
Submitted:
10 July 2023
Posted:
12 July 2023
You are already at the latest version
Abstract
The ironstone outcrops in the rupestrian grasslands, known as cangas, are rich in species of endemic plants. This high endemism and the constant threat because of the area loss and fragmentation make the conservation of the rupestrian grasslands necessary. The rupestrian grasslands that host one of Brazil's largest iron-ore mining areas are Iron Quadrangle. About 89 rare and endangered species of the Iron Quadrangle are exclusive to cangas, including a threatened species Arthrocereus glaziovii. According to the International Union Conservation of Nature (IUCN), this species is considered micro-endemic and endangered and a priority for recovering the Iron Quadrangle's ferruginous rupestrian fields. This work aims to evaluate the potential distribution of A. glaziovii based on abiotic variables and soil elements and characterize the distribution of mining titles that may restrict the areas of occurrence of this species. We used five types of ecological niche model (ENM) algorithms (Bioclim, Domain, MaxEnt, GLM and RandomForest) in two sets of environmental variables for the modeling: 20 climatic variables and 18 edaphic variables. Our results showed that the area of the potential occurrence of A. glaziovii is more extensive and covers areas outside the Iron Quadrangle. However, mining titles currently licensed and in exploration correspond the entire identified area, threatening the species' survival.
Keywords:
Ironstone rupestrian grasslands
; Cactaceae
; geographic distribution
; conservation
; red list
1. Introduction
Campo rupestre is a plant physiognomy of mountain tops in eastern Brazil, characterized by a rich mosaic of herbaceous vegetation and sparse shrubs [1]. The region’s soil is compact, deficient in water and nutrients, and high in heavy metals [2,3]. The rocky outcrops in the rupestrian fields, particularly the iron ones (ferruginous outcrops), are rich in endemic plants and substantially contribute to the biodiversity of the domains in which they are inserted, Cerrado and Atlantic Forest [4,5]. With the region’s climate being considered as high altitude subtropical (dry winters and wet summers), according to the Köppen climate classification [6], the rocky fields present daily high thermal amplitudes (minimum of 14.1°C and maximum of 14.1°C). 23.7ºC) and low relative humidity [7,8]. Thus, soil, relief, and climate conditions are the main factors for establishing its unique and dominant vegetation cover [9].
The Cerrado and the Atlantic Forest are considered hotspots for preserving biodiversity as they harbor great biological diversity, which is highly threatened by human exploitation [10]. This constant threat to rupestrian grasslands, due to the loss and fragmentation of these ecosystems [11], and their high rates of endemism, makes their conservation extremely important, urgent, and necessary. In addition to the threats mentioned above, we find in the rupestrian fields one of Brazil’s largest iron ore extraction areas, the Quadrilátero Ferrífero (Q.F.), placing mining as the main factor of destruction in this region [12]. In the years 1960 to 2012, the Q.F. lost about 40% of its total area of cangas (about 100 ferruginous outcrops) to the extraction of iron ore [5,12]. Today, nearly 225 remaining ferruginous outcrops and 83% of these show signs of loss or degradation by mining [5].
The biodiversity of the Q.F. is currently under intense threat, as they make up one of the most important Brazilian mineral provinces in the world in iron ore [1,13], and the loss of species from mining activities is marked by a high irreversible environmental impact [14]. The intense mining activities practiced for centuries on a large scale continue to impact the flora of the rupestrian fields, as they resulted in areas with exposed subsoil or gravel, which, even after decades, no natural recovery was recorded [15]. Plants from these locations are susceptible to soil removal. They have a low rate of dispersal and slow growth [16], hindering the natural regeneration of these areas of altered rupestrian fields.
The recurrent mineral exploration in the Q.F. has caused severe impacts on the environment, biodiversity and ecosystem services [17,18]. About 89 rare and threatened species in the Q.F.. are plants with high conservation value as they occur exclusively in cangas in the region [12]. One of these species impacted on the mountain tops of rocky fields by mineral exploration is the microendemic and endangered cactus Arthrocereus glaziovii, which has suffered from reducing its area of occurrence [17,19].
The rupestrian fields make up an ecosystem that is clearly at risk, as they are highly endemic; studies are needed to elucidate the distribution of exclusive, threatened, and rare species for their conservation. The geographic distribution modeling, or niche model, is the most indicated tool to predict the potential distribution of species [20,21,22]. Thus, this work aims to investigate i) the potential distribution of A. glaziovii, based on abiotic variables and soil elements; and ii) characterize the distribution of mineral titles that may restrict the areas of occurrence of A. glaziovii.
2. Materials and Methods
2.1. Study species
The Cactaceae family is Neotropical, with distribution almost exclusively on the American continent, and one of its centers of diversity is Brazil [23]. The genus Arthrocereus stands out, comprising six species, all endemic to Brazil. They occur in the shrub and sub-shrub form and have particular habitats. Consequently, they are restricted to localities [24].
Arthrocereus glaziovii (K.Schum.) N.P.Taylor & Zappi is among the three taxa of this genus present in southeastern Brazil and stands out for a rare columnar-looking cactus. The species is considered microendemic, as it has a small number of populations restricted to the ferruginous outcrops of rupestrian fields [25]. This cactus is considered threatened with extinction, according to the International Union for the Conservation of Nature (IUCN) [26], due to anthropogenic activities, which cause the suppression of its habitat by removing soil for mining [8]. The species is considered a priority in the recovery of the ferruginous rocky fields of the Q.F., as it is unique to the region [27].
2.2. Study area
Considering that the microendemic distribution of the species only occurs in places rich in iron ore, the study region is concentrated in the Espinhaço Range, where the Q.F. is located. This set of mountain ranges is located in the Brazilian Central Plateau, with vegetation predominantly of rupestrian fields, is one of the most important mountain ranges in the country, and is home to one of the greatest floristic diversity in South America [16,28,29,30].
In the large iron ore deposits of the mountain ranges that delimit the Q.F., this is where the ferruginous outcrops, known as cangas, are located [17]. The cangas are composed of quartizitic, granitic and hematitic outcrops that occur interspersed, forming different microhabitats, where A. glaziovii occurs [8]) (Figure 1).
