
Submitted:
12 July 2023
Posted:
13 July 2023
You are already at the latest version
Abstract
Two novel dsRNA mycoviruses were found in different isolates of Diplodia fraxini, NW FVA 1581 and NW FVA 1706, which were isolated from a root, associated with stem collar necrosis of Fraxinus excelsior L. Both mycelia are infected by a novel fusagravirus, which was named Diplodia fraxini fusagravirus 1 (DfFV1), and isolate NW FVA 1706 is additionally infected by a novel partitivirus, which was denominated as Diplodia fraxini partitivirus 1 (DfPV1). The one-segmented, bicistronic genome of DfFV1 is composed of about 8,500 bp. Their ORFs are connected by a 1 slippery heptamer sequence and the 3’ terminal ORF is coding for the viral RdRp. The genome of DfPV1 is composed of three, monocistronic dsRNA segments ranging from 1,755 bp (dsRNA 1) over 1,588 bp (dsRNA 2) to 1,233 bp (dsRNA 3). Based on genome organization and phylogenetic positions, DfFV1 was assigned to the proposed family of “Fusagraviridae” and DfPV1 to the genus Gammapartitivirus within the family of Partitiviridae.
Ultra-structural analysis showed that polysomal structures were stabilized in the single infection and none of these structures could be isolated in the double infection. It is assumed that DfFV1 has an opportunistic lifestyle, being either protected by ribosomes or by transcapsidation from particles of DfPV1.
Keywords:
Fusagravirus
; Partitivirus
; Co-infection
; Transcapsidation
; Polysomal encapsulation
; Diplodia fraxini
1. Introduction
The rapid spread of the invasive Ascomycete Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz & Hosoya affects the natural ash species in Europe and their decline increased in the last three decades. Especially in Northern Europe, ash trees already have been eradicated [1]. Different fungi are associated with infected trees and among them, members of the Botryosphaeriaceae are the main species involved in ash dieback etiology [2]. Within this family, Diplodia mutila (Fr.) Mont. (teleomorph: Botryosphaeria stevensii Shoemaker) was one of the most reported species [3,4]. Diplodia fraxini (Fr.) Fr. (Botryodiplodia fraxini (Fr.) Sacc.) has been referred to Diplodia mutila (D. mutila) in previous studies [5], as both species are very similar and closely related. Based on the work of Alves et al. [6], D. mutila and Diplodia fraxini (D. fraxini) can be differentiated by morphological and phylogenetic markers. Diplodia fraxini causes dark brown inner bark lesions on F. excelsior that spread up and down from the site of infection.
In all main taxa of fungi and oomycetes, mycoviruses are widespread [reviewed in 7]. Since the first discovery of a mycovirus which causes morphological alterations in cultivated mushrooms by Hollings [8], the knowledge about mycoviruses has expanded rapidly in the last decade. Mycoviruses can encode their proteasome on positive sense single-stranded RNA (+ssRNA), which was latest discovered to be the most widespread strategy among mycoviruses, on negative sense single-stranded RNA (-ssRNA) or on positive sense single-stranded DNA (+ssDNA). Besides these, many mycoviruses possess a genome which consists of double-stranded RNA (dsRNA) [reviewed in 7].
Within the order of Durnavirales, currently six families are recognized by the International Committee on Taxonomy of Viruses (ICTV): Amalgaviridae, Curvulaviridae, Fusariviridae, Hypoviridae, Picobirnaviridae and Partitiviridae (https://ictv.global/taxonomy). Members of the Family Partitividae encode a RdRp and a capsid protein on two dsRNA segments which range in total from 3,000–4,800 bp. However, additional defective or satellite dsRNA segments may also be present. The segments are separately encapsidated by identical capsid subunits in isometric particles with sizes ranging from 25 to 43 nm [9]. According to the ICTV, members of the family are separated in five genera (Alphapartitivirus, Betapartitivirus, Cryspovirus, Deltapartitivirus, Gammapartitivirus) [9]. Recently, two novel genera, “Epsilonpartitivirus” and “Zetapatitivirus” were proposed by Jiang et al. [10] and Nerva et al. [11]. While partitiviruses found in fungi, plants and insects are accommodated in the genera Alpha-, Beta- and Deltapartitivirus or in the proposed genus “Epsilonpartitivirus”, members of the genus Gammapartitivirus and of the proposed genus “Zetapatitivirus” are exclusively described from fungal hosts [reviewed in 7,9]. Beside their host range, the different genera are distinguished by features of the dsRNA, the size of the segments and the molecular weight (MW) of the capsid protein subunits [9]. While the 3’-terminus of members of the Alpha- and Betapartitiviruses genus is polyadenylated, no poly(A) tail is found on gammapartitiviruses. Several members of the Partitiviridae family are not bipartite but tripartite [12,13]. The function of the third segment is still unknown and it may not be always detectable [12].
