
Submitted:
03 July 2023
Posted:
04 July 2023
You are already at the latest version
Abstract
In vineyards, Botryosphaeria dieback (BD) occurs in young and old plants. In the field, the prevalence and severity of the disease increase proportionally with the age of the vineyards. Among the pathogens that cause (BD), Diplodia seriata is the most prevalent species in Chile and other countries with a Mediterranean climate. To date, no information is available on the sus-ceptibility of adult wood to infection by this pathogen since most of the pathogenicity tests have been carried out on 1- or 2-year-old shoots or detached canes. Therefore, a pathogenicity test was carried out on plants under field conditions, with inoculations in plants, on one-year-old shoots, and 2- and 10-year-old wood in grapevine cv. Cabernet Sauvignon. A pathogenicity test was carried out with two isolates of D. seriata selected in an assay in detached canes that were 2-year-old. The results on plants showed that D. seriata was significantly more aggressive on 10-year-old than on one- or two-year-old tissue. These results were compared with the lesions ob-tained from two-year-old canes. The results of this work are consistent with the damage ob-served in the field, highlight the importance of identifying each pathogenic species that causes (BD) and contribute to the knowledge of the epidemiology of this disease in Mediterranean cli-mates.
Keywords:
Keywords: Plant–pathogen interactions
; Grape trunk diseases
; GTD
; Botryosphaeria dieback
; Pathogenicity
; Tissue age
; Virulence
; Vitis vinifera
; Cabernet Sauvignon.
1. Introduction
Grapevine trunk diseases (GTDs) are the leading cause of vineyard deterioration, causing a decrease in the viability of vineyards, an increase in production costs, and major economic losses in the wine industry [1,2,3,4,5,6]. The three main GTDs are esca disease, Eutypa dieback, and (BD) (BD), which generally attack the structural parts of plants [2,6,7,8,9,10]. (BD) is present on all continents where wine or table grapes are grown [2,5,7,11,12,13,14,15,16]. Regarding the pathogens associated with BD, 26 species have been identified [10], mainly within the genera Botryosphaeria, Diplodia, Lasiodiplodia, and Neofusicoccum [7,10,11,17]. Some genera are more common in different climates, for example, Diplodia in temperate regions or those with cold winters and Lasiodiplodia in tropical and subtropical areas [18,19,20,21,22,23,24]. Diplodia seriata is the most prevalent species in vineyards in Central Chile [5,15,22] as well as in other parts of the world, such as New South Wales [25,26,27], Western Australia [28], California [29] and South Australia [26].
(BD) causes essential losses in the production of grapes for wine, with a decrease from 30 to more than 50% of production depending on the severity of the disease [5]. The loss of production is associated with the damage it causes to the vines due to the death of the wood where fruit develops, such as cankers in the wood, dead spurs, dead arms, and brown discolorations in the vascular area [2,5,7,15,16,30,31,32,33,34]. In the field, the prevalence and severity of (BD) increase proportionally with the age of the vineyards [5,6,7,10,15,33]. However, many available pathogenicity studies have been carried out on one-year-old shoots or two-year-old canes. Another critical issue to consider is the susceptibility and importance of some varieties for the wine industry. In this case, Cabernet Sauvignon is the most planted variety in Chile, and it is appreciated for good quality wines [35].
For this reason, in this study, a pathogenicity test was carried out with five Chilean isolates of D. seriata in 2-year-old detached canes of Vitis vinifera cv. Cabernet Sauvignon, and with the two most representative isolates, a pathogenicity test was carried out on vine plants under field conditions, with inoculations in 1-year-old shoots, 2-year-old wood, and 10-year-old wood of grapevine. In this study, we observed that D. seriata is more aggressive in old tissue of grapevine cv. Cabernet Sauvignon than in young tissue, which contributes to the knowledge of the epidemiology of this disease in Mediterranean climates.
2. Results
2.1. Pathogenicity Tests on Detached Canes (Nonactivated Isolates)
Ninety days after inoculation with D. seriata isolates PUCV 1472, PUCV 1549, PUCV 2120, PUCV 2142, and PUCV 2183, the vascular lesions and canker lengths on detached canes 2 years old were measured.
2.1.1. Damage Assessment and Recovery of Pathogens
The isolates PUCV 2120 and PUCV 2183 produced vascular discoloration, which extended up and down from the point of inoculation, with vascular lesions presenting variable lengths of 12.6 and 6.9 cm, respectively (Figure 1a). The isolates PUCV 1549 and PUCV 2142 produced dark brown canker lesions on inoculated canes, with lengths of 1.0 and 1.1 cm, respectively (Figure 1b). The isolate PUCV 1472 produced neither canker nor vascular discoloration (in this assay, in the assay with the activated isolate, there was some canker and vascular discoloration). The controls (noninoculated canes) did not present damage (no canker and no vascular discoloration). From the lesions produced by the isolates, the pathogen was recovered, and D. seriata was morphologically identified.
2.2. Pathogenicity Tests on Berry Grapes
Fifteen days after inoculation with D. seriata isolates PUCV 1472, PUCV 1549, PUCV 2120, PUCV 2142, and PUCV 2183 on berry grape cv. Red Globe, the diameter of lesions was measured.
2.2.1. Damage Assessment and Recovery of Pathogens
All isolates caused circular soft rot around the point of inoculation on grape berries, with lesion diameters between 2.1 and 2.9 cm and the presence of grayish-white mycelium around the point of inoculation (Figure 2a). The inoculated berries were cut longitudinally, presenting rottenness on the pomace as well. From the zone of advancing lesions, the pathogen was recovered, and D. seriata was morphologically identified. The controls (noninoculated berry grapes) did not present damage (Figure 2b), thus corroborating that all isolates of D. seriata were pathogenic on berry grape cv. Red Globe.
2.3. Pathogenicity Tests in Detached Canes (Activated Isolates)
Ninety days after inoculation with isolates of D. seriata PUCV1472, PUCV1549, PUCV 2120, PUCV 2142 and PUCV 2183 (previously inoculated in berries) in detached 2-year-old canes, vascular and canker lesion lengths were measured.
2.3.1. Damage Assessment and Recovery of Pathogens
All isolates produced dark brown canker lesions on inoculated canes, with lengths between 1.4 and 3.0 cm (Figure 1b,c), and isolates PUCV 2120 and PUCV 2183 causing the longest cankers. Isolates PUCV 1472, PUCV 2120, and PUCV 2183 also produced vascular discoloration, which was expressed up and down from the point of inoculation. The vascular lesions presented variable lengths between 2.6 and 13.3 cm (Figure 1a), and as with the cankers, the isolates PUCV 2120 and PUCV 2183 were the ones that caused the most significant lesions. The controls (noninoculated canes) did not present damage, no canker or vascular discoloration, thus corroborating that all isolates of D. seriata were pathogenic. From the lesions produced by the isolates, the pathogen was recovered, and D. seriata was morphologically identified. With the results obtained in both detached cane assays, the isolates PUCV 2120 and PUCV 2183 were selected for the field trial.
2.4. Pathogenicity Trial in the Field
Five months after inoculation with D. seriata isolates PUCV 2120 and PUCV 2183 on tissues of three different ages (one-year-old, two-year-old and 10-year-old wood) of plants cv. Cabernet Sauvignon, the length of vascular lesions was measured.
2.4.1. Damage Assessment and Pathogen Recovery
Both isolates produced significant vascular lesions compared to controls (T0 and T00), as shown in Figure 3. In young tissue (one-year-old shoots and two-year-old wood), vascular lesions were significantly smaller than those in older tissue (10-year-old wood), Figure 3 and Figure 4a. The vascular lesions developed in the 10-year-old wood were 4.2 and 4.3 cm for PUCV 2183 and PUCV 2120 isolates, respectively, with no significant differences between the aggressiveness of D. seriata isolates evaluated. In young tissue, vascular lesions caused by D. seriata isolates ranged from 1.9-2.7 cm, with no differences between age of tissue (one or two years old) or isolate (PUCV 2120 or PUCV 2183). Only from inoculated plants was the pathogen recovered, both from young and old tissue. This result shows that the vines used in this trial were healthy and that the lesions presented are attributable to inoculations with D. seriata isolates. Inoculated plants did not show foliar symptoms (Figure 4b,c).
