
Submitted:
10 July 2023
Posted:
11 July 2023
You are already at the latest version
Abstract
Febrile seizures (FS) are a relatively common early-life condition that can cause CNS developmental disorders, but the specific mechanisms of action of FS are poorly understood. In this work, we used hyperthermia-induced FS in 10-day-old rats. By recording local field potentials, we showed that the efficiency of glutamatergic synaptic transmission decreased 15 min after FS. This effect was transient, and after 2 days there were no differences between control and post-FS groups. During early ontogeny, the proportion of calcium-permeable (CP)-AMPA receptors in the synapses of principal cortical and hippocampal neurons is high. Therefore, rapid internalization of CP-AMPA receptors may be one of the mechanisms underlying this phenomenon. Using the whole-cell patch clamp method and the selective CP-AMPA receptor blocker IEM-1460, we tested whether the proportion of CP-AMPA receptors changes. We have demonstrated that FS rapidly reduces synaptic CP-AMPA receptors in both the hippocampus and the entorhinal cortex. This process was accompanied by a sharp decrease in calcium permeability of the membrane of principal neurons, which we revealed in experiments with kainate-induced cobalt uptake. Our experiments show that FSs cause rapid changes in the function of the glutamatergic system, which may have compensatory effects that prevent excessive excitotoxicity and neuronal death.
Keywords:
febrile seizures
; hyperthermia
; hippocampus
; GluA2-lacking AMPA receptor
; entorhinal cortex
; IEM-1460
; development
1. Introduction
Febrile seizures (FS) are one of the most common neurological disorders in childhood [1,2,3]. Short febrile seizures (<10 min) are considered relatively harmless, but complex seizures lasting more than 15 min can lead to several neurological disorders in the future [4]. These include the development of pharmacoresistant temporal lobe epilepsy [2,5,6].
Data from an experimental animal model of FS have also shown that prolonged FS leads to long-term changes in the properties of hippocampal neurons with critical consequences for the excitability of the hippocampal network [7]. These changes further increase the susceptibility of animals to limbic seizures throughout life [8]. Evidence from other neonatal seizure models (hypoxia- or flurotil-induced seizures) strongly supports these findings and shows that seizures early in life cause irreversible functional changes in neural networks, rendering the brain vulnerable to later epilepsy and leading to cognitive impairment [9,10,11]. Even a single neonatal seizure can significantly alter the properties of glutamatergic synapses [12]. However, the specific mechanisms that mediate changes in synaptic properties following seizures remain largely unknown.
One of the common consequences of epileptic seizures is alterations in the number and subunit composition of ionotropic glutamate AMPA receptors, which have been found both in the brains of epileptic patients and in a number of experimental animal models [13,14,15,16,17]. Glutamate AMPA receptors consist of four homologous pore-forming subunits (GluA1-GluA4) that are predominantly assembled into diheteromers [18]. The functional properties of AMPA receptors depend on the presence of the GluA2 subunit, the absence of which or the presence of an unedited GluA2 subunit renders the receptor permeable to Ca2+ ions [19,20,21].
Many studies have shown that seizures result in changes in the proportion of CP-AMPA receptors. Most of the investigations have demonstrated an increase in CP-AMPA receptor expression [22,23,24,25,26,27]. The insertion of CP-AMPA receptors during seizures may result in an increase in intracellular calcium concentration with excessive activation of glutamatergic inputs and may even be a cause of neuronal death [28]. Disruptions in the number and composition of AMPA receptors are thought to underlie subsequent cognitive deficits [21,29]. However, there are studies suggesting a decrease in the expression of CP-AMPA receptors [30,31]. Thus, changes in the subunit composition of AMPA receptors and the appearance of CP-AMPA receptors depend on many factors (age of the animal, type of model, time after seizures, etc.). Age may be a key factor in altering the subunit composition of glutamate receptors. In the early stages of development, CP-AMRA receptors are widely expressed in the CNS [20]. But as the animal matures, the expression of CP-AMRA receptors in synapses decreases significantly, and in the brain of adult animals, CP-AMRA receptors are mainly found in interneurons and glial cells [20,21,32,33].
FSs are observed at an age when the expression of CP-AMPA receptors in pyramidal neurons is high, and their hyperactivation during seizures may cause excitotoxicity. However, FSs usually does not cause significant neuronal death, which has been shown in several studies [34,35,36]. Therefore, the aim of our work was to investigate the properties of glutamatergic transmission and to determine the proportion of CP-AMPA receptors in the synapses of rat hippocampus and entorhinal cortex during the acute phase (15 min) and 48 h after FSs. FSs lasting at least 15 min were induced by hyperthermia in 10-day-old (P10) rats.
2. Results
2.1. The Efficacy of Synaptic Neurotransmission at CA3-CA1 is Immediately Reduced after FSs
We first investigated whether FSs affect glutamatergic synaptic neurotransmission in CA3-CA1 of the rat hippocampus. FS were induced in P10 rats. Hippocampal slices were prepared at two time points: 15 min (n = 23 slices, N = 9 rats) and 48 h after FSs (n = 19, N = 9). We compared the results with age-matched control animals (P10: n = 21, N = 10; P12: n = 21, N = 10). We measured the amplitudes of fEPSPs and presynaptic fiber volleys (FV) recorded at the stratum radiatum of CA1 as a function of extrasynaptic stimulation intensity (25–200 μA, Figure 1a-b, e-f). We then plotted the amplitudes of fEPSPs (mainly reflecting the activation of postsynaptic AMPA receptors) against the amplitudes of FV (mainly reflecting the number of CA3 axons that evoked the action potential) for each slice (Figure 1c, g).
The slopes of these input-output (I/O) relationships reflect the composite cellular transfer function between presynaptic action potentials and the postsynaptic membrane response triggered by glutamate release. Thus, the maximum I/O slopes can be considered as a measure of basal synaptic transmission. By fitting the curves with the sigmoidal Gompertz function, we calculated the maximum I/O slopes and found that the I/O curve slopes were significantly smaller in post-FS than control animals at 15 min (control: 4.0 ± 0.4, n = 21; control:2.9 ± 0.3, n = 23; t = 2.4; p < 0.05; Figure 1d) but not at 48 h after FSs (control: 4.4 ± 0.3, n = 21; control: 3.7 ± 0.3, n = 19; t = 1.38; p = 0.18; Figure 1h).
These results suggest that FSs cause a rapid decrease in the efficiency of synaptic transmission in the hippocampus, which may be a compensatory response to the hyperactivation of glutamatergic transmission during FSs.
2.2. FSs Cause a Rapid Decrease in the Proportion of CP-AMPA Receptors in the Principal Neurons of the Entorhinal Cortex and Hippocampus
Since the proportion of CP-AMPA receptors in the synapses of principal cortical and hippocampal neurons is high during early ontogeny, we tested whether the proportion of CP-AMPA receptors changes. To assess this, we first examined the properties of evoked EPSCs in deep entorhinal pyramidal neurons under local electrical stimulation. For this purpose, AMPA-mediated eEPSCs were isolated pharmacologically with NMDA and GABAa receptor antagonists (50 μM AP5, 10 μM; MK-801, 10 μM gabazine). The stimulation current was chosen to elicit a monosynaptic response with an amplitude of 50–150 pA.
Since CP-AMPA receptors exhibit an inwardly rectifying current-voltage relationship, we performed whole-cell patch-clamp recordings of eEPSCs at –40 mV and +40 mV and calculated the rectification index as the ratio of eEPSC amplitudes at +40 and –40 mV (Figure 2a). Next, we evaluated the blocking effect of IEM-1460, the selective antagonist of CP-AMPA receptors (Figure 2b). At 15 minutes after FS, the rectification index of the eEPSC was significantly higher in post-FS rats than in control rats (Figure 2d, f), and application of IEM-1460 induced a significantly greater block of AMPA-mediated eEPSCs in control rats than in rats subjected to FS (Figure 2c, e).
However, no significant change in eEPSC block and rectification index was observed at 48 h after FSs (Figure 2e,f). Taken together, these results indicate that FSs induce a rapid decrease in the expression of CP-AMPA receptors in principal neurons of young rats.
Since only synaptic AMPA receptors were activated in this experiment, we can assume that CP-AMPA receptors were translocated from the postsynaptic density to the extrasynaptic region of the membrane or were internalized within neurons. To determine which of these possibilities is more likely, we performed an additional study in isolated neurons. In this experiment, we used isolated pyramidal neurons from the CA1 region of the hippocampus and a rapid extracellular application of kainate, which allowed us to activate all AMPA receptors on the cell membrane.
We showed that application of 100 µM kainate to isolated hippocampal CA1 pyramidal neurons from control and post-FS rats induced similar weakly desensitizing inward currents. We compared the percentage of inhibition of kainate-induced currents by the selective CP-AMPA receptor channel blocker IEM-1460 in neurons from control and post-FS rats. Representative examples of kainate-induced current inhibition at –80 mV holding voltage by 100 µM IEM-1460 are shown in Figure 3a, c.
