
Submitted:
12 July 2023
Posted:
13 July 2023
You are already at the latest version
Abstract
Breast cancer patients with high levels of HER2 (human epidermal growth factor receptor 2) expression had worse clinical outcomes. Anti-HER2 monoclonal antibody (mAb) is the most important therapeutic modality for HER2-positive breast cancer. We previously immunized mice with the ectodomain of HER2 to create the anti-HER2 mAb, H2Mab-77 (mouse IgG1, kappa). This was then altered to produce H2Mab-77-mG2a-f, an afucosylated mouse IgG2a. In the present work, we examined the reactivity of H2Mab-77-mG2a-f and antitumor effects against breast cancers in vitro and in vivo. BT-474, an endogenously HER2-expressed breast cancer cell line, was identified by H2Mab-77-mG2a-f with a strong binding affinity [a dissociation constant (KD): 5.0 × 10-9 M]. H2Mab-77-mG2a-f could stain HER2 of breast cancer tissues in immunohistochemistry and detect HER2 protein in western blot analysis. Furthermore, H2Mab-77-mG2a-f demonstrated strong antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) for BT-474 cells. MDA-MB-468, a HER2-negative breast cancer cell line, was unaffected by H2Mab-77-mG2a-f. Additionally, in the BT-474-bearing tumor xenograft model, H2Mab-77-mG2a-f substantially suppressed tumor development when compared to the control mouse IgG2a mAb. In contrast, the HER2-negative MDA-MB-468-bearing tumor xenograft model showed no response to H2Mab-77-mG2a-f. These findings point to the possibility of H2Mab-77-mG2a-f as a treatment regimen by showing that it has antitumor effects on HER2-positive breast tumors.
Keywords:
HER2
; monoclonal antibody
; ADCC
; CDC
; antitumor activity
1. Introduction
The receptor tyrosine kinase human epidermal growth factor receptor 2 (HER2, also known as ERBB2) is a type I transmembrane glycoprotein that controls cell development and survival. HER2 can form homodimer and heterodimers with related family members including EGFR, HER3, and HER4. The complexes initiate intracellular signaling pathways like mitogen-activated protein kinase and phosphoinositide 3-kinase/Akt (PI3K/Akt). The extracellular domain of HER2 is composed of four regions, domain I to Ⅳ. Unlike other family receptors, HER2 extracellular domain basically has an active structure, dimer formation with other molecules is possible even without a ligand [1,2]. This structural characteristic is assumed to be the root of HER2’s lack of ligand basis [3]. It should be mentioned that HER2 and HER3 interactions create extremely potent mitogenic signals and have been implicated in cancer progression [4,5,6].
The most frequent kind of cancer among women is breast cancer [7]. 43,170 American women will pass away from breast cancer in 2023, according to 297,790 new cases [7]. 20% of breast cancer patients have an overexpression of HER2, which is linked to a poor prognosis [8,9,10,11]. Carcinogenic HER2 activation is caused by a change in the HER2 receptor gene, more HER2 overexpression and amplification, and less phosphatase activity [12]. Breast tumor incidence and development are correlated with HER2 gene amplification and protein overexpression; this was discovered in 1987 [13]. Angiogenesis, cell cycle progression, survival, angiogenesis, migration, invasion, and tumorigenesis are all accelerated by aberrant activation of HER2 [14,15,16,17]. Breast cancer brain metastases are among the cancer subtypes with an elevated risk of HER2-amplified tumors [18,19]. In order to forecast the effectiveness of anti-HER2 therapy, immunohistochemistry and/or in situ hybridization are often used to evaluate the HER2 status of breast cancer [20]. The development of HER2-targeted medicines has advanced significantly, and HER2 has grown in popularity as a cancer therapeutic target [21].
Trastuzumab emtansine (T-DM1) and trastuzumab deruxtecan (T-DXd) are antibody-drug conjugates (ADCs) in which the payload is attached to trastuzumab, a humanized anti-HER2 monoclonal antibody (mAb) [22,23,24]. Trastuzumab has been used to treat HER2-positive metastatic breast cancer. T-DM1 was authorized for HER2-positive advanced breast cancer by the US Food and Drug Administration (FDA) and the European Medicines Agency in 2013, while T-DXd was approved in the US in 2019 for HER2-positive metastatic or unresectable breast cancer [25,26,27]. Trastuzumab binds to HER2 domain IV, causing antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cell phagocytosis (ADCP), HER2 internalization-mediated destruction, and dimerization inhibition [28,29,30]. The extracellular domain of HER2 has been seen in breast cancer patients’ serum, but trastuzumab binding prevents this HER2 cleavage [31,32]. After connecting to the receptors, the ADCs are internalized by endocytosis, where they eventually release their payloads and cause cell damage. As a result, trastuzumab-based ADCs are potent weapons that combine the dual antitumor action of trastuzumab with a cytotoxic payload. T-DM1 has been shown to preserve trastuzumab effects such as blocking HER2 ectodomain cleavage, inhibiting PI3K/Akt signaling, and engaging immune cells through Fc-gamma (Fcγ) receptors, resulting in ADCC [33]. T-DXd is an ADC of the future that combines deruxtecan and trastuzumab. Due to the bystander impact of its potent payload on surrounding cells, T-DXd has been found to be effective not only on HER2-high tumor cells but also on HER2-low tumor cells [26,34]. T-DXd is being tested in clinical studies to see whether it can be used to treat HER2-positive gastric and non-small-cell lung cancer [35,36].
Targeting different HER2 epitopes may make it easier to stop HER2-dependent cell growth, which would stop the growth of tumors [37]. Pertuzumab, a therapeutic antibody authorized in 2012 for HER2-positive breast cancer, binds to the domain II of HER2, preventing receptor dimerization with partner receptors and related signal transduction [38,39,40]. Pertuzumab, although having ADCC efficacy equal to trastuzumab, does not decrease HER2 shedding [31,40,41]. A potential mechanism for resistance to anti-HER2 therapy includes alteration of the HER2 ectodomain, which results in a reduction of anti-HER2 antibody binding affinity [42]. Anti-HER2 treatments using distinct anti-HER2 mAb variants, such as binding epitopes, or in conjunction with chemotherapy might pave the way for the continued development of HER2-targeted cancer therapies.
We previously created an anti-HER2 mAb (clone H2Mab-77; mouse IgG1, kappa) by immunizing mice with the ectodomain of HER2 or with LN229 glioblastoma cells that overexpress HER2 [43]. Flow cytometry, western blotting, and immunohistochemical tests may all be performed with H2Mab-77. To test its ADCC, complement-dependent cytotoxicity (CDC), and antitumor efficacy in xenograft models, we also altered H2Mab-77 into a core-fucose-deleted and subclass-converted anti-HER2 mAb (H2Mab-77-mG2a-f). The activation of natural killer cells (NK cells), which is enhanced by the antibody’s Fc region binding to FcγRIIIa on NK cells, results in the destruction of target cells, including tumor cells, and facilitates ADCC [44]. A core-fucose deletion in the Fc N-glycan has been shown to improve the binding of Fc to FcγRIIIa on effector NK cells [45,46]. In this work, we assessed the antitumor efficacy of H2Mab-77-mG2a-f against breast cancer using in vitro and in vivo models, as well as its reactivities in flow cytometry, western blotting, and immunohistochemical investigations.
2. Results
2.1. Development of a Core-Fucose-Deficient Anti-HER2 mAb, H2Mab-77-mG2a-f
We previously created an anti-HER2 mAb, H2Mab-77 (mouse IgG1, kappa), by immunizing mice with either the HER2 ectodomain or LN229 glioblastoma cells overexpressing HER2. Flow cytometry, western blotting, and immunohistochemical tests revealed that H2Mab-77 has a high binding affinity.
In this study, we changed the antibody to improve its antitumor efficacy. We altered the subclass of H2Mab-77 from mouse IgG1 to mouse IgG2a by joining the VH and VL chains of H2Mab-77 with the CH and CL chains of mouse IgG2a, respectively, since mouse IgG1 has no ADCC and CDC activity. In addition, utilizing BINDS-09 cells (ExpiCHO-S cells lacking FUT8 fucosyltransferase), an afucosylated mouse IgG2a form of H2Mab-77 (H2Mab-77-mG2a-f) was generated (Figure 1).
2.2. Analysis of H2Mab-77-mG2a-f Reactivity against Breast Adenocarcinoma Cells by Flow Cytometry
In a previous study, the original H2Mab-77 (mouse IgG1, kappa) was found to be suitable for flow cytometry, western blotting, and immunohistochemistry [43]. In this study, an afucosylated form of the anti-HER2 mAb, H2Mab-77-mG2a-f, was generated. Similar to the original H2Mab-77, H2Mab-77-mG2a-f successfully detected exogenously overexpressed HER2 in LN229 (LN229/HER2 cells) at 1 or 10 µg/mL (Figure 2A,B).
Next, we conducted flow cytometric analysis using H2Mab-77-mG2a-f on two breast cancer cell lines, BT-474 and MDA-MB-468. BT-474 represents the luminal B type of breast cancer with positive expression of progesterone receptor (PR+), estrogen receptor (ER+), and HER2. In contrast, MDA-MB-468 is a triple-negative breast cancer lacking expression of PR, ER, and HER2 [47]. H2Mab-77-mG2a-f demonstrated recognition of endogenously expressed HER2 in BT-474 at 1 or 10 µg/mL, while it showed no response to MDA-MB-468 at either concentration (Figure 2C,D).
Subsequently, we evaluated the binding affinity of H2Mab-77-mG2a-f to LN229/HER2 cells and BT-474 cells using flow cytometry. The apparent dissociation constants (KD) of H2Mab-77-mG2a-f to LN229/HER2 and BT-474 cells were determined to be 3.4 × 10−9 M and 5.0 × 10−9 M, respectively (Figure 2E,F). These findings indicate that H2Mab-77-mG2a-f can effectively recognize HER2 in both cell types with a high binding affinity.
2.3. Western Blotting Using H2Mab-77-mG2a-f
Similar to the original H2Mab-77, H2Mab-77-mG2a-f detected the HER2 band with an estimated 180-kDa band in lysates generated from LN229, LN229/HER2, and BT-474 cells; no band was seen in MDA-MB-468 cells, which are HER2-negative (Figure 3A). For the internal control, an anti-IDH1 mAb (clone RcMab-1) was employed (Figure 3B). These findings revealed that H2Mab-77-mG2a-f may be employed in western blotting to assess HER2 expression in cultured cells, including breast cancer cells.
2.4. Immunohistochemical Analyses Using H2Mab-77-mG2a-f against Breast Cancer Tissue
The H2Mab-77-mG2a-f antibody was then utilized to target clinical specimens of human breast cancer tissue array via immunohistochemistry analysis. Similar to the original H2Mab-77, H2Mab-77-mG2a-f identified HER2 on the plasma membrane of breast cancer tissue (Figure 4). Table 1 summarizes the immunohistochemical study of breast cancer. H2Mab-77-mG2a-f effectively stained HER2 in 14 of 63 cases (22%) of breast cancer, showing its use in the immunohistochemical investigation of formalin-fixed paraffin-embedded tumor sections for identifying HER2.
2.5. ADCC and CDC Activities of H2Mab-77-mG2a-f -Mediated in Breast Cancer
We next tested H2Mab-77-mG2a-f for ADCC against BT-474 cells (HER2-expressing breast cancer cell line) and MDA-MB-468 cells (HER2-negative breast cancer cell line). As shown in Figure 5A, H2Mab-77-mG2a-f demonstrated more ADCC (38.9% cytotoxicity) against BT-474 cells than 281-mG2a-f, the control afucosylated mouse IgG2a (5.5% cytotoxicity; P < 0.001). In contrast, the ADCC activity of H2Mab-77-mG2a-f against MDA-MB-468 cells was 9.3% cytotoxicity, which was comparable to that of 281-mG2a-f (10.2% cytotoxicity).