2.5. Maximum Parsimony Analysis
The concatenated ITS and BT phylogenetic analysis included 66 D. seriata isolates, all obtained from V. vinifera, showing that the five isolates used in this assay are in different clades within the species, regardless of virulence or origin. The analyses contained 802 nucleotides, of which 702 were constant and 22 were parsimony informative. The analyses yielded tree #1 out of the 10 most parsimonious trees, with tree length = 53; consistency index = 0.960000, retention index = 0.995781, and composite index = 0.976992 (0.955949) for all sites and parsimony-informative sites (in parentheses). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The MP tree was obtained using the tree-bisection-regrafting (TBR) algorithm with search level 1, in which the initial trees were obtained by the random addition of sequences (10 replicates). This analysis involved 79 nucleotide sequences. There were a total of 802 positions in the final dataset.
Figure 5.
Phylogenetic tree of isolates of D. seriata based on maximum parsimony (MP) analysis of two loci (ITS and β-tubulin). Chilean isolates used in this work with PUCV code. MP bootstrap values are shown above the branches. Spensecermartinsia viticola isolates PD258 (Inderbitzin et al., 2010) and UCP105 (Adesemoye and Eskalen, 2011) were the outgroups used. Values were obtained with Mega X software.
Figure 5.
Phylogenetic tree of isolates of D. seriata based on maximum parsimony (MP) analysis of two loci (ITS and β-tubulin). Chilean isolates used in this work with PUCV code. MP bootstrap values are shown above the branches. Spensecermartinsia viticola isolates PD258 (Inderbitzin et al., 2010) and UCP105 (Adesemoye and Eskalen, 2011) were the outgroups used. Values were obtained with Mega X software.

3. Discussion
This study reports for the first time the comparison of the pathogenicity of the fungus Diplodia seriata on tissues of different ages in cv. Cabernet Sauvignon plants and demonstrates its aggressiveness in old tissue under field conditions. Diplodia seriata is one of the most prevalent pathogens isolated from grapevines with symptoms of (BD), mainly in wine or table grape-producing areas with Mediterranean climates [5,6,15,17,23,25,26,28,36]. Of the isolates obtained from diseased tissue, D. seriata is the most widely distributed in studies carried out from more than 20 years to the present. In a sampling of 11 sites distributed in different vineyards in the Hunter Valley of New South Wales (NSW, Australia), D. seriata was isolated in 90% of them [25]. In California (USA), a survey on different varieties of grapevine (table grapes and wine) indicated that D. seriata had the broadest geographical distribution within the Botryosphaeriaceae found between the wine-growing areas located between Mendocino and Kem [12]. From collections in the wine-growing regions of Western Australia and the Hunter Valley and Mudgee of NSW [17,28], D. seriata accounted for more than 50% of the total number of isolates collected and present in all the regions surveyed. Pitt et al. [26] reported that Diplodia was the most predominant genus. The species D. seriata represented almost 80% of Botryosphaeriaceae isolates collected during the survey in NSW and South Australia. In Catalonia (Spain), the results of GTD-associated fungal isolates showed that D. seriata was isolated in up to 68.4% of the samples in diseased plants, being the most prevalent species isolated [36]. In central Chile, with a Mediterranean climate, results of studies carried out on table grapes and wine have been consistent with those of the above studies. Morales et al. [15] reported an 83.3% prevalence of D. seriata in table grape plants affected by (BD). Additionally, in Chile, Díaz et al. [16] reported that D. seriata (unlike other pathogens found) was present in all the valleys studied in a national survey of Chilean decayed vineyards. Valencia et al. [22] recorded the dissemination of Botryosphaeriaceae conidia, and D. seriata was the main species detected in vineyards of cv. Cabernet Sauvignon. Specifically, in cv. Cabernet Sauvignon, a sampling carried out in the main producing area of this wine variety in Chile (O’Higgins and Maule regions), reported that in 2010, D. seriata was the most prevalent species isolated from symptomatic vines (68%). Eight years later (2018), when sampling the same area, D. seriata continued to be the most prevalent species, increasing its prevalence (91%), and the overall yield losses associated with (BD) were 39% in 2010 and 46% in 2018 [5]. Based on the works mentioned above, most isolates of D. seriata were obtained from symptomatic old wood tissues. However, pathogenicity tests for D. seriata are mainly carried out on young tissue (shoots, one- or two-year-old canes, rotted plants, or young tissue on old plants). In these cases, D. seriata virulence was weak or less significant than that of other Botryosphaeriaceae species, such as Lasiodiplodia theobromae, L. viticola, Neofusicoccum parvum or Diplodia mutila [8,33,37,38], or when compared with other causal agents of GTDs, such as Eutypa lata [39]. This difference in virulence could be due to several factors, including the differential susceptibility of the vine varieties, the conditions, age and tissue type of the host plant, the inoculation method used, the geographic origin of the isolates, and the different incubation periods of the experiments [7,31,32,40,41].
In our work, the pathogenicity of D. seriata isolates was verified in plants, with inoculations on young tissue (one- or two-year-old tissue age) and on old tissue (10-year-old wood) in vine plants. The vascular lesions developed by D. seriata on old tissue (10-year-old wood) were more than twice as long as those in the young tissue. Our results coincide with those described by Morales et al. [15] in old table grape plants in a commercial vineyard. When inoculating shoots <one year, canes >five years old, and mature arms in a 25-year-old vines in commercial vineyards, D. seriata increased the damage by more than 40% in mature arms with respect to the other two ages evaluated, contrary to D. mutila. Additionally, our results are consistent with those reported in naturally affected vineyards. In the field, symptoms of (BD) increase with the age of the vineyard, both in incidence and severity [5,12,15,30]. The extension of the advance of this pathogen in the wood can be explained by the microenvironmental theory [42], where pathogen reaction zones can alone explain the progression of fungi within tree trunks, as fungi follow dehydrated, oxygen-rich zones that occur along the reaction zones near the wounds. The high moisture content and the associated aeration restriction limit the activity of mycelial fungi in felled wood. Additionally, the most significant damage in old tissue can be explained concerning the plant microbiome, which evolves over time and with plant tissues. Microbiome diversity is higher in young vines [1,43,44,45] and in visually healthy tissues than in diseased tissues [46].
In contrast, studies with Neofusicoccum on walnut plants show that the relationship between age and damage is inverse. One-year-old shoots were significantly more sensitive to N. parvum than two-year-old to four-year-old shoots wood [47]. Therefore, correctly identifying the causal agent and studying the interaction with the host is essential.
In our work, when comparing the results of pathogenicity tests on two-year-old tissue, detached cuttings or canes of plants (attached), our study shows a difference in the severity of D. seriata damage. This may be due to the type of inoculum, mycelium (in the first case) or conidia (in the second case), and/or the state of the tissue (detached or attacched). The vascular lesion was much more aggressive in detached cuttings inoculated with mycelium, reaching up to 12 cm, than in canes on plants, where the lesions were up to 5 cm tissue. In the literature, it is described that during the infection of the vine, a series of molecules are activated that provide protection against the spread of the pathogen [46]. The first active defense is the formation of tylosis within the vessels. Responses to GTD fungi are thought to occur due to PAMP-activated immunity. Phytoalexins, compounds of the phenylpropanoid pathway such as resveratrol or viniferin, are found in higher amounts in tissues after GTD fungal infection [48,49,50,51,52]. Phytoalexins can inhibit the growth and colonization of fungi, block some metabolites produced by fungi during infection, or interfere with oxidation‒reduction reactions [52]. In addition to the active defense implemented by the vines, the anatomy of the wood could also be essential. The density of parenchymal rays and their arrangement in space could enhance active responses to infection [54] and the diameter of the vessels could also be a factor [55,56,57]. Contrary, Amponsah et al., [58], showed a faster rate of germination of N. luteum conidia on attached shoots and leaves wounded than on detached and unwounded leaf surfaces, this may be due to the ability of Neofusicoccum species to attack on spring and summer time associated to shoots and leaves damage, unlike D. seriata which mainly affects woody tissue. Regarding the type of inoculum, our results are similar to those reported by Moral et al. [59]. They found that inoculation with a suspension of Botryosphaeria dothidea conidia resulted in more severe disease than inoculation with a plug of mycelium in olive fruits, probably due to the presence of water in the inoculum. In addition, in other Botryosphaeriaceae species, B. dothidea in this case, the physiological characterization of the isolates that cause Dalmatian disease showed that the optimal temperature was 26°C for mycelium growth and 30°C for germination of conidia. Both factors can influence the aggressiveness of the symptoms, which should be studied for D. seriata.