This region supplies the demand for iron ore in Brazil and abroad. The annual Brazilian iron ore production is approximately 400 million tons, and more than half of this production is concentrated in the Q.F. [31]. In 2019, the iron ore production of the largest Brazilian mining company, and third in the world, located in the state of Minas Gerais (M.G.), was 301.9 million tons [32].
2.3. Species distribution modeling
To determine the ecological niche of A. glaziovii, we collected georeferenced specimen records in the Global Biodiversity Information Facility (GBIF) [33] and speciesLink [34] databases in the herbarium from the Federal University of Minas Gerais (UFMG), and in literature data [35]. Records containing wrong geographic coordinates, no coordinates, or incomplete data were excluded from our analysis.
We used ecological niche modeling (ENM) to model the potential distribution of A. glaziovii in the region of the state of M.G., which comprises a large part of the Espinhaço Range, including the Q.F. For the modeling, we use five types of ENM algorithms. It is essential to use several algorithms to ensure that the prediction is as accurate as possible. Each method has a different predictive ability, different degrees of complexity, and stability, generating different responses for each variable. The algorithms were applied to all compiled georeferenced record points (Figure 2; Table S1).
2.4. Climatic and edaphic data
We have compiled two sets of environmental variables for the construction of ENMs: climatic and edaphic variables. For climate models, we used solar radiation and the 19 bioclimatic variables (Table S2) developed by [36] available on Worldclim 2020 [37]. These variables were constructed based on the spatial extrapolation of monthly averages of climate, temperature, and precipitation data collected from meteorological stations worldwide between the years 1970-2000. For the edaphic models we used 18 soil variables (Table S3) obtained from Instituto Brasileiro de Geografia e Estatística (IBGE) in [38]. Climatic data were acquired with a resolution of 30 seconds of arc (1 km² of spatial resolution), and edaphic data were obtained with a resolution of 2.5 minutes of arc (5 km² of spatial resolution). This difference in scale was why we did not perform an overlapping model of climatic and edaphic variables. These datasets were later cut to the Minas Gerais state.
To avoid redundancy in the variables, we performed a pair-by-pair Pearson correlation for each data set (climate and edaphic), available in the Raster package [36] of the R software [39]. Only one variable was selected in the case of 70% or more correlation to avoid the excess of autocorrelated variables, following Dormann et al. 2013. Were these chosen variables submitted to a selection random forest eliminates variables irrelevant to the data and orders the variables in order of importance. This selection was made using the VSURF package [40].
2.5. Modeling and Evaluation Procedures
To carry out the modeling, we chose five algorithms: Bioclim, Domain Generating Algorithm (DGA), General Linear Model (GLM), Maximum Entropy (MaxEnt), and Random Forest (R.F.), commonly used in ENMs analysis. All analyzes were performed using R version 3.5.3 software [39]. As we do not have actual absence points, we created a thousand pseudo-absence points for the studied species to be used in the GLM, MaxEnt, and R.F. models. After analyzing each current model generated by the algorithms, we created a mean between all models in each set (climate and edaphic). We used 80% of the points for the construction of the models, and 20% of the remaining points were used for model validation.
2.4. Mining titles
The layers in vector format of the mineral titles were obtained from Instituto Pristino’s Geoenvironmental Digital Atlas database [41]. We use 52 layers that correspond to areas that are already being mined and those that will be mined in the future.
3. Results
3.1. Selected climatic and edaphic variables
After performing Pearson’s correlation, six of the 20 initial climatic variables were selected (Figures S1 and S2) and submitted to VSURF. Among these six input climate variables, seasonal precipitation (BIO15), mean annual temperature (BIO1), annual precipitation (BIO12), temperature seasonality (BIO4), and annual temperature variation (BIO7) were the most influential according to VSURF importance values (Table 1), and solar radiation was not significant (SRAD).
Of the 18 initial soil variables, ten were selected by Pearson’s correlation (Figure S3) and also submitted to VSURF. Among these ten selected edaphic variables, silt, potassium (K), nitrogen (N), and hydrogen (H) were the four most influential, and only aluminum (Al) did not influence the model. After excluding the irrelevant variables, the others were classified in order of influence (Table 1).
3.2. Current potential environmental suitability for Arthrocereus glaziovii
All models had the area under the curve (AUC) > 0.98, showing its effectiveness. When considering each of the models individually for both climatic and edaphic variables, together, the most restrictive algorithms for the area were Bioclim and R.F., which identified a small space with environmental suitability located in the Q.F. (Figure 3a, d; Figure 4a, d). Domain and MaxEnt were the algorithms that presented a moderately wide distribution, identifying restricted distributions in the center, south, and southeast of Minas Gerais state and mountainous areas of the studied site (Figures 3b, e; Figure 4b, e). The algorithm with the broadest distribution was the GLM, which identified almost every area analyzed with a probability of environmental suitability for both variables (Figure 3c; Figure 4c).
According to the Bioclim climate model, the average annual temperature for the occurrence of A. glaziovii is very restricted (between 16 and 20˚C), annual precipitation ranging between 1500 and 1600 mm, seasonal precipitation between 84 and 86 mm, the temperature in the hottest month between 18.2 and 19.1 ˚C and the annual temperature variation between 1.7 and 1.8˚C. According to the Domain climate model, the restriction of species occurrence was similar, with the average annual temperature between 16 and 21˚C, the annual precipitation with a variation between 1450 and 1650 mm, the seasonal precipitation between 82 and 89 mm, the temperature in the hottest month between 17.5 and 20.1 ˚C and the annual temperature variation between 1.6 and 1.8˚C. For MaxEnt, the most critical climate variable for the niche was the mean annual temperature, followed by seasonal precipitation, annual precipitation, and annual temperature variation. For this model, the temperature of the warmest month was not significant. As for the R.F., the most essential variable for the niche was annual precipitation, followed by annual temperature variation, seasonal precipitation, average annual temperature, and temperature of the hottest month.
By Bioclim’s edaphic model, sand and clay showed high levels, from 41 to 53 and 25 to 27, respectively, then silt with 15 to 20, pH at 5, H from 3 to 4, and Mg 0 to 1. By the Domain model, the soil presented similar or close values, sand between 30 and 60, clay between 15 and 38, silt with 10 to 15, pH also in the value of 5, H with 1 to 6, and Mg again 0 a 1. Nitrogen and K had values equal to 0. For MaxEnt, the most critical edaphic variable for the niche was Mg (50%), followed by K, N, clay, and silt. As for R.F., the essential variable for the niche was the concentration of N, followed by silt, clay, K, and Mg. Sand, H, and water pH did not show significant importance in both models.