Then we looked to see whether H2Mab-77-mG2a-f had CDC against BT-474 and MDA-MB-468 cells. As indicated in Figure 5B, H2Mab-77-mG2a-f induced more cytotoxicity (60.5% cytotoxicity) in BT-474 cells than control afucosylated mouse IgG2a (26.1% cytotoxicity; P < 0.01). In contrast, the CDC activity of H2Mab-77-mG2a-f was 11.4% cytotoxicity against MDA-MB-468 cells, which was comparable to that of 281-mG2a-f (9.3% cytotoxicity). These encouraging findings show that H2Mab-77-mG2a-f greatly improves both ADCC and CDC activities against HER2-expressing breast cancer cells.
2.6. Antitumor Activities of H2Mab-77-mG2a-f in the Mouse Xenografts of Breast Tumor Cells
Tumor formation was observed on day 7 in 16 mice inoculated with either BT-474 or MDA-MB-468 cells. The mice bearing breast cancer were divided into two groups: one group received treatment with H2Mab-77-mG2a-f, and the other group served as the control and received 281-mG2a-f. Intraperitoneal injections of H2Mab-77-mG2a-f (100 μg) and 281-mG2a-f (100 μg) were administered to the respective groups on days 7, 14, and 21 after cell inoculation. Tumor diameters were measured on days 7, 10, 14, 16, 21, 24, and 28 following cell inoculation. In the BT-474-bearing mice, the group treated with H2Mab-77-mG2a-f exhibited significantly less tumor growth on days 24 (P < 0.01) and 28 (P < 0.001) compared to the group treated with 281-mG2a-f (Figure 6A). The reduction in tumor volume achieved by H2Mab-77-mG2a-f treatment was 36.6% on day 28. Conversely, there was no difference in tumor growth between the H2Mab-77-mG2a-f and 281-mG2a-f treatment groups in the MDA-MB-468-bearing mice (Figure 6B). Additionally, the tumor weight of the BT-474-bearing mice treated with H2Mab-77-mG2a-f was significantly lower than that of the mice treated with 281-mG2a-f (45.1% reduction; P < 0.01, Figure 6C). However, no difference in tumor weight on day 28 was observed between the H2Mab-77-mG2a-f and 281-mG2a-f treatment groups in the MDA-MB-468-bearing mice (Figure 6D). Figure 6E,F show the resected tumors from the H2Mab-77-mG2a-f and 281-mG2a-f-treated groups, respectively, on day 28 after inoculation of BT-474 and MDA-MB-468 cells. There were no significant differences in total body weights between the H2Mab-77-mG2a-f and 281-mG2a-f-treated groups in both the BT-474 and MDA-MB-468-bearing xenografts (Figure 7A,B). The appearance of mice treated with H2Mab-77-mG2a-f and 281-mG2a-f on day 28 after cell inoculation is shown in Figure 7C (BT-474) and Figure 7D (MDA-MB-468), respectively. In summary, these results demonstrate the antitumor effects of H2Mab-77-mG2a-f administration against HER2-positive breast cancer xenografts.
3. Discussion
Humanized anti-HER2 mAbs like trastuzumab and pertuzumab have helped patients with HER2-positive breast cancer live longer [23,25,48,49]. Aside from monotherapy with these mAbs, combined treatment with trastuzumab and pertuzumab has been shown to enhance outcomes [50]. The membrane-bound p95 generated by the extracellular cleavage of HER2 by metalloproteinase has kinase activity in HER2-overexpressing cells [31]. The inhibition of this cleavage by trastuzumab may be one of the factors suppressing proliferative signals. Furthermore, the antitumor effects trastuzumab is probably mediated by indirect mechanisms such as ADCC activity [44]. In the future, we will look into the impacts of H2Mab-77-mG2a-f on dimer formation and HER2 shedding to better understand the mechanism of the antitumor action by H2Mab-77-mG2a-f.
The development of significant acquired resistance is a challenge for the therapeutic use of trastuzumab [51,52]. Concerns have been raised that even strong ADCs, such as T-DM1, may develop resistance owing to diminished antigen binding, poor internalization, lysosomal degradation errors, and accelerated cellular drug clearance by drug-efflux pumps [53]. MUC4, a membrane-associated mucin known to mask membrane proteins, has been shown to inhibit trastuzumab binding to HER2 [54]. Furthermore, MUC4 acts as an intramembranous ligand and activator of HER2, resulting in the inhibition of apoptosis in cancer cells [55].
Antibodies have distinct activities depending on their binding epitopes, similar to trastuzumab and pertuzumab. The FDA-approved margetuximab and trastuzumab have comparable epitopes and binding affinity; however, margetuximab has a greater binding capacity to the ADCC activator FcγRIIIa and a lower affinity for the immune activation inhibitor CD32B [56,57,58]. Patients with breast cancer who have low-binding FcγRIIIa alleles may also benefit from a combination of margetuximab and anti-HER2 treatments [59]. When used in conjunction with trastuzumab, the HER2-targeted humanized mAb 1E11 inhibits the growth of HER2-expressing gastric tumors by binding to the HER2 domain IV, which does not overlap with trastuzumab [60]. Therefore, one of the primary strategies for combating drug resistance is the development of antibodies with a variety of features, including the binding epitope. H2Mab-19, H2Mab-41, H2Mab-77, H2Mab-119, H2Mab-139, and H2Mab-181 are anti-HER2 mAbs that we have previously established and proven to have antitumor effects [43,61,62,63,64,65,66]. In our early findings, these mAbs have different epitopes, including the HER2 domains I, III, and IV. Transtuzumab and anti-HER2 antibodies that target several epitopes actually have a stronger antitumor impact than standalone therapies [37].
The modified H2Mab-77-mG2a-f demonstrated ADCC and CDC activities depending on HER2 expression as well as superior antitumor effects in xenograft models of HER2-positive breast cancer (Figure 5 and Figure 6). Combining H2Mab-77-mG2a-f with other HER2-targeted medicines has the potential to overcome resistance to HER2 antibody therapy. HER2-positive breast tumors account for around half of all brain metastases [19,67,68]. In a future investigation, we will confirm the inhibitory impact of H2Mab-77-mG2a-f on this metastasis.
Bispecific antibodies targeting HER2×CD3 (ertumaxomab), HER2×CD16, and HER2×HER3 (zenocutuzumab: MCLA-128 and MM-111) are being developed in addition to naive antibodies [69,70,71,72,73,74]. Recently, progress has been made in the creation of bispecific antibodies that target both the immune checkpoint molecules PD-1/PD-L1 and HER2, with antitumor effects proven in preclinical animals [75,76]. In trastuzumab-resistant cancer models, bispecific antibodies targeting immune checkpoint molecules and HER2 may be more successful than individual mAb treatments. The use of H2Mabs to create bispecific antibodies is another option for increasing anti-HER2 treatment. Furthermore, attention has been drawn to chimeric antigen receptor-T (CAR-T) cell treatment, which possesses both antibody specificity and T cell cytotoxicity [77,78,79]. While the FDA authorized the first CD19 CAR-T treatment for B-cell lymphoma in 2017, no CAR-T therapeutic targeting HER2 has yet to be produced [80]. In preclinical settings, we previously showed that anti-podoplanin CAR-T cells, in which we incorporated the created anti-podoplanin mAb into T cells, had strong antitumor efficacy and may release proinflammatory cytokines [81,82]. The future possibilities of H2Mab-77 for HER2-targeting CAR-T applications are hoped for.
4. Materials and Methods
4.1. Cell Lines
Human breast cancer cell lines (BT-474 and MDA-MB-468) and a human glioblastoma cell line (LN229) were obtained from the American Type Culture Collection (Manassas, VA). To establish a HER2-overexpressed LN229 cell line (LN229/HER2), LN229 cells were transfected with the pCAG/HER2-RAPMAP plasmid using the Neon Transfection System (Thermo Fisher Scientific, Inc., Waltham, MA), as previously [43,83,84] described. All cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Nacalai Tesque, Inc., Kyoto, Japan) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.), 100 μg/mL streptomycin, 100 units/ml penicillin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). The cells were maintained in a humidified incubator at 37°C with 5% CO2 and 95% air atmosphere.
4.2. Animals
To reduce animal hardship and suffering, all animal studies were carried out in compliance with the rules and recommendations. The Institutional Committee for Experiments of the Institute of Microbial Chemistry (Numazu, Japan) authorized animal studies for H2Mab-77-mG2a-f’s antitumor efficacy (approval numbers 2023-001 and 2023-018). Over the course of the trial, mice were kept in a specified pathogen-free environment on an 11 h light/13 h dark cycle with access to food and drink as needed. Throughout the course of the trials, mice were frequently checked for weight and health conditions every two to five days. Humane objectives for euthanasia were established as a loss of original body weight to a point >25% and/or a maximal tumor size >3,000 mm3.
4.3. Antibodies
H2Mab-77, an anti-HER2 mAb, was developed as previously mentioned [43]. We cloned the VH cDNA of H2Mab-77 and the CH of mouse IgG2a into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) in order to change the subclass of H2Mab-77 from mouse IgG1 to mouse IgG2a (H2Mab-77-mG2a). Additionally, the pCAG-Neo vector (FUJIFILM Wako Pure Chemical Corporation) was used to clone the VL cDNA of H2Mab-77 and the CL cDNA of the mouse kappa light chain. Using the ExpiCHO Expression System from Thermo Fisher Scientific, Inc., the vector for H2Mab-77-mG2a was transfected into BINDS-09, FUT8-knockout ExpiCHO-S cells. Using Ab-Capcher (ProteNova Co., Ltd., Kanagawa, Japan), H2Mab-77-mG2a-f, an afucosylated variant of the original antibody, was isolated. For the evaluation of western blotting, RcMab-1, an anti-isocitrate dehydrogenase 1 [IDH1] mAb, was utilized as an internal control. For the investigation of ADCC, CDC, and in vivo antitumor effectiveness, 281-mG2a-f (an afucosylated anti-hamster podoplanin mAb) was employed as an afucosylated reference mouse IgG2a [85,86,87].
4.4. Flow Cytometry
Cells were harvested using 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). Subsequently, they were washed with 0.1% bovine serum albumin (Nacalai Tesque, Inc.) in phosphate-buffered saline (PBS), followed by treatment with H2Mab-77-mG2a-f (1 or 10 µg/mL) for 30 minutes at 4°C. Then, cells were incubated with Alexa Fluor 488-conjugated anti-mouse IgG (1:2000 dilution; Cell Signaling Technology, Inc., Danvers, MA), and fluorescence was measured using an SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
4.5. Determination of the Binding Affinity by Flow Cytometry
After being suspended in 100 μL of serially diluted H2Mab-77-mG2a-f (0.0006–10 µg/mL), the cells were then incubated with 1:200 of Cell Signaling Technology, Inc.’s Alexa Fluor 488-conjugated anti-mouse IgG. The SA3800 Cell Analyzer (Sony Corp.) flow cytometer was then used to gather fluorescence data. The binding isotherms were fitted into the built-in, one-site binding model in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA) to calculate the dissociation constant (KD).
4.6. Western Blotting
Cell pellets were resuspended in phosphate-buffered saline (PBS; Nacalai Tesque, Inc.) containing 1% Triton X-100 (FUJIFILM Wako Pure Chemical Corp.) and 50 μg/mL aprotinin (Nacalai Tesque, Inc.). Cell debris was removed by centrifugation at 21,880 × g for 10 minutes at 4°C. Protein concentration was determined using the bicinchoninic acid (BCA) method. Cell lysates (10 μg) were boiled in sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc.). Proteins were separated on 5-20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corp.) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA). PVDF membranes were blocked with 4% skim milk (Nacalai Tesque, Inc.) for 30 minutes at room temperature. The membranes were then incubated with H2Mab-77-mG2a-f (1 µg/mL) and RcMab-1 (1 µg/mL) for additional 30 minutes at room temperature. Subsequently, the membranes were incubated with peroxidase-conjugated anti-mouse immunoglobulins (diluted 1:2000; Agilent Technologies, Inc., Santa Clara, CA) and peroxidase-conjugated anti-rat immunoglobulins (diluted 1:10000; Sigma-Aldrich Corp.) for 30 minutes. Blots were developed using ImmunoStar LD (FUJIFILM Wako Pure Chemical Corp.) or Pierce™ ECL Plus Western Blotting Substrate (Thermo Fisher Scientific, Inc.) and imaged with a Sayaca-Imager (DRC Co., Ltd., Tokyo, Japan). Image analysis was performed using Qcapture Pro software (DRC Co., Ltd.).
