
Submitted:
07 July 2023
Posted:
10 July 2023
You are already at the latest version
Abstract
The oncogenic potential of human papillomavirus (HPV) may be used to determine the tissue tropism of each HPV type. Cervical cancer develops in the squamo-columar junction of the cervices, and most such lesions are induced by high-risk (HR) HPV types. This suggests that HR types preferentially infect the cervix, whereas the preferential infection site for low-risk (LR) types is not well defined. Determination of HPV tropism when using cytology samples can be uncertain since it is difficult to avoid contamination of cell samples between the cervix and the vagina. Herein, cell samples were carefully collected by independently scraping the cervix and vagina, after which the HPV types were determined. HPV tissue tropism was determined by considering what HPV types were positive at only one of the sites (the cervix or the vagina) as the viruses that preferentially infected that site. This method revealed that all LR types were only identified in vaginal samples, whereas 87% of HR types were identified in cervical sites. Thus, LR types may preferentially infect the vagina, whereas HR types infect the cervix. These findings suggest that preferential tissue tropism of certain HPV types is a probable factor for malignant progression.
Keywords:
human papillomavirus
; tropism
; contamination
; cervical sample
; vaginal sample
1. Introduction
Approximately 50 human papillomavirus (HPV) types have so far been identified from the mucosal epithelium of the uterine cervix, vagina, and vulva [1]. Of these, the World Health Organization suggests that 12 HPVs (HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58, and HPV59) are defined as high-risk (HR) types, and HPV68 is classified as a possible HR type (pHR) by epidemiological evidence up to 2009 [2]. Pathological evidence has demonstrated cervical high-grade squamous intraepithelial lesions (HSIL) and cancer development in the squamocolumnar junction (SCJ) of the uterine cervix [3,4,5]. However, the risk categories are not certain for such HPV types as HPV26, HPV30, HPV34, HPV53, HPV66, HPV67, HPV69, HPV70, HPV73, HPV82, and HPV85 that belong to the α-5, α-6, α-7, α-9, and α-11 phylogenetic classification, and these types are considered as pHR types since they were identified in cervical cancer tissue [6,7,8]. HPV types that do not belong to these groups are likely to be low-risk (LR) types since these types are rarely identified in cancer, and molecular evidence for E6 and E7 proteins of these HPV types suggests a low carcinogenic potential in vivo studies [9,10]. Differences in carcinogenicity are caused by diverse promoter activity and gene expression patterns and functional distinctions between the E6 and E7 proteins [9,10].
Another mechanism for these differences in carcinogenicity may be tissue tropism [11,12]. Most cervical HSIL and cancers develop in the SCJ or transformation zone of the cervix, and many lesions are induced by HR-HPV types [3,4,5]. Several studies have demonstrated that HR-HPV types preferentially infect the columnar/metaplastic cells of the SCJ of the cervix as opposed to the mature squamous epithelium of the vagina [11,12,13,14,15], whereas pHR and LR types prefer the vaginal epithelium [14,16,17,18]. Thus, it is logical that HR types are strongly involved in the development of cervical lesions because of their location in the SCJ where specific cells with stem cell-like potential exist. However, conflicting findings suggest that pHR types with a preference for vaginal epithelium can also be detected in cervical carcinoma [6,7,8,18], whereas HR types with a preference for cervical epithelium are involved in the development of high-grade vaginal intraepithelial lesions (VaIN) [19,20]. In addition, because hypothetical tropism does not imply the absolute infection of a specific site [11,12], the presence of an LR-type lacking a preference for the cervix in a cervical sample cannot be dismissed as contamination from the vagina, which is somewhat confusing considering the possibility of preclinical cervical infection or intraepithelial lesions caused by LR types [20]. The cause of these discrepancies and confusion may be attributed to previous analyses that were inaccurately performed as data were estimated from relative differences in HPV prevalence using samples of uncertain provenance with selection bias reflecting the cervical or vaginal epithelium, e.g., scraped cervical cells from hysterectomized/nonhysterectomized patients [15,16].
Therefore, this study sought to define the HPV types that preferentially infect cervical or vaginal tissues by eliminating cross-contamination between sites from HPV genotyping results of cytology samples from the cervix, upper vaginal, and lower vaginal tissues. Our results supports the hypothesis that LR types preferentially infect vaginal tissue and that HR types preferentially infect cervical tissue while the pHR types do not display a tissue preference.
2. Results
The prevalence and exact 95% CIs of HPV types in the cervical and vaginal (upper or lower) samples from the 264 HPV-positive cases is shown in Table 1. The common HPV types in the cervix were HPV52 (25.0%) and HPV51 (13.6%). The common HPV types in the upper vagina were HPV52 (25.0%) and HPV58 (12.9%), and in the lower vagina were HPV52 (25.0%) and HPV74 (8.7%). The prevalence of HPV tended to decrease with distance from the cervix. Chi-squared tests of independence between each sample and HPV type, HPV risk group, and α phylogenetic group did not identify any significant associations between these groups.
All HPV types detected in the 86 patients with inconsistent HPV type results between cervical and vaginal samples are shown in Table 2. The mean age of these patients was 40 years old (range, 24–65 years old). To eliminate as much contamination between sites as possible, HPV types detected in only one of the samples were identified as the HPV type that preferentially infected that site. The prevalence of HPV types with cervical or vaginal preference in the HPV risk groups and α-phylogenetic groups is shown in Table 3. A strong significant association was found between the HPV risk group and cervical and vaginal preference (χ2(2) = 52.612, p < 0.01). HR types were significantly more common in the cervix than in the vagina, with 87% preferentially infecting the cervix (p < 0.01). HR types were 6.7-fold more common in the cervix than in the vagina. All LR types preferentially infected the vagina (p < 0.01). The prevalence of pHR types was comparable between the cervix and the vagina. A strong significant association with the α phylogenetic group was also found (χ2(4) = 47.375, p < 0.01). The α-5 and α-7 phylogenetic groups preferentially infected the cervix over the vagina, 91.7% (p < 0.05) and 86.8% (p < 0.01), respectively. The α-5 and α-7 phylogenetic groups were 11.0- and 6.6-fold more frequent in the cervix than in the vagina, respectively. The α-11/α-1/α-8/α-10/α-13/α-3/α-15 phylogenetic groups all preferentially infected the vagina (p < 0.01).
Preferentially, assuming that LR types infect the vagina and HR types infect the cervix, the colocalization rates (including possible contamination) of different HPV risk group in 178 cervical and vaginal samples with HPV type concordance in each sample are shown in Table 4. The colocalization rates of LR and HR types in cervical and vaginal samples were 34.8% and 73.6%, respectively.
