Submitted:
03 July 2023
Posted:
03 July 2023
You are already at the latest version
Abstract
Microorganisms present in fermented foods and beverages play a vital role in enhancing both nutritive and non-nutritive components. During the fermentation, microorganisms can either naturally occur in the food through spontaneous fermentation or be intentionally added as starter cultures with a variety of species and strains. In this review, the focus is placed on micronutrients, such as vitamins and minerals, and three major macromolecules – prebiotics, bioactive peptides, and functional fatty acids. During fermentation, non-digestible carbohydrates like fructooligosaccharides and galactooligosaccharides are synthesized, imparting prebiotic properties that support gut health. Enzymatic hydrolysis of proteins by microorganisms yields bioactive peptides with diverse health benefits. Furthermore, fermentation facilitates the formation of fatty acids, including short-chain fatty acids and conjugated linoleic acid, potentially exerting physiological effects. By harnessing the power of microbial fermentation, nutrient-rich and bioactive compounds can be enhanced, offering a promising approach to address global malnutrition concerns.
Keywords:
Fermentation
; vitamins
; minerals
; bioactive compounds
; prebiotics
; conjugated fatty acids
Introduction
Adequate food consumption does not necessarily guarantee sufficient and balanced nutrient intake. Achieving a healthy and balanced diet requires consuming nutrient-rich food, which can be achieved through fortification. Fortification aims to supplement or achieve target levels of nutrients in the population or specific population groups, particularly those affected by hunger and malnutrition. The importance of food fortification is further emphasized during the COVID-19 outbreak. Undernourished individuals with metabolic health issues such as obesity and diabetes face a higher risk associated with COVID-19 [1].
Malnutrition encompasses inadequate, excessive, or imbalanced intake of energy and nutrients. Globally, there are 1.9 billion overweight adults and 462 million underweight adults. Furthermore, 144 million children under the age of five experience stunted growth, while 38.3 million are overweight (WHO, 2020). Reasons for malnutrition include low intake of animal sources in the diet, lack of education, poor economic conditions, and insufficient food availability [2]. According to Chadare et al., (2019), fortification is recognized as a public health strategy to address such issues by balancing nutrient intake without increasing calorie intake. Currently, 83 countries mandating the fortification of cereal grains, 20 fortifying edible oil, and nine fortifying sugar. Some countries also fortify rice, milk, and condiments (Leung, Braverman and Pearce, 2012; Mannar and Hurrel, 2018).
Micronutrients are essential substances that are required in small quantities but are vital for various physiological functions in the human body. They include vitamins (organic compounds) and minerals (inorganic compounds), that differ in term of chemical structure and specific roles. Micronutrients are involved in energy metabolism, immune function, DNA synthesis, and many other processes. On the other hand, non-nutrients are bioactive compounds present in food that go beyond basic nutrition. They encompass a wide range of substances, including bioactive peptides, prebiotics, and conjugated fats. Bioactive peptides are short chains of amino acids that can exert specific physiological effects, such as antioxidant, antihypertensive, and antimicrobial activities. Prebiotics are non-digestible carbohydrates that selectively stimulate the growth and activity of beneficial gut bacteria, promoting gut health. Conjugated fats, such as conjugated linoleic acid (CLA), are modified forms of fatty acids that have been associated with various health benefits, including anti-inflammatory and anticancer properties [4].
Fermentation, a natural process facilitated by microorganisms, has emerged as a promising technique to enhance the biosynthesis and availability of both micronutrients and non-nutrients in food [5]. Fermentation can occur spontaneously or with the use of starter cultures containing specific microbial species [6]. Microbes involved in fermentation can produce or activate certain vitamins, release minerals from complex compounds, generate bioactive peptides through enzymatic hydrolysis, and convert fatty acids into conjugated forms [7]. This transformation not only increases the content of these compounds but also improves their bioavailability, making fermented foods a valuable source of micronutrients and non-nutrients with potential health benefits [7].
Common microbes involved in fermentation include lactic acid bacteria (LAB), yeast (such as Saccharomyces species), and fungi (including Aspergillus, Penicillium, and Fusarium) [8,9,10,11,12]. To ensure food safety, the microbes used in fermentation must be categorized as Generally Recognized as Safe (GRAS). The selection of microorganisms determines their characteristics, such as probiotic properties, antioxidant activity, peptide production, or degradation of antinutritive compounds. The chosen strains should also exist in the intestinal flora for further breakdown [13].
This review aims to demonstrate that fermentation-based biofortification offers a promising approach for enhancing the nutritional value of food through the production of beneficial compounds. The studies discussed in this review highlight the potential of microbial fermentation to produce various nutritional and non-nutritional compounds that have been associated with various health benefits. The choice of specific microorganisms and environmental factors that influence these processes can guide the optimization of fermentation conditions to maximize the production of desired compounds.
Impact of Fermentation on Micronutrients
Micronutrients, as indicated by the term "micro," are essential nutrients required in small quantities that promote health benefits such as growth and immune system support, reducing the risk of infection [14]. They can be classified into four categories: water-soluble vitamins, fat-soluble vitamins, macro-minerals, and trace minerals.
Typically, minerals are not typically synthesized by microorganisms, but their bioavailability can be significantly enhanced through microbial actions. Microorganisms have the capability to break down antinutritional factors that bind to minerals, such as phytic acids, tannins, oxalates, enzyme inhibitors, lectins, and glucosinates [7] (Figure 1). Moreover, microbial activity leads to the production of organic acids, such as lactic acid and acetic acid. These organic acids play a crucial role in breaking down the cell walls of plant-based foods, thereby releasing and making minerals more readily available for absorption. Furthermore, microorganisms secrete digestive enzymes like proteases and lipases, which effectively break down food into smaller molecules, facilitating the enhanced absorption of minerals.
Sadh et al. (2018) demonstrated that fermenting black-eyed pea flour with Aspergillus oryzae at 30°C for 96 hours improved zinc bioavailability from 14.36 ± 0.96% in unfermented flour to 29.63 ± 1.55% in fermented flour, as determined by in vitro assays. A. oryzae is known to possess hydrolytic enzymes such as phytase, which can improve zinc absorption during the fermentation process (Zhang et al., 2015). Similarly, Lazarte et al. (2015) found that fermented cassava tuber exhibited increased zinc bioabsorption from 16.5% to 240% compared to unfermented tuber, with a decrease in pH value from 6.80 to 5.1. The drop in pH value indicates phytate breakdown, as phytic acid activity is optimal at pH values between 4.5 and 5.5 [18].
Microorganisms possess the remarkable ability to synthesize certain nutrients de novo, including essential amino acids and vitamins such as the vitamin B complex (Figure 2). Recent research has demonstrated the ability of different microbial producers to synthesize riboflavin (B2) during the fermentation process. B2 is an important co-enzyme, especially for the formation of flavin adenine dinucleotide (FADH2) that participates as electron carrier during the metabolic processes. The biosynthesis of riboflavin involves a series of enzymatic reactions, starting from the precursor molecule GTP (guanosine triphosphate). Microbes such as bacteria, yeasts, and fungi can produce B2, with certain bacterial species like Bacillus subtilis and Ashbya gossypii being efficient B2 producers. These microorganisms have specific genes encoding the enzymes responsible for the biosynthesis of B2.
Rajendran et al. (2017) found that S. boulardii SAA655, when used as a starter culture in naturally fermented idli batter, increased the B2 content from a static level to 500 ng/g after 14 hours of fermentation. Similarly, Chagas Junior et al., (2021), observed that the addition of starter cultures (S. cerevisiae and Pichia kudriavzevii) in cocoa reduced fermentation time from 96 hours to 72 hours while enhancing B2 production. Thompson et al., (2020) investigated B2 production in a fermented cauliflower and white bean mixture with L. plantarum Lp900. They found a significant increase in riboflavin content (91.6 ± 0.6 μg/100 g fresh weight) compared to the unfermented control (42.83 ± 1.20 μg/100 g fresh weight). Carrizo et al. (2020) also observed that fermentation of quinoa bread with L. plantarum strains increased B2 production (5.1 ± 0.4 μg/g dough) compared to unfermented quinoa bread (2.2 ± 0.1 μg/g dough) after 24 hours at 30°C. Altogether, these result suggests that the addition of starter culture can reduce fermentation time and also enhance the production of B2 [22].
Folate (B9) is important for DNA synthesis, cell division, and red blood cell production. Like B2, B9 can be synthesised by microorganisms de novo through a biosynthetic pathway (Figure 2). This pathway involves a series of enzymatic reactions that convert p-aminobenzoic acid (PABA) and pteridine derivatives into tetrahydrofolate (THF), the active form of vitamin B9. Lactic acid bacteria possess the necessary enzymes, including dihydropteroate synthase (DHPS), dihydrofolate reductase (DHFR), and folylpolyglutamate synthetase (FPGS), to catalyze these reactions.