The results obtained in detached canes (two years old) show that, in general, all the isolates increased their pathogenicity after activation on grape berry cv. Red Globe, being higher in the oldest isolates. For example, with nonactivated isolates, the PUCV 1472 isolate did not cause damage to the cuttings (no vascular lesions and no canker), and in the assay with activated isolates (after inoculation on grape berries), vascular lesions and canker developed. The PUCV 1549 isolate increased the length of the canker by 67% after inoculation in grapes (Figure 1a,b). Concerning this point, studies carried out with postharvest pathogens (i.e., Botrytis cinerea, Colletotrichum, and Alternaria) can explain these results. In this sense, pathogen enzymes are not all constitutive and may require induction by proper substrates [41]. Reveglia et al. [60] demonstrated that the Botryosphaeriaceae family associated with (BD) in Australian grapevine leaves generates secondary metabolites. Martos et al. [61] demonstrated that D. seriata can produce secondary metabolites with phytotoxic properties, and it is possible that it has other mechanisms besides mycelial growth in the host that could be related to its virulence and that this phytotoxicity was strictly dependent on acidic pH substrates. This could explain the lower virulence variability of D. seriata isolates when inoculated on fruits rather than on canes, although this requires further study. In addition, according to the phylogenetic tree obtained, the Diplodia seriata isolates analyzed, all isolated from Vitis vinifera, presented greater variability in the ITS-BT segment with the available D. mutila isolates (also from Vitis vinifera), requiring further analysis.
4. Materials and Methods
4.1. Chemicals, Reagents, and Culture Media
The reagents and culture media used were sodium hypochlorite (SMF Ltda., Chile), agar papa dextrose acidulated (APDA): 20 g L-1 granulated agar (Algas Marinas S.A., Spain), 20 g L-1 mashed potatoes (Nestlé S.A., Switzerland), 22 g L-1 glucose (Vimaroni S.A., Chile), 7 drops of citric acid (Merck S.A., Germany) and 1 liter of sterile distilled water (SDW). Sterile distilled water was obtained by using a distiller (Pobel, Spain) and autoclave (Zonkia, China). The parafilm used was obtained from Bemis, USA.
4.2. Fungal Isolates
Five isolates of D. seriata were used (Figure 6 and Table 1). The isolates belong to the fungal collection of the Phytopathology Laboratory, School of Agronomy, PUCV. The isolates PUCV 1472, PUCV 1549, PUCV 2120, PUCV 2142, and PUCV 2183 were previously obtained from plants with cankers and vascular lesions from commercial vineyards in the wine-growing zone of central Chile, all with a history of BD that was previously molecularly identified [5]. The isolates were classified based on a phylogenetic analysis of the concatenated ITS-BT. A multilocus phylogenetic analysis was performed using maximum parsimony (MP) in MEGAX (Kumar et al., 2018). Bootstrap values were calculated using 1,000 replicates yielding the MP tree using Tree Bisection and Reconnection algorithms. The tree was rooted with Neofusicoccum parvum strain CBS 110302, and other reference isolates of different isolates of D. seriata and D. mutila were used (Table S1). The retention index, rescaled consistency index, and homoplasy index were calculated using MEGA X.
4.3. Pathogenicity Tests on Detached Canes (Nonactivated Isolates and Activated Isolates)
The inoculations were performed on healthy detached 2-year-old canes (15 cm in length) of Vitis vinifera cv. Cabernet Sauvignon. The canes were obtained in the winter of 2020 from an experimental vineyard of the Experimental Station La Palma of Pontifical Catholic University of Valparaíso (PUCV) and maintained at 5°C for three months before use [62].
Nonactivated isolates: detached 2-years-old canes were disinfected (1% sodium hypochlorite for 5 min and 95% ethanol for 30 s) and then triple washed in SDW, according to Morales et al., [15], and an inoculum of 5-mm-diameter mycelial plugs from a 6-day-old on APDA culture was inoculated using an oblique cut made in the bark with a sterile scalpel at the middle of the canes. The inoculated area was covered with parafilm [15,47]. Three detached 2-year-old canes were used for each isolate, and three were used for the control (noninoculated canes, only APDA plug). The assay was performed in duplicate. The canes were incubated for 90 days in an individual humid chamber at 23°C and were distributed according to a completely randomized design (CRD).
Activated isolates: The pathogenicity assay was repeated with the five isolates previously activated in grape berry cv. Red Globe plants. After recovering the isolates from infected berries, we proceeded to repeat what was described above.
4.3.1. Damage Assessment and Pathogen Recovery
After three months of incubation, vascular lesions and canker lengths of the canes were measured up and down from the point of inoculation. For data analysis, the total length of the lesions was considered. Analysis of variance (ANOVA) and Tukey’s test were used for data analysis. Tissue samples were taken from the zone of advancement of the lesions, disinfected with 1% sodium hypochlorite for 5 s, triple washed in SWD, and cultivated in APDA.
4.4. Pathogenicity Tests on Berry Grapes
The inoculations were performed on fresh berry grape cv. Red Globe of Vitis vinifera. Berry grapes were obtained in the summer of 2021 from the experimental vineyards of Estación La Palma of Universidad Católica de Valparaíso (PUCV) and immediately used for the test.
4.4.1. Fungal Isolates and Inoculation
Five isolates of D. seriata were used, the same isolates as in the previous section (Table 1). For the inoculation, berry grapes were disinfected with 1% sodium hypochlorite for 30 seg and then triple-washed in sterile distilled water. A 5-mm-diameter inoculum of a 5-mm-diameter mycelial plug from a 6-day-old grown in APDA culture was inoculated in the middle of the berry through a previously made wound with a sterile needle. The inoculated area was covered with parafilm. Five berry grapes were used for each isolate, and five were used for the control (noninoculated berry grape, only APDA plug). The berry grapes were incubated for 15 days in an individual humid chamber at 23°C and were distributed according to a completely randomized design (CRD). The pathogenicity assay in detached canes was repeated with the five isolates previously activated in grape berries.
4.4.2. Damage Assessment and Pathogen Recovery
After incubation of the berry grapes, fruit rot was measured around the point of inoculation. The diameter of the lesions was considered. Analysis of variance (ANOVA) and Tukey’s test were used for data analysis. Tissue samples were taken from the zone of advancement of the lesions, disinfected with sodium hypochlorite for 5 seg, triple washed in ADE, and cultivated in APDA. The plates were incubated for 7 d at 24°C.
4.5. Pathogenicity Trial in the Field
The experiment was carried out on 10-year-old not grafted cv. Cabernet Sauvignon plants in an experimental vineyard at Experimental Station La Palma of Pontifical Catholic University of Valparaíso (PUCV). The selected vineyard did not present previous GTD symptoms, and it was trained in the bilateral cordon system, with the spur pruned to 4 buds. The inoculations were performed on tissues of three different ages: one-year-old shoots, two-year-old wood, and 10-year-old wood, with each inoculation/age on a different plant (Figure 7).