In experiments with neurons from P10 rats, the percentage of inhibition by IEM-1460 was reduced in post-FS animals compared to the control (post-FS: 13.2 ± 1.7%, n = 20 vs. control: 21.4 ± 1.9%, n = 17, t = 3.2, p < 0.01). These results indicate that seizures in P10 rats decreased the proportion of CP-AMPA receptors on the membrane surface of hippocampal pyramidal neurons. This supports the notion that seizures induce rapid internalization of CP-AMPA receptors. In contrast, in P12 rats, the percentage of inhibition by IEM-1460 was similar in control and post-FS groups (12.1 ± 0.8%, n = 12 vs. 9.6 ± 1.0%, n = 10, t = 1.97, p = 0.06). Thus, in P12 rats, an age-related decrease in the proportion of CP-AMPA receptors in control animals compensated for the difference between the compared groups.
2.4. Kainate-Induced Co2+ Uptake in Hippocampal CA1 and CA3 Areas and in the Entorhinal Cortex is Reduced Immediately after FSs
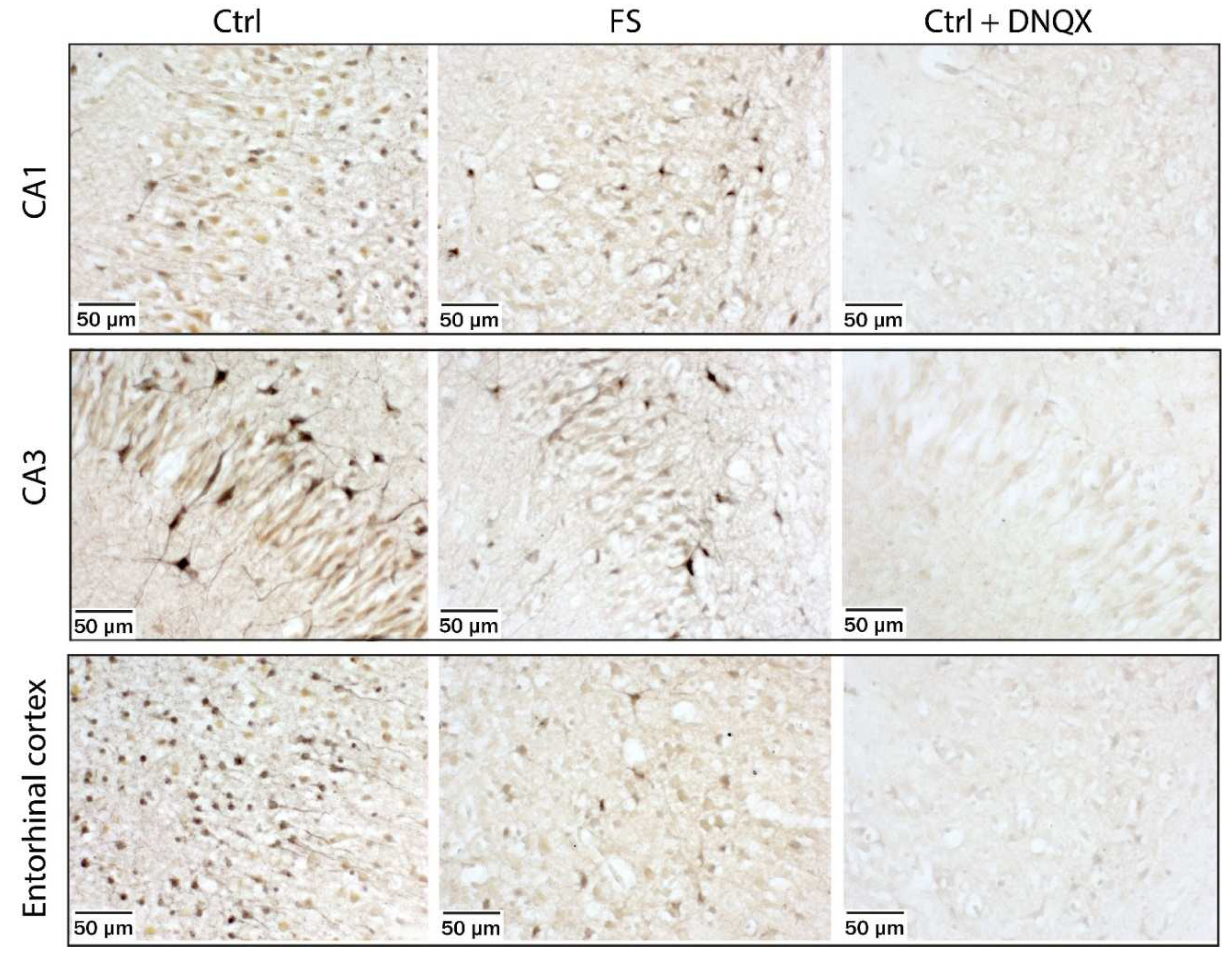
We then used Co2+ labeling [25] to directly visualize CP-AMPA receptor-containing cells in rats 15 min (Figure 4) and 48 h after FSs (Figure 5). Acute brain slices were treated with kainate in the presence of AP-5 and TTX. Co2+-stained cells were observed in all areas of the hippocampus and all layers of the entorhinal cortex. In P10 rats, stronger Co2+ staining was observed in brain sections from control than from post-FS rats. The morphology of the Co2+-stained cells and their location varied. In addition to cells resembling interneurons known to express CP-AMPA receptors, we saw a significant number of highly stained pyramidal neurons in slices from control animals. In slices from post-FS animals, highly stained pyramidal neurons were virtually absent.
To confirm that Co2+ influx was mediated by AMPA receptors, we performed a control experiment in which the AMPA receptor blocker DNQX was also added to the incubation solution. In this case, we observed a complete absence of Co2+-stained neurons (Figure 4). This experiment further confirms that FSs lead to a rapid internalization of CP-AMPA receptors from the membrane of principal neurons.
No differences were observed between P12 control and post-FS rats, either in the CA1 and CA3 areas of the hippocampus or in the entorhinal cortex (Figure 5).
Figure 6.
Kainate-induced Co2+ uptake showed greater Co2+ uptake in hippocampal CA1 and CA3 cell layers and in the entorhinal cortex in acute brain slices from P12 control rats (left panels) and poorer Co2+ uptake in rats 48 hours after FSs (right panels).
Figure 6.
Kainate-induced Co2+ uptake showed greater Co2+ uptake in hippocampal CA1 and CA3 cell layers and in the entorhinal cortex in acute brain slices from P12 control rats (left panels) and poorer Co2+ uptake in rats 48 hours after FSs (right panels).
Thus, the histological data confirm the results of the electrophysiological studies and indicate a decrease in CP-AMPA receptors in rats immediately after seizures.
3. Discussion
The results obtained indicate that FSs lead to a significant reduction in CP-AMPA receptor expression in entorhinal and hippocampal pyramidal neurons of young rats, as shown by whole-cell patch-clamp recordings and measurements of kainite-induced Co2+ uptake. These changes affect the functional properties of the neuronal network by reducing baseline synaptic transmission.
Changes in CP-AMPA receptor expression have been reported in several other seizure models. In the lithium-pilocarpine model, an increase in GluA2 expression relative to the expression of other AMPA receptor subunits was found several hours after seizures [30,31]. However, most studies show the opposite. The decrease of GluA2 expression in the hippocampus was found after kainite-induced seizures [22,26]. One week following pilocarpine-induced seizures, decreased GluA2 gene expression was found in the temporal cortex [37]. The increase of CP-AMPA receptor expression in neocortical principal cells has been observed in in vitro studies [27,38,39].
The present study used the young P10 rats, which still have high levels of CP-AMPA receptor expression in principal neurons of the neocortex and hippocampus [20,33]. Therefore, it was unclear whether FS would further increase CP-AMPA receptor levels or, conversely, cause them to decrease. Using similarly aged rats with high levels of CP-AMPA receptors in principal neurons, it has been shown that hypoxia-induced seizures further increase CP-AMPA receptor expression in synapses [9,17,25]. This elevated expression level of CP-AMPA receptors was observed within days of hypoxia-induced seizures and was confirmed by both electrophysiological (increased inwardly rectifying current-voltage relationships) and histological (enhanced staining in hippocampal slices for AMPA receptor-mediated Co2+ uptake) methods [25]. Increased AMPA receptor-mediated Ca2+ influx may contribute to the chronic epileptogenic effects of perinatal hypoxia-induced seizures and may be associated with disturbances in synaptic plasticity that later lead to epilepsy [40,41]. Thus, the decrease in CP-AMPA receptor expression observed in the present study may be unique to the FS model. In contrast to the hypoxia-induced seizure model, FSs in the model used are much less likely to lead to the development of epilepsy later in life, and the incidence of limbic epilepsy after long FSs and spontaneous seizure severity were influenced by the duration of the initial FS [8].
Rapid changes in synaptic function are often observed after an epileptic seizure [42]. It can be hypothesized that the transient downregulation of CP-AMPA receptor expression may be a compensatory mechanism that helps to minimize the consequences of seizures and results in a lack of transition to acquired epilepsy in a model of FS. This hypothesis is supported by a number of data from other studies. For example, both the seizure-induced increase in CP-AMPA receptor function and the concomitant increase in seizure susceptibility in vivo were prevented by early blockade of AMPA receptors with NBQX [9,17,40] or perampanel [43]. Although the immediate effect of CP-AMPA receptor incorporation into the synapses of principal neurons of the entorhinal cortex may be antiepileptic [44], the increased expression of CP-AMPA receptors after seizures is usually considered a pathological process that supports the transition from seizures to status epilepticus [45,46].