3. Discussion
Herein, we used a highly sensitive HPV polymerase chain reaction (PCR) method to study HPV tissue tropism in cell samples from the cervix and vagina. Our results suggest that the LR and HR types have a significant preference for vaginal and cervical epithelium, respectively, while the pHR type displayed no tissue preference. This study strongly supports the hypothesis that mucosal HPVs have tropism for specific sites within the lower genital tract.
Prior studies of HPV tropism compared women with hysterectomies to those without [15,16] and women younger than 50 to those 50 and older [14]. These comparisons were made on the basis that with the former population, the cervical region is abraded for collection of columnar and metaplastic cells of the cervical SCJ, whereas with the latter populations, only cells from the mature squamous epithelium similar to that of the vagina were collected. However, age-related sampling and hysterectomy-related potential biases may have affected these study results. To eliminate such biases, Castle et al. [17] collected paired vaginal and cervical specimens from the same patients and examined the relationship between vaginal and cervical HPVs at the type-specific level. They revealed that noncarcinogenic HPV types of the α-3/α-15 and α-1/α-8/α-10 phylogenetic groups, which corresponded to LR types, may have tropism for the vaginal epithelium, whereas HR types of the α-9/α-11 groups appeared to have similar affinity for both the vaginal and cervical epithelium. Although their study was similar to our analysis in terms of sampling methods, our results showed a clear preference of HR HPVs or α5/α9 groups for cervical infection and that of LR HPVs or α-11/α-1/α-8/α-10/α-13/α-3/α-15 groups for vaginal infection. The reported susceptibility of both tissues to the α-9/α-11 groups [17] is also explained by the cervix and vagina being preferentially infected by the α-9 and α-11 groups respectively.
We consider that our results differ in part from those of previous studies because we eliminated contamination by vaginal cells in the cervical samples and by cervical cells in the vaginal samples to the extent possible. Thus, the HPV detection results at the three sites showed that the identified HPV types were almost identical. The only way to reveal HPV preference in this situation is to identify HPV types that are uncommon in each sample. We believe that this method of tropism analysis is why we could clarify the tissue susceptibility of each risk group.
Tropism is accepted as a preference for a particular anatomical site and is not considered to be directly related to the development of cancer at that site [11,12]. This supposition is supported given that LR types prefer the vagina but rarely induce the development of vaginal lesions. E6 and E7 proteins of the LR-type are not reportedly involved in the development of HSIL because they do not drive host cell proliferation or transforming these cells into cancer cells [21]. We previously reported that no LR types caused a single-type infection in patients with HSIL and cancer of the cervix [22]. In addition to functional differences in E6 and E7 protein activity between the LR and HR types, the LR-type does not preferentially infect cervical tissue, which might explain why the LR HPV types are less likely to cause persistent infection and induce the development of HSIL.
The genotypes of pHR HPVs are undetected in normal tissues, but because of the low prevalence in patients with cervical cancer, consensus has not emerged to classify these HPVs as HR types [23]. The low prevalence of pHR types in the cervix is probably explained by the lack of susceptibility of the cervix to genotypes of pHR HPVs, as all α-11 genotypes (HPV34 and HPV73) displayed a preference for the vaginal epithelium. pHR types have been demonstrated to preferentially infect the vaginal epithelium [18], which is inconsistent with their involvement in the development of high-grade cervical lesions. This may be explained here as we identified that α-5 (HPV26 and HPV82) and α-9 (HPV67) in the pHR group exhibited tropism for the cervix.
A cervical abrasion sample used for cervical cancer screening is often assumed to contain only cervical cells. However, assuming that LR types preferentially infect the vagina, our colocalization analysis revealed that approximately 30% of cervical specimens contained a mixture of LR type infected cells of vaginal origin. In cervical cancer screening, a preclinical cervical infection is often determined when the Hybrid Capture II (HC) assay is positive and biopsy is negative [24]. Although the specificity of HC varies depending on whether the endpoint is cervical intraepithelial neoplasm (CIN) 2+ or not, some of the reported 7%–25% false positives [25] may require consideration of possible contamination of LR-type infected cells from the vagina to the cervix, given that HC cross-reacts with LR-type and becomes positive [26]. Conversely, the vagina is considered the reservoir for any mucosal HPV types [20], and this may be because LR types preferential infect the vagina while vaginal accumulation of cervix-derived HR and pHR type-infected cells occurs during most phases of the menstrual cycle [27]. Colocalization analysis also revealed HR types in approximately 70% of the vaginal samples, presumably caused by cervical to vaginal contamination. Under vaginal infection by various HPV types, the HR type is mainly known to cause high-grade VaIN and vaginal cancer [19]. Previously, we used manual microdissection of lesions from formalin-fixed, paraffin-embedded tissue specimens to demonstrate that the HR or pHR type is responsible for high-grade VaIN and squamous cell carcinoma [20]. However, the prevalence of vaginal lesions is significantly lower than that of cervical lesions, with an annual incidence of <1 per 100,000 women [28]. This confirms our results that HR types are less likely to preferentially infect the vagina. By contrast, women with a history of CIN2+ have been reported to have higher frequencies of VaIN and vaginal cancer because of persistent infection by HR types [29,30]. Thus, if HR types contribute to the development of cervical lesions they would also infect the vaginal epithelium. Nevertheless, we recently found that HPV types from VaIN and CIN lesions in the same patient differed in 92.3% (24/26) of cases, suggesting that VaIN and CIN develop independently [20]. Therefore, the higher risk of VaIN in patients with CIN is not because of the HPV strains that contribute to the development of CIN as these are more likely to be transmitted because they prefer the vagina. Instead, the immune response of the host [31,32], who is prone to acquiring persistent infection, is similarly impaired in the vagina. Consequently, we believe that HPV tissue tropism is not a significant factor in the development of vaginal lesions but due to the success of any of the various HR or pHR types in the vaginal reservoir in establishing a persistent infection.
This study had some limitations. First, our statistics and inferences were restricted by the limited number of samples included in this study. Furthermore, this study only speculated on the tropism of HPV for the cervix and vagina based on HPV genotyping using cell samples obtained from a small population of women. In addition, our selection criteria for this study may have biased our findings. Unfortunately, this study did not have accurate HPV type data for the biopsied tissues, and we could not assess whether each HPV risk group truly exhibited tropism for each tissue. Nonetheless, given that similar results were obtained in studies using paired scraped cervical and vaginal samples, further research is warranted because additional data could further clarify HPV tropism and the development of cervical and vaginal lesions.
4. Materials and Methods
4.1. Patients and Specimen Collection
A total of 324 randomly selected patients who had been referred for colposcopy because of abnormalities identified during cervical cytologic screening at Tokyo Genki Plaza Health Medical Center from 2020 to 2021 were enrolled in the study. Three types of SurePath™ (Becton Dickinson and Company, Franklin Lakes, NJ, USA) liquid-based cytology (LBC) were used to collect samples from the cervix and upper and lower vagina tissues of the patients.
