicillin-binding proteins in Helicobacter pylori. J. Antimicrob. Chemother. 2000, 45, 591–598. [Google Scholar] [CrossRef]
- Ikeda, F.; Yokota, Y.; Mine, Y.; Tatsuta, M. Activity of cefixime against Helicobacter pylori and affinities for the penicillin-binding proteins. Antimicrob. Agents Chemother. 1990, 34, 2426–2428. [Google Scholar] [CrossRef]
- Costa, K.; Bacher, G.; Allmaier, G.; Dominguez-Bello, M.G.; Engstrand, L.; Falk, P.; de Pedro, M.A.; Portillo, F.G.-D. The Morphological Transition of Helicobacter pylori Cells from Spiral to Coccoid Is Preceded by a Substantial Modification of the Cell Wall. J. Bacteriol. 1999, 181, 3710–3715. [Google Scholar] [CrossRef]
- Kuo, C.-J.; Guo, R.-T.; Lu, I.-L.; Liu, H.-G.; Wu, S.-Y.; Ko, T.-P.; Wang, A.H.-J.; Liang, P.-H. Crystal Structures and Computer Screened Inhibitors of Helicobacter pylori Undecaprenyl Pyrophosphate Synthase. 2007; 305–310. [Google Scholar] [CrossRef]
- Modak, Joyanta K., Alexandra Tikhomirova, Rebecca J. Gorrell, Mohammad M. Rahman, Despina Kotsanas, Tony M. Korman, Jose Garcia-Bustos et al. "Anti-Helicobacter pylori activity of ethoxzolamide". Journal of enzyme inhibition and medicinal chemistry 2019, 34(1), 1660–1667.
- Rahman, M.M.; Tikhomirova, A.; Modak, J.K.; Hutton, M.L.; Supuran, C.T.; Roujeinikova, A. Antibacterial activity of ethoxzolamide against Helicobacter pylori strains SS1 and 26695. Gut Pathog. 2020, 12, 1–7. [Google Scholar] [CrossRef]
- Wang, G.; Maier, S.E.; Lo, L.F.; Maier, G.; Dosi, S.; Maier, R.J. Peptidoglycan Deacetylation in Helicobacter pylori Contributes to Bacterial Survival by Mitigating Host Immune Responses. Infect. Immun. 2010, 78, 4660–4666. [Google Scholar] [CrossRef]
- Zhao, Y., Cai, Y., Chen, Z., Li, H., Xu, Z., Li, W.,... & Sun, Y. SpoT-mediated NapA upregulation promotes oxidative stress-induced Helicobacter pylori biofilm formation and confers multidrug resistance. Antimicrobial agents and chemotherapy 2021, 65(5), e00152-21.
- Wang, G.; Lo, L.F.; Forsberg, L.S.; Maier, R.J. Helicobacter pylori Peptidoglycan Modifications Confer Lysozyme Resistance and Contribute to Survival in the Host. mBio 2012, 3, e00409–12. [Google Scholar] [CrossRef] [PubMed]
- Chou, Feng-Pai, Chia-Tse Tsai, Ya-Sheng Chiou, Yi-Ju Chen, Meng-Erh Li, Ting-Wei Guo, Jason WenJay Lyu, Sheng-Hao Chou, and Tung-Kung Wu. "An enzymatic approach to configurationally rare trans-androsteronyl-α-glucoside and Its potential anticancer application". Chemical Biology & Drug Design 2017, 89(1), 61–66.
- Qaria, M. A., Kumar, N., Hussain, A., Qumar, S., Doddam, S. N., Sepe, L. P., & Ahmed, N. Roles of cholesteryl-α-glucoside transferase and cholesteryl glucosides in maintenance of Helicobacter pylori morphology, cell wall integrity, and resistance to antibiotics. MBio 2018, 9(6), e01523-18.