The average of all models showed a wide potential distribution of the species Figure 4f. According to the average cut-off of all environmental niche models, the potential distribution of A. glaziovii is currently located in the central-southeast and south of M.G., with a particular concentration in the Espinhaço Range (and Q.F.) and northern regions. South of the study area (Serra da Mantiqueira) and north (Figure 3g).
3.3. Mining titles
Overlapping all layers of mineral titles in the Q.F., we observe few remaining areas in this region (Figure 5). It is clear that the distribution of A. glaziovii, both actual and potential, is an area of important mining extraction. The distribution of future mines largely overlaps the occurrence and the suitable regions of the potential distribution of the species. Iron ore extraction is the most frequent, followed by gold ore.
4. Discussion
Our results demonstrate that the area of the potential occurrence of A. glaziovii considering both climatic and edaphic variables is more significant and covers areas outside the Q.F.. However, currently licensed and under exploration mining titles threaten the entire identified area, jeopardizing the species’ survival.
Seasonal and annual rainfall, warmest month temperature, and annual temperature were the abiotic factors determining the geographic distribution of A. glaziovii. Temperature and rainfall are important as they influence the length and intensity of the cacti’s reproductive period [42]. A. glaziovii has an annual flowering period lasting from 1 to 5 months, starting between the end of the dry period and the beginning of the rainy period [43,44]. The temperature has a positive influence on the production of flowers and fruits, but rainfall does not influence this factor [45]. The fruiting of the species takes place in the middle of the rainy season, and both temperature and rainfall positively influence this phenophase [45]. The increase and decrease in temperature influence the germination of the species, with an optimal temperature between 25 and 30ºC, even in the rainy season to establish seedlings [1,46].
The species A. glaziovii currently occurs in the Q.F. region in the center-southeast of the state of M.G., and the Domain and MaxEnt models indicated a potentially larger distribution area for this species in this same region. The Bioclim and R.F. models showed smaller occurrence than the current. The GLM indicated a wide distribution throughout the state of M.G. The cut of the averages of the models stated an occurrence area located to the south (climate model) and to the north of the state of M.G. The potential distribution models of A. glaziovii showed, therefore, that this species has a potential geographic distribution restricted to the central-southeast region of the state of M.G., with a concentration along the Espinhaço Range, to the south and north of the state of M.G.
The area south of M.G. state in the climate models is called Serra da Mantiqueira, and the indication in the model is probably due to being a region with high elevations. According to Koppen’s classification, the climate in this region is subtropical in altitude, equal to that of the species’ current occurrence area [47]. In Serra da Mantiqueira, the vegetation found is called campo de altitude and grows under granitic outcrops of igneous or metamorphic rocks, which differs from the physiognomies occurring on ferruginous outcrops of the Q.F. and quartzite fields of the Espinhaço Range [47]. Although both occur on rocks and have similar climates, they are of different lithotypes, explaining why the edaphic model did not point to this region [47].
Only in the MaxEnt model of climate variables a small area north of the state of M.G. was indicated as a possible area of environmental suitability for the occurrence of A. glaziovii. The north of M.G. is a transition region of Cerrado and Caatinga with hematite in the soil, with a humid tropical climate of savannah. The driest season coincides with winter, and rainfall is less than 60 mm [48]. The soil of this indicated region also presents high concentrations of Fe and the areas of occurrence of the species in the Q.F. However, this element was not analyzed in the models, as no studies prove its influence directly on the plant, indicating that the species has some mechanism of accumulation or tolerance to it [35].
The final indication of the area to the north in the edaphic models is due to other elements of its composition. For the species, Mg, K, N, silt, and clay were the most important elements in determining the distribution model of edaphic variables. K has a positive correlation in the development and growth of A. glaziovii and can promote its germination. At the same time, N in large concentrations decreases the germination rate of the species [35]. Sand and clay contents are directly related to the water retention capacity of canga soils [49]. The northern region soil has high Mg levels, oscillating between high and low K, which would go against the preference found in our results. However, it has high values of silt, with 50% (high) of its composition being clay and 20% (low) of sand, and the pH of the area is slightly acidic, between 5.8 and 6.6, which explains the indication of this area in the model [50].
The overlapping of existing mining titles in the Q.F. indicates that the remaining areas of canga are entirely compromised by mining, and mining tends always to increase and surpass its past productions. Mining has already been identified as the leading cause of the irreversible loss of 40% of canga areas in the last 40 years [12]. For the next 50 years, forecasts indicate a catastrophic scenario for cangas [51]. Climate change would result in losses of up to 82% and mining impacts in losses of up to 60% of the remaining areas of rupestrian fields.
Although abiotic factors indicate an expansion in the distribution of the species, mining activities compromise this possibility with habitat loss and degradation. Fragmented environments are more vulnerable to the adverse effects of impacts, and a large part of the current area of canga is constituted by canga fragmented by losses already suffered [51]. The areas of occurrence and potential distribution of A. glaziovii are of important mining extraction, around 102 cangas have mining activities close to them, and the indirect impacts of mining reach up to 5 km from the mines [12,52,53]. The expansion of the mining sector in rupestrian fields will continue to generate a domino effect of unprecedented impacts to this ecosystem and its endemic species [18,53]. Canga species threatened with extinction by the IUCN, such as A. glaziovii, could quickly disappear and become extinct if action is not taken to preserve them. The transmission of knowledge and awareness of the importance of these areas and their endemic species, and the magnitude of the losses already suffered by their misuse, is necessary for their effective conservation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Correlation of climate variables; Figure S2: Correlation of chosen climatic variables with radiation; Figure S3: Correlation of edaphic variables (soil); Table S1: Occurrence records of the specimens considered in the study; Table S2: Bioclimatic variables from the Worldclim database; Table S3: Edaphic variables (soil) from the IBGE database.
Funding
This research received no funding.
Acknowledgments
We are grateful to Glória Ramos Soares and Lucas Neves Perillo for their comments in the previous version.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheib, A.L. Ecologia da germinação e potencial para formação de banco de sementes de espécies de Arthrocereus A. Berger (Cactaceae) endêmicas dos campos rupestres de Minas Gerais, Brasil. Dissertação de Mestrado, Universidade Federal de Minas Gerais, Belo Horizonte, 2009.