Immediately before colposcopic assessment, a vaginal speculum was used to expose the cervix of each patient, and then the endocervix and portio vaginalis were scraped using an IM sampler instrument (Muto Pure Chemical, Tokyo, Japan) consisting of a spatula and brush and eluted into a collection vial, which was used as the cervical sample. Then, while holding the vaginal speculum open, the bilateral portions from the vaginal vestibule to the upper half of the vaginal canal were scraped with a spatula and collected as the upper vaginal sample, and cells from the lower half of the vaginal canal were collected in this same manner as the lower vaginal sample. Samples were collected after obtaining written informed consent from the patients. The Ethics Committee on Human Research of Kyorin University approved the study protocol, which was implemented in accordance with the approved guidelines. Patients who were negative for HPV in three different samples were excluded, and consequently, 264 patients who were positive for any HPV type in any of the samples were evaluated (Figure 1). The patients were nonpregnant women with a mean age of 40 (range, 22–71) years. Several samples used in the present study had been previously reported in a study of the effects of menstrual cycle on the accumulation of HPV-infected cells exfoliated from the cervix that drift into the vagina [27].
4.2. HPV Genotyping Using LBC Samples
DNA was isolated from the cell pellet of each LBC sample using the hot sodium hydroxide method [33]. Cell pellets were lysed with 50 μL of alkaline lysis solution (25 mM NaOH and 0.2 mM ethylenediaminetetraacetic acid; pH 12.0) for 30 min at 95°C. Lysed cells were then neutralized with 0.04 M Tris-HCl (pH 5.0), centrifuged at 13,200 rpm for 1 min, and directly used as the DNA template for PCR amplification. All HPV-positive samples tested positive for human β-globin DNA, demonstrating that DNA of amplifiable quality was extracted from the specimens. HPV genotyping was performed using a highly sensitive HPV PCR method known as uniplex E6/E7 PCR [34]. This method can identify the E6 or E7 genes of 39 mucosal HPV types, including 12 HR types (HPV16, HPV18, HPV31, HPV33, HPV35, HPV39, HPV45, HPV51, HPV52, HPV56, HPV58, and HPV59), 12 pHR types (HPV26, HPV30, HPV34, HPV53, HPV66, HPV67, HPV68, HPV69, HPV70, HPV73, HPV82, and HPV85), and 15 LR types (HPV6, HPV11, HPV40, HPV42, HPV44, HPV54, HPV55, HPV61, HPV62, HPV71, HPV74, HPV81, HPV84, HPV89, and HPV90), from as few as 100 viral copies, with no cross-reactivity across all HPV genotypes.
4.3. Data Analysis
Prevalence estimates and exact 95% CIs were calculated for each individual HPV type, HPV risk group, and α phylogenetic group detected in cervical, upper vaginal, and lower vaginal samples from 264 patients, and HPV types that preferred the cervix or vagina were then evaluated from the individual HPV types detected in the 86 patients with inconsistent HPV types in the vaginal (upper and lower) samples versus the cervical samples. HPV types detected in only one of the samples were identified as the HPV type that preferentially infected that site. For example, if HPV56 and HPV62 were positive in the cervical sample and only HPV62 was positive in the vaginal upper or lower samples, HPV62 was considered to be a virus that had migrated from one site and contaminated the other; HPV56 was identified as the only virus that preferentially infected the cervix. Differences in the prevalence of each HPV type were compared by using a z test for the difference between two proportions with Bonferroni adjustment for multiple comparisons using Statistical Package for the Social Sciences version 25.0 (SPSS Inc., Chicago, IL, USA). Furthermore, we investigated the colocalization rate, including possible contamination of HPV types with vaginal tropism in the cervical samples and with cervical tropism in the vaginal samples based on the results of tropism analysis.
5. Conclusions
This study evaluated HPV tissue tropism and eliminated cross-contamination of HPV genotyping results using cervical and vaginal cytology samples. The results emphasize that LR types preferentially infect vaginal tissue while HR types preferentially infect cervical tissue. Thus, preferential tissue tropism of certain HPV type is a probable factor for malignant progression.
Author Contributions
Conceptualization, M.O. (Mitsuaki Okodo); methodology, M.O. (Mitsuaki Okodo), R.S., S.M. and Y.I.; validation, M.O. (Mitsuaki Okodo) and K.O.; formal analysis, M.O. (Mitsuaki Okodo) and K.T.; investigation, M.O. (Mitsuaki Okodo), K.O., R.S., S.M. and Y.I.; resources, M.O. (Mizue Oda); data curation, M.O. (Mitsuaki Okodo), K.O. and K.T.; writing—original draft preparation, M.O. (Mitsuaki Okodo); writing—review and editing, M.O. (Mitsuaki Okodo), K.O. and T.S; visualization, K.O.; supervision, M.O. (Mitsuaki Okodo); funding acquisition, M.O. (Mitsuaki Okodo); project administration, M.O. (Mitsuaki Okodo) and M.O. (Mizue Oda), All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by JSPS KAKENHI, grant number 23K09676.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of Human Research of Kyorin University (2020-39, 17 March 2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data and material that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to thank Enago (www.enago.jp) for the English language review.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Van Doorslaer, K.; Li, Z.; Xirasagar, S.; Maes, P.; Kaminsky, D.; Liou, D.; Sun, Q.; Kaur, R.; Huyen, Y.; McBride, A.A. The Papillomavirus Episteme: a major update to the papillomavirus sequence database. Nucleic Acids Res. 2017, 45, D499–D506. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Human papillomaviruses. IARC Monogr. Eval. Carcinog. Risks Hum. 2007, 90, 1–636. [Google Scholar]
