
Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
While DNA serves as the fundamental genetic blueprint for an organism, it is not a static entity. Gene expression, the process by which genetic information is utilized to create functional products like proteins, can be modulated by a diverse range of environmental factors. Epigenetic mechanisms, encompassing DNA methylation, histone modification, and microRNAs, play a pivotal role in mediating the intricate interplay between the environment and gene expression. Intriguingly, alterations in the epigenome have the potential to be inherited across generations. Ethanol abuse poses significant health issues worldwide. Ethanol has the capability to induce changes in the epigenome, which can be inherited by offspring, thus impacting them even in the absence of direct ethanol exposure. This review article reviewed the alterations caused by ethanol on the epigenome. We thoroughly gathered all relevant papers investigating the behavioral and molecular consequences of parental ethanol exposure on their offspring. In conclusion, the comprehensive analysis of the literature supports the notion that ethanol exposure induces lasting epigenetic alterations, which can influence the behavior and health of future generations. This knowledge emphasizes the significance of addressing the potential transgenerational effects of ethanol and highlights the importance of preventive measures to minimize the adverse impact on offspring.
Keywords:
Alcohol
; Ethanol
; Epigenetics change
; phenotype
; offspring
; Epigenome
1. Introduction
Globally, alcohol is one of the most commonly consumed products for entertainment purposes [1]. The number of people aged 15 and over who consume alcohol is 23.48 billion (43% of the population), with men drinking two times as often as women (53.6%/32.3%). The 2018 Global Alcohol and Health Report by the World Health Organization (WHO) highlighted a significant increase in worldwide per capita alcohol consumption (APC) among individuals aged 15 and above, rising from 5.7 to 6.4 liters between 2000 and 2016. This report emphasized the grave public health concern posed by harmful alcohol consumption, which is associated with over 200 diseases and injuries, resulting in approximately 3 million deaths annually on a global scale [2]. According to worldwide data from 2016, alcohol is the seventh leading risk factor in terms of disability-adjusted life years, with an increase of more than 25% since 1990 [3]. Substantial morbidity and mortality are associated with alcohol dependence and alcohol abuse or harmful use [4]. Heavy alcohol consumption is significantly correlated with heart disease [5], stroke, cancer [6], and liver cirrhosis [7] due to its impact on the cardiovascular, gastrointestinal, and immune systems. An excessive amount of alcohol also results in neuronal cell death and neurodegeneration [8], which are the main causes of Parkinson's disease [9], Alzheimer's disease [10], and amyotrophic lateral sclerosis [11]. A strong link is also found between alcohol use disorders and depressive episodes, anxiety, insomnia, suicide, as well as tobacco and drug abuse [4,12,13].
Alcohol dependence has traditionally been regarded as a brain disorder as it alters neuronal function [14,15]. Evidence shows that alcohol affects several neurotransmitter systems in the brain, including those involved in reward and stress circuits [14]. The neurotransmitter systems are composed of dopamine, serotonin, opioid peptides, glutamate, and aminobutyric acid (GABA), which play a crucial role in both positive and negative reinforcement processes that motivate drug seeking and maintain alcohol consumption after alcoholism occurs [15]. Based on the affected neurotransmitters, some pharmacological approaches have been approved by FDA to be implicated in managing alcoholism, such as Naltrexone, Acamprosate, and Disulfiram. Nonetheless, these medications demonstrate a small effect size regarding clinical outcomes and relapse rates, indicating that other biologic processes and peripheral mechanisms might have been involved [16].
Alcohol use disorder (AUD) is identified by a wide range of symptoms that can be further evaluated genetically [17]. These genetic discoveries led to downstream analyses that revealed robust links between alcohol use and physical and mental health. Furthermore, several morphological differences were observed between offspring from families with alcohol dependence and those from control families, including differences in neurotransmitter functioning that occur during childhood, affect regional brain development, and possibly persist into adulthood [18]. There is strong evidence that prenatal alcohol exposure is the main cause of fetal alcohol spectrum disorder (FASD), a condition characterized by severe neurodevelopmental delay and a variety of manifestations, such as growth deficiencies, psychiatric vulnerability, as well as both behavioral and cognitive impairments [19,20].
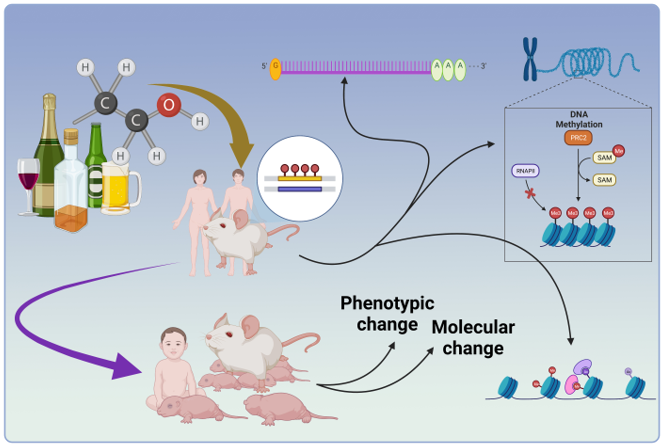
Considering the social, economic, and individual burden exerted by alcohol dependence and its comorbidity with anxiety, it is imperative to understand the underlying cause of the negative affective domains of alcohol addiction [21]. Recently, advances in the study of alcoholism have revealed epigenetic pathways in the brain influenced by several modes of alcohol exposure that lead to changes in neuronal morphology and the development and maintenance of AUDs. In the epigenetic processes, DNA and histones undergo chemical modifications; in particular, DNA and histone methylation, histone acetylation, as well as microRNA expression mechanisms are regulated by exposure to both acute and chronic levels of ethanol [22]. The alteration of the epigenome was claimed to occur in different parts of the brain or the blood in the level of mRNA expression or protein production, resulting in an increased tendency to alcohol consumption [20,23]. Furthermore, these modifications can be impermanent or transferred to the next generations [19]. Rapid advances in molecular techniques and deep insights into epigenomes, transcriptomes, and proteomes have led to the development of novel therapies for drug dependence and alcoholism that target epigenetic mechanisms to manage these conditions [24]. The current review aims to evaluate the intergenerational and transgenerational changes in molecular and behavioral outcomes induced by alcohol exposure. We also summarize the latest findings regarding allostatic alterations within the epigenome as a promising tool for regulating negative affective states in alcohol addiction.
2. Alcohol Abuse and Changes in the epigenome
As mentioned in the introduction, ethanol consumption can significantly impact the epigenome. Exposure to alcohol at different stages of life can disrupt chromatin function and DNA methylation, which affects neuronal plasticity, potentially leading to prolonged effects [25]. Therefore, in this part, we aim to review ethanol consumption's impact on various aspects of the epigenome (Summarized in Table 1). We have taken into account both human and animal studies, as we believe it is crucial to consider findings from different models to gain a comprehensive understanding of the impact of ethanol consumption on the epigenome. By categorizing the manuscripts based on the study subjects, we have ensured a thorough evaluation of the available literature. It is worth noting that since acute and chronic ethanol exposure induces different changes in the epigenome (for example, [26,27]), we only focus on chronic alcohol exposures.
2.1. Human Studies
Many human studies have focused on DNA methylation in the context of epigenetic changes following ethanol exposure. However, it is crucial to consider other aspects as well. Research investigating the impact of ethanol consumption on epigenetic modifications has predominantly examined DNA methylation patterns. Various sample types, such as blood and postmortem brain tissue, have been utilized in these studies.
Several studies have evaluated the effect of ethanol abuse on gene methylation levels using blood samples. In humans who were prenatally exposed to ethanol, 16 affected CpG sites were identified compared to the control group [19]. Philibert et al. (2018, 2014) conducted genome-wide DNA methylation analyses in mononuclear blood cells, finding significant changes in DNA methylation levels between patients with AUD and healthy control subjects [28,29]. Also, distinct DNA methylation patterns based on ethnicity were observed in the individuals with alcohol addiction. This study showed specific genes exhibiting hypermethylation in African American alcohol addicts (POMC and GABRB3), while in European Americans, different genes showed hypermethylation (HTR3A, NCAM1, DRD4, MBD3, HTR2B, and GRIN1) [30]. Indeed, it has been found that alcohol misuse leads to genotype-dependent, allelic-specific methylation at a specific DNA methyl region (DMR) called DMR-DLGAP2, which is associated with reward processing and subsequent alcohol abuse [31]. A significant increase in dopamine D2 receptor gene methylation during alcohol withdrawal/early abstinence was also observed [32]. Studies examining individuals with different drinking patterns demonstrated an increase in DNA methylation of PER2 and POMC genes among binge and heavy drinkers, accompanied by decreased gene expression. These higher levels of DNA methylation were predictors of increased subjective alcohol craving and were associated with the amount of alcohol consumed during the Alcohol Taste Test [23,33]. Krasteva et al. (2020) evaluated DNMT1, DNMT3a, and methyl-CpG-binding domain proteins in the blood of individuals with AUD, finding downregulation of MeCP2 and MBD1 in the alcohol-dependent group [34].
Furthermore, studies have explored DNA methylation patterns in relation to alcohol consumption in various populations. Maeda et al. (2022) conducted a cross-sectional study involving non-drinkers, moderate drinkers, and heavy drinkers, finding significantly lower TXNIP DNA methylation levels in the leukocytes of heavy drinkers compared to non-drinkers and moderate drinkers [35]. Dugué et al. (2021) performed a large sample size study measuring DNA methylation in blood samples from participants, identifying numerous CpGs associated with alcohol intake, including some that exhibited differential methylation in former drinkers compared to current drinkers [36]. Besides, methylation levels of ALDH2 and MTHFR in peripheral blood leukocytes among individuals with alcohol use disorder are higher than in the control group [37]. In investigations carried out by Zhao et al. (2013) on peripheral mononuclear cells, they identified 865 hypomethylated and 716 hypermethylated CG sites in individuals who consumed alcohol. Additionally, gene expression changes were observed in LDH1L2, GAD1, DBH, and GABRP genes under alcohol exposure [38]. Furthermore, global DNA hypermethylation in the mononuclear cells of alcoholics experiencing early alcohol withdrawal was observed [39]. A study by Clark et al. (2022) examined cell-type-specific DNA methylation in individuals with alcohol use disorder and found a significant enrichment of methylation and hydroxymethylation patterns associated with alcohol use disorder in monocytes [40]. In addition, they also identified and replicated a significant finding in DLGAP1, suggesting its potential involvement in alcohol use disorder-related brain impairment.
A study on Finnish twins (1,004 individuals) assessed the relationship between alcohol consumption and epigenome-wide DNA methylation. Significant associations between alcohol consumption and DNA methylation were identified at 24 specific CpG sites. When comparing differences in alcohol consumption and DNA methylation between monozygotic twin siblings (185 pairs), four CpG sites were replicated from the initial analysis, and 23 additional sites were discovered. However, no significant differences in DNA methylation patterns were found when examining qualitative differences in drinking patterns within twin pairs (44 pairs) [41]. Another study on 57 Chinese monozygotic twins and 158 Swedish adoption/twin participants revealed that hypermethylation of cg07326074, located in the tumor-promoting gene C16orf59, was associated with alcohol consumption. These findings highlight the specific DNA methylation pattern at this CpG site in relation to alcohol consumption in monozygotic twins [42].
A recent study conducted on six separate cohorts, encompassing both blood and brain samples, utilized epigenome-wide data. The findings of this study provided insights into the association between alcohol use behaviors and a network of differentially methylated regions, particularly in genes related to glucocorticoid signaling and inflammation [43]. Some studies have explored DNA methylation patterns in the brains of individuals with AUD, revealing important information about the epigenetic changes associated with chronic alcohol consumption. In a survey by Manzardo et al. (2012), DNA was isolated from the frontal cortex of ten alcoholics and ten matched controls. Microarrays containing genomic promoter regions showed no significant differences between alcohol-dependent individuals and controls regarding global methylation parameters or most gene targets. Notably, DNA methylation was also detected in various variants of histone proteins, particularly those associated with gene loci in the HIST2 domain of chromosome 1 [44]. Additionally, profiling the transcriptome of the amygdala and frontal cortex in 17 alcoholics and 15 matched controls revealed changes in chromatin modification in the alcoholic brain. This was evidenced by the association between DNA hypomethylation and increased histone H3K4 trimethylation in alcoholics [45]. Besides DNA methylation, there are studies that evaluate the role of early-onset alcohol abuse on the expression of long non-coding RNA (lncRNA) in the amygdala. It has been found that the levels of BDNF-AS lncRNA increased in the amygdala of early-onset ethanol abuse, whereas it does not show such an increase in those with late-onset AUD. The upregulation of BDNF-AS in the early onset AUD group is linked to a significant decrease in BDNF expression and increased recruitment of EZH2, which is responsible for depositing repressive H3K27 trimethylation (H3K27me3) at regulatory regions within the BDNF gene [46].