In recent literatures Laiño et al. (2013) found that yoghurt supplemented with L. bulgaricus CRL871, S. thermophilus CRL803, and CRL415 strains produced the highest amount of B9 (180 ± 10 mg/L) after 6 hours of fermentation at 42°C, which was 125% higher than commercial yoghurt. Kobayashi et al. (2019) mentioned that S. thermophilus promotes the activity of L. bulgaricus, leading to increased B9 production. Clara et al., (2020) also reported positive results for B9 production after 30 days of cheese fermentation with nine species: L. plantarum, L. casei, L. delbrueckii subsp. bulgaricus, L. paracasei subsp. paracasei, L. rhamnosus, St. thermophilus, Lc. lactis subsp. lactis, E. lactis, and E. faecium.
The comprehensive review on the ability of microorganism in produce cobalamin (B12) was performed by [26]. Vitamin B12 is primarily produced by bacteria during fermentation, with Propionibacterium freudenreichii, L. reuteri, L. plantarum, and Bifidobacterium animalis being notable producers. The biosynthesis of vitamin B12 involves complex pathways, including the conversion of precursors like uroporphyrinogen III. L. reuteri has approximately 30 genes responsible for vitamin B12 synthesis, with the hem gene positioned uniquely in the middle of the gene cluster [27].
Xie et al. (2019) investigated the co-fermentation effect with pH value on the production of B12 in fermented wheat bran. They found that wheat bran fermented with P. freudenreichii DSM 20271 and L. brevis ATCC 14869 showed an increase in B12 production from day 1 (255 ± 31 ng/g dw) to day 3 (332 ± 44 ng/g dw) at 25°C. Lactobacillus strains contribute to food safety by inhibiting the growth of other microbes and promoting acidic conditions in the medium (Selwet, 2020), while P. freudenreichii is a microbial producer of B12 [29]. Wolkers – Rooijackers et al. (2018) also supported the role of P. freudenreichii as a B12 producer during the fermentation of lupin tempeh at 25°C. However, they found lower B12 production (0.01 ± 0 μg/100 g) in fermented lupin tempeh at 25°C for 2 days, while B12 production (30.67 ± 0.17 μg/100 g) was detected at 30°C fermentation. The optimum incubation temperature for B12 synthesis by Propionibacterium spp. is in the range of 30 to 35°C [31]. Overall, these findings demonstrate the potential of fermentation to enhance the production of micronutrients such as B2, B9, vB12, and minerals like iron and zinc. By selecting specific microbial strains, controlling pH, and optimizing incubation temperatures, the nutritional quality of fermented foods can be improved.
Vitamin K exists in multiple dietary forms, including phylloquinone (PK) and menaquinones (MKs). PK, also known as vitamin K-1, is predominantly found in green leafy vegetables, while MKs, including MK-4 to MK-13, are primarily synthesized by bacteria and present in smaller amounts in meat, dairy, and fermented food products. Bacterial synthesis of MKs was discovered in the 1930s, and most bacteria produce MKs with isoprenoid side chains of varying lengths. Bacteria utilize distinct biochemical pathways for MK synthesis, such as the pathway employed by LAB in food fermentations. In this pathway, the napthoquinone ring is synthesized from chorismate, and the isoprenoid side chain is separately formed before being joined to complete MK biosynthesis. The specific bacterial strains and fermentation conditions influence the concentrations and forms of MKs in fermented foods. For instance, cheese and milk products are major dietary sources of long-chain MKs, particularly MK-8 and MK-9, produced by LAB strains. Other fermented foods like sauerkraut and natto also contain long-chain MKs. However, limited comprehensive data exist on MK contents in different foods and regional variations in dairy consumption, necessitating further research to accurately assess MK intake at the individual and population levels [32].
Effect of fermentation on Non-nutrient/Bioactive Compounds
Bioactive compounds can be described as phytochemicals released from the food matrix, which contribute to various health benefits through their metabolism [33]. This concept, originally advocated by Hippocrates 2500 years ago with the statement "Let food be thy medicine and medicine be the food," is now embraced by consumers who are increasingly educated and aware of the impact of their daily food intake. The growing demand for fermented foods in recent years is closely tied to society's increasing concern for health and quality of life. Fermentation is known to transform food crops into edible forms, enriching them with natural bioactive compounds such as non-digestible carbohydrates, proteins, and fatty acids.
Non-Digestible Carbohydrate
Indigestible carbohydrates, also known as soluble fiber, are carbohydrates that cannot be digested by the human digestive system and instead bypass the absorption process in the upper part of the intestine. When these indigestible carbohydrates reach the lower gut, they have the ability to modify the composition and function of microorganisms in the human gut [34]. Consumption of prebiotics by probiotic bacteria leads to several changes, including an increase in the expression or alteration of the composition of short-chain fatty acids, a decrease in luminal colon pH, and a reduction in nitrogenous end-products [35]. These changes can contribute to numerous health benefits, such as improving intestinal health, strengthening the immune system, preventing cardiovascular-related conditions, and enhancing the gut's ability to absorb certain nutrients [36]. In this section, we will focus on two main types of prebiotics: fructooligosaccharides and galactooligosaccharides. These prebiotics play a significant role in promoting gut health and overall well-being.
Prebiotics - Fructooligosaccharide (FOS) and Galactooligosaccharides (GOS)
Prebiotic production is commonly associated with fungi, although certain LAB also have the capability to produce prebiotics like FOS and GOS. The inclusion of prebiotics, specifically oligosaccharides, has been demonstrated to promote the growth of LAB and enhance the production of B-vitamins, especially folate [37,38]. However, rather than being naturally produced (in situ) during food processing, these prebiotics are often added or fortified in food to enhance their functionality. They can directly influence the health properties of the food by providing specific benefits, or indirectly enhance the effectiveness of probiotics present in the food. Table 1 presents a collection of studies that have explored the capacity of microorganisms to increase the content of FOS and GOS in food. The production of prebiotics in situ is more commonly practiced with GOS compared to FOS. Industrial production of FOS is typically carried out under controlled conditions using fungi, while GOS is often produced directly within dairy-based fermented foods, benefiting from the numerous LAB strains that naturally ferment such products.
FOS are a type of inulin-type oligosaccharide commonly used as prebiotics due to their strong bifidogenic activity [39,40]. The high demand for FOS has led to ongoing research in search of new microbes that can produce fructosyltransferase (FTase) with excellent transfructosylation activity, which is crucial for FOS production. Some potent microorganisms involved in the industrial fermentation of FOS includes Aspergillus sp., Penicillium sp., Aureobasidium pullulans, LAB, and some bacterial species [41]. Both submerged and solid-state fermentation methods can be employed for cost-effective and efficient production of short-chain FOS. The food industry often uses FOS as low-calorie sweeteners due to their desirable properties [42].
FOS can be synthesized through a transfructosylation reaction via two pathways: the enzyme ß-D-fructofuranosidase (FFase) or fructosyltransferase (FTase). The process involves two major steps. The first step, catalyzed by FFase (EC 3.2.1.26), irreversibly hydrolyzes the glycosidic bond of sucrose (GF) by cleaving the β (2,1) linkages. FFase acts as both a donor and an acceptor of a fructosyl residue, resulting in the production of glucose and the trisaccharide 1-kestose. The second step is a readily reversible reaction, where 1-kestose or another β (2,1) linked fructan can act as the fructosyl donor, while GF or any fructan can act as the acceptor. This reaction is catalyzed by FTase (EC 2.4.1.9), which specifically cleaves and reforms the β (2,1) linkage between fructosyl-fructose units [39,40].
The physicochemical characteristics of the substrate play a crucial role in FOS production during microbial fermentation. Studies by [55]and [56] highlighted the high FOS production observed in sugar cane bagasse, which was attributed to its high-water absorption index (WAI) facilitating microorganism growth and cell immobilization. However, recent findings from [57]and [58] challenged the notion that physicochemical parameters alone determine FOS production. They observed that certain microorganisms, such as A. tamarii URM4634 and Aspergillus niger LBA 02, exhibited low WAI but still showed high enzyme activity. This suggests that factors beyond physicochemical parameters, including nutrient absorption and enzyme production, contribute to the complex process of FOS production during fermentation.
According to Davani-Davari et al. (2019), during the fermentation process, β-galactosidase can undergo transgalactosylation mechanism to produce GOS. The GOS can be produced through the binding of galactose molecules to lactose where 3-6 pieces of saccharides bind with β (1-4; 1-6) bonds to 2-5 parts of galactose units (Demirci et al., 2017). Optimizing temperature is crucial for maximizing β-galactosidase activity and GOS production in different microbial strains. For instance, Penicillium sp. exhibited high activity at 50°C, while LAB strains, like L. bulgaricus, thrived at 45°C. Moreover, strain selection plays a vital role, with P. acidilactici showing enhanced β-galactosidase activity and GOS production [60].