- de Sanjosé, S.; Brotons, M.; Pavón, M.A. The natural history of human papillomavirus infection. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 2–13. [Google Scholar] [CrossRef]
- Schiffman, M.; Castle, P.E.; Jeronimo, J.; Rodriguez, A.C.; Wacholder, S. Human papillomavirus and cervical cancer. Lancet 2007, 370, 890–907. [Google Scholar] [CrossRef]
- Mirkovic, J.; Howitt, B.E.; Roncarati, P.; Demoulin, S.; Suarez-Carmona, M.; Hubert, P.; McKeon, F.D.; Xian, W.; Li, A.; Delvenne, P.; Crum, C.P.; Michael, H. Carcinogenic HPV infection in the cervical squamo-columnar junction. J. Pathol. 2015, 236, 265–271. [Google Scholar] [CrossRef]
- de Sanjosé, S.; Quint, W.G.; Alemany, L.; Geraets, D.T.; Klaustermeier, J.E.; Lloveras, B.; Tous, S.; Felix, A.; Bravo, L.E.; Shin, H.-R.; Vallejos, C.S.; Ruiz, P.A.d.; Lima, M.A.; Guimera, N.; Clavero, O.; Alejo, M.; Llombart-Bosch, A.; Cheng-Yang, C.; Tatti, S.A.; Kasamatsu, E.; Iljazovic, E.; Odida, M.; Prado, R.; Seoud, M.; Grce, M.; Usubutun, A.; Jain, A.; Suarez, G.A.H.; Lombardi, L.E.; Banjo, A.; Menéndez, C.; Domingo, E.J.; Velasco, J.; Nessa, A.; Chichareon, S.C.B.; Qiao, Y.L.; Lerma, E.; Garland, S.M.; Sasagawa, T.; Ferrera, A.; Hammouda, D.; Mariani, L.; Pelayo, A.; Steiner, I.; Oliva, E.; Meijer, C.J.; Al-Jassar, W.F.; Cruz, E.; Wright, T.C.; Puras, A.; Llave, C.L.; Tzardi, M.; Agorastos, T.; Garcia-Barriola, V.; Clavel, C.; Ordi, J.; Andújar, M.; Castellsagué, X.; Sánchez, G.I.; Nowakowski, A.M.; Bornstein, J.; Muñoz, N.; Bosch, F.X. Human papillomavirus genotype attribution in invasive cervical cancer: a retrospective cross-sectional worldwide study. Lancet Oncol. 2010, 11, 1048–1056. [Google Scholar] [CrossRef]
- Bzhalava, D.; Guan, P.; Franceschi, S.; Dillner, J.; Clifford, G. A systematic review of the prevalence of mucosal and cutaneous human papillomavirus types. Virology 2013, 445, 224–231. [Google Scholar] [CrossRef]
- Halec, G.; Alemany, L.; Lloveras, B.; Schmitt, M.; Alejo, M.; Bosch, F.X.; Tous, S.; Klaustermeier, J.E.; Guimerà, N.; Grabe, N.; Lahrmann, B.; Gissmann, L.; Quint, W.; Bosch, F.X.; de Sanjosé, S.; Pawlita, M. Pathogenic role of the eight probably/possibly carcinogenic HPV types 26, 53, 66, 67, 68, 70, 73 and 82 in cervical cancer. J. Pathol. 2014, 234, 441–451. [Google Scholar] [CrossRef]
- Münger, K.; Howley, P.M. Human papillomavirus immortalization and transformation functions. Virus Res. 2002, 89, 213–228. [Google Scholar] [CrossRef]
- Fehrmann, F.; Laimins, L.A. Human papillomaviruses: targeting differentiating epithelial cells for malignant transformation. Oncogene 2003, 22, 5201–5207. [Google Scholar] [CrossRef]
- De Villiers, E.-M.; Fauquet, C.; Broker, T.R.; Bernard, H.-U.; Zur Hausen, H. Classification of papillomaviruses. Virology 2004, 324, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Egawa, N.; Egawa, K.; Griffin, H.; Doorbar, J. Human papillomaviruses; epithelial tropisms, and the development of neoplasia. Viruses 2015, 7, 3863–3890. [Google Scholar] [CrossRef]
- Baldwin, S.; Santos, C.; Brown, E.M.; Nuño, T.; Giuliano, A.; Davis, J.; Garcia, F. Comparison of type-specific human papillomavirus data from self and clinician directed sampling. Gynecol. Oncol. 2005, 97, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Castle, P.E.; Jeronimo, J.; Schiffman, M.; Herrero, R.; Rodríguez, A.C.; Bratti, M.C.; Hildesheim, A.; Wacholder, S.; Long, L.R.; Neve, L.; Pfeiffer, R.; Burk, R.D. Age-related changes of the cervix influence human papillomavirus type distribution. Cancer Res. 2006, 66, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- D'Souza, G.; Burk, R.D.; Zhong, Y.; Minkoff, H.; Stewart Massad, L.; Xue, X.; Heather Watts, D.; Anastos, K.; Palefsky, J.M.; Levine, A.M.; Colie, C.; Castle, P.E.; Strickler, H.D. Cervicovaginal human papillomavirus (HPV)-infection before and after hysterectomy: evidence of different tissue tropism for oncogenic and nononcogenic HPV types in a cohort of HIV-positive and HIV-negative women. Int. J. Cancer 2012, 131, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Castle, P.E.; Schiffman, M.; Bratti, M.C.; Hildesheim, A.; Herrero, R.; Hutchinson, M.L.; Rodriguez, A.C.; Wacholder, S.; Sherman, M.E.; Kendall, H.; Viscidi, R.P.; Jeronimo, J.; Schussler, J.E.; Burk, R.D. A population-based study of vaginal human papillomavirus infection in hysterectomized women. J. Infect. Dis. 2004, 190, 458–467. [Google Scholar] [CrossRef]
- Castle, P.E.; Rodriguez, A.C.; Porras, C.; Herrero, R.; Schiffman, M.; Gonzalez, P.; Hildesheim, A.; Burk, R.D. A comparison of cervical and vaginal human papillomavirus. Sex. Transm. Dis. 2007, 34, 849–855. [Google Scholar] [CrossRef]