Viruses in the proposed family of “Fusagraviridae” [14] contain a bicistronic dsRNA genome which ranges from 8,500 bp for Trichoderma atroviride mycovirus 1 [15] to 10,200 bp for Cryphonectria naterciae fusagravirus 1 [16]. Viruses of this family were detected in fungi, plants [17] and insects [18]; however, they are, according to Ayllón and Vainio [7] mostly found in ascomycetes. The two ORFs are believed to be separated by a -1 ribosomal frameshifting, which is mediated by a heptameric slippery sequence with the consensus nucleotides (nts) XXXX (any nucleotide) YY (either A or U) and Z (not G) upstream of the 5’-proximal (ORF 1) stop codon and a Recoding Stimulatory Element (RSE) immediately downstream from the slippery site [14,19,20,21,22]. While ORF 1 encodes a hypothetical protein, the 3´-proximal ORF (ORF 2) encodes a protein with RdRp motifs and in some species a Phytoreo_S7 domain was detected [14]. Although for several fusagra-like viruses, the expression of a capsid protein was verified [17] and particles were obtained by sucrose density centrifugation [15], others suggested a capsidless nature of fusagraviruses [18,23,24].
In many publications, species within the genus Botryosphaeria were described to be viral hosts [25,26,27], the first report of a virus in Diplodia sp. was described from Diplodia seriata (De Not) with a multiinfection [28]. Until now, no viruses were described from D. fraxini. In here, we describe the novel partitivirus Diplodia fraxini partitivirus 1 (DfPV1) and two strains of the novel Diplodia fraxini fusagravirus 1 (DfFV1) which we isolated from two independent isolates of D. fraxini. We showed that the single infection of DfFV1 and the double infection of DfFV1 and DfPV1 differ in the formation of their ultrastructural patterns.
2. Materials and Methods
2.1. Fungal Isolates, Propagation And Species Determination
The Diplodia fraxini strains NW-FVA 1581 and NW-FVA 1706 were isolated from trunk tissues of Fraxinus excelsior L. The sampled European ash trees were affected by ash dieback caused by H. fraxineus and exhibited stem collar necrosis and rots. Isolation of both D. fraxini strains and the sample sites were described in Langer [5], in which these strains were referred to Botryosphaeria stevensii (Anamorph: Diplodia mutila). Briefly, NW-FVA 1581 was isolated from wood tissue sampled in Schleswig-Holstein, Germany (Tree 8, N54° 40.264' E9°41.202'). NW-FVA 1706 was sampled from necrotic tissue in Schleswig-Holstein, Germany, (Tree 26, N54° 05.238' E10° 23.159').
Cultivation of mycelium was performed in the dark at room temperature on solid complete medium (CMS) as stated in Leach et al. [29]. For harvesting, the mycelium was separated from the medium by a cellophane sheet.
For the phylogenetic analysis of the fungal isolates, genomic DNA was extracted using the method of Damm et al. [30]. The 5.8S nuclear ribosomal gene with the two flanking internal transcribed spacers (ITS-1 and ITS-2) was amplified using the primer pairs ITS-1F [31] and ITS-4 [32]. Additionally, partial sequences of the translation elongation factor 1α (TEF1-α) and of the β-tubulin gene (TUB) were generated using the primer pairs EF1-728F [33], EF1-1567R [34] and Bt2a+Bt2b [35]. Obtained sequences of PCR products were submitted to GenBank and are displayed in Table 1.
For phylogenetic analysis, three single locus datasets (ITS, EF1-α, TUB), including appropriate reference sequences retrieved from GenBank, were aligned automatically using MAFFT v. 7.308 [36,37] and manually adjusted where necessary. The concatenated ITS-EF1-α-TUB sequence-dataset was analyzed using Bayesian Inference (BI) and Maximum Likelihood (ML). For BI analysis, the best fit model of evolution for each partition was estimated by MEGA7 [38]. Posterior probabilities were determined by Markov Chain Monte Carlo sampling (MCMC) in MrBayes v. 3.2.6 [39,40] as implemented in Geneious R11 [41], using the estimated models of evolution. Four simultaneous Markov chains were run for 1 million generations and trees were sampled every 100th generation. The first 2000 trees, which represent the burn-in phase of the analysis, were discarded and the remaining 8000 trees were used to calculate posterior probabilities in the majority rule consensus tree. The ML analysis was performed by RAxML v. 8.2.11 [42,43] as implemented in Geneious R11 [41], using the GTRGAMMA model with the rapid bootstrapping and search for best scoring ML tree algorithm, including 1000 bootstrap replicates.