4.5.1. Fungal Isolates and Inoculation
Two isolates of D. seriata, PUCV 2120 and PUCV 2183 (Table 1), previously selected in pathogenicity tests on two-year-old detached canes, were used for the field experiment. The inoculation was carried out in July 2021, with 50 µl of inoculum at 1*104 spores*ml-1 on fresh cuts on one-year-old shoots, two-year-old wood, and 10-year-old wood (arm) (Figure 7). The cuts in one-year-old shoots and two-year-old wood were made with pruning shears and in 10-year-old wood with a chainsaw (Stihl, Germany). Spore suspensions were prepared according to Úrbez-Torres et al. [38] and Larach et al. [5]: 5-day-old mycelial plugs of each isolate were placed in Petri dishes containing 2% agar water and needle-autoclaved pine. Plates were incubated in a chamber at room temperature (19–21°C) under near-ultraviolet light (λ = 320 nm) until pycnidium production and conidium development. Mature pycnidia were crushed in sterile distilled water, and the solution obtained was filtered through a sterilized cheesecloth. Five plants were used for each isolate/tissue age, with one inoculation per plant. Five absolute control plants (without cutting or inoculation, T00) and five control plants (cut plus sterile distilled water, T0) were used. Plants were distributed according to CRD.
4.5.2. Damage Assessment and Pathogen Recovery
Five months after inoculation of tissue, vascular lesions and canker length were recorded. For this, the damage produced in each tissue/age was evaluated in the field, and the plant material was cut to measure the lesions. The assay was performed in duplicate. Analysis of variance of two factors (ANOVA) and Tukey’s test were used for data analysis. To recover the fungus, tissue samples were taken from the zone of advancement of the lesions, disinfected, and cultivated in APDA.
5. Conclusions
In this study, we observed that D. seriata is more aggressive in old tissue of grapevines plants cv. Cabernet Sauvignon than in young tissue, verifying the pathogenicity of D. seriata and its role as a causal agent of the disease (BD). These results contribute to the knowledge of the epidemiology of this disease in Mediterranean climates. Additionally, the pathogenicity tests carried out to select the isolates for pathogenicity testing in the field showed different results for vegetative tissue or berry grapes, highlighting the importance of the type of tissue in the pathogenicity or virulence of the D. seriata pathogen. Finally, the damage observed in cuttings is probably due to the combination of the type of inoculum (mycelium or conidia) and the state of the plant material (cut or not cut), which should be studied in depth to determine the extent of its role in the severity of the damage.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Investigation and writing, A.L. ; Original draft preparation, A.L., D.C.-N. ; Formal analysis, X.B., M.S., P.V.-C., E.S. ; Investigation, P.S., A. S, N.R., A.L. ; Statistical analysis: A. L E.S., P.V.-C. ; Writing—review & editing, P.V.-C. and M.S. ; Supervision and Writing—review & editing, X.B.
Funding
This research was funded by FONDECYT REGULAR 1211094 (ANID, Chile) and the operating expenses of Alejandra Larach’s doctoral scholarship, grant number 21180920.
Data Availability Statement
Not applicable here.
Acknowledgments
The authors thank Lorena Tapia for her support in laboratory activities and Iván Cortés for his collaboration in the field maintenance trial.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.; Da Costa, J.; Guerin-Dubrana, L.; Rey, P. Overview of grapevine trunk diseases in France in the 2000s. Phytopathol Mediterr. 2013, 52(2), 262–275. [Google Scholar]
- Fontaine, F.; Pinto, C.; Vallet, J.; Clément, Ch.; Catarina, A.; Spagnolo, A. The effects of grapevine trunk diseases (GTDs) on vine physiology. Eur J Plant Pathol. 2016, 114, 707–721. [Google Scholar]
- Hillis, V.; Lubell, M.; Kaplan, J.; Doll, D.; Baumgartner, K. The role of pest control advisers in preventative management of grapevine trunk diseases. Phytopathol. 2016, 106(4), 339–47. [Google Scholar]
- Kaplan, J.; Travadon, R.; Cooper, M.; Hillis, V.; Lubell, M.; Baumgartner, K. Identifying Economic Hurdles to Early Adoption of Preventative Practices: The Case of Trunk Diseases in California Winegrape Vineyards. AgEcon Search 2016. [Google Scholar]
- Larach, A.; Torres, C.; Riquelme, N.; Valenzuela, M.; Salgado, E.; Seeger, M.; Besoain, X. Yield loss estimation and pathogen identification from (BD) in vineyards of Central Chile over two growing seasons. Phytopathol Mediterr. 2020, 59(3), 537–48. [Google Scholar]
- Kenfaoui, J.; Radouane, N.; Mennani, M.; Tahiri, A.;Ghadraoui, L.; Belabess, Z.; Fontaine, F.; El Hamss, H.; Amiri, S.; Lahlli, R.; Ait, Barka, E. A Panoramic View on Grapevine Trunk Diseases Threats: Case of Eutypa Dieback, (BD), and Esca Disease. J of Fungi. 2022, 8, 595. [Google Scholar]
- Úrbez-Torres, JR.; Gubler, WD. Susceptibility of grapevine pruning wounds to infection by Lasiodiplodia theobromae and Neofusicoccum parvum. Plant Pathol. 2011, 60(2), 261–70. [Google Scholar]
- Úrbez-Torres, JR.; Battany, M.; Bettiga, LJ.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R.; Verdegaal, P.; Gubler, W. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant Dis. 2010, 94(6), 717–24. [Google Scholar]
- Besoain, X. Grapevine Trunk Diseases (GTDs): Impact on Table Grapes and Wine Vineyards in Chile. In: Grapes and Wines - Advances in Production, Processing, Analysis and Valorization. InTech. 2018.
- Gramaje, D.; Urbez-Torres, JR.; Sosnowski, MR. Managing grapevine trunk diseases with respect to etiology and epidemiology: Current strategies and Future prospects. Plant Dis. 2018, 102(1), 12–39. [Google Scholar]
- Billones-Baaijens, R.; Savocchia, S. A review of Botryosphaeriaceae species associated with grapevine trunk diseases in Australia and New Zealand. Aust Plant Pathol. 2019, 48, 3–18. [Google Scholar]
- Rolshausen, P.; Baumgartner, K.; Travador, R.; Funjiyoshi, P.; Pouzoulet, J.; Wilcox, W. Identification of Eutypa spp. Causing Eutypa Dieback o Grapevine in Eastern North America. Plant Dis 2014, 483–491. [Google Scholar]