- Winer, R.L.; Hughes, J.P.; Feng, Q.; O’Reilly, S.; Kiviat, N.B.; Koutsky, L.A. Comparison of incident cervical and vulvar/vaginal human papillomavirus infections in newly sexually active young women. J. Infect. Dis. 2009, 199, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Tsimplaki, E.; Argyri, E.; Michala, L.; Kouvousi, M.; Apostolaki, A.; Magiakos, G.; Papassideri, I.; Panotopoulou, E. Human papillomavirus genotyping and e6/e7 mRNA expression in Greek women with intraepithelial neoplasia and squamous cell carcinoma of the vagina and vulva. J. Oncol. 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Zhang, S.; Saito, M.; Okayama, K.; Okodo, M.; Kurose, N.; Sakamoto, J.; Sasagawa, T. HPV Genotyping by molecular mapping of tissue samples in vaginal squamous intraepithelial neoplasia (VaIN) and vaginal squamous cell carcinoma (VaSCC). Cancers (Basel) 2021, 13, 3260. [Google Scholar] [CrossRef]
- Halec, G.; Schmitt, M.; Dondog, B.; Sharkhuu, E.; Wentzensen, N.; Gheit, T.; Tommasino, M.; Kommoss, F.; Bosch, F.X.; Franceschi, S.; Clifford, G.; Gissmann, L.; Pawlita, M. Biological activity of probable/possible high-risk human papillomavirus types in cervical cancer. Int. J. Cancer 2013, 132, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Okayama, K.; Kimura, H.; Teruya, K.; Ishii, Y.; Fujita, K.; Fujii, M.; Oda, M.; Sasagawa, T.; Okodo, M. Correlation between human papillomavirus codetection profiles and cervical intraepithelial neoplasia in Japanese women. Microorganisms 2020, 8, 1863. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.; Bosch, F.X.; De Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.L.M.; Group, I.A.f.R.o.C.M.C.C.S. Epidemiologic classification of human papillomavirus types associated with cervical cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Saslow, D.; Solomon, D.; Lawson, H.W.; Killackey, M.; Kulasingam, S.L.; Cain, J.; Garcia, F.A.; Moriarty, A.T.; Waxman, A.G.; Wilbur, D.C.; Wentzensen, N.; Downs, L.S.; Spitzer, M.; Moscicki, A.-B.; Franco, E.L.; Stoler, M.H.; Schiffman, M.; Castle, P.E.; Myers, E.R.; Chelmow, D.; Herzig, A.; Kim, J.J.; Kinney, W.; Herschel, W.L.; Waldman, J. American Cancer Society, American Society for Colposcopy and Cervical Pathology, and American Society for Clinical Pathology screening guidelines for the prevention and early detection of cervical cancer. CA Cancer J. Clin. 2012, 62, 147–172. [Google Scholar] [CrossRef]
- Arbyn, M.; Ronco, G.; Anttila, A.; Meijer, C.J.; Poljak, M.; Ogilvie, G.; Koliopoulos, G.; Naucler, P.; Sankaranarayanan, R.; Peto, J. Evidence regarding human papillomavirus testing in secondary prevention of cervical cancer. Vaccine 2012, 30, F88–F99. [Google Scholar] [CrossRef]
- Castle, P.E.; Schiffman, M.; Burk, R.D.; Wacholder, S.; Hildesheim, A.; Herrero, R.; Bratti, M.C.; Sherman, M.E.; Lorincz, A. Restricted cross-reactivity of hybrid capture 2 with nononcogenic human papillomavirus types. Cancer Epidemiol. Biomarkers Prev 2002, 11, 1394–1399. [Google Scholar]
- Okodo, M.; Okayama, K.; Teruya, K.; Tanabe, K.; Ito, C.; Ishii, Y.; Fujii, M.; Kimura, H.; Oda, M. Effects of menstrual cycle on the accumulation of human papillomavirus-infected cells exfoliated from the cervix that drift into the vagina. Microorganisms 2022, 10, 693. [Google Scholar] [CrossRef]
- Khan, M.J.; Massad, L.S.; Kinney, W.; Gold, M.A.; Mayeaux Jr, E.; Darragh, T.M.; Castle, P.E.; Chelmow, D.; Lawson, H.W.; Huh, W.K. A common clinical dilemma: management of abnormal vaginal cytology and human papillomavirus test results. Gynecol. Oncol. 2016, 141, 364–370. [Google Scholar] [CrossRef]
- Murta, E.F.; Neves Junior, M.A.; Sempionato, L.R.F.; Costa, M.C.; Maluf, P.J. Vaginal intraepithelial neoplasia: clinical-therapeutic analysis of 33 cases. Arch. Gynecol. Obstet. 2005, 272, 261–264. [Google Scholar] [CrossRef]
- Frega, A.; Sopracordevole, F.; Assorgi, C.; Lombardi, D.; De Sanctis, V.; Catalano, A.; Matteucci, E.; Milazzo, G.N.; Ricciardi, E.; Moscarini, M. Vaginal intraepithelial neoplasia: a therapeutical dilemma. Anticancer Res. 2013, 33, 29–38. [Google Scholar]
- Kohaar, I.; Thakur, N.; Salhan, S.; Batra, S.; Singh, V.; Sharma, A.; Sodhani, P.; Das, B.C.; Sarkar, D.P.; Bharadwaj, M. TNF α–308G/A polymorphism as a risk factor for HPV associated Cervical Cancer in Indian population. Anal. Cell. Pathol. 2007, 29, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Kohaar, I.; Hussain, S.; Thakur, N.; Tiwari, P.; Nasare, V.; Batra, S.; Singh, V.; Bhambani, S.; Das, B.C.; Sarkar, D.P.; Bharadwaj, M. Association between human leukocyte antigen class II alleles and human papillomavirus-mediated cervical cancer in Indian women. Hum. Immunol. 2009, 70, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Truett, G.E.; Heeger, P.; Mynatt, R.L.; Truett, A.A.; Walker, J.A.; Warman, M.L. Preparation of PCR-quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques 2000, 29, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Okodo, M.; Okayama, K.; Teruya, K.; Sasagawa, T. Uniplex E6/E7 PCR method detecting E6 or E7 genes in 39 human papillomavirus types. J. Med. Virol. 2018, 90, 981–988. [Google Scholar] [CrossRef]
Figure 1.
Stratification of patients evaluated for cervical and vaginal tropism and contamination of cervical samples by human papillomavirus (HPV) with vaginal tropism.
Figure 1.
Stratification of patients evaluated for cervical and vaginal tropism and contamination of cervical samples by human papillomavirus (HPV) with vaginal tropism.