4.7. Immunohistochemical Analysis
After being autoclaved in EnVision FLEX Target Retrieval Solution High pH (Agilent Technologies, Inc.) for 20 min, paraffin-embedded tissue sections from the breast cancer tissue array (Cat. No. T8235721-5, Lot. No. B904111; BioChain, San Francisco, CA, USA) were treated with 3% hydrogen peroxide for 15 minutes at room temperature. Thermo Fisher Scientific, Inc.’s SuperBlock T20 was used to block the tissue sections before H2Mab-77-mG2a-f (10 µg/mL) and the EnVision+ Kit for mice (Agilent Technologies, Inc.) were applied for 60 minutes at room temperature each. 3,3'-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies, Inc.) was used to create the color for 2 minutes at room temperature. Mayer’s hematoxylin solution (FUJIFILM Wako Pure Chemical Corporation) was used for counterstaining. The sections were examined using Leica DMD108 (Leica Microsystems GmbH, Wetzlar, Germany) to capture pictures.
4.8. ADCC of H2Mab-77-mG2a-f
The following evaluation was made of H2Mab-77-mG2a-f’s ability to induce ADCC. Five-week-old female BALB/c nude mice were bought from Charles River Laboratories, Inc. Spleens were extracted aseptically after cervical dislocation euthanasia. Spleen tissues were then processed through a sterile cell strainer (352360, BD Falcon) using a syringe to produce single-cell suspensions. A 10-second exposure to ice-cold, distilled water destroyed erythrocytes. Effector cells were created by resuspending the splenocytes in DMEM with 10% FBS after being cleaned with DMEM. Thermo Fisher Scientific, Inc.’s 10 μg/mL Calcein AM was used to mark the target cells (BT-474 and MDA-MB-468) for the study. In 96-well plates, target cells (2 × 104 cells) were seeded before effector cells (effector to target ratio, 50:1) and 100 μg/mL of either H2Mab-77-mG2a-f or 281-mG2a-f were added. A microplate reader (Power Scan HT; BioTek Instruments, Inc.) with an excitation wavelength of 485 nm and an emission wavelength of 538 nm was used to analyze the Calcein release into the medium after a 4.5-hour incubation at 37°C.
This is how cytolyticity (% lysis) was determined: % lysis is calculated as (E - S)/(M - S) x 100, where “E” denotes the fluorescence in effector and target cell cultures, “S” denotes the spontaneous fluorescence of only target cells, and “M” denotes the maximum fluorescence after treatment with a lysis buffer (10 mM Tris-HCl (pH 7.4), 10 mM EDTA, and 0.5% Triton X-100). The Institutional Committee for trials of the Institute of Microbial Chemistry (Numazu, Japan) authorized animal trials for ADCC of H2Mab-77-mG2a-f (permission number 2023-018).
4.9. CDC of H2Mab-77-mG2a-f
The following procedure was used to assess how well H2Mab-77-mG2a-f induced CDC. With 10 µg/mL Calcein AM, the target cells (BT-474 and MDA-MB-468) were marked. Target cells (2 × 104 cells) were put in 96-well plates with 100 µg/mL of either H2Mab-77-mG2a-f or 281-mG2a-f and rabbit complement (final dilution 1:10; Low-Tox-M Rabbit Complement; Cedarlane Laboratories). The amount of calcein released into the medium was measured during 4.5-hour incubation at 37°C.
4.10. Antitumor Activities of H2Mab-77-mG2a-f in Xenografts of Breast Cancer
Charles River Laboratories, Inc. provided the BALB/c nude mice (female, 5 weeks old). Cells from BT-474 and MDA-MB-468 were mixed with DMEM and BD Biosciences’ Matrigel Matrix Growth Factor Reduced. Subcutaneous injections were then given to the mice’s left flanks. On the seventh post-inoculation day, 100 µg of H2Mab-77-mG2a-f (n = 8) or 281-mG2a-f (n = 8) in 100 µL PBS were administered intraperitoneally. Additional antibody injections were given on days 14 and 21. The tumor diameter was assessed on days 7, 10, 14, 16, 21, 24, and 28 after breast cancer cell implantation. Tumor volumes were calculated in the same manner as previously stated. The weight of the mice was also assessed on days 7, 10, 14, 16, 21, 24, and 28 following breast cancer cell inoculation. When the observations were finished on day 28, the mice were killed, and tumor weights were assessed following tumor excision.
Author Contributions
T.T., H.S., and T.O. performed the experiments. M.K.K. and Y.K. designed the experiments. T.T. and H.S. analyzed the data and wrote the manuscript. All authors have read and agreed to the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP23ama121008 (Y.K.), JP23am0401013 (Y.K.), and JP23ck0106730 (Y.K.).
Institutional Review Board Statement
Animal experiments were approved by the Institutional Committee for Experiments of the Institute of Microbial Chemistry (approval no. 2022-056, 2023-001, and 2023-018).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article and supplementary material.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Pal, S.K.; Pegram, M. Targeting HER2 Epitopes. Semin Oncol 2006, 33, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Agus, D.B.; Akita, R.W.; Fox, W.D.; Lewis, G.D.; Higgins, B.; Pisacane, P.I.; Lofgren, J.A.; Tindell, C.; Evans, D.P.; Maiese, K.; et al. Targeting ligand-activated ErbB2 signaling inhibits breast and prostate tumor growth. Cancer Cell 2002, 2, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Garrett, T.P.; McKern, N.M.; Lou, M.; Elleman, T.C.; Adams, T.E.; Lovrecz, G.O.; Kofler, M.; Jorissen, R.N.; Nice, E.C.; Burgess, A.W.; et al. The crystal structure of a truncated ErbB2 ectodomain reveals an active conformation, poised to interact with other ErbB receptors. Mol Cell 2003, 11, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Wallasch, C.; Weiss, F.U.; Niederfellner, G.; Jallal, B.; Issing, W.; Ullrich, A. Heregulin-dependent regulation of HER2/neu oncogenic signaling by heterodimerization with HER3. Embo j 1995, 14, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Alimandi, M.; Romano, A.; Curia, M.C.; Muraro, R.; Fedi, P.; Aaronson, S.A.; Di Fiore, P.P.; Kraus, M.H. Cooperative signaling of ErbB3 and ErbB2 in neoplastic transformation and human mammary carcinomas. Oncogene 1995, 10, 1813–1821. [Google Scholar]
- Pinkas-Kramarski, R.; Soussan, L.; Waterman, H.; Levkowitz, G.; Alroy, I.; Klapper, L.; Lavi, S.; Seger, R.; Ratzkin, B.J.; Sela, M.; et al. Diversification of Neu differentiation factor and epidermal growth factor signaling by combinatorial receptor interactions. Embo j 1996, 15, 2452–2467. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J Clin 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Cronin, K.A.; Harlan, L.C.; Dodd, K.W.; Abrams, J.S.; Ballard-Barbash, R. Population-based estimate of the prevalence of HER-2 positive breast cancer tumors for early stage patients in the US. Cancer Invest 2010, 28, 963–968. [Google Scholar] [CrossRef]
- Guarneri, V.; Barbieri, E.; Dieci, M.V.; Piacentini, F.; Conte, P. Anti-HER2 neoadjuvant and adjuvant therapies in HER2 positive breast cancer. Cancer Treat Rev 2010, 36 Suppl 3, S62–66. [Google Scholar] [CrossRef]
- Abd El-Rehim, D.M.; Pinder, S.E.; Paish, C.E.; Bell, J.A.; Rampaul, R.S.; Blamey, R.W.; Robertson, J.F.; Nicholson, R.I.; Ellis, I.O. Expression and co-expression of the members of the epidermal growth factor receptor (EGFR) family in invasive breast carcinoma. Br J Cancer 2004, 91, 1532–1542. [Google Scholar] [CrossRef]
- Purdie, C.A.; Baker, L.; Ashfield, A.; Chatterjee, S.; Jordan, L.B.; Quinlan, P.; Adamson, D.J.; Dewar, J.A.; Thompson, A.M. Increased mortality in HER2 positive, oestrogen receptor positive invasive breast cancer: a population-based study. Br J Cancer 2010, 103, 475–481. [Google Scholar] [CrossRef]
- Herter-Sprie, G.S.; Greulich, H.; Wong, K.K. Activating Mutations in ERBB2 and Their Impact on Diagnostics and Treatment. Front Oncol 2013, 3, 86. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, F.; Fontanella, C.; Amoroso, V.; Bianchi, G.V.; Bisagni, G.; Falci, C.; Fontana, A.; Generali, D.; Gianni, L.; Grassadonia, A.; et al. Current challenges in HER2-positive breast cancer. Crit Rev Oncol Hematol 2016, 98, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.F. HER2 and Breast Cancer - A Phenomenal Success Story. N Engl J Med 2019, 381, 1284–1286. [Google Scholar] [CrossRef]
- Arteaga, C.L.; Engelman, J.A. ERBB receptors: from oncogene discovery to basic science to mechanism-based cancer therapeutics. Cancer Cell 2014, 25, 282–303. [Google Scholar] [CrossRef]
- Riecke, K.; Witzel, I. Targeting the Human Epidermal Growth Factor Receptor Family in Breast Cancer beyond HER2. Breast Care (Basel) 2020, 15, 579–585. [Google Scholar] [CrossRef]
- Martin, A.M.; Cagney, D.N.; Catalano, P.J.; Warren, L.E.; Bellon, J.R.; Punglia, R.S.; Claus, E.B.; Lee, E.Q.; Wen, P.Y.; Haas-Kogan, D.A.; et al. Brain Metastases in Newly Diagnosed Breast Cancer: A Population-Based Study. JAMA Oncol 2017, 3, 1069–1077. [Google Scholar] [CrossRef]
- Kuksis, M.; Gao, Y.; Tran, W.; Hoey, C.; Kiss, A.; Komorowski, A.S.; Dhaliwal, A.J.; Sahgal, A.; Das, S.; Chan, K.K.; et al. The incidence of brain metastases among patients with metastatic breast cancer: a systematic review and meta-analysis. Neuro Oncol 2021, 23, 894–904. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.; Schwartz, J.N.; Hagerty, K.L.; Allred, D.C.; Cote, R.J.; Dowsett, M.; Fitzgibbons, P.L.; Hanna, W.M.; Langer, A.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. J Clin Oncol 2007, 25, 118–145. [Google Scholar] [CrossRef]