The fact that "seizure begets seizure" has been confirmed by extensive experimental research [47]. Among the possible mechanisms of epileptogenesis that are related to changes in the level of expression of the CP-AMPA receptors, two main ones can be distinguished. The first is that excessive activity of CP-AMPA receptors and Ca2+ entry through them in neurons that normally express calcium-impermeable AMPA receptors contributes to, or causes, delayed cell death in response to endogenous glutamate [28]. This mechanism is likely to be largely involved in the mature brain, and evidence for this has been obtained in the kainate model of temporal lobe epilepsy and partially in the lithium-pilocarpine model of temporal lobe epilepsy. However, in the immature brain, pyramidal neurons probably have some compensatory mechanisms that prevent their death by excessive activation of CP-AMPA receptors, so that no significant neurodegeneration is observed in either the hypoxic seizure model, which leads to an increase in CP-AMPA receptors, or the febrile seizure model, which leads to a decrease in CP-AMPA receptors. Therefore, we can assume that the alteration of CP-AMPA receptors in early life has no significant effect on neuronal death and is not a key factor in epileptogenesis.
The second mechanism may involve impaired synaptic plasticity. Epileptic seizures are known to induce synaptic plasticity in hippocampal and neocortical neurons. Several reports indicate that in neurons that do not express CP-AMPA receptors under baseline conditions, seizure-like activity promotes the NMDA receptor-dependent form of synaptic potentiation [27,48,49]. However, our previous results indicate that CP-AMPA receptor activity mostly opposes the long-term enhancement of hippocampal synapses [50], in contrast to its role in conventional LTP formation [51,52]. In rats, hypoxia-induced seizures at P10 cause a rapid increase in miniature EPSC amplitude and frequency in CA1 pyramidal neurons due to increased CP-AMPA receptor expression [9], and a significant decrease in the incidence of silent synapses and persistent attenuation of LTP [53]. Moreover, alterations in CP-AMPA receptor expression may impair long-term depression expression, which in turn may disrupt normal hippocampal dendritic spine maturation and pruning during development, resulting in an overabundance of less efficient excitatory synapses [17,54].
We can hypothesize that FS in younger rats expressing CP-AMPA receptors may promote a different form of long-term synaptic plasticity compared to that observed in other seizure models. We have previuosly reported that FSs impair hippocampal LTP due to insufficient NMDAR activity [36]. Several other forms of CP-AMPA receptor-related synaptic plasticity have been described in both principal neurons [55] and interneurons [56]. It was previously reported that high-frequency stimulation of the input to cerebellar stellate cells, which predominantly express CP-AMPA receptors in synapses, leads to the expression of calcium-impermeable AMPA receptors in these cells [57,58,59]. Increased Ca2+ entry through synaptic CP-AMPA receptors, but not NMDA receptors, triggers the insertion of GluA2-containing receptors into the synapse and the removal of GluA2-lacking receptors from the synapse. Acute stress can promote the CP-AMPA receptor-dependent form of LTP in the hippocampus [60].
Taken together, the obtained results suggest that the regulatory mechanisms that determine the prevalence of a particular AMPA receptor subtype may be differentially affected by epileptiform activity, depending on the age of the subject and the seizure model. Our study describes a new form of plasticity in synapses of the entorhinal cortex and hippocampus induced by FSs, which is a rapid decrease in the proportion of CP-AMPA receptors. We believe that this form of plasticity may be one of the compensatory mechanisms that provides a decrease in the level of excitation in neuronal circuits and prevents epileptogenesis.
4. Materials and Methods
4.1. Animals
All experiments were conducted in accordance with local guidelines for the treatment of laboratory animals and were approved by the Ethics Committee of the Sechenov Institute of Evolutionary Physiology and Biochemistry. These recommendations are in full compliance with Russian and international standards for animal research. Wistar rat females and pups were maintained under standard conditions at room temperature with free access to water and food. Only 10 pups were allowed to remain in the litter.
4.2. FS Model
FSs were induced on P10 as previously described [36,61]. Briefly, pups were placed at the bottom of a 10-liter glass chamber for 30 min in which the temperature was maintained at 46°C. The rectal temperature of the animals was measured periodically during the experiments. If the temperature rose above 41°C, the pups were removed from the chamber and placed on a cool surface for 2 min. Behavioral seizures in this paradigm were stereotyped and consisted of heat-induced hyperkinetic arrest followed by facial automatisms, often accompanied by body flexion, followed by myoclonic jerks and clonic convulsions.
After hyperthermia, animals were placed on a cold surface until core temperature returned to the normal range and then returned to the home cage. Only animals with FSs lasting at least 15 min (N = 30 rats), corresponding to complex FSs [62], were included in the study. The littermates used as controls were removed from the cage for this period but kept at room temperature (N = 33 rats).
4.3. Brain Slice Preparation
On the day of testing, rats were killed by decapitation, and the brain was rapidly excised and placed in cold (4°C) oxygenated (95% O2/5% CO2) artificial cerebrospinal fluid (ACSF) containing (in mM): 126 NaCl, 24 NaHCO3, 2.5 KCl, 2CaCl2, 1.25 NaH2PO4, 1MgSO4, and 10 glucose. Slice preparation was performed as previously described [63]. Briefly, a vibratome HM 650V (Microm International, Walldorf, Germany) was used to prepare horizontal brain slices (350 μm) containing the dorsal hippocampus and the entorhinal cortex (EC). Before electrophysiological experiments, the slices were allowed to recover in oxygenated ACSF at 35°C for 1 hour.
4.4. Field Potential Recordings
After incubation, slices were placed in a recording chamber and perfused with a constant flow of ACSF at a rate of 6-7 ml/min at a temperature of 30°C. ACSF-filled glass microelectrodes (0.2–1.0 MΩ) were used to record fEPSPs from the CA1 stratum radiatum. Orthodromic electrical stimulation with a twisted bipolar nichromic stimulation electrode of increasing intensity (25–200 mA) via an A365 stimulus isolator (WPI, USA) was used to evoke synaptic responses. Evoked potentials were recorded using a model 1800 amplifier (A-M Systems, Carlsborg, WA, USA) and digitized and recorded on a personal computer using an NI USB-6211 A/D converter (National Instruments, Austin, Texas, USA) and WinWCP v5 software (University of Strathclyde, UK). Clampfit 10.2 software (Axon Instruments, USA) was used to analyze the electrophysiological data. The amplitudes of fEPSP and FVs were measured. The maximum slope of the input-output (I/O) relationship (fEPSP amplitude vs. FV amplitude) was then calculated for each slice by fitting a sigmoidal Gompertz function as previously described [64].
4.5. Whole-Cell Patch-Clamp Recordings from Entorhinal Cortex Pyramidal Neuron Slices
To visualize the principal neurons of the deep layers of the medial entorhinal cortex, a Nikon FN-1 microscope (Nikon, Tokyo, Japan) with differential interference contrast optics and a video camera (Grasshopper 3 GS3-U3-23S6M-C; FLIR Integrated Imaging Solutions Inc., Wilsonville, OR, USA) were used. Borosilicate glass pipettes (3–5 MΩ) were made using an aP-1000 pipette puller (Sutter Instrument, Novato, CA, USA). The pipetting solution was composed as follows: Cs-methanesulfonate-124 mM, NaCl-10 mM, HEPES-10 mM, EGTA-5 mM, QX314-6 mM, MK-801-3 mM, ATP-Mg-4 mM, GTP-0.3 mM, and spermine-0.2 mM (pH was adjusted to 7.25 by adding CsOH).
The MultiClamp 700B (Molecular Devices, Sunnyvale, CA, USA) patch-clamp amplifier, NI USB-6343 A/D converter (National Instruments, Austin, TX, USA) controlled by WinWCP 5 software (University of Strathclyde, Glasgow, UK) were used for whole-cell patch-clamp recordings. Data were lowpass filtered at 5kHz and digitized at 40kHz. The access resistance was typically 10-20 MΩ. It remained stable (≤30% increase) throughout the experiment in all cells included in the analysis. The liquid junction potential was compensated off-line by subtraction of 7 mV from the value of the holding voltage.
Synaptic responses were evoked by the stimulating bipolar electrode placed at a distance of 50-150 µm from the recorded neuron. AMPA receptor-mediated eEPSCs were recorded in the presence of NMDA and GABAa receptor antagonists (D-AP5, 50 μM; MK-801, 10 μM; gabazine, 10 μM) in the perfusion solution. The proportion of CP-AMPA receptors at the synapses of principal neurons was assessed by applying the selective antagonist of these receptors (IEM-1460, 100 μM) and by calculating the rectification index of eEPSCs. The rectification index was calculated as the ratio of eEPSC amplitudes recorded at +40 mV and at -40 mV.