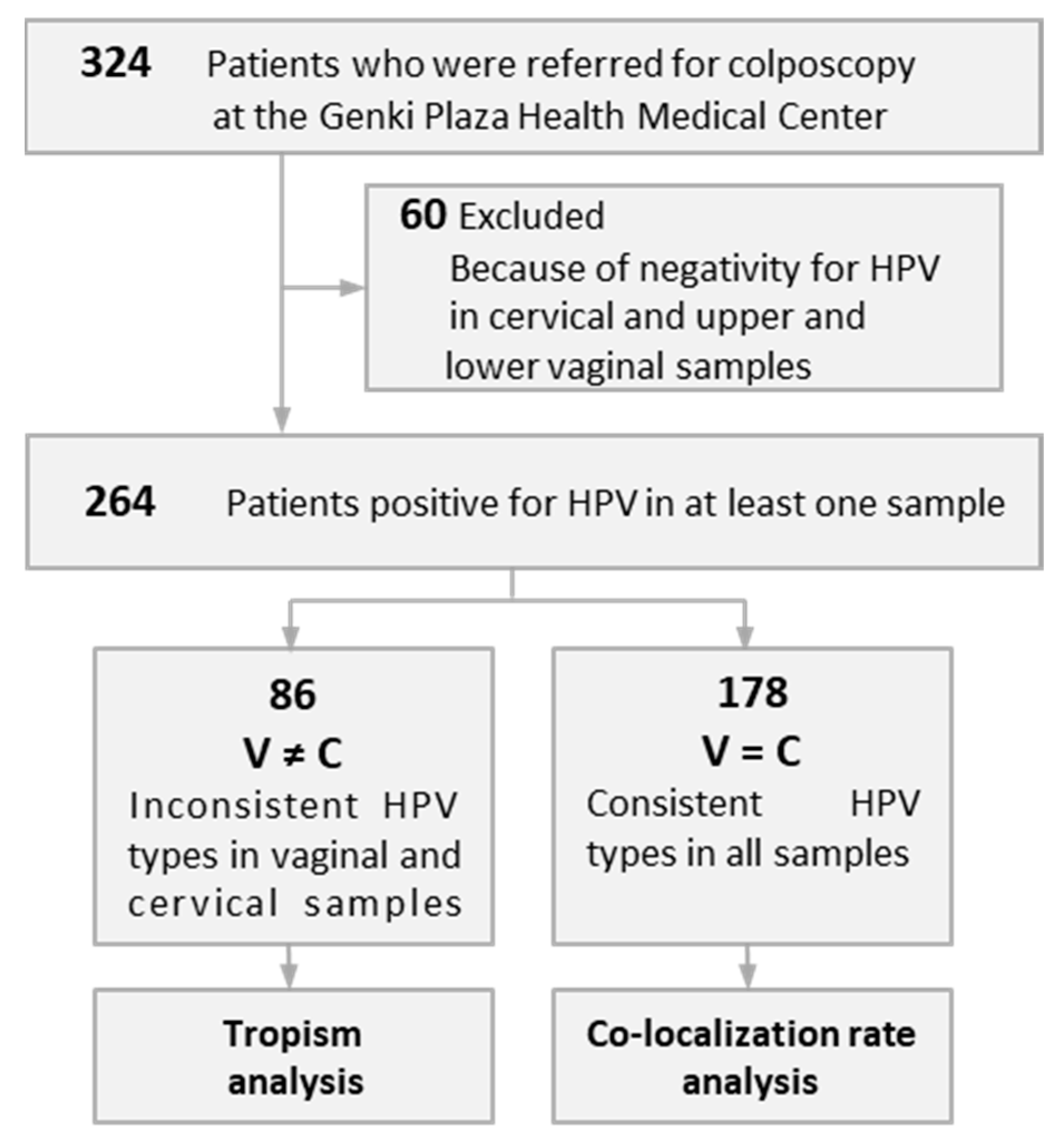

Table 1.
Prevalence of all HPV types detected in cervical samples and upper or lower vaginal samples from 264 patients.
Table 1.
Prevalence of all HPV types detected in cervical samples and upper or lower vaginal samples from 264 patients.
| Prevalence (95% CI) % | |||
|---|---|---|---|
| Cervical samples | Vaginal samples | ||
| Upper | Lower | ||
| HPV types | |||
| HPV16 | 12.9 (8.8–16.9) | 10.6 (6.9–14.3) | 4.5 (2.0–7.1) |
| HPV18 | 1.9 (0.2–3.5) | 1.1 (0.0–2.4) | 0 |
| HPV31 | 9.5 (5.9–13) | 7.6 (4.4–10.8) | 2.7 (0.7–4.6) |
| HPV33 | 2.7 (0.7–4.6) | 2.3 (0.5–4.1) | 1.1 (0.0–2.4) |
| HPV35 | 0.4 (0.1–1.1) | 0.4 (0.0–1.1) | 0.4 (0.0–1.1) |
| HPV39 | 3.8 (1.5–6.1) | 3.4 (1.2–5.6) | 1.9 (0.2–3.5) |
| HPV45 | 1.1 (0.0–2.4) | 1.1 (0.0–2.4) | 0.8 (0.0–1.8) |
| HPV51 | 13.6 (9.5–17.8) | 10.2 (6.6–13.9) | 4.5 (2.0–7.1) |
| HPV52 | 25.0 (19.8–30.2) | 19.3 (14.6–24.1) | 11.4 (7.5–15.2) |
| HPV56 | 9.5 (5.9–13.0) | 8.0 (4.7–11.2) | 5.3 (2.6–8.0) |
| HPV58 | 12.9 (8.8–16.9) | 12.1 (8.2–16.1) | 5.3 (2.6–8.0) |
| HPV59 | 4.2 (1.8–6.6) | 4.5 (2.0–7.1) | 3.4 (1.2–5.6) |
| HPV26 | 0.4 (0.0–1.1) | 0 | 0 |
| HPV30 | 0.8 (0.0–1.8) | 0.8 (0.0–1.8) | 0.4 (0.0–1.1) |
| HPV34 | 1.9 (0.2–3.5) | 2.3 (0.5–4.1) | 1.1 (0.0–2.4) |
| HPV53 | 8.0 (4.7–11.2) | 8.3 (5.0–11.7) | 4.2 (1.8–6.6) |
| HPV66 | 8.3 (5.0–11.7) | 8.3 (5.0–11.7) | 5.7 (2.9–8.5) |
| HPV67 | 3.8 (1.5–6.1) | 2.7 (0.7–4.6) | 1.5 (0.0–3.0) |
| HPV69 | 1.1 (0.0–2.4) | 1.1 (0.0–2.4) | 0.8 (0.0–1.8) |
| HPV68 | 3.0 (1.0–5.1) | 3.0 (1.0–5.1) | 1.5 (0.0–3.0) |
| HPV70 | 1.5 (0.0–3.0) | 0 | 0.8 (0.0–1.8) |
| HPV73 | 0 | 0.4 (0.0–1.1) | 0 |
| HPV82 | 7.6 (4.4–10.8) | 6.8 (3.8–9.9) | 0 |
| HPV6 | 1.5 (0.0–3.0) | 1.5 (0.0–3.0) | 1.5 (0.0–3.0) |
| HPV11 | 0.4 (0.0–1.1) | 0.4 (0.0–1.1) | 0 |
| HPV40 | 2.7 (0.7–4.6) | 3.0 (1.0–5.1) | 0 |
| HPV42 | 6.4 (3.5–9.4) | 7.2 (4.1–10.3) | 4.5 (2.0–7.1) |
| HPV44 | 0.4 (0.0–1.1) | 0.4 (0.0–1.1) | 0.4 (0.0–1.1) |
| HPV54 | 3.4 (1.2–5.6) | 3.8 (1.5–6.1) | 1.9 (0.2–3.5) |
| HPV55 | 1.9 (0.3–3.5) | 2.3 (0.5–4.1) | 1.9 (0.2–3.5) |
| HPV61 | 3.4 (1.2–5.6) | 3.4 (1.2–5.6) | 1.1 (0.0–2.4) |
| HPV62 | 4.9 (2.3–7.5) | 5.7 (2.9–8.5) | 2.7 (0.7–4.6) |
| HPV71 | 2.7 (0.7–4.6) | 2.7 (0.7–4.6) | 0.4 (0.0–1.1) |
| HPV74 | 8.7 (5.3–12.1) | 9.8 (6.3–13.4) | 5.7 (2.9–8.5) |
| HPV81 | 3.4 (1.2–5.6) | 4.2 (1.8–6.6) | 1.9 (0.2–3.5) |
| HPV84 | 0.8 (0.0–1.8) | 1.5 (0.0–3.0) | 1.1 (0.0–2.4) |
| HPV89 | 0.8 (0.0–1.8) | 0.8 (0.0–1.8) | 0.4 (0.0–1.1) |
| HPV90 | 4.5 (2.0–7.1) | 5.3 (2.6–8.0) | 3.4 (1.2–5.6) |
| Risk groups | |||
| High-risk type | 95.1 (92.5–97.7) | 80.7 (75.9–85.4) | 41.3 (35.3–47.2) |
| Possible high-risk type | 34.5 (29.1–40.6) | 35.2 (29.5–41.0) | 17.4 (12.8–22.0) |
| Low-risk type | 39.0 (33.1–44.9) | 50.4 (44.3–56.4) | 26.9 (21.5–32.2) |
| α phylogenetic groups | |||
| α-5 | 21.6 (16.6–26.6) | 18.2 (13.5–22.8) | 6.8 (3.8–9.9) |
| α-7 | 15.2 (10.8–19.5) | 14.8 (10.5–19.1) | 8.3 (5.0–11.7) |
| α-9 | 65.2 (59.4–70.9) | 54.9 (48.9–60.9) | 26.9 (21.5–32.2) |
| α-6 | 25.8 (20.8–31.4) | 25.4 (20.1–30.6) | 15.5 (11.2–19.9) |
| α-11/α-1/α-8/α-10/α-13/α-3/α-15 | 40.9 (35.0–46.8) | 53.0 (47.0–59.1) | 28.0 (22.6–33.4) |
HPV, human papillomavirus.