- Ross, J.S.; Slodkowska, E.A.; Symmans, W.F.; Pusztai, L.; Ravdin, P.M.; Hortobagyi, G.N. The HER-2 receptor and breast cancer: ten years of targeted anti-HER-2 therapy and personalized medicine. Oncologist 2009, 14, 320–368. [Google Scholar] [CrossRef] [PubMed]
- Cobleigh, M.A.; Vogel, C.L.; Tripathy, D.; Robert, N.J.; Scholl, S.; Fehrenbacher, L.; Wolter, J.M.; Paton, V.; Shak, S.; Lieberman, G.; et al. Multinational study of the efficacy and safety of humanized anti-HER2 monoclonal antibody in women who have HER2-overexpressing metastatic breast cancer that has progressed after chemotherapy for metastatic disease. J Clin Oncol 1999, 17, 2639–2648. [Google Scholar] [CrossRef]
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of chemotherapy plus a monoclonal antibody against HER2 for metastatic breast cancer that overexpresses HER2. N Engl J Med 2001, 344, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J Clin Oncol 2002, 20, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Amiri-Kordestani, L.; Blumenthal, G.M.; Xu, Q.C.; Zhang, L.; Tang, S.W.; Ha, L.; Weinberg, W.C.; Chi, B.; Candau-Chacon, R.; Hughes, P.; et al. FDA approval: ado-trastuzumab emtansine for the treatment of patients with HER2-positive metastatic breast cancer. Clin Cancer Res 2014, 20, 4436–4441. [Google Scholar] [CrossRef]
- Nakada, T.; Sugihara, K.; Jikoh, T.; Abe, Y.; Agatsuma, T. The Latest Research and Development into the Antibody-Drug Conjugate, [fam-] Trastuzumab Deruxtecan (DS-8201a), for HER2 Cancer Therapy. Chem Pharm Bull (Tokyo) 2019, 67, 173–185. [Google Scholar] [CrossRef]
- Zheng, Y.; Zou, J.; Sun, C.; Peng, F.; Peng, C. Ado-tratuzumab emtansine beyond breast cancer: therapeutic role of targeting other HER2-positive cancers. Front Mol Biosci 2023, 10, 1165781. [Google Scholar] [CrossRef]
- Hudis, C.A. Trastuzumab--mechanism of action and use in clinical practice. N Engl J Med 2007, 357, 39–51. [Google Scholar] [CrossRef]
- Cho, H.S.; Mason, K.; Ramyar, K.X.; Stanley, A.M.; Gabelli, S.B.; Denney, D.W., Jr.; Leahy, D.J. Structure of the extracellular region of HER2 alone and in complex with the Herceptin Fab. Nature 2003, 421, 756–760. [Google Scholar] [CrossRef]
- Shi, Y.; Fan, X.; Deng, H.; Brezski, R.J.; Rycyzyn, M.; Jordan, R.E.; Strohl, W.R.; Zou, Q.; Zhang, N.; An, Z. Trastuzumab triggers phagocytic killing of high HER2 cancer cells in vitro and in vivo by interaction with Fcγ receptors on macrophages. J Immunol 2015, 194, 4379–4386. [Google Scholar] [CrossRef]
- Molina, M.A.; Codony-Servat, J.; Albanell, J.; Rojo, F.; Arribas, J.; Baselga, J. Trastuzumab (herceptin), a humanized anti-Her2 receptor monoclonal antibody, inhibits basal and activated Her2 ectodomain cleavage in breast cancer cells. Cancer Res 2001, 61, 4744–4749. [Google Scholar]
- Loo, L.; Capobianco, J.A.; Wu, W.; Gao, X.; Shih, W.Y.; Shih, W.H.; Pourrezaei, K.; Robinson, M.K.; Adams, G.P. Highly sensitive detection of HER2 extracellular domain in the serum of breast cancer patients by piezoelectric microcantilevers. Anal Chem 2011, 83, 3392–3397. [Google Scholar] [CrossRef]
- Junttila, T.T.; Li, G.; Parsons, K.; Phillips, G.L.; Sliwkowski, M.X. Trastuzumab-DM1 (T-DM1) retains all the mechanisms of action of trastuzumab and efficiently inhibits growth of lapatinib insensitive breast cancer. Breast Cancer Res Treat 2011, 128, 347–356. [Google Scholar] [CrossRef]
- Ogitani, Y.; Hagihara, K.; Oitate, M.; Naito, H.; Agatsuma, T. Bystander killing effect of DS-8201a, a novel anti-human epidermal growth factor receptor 2 antibody-drug conjugate, in tumors with human epidermal growth factor receptor 2 heterogeneity. Cancer Sci 2016, 107, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Li, B.T.; Smit, E.F.; Goto, Y.; Nakagawa, K.; Udagawa, H.; Mazières, J.; Nagasaka, M.; Bazhenova, L.; Saltos, A.N.; Felip, E.; et al. Trastuzumab Deruxtecan in HER2-Mutant Non-Small-Cell Lung Cancer. N Engl J Med 2022, 386, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Shitara, K.; Bang, Y.J.; Iwasa, S.; Sugimoto, N.; Ryu, M.H.; Sakai, D.; Chung, H.C.; Kawakami, H.; Yabusaki, H.; Lee, J.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Positive Gastric Cancer. N Engl J Med 2020, 382, 2419–2430. [Google Scholar] [CrossRef] [PubMed]
- Spiridon, C.I.; Ghetie, M.A.; Uhr, J.; Marches, R.; Li, J.L.; Shen, G.L.; Vitetta, E.S. Targeting multiple Her-2 epitopes with monoclonal antibodies results in improved antigrowth activity of a human breast cancer cell line in vitro and in vivo. Clin Cancer Res 2002, 8, 1720–1730. [Google Scholar] [PubMed]
- Harbeck, N.; Beckmann, M.W.; Rody, A.; Schneeweiss, A.; Müller, V.; Fehm, T.; Marschner, N.; Gluz, O.; Schrader, I.; Heinrich, G.; et al. HER2 Dimerization Inhibitor Pertuzumab - Mode of Action and Clinical Data in Breast Cancer. Breast Care (Basel) 2013, 8, 49–55. [Google Scholar] [CrossRef]
- Barthélémy, P.; Leblanc, J.; Goldbarg, V.; Wendling, F.; Kurtz, J.E. Pertuzumab: development beyond breast cancer. Anticancer Res 2014, 34, 1483–1491. [Google Scholar]
- Franklin, M.C.; Carey, K.D.; Vajdos, F.F.; Leahy, D.J.; de Vos, A.M.; Sliwkowski, M.X. Insights into ErbB signaling from the structure of the ErbB2-pertuzumab complex. Cancer Cell 2004, 5, 317–328. [Google Scholar] [CrossRef]
- Clynes, R.A.; Towers, T.L.; Presta, L.G.; Ravetch, J.V. Inhibitory Fc receptors modulate in vivo cytotoxicity against tumor targets. Nat Med 2000, 6, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Nitta, H.; Li, Z. HER2 Intratumoral Heterogeneity in Breast Cancer, an Evolving Concept. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Fujii, Y.; Kaneko, M.K.; Yamada, S.; Nakamura, T.; Yanaka, M.; Saidoh, N.; Chang, Y.W.; Handa, S.; Takahashi, M.; et al. H2Mab-77 is a Sensitive and Specific Anti-HER2 Monoclonal Antibody Against Breast Cancer. Monoclon Antib Immunodiagn Immunother 2017, 36, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Tsao, L.C.; Force, J.; Hartman, Z.C. Mechanisms of Therapeutic Antitumor Monoclonal Antibodies. Cancer Res 2021, 81, 4641–4651. [Google Scholar] [CrossRef]
- Shinkawa, T.; Nakamura, K.; Yamane, N.; Shoji-Hosaka, E.; Kanda, Y.; Sakurada, M.; Uchida, K.; Anazawa, H.; Satoh, M.; Yamasaki, M.; et al. The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem 2003, 278, 3466–3473. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Ohishi, T.; Nakamura, T.; Inoue, H.; Takei, J.; Sano, M.; Asano, T.; Sayama, Y.; Hosono, H.; Suzuki, H.; et al. Development of Core-Fucose-Deficient Humanized and Chimeric Anti-Human Podoplanin Antibodies. Monoclon Antib Immunodiagn Immunother 2020, 39, 167–174. [Google Scholar] [CrossRef]
- Song, K.H.; Trudeau, T.; Kar, A.; Borden, M.A.; Gutierrez-Hartmann, A. Ultrasound-mediated delivery of siESE complexed with microbubbles attenuates HER2+/- cell line proliferation and tumor growth in rodent models of breast cancer. Nanotheranostics 2019, 3, 212–222. [Google Scholar] [CrossRef]
- Valabrega, G.; Montemurro, F.; Aglietta, M. Trastuzumab: mechanism of action, resistance and future perspectives in HER2-overexpressing breast cancer. Ann Oncol 2007, 18, 977–984. [Google Scholar] [CrossRef]
- Amiri-Kordestani, L.; Wedam, S.; Zhang, L.; Tang, S.; Tilley, A.; Ibrahim, A.; Justice, R.; Pazdur, R.; Cortazar, P. First FDA approval of neoadjuvant therapy for breast cancer: pertuzumab for the treatment of patients with HER2-positive breast cancer. Clin Cancer Res 2014, 20, 5359–5364. [Google Scholar] [CrossRef]
- Swain, S.M.; Kim, S.B.; Cortés, J.; Ro, J.; Semiglazov, V.; Campone, M.; Ciruelos, E.; Ferrero, J.M.; Schneeweiss, A.; Knott, A.; et al. Pertuzumab, trastuzumab, and docetaxel for HER2-positive metastatic breast cancer (CLEOPATRA study): overall survival results from a randomised, double-blind, placebo-controlled, phase 3 study. Lancet Oncol 2013, 14, 461–471. [Google Scholar] [CrossRef]
- Derakhshani, A.; Rezaei, Z.; Safarpour, H.; Sabri, M.; Mir, A.; Sanati, M.A.; Vahidian, F.; Gholamiyan Moghadam, A.; Aghadoukht, A.; Hajiasgharzadeh, K.; et al. Overcoming trastuzumab resistance in HER2-positive breast cancer using combination therapy. J Cell Physiol 2020, 235, 3142–3156. [Google Scholar] [CrossRef]
- Vivekanandhan, S.; Knutson, K.L. Resistance to Trastuzumab. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Khoury, R.; Saleh, K.; Khalife, N.; Saleh, M.; Chahine, C.; Ibrahim, R.; Lecesne, A. Mechanisms of Resistance to Antibody-Drug Conjugates. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Nagy, P.; Friedländer, E.; Tanner, M.; Kapanen, A.I.; Carraway, K.L.; Isola, J.; Jovin, T.M. Decreased accessibility and lack of activation of ErbB2 in JIMT-1, a herceptin-resistant, MUC4-expressing breast cancer cell line. Cancer Res 2005, 65, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Carraway, K.L.; Perez, A.; Idris, N.; Jepson, S.; Arango, M.; Komatsu, M.; Haq, B.; Price-Schiavi, S.A.; Zhang, J.; Carraway, C.A. Muc4/sialomucin complex, the intramembrane ErbB2 ligand, in cancer and epithelia: to protect and to survive. Prog Nucleic Acid Res Mol Biol 2002, 71, 149–185. [Google Scholar] [CrossRef] [PubMed]