- Marcus EA et al. The effects of varying acidity on Helicobacter pylori growth and bactericidal efficacy of ampicillin. Aliment Pharmacol Ther 2012, 36, 972. [CrossRef]
- Wong, E.H.J.; Ng, C.G.; Goh, K.L.; Vadivelu, J.; Ho, B.; Loke, M.F. Metabolomic analysis of low and high biofilm-forming Helicobacter pylori strains. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Njume, C.; Afolayan, A.J.; Samie, A.; Ndip, R.N. Inhibitory and Bactericidal Potential of Crude Acetone Extracts of Combretum molle (Combretaceae) on Drug-resistant Strains of Helicobacter pylori. J. Heal. Popul. Nutr. 2011, 29, 438–445. [Google Scholar] [CrossRef]
- Loh, J.T.; Gupta, S.S.; Friedman, D.B.; Krezel, A.M.; Cover, T.L. Analysis of Protein Expression Regulated by the Helicobacter pylori ArsRS Two-Component Signal Transduction System. J. Bacteriol. 2010, 192, 2034–2043. [Google Scholar] [CrossRef]
- Allen, M.G.; Bate, M.Y.; Tramonte, L.M.; Avalos, E.Y.; Loh, J.; Cover, T.L.; Forsyth, M.H. Regulation of Helicobacter pylori Urease and Acetone Carboxylase Genes by Nitric Oxide and the CrdRS Two-Component System. Microbiol. Spectr. 2023, 11, e0463322. [Google Scholar] [CrossRef] [PubMed]
- Brahmachary, P.; Wang, G.; Benoit, S.L.; Weinberg, M.V.; Maier, R.J.; Hoover, T.R. The human gastric pathogen Helicobacter pylori has a potential acetone carboxylase that enhances its ability to colonize mice. BMC Microbiol. 2008, 8, 14–14. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, M.A.; Ares, M.; Von Bargen, K.; Panunzi, L.G.; Martínez-Cruz, J.; Valdez-Salazar, H.-A.; Jiménez-Galicia, C.; Torres, J. Gene Expression Profiling of Transcription Factors of Helicobacter pylori under Different Environmental Conditions. Front. Microbiol. 2017, 8, 615. [Google Scholar] [CrossRef] [PubMed]
- Servetas, S.L.; Carpenter, B.M.; Haley, K.P.; Gilbreath, J.J.; Gaddy, J.A.; Merrell, D.S. Characterization of Key Helicobacter pylori Regulators Identifies a Role for ArsRS in Biofilm Formation. J. Bacteriol. 2016, 198, 2536–2548. [Google Scholar] [CrossRef]
- Raj, D.S.; Kesavan, D.K.; Muthusamy, N.; Umamaheswari, S. Efflux pumps potential drug targets to circumvent drug Resistance – Multi drug efflux pumps of Helicobacter pylori. Mater. Today: Proc. 2021, 45, 2976–2981. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Hojo, F.; Kamiya, S. Effect of Helicobacter pylori biofilm formation on susceptibility to amoxicillin, metronidazole and clarithromycin. Microb. Pathog. 2019, 132, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Cai, Y.; Chen, Z.; Gao, S.; Geng, X.; Li, Y.; Jia, J.; Sun, Y. Bifunctional Enzyme SpoT Is Involved in Biofilm Formation of Helicobacter pylori with Multidrug Resistance by Upregulating Efflux Pump Hp1174 ( gluP ). Antimicrob. Agents Chemother. 2018, 62, e00957–18. [Google Scholar] [CrossRef]
- Cai, Y., Wang, C., Chen, Z., Xu, Z., Li, H., Li, W., & Sun, Y. Transporters HP0939, HP0497, and HP0471 participate in intrinsic multidrug resistance and biofilm formation in Helicobacter pylori by enhancing drug efflux. Helicobacter 2020, 25(4), e12715.