A consistent trend of increased demethylation was observed in the sperm DNA of individuals who consumed alcohol, particularly at two specific imprinted loci. The most significant difference was found at the IG-DMR locus, where individuals who heavily consumed alcohol showed significant demethylation compared to those who did not drink alcohol. These findings raise the hypothesis that if these epigenetic changes are transmitted through fertilization, they could disrupt the appropriate gene expression levels required for normal prenatal development [47]. While the level of alcohol use did not show a significant association with the High DNA Stainability or DNA Fragmentation Index of sperm in a study by Trautman et al. [48], another survey by Blaseg et al. revealed that in non-drinkers, the levels of High DNA Stainability increased [49].
All this evidence indicates that alcohol exposure can lead to alterations in the epigenome. Considering that epigenetic changes can be inherited, it underscores the importance of implementing reprogramming strategies in public health policies.
2.2. Animal Studies
Several studies on animal models have demonstrated that alcohol exposure can lead to alterations in DNA methylation patterns, histone modification, and miRNAs. It has been revealed a notable reduction in DNA methylation levels in the leptin promoter region of animals consuming alcohol compared to those who had not been exposed to alcohol. However, after a period of 6 days without alcohol consumption, this disparity in methylation levels ceased to exist [50]. Adolescent intermittent ethanol (AIE) exposure has been found to cause significant alterations in DNA methylation mechanisms within the amygdaloid circuitry. Following repeated exposure to ethanol during adolescence, there is a decrease in the activity of DNMT enzymes, accompanied by an increase in the expression of the growth factors and DNA-damage-inducible (Gadd45) family of proteins, which facilitate active DNA demethylation. However, when withdrawal from repeated ethanol exposure occurs after 24 hours, DNMT activity in the amygdala increases. This heightened DNMT activity, along with elevated Dnmt3b mRNA levels and reduced Gadd45 mRNA expression, is maintained throughout development within the amygdala [51].
Additionally, exposure to AIE in rats resulted in hyperacetylation of H3K9/14 in the promoter of the Melanocortin 4 receptor (mc4r) in the amygdala, as well as in the promoters of both Mc4r and Pomc in the hypothalamus. In contrast, the promoter of the Npy gene exhibited lower levels of acetylation following adolescent ethanol exposure [52]. In Rhesus macaques, ethanol self-alcohol administration impacted 17 significant DMRs. Using three different DMR-associated genes for analysis, it was found that excessive alcohol consumption increased JAKMIP1-DMR and GPR39-DMR methylation and decreased LRP5-DMR methylation [53]. Another animal study demonstrated that animals with a history of alcohol dependence exhibited elevated levels of global DNA methylation in the medial prefrontal cortex (mPFC). This increase in DNA methylation was found to be associated with alcohol-seeking behavior, specifically through the silencing of Syt2 [54]. In addition, exposure to ethanol during post-natal days 2-6 led to reduced global DNA methylation along with globally increased acetylation of histone 3 lysine 9 (H3K9ac) in hypothalamic microglia, which resulted in elevated levels of pro-inflammatory cytokines [55].
It has been reported that histone modification undergoes changes in both short-term and long-term ethanol exposure in the mPFC. Following short-term exposure, the levels of AcH3K9, AcH3K5, and AcH4K12 increased, while the levels of AcH3K9 decreased. In contrast, during long-term exposure, only the levels of AcH4K5 increased in the mPFC [56]. Further, mice exposed to chronic ethanol showed reduced levels of AcH3K14 expression in the basal forebrain (BF) and lateral hypothalamus [57]. Acute ethanol exposure can potentially influence histone modification apart from chronic exposure. Studies have shown that acute ethanol exposure leads to decreased H3K9me2 and G9a protein levels in the amygdala. Additionally, it causes a reduction in the occupancy levels of these proteins near the CREB and Npy genes [58].
During the adolescent stage, exposure to ethanol was found to cause a reduction in the levels of mRNA associated with H3 lysine 9 dimethylation (H3K9me2) in the amygdala. Moreover, it led to an elevation in the presence of H3K9me2, specifically at the promoter region of BDNF exon IV [27]. The miRNA-137 was implicated in the process of chromatin remodeling at BDNF exon IV [59]. Evidence suggests that adolescent alcohol exposure leads to the regulation of enhancer RNA (eRNA) through epigenetic reprogramming in the amygdala. This regulation is particularly observed at the Arc synaptic activity response element site [60]. It has been observed that ethanol can also impact miRNA expression. Specifically, acute administration of ethanol was found to decrease the levels of several miRNAs (miR-29b, miR-154, miR-7a-2, miR-185, miR-7c-1, miR-485, miR-6215, miR-29a, miR-374, miR-433, miR-1843a, miR-135a) in the hippocampus. These alterations in miRNA expression were accompanied by changes in the levels of cytokines and growth factors [61]. Furthermore, Rossetto et al. (2019) demonstrated that prolonged exposure to ethanol increased the levels of specific miRNAs, including miR-155-5p, miR-146a-5p, miR-126-3p, and miR-132-3p, in the cerebellum. These miRNAs have been found to play a role in inflammatory pathways [62].
There are studies that explore the impact of ethanol exposure on epigenetic changes in sperm, considering that these changes can be inherited by subsequent generations. Research has demonstrated a decrease in the levels of DNMT1 in the sperm of rodents exposed to chronic ethanol through intragastric administration [63]. Furthermore, multiple studies have indicated that DNA methylation levels at imprinted gene loci are decreased in the sperm of mice subjected to chronic alcohol treatment [64,65,66]. Similar findings have also been observed in men diagnosed with AUD [47]. Prenatal exposure to ethanol has been shown to increase methylation levels at the promoters of the POMC gene in sperm. This epigenetic alteration can subsequently influence the functioning of the hypothalamic-pituitary-adrenal axis in the offspring [67]. Furthermore, Rahimipour et al. (2013) demonstrated that chronic ethanol exposure in mice had detrimental effects on the integrity of sperm chromatin and DNA [68]. Chronic intermittent alcohol exposure has been shown to induce changes in multiple miRNA and tDR (tRNA-derived RNA) species in sperm, as demonstrated by Rompala et al. [69]. Notably, among the small RNAs that exhibited increased levels in the sperm due to alcohol exposure, miR10a stood out. This particular microRNA is known to be significantly enriched in sperm compared to oocytes [70] and is suggested to function as a posttranscriptional regulator of the alcohol-related gene BDNF [71]. Furthermore, ethanol exposure has been shown to elevate the levels of H3K4me3 (trimethylation of histone H3 at lysine 4) in sperm. This increase in H3K4me3 leads to enhanced CTCF (CCCTC-binding factor) binding in male placentae [72].
In summary, ethanol exposure during different developmental stages, such as adolescence or adulthood, can induce changes in DNA methylation, histone modification, and miRNAs in the brain and germ cells. These epigenetic changes can influence the phenotype of individuals who abuse alcohol and have the potential to be transmitted to future generations.
3. Transgenerational and Intergenerational Effects of Alcohol
It has been indicated that alcohol abuse by parents can affect the health and development of future generations. Studies have shown that alcohol use during pregnancy causes fetal alcohol spectrum disorders (FASDs), causing a range of physical, cognitive, and behavioral impairments in offspring [73]. However, emerging research suggests that the effects of alcohol may extend beyond immediate offspring and be transmitted to subsequent generations. As mentioned in the previous section, ethanol consumption induced changes in the epigenome. These epigenetic modifications can influence gene expression patterns and contribute to transmitting alcohol-related traits and vulnerabilities across generations. Studies have revealed that parental alcohol consumption alters offspring's gene expression patterns and phenotypes. Here, we summarized animal studies conducted on the transgenerational effects of alcohol (Table 2 and Table 3). It should be noted that understanding the transgenerational effects of alcohol is crucial for implementing effective prevention strategies and interventions to break the cycle of alcohol-related harm and promote healthier outcomes for future generations.
According to a study conducted by Hill et al., which focused on a long-term follow-up of three generations of individuals with a family history of alcohol dependence, ancestral exposure to alcohol can have lasting effects on epigenetic processes. Specifically, it was found that DNA methylation, which controls the expression of genes involved in cancer development, such as HRAS and TP53, can be impacted. The study revealed that offspring with increased loading of parental alcohol dependence displayed hypomethylation of the HRAS gene, indicating potentially increased expression of this oncogene. Moreover, the hypomethylation of HRAS CpG sites was associated with a higher prevalence of familial alcohol dependence across multiple generations. Based on the findings, it is indicated that a genetic predisposition to alcohol dependence within families leads to alterations in the methylation patterns of the DRD2 gene. These changes have implications for reducing the volumes of certain brain regions in the left hemisphere, specifically the fusiform, insula, and temporal areas, which are known to be involved in social cognition. While there is a correlation between prenatal substance use and familial risk for alcohol dependence, it is improbable that these factors alone can account for the observed decrease in volume within these brain regions [18].
3.1. Maternal Care and Pups Development in Preconception Model of Ethanol Exposure
Evidence showed that preconception ethanol exposure affects maternal caring, mortality rate, and pups' appearance characteristics. Maternal ethanol exposure three weeks before mating did not affect the behavior of mothers in terms of licking, grooming, and arched-back nursing. Additionally, the duration of estrus returned to its normal length two weeks after the withdrawal of alcohol [74]. Liang et al. (2014) found that the mortality rate increased among the offspring of ethanol-exposed sires [75]. Although the results obtained from our lab indicated that maternal and biparental ethanol exposure 10 days before mating decreased maternal care behavior and increased the mortality rate in the offspring during the 21 days after birth (submitted data),
Sex ratio, the number of pups, and pregnancy duration also were investigated in the ethanol-exposed animals. In a study by Jabbar et al. (2016), maternal exposure to alcohol resulted in longer pregnancy duration and decreased body weight. Still, it did not affect how mothers cared for their babies [74]. Furthermore, there was no influence of paternal alcohol exposure on the number of pups, sex ratio, and mortality rate of litters [76,77,78]. It was indicated that exposure of male mice to ethanol 12 hours before mate increased mortality rates and the number of pups [79]. This controversy might be attributed to acute vs. chronic ethanol exposure.
The body weight of pups was found to be lower than usual when either parent was exposed to ethanol during adolescence and adulthood (PND 37-44 and PND 67-74) 24 hours before mating [80]. Male mice exposed to acute ethanol 12 hours before mating exhibited a reduced total number of pups, decreased body weights after birth, and an increased mortality rate during the first three weeks after birth [79]. There is evidence showing that paternal alcohol exposure before gestation did not affect the body weight of male or female offspring [75,76,81,82,83]. Ceccanti and colleagues [78] found that exposure of male mice to ethanol 2 months before gestation reduced pup's body weight, which was in agreement with a study conducted by Ledig et al. [84]. Besides, adolescent exposure to ethanol in male rats (2 weeks before mating) did not alter developmental landmarks in offspring [85].
Overall, these findings demonstrate that preconception ethanol exposure can have significant effects on maternal behavior, mortality rates, pup characteristics, and body weight. However, there are variations in the outcomes depending on factors such as timing, duration, parental sex (maternal or paternal exposure), and whether the exposure is acute or chronic.
3.2. phenotype Changes Followed by Preconception Parental Ethanol Exposure
Phenotypic changes resulting from parental drug exposure have garnered recent interest in the field of research. This part aims to provide an overview of the behavioral changes observed following parental ethanol exposure before gestation. These changes shed light on the potential effects of parental alcohol consumption on offspring development and behavior.
3.2.1. Anxiety-like Behavior
Several studies have revealed that parental alcohol exposure prior to gestation can lead to alterations in anxiety-related behavior in offspring. Importantly, these changes have been found to be dependent on the sex of the offspring. The findings from these studies indicate that when either the mother or the father is exposed to alcohol before conception, it can impact the anxiety levels of their offspring. The specific behavioral changes observed in the progeny differ between males and females. Exposing male mice to ethanol for 5 weeks prior to breeding with a drug-naïve female resulted in an increased anxiolytic effect of ethanol in male –but not female- offspring. However, no significant change in the anxiety-like behavior at baseline was observed in the EPM test [86]. Furthermore, Jabbar et al. [74] demonstrated that maternal ethanol exposure three weeks before gestation increased anxiety-like behavior, specifically in male offspring, as observed in both the EPM and OFT. Contrary, there is other finding that have demonstrated an increase in basal levels of anxiety specifically in the female progeny of male mice exposed to ethanol 30 days before mating [75]. Evidence revealed that male CD1 mice exposed to ethanol prenatally showed that pre-pubescent offspring (F1 and F2) displayed no change in anxiety-like behavior compared to the control group [87]. In another study, the role of exposing male rats to ethanol 8 weeks before conception on the anxiolytic effect of ethanol in male and female offspring was evaluated. It has been found that after ethanol administration, anxiety decreased in the male progenies of ethanol-sired rats, but no significant change was observed in the female progenies. However, paternal alcohol exposure does not impact the baseline anxiety-like behavior, as measured by the time spent in the center of the open field test, in rats of either sex [88].