- Royce, M.; Osgood, C.L.; Amatya, A.K.; Fiero, M.H.; Chang, C.J.G.; Ricks, T.K.; Shetty, K.A.; Kraft, J.; Qiu, J.; Song, P.; et al. FDA Approval Summary: Margetuximab plus Chemotherapy for Advanced or Metastatic HER2-Positive Breast Cancer. Clin Cancer Res 2022, 28, 1487–1492. [Google Scholar] [CrossRef]
- Nordstrom, J.L.; Gorlatov, S.; Zhang, W.; Yang, Y.; Huang, L.; Burke, S.; Li, H.; Ciccarone, V.; Zhang, T.; Stavenhagen, J.; et al. Anti-tumor activity and toxicokinetics analysis of MGAH22, an anti-HER2 monoclonal antibody with enhanced Fcγ receptor binding properties. Breast Cancer Res 2011, 13, R123. [Google Scholar] [CrossRef]
- Rugo, H.S.; Im, S.A.; Cardoso, F.; Cortés, J.; Curigliano, G.; Musolino, A.; Pegram, M.D.; Wright, G.S.; Saura, C.; Escrivá-de-Romaní, S.; et al. Efficacy of Margetuximab vs Trastuzumab in Patients With Pretreated ERBB2-Positive Advanced Breast Cancer: A Phase 3 Randomized Clinical Trial. JAMA Oncol 2021, 7, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Mandó, P.; Rivero, S.G.; Rizzo, M.M.; Pinkasz, M.; Levy, E.M. Targeting ADCC: A different approach to HER2 breast cancer in the immunotherapy era. Breast 2021, 60, 15–25. [Google Scholar] [CrossRef]
- Ko, B.K.; Lee, S.Y.; Lee, Y.H.; Hwang, I.S.; Persson, H.; Rockberg, J.; Borrebaeck, C.; Park, D.; Kim, K.T.; Uhlen, M.; et al. Combination of novel HER2-targeting antibody 1E11 with trastuzumab shows synergistic antitumor activity in HER2-positive gastric cancer. Mol Oncol 2015, 9, 398–408. [Google Scholar] [CrossRef]
- Yamada, S.; Itai, S.; Nakamura, T.; Chang, Y.W.; Harada, H.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Establishment of H(2)Mab-119, an Anti-Human Epidermal Growth Factor Receptor 2 Monoclonal Antibody, Against Pancreatic Cancer. Monoclon Antib Immunodiagn Immunother 2017, 36, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Yamada, S.; Itai, S.; Kato, Y. Development of an Anti-HER2 Monoclonal Antibody H2Mab-139 Against Colon Cancer. Monoclon Antib Immunodiagn Immunother 2018, 37, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ohishi, T.; Takei, J.; Nakamura, T.; Sano, M.; Asano, T.; Sayama, Y.; Hosono, H.; Kawada, M.; Kaneko, M.K. An Anti-Human Epidermal Growth Factor Receptor 2 Monoclonal Antibody H2Mab-19 Exerts Antitumor Activity in Mouse Colon Cancer Xenografts. Monoclon Antib Immunodiagn Immunother 2020, 39, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ohishi, T.; Yamada, S.; Itai, S.; Takei, J.; Sano, M.; Nakamura, T.; Harada, H.; Kawada, M.; Kaneko, M.K. Anti-Human Epidermal Growth Factor Receptor 2 Monoclonal Antibody H2Mab-41 Exerts Antitumor Activity in a Mouse Xenograft Model of Colon Cancer. Monoclon Antib Immunodiagn Immunother 2019, 38, 157–161. [Google Scholar] [CrossRef]
- Kato, Y.; Ohishi, T.; Sano, M.; Asano, T.; Sayama, Y.; Takei, J.; Kawada, M.; Kaneko, M.K. H(2)Mab-19 Anti-Human Epidermal Growth Factor Receptor 2 Monoclonal Antibody Therapy Exerts Antitumor Activity in Pancreatic Cancer Xenograft Models. Monoclon Antib Immunodiagn Immunother 2020, 39, 61–65. [Google Scholar] [CrossRef]
- Kato, Y.; Ohishi, T.; Takei, J.; Nakamura, T.; Kawada, M.; Kaneko, M.K. An Antihuman Epidermal Growth Factor Receptor 2 Monoclonal Antibody (H(2)Mab-19) Exerts Antitumor Activity in Glioblastoma Xenograft Models. Monoclon Antib Immunodiagn Immunother 2020, 39, 135–139. [Google Scholar] [CrossRef]
- Azim, H.A.; Azim, H.A., Jr. Systemic treatment of brain metastases in HER2-positive breast cancer: current status and future directions. Future Oncol 2012, 8, 135–144. [Google Scholar] [CrossRef]
- Leone, J.P.; Leone, B.A. Breast cancer brain metastases: the last frontier. Exp Hematol Oncol 2015, 4, 33. [Google Scholar] [CrossRef]
- Turini, M.; Chames, P.; Bruhns, P.; Baty, D.; Kerfelec, B. A FcγRIII-engaging bispecific antibody expands the range of HER2-expressing breast tumors eligible to antibody therapy. Oncotarget 2014, 5, 5304–5319. [Google Scholar] [CrossRef]
- Diermeier-Daucher, S.; Ortmann, O.; Buchholz, S.; Brockhoff, G. Trifunctional antibody ertumaxomab: Non-immunological effects on Her2 receptor activity and downstream signaling. MAbs 2012, 4, 614–622. [Google Scholar] [CrossRef]
- Haense, N.; Atmaca, A.; Pauligk, C.; Steinmetz, K.; Marmé, F.; Haag, G.M.; Rieger, M.; Ottmann, O.G.; Ruf, P.; Lindhofer, H.; et al. A phase I trial of the trifunctional anti Her2 × anti CD3 antibody ertumaxomab in patients with advanced solid tumors. BMC Cancer 2016, 16, 420. [Google Scholar] [CrossRef]
- Schram, A.M.; Odintsov, I.; Espinosa-Cotton, M.; Khodos, I.; Sisso, W.J.; Mattar, M.S.; Lui, A.J.W.; Vojnic, M.; Shameem, S.H.; Chauhan, T.; et al. Zenocutuzumab, a HER2xHER3 Bispecific Antibody, Is Effective Therapy for Tumors Driven by NRG1 Gene Rearrangements. Cancer Discov 2022, 12, 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Fontana, E.; Torga, G.; Fostea, R.; Cleator, S.; Wasserman, E.; Murat, A.; Arkenau, H.T. Sustained Tumor Regression With Zenocutuzumab, a Bispecific Antibody Targeting Human Epidermal Growth Factor Receptor 2/Human Epidermal Growth Factor Receptor 3 Signaling, in NRG1 Fusion-Positive, Estrogen Receptor-Positive Breast Cancer After Progression on a Cyclin-Dependent Kinase 4/6 Inhibitor. JCO Precis Oncol 2022, 6, e2100446. [Google Scholar] [CrossRef]
- McDonagh, C.F.; Huhalov, A.; Harms, B.D.; Adams, S.; Paragas, V.; Oyama, S.; Zhang, B.; Luus, L.; Overland, R.; Nguyen, S.; et al. Antitumor activity of a novel bispecific antibody that targets the ErbB2/ErbB3 oncogenic unit and inhibits heregulin-induced activation of ErbB3. Mol Cancer Ther 2012, 11, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Cui, Y.; Liu, X.; Liu, G.; Dong, X.; Tang, L.; Hung, Y.; Wang, C.; Feng, M.Q. A bispecific antibody targeting HER2 and PD-L1 inhibits tumor growth with superior efficacy. J Biol Chem 2021, 297, 101420. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.L.; Zhu, H.X.; Deng, L.; Meng, X.Q.; Li, K.; Xu, W.; Zhao, L.; Liu, Y.Q.; Zhu, Z.P.; Huang, H.M. Bispecific antibody simultaneously targeting PD1 and HER2 inhibits tumor growth via direct tumor cell killing in combination with PD1/PDL1 blockade and HER2 inhibition. Acta Pharmacol Sin 2022, 43, 672–680. [Google Scholar] [CrossRef]
- Boulch, M.; Cazaux, M.; Loe-Mie, Y.; Thibaut, R.; Corre, B.; Lemaître, F.; Grandjean, C.L.; Garcia, Z.; Bousso, P. A cross-talk between CAR T cell subsets and the tumor microenvironment is essential for sustained cytotoxic activity. Sci Immunol 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Lugtenburg, P.J.; Kersten, M.J.; Subklewe, M.; Spanjaart, A.; Shah, N.N.; Kerbauy, L.N.; Roddie, C.; Pennings, E.R.A.; Mahuad, C.; et al. Cellular therapy in lymphoma. Hematol Oncol 2023. [Google Scholar] [CrossRef]
- Shin, M.H.; Oh, E.; Kim, Y.; Nam, D.H.; Jeon, S.Y.; Yu, J.H.; Minn, D. Recent Advances in CAR-Based Solid Tumor Immunotherapy. Cells 2023, 12. [Google Scholar] [CrossRef]
- Mullard, A. FDA approves first CAR T therapy. Nat Rev Drug Discov 2017, 16, 669. [Google Scholar] [CrossRef]
- Shiina, S.; Ohno, M.; Ohka, F.; Kuramitsu, S.; Yamamichi, A.; Kato, A.; Motomura, K.; Tanahashi, K.; Yamamoto, T.; Watanabe, R.; et al. CAR T Cells Targeting Podoplanin Reduce Orthotopic Glioblastomas in Mouse Brains. Cancer Immunol Res 2016, 4, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A.; Waseda, M.; Ishii, T.; Kaneko, M.K.; Kato, Y.; Kaneko, S. Improved anti-solid tumor response by humanized anti-podoplanin chimeric antigen receptor transduced human cytotoxic T cells in an animal model. Genes Cells 2022, 27, 549–558. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016, 35, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.K.; Ogasawara, S.; Yamada, S.; Yanaka, M.; Nakamura, T.; Saidoh, N.; Yoshida, K.; Honma, R.; Kato, Y. Development of RAP Tag, a Novel Tagging System for Protein Detection and Purification. Monoclon Antib Immunodiagn Immunother 2017, 36, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Ikota, H.; Nobusawa, S.; Arai, H.; Kato, Y.; Ishizawa, K.; Hirose, T.; Yokoo, H. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015, 32, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y. Specific monoclonal antibodies against IDH1/2 mutations as diagnostic tools for gliomas. Brain Tumor Pathol 2015, 32, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Nanamiya, R.; Suzuki, H.; Takei, J.; Li, G.; Goto, N.; Harada, H.; Saito, M.; Tanaka, T.; Asano, T.; Kaneko, M.K.; et al. Development of Monoclonal Antibody 281-mG(2a)-f Against Golden Hamster Podoplanin. Monoclon Antib Immunodiagn Immunother 2022, 41, 311–319. [Google Scholar] [CrossRef]
Figure 1.
Generation of H2Mab-77-mG2a-f (mouse IgG2a) and H2Mab-77-mG2a-f (core-fucose-deleted mouse IgG2a) from the original anti-HER2 mAb, H2Mab-77 (mouse IgG1).
Figure 1.
Generation of H2Mab-77-mG2a-f (mouse IgG2a) and H2Mab-77-mG2a-f (core-fucose-deleted mouse IgG2a) from the original anti-HER2 mAb, H2Mab-77 (mouse IgG1).
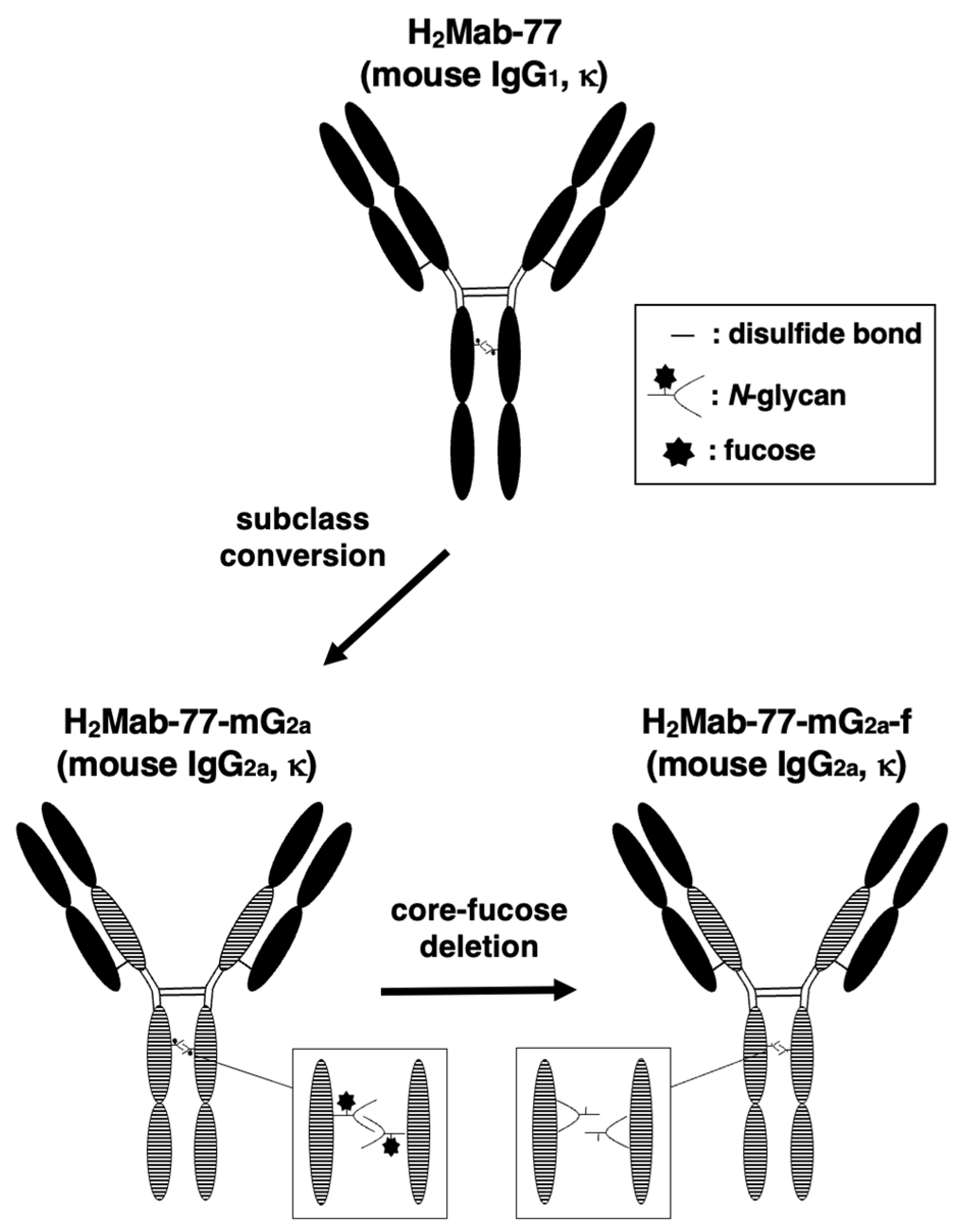
Figure 2.