- Giulietti, A.M.; Pirani, J.R.; Harley, R.M. Espinhaço range region, eastern Brazil. In: Centres of plant diversity: a guide and strategy for their conservation, 1st ed.; Davis, S.D.; Heywood, V.H.; Herrera-MacBryde, O.; Villa-Lobos, J.; Hamilton, A.C., Eds.; WWF/IUCN: Cambridge, UK, 1997; pp. 397–404.
- Porto, M. L.; Silva, M. F. F. Tipos de vegetação metalófila em áreas da Serra de Carajás e de Minas Gerais. Acta Botanica Brasilica 1989, 3, 13-21. [CrossRef]
- Jacobi, C.M.; Carmo, F.F. Diversidade dos campos rupestres ferruginosos no Quadrilátero Ferrífero, MG. Megadiversidade 2008, 4, 1-12.
- Jacobi C.M.; Carmo F.F. Plantas Vasculares sobre Cangas. In Diversidade Florística nas cangas do Quadrilátero Ferrífero, 1st ed.; Jacobi C.M., Carmo F.F., Eds.; Código Editora: Belo Horizonte, Brazil, 2012; pp. 31– 42.
- Alvares C.A., Stape J.L., Sentelhas P.C., Gonçalves J.L.M., Sparovek G. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift 2013, 22, 711-728. [CrossRef]
- Gianotti, A.R.C.; Souza, M.J.H.; Machado, L.M.; Pereira, I.M.; Vieira, A.D. Magalhães, M. R. Análise microclimática em duas fitofisionomias do cerrado no alto Vale do Jequitinhonha, Minas Gerais. Revista Brasileira de Meteorologia 2013, 28, 246-256. [CrossRef]
- Jacobi, C.M.; Carmo, F.F.; Vincent, R.C.; Stehmann, J. R. Plant communities on the ironstone outcrops - a diverse and endangered Brazilian ecosystem. Biodiversity and Conservation 2007, 16, 2185-2200. [CrossRef]
- Lima, L. R.; Pirani, J.R. O gênero Croton L. (Euphorbiaceae) na cadeia do espinhaço, Minas Gerais, Brasil. Bol Bot. 2003, 21, 299-344. [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853-858. [CrossRef]
- Oliveira, A.S.; Resende, M.G.; Silva, W.H.O. Análise da fragmentação florestal da região centro sul do estado de Minas Gerais através de estimativa de métricas de paisagem, Proceedings of the Congresso Nacional de Botânica, Belo Horizonte, Brazil, 2013; Belo Horizonte, Brazil, 2013, Abstract Number id6220.
- Carmo, F.F. Importância Ambiental e Estado de Conservação dos Ecossistemas de Cangas no Quadrilátero Ferrífero e Proposta de Áreas-Alvo para a Investigação e Proteção da Biodiversidade em Minas Gerais. Dissertação de Mestrado, Universidade Federal de Minas Gerais, Belo Horizonte, 2010.
- Salles, D.M.; Carmo, F.F.; Jacobi, C.M. Habitat loss challenges the conservation of endemic plants in mining-targeted Brazilian mountains. Environmental Conservation 2018, 46, 140-146. [CrossRef]
- Jacobi, C.M.; Carmo, F.F.; Campos, I.C. Soaring extinction threats to endemic plants in Brazilian metal-rich regions. AMBIO 2011, 40, 540-543. [CrossRef]
- Negreiros, D.; Fernandes, G.W; Silveira, F.A.O.; Chalub, C. Seedling growth and biomass allocation of endemic and threatened shrubs of rupestrian fields. Acta Oecologica 2009, 35, 301-310. [CrossRef]
- Silveira, F.A.O.; Negreiros, D.; Barbosa, N.P.U.; Buisson, E.; Carmo, F.F.; Carstensen, D.W.; Conceição, A.A.; Cornelissen, T.G.; Echternacht, L.; Fernandes, G.W.; Garcia, Q.S.; Guerra, T.J.; Jacobi, C.M.; Lemos-Filho, J.P.; Le Stradic, S.; Morellato, L.P.C.; Neves, F.S.; Oliveira, R.S.; Schaefer, C.E.; Viana, P.L.; Lambers, H. Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant and Soil 2016, 403, 129–152. [CrossRef]
- Jacobi, C.M.; Carmo, F.F. The contribution of ironstone outcrops to plant diversity in the Iron Quadrangle, a threatened Brazilian landscape. AMBIO 2008, 37, 324-326. [CrossRef]
- Fernandes, G.W.; Ribeiro, S.P. Deadly conflicts: mining, people, and conservation. Perspectives in Ecology and Conservation 2017, 15, 141–144. [CrossRef]
- Zappi, D.; Taylor, N.; Silva, S.R.; Machado, M.; Moraes, E.M.; Calvente, A.; Cruz, B.; Correia, D.; Larocca, J.; Assis, J.G.A.; Aona, L.; Menezes, M.O.T.; Meiado, M.; Marchi, M.N.; Santos, M.R.; Bellintani, M.; Coelho, P.; Nahoum, P.I.; Resende, S. Plano de ação nacional para a conservação das Cactaceas. 1st ed.; Instituto Chico Mendes de Conservação da Biodiversidade, ICMBio: Brasília, Brazil, 2011, pp. 1-58.
- Nabout, J.C.; Júnior, P.M.; Bini, L.M.; Diniz-Filho, J.A.F. Distribuição geográfica potencial de espécies americanas do caranguejo "violinista"; (Uca spp.) (Crustacea, Decapoda) com base em modelagem de nicho ecológico. Iheringia 2009, 92-98. [CrossRef]
- Sousa, N.P.R. Estrutura hierárquica na resposta das distribuições geográficas de plantas do Cerrado a mudanças climáticas. Dissertação de Mestrado, Universidade Federal de Goiás, Goiânia, 2011.
- Carvalho, M.C.; Gomide, L.R., Acerbi-Júnior, F.W.; Tng, D. Potential and future geographical distribution of Eremanthus erythropappus (DC.) MacLeish: a tree threatened by climate change. Floresta e Ambiente 2019, 26, e20180455. [CrossRef]
- REFLORA - Plantas do Brasil: resgate histórico e herbário virtual para o conhecimento e conservação da flora brasileira. Available online: http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB1417 (acessed in 21/06/2019).