- Hathroubi, S.; Zerebinski, J.; Clarke, A.; Ottemann, K.M. Helicobacter Pylori Biofilm Confers Antibiotic Tolerance in Part via A Protein-Dependent Mechanism. Antibiotics 2020, 9, 355. [Google Scholar] [CrossRef]
- Jolaiya, Tolulope Funbi, Muinah Adenike Fowora, Charles Onyekwere, Rose Ugiagbe, Ifeanyi Ifeoma Agbo, Olufunmilayo Lesi, Dennis Amojuayi Ndububa et al. "Gastroenterology and Hepatology Research". Journal of GHR 2020, 9(4), 3283–3289.
- Jiang, M.; Ma, L.; Huang, Y.; Wu, H.; Dou, J.; Zhou, C. Antimicrobial activities of peptide Cbf-K16 against drug-resistant Helicobacter pylori infection in vitro and in vivo. Microb. Pathog. 2020, 138, 103847. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Jiang, A.-M.; Ma, Z.-Y.; Li, X.-B.; Xiong, Y.-Y.; Dou, J.-F.; Wang, J.-F. The Synthetic Antimicrobial Peptide Pexiganan and Its Nanoparticles (PNPs) Exhibit the Anti-Helicobacter pylori Activity in Vitro and in Vivo. Molecules 2015, 20, 3972–3985. [Google Scholar] [CrossRef]
- Parreira, P.; Monteiro, C.; Graça, V.; Gomes, J.; Maia, S.; Gomes, P.; Gonçalves, I.C.; Martins, M.C.L. Surface Grafted MSI-78A Antimicrobial Peptide has High Potential for Gastric Infection Management. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- de la Fuente-Núñez, C.; Reffuveille, F.; Mansour, S.C.; Reckseidler-Zenteno, S.L.; Hernández, D.; Brackman, G.; Coenye, T.; Hancock, R.E. D-Enantiomeric Peptides that Eradicate Wild-Type and Multidrug-Resistant Biofilms and Protect against Lethal Pseudomonas aeruginosa Infections. Chem. Biol. 2015, 22, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Mansour, S.C.; de la Fuente-Núñez, C.; Hancock, R.E.W. Peptide IDR-1018: modulating the immune system and targeting bacterial biofilms to treat antibiotic-resistant bacterial infections. J. Pept. Sci. 2014, 21, 323–329. [Google Scholar] [CrossRef]
- Wells, D.H.; Gaynor, E.C. Helicobacter pylori Initiates the Stringent Response upon Nutrient and pH Downshift. J. Bacteriol. 2006, 188, 3726–9. [Google Scholar] [CrossRef]
- Ji, J.; Yang, H. In Vitro Effects of Lactobacillus plantarum LN66 and Antibiotics Used Alone or in Combination on Helicobacter pylori Mature Biofilm. Microorganisms 2021, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Salas-Jara, M.J.; Sanhueza, E.A.; Retamal-Díaz, A.; González, C.; Urrutia, H.; García, A. Probiotic Lactobacillus fermentum UCO-979C biofilm formation on AGS and Caco-2 cells and Helicobacter pylori inhibition. Biofouling 2016, 32, 1245–1257. [Google Scholar] [CrossRef]
- Jin, F.; Yang, H. Effects of Lactobacillus salivarius LN12 in Combination with Amoxicillin and Clarithromycin on Helicobacter pylori Biofilm In Vitro. Microorganisms 2021, 9, 1611. [Google Scholar] [CrossRef]
- Dufour, C.; Wink, J.; Kurz, M.; Kogler, H.; Olivan, H.; Sablé, S.; Heyse, W.; Gerlitz, M.; Toti, L.; Nußer, A.; et al. Isolation and Structural Elucidation of Armeniaspirols A-C: Potent Antibiotics against Gram-Positive Pathogens. Chem. – A Eur. J. 2012, 18, 16123–16128. [Google Scholar] [CrossRef]
- Jia, J., Zhang, C., Liu, Y., Huang, Y., Bai, Y., Hang, X.,... & Bi, H. Armeniaspirol A: a novel anti-Helicobacter pylori agent. Microbial biotechnology 2022, 15(2), 442–454.