Other investigations for assessing the anxiety-like behavior in the offspring of ethanol-exposed parents using OFT also showed different results. For example, male offspring sired by ethanol-exposed fathers exhibited a longer duration of stay in the center of the test arena compared to their non-ethanol-exposed siblings. However, the overall distance traveled showed no significant variation between the two groups. Additionally, the resting time in the center increased, indicating an increase in the amount of time spent in a less anxious state, while the resting time in the perimeter decreased, suggesting a decline in anxiety-like behavior in the aforementioned offspring. In this study, however, no differences in EPM parameters were observed [89]. Another investigation evaluating anxiety behavior in the progeny of male rats exposed to ethanol before gestation revealed no significant changes in the results of OFT in both male and female offspring of ethanol-exposed males, specifically exposed 8 weeks before mating [90].
In summary, the effects of ethanol exposure on anxiety-like behavior in offspring appear to be influenced by various factors, including the timing of exposure, parental sex, and offspring sex. These findings highlight the complex and sex-dependent nature of ethanol's impact on anxiety-like behavior in offspring.
3.2.2. Learning and Memory
Memory impairment is a common consequence of chronic alcohol abuse, affecting both short-term and long-term memory processes. Research studies have shown that alcohol-related memory impairment is linked to structural and functional changes in the brain, particularly in the hippocampus and prefrontal cortex [91]. Several studies have investigated the effects of paternal ethanol exposure on memory and cognitive abilities in offspring. Abel et al. [82] reported that paternal ethanol exposure, up to 10 days before pregnancy, had a negative impact on memory in the offspring, as assessed through the passive avoidance test. Similarly, ethanol exposure to the father for 39 days and two weeks before conception impaired memory in the radial arm maze in the offspring, indicating compromised cognitive function [85]. Furthermore, Liang et al. [75] conducted the Morris water maze (MWM) test and observed that both male and female offspring of ethanol-exposed sires, exposed 1 month before copulating, exhibited deficits in both learning and memory abilities. These findings collectively suggest that paternal ethanol exposure can have detrimental effects on offspring's cognitive function and memory performance. It is worth noting that these studies highlight the importance of the timing of ethanol exposure, as different time points before mating or conception resulted in distinct cognitive impairments in the offspring.
3.2.3. Depression
The findings from a comprehensive meta-analysis of epidemiological data indicate a strong association between alcohol use disorder (AUD) and an elevated risk of depression [92]. Additionally, a study conducted on adult male mice revealed that exposing them to ethanol for one month, including one month before gestation, led to an increase in depressive-like behavior, specifically in female offspring, while no significant effect was observed in male offspring [75]. Besides, in the first and second generations of male CD1 mice prenatally exposed to ethanol, the depressive-like behavior increased [87]. Our laboratory conducted experiments that further support the association between maternal ethanol exposure and increased depressive-like behavior in offspring. In our study, we exposed female rats to ethanol for a duration of 30 days, with a subsequent period of 10 days of abstinence before mating. Our research demonstrated a significant increase in depressive-like behavior in the progeny of these maternal ethanol-exposed female rats. This finding suggests that maternal ethanol exposure during a critical period can have lasting effects on the emotional well-being of the offspring, specifically manifesting as depressive-like behavior (submitted data).
3.2.4. Reward
Parental ethanol abuse has been shown to impact the reward system in their offspring significantly. Studies have indicated that exposure to ethanol by either the mother or father can lead to alterations in the neural circuitry responsible for processing rewards in the offspring. This can result in changes in their behavioral responses to rewarding stimuli. For example, the offspring of parents with an ethanol abuse history may exhibit an increased preference for ethanol-related cues or substances, showing a heightened sensitivity to the rewarding effects of alcohol. These changes in the reward system highlight the intergenerational consequences of parental ethanol abuse and emphasize the importance of understanding the long-term effects on offspring behavior and susceptibility to substance abuse [93]. Here, we have compiled a summary of studies investigating the impact of parental ethanol abuse prior to mating on alterations in the reward system of the offspring.
According to Ceccanti et al. (2016), it has been suggested that if male mice were exposed to ethanol two months before mating, it could influence the ethanol-induced conditioned place preference (CPP) in their offspring. The study found that the offspring of mice exposed to alcohol displayed higher conditioning scores in the low dose (0.5 g/kg) group compared to the control group. However, in the high dose (1.5 g/kg) group, they exhibited aversion [78]. Male offspring sired by mice exposed to ethanol vapor for five weeks, with conception occurring immediately after the last exposure, demonstrated a greater preference for ethanol consumption in the two-bottle choice test. However, no such preference was observed in female offspring sired by ethanol-exposed mice [65]. In the context of chronic stress, the offspring of ethanol-exposed mice did not exhibit any differences in ethanol preference compared to the control group [94]. Furthermore, it has been reported that exposure to ethanol for 12 weeks, with a 48-hour window before conception, resulted in decreased ethanol consumption in male offspring but not in female offspring, as assessed through the drinking in the dark (DID) method [95].
In a study conducted by Nieto et al. (2022), male Wistar rats were subjected to chronic intermittent ethanol exposure in vapor chambers for 6 weeks, with a daily duration of 16 hours and 5 days per week, and mate after eight weeks. The alcohol-sired offspring consumed less alcohol (at 5% and 10% concentrations) in the alcohol self-administration task without any differences in their response to food [96]. These findings suggest that paternal alcohol exposure before conception provides protection against the initial reinforcing effects of alcohol.
3.2.5. Other Phenotypic Changes
Parental alcohol abuse has been found to affect various neurobehavioral reflexes, such as the righting reflex, geotactic negative reflex, balance, and gait, as documented by Jamerson et al. [77]. Further, parental ethanol exposure causes behavioral abnormalities in mice models, including abnormal sensorimotor processing that extends to the F2 generation [87].Rompala et al. [94] discovered that male offspring from B6 mice exposed to ethanol for five weeks displayed resistance to stress-induced polydipsia. In another study, impulsivity was assessed using the electro-foot shock-aversive water drinking test, revealing that the offspring of male mice exposed to ethanol (for seven weeks, with exposure occurring one week before gestation) exhibited increased impulsivity. Moreover, these male offspring also displayed ADHD-like behavior [97]. It has been observed that alcohol-sired rats exhibited more aggressive-like behavior and reduced defensive/fearful behavior [98]. Furthermore, paternal ethanol exposure two months before gestation resulted in increased locomotion, decreased latency in the novelty-seeking test, and increased time spent in the light compartment during the light-dark test [84]. A morphometric study has revealed that paternal alcohol exposure, both during and before conception, leads to an increase in the thickness of cortical sections I–IV and V–VI. This finding suggests that paternal alcohol consumption may have a significant impact on the structural development of specific layers within the cortex. The increased cortical thickness in these sections could potentially influence neuronal connectivity, synaptic plasticity, and overall brain function [77].
There is a study conducted on zebrafish by Suresh et al. [99]. They utilized zebrafish to eliminate the influence of maternal care on the transgenerational inheritance of ethanol. The zebrafish were exposed to ethanol seven hours before mating, and subsequent behavioral alterations were observed in the offspring. The study revealed that the ethanol-exposed offspring displayed increased immobility and exhibited more frequent turns than the control group. These findings suggest that these behavioral changes may indicate anxiety and exploratory behavior alterations.
3.3. Parental Ethanol Exposure and Endocrine Changes in the Offspring
Apart from the behavioral changes resulting from parental ethanol exposure, it has been noted that ethanol exposure can also modify the levels of endocrine hormones in the offspring. Plasma corticosterone levels in male offspring were found to be elevated due to maternal alcohol exposure before gestation. Additionally, both males and females showed increased Adrenocorticotropic hormone (ACTH) and corticosterone levels in response to the lipopolysaccharide (LPS) challenge. In addition, male and female offspring of female rats exposed to ethanol preconceptionally exhibited a reduction in hypothalamic β-endorphin, along with an increase in corticotropin-releasing hormone (CRH) [74]. These offspring also displayed heightened levels of inflammatory cytokines and cellular apoptosis in the pancreas, disrupted insulin production and functionality in the liver, and a decreased count of proopiomelanocortin neurons in the hypothalamus, which caused hyperglycemia and hypoinsulinemia. However, when proopiomelanocortin neurons were replenished in these animals, it led to the restoration of glucose levels and the re-establishment of homeostasis [100].
When both parents were exposed to ethanol one day before mating, it caused a reduction in corticosterone levels, specifically in male offspring, whereas female offspring did not demonstrate significant changes. Furthermore, the male progeny exhibited a decrease in circulating levels of luteinizing hormone (LH), while both male and female offspring displayed decreased testosterone levels. Notably, no notable alterations were observed in the morphology of the testes [80]. Consistent with these findings, it has been shown that male offspring fathered by males exposed to ethanol, particularly when exposed 10 days prior to mating, displayed reduced testosterone levels [82]. Paternal ethanol exposure for 5 weeks just before conception attenuates the response of the HPA-axis (reduced corticosterone levels) to acute restraint stress, specifically in male offspring [94].
In conclusion, parental ethanol exposure can modify endocrine hormone levels, such as increased corticosterone levels, hyperglycemia, hypoinsulinemia, and decreased circulating luteinizing hormone levels in male offspring. These findings highlight the complex impact of ethanol exposure on endocrine and stress response systems in offspring, with potential gender-specific effects.
3.4. Parental Ethanol Exposure before Gestation and Changes in Gene Expression
Preconception ethanol exposure, whether it occurs in the mother or the father, has the potential to impact gene expression levels in their offspring. Research suggests that the process of epigenome regulation can influence these alterations in gene expression. The inherited epigenetic modifications can have long-lasting effects on gene expression patterns in the offspring. They can either activate or suppress specific genes, leading to changes in cellular processes and potentially influencing behavior. Here, we summarize the molecular changes that occurred in the brain of offspring.
3.4.1. Epigeneome Changes
It has been declared that sires exposed to alcohol (for 6 weeks and 8 weeks free before gestation) exhibit higher levels of global DNA methylation in sperm compared to control sires. However, there are no statistically significant differences in global DNA methylation levels between the sire groups in the NAc or mPFC. Furthermore, global DNA methylation levels in sperm do not differ between male offspring sired by alcohol-exposed and control parents. Similarly, there were no significant effects observed in global DNA methylation between alcohol-exposed offspring and the control group in the prefrontal cortex (PFC) and nucleus accumbens [90]. In a study conducted by Nieto et al. (2022), male Wistar rats were subjected to chronic intermittent ethanol exposure. After eight weeks, these rats were bred with females that had not been exposed to alcohol. The alcohol-exposed sires showed lower levels of Bdnf DNA methylation in the Nucleus Accumbens (NAc) and higher levels in the mPFC. Although the same methylation pattern was not observed in the offspring, both male and female offspring sired by alcohol-exposed males exhibited abnormal Bdnf DNA methylation patterns compared to offspring sired by control males. However, the dysregulated Bdnf methylation pattern in reward-related circuitry observed in the alcohol-exposed sires was not replicated in the offspring [96].
An analysis of the thalamus genome in offspring rats revealed that exposure to alcohol by either parent led to increasing hypermethylation of differentially methylated cytosines (DMCs). The total number of hypermethylated DMCs was higher in the offspring when both parents had alcohol exposure than when only one parent had alcohol exposure. Interestingly, genes that were differentially methylated when only one parent was exposed to ethanol remained unchanged in the offspring of both parents exposed to alcohol. Additionally, the methylation patterns of DMCs varied across the chromosome depending on the alcohol exposure status of the parents. Specifically, both parents' alcohol exposure caused hypermethylation of chromosome X, while only one parent's alcohol exposure resulted in increased methylation on chromosome X [101]. The study conducted by Liang et al. reported an increase in the methylation of H19 and Peg3 in sperms of ethanol-exposed mice. Evaluating of cerebral cortex in the offspring revealed that there were no changes in the methylation of H19, PEG3, and Ndn genes, while the methylation of the snrpn gene decreased in the alcohol-exposed offspring [75].