Flow cytometric analysis using H2Mab-77-mG2a-f. (A) LN229/HER2, (B) LN229, (C) BT-474, and (D) MDA-MB-468 cells were treated with H2Mab-77-mG2a-f (red) or buffer control (black), followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were analyzed using the SA3800 Cell Analyzer. The binding affinity of H2Mab-77-mG2a-f was determined by flow cytometry in (E) LN229/HER2 and (F) BT-474 cells. Serially diluted H2Mab-77-mG2a-f was added to the cells, followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer, and the dissociation constant (KD) was calculated using GraphPad PRISM 6.
Figure 2.
Flow cytometric analysis using H2Mab-77-mG2a-f. (A) LN229/HER2, (B) LN229, (C) BT-474, and (D) MDA-MB-468 cells were treated with H2Mab-77-mG2a-f (red) or buffer control (black), followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were analyzed using the SA3800 Cell Analyzer. The binding affinity of H2Mab-77-mG2a-f was determined by flow cytometry in (E) LN229/HER2 and (F) BT-474 cells. Serially diluted H2Mab-77-mG2a-f was added to the cells, followed by Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer, and the dissociation constant (KD) was calculated using GraphPad PRISM 6.
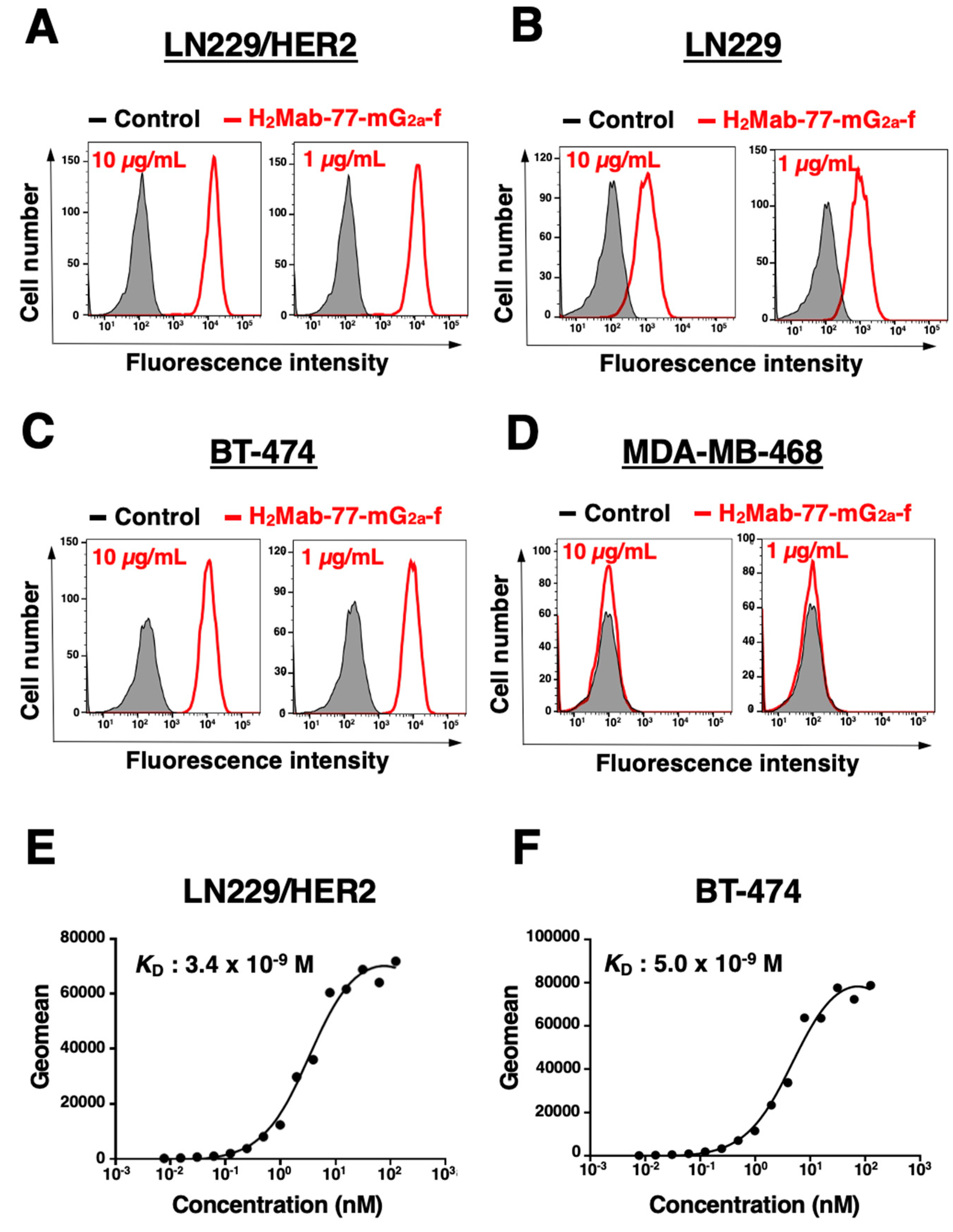
Figure 3.
Detection of HER2 by western blotting using H2Mab-77-mG2a-f. Cell lysates were electrophoresed and transferred onto PVDF membranes. After blocking, the PVDF membranes were incubated with H2Mab-77-mG2a-f (1 µg/mL) (A) or an anti-IDH1 monoclonal antibody (clone RcMab-1, 1 µg/mL) (B), followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins or peroxidase-conjugated anti-rat immunoglobulins. Blots were developed using ImmunoStar LD or ECL Plus Western Blotting Substrate and imaged with a Sayaca-Imager.
Figure 3.
Detection of HER2 by western blotting using H2Mab-77-mG2a-f. Cell lysates were electrophoresed and transferred onto PVDF membranes. After blocking, the PVDF membranes were incubated with H2Mab-77-mG2a-f (1 µg/mL) (A) or an anti-IDH1 monoclonal antibody (clone RcMab-1, 1 µg/mL) (B), followed by incubation with peroxidase-conjugated anti-mouse immunoglobulins or peroxidase-conjugated anti-rat immunoglobulins. Blots were developed using ImmunoStar LD or ECL Plus Western Blotting Substrate and imaged with a Sayaca-Imager.
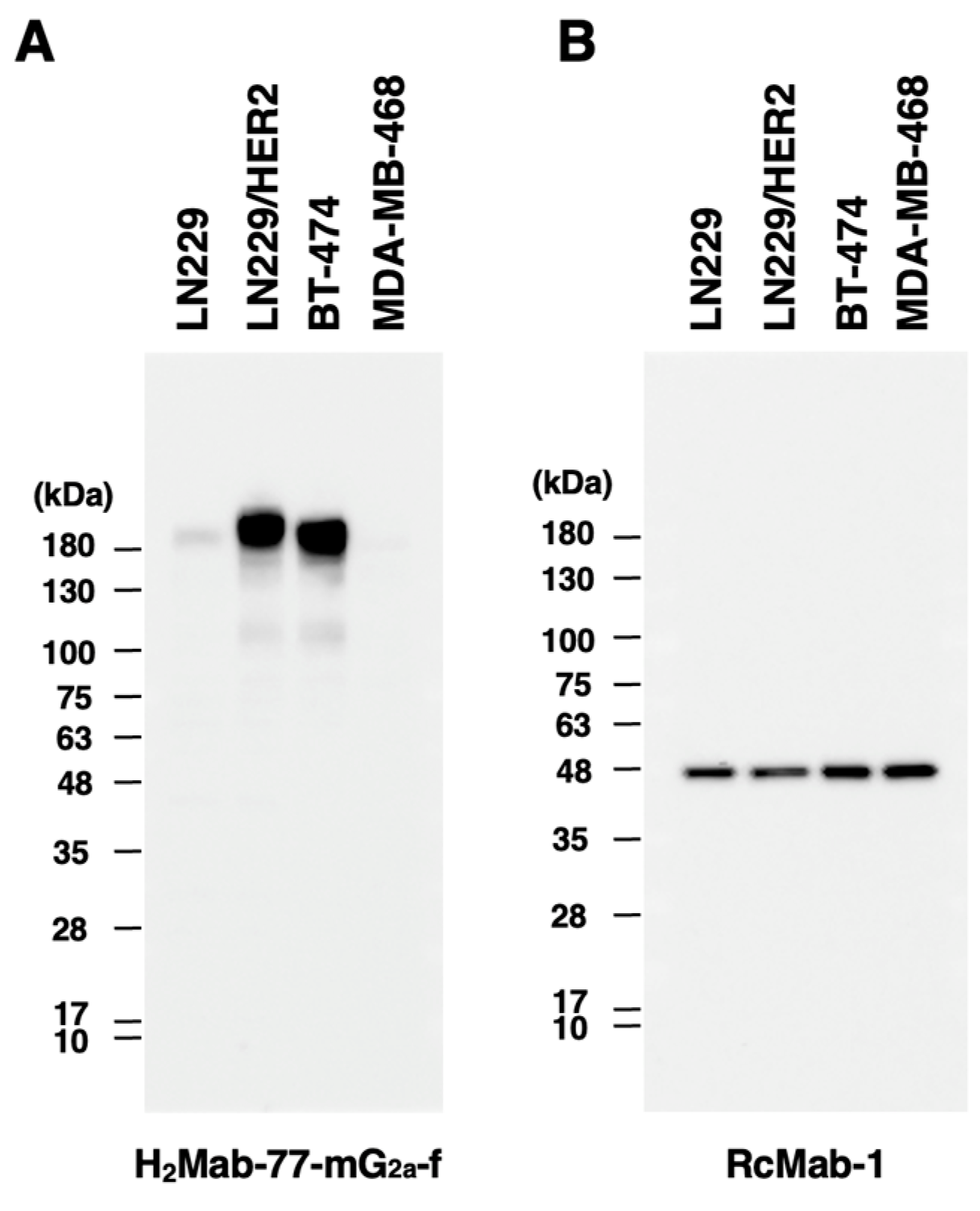
Figure 4.
(A,B) Detection of HER2 in breast cancer specimens using immunohistochemical analysis with H2Mab-77-mG2a-f. Tissue sections from patients with human breast cancer were incubated with H2Mab-77-mG2a-f at a concentration of 10 µg/mL and then treated with the EnVision+ kit. Scale bar: 100 µm.
Figure 4.
(A,B) Detection of HER2 in breast cancer specimens using immunohistochemical analysis with H2Mab-77-mG2a-f. Tissue sections from patients with human breast cancer were incubated with H2Mab-77-mG2a-f at a concentration of 10 µg/mL and then treated with the EnVision+ kit. Scale bar: 100 µm.