4.6. Patch-Clamp Recordings of Membrane Currents from Isolated CA1 Pyramidal Neurons
Transverse hippocampal slices (250 µm thick) were prepared using a vibratome (Campden Instruments Ltd, Loughborough, UK) and stored in a solution containing (in mM) 124 NaCl, 5 KCl, 1.3 CaCl2, 2.0 MgSO4, 26 NaHCO3, 1.24 NaH2PO4, and 10 D-glucose, aerated with carbogen (95% O2, 5% CO2). Single pyramidal neurons were isolated from slices by vibrodissociation. Membrane currents generated in response to applications of 100 μM kainate at a holding voltage of -80 mV were recorded using the whole-cell patch-clamp technique. During the experiments, a series resistance of approximately 20 MΩ was 70-80% compensated. The currents were recorded using an EPC-8 amplifier (HEKA Electronics, Lambrecht, Germany). They were filtered at 5 kHz, sampled, and stored on a personal computer. Drugs were applied with the RSC-200 perfusion system (BioLogic Science Instruments, Claix, France). The system was computer controlled. Solution exchange time in whole-cell mode was 50-60 ms. The extracellular solution contained (in mM): 143 NaCl, 5 KCl, 2 MgSO4, 2.5 CaCl2, 18 D-glucose, and 10 HEPES (pH was adjusted to 7.4 with HCl). The pipette solution contained (in mM): 100 CsF, 40 CsCl, 5 NaCl, 0.5 CaCl2, 5 EGTA, and 10 HEPES (pH was adjusted to 7.2 with CsOH). The experiments were carried out at room temperature (22-24°C).
4.7. The Kainate-Induced Cobalt Uptake Method
Horizontal brain slices (400 μm) were cut with a HM 650 V vibratome (Microm, Walldorf, Germany) as described above. Sections were left to recover for 1 h at 28°C in ACSF containing TTX (0.5 μM). All subsequent incubations were performed at 20°C. Slices were transferred for 15 min to low-sodium, low-calcium Krebs' solution (in mM: 50 NaCl, 2.5 KCl, 26 NaHCO3, 1.25 NaH2PO4, 25 glucose, 0.5 CaCl2, 2 MgCl2) containing TTX (0.5 μM) and D-AP5 (100 μM). As a control, some slices were incubated with the added AMPA receptor antagonist DNQX (10 μM).
The slices were then stimulated with kainate (20 μM) for 20 min in Krebs' solution containing CoCl2 (1.5 mM). The slices were then washed for 10 min in Krebs' solution without divalent ions (in mM: 135 sucrose, 50 NaCl, 2.5 KCl, 26 NaHCO3, 1.25 NaH2PO4, 25 glucose) containing EDTA (0.5 mM) and then washed for 5 min in Krebs' solution without divalent ions. The slices were then incubated in Krebs' solution without divalent ions containing 0.12% (NH4)2S for 5 min to precipitate intracellular Co2+. The slices were then washed in Krebs' solution without divalent ions for 5 min and fixed in 4% paraformaldehyde (PFA) in 0.1 M phosphate-buffered saline (PBS) overnight at 4°C.
The next day, the slices were washed in 0.1 M PBS. They were then incubated in 30% sucrose in 0.1 M PBS. Slices were embedded in O.C.T. compound (Sakura Finetek USA), frozen in cooled (<-50°C) isopentane (78-78-4, Isopentane Solution, Sigma-Aldrich, St. Louis, MO, USA), and stored at -80°C. Specimens were sectioned at 20 μm thickness on a Bright OTF5000 cryostat (Bright Instrument Co Ltd, Huntingdon, UK) and mounted on Super Frost Plus-coated microscope slides (J1800AMNZ, Fisher Scientific UK Ltd, Loughborough, UK). For silver enhancement, sections were incubated in 2% Na2WO4 for 10 min and then in developer solution [8 parts AgNO3 solution: 1% Triton X-100, 7.5% CH3COOH, 30.3 mM CH3COONa, 2.94 mM AgNO3; 1 part 5% Na2WO4; 1 part 0.25% ascorbic acid] for 15 min in the dark and washed in 2% Na2WO4, dehydrated in increasing concentrations of ethanol solutions. Finally, the sections were cleared with microclearing (Diapath, Martinengo, Italy) and coverslipped with VitroGel (Ergo Production, Moscow, Russia).
Micrographs of stained hippocampal neurons in CA1, CA3, and entorhinal cortex layers from Co2+ uptake experiments were obtained using a Zeiss microscope (Carl Zeiss AG, Göttingen, Germany) at ×400 magnification.
4.8. Statistical Analysis
Statistical analysis and graphing of results were performed using OriginPro 8 (OriginLab Corporation, Northampton, MA, USA) and SigmaPlot 12.5 (Systat Software Inc., Palo Alto, CA, USA). Dixon's Q test (at 90% confidence level) was used to identify and reject outliers. Normality of the sample data was assessed using the Kolmogorov-Smirnov test. Statistical significance was assessed using Student's t-test and ANOVA, as described in the text. All data are presented as mean ± standard error of the mean. p < 0.05 was considered statistically significant.
Author Contributions
Formal analysis, Tatyana Postnikova, Alexandra Griflyuk, Arseniy Zhigulin, Oleg Barygin, Dmitry Amakhin,; Funding acquisition, Tatyana Postnikova; Investigation, Tatyana Postnikova, Alexandra Griflyuk, Arseniy Zhigulin, Elena Soboleva; Methodology, Tatyana Postnikova, Alexandra Griflyuk, Oleg Barygin, Dmitry Amakhin, Aleksey Zaitsev; Validation, Tatyana Postnikova and Aleksey Zaitsev; Writing – original draft, Tatyana Postnikova, Alexandra Griflyuk, Arseniy Zhigulin, Elena Soboleva, Oleg Barygin, Dmitry Amakhin, Aleksey Zaitsev; Writing – review & editing, Tatyana Postnikova and Aleksey Zaitsev.
Funding
This research was funded by the Russian Science Foundation, grant number 23-25-00143.
Institutional Review Board Statement
"The study was conducted according to the EU Directive 2010/63/EU for animal experiments and approved by the Ethics Committee of the Sechenov Institute of Evolutionary Physiology and Biochemistry of the Russian Academy of Sciences (Ethical permit number 13-k-a, 15 February 2018).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Silverstein, F.S.; Jensen, F.E. Neonatal Seizures. Ann Neurol 2007, 62, 112–120. [Google Scholar] [CrossRef]
- Mewasingh, L.D.; Chin, R.F.M.; Scott, R.C. Current Understanding of Febrile Seizures and Their Long-Term Outcomes. Dev Med Child Neurol 2020, 62, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Chen, Z. Generation of Febrile Seizures and Subsequent Epileptogenesis. Neurosci Bull 2016, 32, 481–492. [Google Scholar] [CrossRef]
- Waruiru, C.; Appleton, R. Febrile Seizures: An Update. Arch Dis Child 2004, 89, 751–756. [Google Scholar] [CrossRef]
- Cendes, F. Febrile Seizures and Mesial Temporal Sclerosis. Curr Opin Neurol 2004, 17, 161–164. [Google Scholar] [CrossRef]
- Lewis, D. V; Shinnar, S.; Hesdorffer, D.C.; Bagiella, E.; Bello, J.A.; Chan, S.; Xu, Y.; MacFall, J.; Gomes, W.A.; Moshé, S.L.; et al. Hippocampal Sclerosis after Febrile Status Epilepticus: The FEBSTAT Study. Ann Neurol 2014, 75, 178–185. [Google Scholar] [CrossRef]
- Brewster, A.; Bender, R.A.; Chen, Y.; Dube, C.; Eghbal-Ahmadi, M.; Baram, T.Z. Developmental Febrile Seizures Modulate Hippocampal Gene Expression of Hyperpolarization-Activated Channels in an Isoform- and Cell-Specific Manner. J Neurosci 2002, 22, 4591–4599. [Google Scholar] [CrossRef]
- Dubé, C.M.; Ravizza, T.; Hamamura, M.; Zha, Q.; Keebaugh, A.; Fok, K.; Andres, A.L.; Nalcioglu, O.; Obenaus, A.; Vezzani, A.; et al. Epileptogenesis Provoked by Prolonged Experimental Febrile Seizures: Mechanisms and Biomarkers. J Neurosci 2010, 30, 7484–7494. [Google Scholar] [CrossRef]
- Rakhade, S.N.; Zhou, C.; Aujla, P.K.; Fishman, R.; Sucher, N.J.; Jensen, F.E. Early Alterations of AMPA Receptors Mediate Synaptic Potentiation Induced by Neonatal Seizures. The Journal of Neuroscience 2008, 28, 7979–7990. [Google Scholar] [CrossRef]