It has been indicated that prenatal exposure to ethanol changes Pomc DNA methylation in the hypothalamus and sperm across generations (two generations) via the male germline, which suppression of pomc expression involves the hyper stress response observed in the male germlines of F1 and F2 rats [67]. Preconception ethanol exposure in sires leads to a reduction in DNA methylation levels of the BDNF gene in the ventral tegmental area of male offspring. However, no effect was observed in the mPFC or in female offspring. Indeed, the levels of BDNF expression increased in these progenies, followed by a reduction in DNA methylation [65]. Paternal ethanol exposure during late adolescence, adulthood, and one week before conception has been found to decrease DNMT1 and MeCP2 levels, as well as DAT expression in the mPFC and striatum of male offspring. Notably, studies have demonstrated that the reduction in DAT expression is correlated with an increase in methylation levels within the promoter region of this gene [83]. It was found that the methylation of the crfr1 (corticotropin-releasing factor receptor 1) gene decreased in the hippocampus and amygdala of male offspring born to ethanol-exposed female rats. Conversely, the methylation levels of pomc increased in the hypothalamus. These findings suggest that maternal ethanol exposure can impact the expression and epigenetic regulation of stress-related genes in specific brain regions of offspring, potentially contributing to altered stress responses and neurobehavioral outcomes [102].
In conclusion, while some studies have reported higher global DNA methylation levels in sperm of alcohol-exposed sires, these differences did not extend to specific brain regions such as the nucleus accumbens or mPFC in offspring. Additionally, alterations in DNA methylation profiles have been observed in the offspring of parents who engage in binge-pattern alcohol consumption during adolescence, suggesting intergenerational effects.
3.4.2. Changes in Gene Expression
In addition to the changes in the epigenome, numerous studies have evaluated the gene expression levels in the brain of offspring born to parents who were exposed to ethanol preconceptionally. By examining gene expression levels, researchers seek to uncover molecular mechanisms that may underlie the observed phenotypic and behavioral alterations in these offspring. Through these investigations, a deeper understanding of the molecular consequences of preconceptional ethanol exposure and its potential long-term effects on offspring development is being attained.
It has been shown that pre-pubertal and adulthood ethanol exposure before conception changes some genes' expression levels in the offspring's hypothalamus. These genes involved in neurodevelopment and synaptic plasticity (FGF13, BMP1, SERPINI1, RELN, PAK3, IGF2R, VAMP3, FGFR3), metabolic functions (GnRH, APOE), transcriptional regulation, and post-translational processing (EGR2, DICER1, WBP4, SUMO2), as well as chromatin modifications (HDAC3, SIRT2, DNMT1) [103]. In a study conducted by Ceccanti et al. [78], it was demonstrated that paternal ethanol exposure two months before conception did not have an impact on dopamine receptors (D1 and D2) in various brain regions, including the PFC, hippocampus, hypothalamus, and olfactory bulb. However, BDNF levels increased while nerve growth factor (NGF) and P75 levels decreased in the PFC. Moreover, there were no changes in Trk-B expression in the male offspring's brain, but Trk-A expression increased in the hippocampus. Additionally, it was found that only the olfactory bulb showed an increase in dopamine transporter (DAT) levels [78]. Maternal ethanol exposure, specifically for a duration of four weeks, three weeks prior to mating, has been shown to induce changes in the expression levels of stress regulatory genes in the brains of offspring, as reported by Jabbar et al. (2016) [74]. Notably, the mRNA expression levels of crf (corticotropin-releasing factor) increased in the hypothalamus, while pomc mRNA levels decreased. Additionally, the mRNA expression of crfr1 increased in the hippocampus and amygdala [102]. In addition to alterations in gene expression in the brain, male exposure to ethanol before conception is associated with various adverse effects, including reduced fetal growth, decreased placental efficiency, abnormal cholesterol trafficking, alterations in genetic pathways related to hepatic fibrosis, and disruptions in gene expression [104].
In summary, these investigations have revealed alterations in the expression levels of genes involved in neurodevelopment, synaptic plasticity, metabolic functions, transcriptional regulation, and chromatin modifications. Furthermore, specific brain regions have shown changes in the expression of genes related to stress regulation and neurotransmitter systems.
4. Conclusion
Ethanol-induced alterations in DNA methylation patterns have been observed in both the brain and germ cells in humans. In preclinical studies, it has been demonstrated that ethanol exposure can also induce alterations in histone modifications and miRNAs. These changes can result in long-lasting effects on gene expression patterns and contribute to the phenotypic outcomes observed in individuals who abuse alcohol. Importantly, these epigenetic modifications have the potential to be transmitted across generations, meaning that the effects of ethanol exposure can extend beyond the individual experiencing the exposure.
These findings highlight the complexity of the epigenetic changes induced by ethanol exposure and their potential implications for the affected individuals and their offspring. Understanding these epigenetic alterations is crucial for developing effective prevention and intervention strategies for alcohol-related disorders. Additionally, recognizing the transgenerational impact of ethanol-induced epigenetic changes underscores the importance of public health policies aimed at reducing alcohol consumption and promoting healthy lifestyles across generations.
Abbreviation
ACTH: Adrenocorticotropic hormone; ADHD: Attention-Deficit / Hyperactivity Disorder; AUD: Alcohol use disorder; AIE: Adolescent intermittent ethanol, ALDH2: Mitochondrial Aldehyde Dehydrogenase, BDNF: Lower brain-derived neurotrophic factor; BF: Basal Forebrain, CREB: cAMP-responsive element binding protein, CTCF: CCCTC-binding factor, CPP: Conditioned place preference; CRH: Corticotropin-releasing hormone; Crfr1: Corticotropin-releasing factor receptor-1; DAT: Dopamine transporter; DID: Drinking in the dark; DBH: Dopamine Beta (β)-hydroxylase, DMR: DNA Methyl Region, DNMT: DNA Methyl Transferase, eRNA: enhancer RNA, EZH2: Enhancer Of Zeste 2 Polycomb Repressive Complex 2 Subunit, EPM: Elevated plus maze; FASD: Fetal alcohol spectrum disorders; F1: First filial generation; F2: Second filial generation; GAD: Glutamic Acid Decarboxylase, HIST: Histone Gene Cluster, LH: Luteinizing hormone; LDH: Lactate Dehydrogenase, LNC-RNA: long non-coding RNA, LPS: Lipopolysaccharide; MC4R: Melanocortin 4 Receptor, MeCP2: Methyl CpG Binding Protein 2, miRNA: micro RNA, MPFC: Medial Prefrontal Cortex, MTHFR: Methylenetetrahydrofolate Reductase, MWM: Morris water maze; NAc: Nucleus Accumbens; NGF: Nerve growth factor; NPY: Neuropeptide Y, OFT: Open field test; PER2: Period2, PAT: Passive avoidance test; Pomc: Pro-opiomelanocortin; PND: Post-natal day; PFC: Prefrontal cortex; SYT2: Synaptotagmin 2, tDR: tRNA-derived RNA, TXNIP: Thioredoxin-interacting Protein ; TBC: Two-bottle choice;
References
- Ilhan, M.N.; Yapar, D. Alcohol consumption and alcohol policy. Turk. J. Med Sci. 2020, 50, 1197–1202. [Google Scholar] [CrossRef]
- Witkiewitz, K.; Litten, R.Z.; Leggio, L. Advances in the science and treatment of alcohol use disorder. Sci. Adv. 2019, 5, eaax4043. [Google Scholar] [CrossRef] [PubMed]
- Axley, P.D.; Richardson, C.T.; Singal, A.K. Epidemiology of Alcohol Consumption and Societal Burden of Alcoholism and Alcoholic Liver Disease. Clin. Liver Dis. 2019, 23, 39–50. [Google Scholar] [CrossRef] [PubMed]
- A Schuckit, M. Alcohol-use disorders. Lancet 2009, 373, 492–501. [Google Scholar] [CrossRef]
- Roerecke, M. Alcohol’s Impact on the Cardiovascular System. Nutrients 2021, 13, 3419. [Google Scholar] [CrossRef] [PubMed]
- Boffetta, P.; Hashibe, M. Alcohol and cancer. Lancet Oncol. 2006, 7, 149–156. [Google Scholar] [CrossRef]
- Wu, X.; Fan, X.; Miyata, T.; Kim, A.; Ross, C.K.C.-D.; Ray, S.; Huang, E.; Taiwo, M.; Arya, R.; Wu, J.; et al. Recent Advances in Understanding of Pathogenesis of Alcohol-Associated Liver Disease. Annu. Rev. Pathol. Mech. Dis. 2023, 18, 411–438. [Google Scholar] [CrossRef]
- Peng, B.; Yang, Q.; Joshi, R.B.; Liu, Y.; Akbar, M.; Song, B.-J.; Zhou, S.; Wang, X. Role of Alcohol Drinking in Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2020, 21, 2316. [Google Scholar] [CrossRef]
- Marras, C., C. G. Canning and S. M. Goldman. "Environment, lifestyle, and parkinson's disease: Implications for prevention in the next decade." Movement Disorders 34 (2019): 801-11.
- Mohebichamkhorami, F.; Niknam, Z.; Khoramjouy, M.; Heidarli, E.; Ghasemi, R.; Hosseinzadeh, S.; Mohseni, S.S.; Hajikarim-Hamedani, A.; Heidari, A.; Ghane, Y.; et al. Brain Homogenate of a Rat Model of Alzheimer's Disease Modifies the Secretome of 3D Cultured Periodontal Ligament Stem Cells: A Potential Neuroregenerative Therapy. Iran. J. Pharm. Res. 2022, 21, e133668. [Google Scholar] [CrossRef]
- de Jong, S.W.; Huisman, M.H.B.; Sutedja, N.A.; van der Kooi, A.J.; de Visser, M.; Schelhaas, H.J.; Fischer, K.; Veldink, J.H.; Berg, L.H.v.D. Smoking, Alcohol Consumption, and the Risk of Amyotrophic Lateral Sclerosis: A Population-based Study. Am. J. Epidemiology 2012, 176, 233–239. [Google Scholar] [CrossRef]
- Shafiee, A.; Oraii, A.; Jalali, A.; Alaeddini, F.; Saadat, S.; Masoudkabir, F.; Tajdini, M.; Ashraf, H.; Omidi, N.; Heidari, A.; et al. Epidemiology and prevalence of tobacco use in Tehran; a report from the recruitment phase of Tehran cohort study. BMC Public Heal. 2023, 23, 1–14. [Google Scholar] [CrossRef]
- Salehi, N., S. M. Fanaei, S. Hosseini and M. Mohseny. "The prevalence of anxiety and its associated factors among medical interns during the covid-19 outbreak in iran." International Journal of Body, Mind and Culture 9 (2022): 343-50. Available online: https://ijbmc.org/index.php/ijbmc/article/view/387.