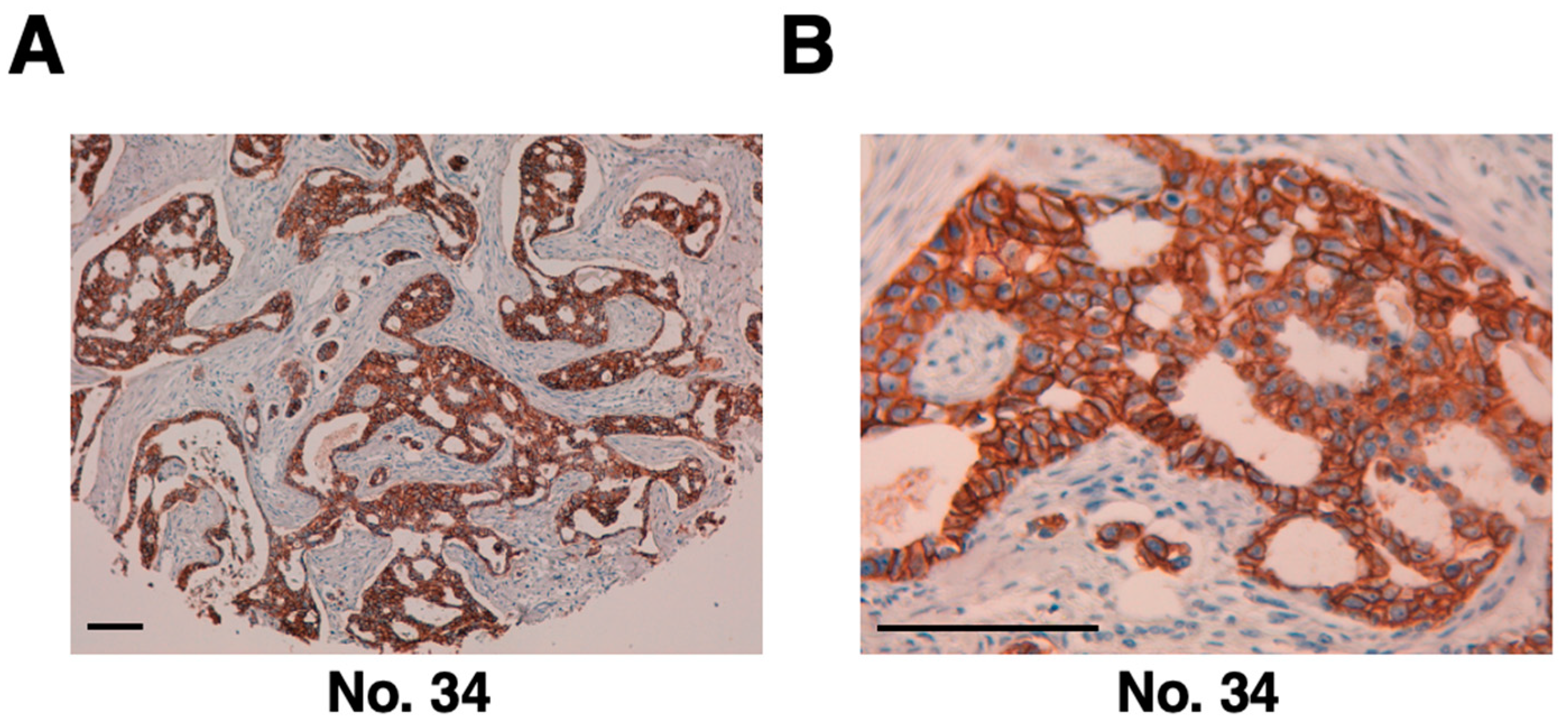
Figure 5.
Investigation of the ADCC and CDC activities elicited by H2Mab-77-mG2a-f. (A) Antibody-dependent cellular cytotoxicity (ADCC) elicited by H2Mab-77-mG2a-f and 281-mG2a-f (control antibodies) targeting BT-474 and MDA-MB-468 cells. (B) Complement-dependent cytotoxicity (CDC) elicited by H2Mab-77-mG2a-f and 281-mG2a-f (control antibodies) targeting BT-474 and MDA-MB-468 cells. Values are presented as the mean ± SEM. Statistical significance is indicated by asterisks (***P < 0.001, **P < 0.01, n.s., not significant, unpaired t-test). ADCC, antibody-dependent cellular cytotoxicity; CDC, complement-dependent cytotoxicity.
Figure 5.
Investigation of the ADCC and CDC activities elicited by H2Mab-77-mG2a-f. (A) Antibody-dependent cellular cytotoxicity (ADCC) elicited by H2Mab-77-mG2a-f and 281-mG2a-f (control antibodies) targeting BT-474 and MDA-MB-468 cells. (B) Complement-dependent cytotoxicity (CDC) elicited by H2Mab-77-mG2a-f and 281-mG2a-f (control antibodies) targeting BT-474 and MDA-MB-468 cells. Values are presented as the mean ± SEM. Statistical significance is indicated by asterisks (***P < 0.001, **P < 0.01, n.s., not significant, unpaired t-test). ADCC, antibody-dependent cellular cytotoxicity; CDC, complement-dependent cytotoxicity.
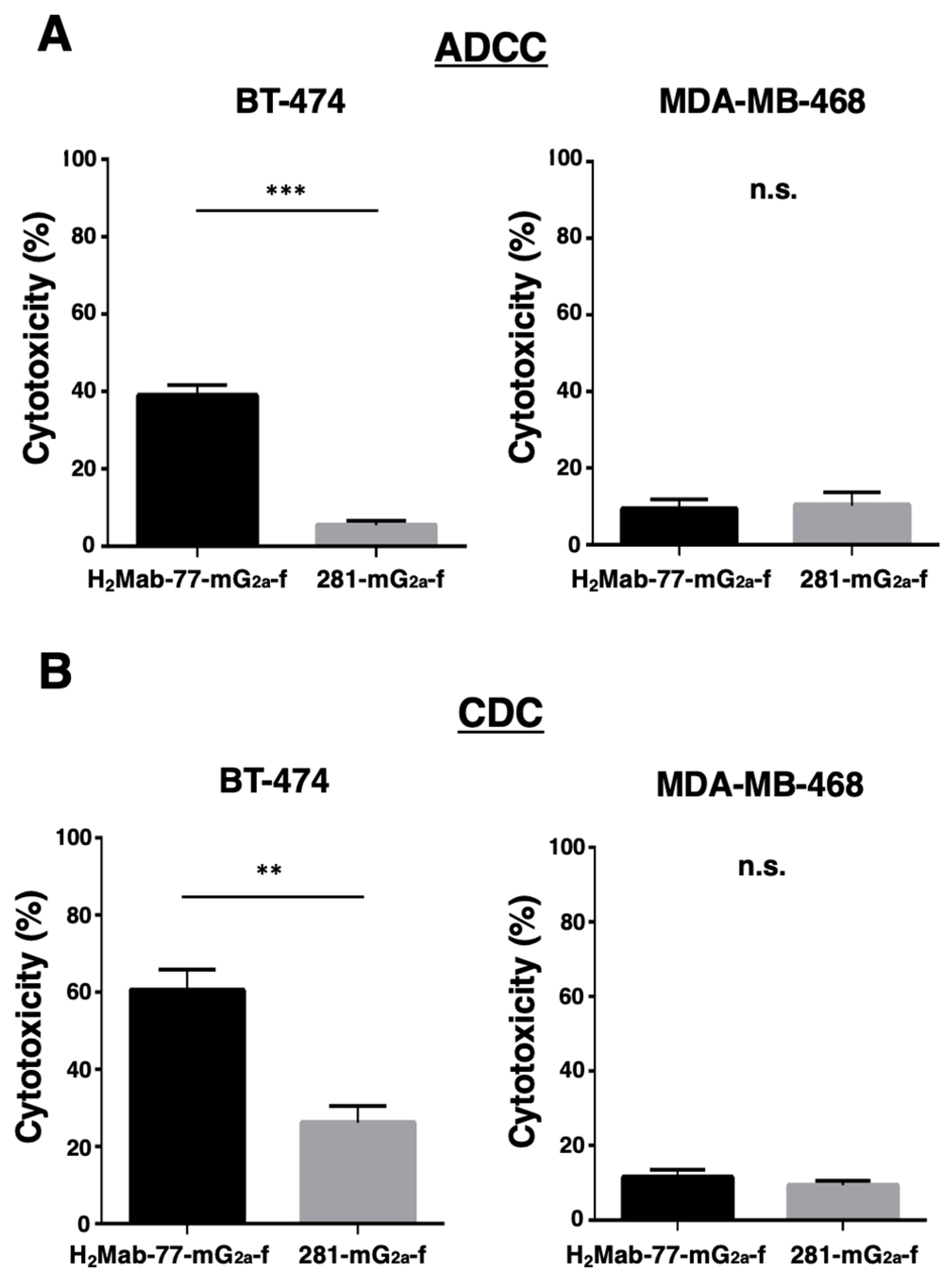
Figure 6.
Evaluation of the antitumor activity of H2Mab-77-mG2a-f in breast cancer xenograft models. (A) BT-474 cells (5 × 106 cells) and (B) MDA-MB-468 cells (5 × 106 cells) were subcutaneously transplanted into the left flanks of mice. On day 7 after transplantation, 100 μg of H2Mab-77-mG2a-f and 281-mG2a-f (control) were intraperitoneally injected into mice. Additional antibody treatments were conducted on days 14 and 21. Tumor diameters were measured on days 7, 10, 14, 16, 21, 24, and 28 after the inoculation of tumor cells. Values are presented as the mean ± SEM. Statistical significance is indicated by asterisks (***P < 0.001, **P < 0.01, n.s., not significant, ANOVA, and Sidak’s multiple comparisons test). (C) Tumors of BT-474 and (D) MDA-MB-468 xenografts were resected from H2Mab-77-mG2a-f and 281-mG2a-f (control) groups. Tumor weight on day 28, tumor weight was measured from the excised xenografts. Values are presented as the mean ± SEM. Statistical significance is denoted by asterisks (**P < 0.01, n.s., not significant, unpaired t-test). (E) Resected tumors of BT-474 and (F) MDA-MB-468 xenografts from H2Mab-77-mG2a-f and 281-mG2a-f (control) groups on day 28. Scale bar: 1 cm.
Figure 6.
Evaluation of the antitumor activity of H2Mab-77-mG2a-f in breast cancer xenograft models. (A) BT-474 cells (5 × 106 cells) and (B) MDA-MB-468 cells (5 × 106 cells) were subcutaneously transplanted into the left flanks of mice. On day 7 after transplantation, 100 μg of H2Mab-77-mG2a-f and 281-mG2a-f (control) were intraperitoneally injected into mice. Additional antibody treatments were conducted on days 14 and 21. Tumor diameters were measured on days 7, 10, 14, 16, 21, 24, and 28 after the inoculation of tumor cells. Values are presented as the mean ± SEM. Statistical significance is indicated by asterisks (***P < 0.001, **P < 0.01, n.s., not significant, ANOVA, and Sidak’s multiple comparisons test). (C) Tumors of BT-474 and (D) MDA-MB-468 xenografts were resected from H2Mab-77-mG2a-f and 281-mG2a-f (control) groups. Tumor weight on day 28, tumor weight was measured from the excised xenografts. Values are presented as the mean ± SEM. Statistical significance is denoted by asterisks (**P < 0.01, n.s., not significant, unpaired t-test). (E) Resected tumors of BT-474 and (F) MDA-MB-468 xenografts from H2Mab-77-mG2a-f and 281-mG2a-f (control) groups on day 28. Scale bar: 1 cm.