- Gonzaga, D.R.; Souza, M.A.; Neto, L.M.; Peixoto, A.L.; Mendonça, C.B.F.; Gonçalves-Esteves, V. The systematic value of pollen morphology in Arthrocereus A. Berger (Cactaceae, Cactoideae). Review of Palaeobotany and Palynology 2019, 269, 33-41. [CrossRef]
- Zappi, D.C.; Taylor, N.P. Cactaceae. In Diversidade Florística nas Cangas do Quadrilátero Ferrífero, 1st ed.; Jacobi, C.M.; Carmo, F.F., Eds.; Código Editora: Belo Horizonte, Brazil, 2012.
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/search?query=Arthrocereus%20glaziovii&searchType=species (acessed in 22/07/2019).
- Skirycz, A.; Castilho A.; Chaparro, C.; Carvalho N.; Tzotzos G., Siqueira J.O. (2014). Canga biodiversity, a matter of mining. Frontiers in Plant Science 2014, 5, 1-9. [CrossRef]
- King, L.C. A geomorfologia do Brasil Oriental. Rio de Janeiro, Revista Brasileira de Geografia 1956, 18, 147-265.
- Stannard, B.L.; Harvey, Y.B; Harley, R.M. Flora of the Pico das Almas: Chapada Diamantina - Bahia, Brazil, 1st ed.; Royal Botanic Gardens: Kew, London, England, pp. 1-853.
- Giulietti, A.M.; Pirani, J.R.; Harley, R.M. Espinhaço range region, eastern Brazil. In Centres of plant diversity: a guide and strategy for their conservation, Davis, S.D.; Heywood, V.H., Eds.; Gland : IUCN / WWF: Cambridge, England, v.3, pp. 1-562.
- IBRAM - Instituto Brasileiro de Mineração. Minério de Ferro. In Informações Sobre Economia Mineral Brasileira, 1st ed.; Rodrigues, C.P., Costa, E.R., Eds.; Instituto Brasileiro de Mineração: Brasília, Brazil, 2015; pp. 22–25.
- Bianchetti, M. Vale contabiliza recuo de 41,2% na extração de minério em MG. In: Diário do comércio. Available online: https://diariodocomercio.com.br (acessed in 20/01/2021).
- Global Biodiversity Information Facility (GBIF). Available online: https://www.gbif.org/ (acessed in 13/01/2019).
- speciesLink. Available online: https://specieslink.net/ (acessed in 13/01/2019).
- Climaco, L.F.S. Variabilidade fenotípica da espécie microendêmica Arthrocereus glaziovii Zappy & Taylor (Cactaceae) em campos rupestres ferruginosos. Dissertação de Mestrado, Universidade Federal de Ouro Preto, Ouro Preto, 2017.
- Hijmans, R.J.; Etten, J. V. Raster: Geographic analysis and modeling with raster data. R package version 2.0-12. 2012. Available online: http://CRAN.R-project.org/package=raster (acessed in 20/12/2022).
- Worldclim 2020. Available online: http://www.worldclim.org (acessed in 15/05/2019).
- Arruda, D. M.; Fernandes-Filho, E. I.; Solar, R.R.; Schaefer, C. E. Combining climatic and soil properties better predicts covers of Brazilian biomes. The Science of Nature 2017, 104, 1-10. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. https://www.R-project.org/. 2017.
- Genuer, R.; Poggi, J. M.; Malot, T. M. VSURF: Variable Selection Using Random Forests. R package version 1.0.4. 2018. Available online: https://CRAN.R-project.org/package=VSURF (acessed in 20/12/2022).
- Instituto Pristino’s Geoenvironmental Digital Atlas database. Available online: https://www.institutopristino.org.br (acessed in 20/12/2019).
- Petit, S.The reproductive phenology of three sympatric species of columnar cacti on Curaçao. Journal of Arid Environments 2001, 49, 521–531. [CrossRef]
- Lima, A.L.A. Padrões fenológicos de espécies lenhosas e cactáceas em uma área do semi-árido do Nordeste do Brasil. Dissertação de Mestrado, Universidade Federal Rural de Pernambuco, Recife, 2007.
- Alencar, J.C.; Almeida, R.A.; Fernandes, N.P. Fenologia de espécies florestais em floresta tropical úmida de terra firme na Amazônia Central. Acta Amazônica 1979, 9, 163-198. [CrossRef]
- Oliveira, D.V. Aspectos da história de vida de Arthrocereus glaziovii (K.Schum.) N.P.Taylor & Zappi (Cactaceae), uma espécie endêmica do Quadrilátero Ferrífero, Minas Gerais, Brasil. Dissertação de Mestrado, Universidade Federal de Ouro Preto, Ouro Preto, 2017.
- Cheib, A.L.; Garcia, Q.S. Longevity and germination ecology of seeds of endemic Cactaceae species from high-altitude sites in southeastern Brazil. Seed Science Research 2012, 22, 45-53. [CrossRef]
- Gonçalves, L.N. Campos de altitude do maciço Marins-Itaguaré, Serra da Mantiqueira SP/MG: composição florística, fitogeografia e estrutura da vegetação. Dissertação de Mestrado. Universidade Federal de Juíz de Fora, Juíz de Fora, 2019.
- Sá Júnior, A. Aplicação da classificação de Köppen para o zoneamento climático do estado de Minas Gerais. Dissertação de Mestrado. Universidade Federal de Lavras, Lavras, 2009.
- Costa, S.A.D. Caracterização química, física, mineralógica e classificação de solos ricos em ferro do Quadrilátero Ferrífero. Dissertação de Mestrado (Solos e Nutrição de Plantas). Universidade Federal de Viçosa, Viçosa, 2003.
- Souza, V. N. R.; Neto, J. E. E.; Matrangolo, C. A. R.; Magalhães, W. T.; Fogaça, C. A.; Figueiredo, M. A. P.; Figueiredo, L. H. A. Caracterização de diferentes solos eutróficos na região norte de Minas Gerais. Revista Intercâmbio 2019, 15, 106-122. [CrossRef]
- Fernandes, G.W.; Barbosa, N.P.U.; Alberton, B.; Barbieri, A.; Dirzo, R.; Goulart, F.; Guerra, T.J.; Morellato, L.P.C.; Solar, R.R.C. The deadly route to collapse and the uncertain fate of Brazilian rupestrian grasslands. Biodiversity and Conservation 2018, 27, 2587-2603.