- Banerjee, N. Neurotransmitters in alcoholism: A review of neurobiological and genetic studies. Indian J. Hum. Genet. 2014, 20, 20–31. [Google Scholar] [CrossRef]
- Leclercq, S.; de Timary, P.; Delzenne, N.M.; Stärkel, P. The link between inflammation, bugs, the intestine and the brain in alcohol dependence. Transl. Psychiatry 2017, 7, e1048–e1048. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.A. Update on neuropharmacological treatments for alcoholism: Scientific basis and clinical findings. Biochem. Pharmacol. 2008, 75, 34–56. [Google Scholar] [CrossRef]
- Sanchez-Roige, S.; Palmer, A.A.; Clarke, T.-K. Recent Efforts to Dissect the Genetic Basis of Alcohol Use and Abuse. Biol. Psychiatry 2019, 87, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.Y.; Sharma, V.K. DRD2 methylation and regional grey matter volumes in young adult offspring from families at ultra-high risk for alcohol dependence. Psychiatry Res. Neuroimaging 2019, 286, 31–38. [Google Scholar] [CrossRef]
- Abrishamcar, S.; Chen, J.; Feil, D.; Kilanowski, A.; Koen, N.; Vanker, A.; Wedderburn, C.J.; Donald, K.A.; Zar, H.J.; Stein, D.J.; et al. DNA methylation as a potential mediator of the association between prenatal tobacco and alcohol exposure and child neurodevelopment in a South African birth cohort. Transl. Psychiatry 2022, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Miozzo, F.; Arnould, H.; de Thonel, A.; Schang, A.-L.; Sabéran-Djoneidi, D.; Baudry, A.; Schneider, B.; Mezger, V. Alcohol exposure promotes DNA methyltransferase DNMT3A upregulation through reactive oxygen species-dependent mechanisms. Cell Stress Chaperon. 2018, 23, 115–126. [Google Scholar] [CrossRef]
- Pandey, S.C.; Kyzar, E.J.; Zhang, H. Epigenetic basis of the dark side of alcohol addiction. Neuropharmacology 2017, 122, 74–84. [Google Scholar] [CrossRef]
- Ponomarev, I. Epigenetic Control of Gene Expression in the Alcoholic Brain. Alcohol Res. Curr. Rev. 2013, 35, 69–76. [Google Scholar]
- Gangisetty, O.; Sinha, R.; Sarkar, D.K. Hypermethylation of Proopiomelanocortin and Period 2 Genes in Blood Are Associated with Greater Subjective and Behavioral Motivation for Alcohol in Humans. Alcohol. Clin. Exp. Res. 2019, 43, 212–220. [Google Scholar] [CrossRef]
- Ciafre', S.; Carito, V.; Ferraguti, G.; Greco, A.; Chaldakov, G.N.; Fiore, M.; Ceccanti, M. How alcohol drinking affects our genes: an epigenetic point of view. Biochem. Cell Biol. 2019, 97, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Vetreno, R.P.; Broadwater, M.A.; Robinson, D.L. Adolescent Alcohol Exposure Persistently Impacts Adult Neurobiology and Behavior. Pharmacol. Rev. 2016, 68, 1074–1109. [Google Scholar] [CrossRef]
- Zhang, H.; Kyzar, E.J.; Bohnsack, J.P.; Kokare, D.M.; Teppen, T.; Pandey, S.C. Adolescent alcohol exposure epigenetically regulates CREB signaling in the adult amygdala. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Kyzar, E.J.; Zhang, H.; Sakharkar, A.J.; Pandey, S.C. Adolescent alcohol exposure alters lysine demethylase 1 (LSD1) expression and histone methylation in the amygdala during adulthood. Addict. Biol. 2017, 22, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- A Philibert, R.; Penaluna, B.; White, T.; Shires, S.; Gunter, T.; Liesveld, J.; Erwin, C.; Hollenbeck, N.; Osborn, T. A pilot examination of the genome-wide DNA methylation signatures of subjects entering and exiting short-term alcohol dependence treatment programs. Epigenetics 2014, 9, 1212–1219. [Google Scholar] [CrossRef]
- Philibert, R.; Dogan, M.; Noel, A.; Miller, S.; Krukow, B.; Papworth, E.; Cowley, J.; Knudsen, A.; Beach, S.R.H.; Black, D. Genome-wide and digital polymerase chain reaction epigenetic assessments of alcohol consumption. Am. J. Med Genet. Part B: Neuropsychiatr. Genet. 2018, 177, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Herman, A.I.; Kranzler, H.R.; Anton, R.F.; Zhao, H.; Zheng, W.; Gelernter, J. Array-Based Profiling of DNA Methylation Changes Associated with Alcohol Dependence. Alcohol. Clin. Exp. Res. 2013, 37, E108–E115. [Google Scholar] [CrossRef]
- Meng, W.; Sjöholm, L.K.; Kononenko, O.; Tay, N.; Zhang, D.; Sarkisyan, D.; Geske, J.R.; Ing, A.; Qiu, W.; Watanabe, H.; et al. Genotype-dependent epigenetic regulation of DLGAP2 in alcohol use and dependence. Mol. Psychiatry 2019, 26, 4367–4382. [Google Scholar] [CrossRef]
- Hillemacher, T.; Rhein, M.; Burkert, A.; Heberlein, A.; Wilhelm, J.; Glahn, A.; Muschler, M.A.N.; Kahl, K.G.; Kornhuber, J.; Bleich, S.; et al. DNA-methylation of the dopamin receptor 2 gene is altered during alcohol withdrawal. Eur. Neuropsychopharmacol. 2019, 29, 1250–1257. [Google Scholar] [CrossRef]
- Muschler, M.A.N.; Hillemacher, T.; Kraus, C.; Kornhuber, J.; Bleich, S.; Frieling, H. DNA methylation of the POMC gene promoter is associated with craving in alcohol dependence. J. Neural Transm. 2010, 117, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, M.; Koycheva, Y.; Taseva, T.; Simeonova, S. Changes in the Expression of DNA Methylation Related Genes in Leukocytes of Persons with Alcohol and Drug Dependence. Acta Medica Bulg. 2020, 47, 11–17. [Google Scholar] [CrossRef]
- Maeda, K.; Yamada, H.; Munetsuna, E.; Fujii, R.; Yamazaki, M.; Ando, Y.; Mizuno, G.; Ishikawa, H.; Ohashi, K.; Tsuboi, Y.; et al. Association of drinking behaviors with TXNIP DNA methylation levels in leukocytes among the general Japanese population. Am. J. Drug Alcohol Abus. 2022, 48, 302–310. [Google Scholar] [CrossRef]
- Dugue, P.A.; Wilson, R.; Lehne, B.; Jayasekara, H.; Wang, X.; Jung, C.H.; Joo, J.E.; Makalic, E.; Schmidt, D.F.; Baglietto, L.; et al. Alcohol consumption is associated with widespread changes in blood DNA methylation: Analysis of cross-sectional and longitudinal data. Addict. Biol. 2021, 26, e12855. [Google Scholar] [CrossRef]
- Soundararajan, S.; Agrawal, A.; Purushottam, M.; Anand, S.D.; Shankarappa, B.; Sharma, P.; Jain, S.; Murthy, P. Changes in DNA methylation persist over time in males with severe alcohol use disorder—A longitudinal follow-up study. Am. J. Med Genet. Part B: Neuropsychiatr. Genet. 2021, 186, 183–192. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, R.; Li, W.; Liao, Y.; Tang, J.; Miao, Q.; Hao, W. Genome-wide DNA methylation patterns in discordant sib pairs with alcohol dependence. Asia-Pacific Psychiatry 2013, 5, 39–50. [Google Scholar] [CrossRef]
- Lenz, B.; Reulbach, U.; Kornhuber, J.; Bleich, S.; Bönsch, D. Homocysteine associated genomic DNA hypermethylation in patients with chronic alcoholism. J. Neural Transm. 2004, 111, 1611–1616. [Google Scholar] [CrossRef]
- Clark, S.L.; Chan, R.F.; Zhao, M.; Xie, L.Y.; Copeland, W.E.; Penninx, B.W.J.H.; Aberg, K.A.; Oord, E.J.C.G.v.D. Dual methylation and hydroxymethylation study of alcohol use disorder. Addict. Biol. 2022, 27, e13114–e13114. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, M.; Bollepalli, S.; Cazaly, E.; Salvatore, J.E.; Barr, P.; Rose, R.J.; Dick, D.; Kaprio, J.; Ollikainen, M. Associations of Alcohol Consumption With Epigenome-Wide DNA Methylation and Epigenetic Age Acceleration: Individual-Level and Co-twin Comparison Analyses. Alcohol. Clin. Exp. Res. 2021, 45, 318–328. [Google Scholar] [CrossRef]
- Lu, M.; Xueying, Q.; Hexiang, P.; Wenjing, G.; Hägg, S.; Weihua, C.; Chunxiao, L.; Canqing, Y.; Jun, L.; Zengchang, P.; et al. Genome-wide associations between alcohol consumption and blood DNA methylation: evidence from twin study. Epigenomics 2021, 13, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Lohoff, F.W.; Roy, A.; Jung, J.; Longley, M.; Rosoff, D.B.; Luo, A.; O’connell, E.; Sorcher, J.L.; Sun, H.; Schwandt, M.; et al. Epigenome-wide association study and multi-tissue replication of individuals with alcohol use disorder: evidence for abnormal glucocorticoid signaling pathway gene regulation. Mol. Psychiatry 2021, 26, 2224–2237. [Google Scholar] [CrossRef] [PubMed]
- Manzardo, A.; Henkhaus, R.; Butler, M. Global DNA promoter methylation in frontal cortex of alcoholics and controls. Gene 2012, 498, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Ponomarev, I.; Wang, S.; Zhang, L.; Harris, R.A.; Mayfield, R.D. Gene Coexpression Networks in Human Brain Identify Epigenetic Modifications in Alcohol Dependence. J. Neurosci. 2012, 32, 1884–1897. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, J.P.; Teppen, T.; Kyzar, E.J.; Dzitoyeva, S.; Pandey, S.C. The lncRNA BDNF-AS is an epigenetic regulator in the human amygdala in early onset alcohol use disorders. Transl. Psychiatry 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Ouko, L.A.; Shantikumar, K.; Knezovich, J.; Haycock, P.; Schnugh, D.J.; Ramsay, M. Effect of Alcohol Consumption on CpG Methylation in the Differentially Methylated Regions ofH19andIG-DMRin Male Gametes-Implications for Fetal Alcohol Spectrum Disorders. Alcohol. Clin. Exp. Res. 2009, 33, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Trautman, A.; Gurumoorthy, A.; Hansen, K.A. Effects of alcohol use on sperm chromatin structure, a retrospective analysis. Basic Clin. Androl. 2023, 33, 1–15. [Google Scholar] [CrossRef]
- Blaseg, E.; Von Wald, T.; Hansen, K.A. Vitamin D levels and human sperm DNA fragmentation: a prospective, cohort study. Basic Clin. Androl. 2022, 32, 1–7. [Google Scholar] [CrossRef]
- Wieting, J.; Rhein, M.; Hillemacher, T.; Lichtinghagen, R.; Hoppe, V.; Müschen, L.; Glahn, A.; Frieling, H.; Bleich, S.; Muschler, M. DNA Methylation of the Leptin Gene Promoter is Altered by Chronic Alcohol Exposure in an Animal Model for Alcohol Dependence. Eur. Addict. Res. 2019, 25, 49–55. [Google Scholar] [CrossRef]
- Sakharkar, A.J.; Kyzar, E.J.; Gavin, D.P.; Zhang, H.; Chen, Y.; Krishnan, H.R.; Grayson, D.R.; Pandey, S.C. Altered amygdala DNA methylation mechanisms after adolescent alcohol exposure contribute to adult anxiety and alcohol drinking. Neuropharmacology 2019, 157, 107679–107679. [Google Scholar] [CrossRef] [PubMed]
- Kokare, D.M.; Kyzar, E.J.; Zhang, H.; Sakharkar, A.J.; Pandey, S.C. Adolescent Alcohol Exposure-Induced Changes in Alpha-Melanocyte Stimulating Hormone and Neuropeptide Y Pathways via Histone Acetylation in the Brain During Adulthood. Int. J. Neuropsychopharmacol. 2017, 20, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Cervera-Juanes, R.; Wilhelm, L.J.; Park, B.; A Grant, K.; Ferguson, B. Alcohol-dose-dependent DNA methylation and expression in the nucleus accumbens identifies coordinated regulation of synaptic genes. Transl. Psychiatry 2017, 7, e994–e994. [Google Scholar] [CrossRef]
- Barbier, E.; Tapocik, J.D.; Juergens, N.; Pitcairn, C.; Borich, A.; Schank, J.R.; Sun, H.; Schuebel, K.; Zhou, Z.; Yuan, Q.; et al. DNA Methylation in the Medial Prefrontal Cortex Regulates Alcohol-Induced Behavior and Plasticity. J. Neurosci. 2015, 35, 6153–6164. [Google Scholar] [CrossRef]
- Chastain, L.G.; Franklin, T.; Gangisetty, O.; Cabrera, M.A.; Mukherjee, S.; Shrivastava, P.; Jabbar, S.; Sarkar, D.K. Early life alcohol exposure primes hypothalamic microglia to later-life hypersensitivity to immune stress: possible epigenetic mechanism. Neuropsychopharmacology 2019, 44, 1579–1588. [Google Scholar] [CrossRef]
- Montesinos, J.; Pascual, M.; Rodríguez-Arias, M.; Miñarro, J.; Guerri, C. Involvement of tlr4 in the long-term epigenetic changes, rewarding and anxiety effects induced by intermittent ethanol treatment in adolescence. Brain Behav. Immun. 2016, 53, 159–171. [Google Scholar] [CrossRef]
- Sharma, R.; Sahota, P.; Thakkar, M.M. Chronic alcohol exposure reduces acetylated histones in the sleep-wake regulatory brain regions to cause insomnia during withdrawal. Neuropharmacology 2020, 180, 108332–108332. [Google Scholar] [CrossRef]