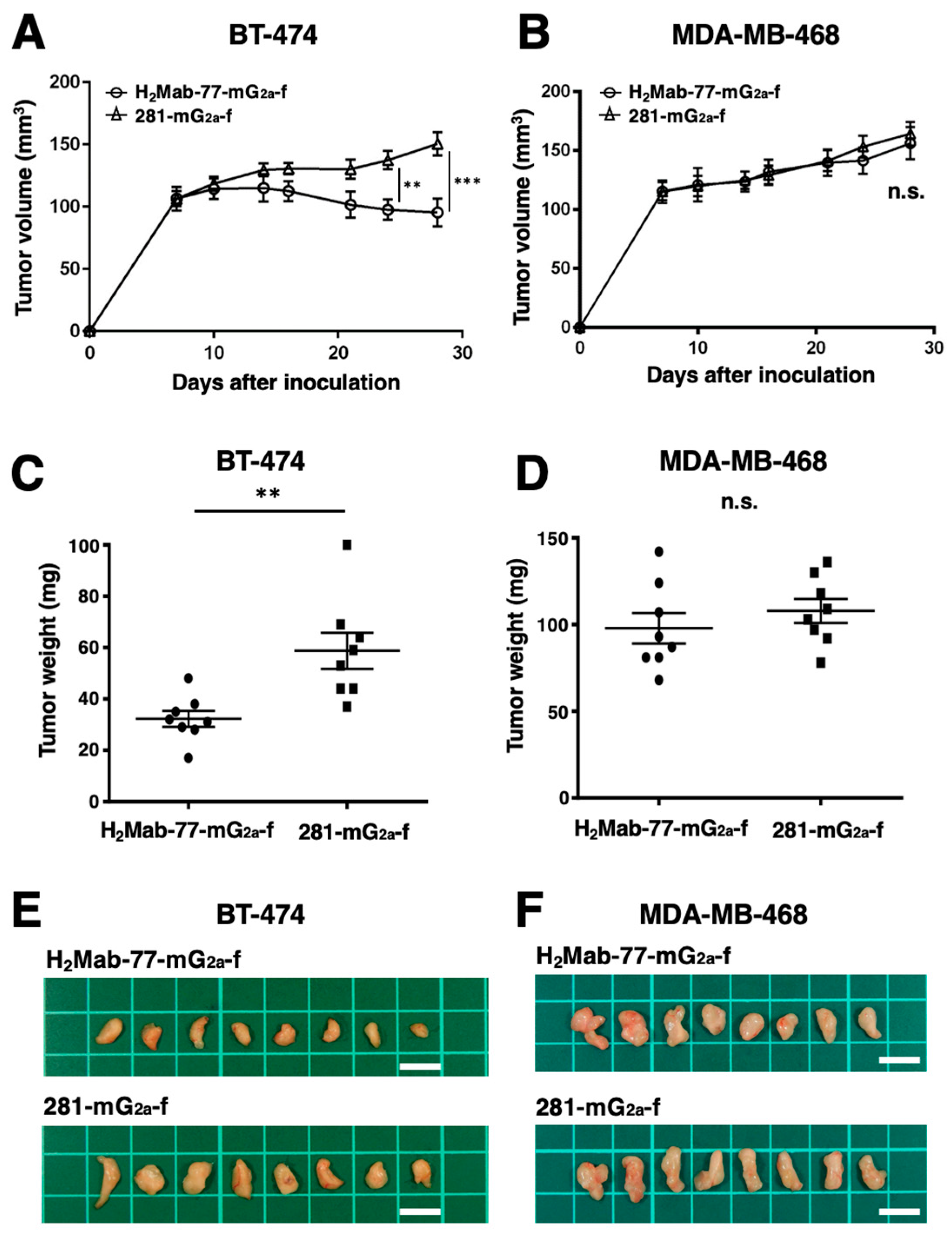
Figure 7.
Monitoring the body weights and appearances of mice bearing breast tumor xenografts. (A) Body weights of mice inoculated with BT-474 and (B) MDA-MB-468 xenografts in H2Mab-77-mG2a-f and 281-mG2a-f (control) groups were recorded on days 7, 10, 14, 16, 21, 24, and 28. Values are mean ± SEM. No statistical significance is indicated by n.s. (ANOVA and Sidak’s multiple comparisons test). (C) Body appearance of BT-474 and (D) MDA-MB-468-implanted mice in H2Mab-77-mG2a-f and 281-mG2a-f (control) groups on day 28, respectively (scale bar: 1 cm).
Figure 7.
Monitoring the body weights and appearances of mice bearing breast tumor xenografts. (A) Body weights of mice inoculated with BT-474 and (B) MDA-MB-468 xenografts in H2Mab-77-mG2a-f and 281-mG2a-f (control) groups were recorded on days 7, 10, 14, 16, 21, 24, and 28. Values are mean ± SEM. No statistical significance is indicated by n.s. (ANOVA and Sidak’s multiple comparisons test). (C) Body appearance of BT-474 and (D) MDA-MB-468-implanted mice in H2Mab-77-mG2a-f and 281-mG2a-f (control) groups on day 28, respectively (scale bar: 1 cm).
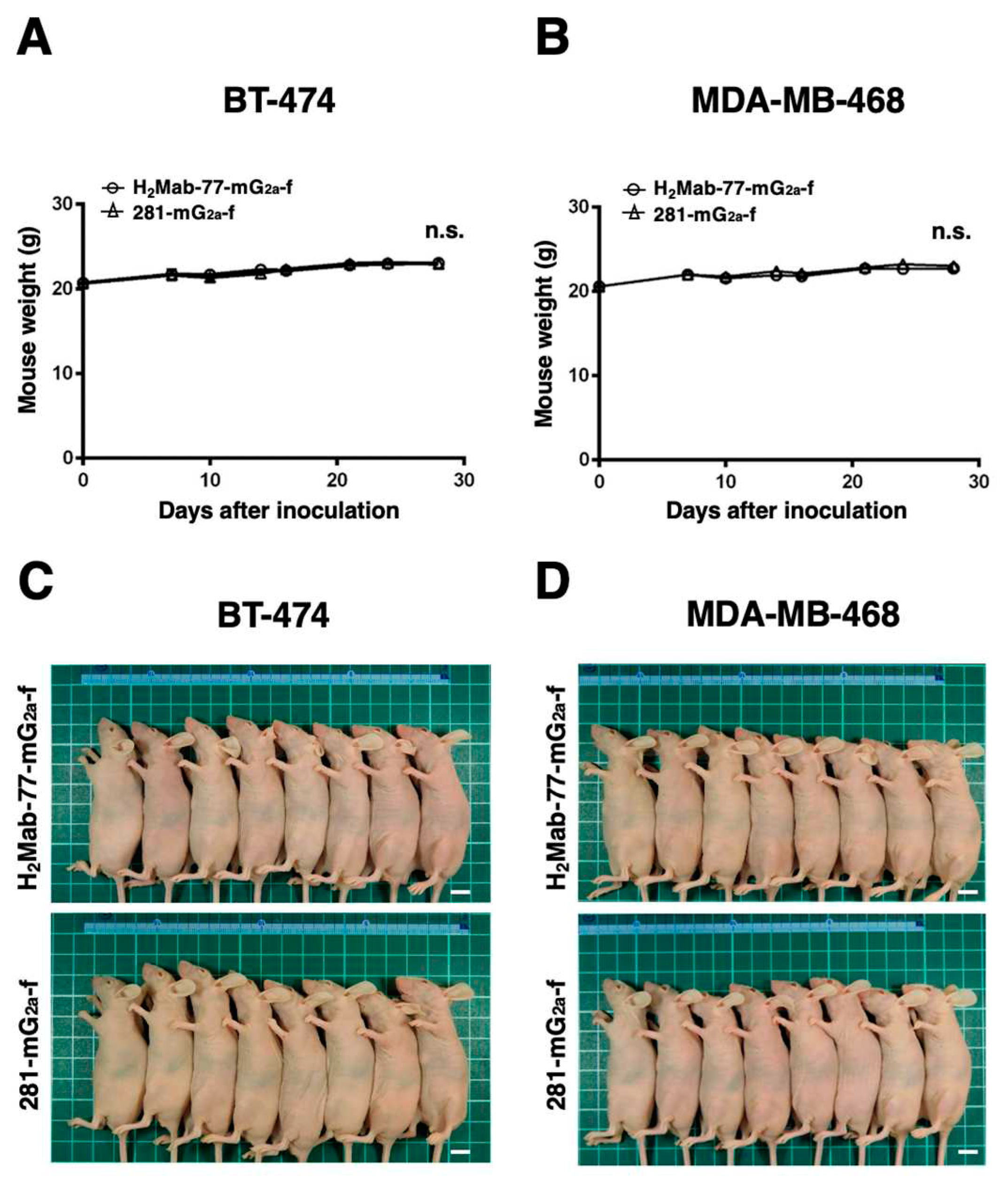

Table 1.
Immunohistochemical analysis using H2Mab-77-mG2a-f against breast cancer tissue array.
| No. | Sex | Age | Pathological Diagnosis | Differentiation | TNM | H2Mab-77-mG2a-f |
|---|---|---|---|---|---|---|
| 1 | F | 44 | Invasive ductal carcinoma | Moderately | T2N2M1 | - |
| 2 | F | 58 | Medullary carcinoma | Moderately | T2N2M1 | - |
| 3 | F | 40 | Invasive ductal carcinoma | Moderately | T2N1M0 | 2+ |
| 4 | F | 52 | Invasive ductal carcinoma | Moderately | T2N2M1 | - |
| 5 | F | 60 | Invasive ductal carcinoma | Moderately | T2N1M1 | - |
| 6 | F | 57 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 7 | F | 48 | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 8 | F | 66 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 9 | F | 58 | Adenocarcinoma | Moderately | T2N2M1 | - |
| 10 | F | 63 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 11 | F | 32 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 12 | F | 59 | Invasive lobular carcinoma | Well | T2N2M0 | - |
| 13 | F | 44 | Invasive lobular carcinoma | Well | T2N2M0 | - |
| 14 | F | 60 | Invasive lobular carcinoma | Moderately | T2N1M0 | - |
| 15 | F | 44 | Invasive ductal carcinoma | Moderately | T2N2M0 | 3+ |
| 16 | F | 82 | Invasive ductal carcinoma | Moderately | T2N1M1 | - |
| 17 | F | 58 | Adenocarcinoma | Moderately | T2N1M1 | 2+ |
| 18 | F | 57 | Invasive ductal carcinoma | Poorly | T3N3M0 | - |
| 19 | F | 41 | Invasive ductal carcinoma | Moderately | T2N1M0 | - |
| 20 | F | 44 | Invasive ductal carcinoma | Moderately | T2N2M0 | 1+ |
| 21 | F | 78 | Invasive ductal carcinoma | Moderately | T2N1M0 | - |
| 22 | F | 60 | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 23 | F | N/A | Invasive ductal carcinoma | Moderately | T2N1M1 | 2+ |
| 24 | F | 46 | Invasive ductal carcinoma | Moderately | T2N3M1 | - |
| 25 | F | 41 | Invasive ductal carcinoma | Moderately | T2N2M0 | - |
| 26 | F | 59 | Invasive ductal carcinoma | Poorly | T2N0M0 | - |
| 27 | F | 45 | Invasive ductal carcinoma | Poorly | T2N0M0 | - |
| 28 | F | 43 | Invasive ductal carcinoma | N/A | T2N1M1 | - |
| 29 | F | 26 | Fibroadenoma | N/A | T1N0M0 | - |
| 30 | F | 40 | Invasive ductal carcinoma | N/A | T1N0M0 | - |
| 31 | F | 38 | Fibroadenoma | N/A | T2N0M0 | - |
| 32 | F | 51 | Invasive ductal carcinoma | Moderately | T2N2M0 | - |
| 33 | F | 45 | Invasive ductal carcinoma | Poorly | T2N0M0 | 2+ |
| 34 | F | 45 | Invasive ductal carcinoma | Poorly | T2N1M0 | 3+ |
| 35 | F | 47 | Invasive ductal carcinoma | Moderately | T2N1M0 | - |
| 36 | F | 55 | Invasive ductal carcinoma | Moderately | T2N3M1 | 1+ |
| 37 | F | 58 | Invasive ductal carcinoma | Moderately | T3N3M0 | - |
| 38 | F | 47 | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 39 | F | 38 | Invasive ductal carcinoma | Poorly | T2N0M0 | - |
| 40 | F | 40 | Invasive ductal carcinoma | Poorly | T2N0M0 | - |
| 41 | F | 57 | Invasive ductal carcinoma | Poorly | T2N0M0 | - |
| 42 | F | 42 | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 43 | F | 60 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 44 | F | 58 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 45 | F | 41 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 46 | F | 50 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 47 | F | 60 | Invasive ductal carcinoma | Moderately | T2N2M1 | - |
| 48 | F | 53 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 49 | F | 65 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 50 | F | 43 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 51 | F | 57 | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 52 | F | 37 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 53 | F | 50 | Invasive ductal carcinoma | Moderately | T2N3M0 | - |
| 54 | F | 48 | Invasive ductal carcinoma | Poorly | T2N1M0 | - |
| 55 | F | 50 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 56 | F | 53 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 57 | F | 49 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 58 | F | 65 | Invasive ductal carcinoma | Moderately | T2N1M0 | - |
| 59 | F | 43 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 60 | F | 58 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 61 | F | 48 | Invasive ductal carcinoma | Moderately | T2N0M0 | - |
| 62 | F | N/A | Invasive ductal carcinoma | Moderately | N/A | 1+ |
| 63 | F | N/A | Invasive ductal carcinoma | Moderately | N/A | - |
N/A, not available; TNM, tumor node metastasis; F, female. -, no stain; 1+, weak intensity; 2+, moderate intensity; 3+, strong intensity
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