Bioactive peptides (BP) are specific protein fragments that contribute to overall body health. The activity of these peptides is influenced by their inherent amino acid composition and sequence. Initially inactive in the parent protein, BP can be generated through microbial fermentation, leading to functional food benefits, as highlighted by Taniguchi et al., (2018). Various microorganisms possess different proteolytic systems, and common probiotic strains like L. plantarum, L. acidophilus, Aspergillus oryzae, and B. subtilis are known to undergo proteolytic [62]
Fermentation of various food substrates using specific probiotic strains has been shown to enhance the production of BP with potential health benefits. Ayyash et. al (2018) investigated the fermentation of camel milk using L. lactis KX881782 and observed a significant inhibition of α-amylase and α-glucosidase, indicating potential antidiabetic activity. Similarly, Flores-Medellín et al. (2020) found that fermented black bean with B. subtilis exhibited beneficial effects in obesity and type 2 diabetes mellitus. The mechanism of action of BP involves the inactivation of specific enzymes such as α-amylase, α-glucosidase, and dipeptidyl peptidase-IV (DPP-IV), as discussed by Yan et al. (2019). Mazorra-Manzano et al. (2020) demonstrated an increase in ACE-inhibitory activity in cheese whey after fermentation, while Wu et. al. (2017) highlighted the inhibition of angiotensin-I-converting enzyme (ACE) as a mechanism for antihypertensive effects. Furthermore, Najafian and Babji (2018) and Wu et al. (2017) reported antioxidant activities in fermented fish and salmon skin, respectively. These findings underscore the potential of fermentation in generating BP with various health-promoting properties.
The production of BP through microbial fermentation is influenced by the duration of fermentation, as observed by Sharma et al., (2020). They found that the highest proteolytic activity (30.38 mg leucine/mL) in flaxseed milk supplemented with L. plantarum NCDC 374 was detected at 126 hours of fermentation, with a subsequent decrease in proteolytic activity at later stages. Similar trends were reported by Liu et al. (2017) in their study on defatted wheat germ (DWG) fermented with B. subtilis 10160. The peptide content increased from 0 hours (4.31 ± 0.59%) to 48 hours (29.68 ± 0.98%) and then decreased to 25.80 ± 1.49% at 72 hours of fermentation. This observation aligns with the findings of Ying and Voo (2020), who emphasized that the fermentation process can enhance bioactive peptide levels in food and beverages through the proteolytic activity of microorganisms. Maffioli et al., (2020) discussed the interaction between endopeptidases and exopeptidases, noting that with longer fermentation durations, peptides produced by endopeptidases can further hydrolyze into amino acids, resulting in lower peptide levels (Figure 2).
Furthermore, Panchal et al., (2020) and Peres et al. (2017) both reported the highest proteolytic activity and bioactive peptide production in goat milk fermented with L. fermentum and L. helveticus IMAU80872, respectively, under specific fermentation conditions. It has been suggested by Raveschot et al. (2018) that Lactobacillus strains develop proteolytic systems to hydrolyze proteins into various bioactive peptides (Figure 3). However, the BP profiles can vary among different Lactobacillus strains due to differences in CEP gene expression, CEP gene mutations, and enzymatic activities under specific optimum conditions.
Short-Chain Fatty Acids (SCFAs) and Conjugated Fatty Acids (CLA)
Short-chain fatty acids (SCFAs) are end-products of microbial fermentation in the gut and play a crucial role in host health. Acetate, propionate, and butyrate are the main SCFAs produced through the breakdown of dietary fibers and other complex carbohydrates by gut microbiota. SCFAs have diverse physiological effects, including energy metabolism, immune modulation, and maintenance of gut barrier integrity. However, various fermented foods, such as from dairy and vegetable-based fermented foods were shown to also be rich in these substrates. Dietary SCFA has been shown to exert multiple health benefits, such as reduced risk of metabolic perturbations, obesity, improved lipid synthesis and microbial composition [76].
Numerous studies have investigated the production of short-chain fatty acids (SCFAs) through microbial fermentation in various food substrates. For instance, Jia et al. (2016) observed increased SCFA production (11.74 ± 0.08 mg mL−1) during the fermentation of goat milk yogurt with Lactobacillus rhamnosus GG. Similarly, Hu et al. (2019) reported elevated levels of acetic acid, propionic acid, and butyric acid in carrot juice fermented with L. rhamnosus GG. However, Wang et al. (2020) argued that the correlation between SCFA production and L. rhamnosus strains may not be direct.
Lee et al. (2020) demonstrated higher SCFA production in fermented rice liquors supplemented with barley compared to control alcoholic drinks. The supplemented group showed increased butyric acid (0.09 nmol/g dried fecal), propionic acid (0.11 nmol/g dried fecal), and acetic acid (0.45 nmol/g dried fecal) levels, while the control group had lower levels (butyric acid: 0.05 nmol/g dried fecal, propionic acid: 0.06 nmol/g dried fecal, acetic acid: 0.10 nmol/g dried fecal). Similarly, Pérez-Burillo et al. (2019) found that fermenting salami with citrus fibers enhanced SCFA production (acetate: 66%, propionate: 20%, butyrate: 58%) compared to the control.
These studies suggest that fermentation with dietary fiber can serve as an effective substrate for probiotics, leading to increased SCFA production [82]. The modulation of gut flora by fermented beverages has potential implications, as SCFAs have been found to activate G protein-coupled receptors (GPCRs) and influence glycemic response, improving glucose tolerance and insulin release Alexander et al., (2019) and. Butyrate, a major SCFA derived from Acetyl-CoA through the glycolysis of dietary carbohydrates, plays a significant role in these physiological effects (Erik et al., 2018).
Fermentation also plays a role in the conversion of linoleic acid (LA) into conjugated linoleic acid (CLA), a beneficial modified fat. Microorganisms such as Bifidobacterium, Enterococcus, Lactobacillus, Lactococcus, Propionibacterium, and Streptococcus can convert LA into CLA through linoleate isomerase enzyme activity. CLA synthesis can occur through direct isomerization of LA or via 10-hydroxy-octadecenoic acid in certain Lactobacillus, Propionibacterium, Bifidobacterium, and Clostridium-like bacteria found in the human gut [85]. CLA production has been associated with reducing LDL cholesterol levels, enhancing immune function and osteogenesis, preventing hyperinsulinemia, and protecting against colon cancer [86].
The production of conjugated CLA through microbial fermentation has been investigated in various food products. For instance, Hwang et al. (2021) observed the production of cis-9, trans-11 CLA and trans-10, cis-12 CLA during the fermentation of soybean powder yogurt (SPY) with L. plantarum, while no CLA production was detected with L. brevis WCP02. Similarly, [88] found efficient CLA production in semi-hydrated sausages fermented with L. plantarum AB20–961. Furthermore, Palachum et al., (2020) developed gummy jelly using guava pulp fermented with probiotic L. plantarum WU-P19. L. plantarum has been recognized as a probiotic capable of producing CLA Khosravi et al. (2015), along with other microorganisms such as Bifidobacterium, Lactobacillus, Streptococcus, Propionibacterium, and Clostridium [85]
Temperature has been identified as a factor influencing CLA production. Amiri et al. (2020) demonstrated that the highest CLA production (105.08 μg/ml) in whey cheese supplemented with Bifidobacterium lactis BB12 occurred at 34°C for 60 hours. In contrast, at the same fermentation duration (60 hours) but at 42°C, the CLA production was lower (67.66 μg/ml). Another study by Amiri et al. (2020) showed that the highest CLA production during the fermentation of cheese whey with L. acidophilus was at 38°C for 60 hours (38.69 μg/ml), while at 34°C, the production was lower (19.01 μg/ml). Temperature plays a crucial role in microbial growth and metabolism during fermentation. The optimum temperature for the growth and metabolism of L. acidophilus in synthesizing CLA from linoleic acid (LA) is around 37°C [91]. The temperature also affects the isomerase capability of lactic acid bacteria in transforming LA into CLA, as high temperatures can denature the isomerase enzyme and reduce CLA production [92].
Conclusion and Future Prospects
Future directions in this field involve further exploration of the potential of different microorganisms and their interactions to enhance the production of these components. Additionally, investigating the impact of fermentation on other bioactive compounds and nutritional profiles would provide a more comprehensive understanding of the benefits of fermented biofortified foods. Advances in microbiome research and functional genomics can also contribute to a deeper understanding of the mechanisms involved in microbial fermentation and its impact on human health. Finally, efforts should be directed towards scaling up and translating these findings into practical applications, such as developing novel functional food products that can be easily incorporated into everyday diets, to address nutritional deficiencies and promote human health.