- Chen, K.; Baram, T.Z.; Soltesz, I. Febrile Seizures in the Developing Brain Result in Persistent Modification of Neuronal Excitability in Limbic Circuits. Nat Med 1999, 5, 888–894. [Google Scholar] [CrossRef]
- Cornejo, B.J.; Mesches, M.H.; Benke, T.A. A Single Early-Life Seizure Impairs Short-Term Memory but Does Not Alter Spatial Learning, Recognition Memory, or Anxiety. Epilepsy and Behavior 2008, 13, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, B.J.; Mesches, M.H.; Coultrap, S.; Browning, M.D.; Benke, T.A. A Single Episode of Neonatal Seizures Permanently Alters Glutamatergic Synapses. Ann Neurol 2007, 61, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Egbenya, D.L.; Hussain, S.; Lai, Y.-C.; Xia, J.; Anderson, A.E.; Davanger, S. Changes in Synaptic AMPA Receptor Concentration and Composition in Chronic Temporal Lobe Epilepsy. Mol Cell Neurosci 2018, 92, 93–103. [Google Scholar] [CrossRef]
- Ekonomou, A.; Smith, A.L.; Angelatou, F. Changes in AMPA Receptor Binding and Subunit Messenger RNA Expression in Hippocampus and Cortex in the Pentylenetetrazole-Induced ‘Kindling’ Model of Epilepsy. Molecular Brain Research 2001, 95, 27–35. [Google Scholar] [CrossRef]
- Cruz Del Angel, Y.; Orfila, J.E.; Herson, P.S.; Brooks-Kayal, A.; González, M.I. Down-Regulation of AMPA Receptors and Long-Term Potentiation during Early Epileptogenesis. Epilepsy & Behavior 2021, 124, 108320. [Google Scholar] [CrossRef]
- Mathern, G.W.; Pretorius, J.K.; Kornblum, H.I.; Mendoza, D.; Lozada, A.; Leite, J.P.; Chimelli, L.M.C.; Fried, I.; Sakamoto, A.C.; Assirati, J.A.; et al. Human Hippocampal AMPA and NMDA MRNA Levels in Temporal Lobe Epilepsy Patients. Brain 1997, 120, 1937–1959. [Google Scholar] [CrossRef]
- Lippman-Bell, J.J.; Zhou, C.; Sun, H.; Feske, J.S.; Jensen, F.E. Early-Life Seizures Alter Synaptic Calcium-Permeable AMPA Receptor Function and Plasticity. Molecular and Cellular Neuroscience 2016, 76, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Shi, Y.; Jackson, A.C.; Bjorgan, K.; During, M.J.; Sprengel, R.; Seeburg, P.H.; Nicoll, R.A. Subunit Composition of Synaptic AMPA Receptors Revealed by a Single-Cell Genetic Approach. Neuron 2009, 62, 254–268. [Google Scholar] [CrossRef]
- Dingledine, R.; Borges, K.; Bowie, D.; Traynelis, S.F. The Glutamate Receptor Ion Channels. Pharmacol Rev 1999, 51, 7–61. [Google Scholar]
- Henley, J.M.; Wilkinson, K.A. Synaptic AMPA Receptor Composition in Development, Plasticity and Disease. Nat Rev Neurosci 2016, 17, 337–350. [Google Scholar] [CrossRef]
- Isaac, J.T.R.; Ashby, M.C.; McBain, C.J. The Role of the GluR2 Subunit in AMPA Receptor Function and Synaptic Plasticity. Neuron 2007, 54, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.K. Selective Reduction of GluR2 Protein in Adult Hippocampal CA3 Neurons Following Status Epilepticus but Prior to Cell Loss. Hippocampus 1998, 8, 511–525. [Google Scholar] [CrossRef]
- Grooms, S.Y. Status Epilepticus Decreases Glutamate Receptor 2 MRNA and Protein Expression in Hippocampal Pyramidal Cells before Neuronal Death. Proceedings of the National Academy of Sciences 2000, 97, 3631–3636. [Google Scholar] [CrossRef]
- Malkin, S.L.; Amakhin, D. V; Veniaminova, E.A.; Kim, K.K.; Zubareva, O.E.; Magazanik, L.G.; Zaitsev, A. V Changes of AMPA Receptor Properties in the Neocortex and Hippocampus Following Pilocarpine-Induced Status Epilepticus in Rats. Neuroscience 2016, 327, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.M.; Koh, S.; Rio, C.; Wang, C.; Lamperti, E.D.; Sharma, D.; Corfas, G.; Jensen, F.E. Decreased Glutamate Receptor 2 Expression and Enhanced Epileptogenesis in Immature Rat Hippocampus after Perinatal Hypoxia-Induced Seizures. The Journal of Neuroscience 2001, 21, 8154–8163. [Google Scholar] [CrossRef]
- Sommer, C.; Roth, S.U.; Kiessling, M. Kainate-Induced Epilepsy Alters Protein Expression of AMPA Receptor Subunits GluR1, GluR2 and AMPA Receptor Binding Protein in the Rat Hippocampus. Acta Neuropathol 2001, 101, 460–468. [Google Scholar] [CrossRef]
- Amakhin, D. V.; Soboleva, E.B.; Ergina, J.L.; Malkin, S.L.; Chizhov, A. V.; Zaitsev, A. V. Seizure-Induced Potentiation of AMPA Receptor-Mediated Synaptic Transmission in the Entorhinal Cortex. Front Cell Neurosci 2018, 12, 486. [Google Scholar] [CrossRef]
- Pellegrini-Giampietro, D.E.; Gorter, J.A.; Bennett, M. V; Zukin, R.S. The GluR2 (GluR-B) Hypothesis: Ca(2+)-Permeable AMPA Receptors in Neurological Disorders. Trends Neurosci 1997, 20, 464–470. [Google Scholar] [CrossRef]
- Sogawa, Y.; Monokoshi, M.; Silveira, D.C.; Ho Cha, B.; Roberta Cilio, M.; McCabe, B.K.; Liu, X.; Hu, Y.; Holmes, G.L. Timing of Cognitive Deficits Following Neonatal Seizures: Relationship to Histological Changes in the Hippocampus. Developmental Brain Research 2001, 131, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Jiang, L.; Chen, H.; Zhang, X.P. Expression of AMPA Receptor Subunits in Hippocampus after Status Convulsion. Child’s Nervous System 2012, 28, 911–918. [Google Scholar] [CrossRef]
- Russo, I.; Bonini, D.; Via, L. La; Barlati, S.; Barbon, A. AMPA Receptor Properties Are Modulated in the Early Stages Following Pilocarpine-Induced Status Epilepticus. Neuromolecular Med 2013, 15, 324–338. [Google Scholar] [CrossRef]
- Pandey, S.P.; Rai, R.; Gaur, P.; Prasad, S. Development- and Age-Related Alterations in the Expression of AMPA Receptor Subunit GluR2 and Its Trafficking Proteins in the Hippocampus of Male Mouse Brain. Biogerontology 2015, 16, 317–328. [Google Scholar] [CrossRef]
- Kumar, S.S.; Bacci, A.; Kharazia, V.; Huguenard, J.R. A Developmental Switch of AMPA Receptor Subunits in Neocortical Pyramidal Neurons. Journal of Neuroscience 2002, 22, 3005–3015. [Google Scholar] [CrossRef] [PubMed]
- Toth, Z.; Yan, X.-X.X.; Haftoglou, S.; Ribak, C.E.; Baram, T.Z. Seizure-Induced Neuronal Injury: Vulnerability to Febrile Seizures in an Immature Rat Model. The Journal of Neuroscience 1998, 18, 4285–4294. [Google Scholar] [CrossRef]
- Tanabe, T.; Hara, K.; Shimakawa, S.; Fukui, M.; Tamai, H. Hippocampal Damage after Prolonged Febrile Seizure: One Case in a Consecutive Prospective Series. Epilepsia 2011, 52, 837–840. [Google Scholar] [CrossRef]
- Postnikova, T.Y.; Griflyuk, A. V.; Amakhin, D. V.; Kovalenko, A.A.; Soboleva, E.B.; Zubareva, O.E.; Zaitsev, A. V. Early Life Febrile Seizures Impair Hippocampal Synaptic Plasticity in Young Rats. Int J Mol Sci 2021, 22, 8218. [Google Scholar] [CrossRef] [PubMed]
- Zubareva, O.E.; Kovalenko, A.A.A.; Kalemenev, S. V.; Schwarz, A.P.; Karyakin, V.B.; Zaitsev, A. V. Alterations in MRNA Expression of Glutamate Receptor Subunits and Excitatory Amino Acid Transporters Following Pilocarpine-Induced Seizures in Rats. Neurosci Lett 2018, 686, 94–100. [Google Scholar] [CrossRef]
- Rajasekaran, K.; Todorovic, M.; Kapur, J. Calcium-Permeable AMPA Receptors Are Expressed in a Rodent Model of Status Epilepticus. Ann Neurol 2012, 72, 91–102. [Google Scholar] [CrossRef]
- Joshi, S.; Rajasekaran, K.; Sun, H.; Williamson, J.; Kapur, J. Enhanced AMPA Receptor-Mediated Neurotransmission on CA1 Pyramidal Neurons during Status Epilepticus. Neurobiol Dis 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Lippman-Bell, J.J.; Rakhade, S.N.; Klein, P.M.; Obeid, M.; Jackson, M.C.; Joseph, A.; Jensen, F.E. AMPA Receptor Antagonist NBQX Attenuates Later-Life Epileptic Seizures and Autistic-like Social Deficits Following Neonatal Seizures. Epilepsia 2013, 54, 1922–1932. [Google Scholar] [CrossRef]