- Durán, A.P.; Rauch, J.; Gaston, K.J. Global spatial coincidence between protected areas and metal mining activities. Biological Conservation 2013, 160, 272-278. [CrossRef]
- Pena, J.C.C.; Goulart, F.; Fernandes, G.W.; Hoffmann, D.; Leite, F.S.F.; Santos, N.B.; Soares-Filho, B.; Sobral-Souza, T.; Vancine, M.H.; Rodrigues, M. Impacts of mining activities on the potential geographic distribution of eastern Brazil mountaintop endemic species. Perspectives in Ecology and Conservation 2017, 15, 172–178. [CrossRef]
Figure 1.
Regions of occurrence of cangas in the state of Minas Gerais. Source: Pristino Institue.
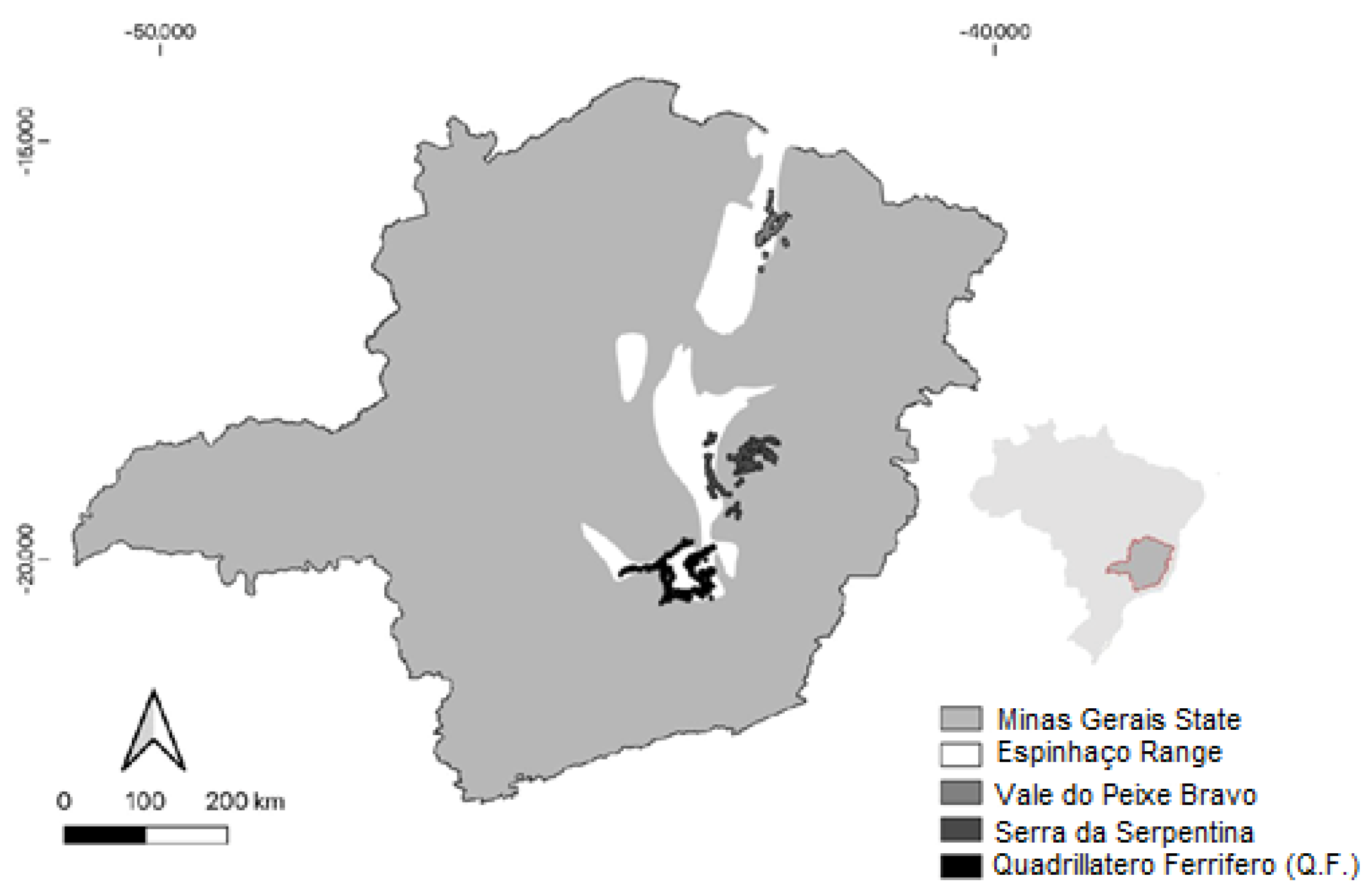
Figure 2.
Location of the studied species, Arthrocereus glaziovii (red circles indicate the occurrence locations used for ecological niche modeling).
Figure 2.
Location of the studied species, Arthrocereus glaziovii (red circles indicate the occurrence locations used for ecological niche modeling).