Acknowledgement
This research was supported by Universiti Putra Malaysia Inisiatif Putra Siswazah Grant, with a reference of UPM.RMC.800-2/1/2022/GP-IPS/9740400.
Conflicts of Interest
No conflict of interest.
References
- Huizar, M.I.; Arena, R.; Laddu, D.R. The Global Food Syndemic: The Impact of Food Insecurity, Malnutrition and Obesity on the Healthspan amid the COVID-19 Pandemic. Prog. Cardiovasc. Dis. 2020, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Alemu, E.A. Malnutrition and Its Implications on Food Security. 2020, 509–518. [CrossRef]
- Chadare, F.J.; Idohou, R.; Nago, E.; Affonfere, M.; Agossadou, J.; Fassinou, T.K.; Kénou, C.; Honfo, S.; Azokpota, P.; Linnemann, A.R.; et al. Conventional and Food-to-Food Fortification: An Appraisal of Past Practices and Lessons Learned. Food Sci. Nutr. 2019, 7, 2781–2795. [Google Scholar] [CrossRef] [PubMed]
- Ellies, W.; Rolfes, S. Understanding Nutrition; 15th, *!!! REPLACE !!!* (Eds.) 2019.
- Rahim, M.H.A.; Hazrin-Chong, N.H.; Harith, H.H.; Wan-Mohtar, W.A.A.Q.I.; Sukor, R. Roles of Fermented Plant-, Dairy- and Meat-Based Foods in the Modulation of Allergic Responses. Food Sci. Hum. Wellness 2023, 12, 691–701. [Google Scholar] [CrossRef]
- Abedelazeez, K.J.D.; Solehah, M.Z.N.; Jaafar, A.H.; Meor Hussin, A.S.; Wan-Mohtar, W.A.A.Q.I.; Abd Rahim, M.H. Production, Organoleptic, and Biological Activities of Belacan (Shrimp Paste) and Pekasam (Fermented Freshwater Fish), the Ethnic Food from the Malay Archipelago. Sains Malaysiana 2023, 52, 1217–1230. [Google Scholar]
- Faizal, F.A.; Ahmad, N.H.; Yaacob, J.S.; Abdul-Halim Lim, S.; Yaacob, J.S.; Abd Rahim, M.H. Food Processing to Reduce Anti-Nutrients in Plant-Based Food. Int. Food Res. J. 2023, 30, 25–45. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.A.A.Q.I.; Ab Kadir, S.; Halim-Lim, S.A.; Ilham, Z.; Hajar-Azhari, S.; Saari, N. Vital Parameters for High Gamma-Aminobutyric Acid (GABA) Production by an Industrial Soy Sauce Koji Aspergillus Oryzae NSK in Submerged-Liquid Fermentation. Food Sci. Biotechnol. 2019, 28, 1747–1757. [Google Scholar] [CrossRef]
- Wan-Mohtar, W.A.A.Q.I.; Ab Kadir, S.; Saari, N. The Morphology of Ganoderma Lucidum Mycelium in a Repeated-Batch Fermentation for Exopolysaccharide Production. Biotechnol. Reports 2016, 11, 2–11. [Google Scholar] [CrossRef]
- Hajar-Azhari, S.; Wan-Mohtar, W.A.A.Q.I.; Ab Kadir, S.; Rahim, M.H.A.; Saari, N. Evaluation of a Malaysian Soy Sauce Koji Strain Aspergillus Oryzae NSK for γ-Aminobutyric Acid (GABA) Production Using Different Native Sugars. Food Sci. Biotechnol. 2018, 27, 479–488. [Google Scholar] [CrossRef]
- Shin Yee, C.; Sohedein, M.N.A.; Poh Suan, O.; Weng Loen, A.W.; Abd Rahim, M.H.; Soumaya, S.; Ilham, Z.; Wan-Mohtar, W.A.A.Q.I. The Production of Functional γ-Aminobutyric Acid Malaysian Soy Sauce Koji and Moromi Using the Trio of Aspergillus Oryzae NSK, Bacillus Cereus KBC, and the Newly Identified Tetragenococcus Halophilus KBC in Liquid-State Fermentation. Futur. Foods 2021, 4, 100055. [Google Scholar] [CrossRef]
- Abd Rahim, M.H. Production Of Lovastatin, (+)-Geodin And Sulochrin By Aspergillus Terreus ATCC 20542 Using Pure And Crude Glycerol, The University of Sydney, 2015.
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as Vitamin Suppliers to Their Host: A Gut Microbiota Perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System–Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Sadh, P.K.; Kumar, S.; Chawla, P.; Duhan, J.S. Fermentation: A Boon for Production of Bioactive Compounds by Processing of Food Industries Wastes (By-Products); 2018; Vol. 23; ISBN 9116662431.
- Zhang, Z.; Xia, L.; Wang, F.; Lv, P.; Zhu, M.; Li, J.; Chen, K. Lignin Degradation in Corn Stalk by Combined Method of H2O2 Hydrolysis and Aspergillus Oryzae CGMCC5992 Liquid-State Fermentation. Biotechnol. Biofuels 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lazarte, C.E.; Vargas, M.; Granfeldt, Y. Zinc Bioavailability in Rats Fed a Plant-Based Diet: A Study of Fermentation and Zinc Supplementation. Food Nutr. Res. 2015, 59. [Google Scholar] [CrossRef]
- Amiri, S.; Rezaei Mokarram, R.; Sowti Khiabani, M.; Rezazadeh Bari, M.; Alizadeh Khaledabad, M. In Situ Production of Conjugated Linoleic Acid by Bifidobacterium Lactis BB12 and Lactobacillus Acidophilus LA5 in Milk Model Medium. Lwt 2020, 132, 109933. [Google Scholar] [CrossRef]
- Chagas Junior, G.C.A.; Ferreira, N.R.; Gloria, M.B.A.; Martins, L.H. da S.; Lopes, A.S. Chemical Implications and Time Reduction of On-Farm Cocoa Fermentation by Saccharomyces Cerevisiae and Pichia Kudriavzevii. Food Chem. 2021, 338, 127834. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.O.; Önning, G.; Holmgren, K.; Strandler, H.S.; Hultberg, M. Fermentation of Cauliflower and White Beans with Lactobacillus Plantarum – Impact on Levels of Riboflavin, Folate, Vitamin B12, and Amino Acid Composition. Plant Foods Hum. Nutr. 2020, 75, 236–242. [Google Scholar] [CrossRef]
- Carrizo, S.L.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Rollán, G.C. Quinoa Pasta Fermented with Lactic Acid Bacteria Prevents Nutritional Deficiencies in Mice. Food Res. Int. 2020, 127, 108735. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, W.; Shen, M.; Kang, Z.; Wang, J.; Quan, Z.; Xiao, J.; Zhao, S.; Liu, D.; Cao, L. Effect of Natural Fermentation of Sorghum on Resistant Starch Molecular Structure and Fermentation Property. J. Chem. 2020, 2020. [Google Scholar] [CrossRef]
- Laiño, J.E.; Juarez del Valle, M.; Savoy de Giori, G.; LeBlanc, J.G.J. Development of a High Folate Concentration Yogurt Naturally Bio-Enriched Using Selected Lactic Acid Bacteria. LWT - Food Sci. Technol. 2013, 54, 1–5. [Google Scholar] [CrossRef]
- Innate, T.; Cells, L. Streptococcus Thermophilus 1131 Induce the Expression of the REG3 Family in the Small Intestine of Mice via the Stimulation of Dendritic Cells And. 2019, 1–11.
- Clara, A.; Silvetti, T.; Milena, B. Folate Production by LAB and Their Potential for Dairy Product Bio-Enrichment by Fermentation.