- Burruano, S.; Mondello, V.; Conigliario, G.; Alfonzo, A.; Spagnolo, A.; Mugnai, L. Grapevine decline in Italy caused by Lasiodiplodia theobromae. Phytopathol Mediterr. 2008, 47, 132–136. [Google Scholar]
- Reis, P.; Magnin-Robert, M.; Nascimento, T.; Spagnolo, A.; Abou-Mansour, E.; Fioretti, C.; Clément, Ch.; Rego, C.; Fontaine, F. Reproducing (BD) foliar symptoms in a simple model system. Plant Dis. 2016, 100(6), 1071–9. [Google Scholar] [PubMed]
- Morales, A.; Latore, B.; Piontelli, E.; Besoain, X. Botryosphaeriaceae Species affecting table grape vineyards in Chile and cultivar susceptibility. Cien Investig Agrar. 2012, 39(3), 445–445. [Google Scholar]
- Díaz, GA.; Auger, J.; Besoain, X.; Bordeu, E.; Latorre, B.A. Prevalence and pathogenicity of fungi associated with grapevine trunk diseases in Chilean vineyards. Cienc Investig Agrar. 2013, 40(2), 327–39. [Google Scholar]
- Savocchia, S.; Steel, CC.; Stodart, BJ.; Somers, A. Pathogenicity of Botryosphaeria species isolated from declining grapevines in sub-tropical regions of Eastern Australia. Vitis. 2007, 46. [Google Scholar]
- Burgess, T.; Wingfield, MJ. Quarantine is important in restricting the spread of exotic seed-borne tree pathogens in the southern hemisphere. International Foresty Review. 2002, 4, 56. [Google Scholar]
- Mohali, S.; Burgess, TI.; Wingfield, MJ. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. Path. 2005, 35, 385–396. [Google Scholar]
- Burgess, TI.; Barber, PA.; Mohali, S.; Pegg, G.; De Beer, W.; Wingfield, MJ. Three new Lasiodiplodia spp. from the tropics, recognized based on DNA sequence comparisons and morphology. Mycologia 2006, 98((3)), 423–435. [Google Scholar]
- Kuntzmann, P.; Villaume, S.; Larignon, P.; Bertsch, C. Esca, BDA and Eutypiosis: foliar symptoms, trunk lesions and fungi observed in diseased vinestocks in two vineyards in Alsace. Vitis. 2010, 49(2), 71–76. [Google Scholar]
- Valencia, D.; Torres, C.; Camps, R.; López, E.; Celis-Diez, JL.; Besoain, X. Dissemination of Botryosphaeriaceae conidia in vineyards in the semiarid Mediterranean climate of the Valparaíso Region of Chile. Phytopathol Mediterr. 2015, 54(2), 394–402. [Google Scholar]
- Burgess, T.; Pei, T.Y.; Garnas, J.; Edwards, J.; Scarlett, K.; Shuttleworth, L.; Daniel, R.; Dann, E.; Parkinson, L.; Dihn, Q.; Shivas, R.; Jami, F. Current status of the Botryosphaeriaceae in Australia. Aust Plant Pathology. 2018, 48, 35–44. [Google Scholar]
- Reis, P.; Gaspar, A.; Alves, A.; Fontaine, F.; Lourenco, I.; Saramago, J.; Mota, M.; Rego, C. Early season symptoms on stem, inflorescences and flowers of grapevine associated with Botryosphaeriaceae species. Plants. 2020, 11, 1–14. [Google Scholar]
- Castillo-Pando, M.; Somers, A.; Green, CD.; Priest, M.; Sriskanthades, M. Fungi associated with dieback of Semillon grapevines in the Hunter Valley of New South Wales. Aust Plant Pathology. 2001, 30, 59–63. [Google Scholar] [CrossRef]
- Pitt, WM.; Huang, R.; Steel, CC.; Savocchia, S. Identification, distribution and current taxonomy of Botryosphaeriaceae species associated with grapevine decline in New South Wales and South Australia. Aust J Grape Wine Res. 2010, 16(1), 258–71. [Google Scholar]
- Qiu, Y.; Steel, CC.; Ash, GJ.; Savocchia, S. Survey of Botryosphaeriaceae associated with grapevine decline in the Hunter Valley and Mudgee grape growing regions of New South Wales. Aust Plant Pathol. 2011, 40(1), 1–11. [Google Scholar]
- Taylor, A.; Hardy, G.E.S.J.; Wood, P.; Burgess, T. Identification and pathogenicity of Botryosphaeria species associated with grapevine decline in Western Australia. Aust Plant Pathol. 2005, 34(2), 187–95. [Google Scholar] [CrossRef]
- Úrbez-Torres, JR.; Leavitt, GM.; Voegel, TM.; Gubler, WD. Identification and distribution of Botryosphaeria spp. associated with grapevine cankers in California. Plant Dis. 2006, 90, 1490–503. [Google Scholar]
- Larignon, P.; Fulchic, R.; Cere, L.; Dubos, B. Observation on black arm in French vineyards. Phytopathol Mediterr. 2001, 40, 336–342. [Google Scholar]
- Van, Niekerk, JM.; Crous, PW.; Groenewald. JZ.; Fourie. PH.; Halleen, F. DNA Phylogeny, Morphology and Pathogenicity of Botryosphaeria species on Grapevines. Mycologia 2004, 96, 781–798.
- Baskarathevan, J.; Jaspers, M.; Jones, EE.; Ridgway, HJ. Incidence and distribution of botryosphaeriaceous species in New Zealand vineyards. Eur J Plant Pathol. 2012, 132, 549–60. [Google Scholar]
- Bertsch, C.; Ramírez-Suero, M.; Magnin-Robert, M.; Larignon, P.; Chong, J.; Abou-Mansour, E.; Spagnolo, A.; Clément, A.; Fontaine, F. Grapevine trunk diseases: Complex and still poorly understood. Plant Pathol. 2013, 62, 243–65. [Google Scholar]
- Spagnolo, A.; Larignon, P.; Magnin-Robert, M.; Hovasse, A.; Cilindre, C.; Van Dorsselaer, A.; Clément, C.; Schaeffer-Reiss, C.; Fontaine, F. Flowering as the most highly sensitive period of grapevine (Vitis vinifera L. cv Mourvédre) to the (BD) agents Neofusicoccum parvum and Diplodia seriata infection. Int J Mol Sci 2014, 15((6)), 9644–9669. [Google Scholar] [PubMed]
- ODEPA. 2022. Oficina de Estudios y Políticas Agrarias. Statistics Productive. (accessed: February 2022, from www.odepa.cl). 20 February.
- Luque J, Elena G, Garcia-Figueres F, Reyes J, Barrios G, Legorburu FJ. Natural infections of pruning wounds by fungal trunk pathogens in mature grapevines in Catalonia (Northeast Spain). Aust J Grape Wine Res. 2014, 20, 134–143.
- Bellée, A.; Comont, G.; Nivault, A.; Abou-Mansour, E.; Coppin, C.; Dufour, MC.; Corio-Costet, M. Life traits of four Botryosphaeriaceae species and molecular responses of different grapevine cultivars or hybrids. Plant Pathol. 2017, 66(5), 763–76. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Leavitt, G.M.; Guerrero, J.C.; Guevara., J.; Gubler, W.D. Identification and pathogenicity of Lasiodiplodia theobromae and Diplodia seriata, the causal agents of bot canker disease of Grapevines in Mexico. Plant Dis 2008, 92((4)), 519–529. [Google Scholar]
- Sosnowski, MR.; Ayres, MR.; McCarthy, MG.; Scott, ES. Winegrape cultivars (Vitis vinifera) vary in susceptibility to the grapevine trunk pathogens Eutypa lata and Diplodia seriata. Aust J Grape Wine. 2022, 28(1), 166–74. [Google Scholar]