- Talos, D.M.; Sun, H.; Zhou, X.; Fitzgerald, E.C.; Jackson, M.C.; Klein, P.M.; Lan, V.J.; Joseph, A.; Jensen, F.E. The Interaction between Early Life Epilepsy and Autistic-Like Behavioral Consequences: A Role for the Mammalian Target of Rapamycin (MTOR) Pathway. PLoS One 2012, 7, e35885. [Google Scholar] [CrossRef]
- Zaitsev, А. V.; Amakhin, D. V.; Dyomina, A. V.; Zakharova, M. V.; Ergina, J.L.; Postnikova, T.Y.; Diespirov, G.P.; Magazanik, L.G. Synaptic Dysfunction in Epilepsy. J Evol Biochem Physiol 2021, 57, 542–563. [Google Scholar] [CrossRef]
- Roberts, N.S.; Handy, M.J.; Ito, Y.; Hashimoto, K.; Jensen, F.E.; Talos, D.M. Anti-Seizure Efficacy of Perampanel in Two Established Rodent Models of Early-Life Epilepsy. Epilepsy and Behavior 2023, 143. [Google Scholar] [CrossRef] [PubMed]
- Amakhin, D. V.; Soboleva, E.B.; Chizhov, A. V.; Zaitsev, A. V. Insertion of Calcium-Permeable AMPA Receptors during Epileptiform Activity In Vitro Modulates Excitability of Principal Neurons in the Rat Entorhinal Cortex. Int J Mol Sci 2021, 22, 12174. [Google Scholar] [CrossRef]
- Joshi, S.; Kapur, J. Mechanisms of Status Epilepticus: α-Amino-3-Hydroxy-5-Methyl-4-Isoxazolepropionic Acid Receptor Hypothesis. Epilepsia 2018, 59, 71–81. [Google Scholar] [CrossRef]
- Adotevi, N.; Lewczuk, E.; Sun, H.; Joshi, S.; Dabrowska, N.; Shan, S.; Williamson, J.; Kapur, J. α-Amino-3-Hydroxy-5-Methyl-4-Isoxazolepropionic Acid Receptor Plasticity Sustains Severe, Fatal Status Epilepticus. Ann Neurol 2020, 87, 84–96. [Google Scholar] [CrossRef]
- Ben-Ari, Y. Epilepsies and Neuronal Plasticity: For Better or for Worse? Dialogues Clin Neurosci 2008, 10, 17–27. [Google Scholar] [CrossRef]
- Abegg, M.H.; Savic, N.; Ehrengruber, M.U.; McKinney, R.A.; Gähwiler, B.H. Epileptiform Activity in Rat Hippocampus Strengthens Excitatory Synapses. Journal of Physiology 2004, 554, 439–448. [Google Scholar] [CrossRef]
- Debanne, D.; Thompson, S.M.; Gähwiler, B.H. A Brief Period of Epileptiform Activity Strengthens Excitatory Synapses in the Rat Hippocampus in Vitro. Epilepsia 2006, 47, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, T.Y.; Amakhin, D. V.; Trofimova, A.M.; Zaitsev, A. V. Calcium-Permeable AMPA Receptors Are Essential to the Synaptic Plasticity Induced by Epileptiform Activity in Rat Hippocampal Slices. Biochem Biophys Res Commun 2020, 529, 1145–1150. [Google Scholar] [CrossRef]
- Plant, K.; Pelkey, K.A.; Bortolotto, Z.A.; Morita, D.; Terashima, A.; McBain, C.J.; Collingridge, G.L.; Isaac, J.T.R. Transient Incorporation of Native GluR2-Lacking AMPA Receptors during Hippocampal Long-Term Potentiation. Nat Neurosci 2006, 9, 602–604. [Google Scholar] [CrossRef]
- Park, M. AMPA Receptor Trafficking for Postsynaptic Potentiation. Front Cell Neurosci 2018, 12, 361. [Google Scholar] [CrossRef]
- Zhou, C.; Bell, J.J.L.; Sun, H.; Jensen, F.E. Hypoxia-Induced Neonatal Seizures Diminish Silent Synapses and Long-Term Potentiation in Hippocampal CA1 Neurons. Journal of Neuroscience 2011, 31, 18211–18222. [Google Scholar] [CrossRef]
- Lippman-Bell, J.J.; Handy, M.; Nieder, C.G.; Getzfread, M.; Jensen, F.E. Altered Hippocampal Dendritic Spine Maturation after Hypoxia-Induced Seizures in Neonatal Rats. Molecular and Cellular Neuroscience 2021, 113. [Google Scholar] [CrossRef]
- Asrar, S.; Zhou, Z.; Ren, W.; Jia, Z. Ca2+ Permeable AMPA Receptor Induced Long-Term Potentiation Requires PI3/MAP Kinases but Not Ca/CaMDependent Kinase II. PLoS One 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, D.M.; Lamsa, K.P. Long-Term Synaptic Plasticity in Hippocampal Interneurons. Nat Rev Neurosci 2007, 8, 687–699. [Google Scholar] [CrossRef]
- Liu, S.J.; Cull-Candy, S.G. Activity-Dependent Change in AMPA Receptor Properties in Cerebellar Stellate Cells. Journal of Neuroscience 2002, 22, 3881–3889. [Google Scholar] [CrossRef]
- Liu, S.J.; Cull-Candy, S.G. Subunit Interaction with PICK and GRIP Controls Ca2+ Permeability of AMPARs at Cerebellar Synapses. Nat Neurosci 2005, 8, 768–775. [Google Scholar] [CrossRef]
- Liu, S.-Q.J.; Cull-Candy, S.G. Synaptic Activity at Calcium-Permeable AMPA Receptors Induces a Switch in Receptor Subtype. Nature 2000, 405, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, G.; Jo, J.; Hogg, E.L.; Piers, T.; Kim, D.H.; Seaton, G.; Seok, H.; Bru-Mercier, G.; Son, G.H.; Regan, P.; et al. Acute Stress Causes Rapid Synaptic Insertion of Ca2+ -Permeable AMPA Receptors to Facilitate Long-Term Potentiation in the Hippocampus. Brain 2013, 136, 3753–3765. [Google Scholar] [CrossRef] [PubMed]
- Griflyuk, A. V.; Postnikova, T.Y.; Zaitsev, A. V. Prolonged Febrile Seizures Impair Synaptic Plasticity and Alter Developmental Pattern of Glial Fibrillary Acidic Protein (GFAP)-Immunoreactive Astrocytes in the Hippocampus of Young Rats. Int J Mol Sci 2022, 23, 12224. [Google Scholar] [CrossRef] [PubMed]
- Notenboom, R.G.E.; Ramakers, G.M.J.J.; Kamal, A.; Spruijt, B.M.; De Graan, P.N.E. Long-Lasting Modulation of Synaptic Plasticity in Rat Hippocampus after Early-Life Complex Febrile Seizures. European Journal of Neuroscience 2010, 32, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Postnikova, T.Y.; Amakhin, D. V.; Trofimova, A.M.; Smolensky, I. V.; Zaitsev, A. V. Changes in Functional Properties of Rat Hippocampal Neurons Following Pentylenetetrazole-Induced Status Epilepticus. Neuroscience 2019, 399, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Zhuravin, I.A.; Dubrovskaya, N.M.; Vasilev, D.S.; Postnikova, T.Y.; Zaitsev, A. V. Prenatal Hypoxia Produces Memory Deficits Associated with Impairment of Long-Term Synaptic Plasticity in Young Rats. Neurobiol Learn Mem 2019, 164, 107066. [Google Scholar] [CrossRef]
Figure 1.
Stimulus-response relationships for fEPSP amplitudes (a, d) and presynaptic fiber volley (FV) amplitudes (e, f) recorded from hippocampal CA1 area 15 min (a-d) and 48 h (e-h) after FSs. According to repeated measures ANOVA, the magnitude of fEPSP amplitude (a, e) in rat hippocampal slices did not differ from that in control animals either 15 min (F7,294 = 0.22, p = 0.98) or 48 h (F7,266 = 0.96, p = 0.46) after FSs. FV amplitudes did not change significantly either 15 min (F7,294 = 0.43; p = 0.88) or 48 h (F7,266 = 0.14, p = 0.99) after FSs. Each point represents the mean ± standard error of the mean. (c, g) Representative examples of I/O relationships between the fEPSP and FV amplitudes in hippocampal slices. The maximum I/O slope is significantly reduced 15 min (d), but not at 48 h (h) after FS; t-test: * - р<0.05. Ctrl – control rat, FS – post-FS rats.
Figure 1.
Stimulus-response relationships for fEPSP amplitudes (a, d) and presynaptic fiber volley (FV) amplitudes (e, f) recorded from hippocampal CA1 area 15 min (a-d) and 48 h (e-h) after FSs. According to repeated measures ANOVA, the magnitude of fEPSP amplitude (a, e) in rat hippocampal slices did not differ from that in control animals either 15 min (F7,294 = 0.22, p = 0.98) or 48 h (F7,266 = 0.96, p = 0.46) after FSs. FV amplitudes did not change significantly either 15 min (F7,294 = 0.43; p = 0.88) or 48 h (F7,266 = 0.14, p = 0.99) after FSs. Each point represents the mean ± standard error of the mean. (c, g) Representative examples of I/O relationships between the fEPSP and FV amplitudes in hippocampal slices. The maximum I/O slope is significantly reduced 15 min (d), but not at 48 h (h) after FS; t-test: * - р<0.05. Ctrl – control rat, FS – post-FS rats.