Table 2.
HPV types that preferentially infected the cervix or vagina.
| Cases | Age | HPV types | All HPV types detected | |||
|---|---|---|---|---|---|---|
| Preferentially infected the cervix | Preferentially infected the vagina | Cervical samples | Vaginal samples | |||
| Upper | Lower | |||||
| 1 | 51 | 56 (HR, α-6) | − | 56, 62 | 62 | Negative |
| 2 | 37 | − | 81 (LR, α-3) | Negative | 81 | 81 |
| 3 | 31 | 31 (HR, α-9) | − | 31, 81, 82 | 81, 82 | Negative |
| 4 | 34 | − | 16 (HR, α-9) | 39, 56, 71 | 16, 39, 56, 71 | 39, 56 |
| 5 | 24 | − | 90 (LR, α-15) | 59 | 59, 90 | 59, 90 |
| 6 | 41 | 16 (HR, α-9) | − | 16 | Negative | Negative |
| 7 | 50 | − | 84 (LR, α-3) | Negative | 84 | 84 |
| 8 | 31 | 52 (HR, α-9) | − | 16, 45, 52, 40 | 16, 45, 40 | 16, 45, 40 |
| 9 | 40 | − | 56 (HR, α-6) | Negative | 56 | 56 |
| 10 | 50 | − | 66 (pHR, α-6) | 51, 56, 58 | 51, 56, 58 | 51, 56, 66 |
| 11 | 35 | 52 (HR, α-9) | − | 52, 53, 74 | 53, 74 | 53, 74 |
| 12 | 26 | − | 59 (HR, α-7) | 56, 66, 74, 90 | 56, 59, 66, 74, 90 | 56, 59, 66, 74, 90 |
| 13 | 40 | 56 (HR, α-6) | − | 56 | Negative | Negative |
| 14 | 50 | 51 (HR, α-5) | − | 51, 58 | 58 | 58 |
| 15 | 45 | − | 34 (pHR, α-11) | Negative | Negative | 34 |
| 16 | 26 | 58 (HR, α-9) | − | 52, 58 | 52 | 52 |
| 17 | 58 | − | 52 (HR, α-9) | 34 | 52, 34 | 34 |
| 18 | 45 | − | 34 (pHR, α-11) | 31,74 | 31, 34, 74 | Negative |
| 19 | 26 | − | 54 (LR, α-13), 66 (pHR, α-5), 74 (LR, α-10), 90 (LR, α-15) | 56 | 56, 54, 66 | 56, 54, 66, 74, 90 |
| 20 | 42 | 39 (HR, α-7) | − | 39, 58 | 58 | 58 |
| 21 | 43 | 51 (HR, α-5) | − | 51 | Negative | Negative |
| 22 | 31 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 23 | 47 | 16 (HR, α-9) | − | 16 | Negative | Negative |
| 24 | 42 | 51 (HR, α-5) | − | 51 | Negative | Negative |
| 25 | 30 | − | 40 (LR, α-8) | 39 | 39, 40 | 39, 40 |
| 26 | 45 | 31 (HR, α-9) | − | 31,74 | 74 | 74 |
| 27 | 28 | 16 (HR, α-9) | − | 16, 52, 56, 55, 74, 81 | 52, 56, 55, 74, 81 | 52, 56, 55, 74, 81 |
| 28 | 43 | 58 (HR, α-9) | − | 52, 58 | 52 | 52 |
| 29 | 56 | 16 (HR, α-9) | − | 16, 42 | 42 | 42 |
| 30 | 53 | − | 58 (HR, α-9), 82 (pHR, α-5) | 45, 51, 90 | 45, 51, 58, 82, 90 | 90 |
| 31 | 25 | 56 (HR, α-6) | − | 56, 34 | 34 | 34 |
| 32 | 32 | 16 (HR, α-9) | − | 16 | Negative | Negative |
| 33 | 46 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 34 | 53 | 51 (HR, α-5), 82 (pHR, α-5) | − | 51, 82 | Negative | Negative |
| 35 | 34 | 82 (pHR, α-5) | − | 51, 82 | 51 | 51 |
| 36 | 53 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 37 | 29 | 52 (HR, α-9) | − | 33, 52 | 33 | Negative |
| 38 | 48 | − | 62 (LR, α-3) | 52, 53, 81 | 52, 53, 62, 81 | 53, 62 |
| 39 | 51 | 58 (HR, α-9) | − | 51, 58 | 51 | 51 |
| 40 | 32 | − | 81 (LR, α-3) | 31 | 31, 81 | 31, 81 |
| 41 | 31 | − | 42 (LR, α-1) | 58 | 58 | 58, 42 |
| 42 | 50 | 66 (pHR, α-6) | − | 40, 66 | 40 | Negative |
| 43 | 51 | 51 (HR, α-5) | − | 51, 56, 58 | 56, 58 | 56 |
| 44 | 40 | 56 (HR, α-6) | − | 56 | Negative | Negative |
| 45 | 49 | 67 (pHR, α-9) | − | 67 | Negative | Negative |
| 46 | 48 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 47 | 59 | − | 74 (LR, α-10) | 44 | 44, 74 | 44 |
| 48 | 51 | − | 11 (LR, α-10) | 35, 67 | 35, 11, 67 | 35 |
| 49 | 33 | 31 (HR, α-9) | − | 31, 58, 74 | 58, 74 | 58, 74 |
| 50 | 26 | − | 55 (LR, α-10) | 52, 56, 59, 30, 54, 74 | 52, 56, 59, 30, 54, 55, 74 | 52, 56, 59, 30, 54, 55, 74 |
| 51 | 37 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 52 | 37 | − | 42 (LR, α-1), 62 (LR, α-3) | 51, 53, 82 | 51, 42, 53, 62, 82 | 51, 42, 53, 62, 82 |
| 53 | 54 | 31 (HR, α-9) | 58 (HR, α-9), 66 (pHR, α-6) | 31, 51, 82, 6b | 51, 58, 6b, 82 | 6b, 66, 82 |
| 54 | 57 | 16 (HR, α-9) | − | 16, 42 | 42 | 42 |
| 55 | 31 | 52 (HR, α-9) | − | 31, 52, 58, 70 | 31, 58, 70 | 31, 58, 70 |
| 56 | 40 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 57 | 51 | 82 (pHR, α-5) | − | 51, 58, 82 | 51, 58 | 51, 58 |