- Fang, H.; Kang, J.; Zhang, D. Microbial Production of Vitamin B12: A Review and Future Perspectives. Microb. Cell Fact. 2017, 16, 15. [Google Scholar] [CrossRef]
- Santos, F.; Vera, J.L.; van der Heijden, R.; Valdez, G.; de Vos, W.M.; Sesma, F.; Hugenholtz, J. The Complete Coenzyme B12 Biosynthesis Gene Cluster of Lactobacillus Reuteri CRL1098. Microbiology 2008, 154, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Coda, R.; Chamlagain, B.; Varmanen, P.; Piironen, V.; Katina, K. Co-Fermentation of Propionibacterium Freudenreichiiand Lactobacillus Brevisin Wheat Bran for in Situproduction of Vitamin B12. Front. Microbiol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deptula, P.; Chamlagain, B.; Edelmann, M.; Sangsuwan, P.; Nyman, T.A.; Savijoki, K.; Piironen, V.; Varmanen, P. Food-like Growth Conditions Support Production of Active Vitamin B12 by Propionibacterium Freudenreichii 2067 without DMBI, the Lower Ligand Base, or Cobalt Supplementation. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wolkers – Rooijackers, J.C.M.; Endika, M.F.; Smid, E.J. Enhancing Vitamin B12 in Lupin Tempeh by in Situ Fortification. LWT 2018, 96, 513–518. [Google Scholar] [CrossRef]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kieliszek, M.; Ścibisz, I. Propionibacterium Spp.—Source of Propionic Acid, Vitamin B12, and Other Metabolites Important for the Industry. Appl. Microbiol. Biotechnol. 2018, 102, 515–538. [Google Scholar] [CrossRef]
- Walther, B.; Karl, J.P.; Booth, S.L.; Boyaval, P. Menaquinones, Bacteria, and the Food Supply: The Relevance of Dairy and Fermented Food Products to Vitamin K Requirements. Adv. Nutr. 2013, 4, 463–473. [Google Scholar] [CrossRef]
- Pateiro, M.; Munekata, P.E.S.; Tsatsanis, C.; Domínguez, R.; Zhang, W.; Barba, F.J.; Lorenzo, J.M. Evaluation of the Protein and Bioactive Compound Bioaccessibility/Bioavailability and Cytotoxicity of the Extracts Obtained from Aquaculture and Fisheries by-Products. Adv. Food Nutr. Res. 2020, 92, 97–125. [Google Scholar] [CrossRef]
- Desai, N.M.; Martha, G.S.; Harohally, N. V.; Murthy, P.S. Non-Digestible Oligosaccharides of Green Coffee Spent and Their Prebiotic Efficiency. Lwt 2020, 118. [Google Scholar] [CrossRef]
- Kumar, J.; Rani, K.; Datt, C. Molecular Link between Dietary Fibre, Gut Microbiota and Health. Mol. Biol. Rep. 2020, 47, 6229–6237. [Google Scholar] [CrossRef]
- Xia, W.; Khan, I.; Li, X. ang; Huang, G.; Yu, Z.; Leong, W.K.; Han, R.; Ho, L.T.; Wendy Hsiao, W.L. Adaptogenic Flower Buds Exert Cancer Preventive Effects by Enhancing the SCFA-Producers, Strengthening the Epithelial Tight Junction Complex and Immune Responses. Pharmacol. Res. 2020, 159, 104809. [Google Scholar] [CrossRef]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Somboonpanyakul, P. B Vitamins and Prebiotic Fructooligosaccharides of Cashew Apple Fermented with Probiotic Strains Lactobacillus Spp., Leuconostoc Mesenteroides and Bifidobacterium Longum. Process Biochem. 2018, 70, 9–19. [Google Scholar] [CrossRef]
- Albuquerque, M.A.C.; Bedani, R.; LeBlanc, J.G.; Saad, S.M.I. Passion Fruit By-Product and Fructooligosaccharides Stimulate the Growth and Folate Production by Starter and Probiotic Cultures in Fermented Soymilk. Int. J. Food Microbiol. 2017, 261, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Hajar-Azhari, S.; Hafiz Abd Rahim, M.; Razid Sarbini, S.; Muhialdin, B.J.; Olusegun, L.; Saari, N. Enzymatically Synthesised Fructooligosaccharides from Sugarcane Syrup Modulate the Composition and Short-Chain Fatty Acid Production of the Human Intestinal Microbiota. Food Res. Int. 2021, 149, 110677. [Google Scholar] [CrossRef] [PubMed]
- Hajar-Azhari, S.; Rahim, M.H.A.; Wan-Mohtar, W.A.A.Q.I.; Sarbini, S.R.; Saari, N. Novel Fructooligosaccharide Conversion from Sugarcane Syrup Using a Specialised Enzymatic PH-Stat Bioreactor. Process Biochem. 2020, 95, 55–63. [Google Scholar] [CrossRef]
- Bhadra, S.; Chettri, D.; Verma, A.K. Microbes in Fructooligosaccharides Production. Bioresour. Technol. Reports 2022, 20, 101159. [Google Scholar] [CrossRef]
- Sánchez-Martínez, M.J.; Soto-Jover, S.; Antolinos, V.; Martínez-Hernández, G.B.; López-Gómez, A. Manufacturing of Short-Chain Fructooligosaccharides: From Laboratory to Industrial Scale. Food Eng. Rev. 2020, 12, 149–172. [Google Scholar] [CrossRef]
- Celligoi, M.A.; Baratella, P. Optimization of High Production of Fructooligosaccharides by Sucrose Fermentation of Bacillus Subtilis Natto CCT 7712. Am. J. Food Technol. 2014, 9, 144. [Google Scholar]
- Tezgel, N.; Kırtel, O.; Van den Ende, W.; Toksoy Oner, E. Fructosyltransferase Enzymes for Microbial Fructan Production. Microb. Enzym. Roles Appl. Ind. 2020, 1–39. [Google Scholar]
- Xie, Y.; Zhou, H.; Liu, C.; Zhang, J.; Li, N.; Zhao, Z.; Sun, G.; Zhong, Y. A Molasses Habitat-Derived Fungus Aspergillus Tubingensis XG21 with High β-Fructofuranosidase Activity and Its Potential Use for Fructooligosaccharides Production. AMB Express 2017, 7, 128. [Google Scholar] [CrossRef]
- Guerra, L.; Ureta, M.; Romanini, D.; Woitovich, N.; Gómez-Zavaglia, A.; Clementz, A. Enzymatic Synthesis of Fructooligosaccharides: From Carrot Discards to Prebiotic Juice. Food Res. Int. 2023, 170, 112991. [Google Scholar] [CrossRef]
- de Jesus, L.F.M.C.; Guimarães, L.H.S. Production of β-Galactosidase by Trichoderma Sp. through Solid-State Fermentation Targeting the Recovery of Galactooligosaccharides from Whey Cheese. J. Appl. Microbiol. [CrossRef]
- Bassetto, R.Z.; Cabral, P.S.; Silveira, M.H.L.; Almeida, M.M.; Chiquetto, N.C. Optimization of β-Galactosidase Production from Penicillium Sp. for Synthesis of Galactooligosaccharides. Int. Food Res. J. 2017, 24, 1793–1798. [Google Scholar]
- Fara, A.; Sabater, C.; Palacios, J.; Requena, T.; Montilla, A.; Zárate, G. Prebiotic Galactooligosaccharides Production from Lactose and Lactulose by: Lactobacillus Delbrueckii Subsp. Bulgaricus CRL450. Food Funct. 2020, 11, 5875–5886. [Google Scholar] [CrossRef] [PubMed]
- Zokaityte, E.; Cernauskas, D.; Klupsaite, D.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Ruzauskas, M.; Gruzauskas, R.; Juodeikiene, G.; Rocha, J.M.; et al. Bioconversion of Milk Permeate with Selected Lactic Acid Bacteria Strains and Apple By-Products into Beverages with Antimicrobial Properties and Enriched with Galactooligosaccharides. Microorganisms 2020, 8, 1–26. [Google Scholar] [CrossRef]
- Oh, N.S.; Kim, K.; Oh, S.; Kim, Y. Enhanced Production of Galactooligosaccharides Enriched Skim Milk and Applied to Potentially Synbiotic Fermented Milk with <italic>Lactobacillus Rhamnosus</Italic> 4B15. Food Sci. Anim. Resour. 2019, 39, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Vénica, C.I.; Bergamini, C. V; Rebechi, S.R.; Perotti, M.C. Galacto-Oligosaccharides Formation during Manufacture of Different Varieties of Yogurt. Stability through Storage. LWT - Food Sci. Technol. 2015, 63, 198–205. [Google Scholar] [CrossRef]
- Yañez-Ñeco, C. V; Rodriguez-Colinas, B.; Amaya-Delgado, L.; Ballesteros, A.O.; Gschaedler, A.; Plou, F.J.; Arrizon, J. Galactooligosaccharide Production from Pantoea Anthophila Strains Isolated from “Tejuino”, a Mexican Traditional Fermented Beverage. Catalysts 2017, 7. [Google Scholar] [CrossRef]