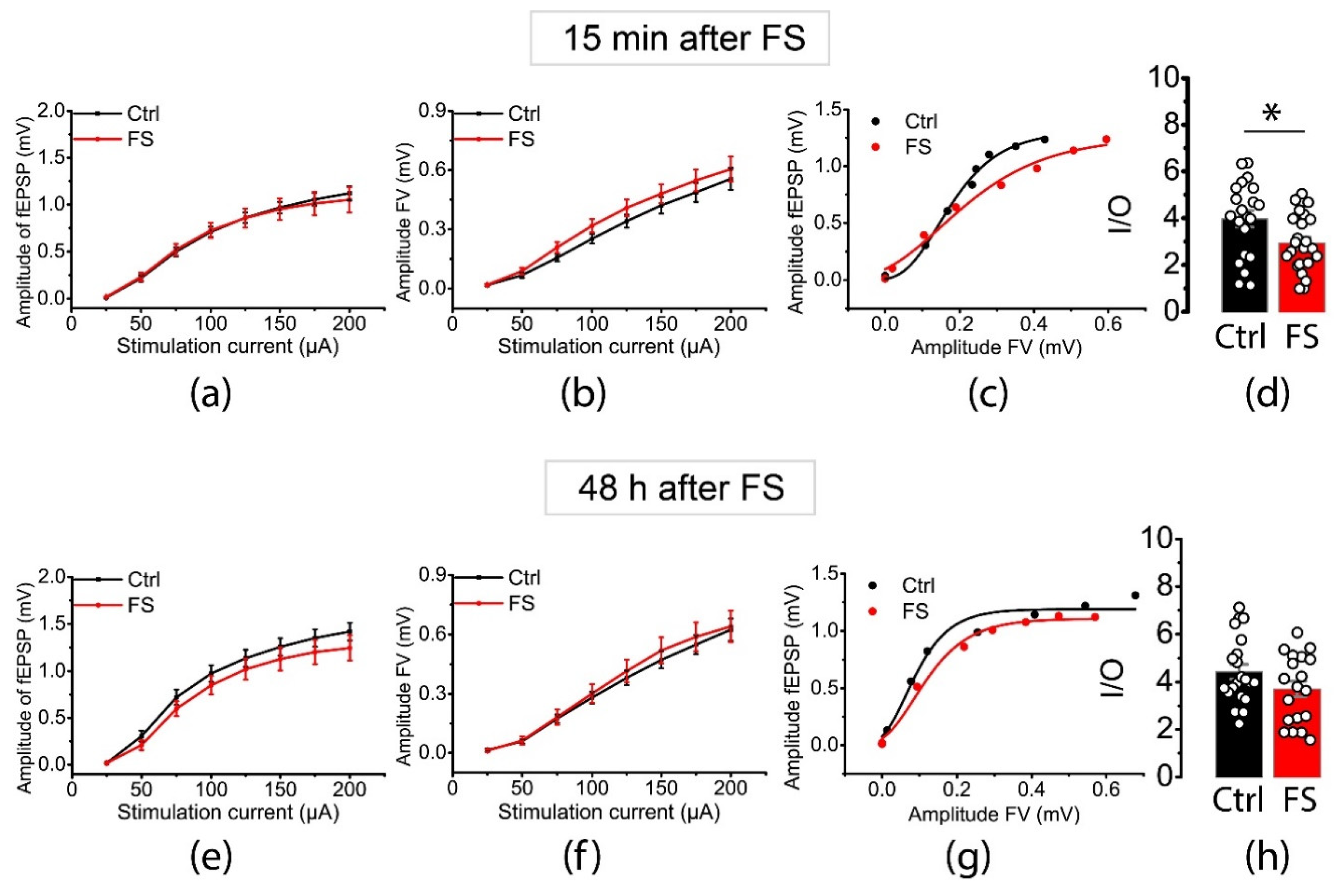
Figure 2.
Altered expression of CP-AMPA receptors in the postsynaptic membrane of entorhinal cortex pyramidal neurons after FSs. (a) Representative examples of AMPA receptor-mediated eEPSCs recorded at –40 and +40 mV in P10 rats. Note that in the control recordings, the negative current has a higher absolute amplitude than the positive current, which is not the case in the recordings from the post-FS rats. (b) Representative examples illustrating the blocking effect of IEM-1460 on eEPSCs in P10 rats. Note the reduced effect of the antagonist in post-FS rats. (c) The block of eEPSCs by IEM-1460 and (d) the rectification index of eEPSCs in P10 rats. IEM-1460 exerts a weaker block of eEPSCs in the post-FS group compared to age-matched controls (t-test, * p = 0.01, n = 14 and 16 for control and post-FS groups, respectively). The rectification index was greater in post-FS rats compared to controls (t-test, * p = 0.02, n = 15 and 19 for control and post-FS groups, respectively); (e) block of eEPSCs by IEM-1460 and (f) rectification index of eEPSCs in P12 rats. No significant differences were found between control and post-FS groups (n = 5 and 5 for (e); n = 12 and 13 for (f)).
Figure 2.
Altered expression of CP-AMPA receptors in the postsynaptic membrane of entorhinal cortex pyramidal neurons after FSs. (a) Representative examples of AMPA receptor-mediated eEPSCs recorded at –40 and +40 mV in P10 rats. Note that in the control recordings, the negative current has a higher absolute amplitude than the positive current, which is not the case in the recordings from the post-FS rats. (b) Representative examples illustrating the blocking effect of IEM-1460 on eEPSCs in P10 rats. Note the reduced effect of the antagonist in post-FS rats. (c) The block of eEPSCs by IEM-1460 and (d) the rectification index of eEPSCs in P10 rats. IEM-1460 exerts a weaker block of eEPSCs in the post-FS group compared to age-matched controls (t-test, * p = 0.01, n = 14 and 16 for control and post-FS groups, respectively). The rectification index was greater in post-FS rats compared to controls (t-test, * p = 0.02, n = 15 and 19 for control and post-FS groups, respectively); (e) block of eEPSCs by IEM-1460 and (f) rectification index of eEPSCs in P12 rats. No significant differences were found between control and post-FS groups (n = 5 and 5 for (e); n = 12 and 13 for (f)).
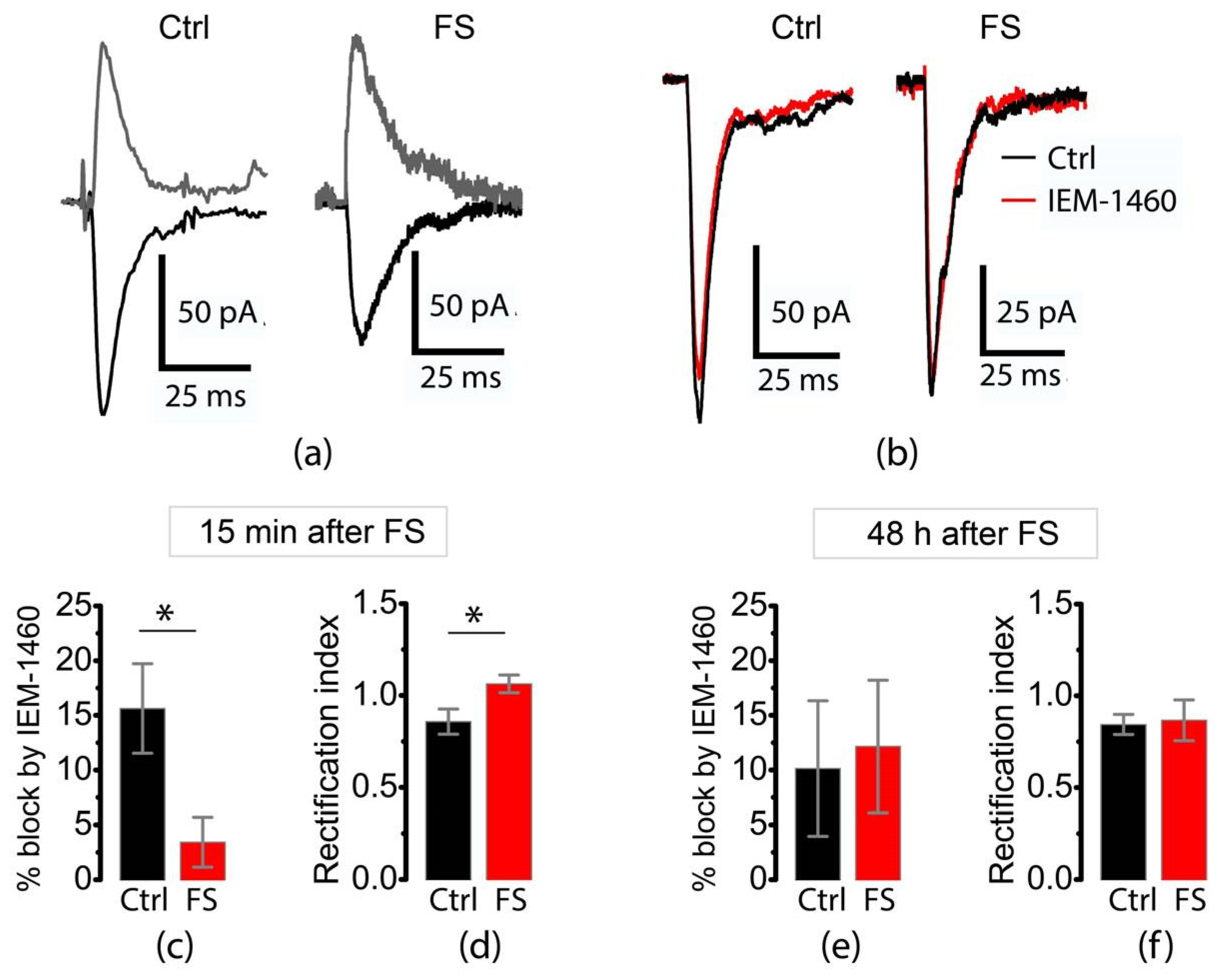
Figure 3.