- Van de Wouw, AP.; Howlett, BJ. Fungal pathogenicity genes in the age of “omics”. Mol Plant Pathol. 2011, 12(5), 507–14. [Google Scholar]
- Elena, G.; Garcia-Figueres, F.; Reigada, S.; Luque, J. Intraspecific variation in Diplodia seriata isolates occurring on grapevines in Spain. Plant Pathol. 2015, 64(3), 680–9. [Google Scholar]
- Boddy, L.; Rayner, A. Origins of decay in living deciduous trees: the role of moisture content and a re-appraisal of the expanded concept of tree decay. New Phytologist. 1983, 94(4), 623–41. [Google Scholar]
- Rey, P.; Lecomte, P.; Guérin-Dubrana, L.; Corio-Costel, F.; Fonteine, F.; Gomes, E.; Goutouly, JP.; Hofdtetter, V.; Legorburu, X.; De La Rocque, B. Maladies du bois de la vigne situation enjeux et perpective. Phytoma. 2011, 640. [Google Scholar]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar]
- Andreolli, M.; Lampis, S.; Zapparoli, G.; Angelini, E.; Vallini, G. Diversity of bacterial endophytes in 3- and 15-year-old grapevines of Vitis vinifera cv. Corvina and their potential for plant growth promotion and phytopathogen control. Microbiological. 2016, 183, 42–52. [Google Scholar]
- Claverie, M.; Notaro, M.; Fontaine, F.; Wery, J. Current knowledge on Grapevine Trunk Diseases with complex etiology: a systemic approach. Phytopathol Mediterr. 2020, 59, 29–53. [Google Scholar] [CrossRef]
- Úrbez-Torres, JR.; Bruez, E.; Hurtado, J.; Gubler, WD. Effect of temperature on conidial germination of Botryosphaeriaceae species infecting grapevines. Plant Dis. 2010, 94(12), 1476–84. [Google Scholar]
- Amalfitano, C.; Evidente, A.; Surico, G.; Tegli, S.; Bertelli, E.; Mugnai, L. Phenols and Stilbene Polyphenols in the Wood of Esca-Diseased Grapevines. Phytopathol Mediterr. 2000, 39, 178–183. [Google Scholar]
- Del Rio, J.; Gonzalez, A.; Fuster, M.; Botia, J.; Gomez, P.; Frias, V.; Ortuño, A. Tylose formation and changes in phenolic compounds of grape roots infected with Phaeomoniella chlamydospore and Phaeocremonium species. Phytopathol Mediterr. 2001, 40, 394–399. [Google Scholar]
- Bruno, G.; Sparapano, L. Effects of three esca-associated fungi on Vitis vinifera L.: I. Characterization of secondary metabolites in culture media and host responses to the pathogens in calli. Physiol Mol Plant Pathol 2006, 69, 209–223. [Google Scholar]
- Martin, N.; Vesentini, D.; Rego, C.; Monteiro, S.; Oliveira, H.; Ferreira, RB. Phaeomoniella chlamydospora infection induces changes in phenolic compounds content in Vitis vinifera. Phytopathol Mediterr. 2009, 48, 101–116. [Google Scholar]
- Spagnolo, A.; Magnin-Robert, M.; Alayi, TD.; Cilindre, C.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Clément, C.; Larignon, P.; Ramirez-Suero, M.; Chong, J.; Bertsch, C.; Abou-Monsour, E.; Fontaine, F. Differential responses of three grapevine cultivars to (BD). Phytopathol 2014, 104, 1021–1031. [Google Scholar]
- Gómez, P.; Báidez, AG.; Ortuño, A.; Del Río, JA. Grapevine xylem response to fungi involved in trunk diseases. Annals of Applied Biology. 2016, 169(1), 116–24. [Google Scholar]
- Pearce, RB. Antimicrobial defences in the wood of living trees. New Phytologist. 1996, 132, 203–33. [Google Scholar]
- Pouzoulet, J.; Pivovaroff, AL.; Santiago, LS.; Rolshausen, PE. Can vessel dimension explain tolerance toward fungal vascular wilt diseases in woody plants? Lessons from Dutch elm disease and esca disease in grapevine. Front Plant Sci 2014, 5, 253. [Google Scholar]
- Ramírez, M.; Pérez, L.M.; Montealegre, J.R. Susceptibility of different grapevine (Vitis vinifera L.) cultivars to Diplodia seriata and Diplodia mutila. Cien. Inv. Agr. 2018, 45, 93–98. [Google Scholar]
- Pouzoulet, J.; Scudiero, E.; Schiavon, M.; Rolshausen, P.E. Xylem Vessel Diameter Affects the Compartmentalization of the Vascular Pathogen Phaeomoniella chlamydospora in Grapevine. Front Plant Sci 2017, 21, 1442. [Google Scholar]
- Amponsah, NT.; Jones, EE.; Ridgway, HJ.; Jaspers, M. Infection and disease progression of Neofusicoccum luteum in grapevine plants. Plant Health Management:An Integrated Approach 2009, 70. [Google Scholar]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, TJ. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathol. 2010, 100(12), 1340–51. [Google Scholar] [CrossRef]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Cimmino, A.; Evidente, A. Phytotoxic metabolites by nine species of Botryosphaeriaceae involved in grapevine dieback in Australia and identification of those produced by Diplodia mutila, Diplodia seriata, Neofusicoccum australe and Neofusicoccum luteum. Nat Prod Res. 2019, 33(15), 2223–9. [Google Scholar]
- Martos, S.; Andolfi, A.; Luque, J.; Mugnai, L.; Surico, G.; Evidente, A. Production of phytotoxic metabolites by five species of Botryosphaeriaceae causing decline on grapevines, with special interest in the species Neofusicoccum luteum and N. parvum. Eur J Plant Pathol. 2008, 121(4), 451–61. [Google Scholar]
- Ramírez, M.; Pérez, LM.; Montealegre, JR. Susceptibility of different grapevine (Vitis vinifera L.) cultivars to Diplodia seriata and Diplodia mutila. Cienc Investig Agrar 2018, 45, 93–98. [Google Scholar] [CrossRef]
Figure 1.
Severity of the damage of the symptoms caused by D. seriata on detached canes of two-year-old Vitis vinifera cv. Cabernet Sauvignon. (a) Mean vascular lesion length. (b) Means canker lesion length. (c) Vascular and canker lesions on cuttings with activated isolates. Means with a common letter are not significantly different, according to Tukey’s test (P ≤ 0.05). The light blue arrow indicates the point of inoculation. The red line describes vascular discoloration.
Figure 1.
Severity of the damage of the symptoms caused by D. seriata on detached canes of two-year-old Vitis vinifera cv. Cabernet Sauvignon. (a) Mean vascular lesion length. (b) Means canker lesion length. (c) Vascular and canker lesions on cuttings with activated isolates. Means with a common letter are not significantly different, according to Tukey’s test (P ≤ 0.05). The light blue arrow indicates the point of inoculation. The red line describes vascular discoloration.
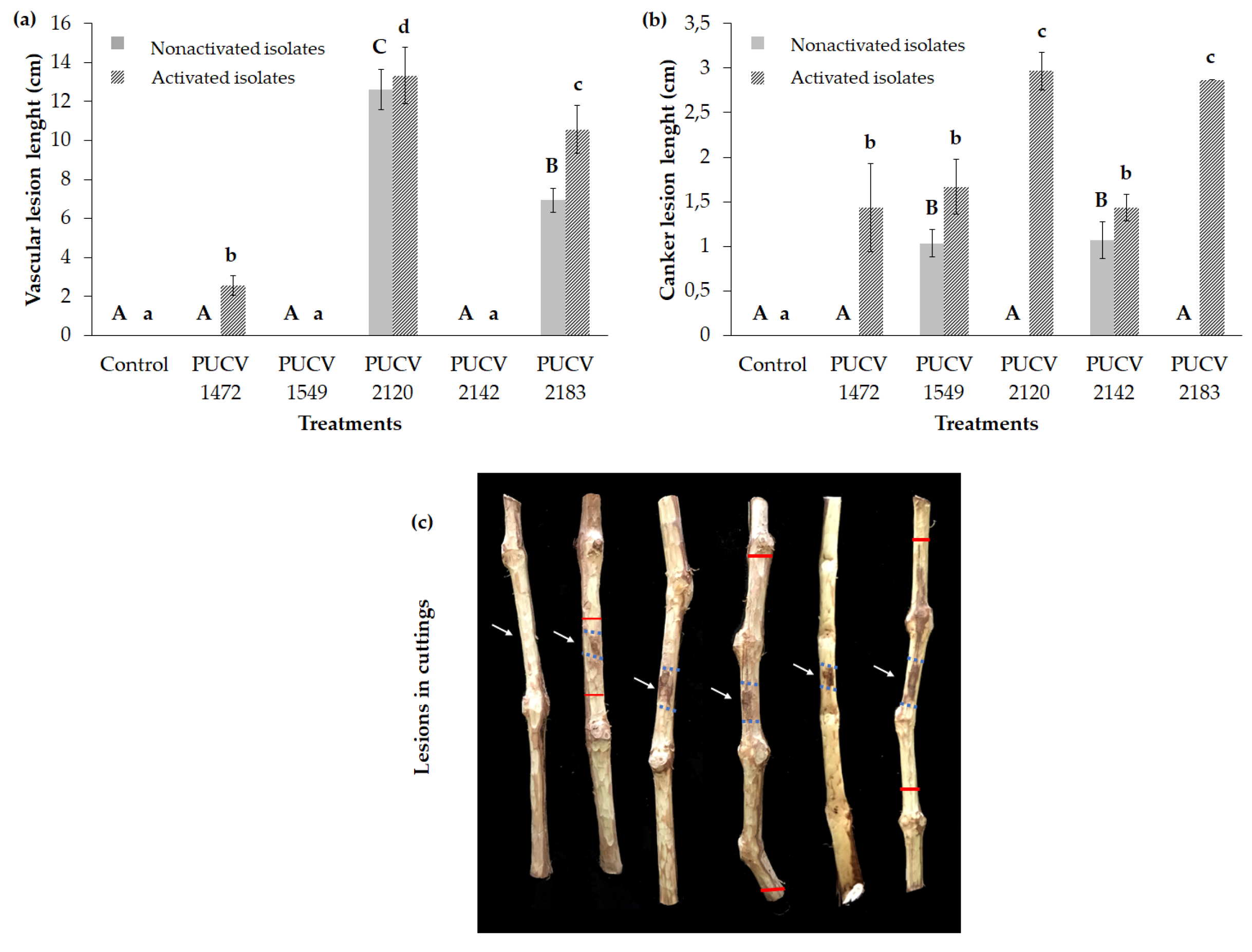
Figure 2.