- Yu, M.; Wang, X.; Ling, F.; Wang, H.; Zhang, P.; Shao, S. Atractylodes lancea volatile oils attenuated helicobacter pylori NCTC11637 growth and biofilm. Microb. Pathog. 2019, 135, 103641. [Google Scholar] [CrossRef] [PubMed]
- Wylie, M.R.; Windham, I.H.; Blum, F.C.; Wu, H.; Merrell, D.S. In vitro antibacterial activity of nimbolide against Helicobacter pylori. J. Ethnopharmacol. 2022, 285, 114828–114828. [Google Scholar] [CrossRef]
- Prasad, A.; Devi, A.T.; Prasad, M.N.N.; Zameer, F.; Shruthi, G.; Shivamallu, C. Phyto anti-biofilm elicitors as potential inhibitors ofHelicobacter pylori. 3 Biotech 2019, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Huang, Y.; Hang, X.; Tong, Q.; Zeng, L.; Jia, J.; Zhang, G.; Bi, H. Dihydrotanshinone I Is Effective against Drug-Resistant Helicobacter pylori In Vitro and In Vivo. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef] [PubMed]
- Tran Trung, H.; Truong Thi Huynh, H.; Nguyen Thi Thuy, L.; Nguyen Van Minh, H.; Thi Nguyen, M.N.; Luong Thi, M.N. Growth-Inhibiting, Bactericidal, Antibiofilm, and Urease Inhibitory Activities of Hibiscus rosa sinensis L. Flower Constituents toward Antibiotic Sensitive- and Resistant-Strains of Helicobacter pylori. ACS Omega 2020, 5, 20080–20089. [Google Scholar] [CrossRef]
- Grande, R.; Carradori, S.; Puca, V.; Vitale, I.; Angeli, A.; Nocentini, A.; Bonardi, A.; Gratteri, P.; Lanuti, P.; Bologna, G.; et al. Selective Inhibition of Helicobacter pylori Carbonic Anhydrases by Carvacrol and Thymol Could Impair Biofilm Production and the Release of Outer Membrane Vesicles. Int. J. Mol. Sci. 2021, 22, 11583. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Masanam, E.; Ramkumar, V.S.; Baskaraligam, V.; Selvaraj, G. Influence of N -acylhomoserine lactonase silver nanoparticles on the quorum sensing system of Helicobacter pylori : A potential strategy to combat biofilm formation. J. Basic Microbiol. 2020, 60, 207–215. [Google Scholar] [CrossRef]
- Grande R, Sisto F, Puca V, et al. Antimicrobial and antibiofilm activities of new synthesized silver Ultra-NanoClusters (SUNCs) against helicobacter pylori. Front Microbiol. 2020, 11, 1705. [CrossRef]
- Huang, Y.; Li, R.; Dai, Y.; Qin, C.; He, J.; Yang, S.; Wang, T.; Su, Y.; Jia, L.; Zhao, W. Rhamnolipid-assisted black phosphorus nanosheets with efficient isolinderalactone loading against drug resistant Helicobacter pylori. Mater. Des. 2022, 216, 110536. [Google Scholar] [CrossRef]
- Arif, M.; Ahmad, R.; Sharaf, M.; Samreen; Muhammad, J. ; Abdalla, M.; Eltayb, W.A.; Liu, C.-G. Antibacterial and antibiofilm activity of mannose-modified chitosan/PMLA nanoparticles against multidrug-resistant Helicobacter pylori. Int. J. Biol. Macromol. 2022, 223, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Parreira, P.; Monteiro, C.; Graça, V.; Gomes, J.; Maia, S.; Gomes, P.; Gonçalves, I.C.; Martins, M.C.L. Surface Grafted MSI-78A Antimicrobial Peptide has High Potential for Gastric Infection Management. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