- Berkel, T.D.M.; Zhang, H.; Teppen, T.; Sakharkar, A.J.; Pandey, S.C. Essential Role of Histone Methyltransferase G9a in Rapid Tolerance to the Anxiolytic Effects of Ethanol. Int. J. Neuropsychopharmacol. 2019, 22, 292–302. [Google Scholar] [CrossRef]
- Kyzar, E.J.; Bohnsack, J.P.; Zhang, H.; Pandey, S.C. MicroRNA-137 Drives Epigenetic Reprogramming in the Adult Amygdala and Behavioral Changes after Adolescent Alcohol Exposure. eneuro 2019, 6. [Google Scholar] [CrossRef]
- Kyzar, E.J.; Zhang, H.; Pandey, S.C. Adolescent Alcohol Exposure Epigenetically Suppresses Amygdala Arc Enhancer RNA Expression to Confer Adult Anxiety Susceptibility. Biol. Psychiatry 2019, 85, 904–914. [Google Scholar] [CrossRef]
- Barney, T.M.; Vore, A.S.; Deak, T. Acute Ethanol Challenge Differentially Regulates Expression of Growth Factors and miRNA Expression Profile of Whole Tissue of the Dorsal Hippocampus. Front. Neurosci. 2022, 16, 884197. [Google Scholar] [CrossRef]
- Rossetto, I.; Cagnon, V.; Lizarte, F.; Tirapelli, L.; Tirapelli, D.; Arantes, R.; Chuffa, L.; Martinez, F.; Martinez, M. Ethanol and caffeine consumption modulates the expression of miRNAs in the cerebellum and plasma of UChB rats. Life Sci. 2019, 229, 180–186. [Google Scholar] [CrossRef]
- Bielawski, D.M.; Zaher, F.M.; Svinarich, D.M.; Abel, E.L. Paternal alcohol exposure affects sperm cytosine methyltransferase messenger RNA levels. Alcohol. Clin. Exp. Res. 2002, 26, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Knezovich, J.G.; Ramsay, M. The Effect of Preconception Paternal Alcohol Exposure on Epigenetic Remodeling of the H19 and Rasgrf1 Imprinting Control Regions in Mouse Offspring. Front. Genet. 2012, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Finegersh, A.; Homanics, G.E. Paternal Alcohol Exposure Reduces Alcohol Drinking and Increases Behavioral Sensitivity to Alcohol Selectively in Male Offspring. PLOS ONE 2014, 9, e99078. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Diao, L.; Liu, J.; Jiang, N.; Zhang, J.; Wang, H.; Zhou, W.; Huang, G.; Ma, D. Paternal ethanol exposure and behavioral abnormities in offspring: Associated alterations in imprinted gene methylation. Neuropharmacology 2014, 81, 126–133. [Google Scholar] [CrossRef]
- Govorko, D.; Bekdash, R.A.; Zhang, C.; Sarkar, D.K. Male Germline Transmits Fetal Alcohol Adverse Effect on Hypothalamic Proopiomelanocortin Gene Across Generations. Biol. Psychiatry 2012, 72, 378–388. [Google Scholar] [CrossRef]
- Rahimipour, M.; Talebi, A.R.; Anvari, M.; Sarcheshmeh, A.A.; Omidi, M. Effects of different doses of ethanol on sperm parameters, chromatin structure and apoptosis in adult mice. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 423–428. [Google Scholar] [CrossRef]
- Rompala, G.R.; Mounier, A.; Wolfe, C.M.; Lin, Q.; Lefterov, I.; Homanics, G.E. Heavy Chronic Intermittent Ethanol Exposure Alters Small Noncoding RNAs in Mouse Sperm and Epididymosomes. Front. Genet. 2018, 9, 32. [Google Scholar] [CrossRef]
- Yang, Q.; Lin, J.; Liu, M.; Li, R.; Tian, B.; Zhang, X.; Xu, B.; Liu, M.; Zhang, X.; Li, Y.; et al. Highly sensitive sequencing reveals dynamic modifications and activities of small RNAs in mouse oocytes and early embryos. Sci. Adv. 2016, 2, e1501482–1501482. [Google Scholar] [CrossRef]
- Jiajie, T.; Yanzhou, Y.; Hoi-Hung, A.C.; Zi-Jiang, C.; Wai-Yee, C. Conserved miR-10 family represses proliferation and induces apoptosis in ovarian granulosa cells. Sci. Rep. 2017, 7, srep41304. [Google Scholar] [CrossRef]
- Bedi, Y.S.; Wang, H.; Thomas, K.N.; Basel, A.; Prunier, J.; Robert, C.; Golding, M.C. Alcohol induced increases in sperm Histone H3 lysine 4 trimethylation correlate with increased placental CTCF occupancy and altered developmental programming. Sci. Rep. 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Riley, E.P.; McGee, C.L. Fetal Alcohol Spectrum Disorders: An Overview with Emphasis on Changes in Brain and Behavior. Exp. Biol. Med. 2005, 230, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, S.; Chastain, L.G.; Gangisetty, O.; A Cabrera, M.; Sochacki, K.; Sarkar, D.K. Preconception Alcohol Increases Offspring Vulnerability to Stress. Neuropsychopharmacology 2016, 41, 2782–2793. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Diao, L.; Liu, J.; Jiang, N.; Zhang, J.; Wang, H.; Zhou, W.; Huang, G.; Ma, D. Paternal ethanol exposure and behavioral abnormities in offspring: Associated alterations in imprinted gene methylation. Neuropharmacology 2014, 81, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.L.; Burling, T.A.; Lochry, E.A.; Sutker, P.B. The effect of paternal alcohol consumption on fetal development in mice. Drug Alcohol Depend. 1982, 9, 89–95. [Google Scholar] [CrossRef]
- A Jamerson, P.; Wulser, M.J.; Kimler, B.F. Neurobehavioral effects in rat pups whose sires were exposed to alcohol. Dev. Brain Res. 2004, 149, 103–111. [Google Scholar] [CrossRef]
- Coccurello, R.; Carito, V.; Ciafrè, S.; Ferraguti, G.; Giacovazzo, G.; Mancinelli, R.; Tirassa, P.; Chaldakov, G.N.; Pascale, E.; Ceccanti, M.; et al. Paternal alcohol exposure in mice alters brain NGF and BDNF and increases ethanol-elicited preference in male offspring. Addict. Biol. 2016, 21, 776–787. [Google Scholar] [CrossRef]
- Meek, L.R.; Myren, K.; Sturm, J.; Burau, D. Acute paternal alcohol use affects offspring development and adult behavior. Physiol. Behav. 2007, 91, 154–160. [Google Scholar] [CrossRef]
- Asimes, A.; Kim, C.K.; Cuarenta, A.; Auger, A.P.; Pak, T.R. Binge Drinking and Intergenerational Implications: Parental Preconception Alcohol Impacts Offspring Development in Rats. J. Endocr. Soc. 2018, 2, 672–686. [Google Scholar] [CrossRef]
- Rompala, G.R.; Finegersh, A.; Homanics, G.E. Paternal preconception ethanol exposure blunts hypothalamic-pituitary-adrenal axis responsivity and stress-induced excessive fluid intake in male mice. Alcohol 2016, 53, 19–25. [Google Scholar] [CrossRef]
- Abel, E.L.; Lee, J.A. Paternal Alcohol Exposure Affects Offspring Behavior but not Body or Organ Weights in Mice. Alcohol. Clin. Exp. Res. 1988, 12, 349–355. [Google Scholar] [CrossRef]
- Kim, P.; Choi, C.S.; Park, J.H.; Joo, S.H.; Kim, S.Y.; Ko, H.M.; Kim, K.C.; Jeon, S.J.; Park, S.H.; Han, S.-H.; et al. Chronic exposure to ethanol of male mice before mating produces attention deficit hyperactivity disorder-like phenotype along with epigenetic dysregulation of dopamine transporter expression in mouse offspring. J. Neurosci. Res. 2014, 92, 658–670. [Google Scholar] [CrossRef]
- Ledig, M.; Misslin, R.; Vogel, E.; Holownia, A.; Copin, J.; Tholey, G. Paternal alcohol exposure: developmental and behavioral effects on the offspring of rats. Neuropharmacology 1998, 37, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.F.; Cicero, T.J.; Kettinger, L.; Meyer, E.R. Paternal alcohol consumption in the rat impairs spatial learning performance in male offspring. Psychopharmacol. 1991, 105, 289–302. [Google Scholar] [CrossRef]
- Finegersh, A.; Homanics, G.E. Paternal Alcohol Exposure Reduces Alcohol Drinking and Increases Behavioral Sensitivity to Alcohol Selectively in Male Offspring. PLOS ONE 2014, 9, e99078. [Google Scholar] [CrossRef]
- Bottom, R.T.; Kozanian, O.O.; Rohac, D.J.; Erickson, M.A.; Huffman, K.J. Transgenerational Effects of Prenatal Ethanol Exposure in Prepubescent Mice. Front. Cell Dev. Biol. 2022, 10, 812429. [Google Scholar] [CrossRef] [PubMed]
- Koabel, J.; McNivens, M.; McKee, P.; Pautassi, R.; Bordner, K.; Nizhnikov, M. The offspring of alcohol-exposed sires exhibit heightened ethanol intake and behavioral alterations in the elevated plus maze. Alcohol 2021, 92, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Beeler, E.; Nobile, Z.L.; Homanics, G.E. Paternal Preconception Every-Other-Day Ethanol Drinking Alters Behavior and Ethanol Consumption in Offspring. Brain Sci. 2019, 9, 56. [Google Scholar] [CrossRef]
- Nieto, S.J.; Harding, M.J.; Nielsen, D.A.; Kosten, T.A. Paternal alcohol exposure has task- and sex-dependent behavioral effect in offspring. Alcohol. Clin. Exp. Res. 2022, 46, 2191–2202. [Google Scholar] [CrossRef]
- Sullivan, E.V.; Pfefferbaum, A. Neurocircuitry in alcoholism: a substrate of disruption and repair. Psychopharmacol. 2005, 180, 583–594. [Google Scholar] [CrossRef]
- Boden, J.M.; Fergusson, D.M. Alcohol and depression. Addiction 2011, 106, 906–914. [Google Scholar] [CrossRef]
- Orford, J.; Velleman, R. Offspring of parents with drinking problems: drinking and drug-taking as young adults. Addiction 1990, 85, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Rompala, G.R.; Finegersh, A.; Homanics, G.E. Paternal preconception ethanol exposure blunts hypothalamic-pituitary-adrenal axis responsivity and stress-induced excessive fluid intake in male mice. Alcohol 2016, 53, 19–25. [Google Scholar] [CrossRef]
- Beeler, E.; Nobile, Z.L.; Homanics, G.E. Paternal Preconception Every-Other-Day Ethanol Drinking Alters Behavior and Ethanol Consumption in Offspring. Brain Sci. 2019, 9, 56. [Google Scholar] [CrossRef]
- Nieto, S.J.; Haile, C.N.; Quave, C.B.; Harding, M.J.; Nielsen, D.A.; Meisch, R.A.; Kosten, T.A. Paternal alcohol exposure reduces acquisition of operant alcohol self-administration and affects Bdnf DNA methylation in male and female offspring. Addict. Biol. 2022, 27, e13078. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Choi, C.S.; Park, J.H.; Joo, S.H.; Kim, S.Y.; Ko, H.M.; Kim, K.C.; Jeon, S.J.; Park, S.H.; Han, S.-H.; et al. Chronic exposure to ethanol of male mice before mating produces attention deficit hyperactivity disorder-like phenotype along with epigenetic dysregulation of dopamine transporter expression in mouse offspring. J. Neurosci. Res. 2014, 92, 658–670. [Google Scholar] [CrossRef]
- Meek, L.R.; Myren, K.; Sturm, J.; Burau, D. Acute paternal alcohol use affects offspring development and adult behavior. Physiol. Behav. 2007, 91, 154–160. [Google Scholar] [CrossRef]
- Suresh, S.; Abozaid, A.; Tsang, B.; Gerlai, R. Exposure of parents to alcohol alters behavior of offspring in zebrafish. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2021, 111, 110143. [Google Scholar] [CrossRef] [PubMed]
- Al-Yasari, A.; Jabbar, S.; A Cabrera, M.; Rousseau, B.; Sarkar, D.K. Preconception Alcohol Exposure Increases the Susceptibility to Diabetes in the Offspring. Endocrinology 2021, 162. [Google Scholar] [CrossRef] [PubMed]
- Asimes, A.; Torcaso, A.; Pinceti, E.; Kim, C.K.; Zeleznik-Le, N.J.; Pak, T.R. Adolescent binge-pattern alcohol exposure alters genome-wide DNA methylation patterns in the hypothalamus of alcohol-naïve male offspring. Alcohol 2017, 60, 179–189. [Google Scholar] [CrossRef]
- Jabbar, S.; Chastain, L.G.; Gangisetty, O.; A Cabrera, M.; Sochacki, K.; Sarkar, D.K. Preconception Alcohol Increases Offspring Vulnerability to Stress. Neuropsychopharmacology 2016, 41, 2782–2793. [Google Scholar] [CrossRef]
- Przybycien-Szymanska, M.M.; Rao, Y.S.; Prins, S.A.; Pak, T.R. Parental Binge Alcohol Abuse Alters F1 Generation Hypothalamic Gene Expression in the Absence of Direct Fetal Alcohol Exposure. PLOS ONE 2014, 9, e89320. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.C.; Skiles, W.M.; Chronister, S.S.; Wang, H.; Sutton, G.I.; Bedi, Y.S.; Snyder, M.; Long, C.R.; Golding, M.C. DNA methylation-independent growth restriction and altered developmental programming in a mouse model of preconception male alcohol exposure. Epigenetics 2017, 12, 841–853. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Ethanol-induced epigenetics changes.