Soft rot lesions caused by D. seriata on Vitis vinifera cv. Red Globe. a) Circular and sunken rot around the point of inoculation with the different isolates. b) Internal damage of inoculated berry grapes, control berries (noninoculated) and berries isolated with recovery of D. seriata from inoculated berry grapes.
Figure 2.
Soft rot lesions caused by D. seriata on Vitis vinifera cv. Red Globe. a) Circular and sunken rot around the point of inoculation with the different isolates. b) Internal damage of inoculated berry grapes, control berries (noninoculated) and berries isolated with recovery of D. seriata from inoculated berry grapes.
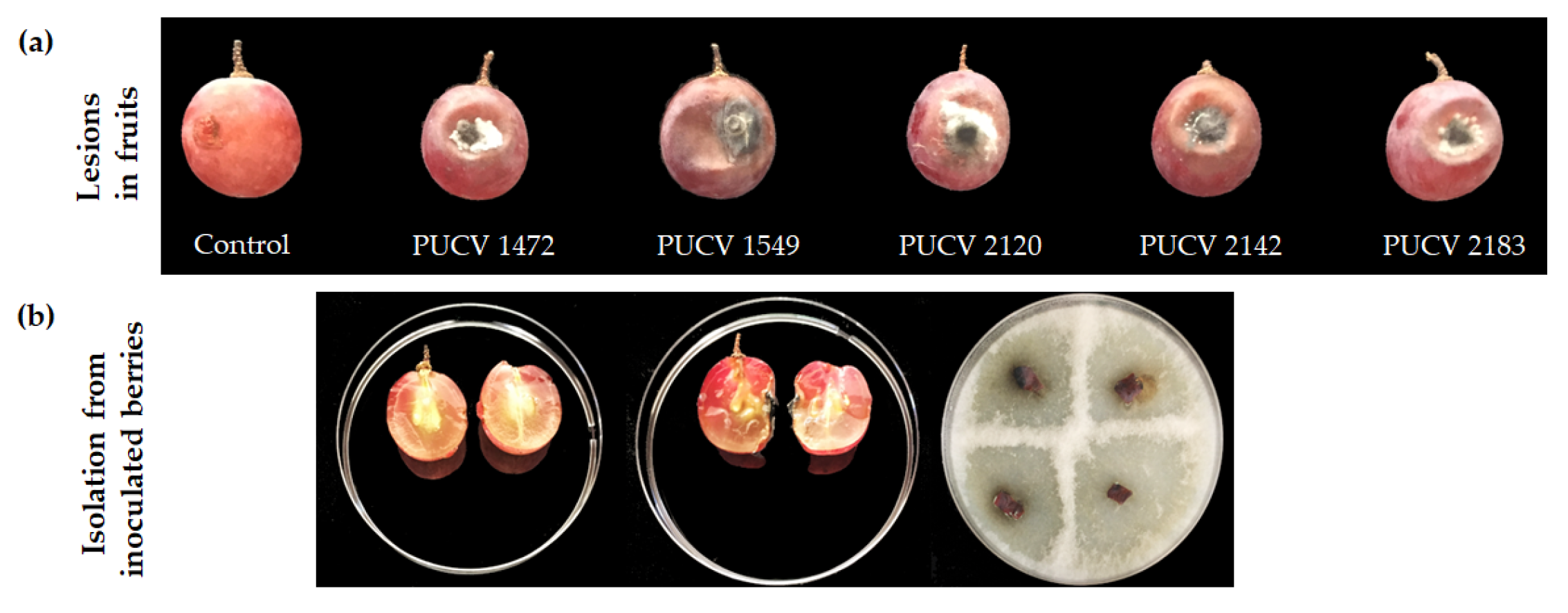
Figure 3.
Damage severity of symptoms (vascular lesion length) caused by Diplodia seriata on a tissue of different ages of Vitis vinifera cv. Cabernet Sauvignon. T00: without cutting or inoculation; T0: cutting plus sterile distilled water. PUCV 2120: D. seriata isolate. PUCV 2183: D. seriata isolate.
Figure 3.
Damage severity of symptoms (vascular lesion length) caused by Diplodia seriata on a tissue of different ages of Vitis vinifera cv. Cabernet Sauvignon. T00: without cutting or inoculation; T0: cutting plus sterile distilled water. PUCV 2120: D. seriata isolate. PUCV 2183: D. seriata isolate.
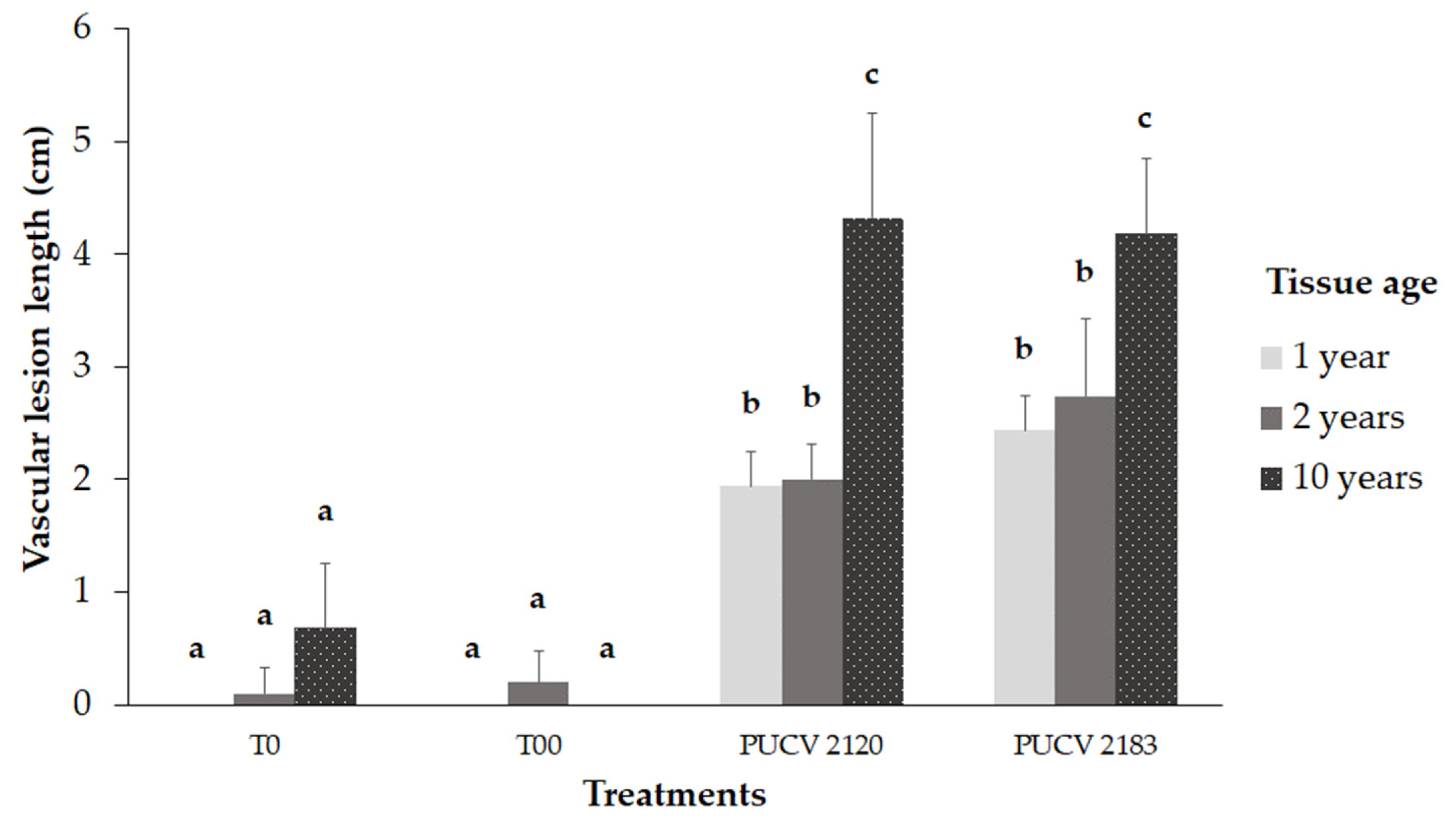
Figure 4.