- Fischer, C.; Kleinschmidt, T. Valorisation of Sweet Whey by Fermentation with Mixed Yoghurt Starter Cultures with Focus on Galactooligosaccharide Synthesis. Int. Dairy J. 2021, 119, 105068. [Google Scholar] [CrossRef]
- de la Rosa, O.; Múñiz-Marquez, D.B.; Contreras-Esquivel, J.C.; Wong-Paz, J.E.; Rodríguez-Herrera, R.; Aguilar, C.N. Improving the Fructooligosaccharides Production by Solid-State Fermentation. Biocatal. Agric. Biotechnol. 2020, 27, 101704. [Google Scholar] [CrossRef]
- Ganaie, M.A.; Soni, H.; Naikoo, G.A.; Santos Oliveira, L.T.; Rawat, H.K.; Mehta, P.K.; Narain, N. Screening of Low Cost Agricultural Wastes to Maximize the Fructosyltransferase Production and Its Applicability in Generation of Fructooligosaccharides by Solid State Fermentation. Int. Biodeterior. Biodegrad. 2017, 118, 19–26. [Google Scholar] [CrossRef]
- Oliveira, R.L. de; Silva, M.F. da; Converti, A.; Porto, T.S. Production of β-Fructofuranosidase with Transfructosylating Activity by Aspergillus Tamarii URM4634 Solid-State Fermentation on Agroindustrial by-Products. Int. J. Biol. Macromol. 2020, 144, 343–350. [Google Scholar] [CrossRef]
- Ohara, A.; Soares de Castro, R.J.; Goia Nishide, T.; Gonçalves Dias, F.F.; Pavan Bagagli, M.; Harumi Sato, H. Invertase Production by Aspergillus Niger under Solid State Fermentation: Focus on Physical-Chemical Parameters, Synergistic and Antagonistic Effects Using Agro-Industrial Wastes. Biocatal. Agric. Biotechnol. 2015, 4, 645–652. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 1–27. [Google Scholar] [CrossRef]
- Chanalia, P.; Gandhi, D.; Attri, P.; Dhanda, S. Purification and Characterization of β-Galactosidase from Probiotic Pediococcus Acidilactici and Its Use in Milk Lactose Hydrolysis and Galactooligosaccharide Synthesis. Bioorg. Chem. 2018, 77, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, M.; Aida, R.; Saito, K.; Ochiai, A.; Takesono, S.; Saitoh, E.; Tanaka, T. Identi Fi Cation and Characterization of Multifunctional Cationic Peptides from Traditional Japanese Fermented Soybean Natto Extracts. J. Biosci. Bioeng. 2018, xx. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of Bioactive Peptides by Lactobacillus Species: From Gene to Application. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Al-Dhaheri, A.S.; Al Mahadin, S.; Kizhakkayil, J.; Abushelaibi, A. In Vitro Investigation of Anticancer, Antihypertensive, Antidiabetic, and Antioxidant Activities of Camel Milk Fermented with Camel Milk Probiotic: A Comparative Study with Fermented Bovine Milk. J. Dairy Sci. 2018, 101, 900–911. [Google Scholar] [CrossRef]
- Flores-Medellín, S.A.; Camacho-Ruiz, R.M.; Guízar-González, C.; Rivera-Leon, E.A.; Llamas-Covarrubias, I.M.; Mojica, L. Protein Hydrolysates and Phenolic Compounds from Fermented Black Beans Inhibit Markers Related to Obesity and Type-2 Diabetes. Legum. Sci. 2020, 1–17. [Google Scholar] [CrossRef]
- Yan, J.; Zhao, J.; Yang, R.; Zhao, W. Bioactive Peptides with Antidiabetic Properties: A Review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese Whey Fermentation by Its Native Microbiota: Proteolysis and Bioactive Peptides Release with ACE-Inhibitory Activity. Fermentation 2020, 6, 1–12. [Google Scholar] [CrossRef]
- Wu, J.; Liao, W.; Udenigwe, C.C. Revisiting the Mechanisms of ACE Inhibitory Peptides from Food Proteins. Trends Food Sci. Technol. 2017, 69, 214–219. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Fractionation and Identification of Novel Antioxidant Peptides from Fermented Fish (Pekasam). J. Food Meas. Charact. 2018, 12, 2174–2183. [Google Scholar] [CrossRef]
- Wu, R.; Chen, L.; Liu, D.; Huang, J.; Zhang, J.; Xiao, X.; Lei, M.; Chen, Y.; He, H. Preparation of Antioxidant Peptides from Salmon Byproducts with Bacterial Extracellular Proteases. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, D.; Kaur, S.; Borah, A. Optimization of Flaxseed Milk Fermentation for the Production of Functional Peptides and Estimation of Their Bioactivities. Food Sci. Technol. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chen, Z.; Shao, J.; Wang, C.; Zhan, C. Effect of Fermentation on the Peptide Content, Phenolics and Antioxidant Activity of Defatted Wheat Germ. Food Biosci. 2017, 20, 141–148. [Google Scholar] [CrossRef]
- Ying, A.; Voo, H. Bioactive Peptides from Food Fermentation : A Comprehensive Review of Their Sources, Bioactivities, Applications, and Future Development. 2020, 1–61. [CrossRef]
- Maffioli, E.; Jiang, Z.; Nonnis, S.; Negri, A.; Romeo, V.; Lietz, C.B.; Hook, V.; Ristagno, G.; Baselli, G.; Kistler, E.B.; et al. High-Resolution Mass Spectrometry-Based Approaches for the Detection and Quantification of Peptidase Activity in Plasma. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Panchal, G.; Hati, S.; Sakure, A. Characterization and Production of Novel Antioxidative Peptides Derived from Fermented Goat Milk by L. Fermentum; Elsevier Ltd, 2020; Vol. 119; ISBN 9409669561.
- Peres, C.M.; Peres, C.; Garcia, H.S. Assessment of Multifunctional Activity of Bioactive Peptides Derived from Fermented Milk by Specific Lactobacillus Plantarum Strains. J. Dairy Sci. 2017, 1–11. [Google Scholar] [CrossRef]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary Short-Chain Fatty Acid Intake Improves the Hepatic Metabolic Condition via FFAR3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef]
- Jia, R.; Chen, H.; Chen, H.; Ding, W. Effects of Fermentation with Lactobacillus Rhamnosus GG on Product Quality and Fatty Acids of Goat Milk Yogurt. J. Dairy Sci. 2016, 99, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zeng, F.; Wu, L.; Wan, X.; Chen, Y.; Zhang, J.; Liu, B. Fermented Carrot Juice Attenuates Type 2 Diabetes by Mediating Gut Microbiota in Rats. Food Funct. 2019, 10, 2935–2946. [Google Scholar] [CrossRef]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus Rhamnosus Strains Relieve Loperamide-Induced Constipation via Different Pathways Independent of Short-Chain Fatty Acids. Front. Cell. Infect. Microbiol. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Lee, J.E.; Ha, J.S.; Park, H.Y.; Lee, E. Alteration of Gut Microbiota Composition by Short-Term Low-Dose Alcohol Intake Is Restored by Fermented Rice Liquor in Mice. Food Res. Int. 2020, 128, 108800. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Mehta, T.; Pastoriza, S.; Kramer, D.L.; Paliy, O.; Rufián-Henares, J.Á. Potential Probiotic Salami with Dietary Fiber Modulates Antioxidant Capacity, Short Chain Fatty Acid Production and Gut Microbiota Community Structure. Lwt 2019, 105, 355–362. [Google Scholar] [CrossRef]
- Bai, Y.; Zhao, J. biao; Tao, S. yu; Zhou, X. jian; Pi, Y.; Gerrits, W.J.J.; Johnston, L.J.; Zhang, S. yi; Yang, H. jian; Liu, L.; et al. Effect of Dietary Fiber Fermentation on Short-Chain Fatty Acid Production and Microbial Composition in Vitro. J. Sci. Food Agric. 2020, 100, 4282–4291. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Swanson, K.S.; Fahey, G.C.; Garleb, K.A. Perspective : Physiologic Importance of Short-Chain Fatty Acids from Nondigestible Carbohydrate Fermentation. 2019, 576–589.