| Study | Species | Site of Investigation | DNA Methylation | Histone Modification | miRNA |
|---|---|---|---|---|---|
| Trautman, 2023 [48] | Human | Sperm | Alcohol use did not show a significant association with the High DNA Stainability or DNA Fragmentation Index of sperm | N/A | N/A |
| Abrishamcar, 2022 [19] | Human | Cord blood samples | Identified 6 CpG sites associated with mQLT Hypermethylation of MAD1L1, CAMTA1, and ALDH1A genes |
N/A | N/A |
| Clark, 2022 [40] | Human | Post-mortem brain samples | Epigenomic deconvolution was utilized to conduct methylome-wide association studies specific to cell types within granulocytes, T-cells, B-cells, and monocytes in 1,132 blood samples. | N/A | N/A |
| Blaseg, 2022 [49] | Human/male | Serum and sperm | In non-drinkers, the levels of High DNA Stainability increased | N/A | N/A |
| Bedi, 2022 [72] | Mice/male | Sperm | No significant alterations in s-adenosylmethionine or DNA methylation levels were detected in the study | Widespread elevation in sperm H3K4me3 enrichment was observed | N/A |
| Barney, 2022 [61] | Rats | Blood and brain (amygdala and dorsal hippocampus) | N/A | N/A | Male and female rats were administered saline or ethanol (3.5 g/kg), and their brains were collected three hours after the injection. The purpose was to evaluate the expression of growth factor, cytokine, or miRNA |
| Maeda, 2021 [35] | Human | Blood samples(leukocytes) | A cross-sectional study was conducted on 404 individuals, comprising 176 males and 228 females. The participants were categorized into non-drinkers, moderate drinkers, and heavy drinkers based on their self-reported drinking habits. The pyrosequencing assay was utilized to determine the levels of TXNIP DNA methylation in leukocytes | N/A | N/A |
| Stephenson, 2021 [41] | Human | Whole-blood samples | The sample was comprised of individuals sourced from the Finnish Twin Cohorts. Participants self-reported their alcohol consumption within the previous week, and their epigenome-wide DNA methylation was evaluated in whole-blood samples | N/A | N/A |
| Soundararajan, 2021 [37] | Human/male | Blood samples | alterations in DNA methylation patterns within peripheral blood leukocytes of individuals with AUD who were actively seeking treatment •gene-specific methylation at ALDH2/MTHFR and global methylation |
N/A | N/A |
| Lu, 2021 [42] | Human | Whole blood samples | • Total of 57 pairs of monozygotic (MZ) twins who exhibited discordance in alcohol consumption, sourced from the Chinese National Twin Registry • 158 pairs of both monozygotic (MZ) and dizygotic twins were assessed from the Swedish Adoption/Twin Study of Aging |
N/A | N/A |
| Lohoff, 2021 [43] | Human | Blood samples | • Extensive analysis of DNA methylation epigenome-wide association in AUD, comprising 625 participants. A cross-tissue/cross-phenotypic approach was employed to replicate the top hit (N = 4798) and identify novel epigenetic targets relevant to AUD | N/A | N/A |
| Sharma, 2020 [57] | Mice/male (C57BL/6J) |
Basal forebrain Lateral hypothalamus |
N/A | Expression of acetylated histones, H3 lysine 14 (AcH3K14), was examined in two major sleep-wake regulatory brain regions | N/A |
| Krasteva, 2020 [34] | Human | Blood samples | The expression pattern of genes associated with DNA methylation, such as DNMT1 and DNMT3a methyltransferases, as well as MeCP2, MBD1, MBD2, MBD3, and MBD4 methyl CpG binding domain proteins, was assessed in blood samples obtained from individuals with alcohol and drug dependence, in comparison to those who abstain from substance use and are in good health. | N/A | N/A |
| Dugué, 2019 [36] | Human | Blood samples | Using the Illumina HumanMethylation450 BeadChip, DNA methylation was measured in blood samples from 5606 Melbourne Collaborative Cohort Study participants | N/A | N/A |
| Chastain, 2019 [55] | Rat/male | Hypothalamic microglia | Decreased global DNA methylation in PND 2–6 alcohol-exposed |
Increased global acetylation of histone 3 lysine 9 (H3K9a) in PND 2–6 alcohol-exposed | N/A |
| Gangisetty, 2019 [23] | Human | Peripheral blood sample | Kyzar, 2019 [60] | Rat/male | Amygdala |
| Hillemacher, 2019 [32] | Human | Blood samples | A significant increase in DRD2-gene methylation during alcohol withdrawal/early abstinence | N/A | N/A |
| Rossetto, 2019 [62] | Rat | Cerebellum and plasma | N/A | N/A | Increased the levels of specific miRNAs, including miR-155-5p, miR-146a-5p, miR-126-3p, and miR-132-3p, in the cerebellum |
| Kyzar, 2019 [61] | Rat | Amygdala | N/A | N/A | miRNA 137 was implicated in the process of chromatin remodeling at BDNF exon IV |
| Kyzar, 2019 [60] | Rat/male | Amygdala | Decreased Arc eRNA expression | Increased H3K27me3 occupancy at the Arc SARE site and five other locations across the Arc promoter and gene body in the amygdala of AIE adult rats compared with AIS rats | N/A |
| Meng, 2019 [31] | Cohort1: human/male Cohort2: human |
Cohort 1: Brain tissues Cohort 2: Whole blood samples |
Methylation within DMR-DLGAP2 | N/A | N/A |
| Sakharkar, 2019 [51] | Rat/male | Amygdala | One hour after exposure: decreased DNMT activity and DNMT3b expression, increased inducible protein 45 mRNA expressions in AIE rats compared to AIS rats 24 hours after exposure: increased DNMT activity but normalized Gadd45 and DNMT3b mRNA expression in AIE rats compared to AIS rats Adulthood: increased DNA methylation of Npy and BDNF exon IV is in the AIE compared to AIS |
N/A | N/A |
| Wieting, 2019 [50] | Rat/male | Blood samples | Increased leptin gene promotor methylation | N/A | N/A |
| Bohnsack, 2019 [46] | Human | Amygdala | N/A | N/A | miRNA 137 was implicated in the process of chromatin remodeling at BDNF exon IV |
| Berkel, 2019 [58] | Rat/male | Amygdala | N/A | Decrease in the levels of H3K9me2 and G9a protein in the amygdala | N/A |
| Zhang, 2018 [26] | Rat/male | Amygdala | N/A | Lower occupancy of acetylated histone H3K9/14 proteins at particular sites in the Creb1, Cbp, and p300 gene promoter regions in amygdala | N/A |
| Philibert, 2018 [29] | Human | Whole blood DNA | Genome-wide methylation | N/A | N/A |
| Rompala, 2018 [69] | Mice/male | Sperm | N/A | N/A | Chronic intermittent alcohol exposure has induced changes in multiple miRNA and tDR (tRNA-derived RNA) species in sperm |
| Cervera-Juanes, 2017 [53] | Rhesus macaques/male | Nucleus accumbens | 17 DMRs, including 14 with methylation levels associated with alcohol consumption Decreased LRP5-DMR methylation Increased JAKMIP1-DMR and GPR39-DMR methylation |
N/A | N/A |
| Kyzar, 2017 [27] | Rat | Amygdala | N/A | Exposure to ethanol causes a reduction in the levels of messenger RNA (mRNA) associated with H3 lysine 9 demethylation (H3K9me2) in the amygdala | N/A |
| Jiajie, 2017 [71] | Human, mouse, and rat | Ovarian granulosa cells | N/A | N/A | miRNA is suggested to function as a posttranscriptional regulator of the alcohol-related gene BDNF |
| Kokare, 2017 [52] | Rat/male | Amygdala and hypothalamus | N/A | Decreased histone H3K9/14 acetylation in the neuropeptide Y promoter in the amygdala, increased histone H3K9/14 acetylation in the melanocortin four receptor gene promoter in the amygdala, and the melanocortin four receptor and pro-opiomelanocortin promoters in the hypothalamus of adolescent intermittent ethanol-exposed adult rats compared with controls | N/A |
| Montesinos, 2016 [56] | Mice/female | Medial prefrontal cortex | Increased expression of BDNF and FosB in wild-type mice | Up-regulated acetylation levels at H3 (K9)and H4 (K5, K12), decreased the tri-methylation of the H3 (K4) levels in adolescent mice after ethanol treatment in wild-type mice | N/A |
| Yang, 2016 [70] | Mice | Oocytes and early embryos | N/A | N/A | The small RNAs that exhibited increased levels of sperm due to alcohol exposure |
| Crews, 2016 [25] | Human | Review | Alcohol at different stages of life can disrupt chromatin function and DNA methylation | N/A | N/A |
| Barbier, 2015 [54] | Rat/male | Medial prefrontal cortex | Global DNA methylation in the medial prefrontal cortex Hypermethylation of the promoter region and exon 1 of Syt2 and on the promoter region of Cacna1a in the medial prefrontal cortex |
N/A | N/A |
| Philibert, 2014 [28] | Human | Blood cells | genome-wide DNA methylation analyses in mononuclear blood cells, finding significant changes in DNA methylation levels between patients with alcohol use disorder and healthy control subjects | N/A | N/A |
| Liang, 2014 [66] | Mice/male | Sperm and in the cerebral cortices | DNA methylation levels at imprinted gene loci are decreased in the sperm of mice subjected to chronic alcohol treatment | N/A | N/A |
| Finegersh, 2014 [65] | Mice/male | Sperm | DNA methylation levels at imprinted gene loci are decreased in the sperm of mice subjected to chronic alcohol treatment | N/A | N/A |
| Zhao, 2013 [38] | Human | Peripheral blood mononuclear cells | 865 hypomethylated and 716 hypermethylated CG sites in individuals who consumed alcohol | N/A | N/A |
| Zhang, 2013 [30] | Human | Peripheral blood cells | DNA methylation patterns based on ethnicity were observed in individuals with alcohol addiction | N/A | N/A |
| Ponomarev, 2012 [45] | Human | Amygdala and frontal cortex | evidenced by the association between DNA hypomethylation and increased histone H3K4 trimethylation in alcoholics | N/A | |
| Manzardo, 2012 [44] | Human | Human | No significant differences between alcohol-dependent individuals and controls in terms of global methylation parameters or most gene targets | DNA methylation was detected in various variants of histone proteins, particularly those associated with gene loci in the HIST2 domain of chromosome 1 | N/A |
| Knezovich, 2012 [64] | Mice/male | Sperm | DNA methylation levels at imprinted gene loci are decreased in the sperm of mice subjected to chronic alcohol treatment | N/A | N/A |
| Muschler, 2010 [33] | Human | Hypothalamic–pituitary–adrenal axis in the brain | Different drinking patterns demonstrated an increase in DNA methylation of POMC genes among binge and heavy drinkers | N/A | N/A |
| Ouko, 2009 [47] | Human | Sperm | Increase methylation levels at the promoters of the POMC gene in sperm | N/A | N/A |
| Bönsch, 2004 [39] | Human | Peripheral blood cells | Global DNA hypermethylation in the mononuclear cells | N/A | N/A |
| Bielawski, 2002 [63] | Rat/male | Sperm | Decrease in the levels of DNMT1 in the sperm of rodents | N/A | N/A |
Abbreviations: ACC: Anterior cingulate cortex; AIE: Adolescent intermittent ethanol; AIS: Adolescent intermittent saline; AUD: Alcohol use disorder; BA9: Brodmann area 9; BDNF: Brain-derived neurotrophic factor; CN: Caudate nucleus; DMR: significant differential methylation region; DNMT: DNA methyl-transferase; Npy : Neuropeptide Y; POMC: Pro-opiomelanocortin gene, PUT: Putamen, VS: Ventral Striatum.
Table 2.