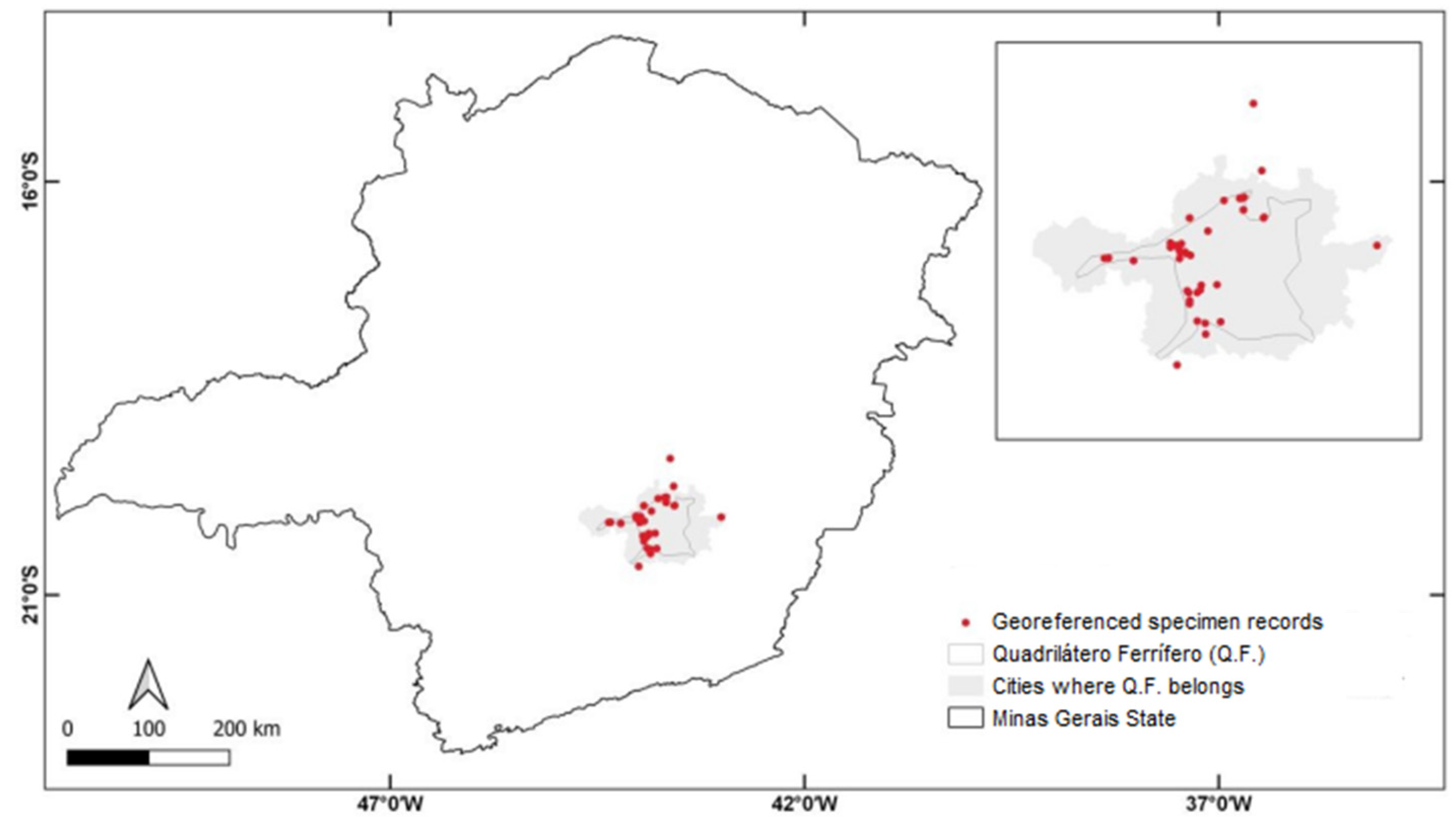
Figure 3.
Arthrocereus glaziovii prediction models for the climatic variables of the algorithms: (a) Bioclim, (b) Domain, (c) GLM, (d) R.F., (e) MaxEnt, (f) average between all algorithms, (g) potential geographic distribution of the species generated from the average of the algorithms.
Figure 3.
Arthrocereus glaziovii prediction models for the climatic variables of the algorithms: (a) Bioclim, (b) Domain, (c) GLM, (d) R.F., (e) MaxEnt, (f) average between all algorithms, (g) potential geographic distribution of the species generated from the average of the algorithms.
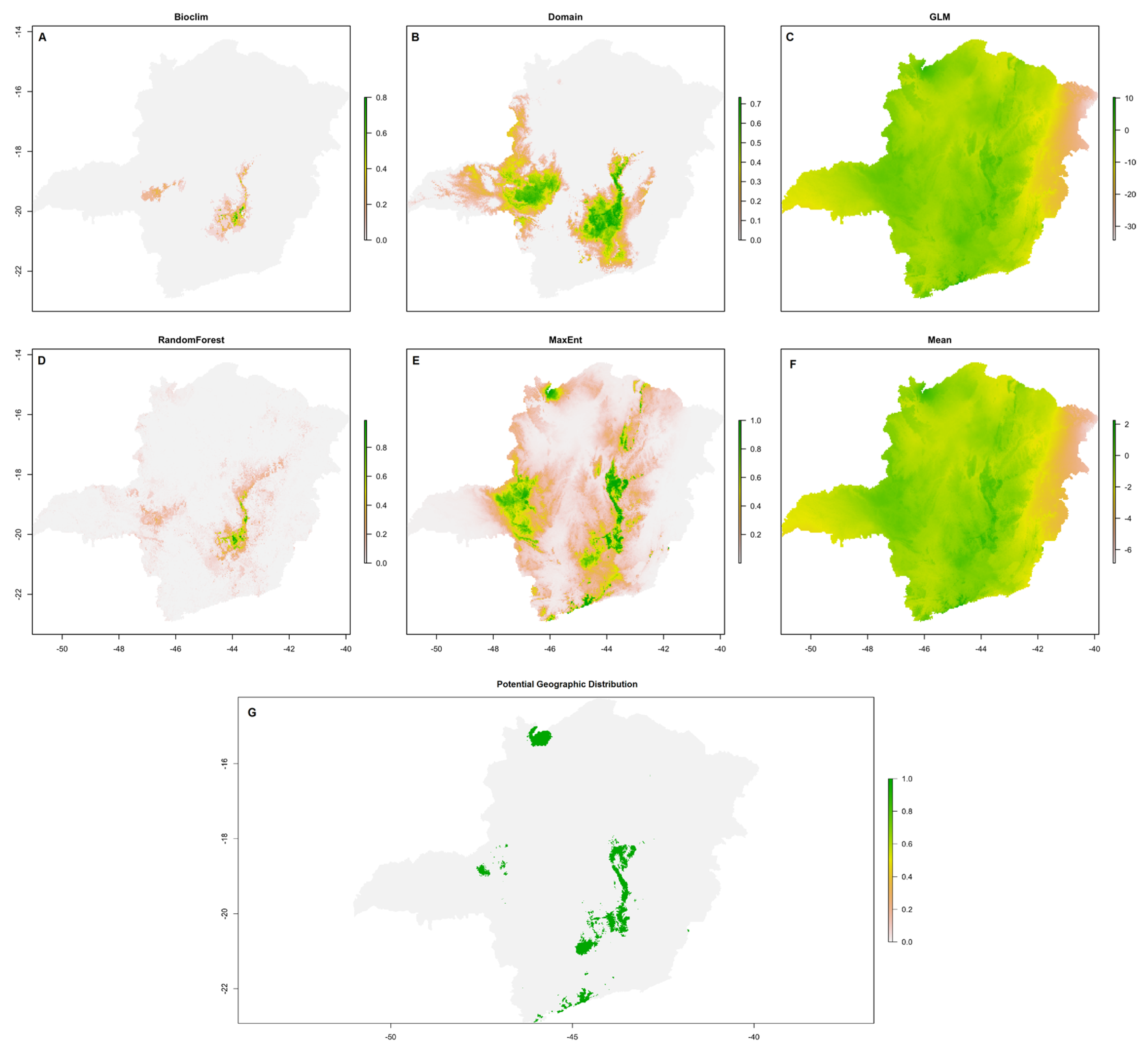
Figure 4.