Inhibition of kainite-induced currents in isolated pyramidal neurons from CA1 hippocampus by IEM-1460. Representative examples of inhibition of CP-AMPA receptors (a) 15 min and (с) 48 h after FSs. Mean percentage blockade by IEM-1460 (100 μM) of kainite-induced currents (b) 15 min and (d) 48 h after FS. Each point represents one value obtained for each neuron. All data are presented as mean ± standard error of the mean; t-test: ** p < 0.01.
Figure 3.
Inhibition of kainite-induced currents in isolated pyramidal neurons from CA1 hippocampus by IEM-1460. Representative examples of inhibition of CP-AMPA receptors (a) 15 min and (с) 48 h after FSs. Mean percentage blockade by IEM-1460 (100 μM) of kainite-induced currents (b) 15 min and (d) 48 h after FS. Each point represents one value obtained for each neuron. All data are presented as mean ± standard error of the mean; t-test: ** p < 0.01.
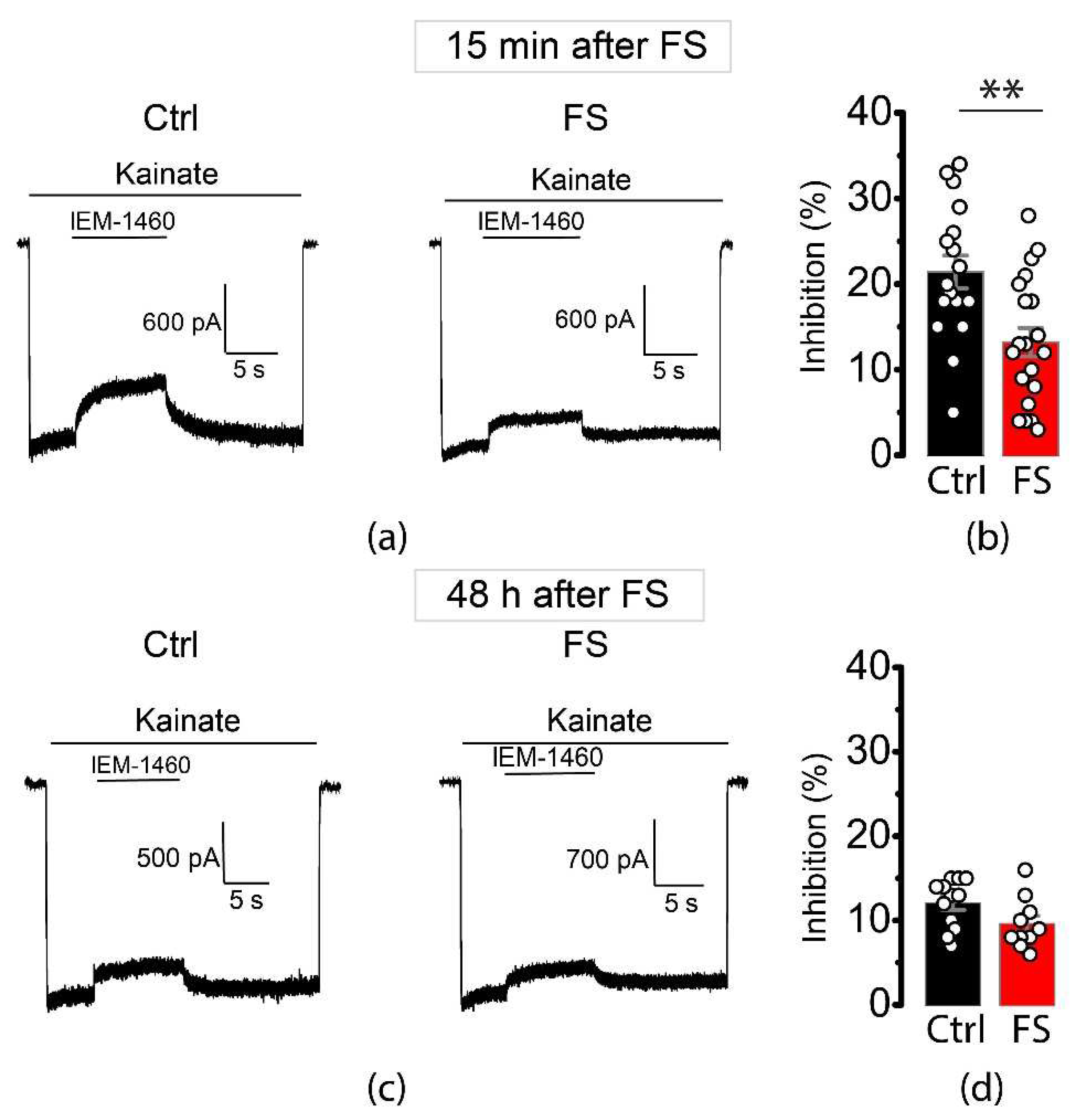
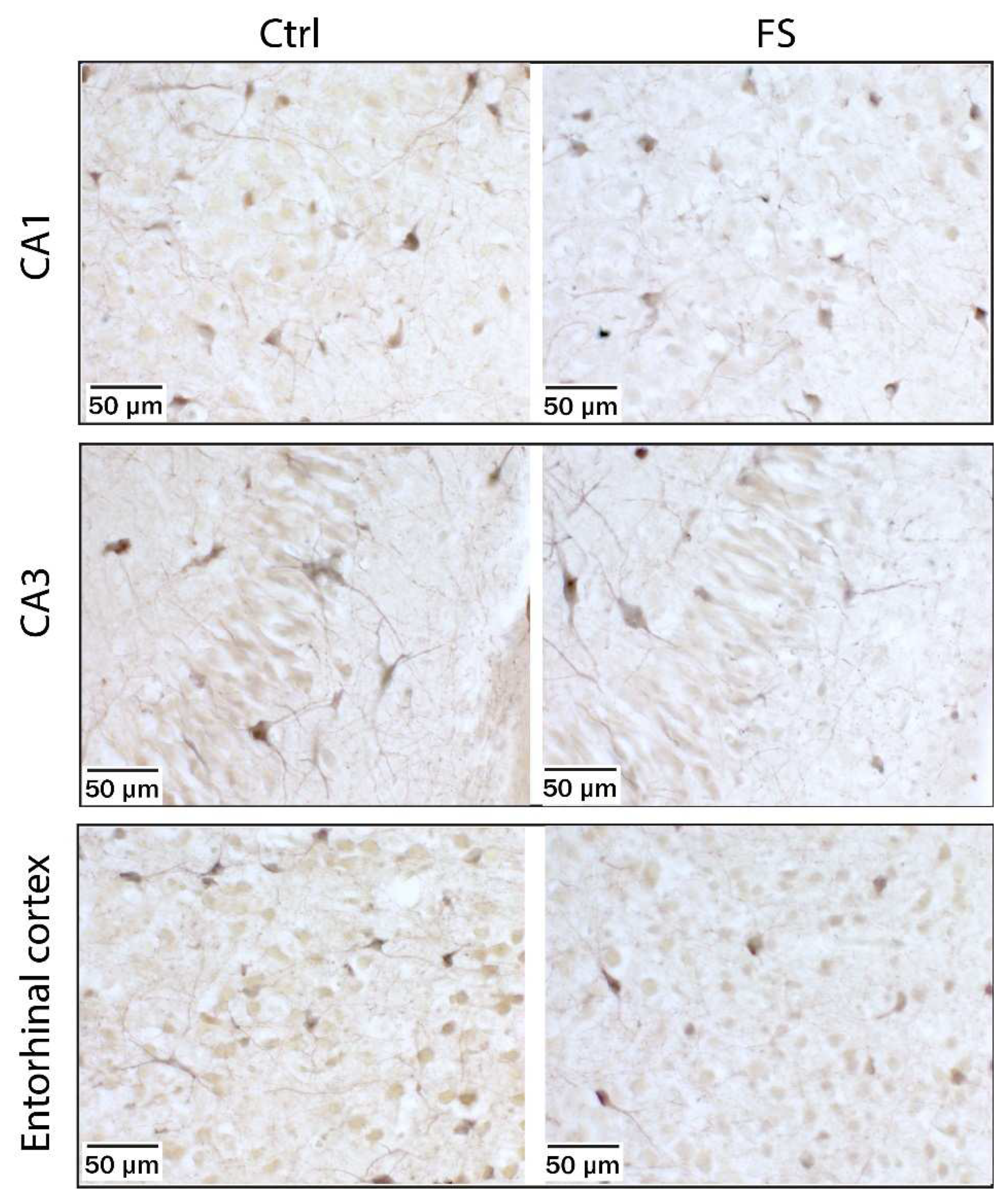
Figure 5.
Kainate-induced Co2+ uptake showed greater Co2+ uptake in hippocampal CA1 and CA3 cell layers and in the entorhinal cortex in acute brain slices from P10 control rats (left panels), and poorer Co2+ uptake in rats 15 min after FSs (middle panels). The AMPA and kainate receptor antagonist DNQX (20 μM) completely blocked Co2+ uptake in CA1, CA3 and entorhinal cortex of rats (right panels).
Figure 5.
Kainate-induced Co2+ uptake showed greater Co2+ uptake in hippocampal CA1 and CA3 cell layers and in the entorhinal cortex in acute brain slices from P10 control rats (left panels), and poorer Co2+ uptake in rats 15 min after FSs (middle panels). The AMPA and kainate receptor antagonist DNQX (20 μM) completely blocked Co2+ uptake in CA1, CA3 and entorhinal cortex of rats (right panels).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