- Erik, K.; Knudsen, B.; Lærke, H.N.; Hedemann, M.S.; Nielsen, T.S.; Ingerslev, A.K.; Søvsø, D.; Nielsen, G.; Theil, P.K.; Purup, S.; et al. Intestinal Barrier Function and Inflammation. [CrossRef]
- Salsinha, A.S.; Pimentel, L.L.; Fontes, A.L.; Gomes, A.M.; Rodríguez-Alcalá, L.M. Microbial Production of Conjugated Linoleic Acid and Conjugated Linolenic Acid Relies on a Multienzymatic System. Microbiol. Mol. Biol. Rev. 2018, 82, 1–21. [Google Scholar] [CrossRef]
- Purba, R.A.P.; Paengkoum, P.; Paengkoum, S. The Links between Supplementary Tannin Levels and Conjugated Linoleic Acid (CLA) Formation in Ruminants: A Systematic Review and Meta-Analysis. PLoS One 2020, 15, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.E.; Kim, S.C.; Kim, D.H.; Lee, H.Y.; Suh, H.K.; Cho, K.M.; Lee, J.H. Enhancement of Isoflavone Aglycone, Amino Acid, and CLA Contents in Fermented Soybean Yogurts Using Different Strains: Screening of Antioxidant and Digestive Enzyme Inhibition Properties. Food Chem. 2021, 340, 128199. [Google Scholar] [CrossRef] [PubMed]
- Özer, C.O.; Kılıç, B. Optimization of PH, Time, Temperature, Variety and Concentration of the Added Fatty Acid and the Initial Count of Added Lactic Acid Bacteria Strains to Improve Microbial Conjugated Linoleic Acid Production in Fermented Ground Beef. Meat Sci. 2021, 171, 108303. [Google Scholar] [CrossRef]
- Palachum, W.; Choorit, W.; Manurakchinakorn, S.; Chisti, Y. Guava Pulp Fermentation and Processing to a Vitamin B12-Enriched Product. J. Food Process. Preserv. 2020, 44, 1–15. [Google Scholar] [CrossRef]
- Khosravi, A.; Safari, M.; Khodaiyan, F.; Gharibzahedi, S.M.T. Bioconversion Enhancement of Conjugated Linoleic Acid by Lactobacillus Plantarum Using the Culture Media Manipulation and Numerical Optimization. J. Food Sci. Technol. 2015, 52, 5781–5789. [Google Scholar] [CrossRef]
- Soto, C.; Soto, C. Lactobacillus Plantarum as Source of Conjugated Linoleic Acid: Effect of PH, Incubation Temperature and Inulin Incorporation. J. Biochem. Technol. 2013, 5, 649–653. [Google Scholar]
- Van Nieuwenhove, C.P.; Teran, V.; Nelina, S. Conjugated Linoleic and Linolenic Acid Production by Bacteria: Development of Functional Foods. Probiotics 2012. [Google Scholar] [CrossRef]
Figure 1.
The possible mechanisms of degradation of antinutrients in plant-based food [7].
Figure 1.
The possible mechanisms of degradation of antinutrients in plant-based food [7].
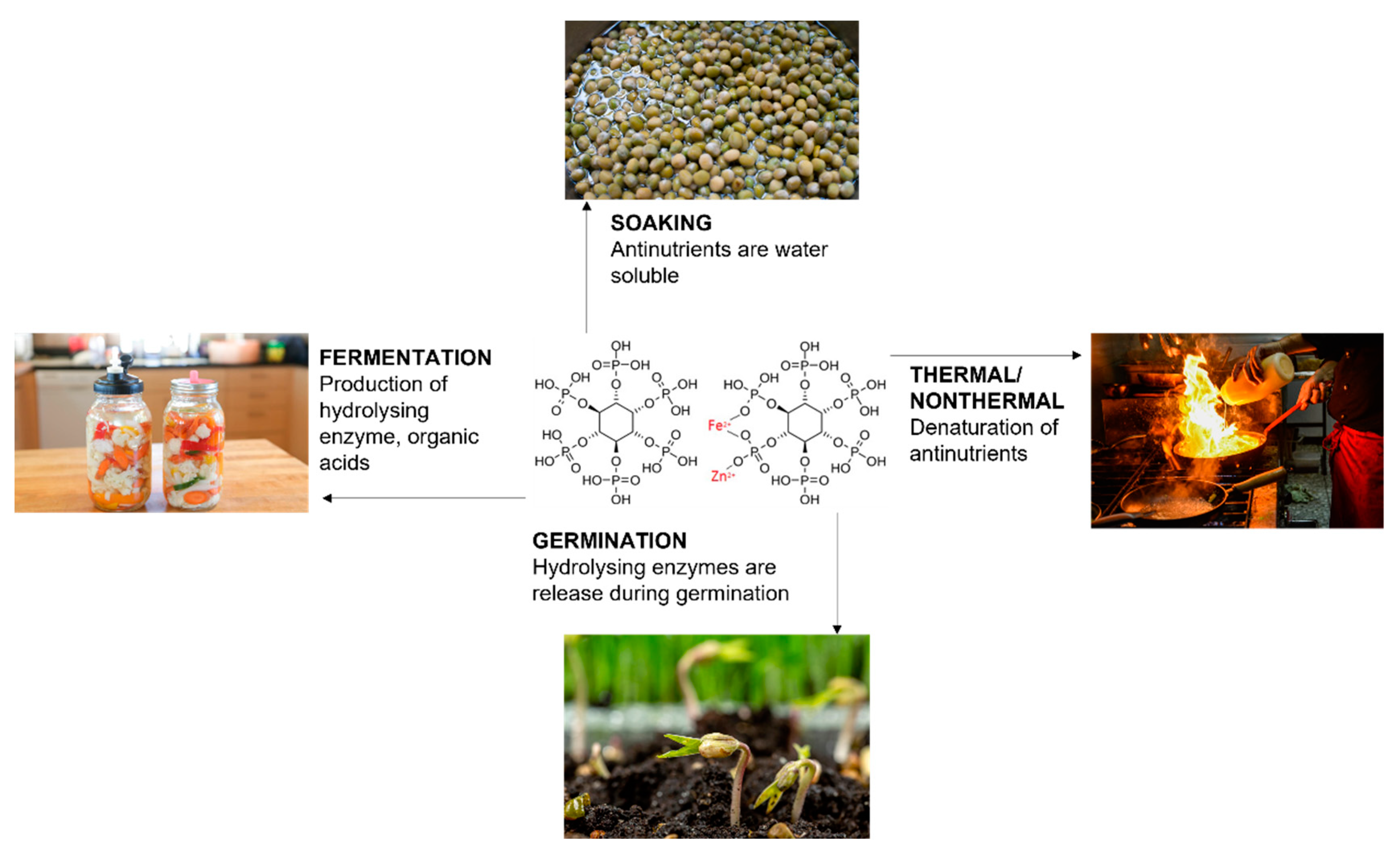
Figure 2.
The possible de novo biosynthesis of nutrients in microorganisms. The figure is based on an unpublished data (under review).
Figure 2.
The possible de novo biosynthesis of nutrients in microorganisms. The figure is based on an unpublished data (under review).
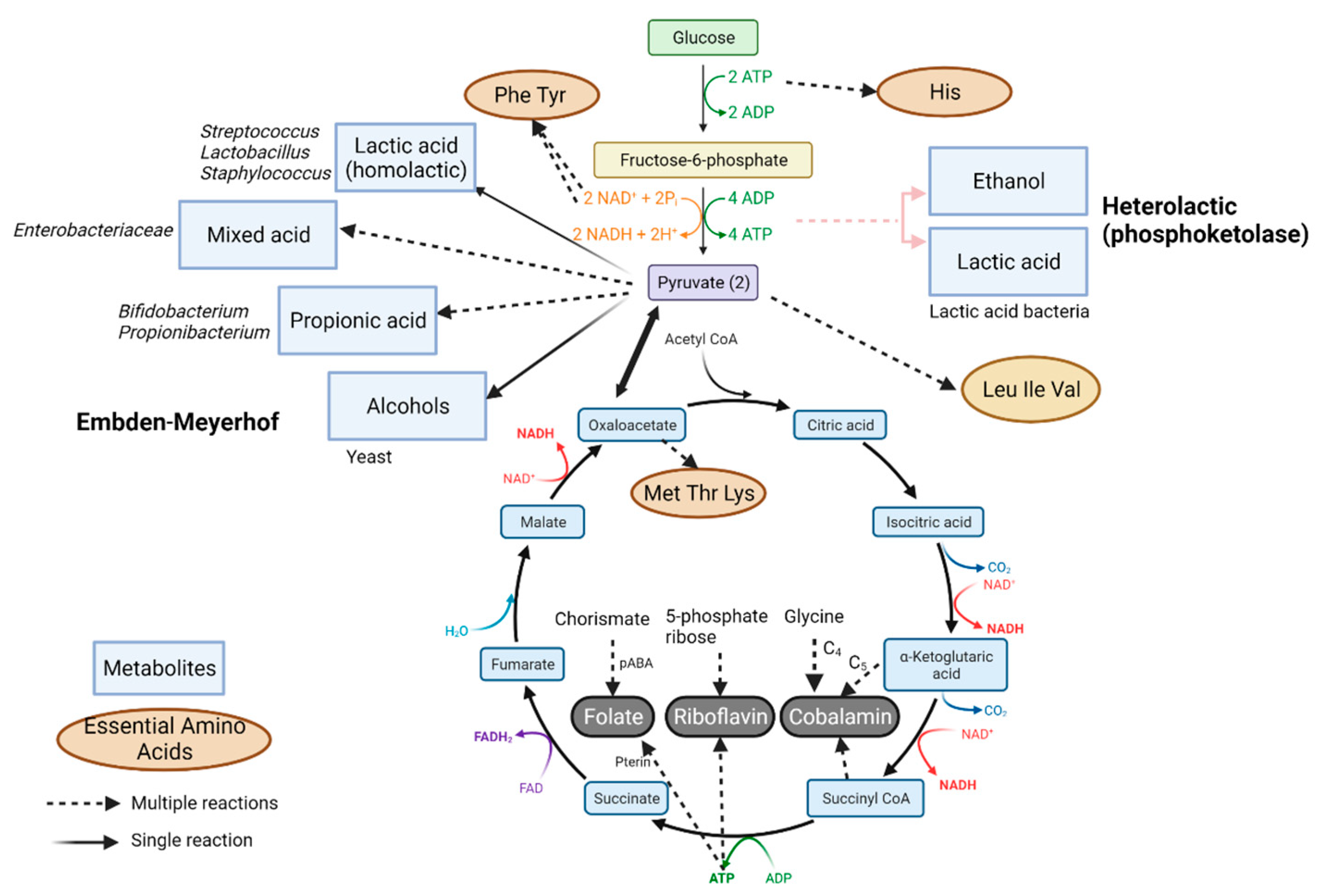
Figure 3.