| 58 | 65 | 53 (pHR, α-6) | − | 53 | Negative | Negative |
| 59 | 28 | − | 53 (pHR, α-6), 74 (LR, α-10) | 16, 52 | 16, 52, 53, 74 | 16, 52, 53, 74 |
| 60 | 46 | − | 74 (LR, α-10) | 31 | 31, 74 | 74 |
| 61 | 30 | 66 (pHR, α-6) | − | 53, 66 | 53 | 53 |
| 62 | 37 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 63 | 32 | 31 (HR, α-9) | − | 31 | Negative | Negative |
| 64 | 25 | − | 84 (LR, α-3) | Negative | 84 | 84 |
| 65 | 38 | 18 (HR, α-7) | − | 18 | Negative | Negative |
| 66 | 47 | − | 66 (pHR, α-6), 90 (LR, α-15) | 16 | 16, 66, 90 | Negative |
| 67 | 54 | 52 (HR, α-9) | − | 52 | Negative | Negative |
| 68 | 28 | − | 90 (LR, α-15) | 42 | 42 | 42, 90 |
| 69 | 40 | 16 (HR, α-9) | 59 (HR, α-7) | 16, 74 | 59, 74 | 74 |
| 70 | 43 | − | 53 (pHR, α-6) | 18 | 18, 53 | 18, 53 |
| 71 | 33 | 51 (HR, α-5) | − | 51, 74 | 74 | 74 |
| 72 | 42 | − | 67 (pHR, α-9) | Negative | Negative | 67 |
| 73 | 30 | − | 74 (LR, α-10) | 16, 39 | 16, 39 | 16, 39, 74 |
| 74 | 49 | 67 (pHR, α-9) | − | 67 | Negative | Negative |
| 75 | 26 | 56 (HR, α-6) | − | 56 | Negative | Negative |
| 76 | 34 | 51 (HR, α-5) | − | 51 | Negative | Negative |
| 77 | 27 | − | 54 (LR, α-13) | 34 | 34, 54 | 34, 54 |
| 78 | 50 | 66 (pHR, α-6) | − | 40, 66 | 40 | 40 |
| 79 | 46 | − | 73 (pHR, α-11) | 54, 61 | 61, 73 | 54,61, 73 |
| 80 | 51 | 58 (HR, α-9) | − | 51, 56, 58 | 51, 56 | 51, 56 |
| 81 | 35 | 26 (pHR, α-5) | − | 26, 90 | 90 | Negative |
| 82 | 56 | − | 89 (LR, α-3) | Negative | Negative | 89 |
| 83 | 26 | 52 (HR, α-9) | − | 52, 59 | 59 | Negative |
| 84 | 29 | 33(HR, α-9), 52 (HR, α-9) | − | 33, 52 | Negative | Negative |
| 85 | 28 | − | 42 (LR, α-1) | 16, 54, 82 | 16, 42, 54, 82 | 16, 54, 82 |
| 86 | 27 | 18 (HR, α-7), 59 (HR, α-7) | − | 18, 31, 52, 59, 55, 67, 74, 90 | 31, 52, 55, 67, 74, 90 | 31, 55, 67, 74, 90 |
In the three columns on the right, all HPV types detected in cervical and vaginal samples are compared, and HPV types detected in only one of the samples are shown in bold as the HPV type that preferentially infected that site. The HPV types per case are shown in the order of the HR type and then other type. In the left column, HPV types are indicated for the preferentially infected sites. HPV, human papillomavirus; LR, low-risk; HR, high-risk; pHR, possible high-risk.
Table 3.
Prevalence of HPV that preferentially infected the cervix or vagina in HPV risk groups and α-phylogenetic groups.
Table 3.
Prevalence of HPV that preferentially infected the cervix or vagina in HPV risk groups and α-phylogenetic groups.
| Preferentially infected | Ratio(cervix / vagina) | |||
|---|---|---|---|---|
| N | Cervix | Vagina | ||
| Risk groups | ||||
| High-risk type | 54 | 87.0%** | 13.0% | 6.7 |
| Possible high-risk type | 21 | 47.6% | 52.4% | 0.9 |
| Low-risk type | 24 | 0% | 100.0%** | 0.0 |
| α phylogenetic groups | ||||
| α-5 | 12 | 91.7%* | 8.3% | 11.0 |
| α-7 | 6 | 66.7% | 33.3% | 2.0 |
| α-9 | 38 | 86.8%** | 13.2% | 6.6 |
| α-6 | 16 | 56.3% | 43.8% | 1.3 |
| α-11/α-1/α-8/α-10/α-13/α-3/α-15 | 27 | 0% | 100.0%** | 0.0 |
**p < 0.01, *p < 0.05, HPV, human papillomavirus.
Table 4.
Colocalization rate (including possible contamination) of different HPV risk groups in the cervical samples or in the vaginal samples in 178 patients.
Table 4.
Colocalization rate (including possible contamination) of different HPV risk groups in the cervical samples or in the vaginal samples in 178 patients.
| Combination of infectingHPV types | Samples (n) | Colocalization rate by LR typesin cervical samples | Colocalization rate by HR typesin vaginal samples | ||
|---|---|---|---|---|---|
| LR | 18 | 34.8% | − | ||
| LR | + pHR | 8 | |||
| LR | + HR | 24 | 73.6% | ||
| LR | + HR | + pHR | 12 | ||
| HR | 79 | − | |||
| HR | + pHR | 16 | |||
| pHR | 21 | − | |||
HPV, human papillomavirus; LR, low-risk; HR, high-risk; pHR, possible high-risk.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