Arthrocereus glaziovii prediction models for the edaphic variables of the algorithms: (a) Bioclim, (b) Domain, (c) GLM, (d) R.F., (e) MaxEnt, (f) average between all algorithms, (g) potential geographic distribution of the species generated from the average of the algorithms.
Figure 4.
Arthrocereus glaziovii prediction models for the edaphic variables of the algorithms: (a) Bioclim, (b) Domain, (c) GLM, (d) R.F., (e) MaxEnt, (f) average between all algorithms, (g) potential geographic distribution of the species generated from the average of the algorithms.
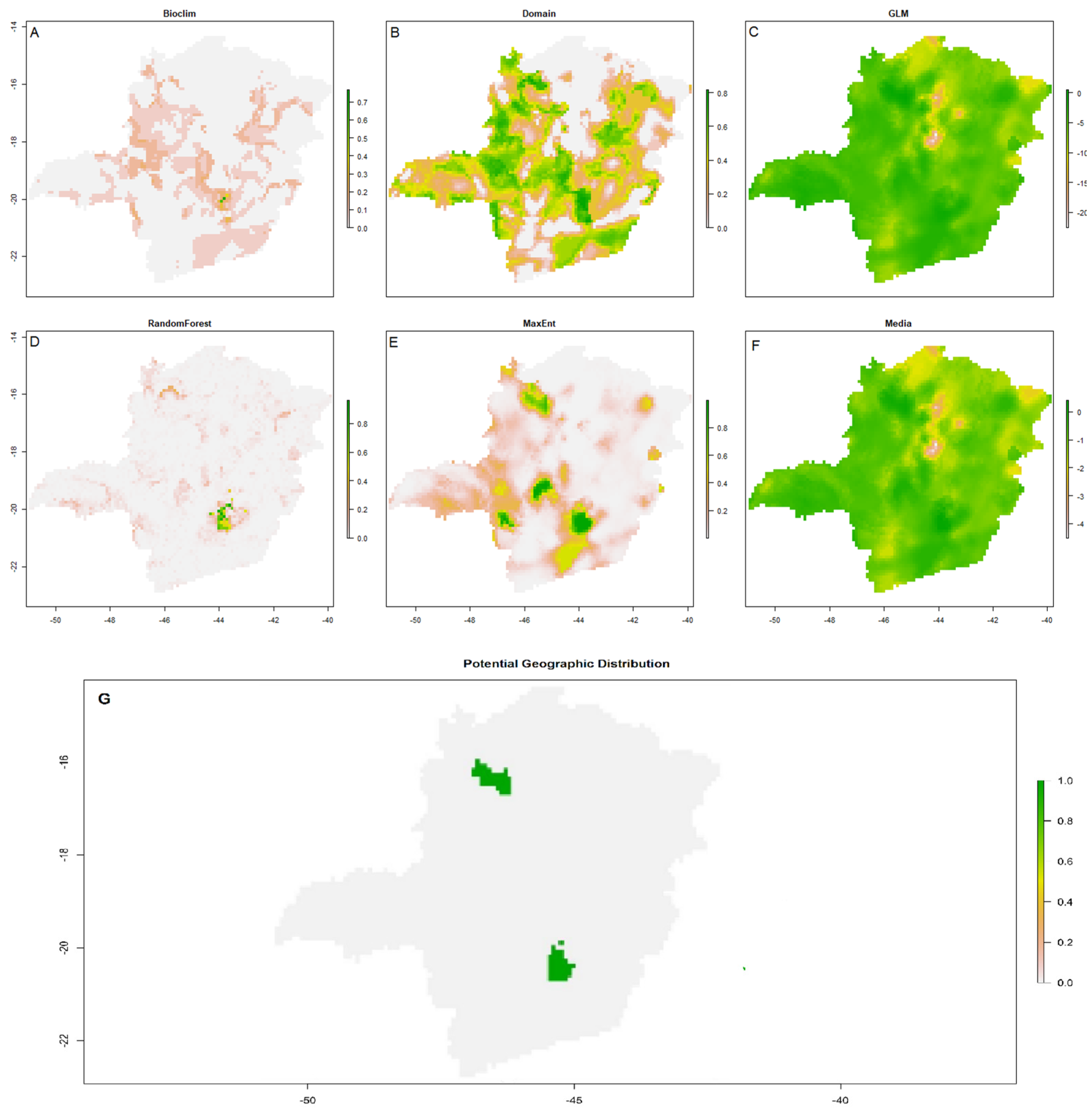
Figure 5.
Mining titles currently licensed in the Quadrilátero Ferrífero region. Source: Pristine Institute.
Figure 5.
Mining titles currently licensed in the Quadrilátero Ferrífero region. Source: Pristine Institute.

Table 1.
Climatic and edaphic variables used as predictors in ENM, in order of importance.
| Climatics1 | Edaphics2 |
|---|---|
| Seasonal Precipitation (BIO15) | Silt |
| Mean Annual Temperature (BIO1) | Potassium (K) |
| Annual Precipitation (BIO12) | Nitrogen (N) |
| Temperature Seasonality (BIO4) | Hydrogen (H) |
| Annual Temperature Variation (BIO7) | Clay |
| Sand | |
| Water pH | |
| Magnesium |
1 Wordclim. 2 IBGE.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