(A) The proteolytic system of Lactobacillus spp. involves cell envelope proteinases (CEPs) and Prt enzymes that initiate protein hydrolysis. Peptides are transported inside the cell and further converted into free amino acids by peptidases. (B) the structure of CEPs from various Lactobacillus species, including different gene domains. The figure is adapted from by Raveschot et al. (2018).
Figure 3.
(A) The proteolytic system of Lactobacillus spp. involves cell envelope proteinases (CEPs) and Prt enzymes that initiate protein hydrolysis. Peptides are transported inside the cell and further converted into free amino acids by peptidases. (B) the structure of CEPs from various Lactobacillus species, including different gene domains. The figure is adapted from by Raveschot et al. (2018).
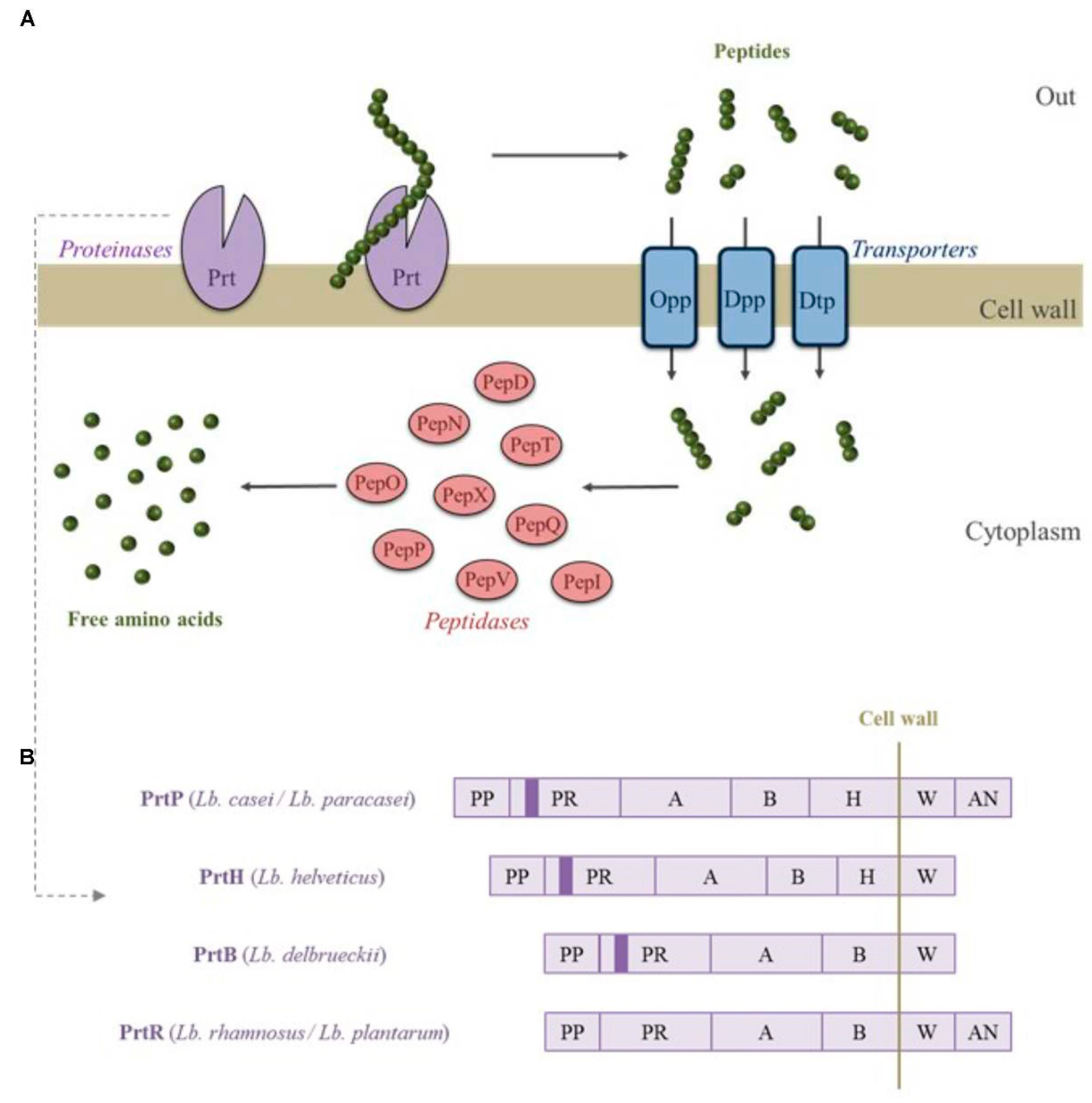

Table 1.
The prebiotic production from fermented food and beverages.
| Food item(s) | Microorganisms | Finding (s) | References |
|---|---|---|---|
| Fructooligosaccharides (FOS) | |||
| Cashew apple juice | L. acidophilus, L. casei, L. plantarum, Leuconostoc mesenteroides, and Bifidobacterium longum | The juice contains increased vitamin B-complex and prebiotics like FOS and oligosaccharides | [37] |
| Natto | Bacillus subtilis natto CCT 7712 | The optimal conditions for FOS formation ( 98.86 g/L) were a sucrose concentration of 300 g/L, pH 7.7, and agitation at 234 rpm | [43] |
| Sugarcane syrup | None (enzymatic synthesis) | Sucrose-rich sugarcane syrup have a prebiotic effect by promoting the growth of beneficial bacteria (Bifidobacterium) and the production of short-chain fatty acids | [39,40] |
| Fermented foods | Lactic acid bacteria and fungi | Inulosucrase catalyzes the biosynthesis of FOS by elongating fructan chains through the addition of fructosyl units, forming β-2,1-glycosidic bonds | [44] |
| Sugarcane Molasses | Aspergillus tubingensis XG21 | High fructofuranosidase (FFase) activity and the ability to synthesize FOS from sugarcane molasses | [45] |
| Carrot juice | A. niger | Carrot juice was utilized as a matrix for producing FOS through the use of a fructosyltransferase enzyme from A. niger | [46] |
| Galactooligosaccharides (GOS) | |||
| Whey cheese | Trichoderma sp. | The highest β-galactosidase activity at 550C | [47] |
| Wheat bran | Penicillium sp. | Highest β-galactosidase activity at the temperature 500C (1.60 IU.mL-1) | [48] |
| Yoghurt | L.bulgaricus CRL450 | Highest GOS production (41.3 %) at fermentation 450C | [49] |
| Fermented milk permeates with apple) | Pediococcus acidilactici | Highest GOS production production (26.80 mg/100mL) at fermentation 30 ◦C for 48 h | [50] |
| Skim milk | Lactobacillus | Direct transformation of lactose into GOS in skim milk Lactobacillus, and utilisation of GOS by Bifidobacterium spp. that lead to the enhanced fermentation and nutritional characteristics | [51] |
| Fermented foods (of Argentina) | Lactobacillus delbrueckii subsp. bulgaricus CRL450 | Out of 20 strains tested, 15 were able to grow on lactose and exhibited β-galactosidase activity, while 11 of them synthesized GOS | [49] |
| Yogurt | Kluyveromyces lactis (YNL-2, GODO) and Lactobacillus acidophilus La-5 | Different types of yogurts with reduced lactose content and enriched with GOS were successfully developed | [52] |
| Tejuino | Pantoea anthophila | The β-galactosidases demonstrated significant transgalactosylation activity, predominantly forming β(1 → 3) and β(1 → 6) linkages | [53] |
| Sweet whey | L. acidophilus, Streptococcus thermophilus, Bifidobacterium lactis, L. delbrueckii subsp. bulgaricus and S. thermophilus | Transgalactosylation was higher in sweet whey compared to control and L. bulgaricus and S. thermophilus showed highest GOS yield | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