Symptoms developed on Vitis vinifera cv. Cabernet Sauvignon plants five months after inoculation with D. seriata. a) Left to right: two-year-old tissue vascular lesion and 10-year-old tissue vascular lesion caused by the PUCV 2021 isolate and 10-year-old tissue of noninoculated plants (T0). b) General view of the field trial five months after the inoculation of the plants. c) Plant control (T0: noninoculated and d) Plant inoculated with D. seriata (PUCV 2120) on 10-year-old wood, no visible foliar symptoms. The white arrow indicates the inoculation point. Red lines delimit vascular discoloration. Scale bar a-b = 2 cm.
Figure 4.
Symptoms developed on Vitis vinifera cv. Cabernet Sauvignon plants five months after inoculation with D. seriata. a) Left to right: two-year-old tissue vascular lesion and 10-year-old tissue vascular lesion caused by the PUCV 2021 isolate and 10-year-old tissue of noninoculated plants (T0). b) General view of the field trial five months after the inoculation of the plants. c) Plant control (T0: noninoculated and d) Plant inoculated with D. seriata (PUCV 2120) on 10-year-old wood, no visible foliar symptoms. The white arrow indicates the inoculation point. Red lines delimit vascular discoloration. Scale bar a-b = 2 cm.

Figure 6.
Locality origin of the Diplodia seriata isolates. On the left, the O’Higgins and Maule regions are highlighted on the map of Chile. On the right are the localities of origin of each isolate of D. seriata and its year of isolation.
Figure 6.
Locality origin of the Diplodia seriata isolates. On the left, the O’Higgins and Maule regions are highlighted on the map of Chile. On the right are the localities of origin of each isolate of D. seriata and its year of isolation.
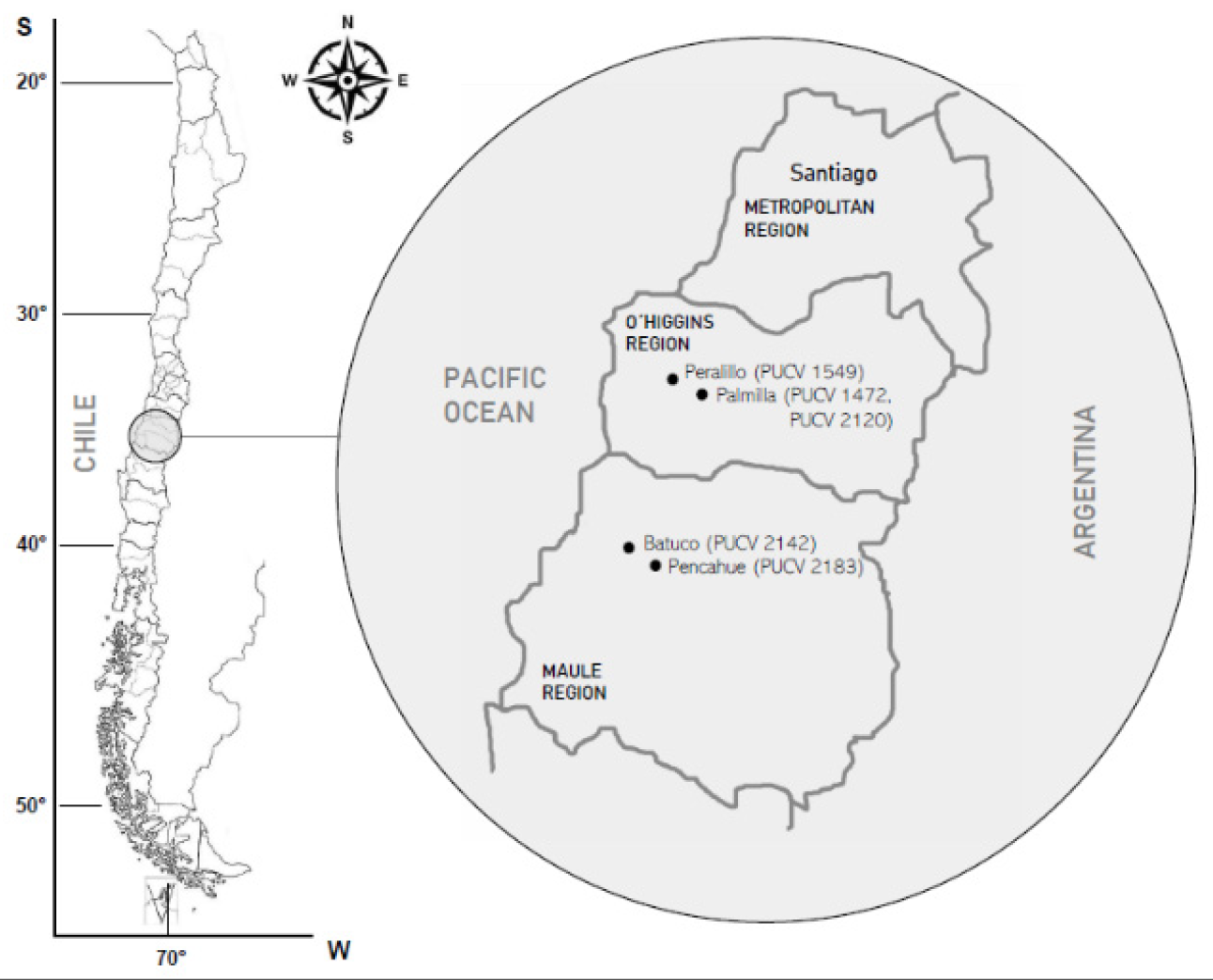
Figure 7.
Inoculation with conidia solutions of D. seriata in grapevine plants cv. Cabernet Sauvignon. a) Inoculation of two-year-old shoots on fresh pruning cut. b) Inoculation of 10-year-old wood (arm) freshly cut by chainsaw.
Figure 7.
Inoculation with conidia solutions of D. seriata in grapevine plants cv. Cabernet Sauvignon. a) Inoculation of two-year-old shoots on fresh pruning cut. b) Inoculation of 10-year-old wood (arm) freshly cut by chainsaw.


Table 1.
Chilean isolates of Diplodia seriata obtained from canker and vascular lesions of Vitis vinifera cv. Cabernet Sauvignon used for pathogenicity tests.
Table 1.
Chilean isolates of Diplodia seriata obtained from canker and vascular lesions of Vitis vinifera cv. Cabernet Sauvignon used for pathogenicity tests.
| Isolate of Diplodia seriata* | Year of Collection | GenBank Access No. | ||
|---|---|---|---|---|
| ITS | BT | EF1-α | ||
| PUCV 1472 | 2010 | KM372581 | KP762454 | - |
| PUCV 1549 | 2010 | KM580514 | KP762464 | - |
| PUCV 2120 | 2018 | MT023573 | MT063140 | MT120827 |
| PUCV 2142 | 2018 | MT023574 | MT063141 | MT120827 |
| PUCV 2183 | 2018 | MT023587 | MT063154 | - |
*Larach et al. [5].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