Phenotype and behavioral changes induced by parental ethanol exposure.
| Study | Species/Gender of Alcoholic Subjects | Ethanol Exposure Protocol | Abstinent Time | Studied Generation(s) | Phenotype and Molecular Changes |
|---|---|---|---|---|---|
| Nieto, 2022 [96] | Rat/Male | 5 days each week for 6 weeks | Mating 8 weeks later | Male and female/ F1 | Lower levels of BDNF DNA methylation in the NAc and higher levels in the mPFC in alcohol-exposed sires. Aaberrant BDNF DNA methylation patterns in alcohol-sired offspring of both sexes compared to control-sired offspring. |
| Nieto, 2022 [90] | Rat/Male | For 6 weeks (5 days/week) | Mating 8 weeks after exposure | Male and female/F1 | EPM: Less anxiety-like behavior in alcohol-sired males that were not affected by alcohol administration. OFT: No significant differences in OFT results in both male and female offspring Paternal alcohol exposure provides protection against the initial reinforcing effects of alcohol. Anxiolytic effects of alcohol in the open field in male offspring only with no paternal alcohol effect No change in locomotor activity in either sex as a result of either paternal alcohol exposure or alcohol administration Sex-specific effects of paternal alcohol exposure in the rotarod test Blunted sensitivity to the alcohol's motor-impairing effects in alcohol-sired male offspring, enhanced sensitivity in alcohol-sired female offspring |
| Bottom, 2022 [87] | Mice/Male | Prenatally exposed | Mate during adulthood | Male and female/F1, F2 | Sensory processing deficits, increased risk-taking behavior CD1 mice exposed to ethanol prenatally resulted in no change in anxiety-like behavior in offspring male CD1 mice prenatally exposed to ethanol, the depressive-like behavior increased Behavioral abnormalities are linked to parental ethanol exposure in mice, including abnormal sensorimotor processing No evidence of heritable, transgenerational phenotypes following a multi-level analysis of several key neocortical components, including sensory area connectivity, gene expression, and spine density analysis Decreased brain/body weights, altered Id2 expression, and reduced spine density in S1 neurons in F1 |
| Al-Yasari, 2021 [100] | Rat/Female | 30 days of treatment | 3 weeks after the last exposure | Male and female/ F1 | An altered stress gene network involving glucose metabolism in oocytes in female rats fed with alcohol in the liquid diet, but not with the isocaloric liquid diet, prior to conceptionF1: Significant hyperglycemia and hypoinsulinemia in the offspring born from preconception alcohol-fed mothers when they were adults, increased levels of inflammatory cytokines and cellular apoptosis in the pancreas, altered insulin production and actions in the liver, and a reduced number of proopiomelanocortin neurons in the hypothalamus |
| Suresh, 2021 [99] | Zebrafish/Female and male | For 7 days | Mating 7 hours after last exposure | Male and female/F1 | Increased duration and reduced frequency of immobility, increased turn angle, and increased intra-individual variance of turn angle in offspring of alcohol-exposed parents displayed anxiety and exploratory behavior alterations. |
| Koabel, 2021 [88] | Rat/Male | For 6 weeks, 5 days per week | Mating 8 weeks after last exposure | Male and female/F1 | Ethanol administration decreased anxiety in male progenies of ethanol-sired rats but not in female progenies OFT: Paternal alcohol exposure does not affect rats of either sex's baseline anxiety-like behavior |
| Rossetto, 2019 [62] | UChB rat/Male | Ethanol: 70 days Caffeine: 55 days |
NA | NA | Expression of miRNAs associated with the inflammatory process in the tissue and plasma of the UChB rats after ethanol and caffeine exposure. Increased expression of the analyzed miRNAs155-5p, -146a-5p, -126-3p, -132-3p for the cerebellar tissue in the Ethanol group, reduced expression of them in the Ethanol+caffeine groupو elevated levels of miR-126-3p and miR-132-3p and decreased levels of miR-155-5p in plasma as a result of caffeine exposure |
| Hill, 2019 [18] | Human | NA | NA | Third-generation offspring (F3) with and without a family background of alcoholdependence | Familial diathesis for alcohol dependence confers changes in methylation of the DRD2 gene that have implications for the reduction of volumes of the fusiform, insula, and temporal regions in the left hemisphere, structures involved in social cognition. Strong collinearity between prenatal use of substances and familial risk for alcohol dependence; it is unlikely that these would have been responsible for the reduction in volume seen in these regions. |
| Chang, 2019 [104] | Mice/Male | For 70 days | Mating after the final exposure | Male and female/ F1 | Fetal growth restriction, decreased placental efficiency, abnormalities in cholesterol trafficking, sex-specific alterations in the genetic pathways regulating hepatic fibrosis, and disruptions in the regulation of imprinted genes (decreased expression of Gnas and Grb10 within the placentas of male offspring and Sgce in the female offspring of EtOH-exposed males, significant reductions in transcripts encoding Cdkn1c, Dcn, and Gnas in male offspring and Meg3 in female offspring in the fetal liver) |
| Beeler, 2019 [89] | B6 mice/Male | For 12 weeks, every other day two bottle choice | Mating 48 hours after the last exposure | Male and female/F1 | OFT: distance did not change. In male: duration time in center increased. No change in total resting time, increased resting time in center and decreased in periphery EPM: no change in parameters DID: reduced ethanol binge-like drinking in male, but not female offspring No preference for sweet and bitter taste |
| Asimes, 2018 [80] | Rat/Male and female | PND 37-44 and PND 67-74 | Mating 24 hours after the last treatment | Male and female/F1 | Hypothalamic GnRH mRNA decreased (male and female)Circulating LH: decreased only in male Circulating testosterone decreased in male and female Tetis: No change in gross morphology or number of elongated spermatids Corticosterone decreased in males but not females, with no change in GR, CRF, AVP Both parents exposed to ethanol 24 hours before mating resulted in significantly lower pup body weights during adolescence and adulthood (PND 37-44 and PND 67-74) Small body weight, fewer play behavior |
| Asimes, 2017 [101] | Rat/Male and female | binge-ethanol exposure paradigm during early (postnatal day 37-44) and late (67-74) adolescent development | Mating 24 hours after the last ethanol dose | Male and female/F1 | Differential DNA methylation patterns in the hypothalamus in offsprings of alcohol-exposed parents DMCs were distinct between offspring depending on which parent was exposed to ethanol. Novel DMCs in offspring with both parents exposed to ethanol DMCs mostly in intergenic, intronic and coding functional regions, no direct correlation between DMCs and gene mRNA expression No change in sex ratio and litter size |
| Rompala, 2016 [94] | C57 mice/Male | 5 weeks of exposure (inhalation) | Mating after the final exposure | Male and female/F1 | Decreased corticosterone secretion after restrain stress only in male Increased fluid intake in response to restrain stress (only in male) CPP: No change in ethanol preference during chronic variable stress No change in bodyweight Polydipsia induced by stress was not observed in male offspring |
| Jabbar, 2016 [74] | Rat/Female | For 4 weeks | Mating 3 weeks after the last treatment | Male and female/F1 | Plasma corticosterone increased in male, ACTH no change After LPS: plasma corticosterone and ACTH increased in both male and female Hypothalamus: CRH increased, β-endorphin decreased, AVP no change CRF mRNA increased, POMC mRNA increasedHippocampus and amygdala: Crfr1 decreased normalize after administration of methylation-blocker Maternal ethanol exposure three weeks before mating did not affect the behavior of mothers in terms of licking, grooming, and arched-back nursing Maternal exposure to alcohol resulted in a longer pregnancy duration and decreased body weight Maternal exposure to alcohol did not affect mothers' care Anxiety: increased in males but not females (EPM and OFT), normalized after administration of methylation-blocker |
| Ceccanti, 2016 [78] | CD1 mice/Male | For 60 days | Mating 2 months after the last exposure | Male/F1 | PFC: BDNF increased, P75 and NGF decreased, Trk, DAT, D1 and D2DR no change Hippocampus: TrkA increased, BDNF, NGF, TrkB, P75, D1D2DR, DAT no change Hypothalamus: No change Olfactory bulb: DAT increased, other no change The number of pups, sex ratio, and mortality rate of pups were not affected by paternal alcohol exposure Ethanol CPP: condition with low dose (0.5) but not high dose (1.5) Bodyweight decreased, mortality rate and number of pups did not change. |
| Liang, 2014 [66] | Mice/Male | Every 2 days for 4 weeks | Mating after one month | Male and female/F1 | F0: sperms: increased methylation of H19, Peg3 F1 cerebral cortex: No change in methylation of H19, Peg 3, Ndn, decreased methylation of Snrpn Mortality rate increased among the offspring of ethanol-exposed sires The offspring's body weight was not affected by paternal alcohol exposure before gestation Weight and sex ratio no change EPM: Increased anxiety only in females Depressive-like behavior: increased only in females MWM: memory and learning impaired in males and females |
| Kim, 2014 [97] | ICR mice/Male | For 7 weeks (at 5 weeks of age) | Mating 1 week after the final treatment | Male/F1 | DAT decreased in mPFC and striatum DNMT1 and MeCP2 decreased in mPFC and striatumMethylation in DAT promoter increased The offspring's body weight was not affected by paternal alcohol exposure before gestation OFT: increased locomotor activity Electro-foot shock aversive water drinking: increased impulsivity Attention/spontaneous alternation behavior in Y-maze test increased (ADHD-like behavior) |
| Finegersh, 2014 [86] | C57 mice/Male | 5 weeks of exposure (inhalation) | Mating after the final exposure | Male and female/ F1 | F0: Decreased DNA methylation at the BDNF exon Ixa F1 (VTA): Decreased methylation at the bdnf exon IXa, no change in bdnf exon IV, dlk-1 F1 VTA: Increased BDNF expression in exon IX only in male offspring mPFC: no change |
| Finegersh, 2014 [65] | C57 mice/Male | 5 weeks of exposure (inhalation) | Mating after the final exposure | Male and female/F1 | TBC test: decrease ethanol preference in male-but, not femalesEPM: no change in basal anxiety-like behavior The increased anxiolytic effect of ethanol only in male OFT: no change accelerating rotarod: no change Ethanol clearance: no change |
| Przybycien-Szymanska, 2014 [103] | Rat/ Male and female | PND 37-44 | Mating on PND 75 | Male and female/ F1 | Genome-wide microarray analysis: changes in gene expression involved in neurogenesis and synaptic plasticity in the hypothalamus on PND 7 FGF 13 decreased, BMP1 increased in males, RELN increased in females, SERPINI1 increased in males, PAK3 decreased, IGF2R increased in males, VAMP3 increased in males, GnRH increased in males, and ApoE decreased. |
| Knezovich, 2012 [64] | Mice/Male | For 5 weeks | Mating at week 6 | Male/ F1 | Two paternally methylated ICRs (H19 and Rasgrf1): decreased in somatic DNA |
| Meek, 2007 [79] | Mice/Male | A single dose | Mating 12-24 hours after exposure | Male and female/F1 | Male mice exposed to ethanol 12 hours before mating had a higher mortality rate and fewer pups Delay in: surface righting, clinging, the tail-pull reflex, rotation, linear movement, climbing an inclined surface, stretched attention, flatback, freezing, and defensive burying Increased aggressive behavior Decreased defensive/fearful behaviors |
| Ledig, 1998 [84] | Rat/Male | For 3 months | Mating 2 months after the exposure | Male and female/F1 | Male mice exposed to acute ethanol 12 hours before mating exhibited a reduced total number of pups In the cultured glia: protein levels did not change during 3 weeks after culture but decreased in 4th week Enolase increased in the first week, with no change until 4th week Glutamine synthase decreased in the first and second weeks SOD decreased in the second week but did not change on remained weeks OFT: locomotion increased Novelty-seeking: locomotion and rearing increased, the latency decreased Light-dark test: time in light increased |
| Wozniak, 1991 [85] | Rat/Male | PND 30, For 39 days | Mating 2 weeks after last treatment | Male and female/F1 | adolescent exposure to ethanol in male rats (2 weeks before mating) resulted in no alteration in developmental landmarks and body weight No change in the NOR test Impaired spatial learning performance in males, but not female in T-maze and RAM |
| Abel, 1988 [82] | Randall, 1982 | Mice/Male | For 5 weeks | Mating 24 hours after the last exposure | Male F1: Decreased serum testosterone level The offspring's body weight was not affected by paternal alcohol exposure before gestation PAT: Memory impairment in offspring following paternal ethanol exposure Decreased required trials to learn a passive avoidance task Longer latencies to reach the choice point in a T-maze paternal ethanol exposure increased locomotion, decreased latency in the novelty-seeking test, and increased time spent in the light compartment during the light-dark test. |
| Randall, 1982 [76] | Mice/Male | For 5 weeks | Mating 24 hours after the last exposure | Male and female/F1 | The number of pups, sex ratio, and mortality rate of pups were not affected by paternal alcohol exposure No change in weight gain, sex ratio, and malformation |
ADHD: Attention-Deficit / Hyperactivity Disorder; AVP: Arginine vasopressin; BDNF: Lower brain-derived neurotrophic factor; ; Crfr1: Corticotropin-releasing factor receptor-1; CPP: Conditioned place preference; DAT: Dopamine transporter; DID: Drinking in the dark; DMC: Differentially methylated cytosine; CRF: Corticotropin-releasing factor; CRH: Corticotropin-releasing hormone; EPM: Elevated plus maze; FASD: Fetal alcohol spectrum disorders;; GnRH: Gonadotropin hormone-releasing hormone; GR: Glucocorticoid receptor; LH: Luteinizing hormone; mPFC: medial Prefrontal cortex; NA: Not applicable; NAc: Nucleus Accumbens; NOR: Novel object recognition; PAT: Passive avoidance test; PatEE: Parental ethanol exposure; PC:EtOH: periconceptional ethanol; POMC: Pro-opiomelanocortin; PND: Post-natal day; MWM: Morris water maze; OFT: Open filed test; RAM: Radial arm maze; TBC: Two bottle choice;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