2.2. DsRNA Extraction, Virus-Like Particle Purification, Protein Analysis and Electron Microscopy
Double stranded RNA was extracted from mycelium using the dsRNA Extraction kit (iNtRON Biotechnology, Seongnam-Si, South Korea) and was analyzed by 1 % (w/v) agarose gel electrophoresis. Virus like particles (VLPs) and polysomes (PSs) were extracted as described for the betachrysovirus Fusarium graminearum virus-China 9 (FgV-ch9) in Lutz et al. [44]. The dsRNA of VLPs and PSs was extracted by peqGOLD TriFastTM (VWR life sciences, Radnor, Pennsylvania, USA) according to the manufacturer’s protocol. The protein patterns of VLPs and PSs were analyzed by a 12.5 % (w/v) SDS-PAGE visualized by Coomassie-Brilliant Blue staining. Bands were cut from the gel and sequenced with LC-MS/MS by a nano-liquid chromatography system (Dionex UltiMate 3000 RSLCnano, ThermoFisher Scientific, Waltham, Massachusetts, USA) and analyzed by means of the Proteome Discoverer 2.0 (ThermoFisher Scientific) by the Universitätsklinikum Hamburg-Eppendorf (UKE, Hamburg, Germany). VLPs and PSs were examined by electron microscopy (LEO 906E, Zeiss, Germany) with 2 % (w/v) uranyl acetate contrasting.
2.3. Virus Sequence Determination
From VLPs isolated dsRNA was submitted to Next-Generation Sequencing. The libraries were prepared according to Nextera XT DNA Library Preparation Kit (Illumina Inc., San Diego, CA, USA) and run on a NextSeq 2000 (Illumina Inc., San Diego, CA, USA) instrument at the Leibniz Institute DSMZ (Braunschweig, Germany) as pair-end reads (2 × 151). De novo assembly of contigs was performed by using Geneious Prime software (Biomatters, Auckland, New Zealand, version 2021.2.2). The extreme 5′- and 3′-termini were determined by single-primer amplification technique (SPAT) using an oligonucleotide with a phosphorylated 5′-terminus and a 2′,3′-dideoxyC-group (23ddC) at the 3′-terminus as a blocker to prevent self-ligation (5′-PO4-TCTCTTCGTGGGCTCTTGCG-23ddC-3′) according to Zhong et al. [45], RT and PCR. Sequences of further primers used for sequencing are displayed in Table S1. Amplicons were cloned into pGEM®-T Vector (Promega Corporation, Fitchburg, Wisconsin, USA) and sequenced. Nucleic acid sequences and ORFs were analyzed by SnapGene (GSL Biotech, San Diego, California, USA) and BLAST on the NCBI website [46]. Alignments of protein sequences and phylogenetic analysis were performed using MEGA X (version 10.2.4; 38,47) with the respective algorithm. The 5’- and 3’-termini of the dsRNA segments of DfPV1 were aligned using the Muscle algorithm [48,49,50,51] in default settings. Alignments for the ML analysis were prepared with the Clustal Omega algorithm [48,52,53,54] in default settings. A bootstrap test was conducted with 1000 replicates for the construction of a ML tree. For DfFV1a and DfFV1b, the model by Le and Gascuel [55] with frequencies and gamma distribution of 5 (LG+G+F) was used. The ML tree of DfPV1, was constructed using the model by Le and Gascuel [55] and a gamma distribution of 5 (LG+G). Phylogenetic analysis was carried out after sequence alignment of the RdRp of virus sequences found by BLASTp with an E-value of 0.0 and were adjusted by hand where necessary. The ML tree of DfFV1a and DfFV1b was rooted with sequences of RdRps of the Magnaporthe oryzae Chrysovirus 1 D/B (MoCV1-D/B) of the Chrysoviridae family [56,57]. The ML tree of DfPV1 was rooted by the use of RdRps of Heterobasidion partitivirus 3 and 12 from the genus Alphapartitivirus [58,59]. Figures were generated and edited by Unipro UGENE (ugene.net, version 1.32.0), INKSCAPE (inkscape.org, version 1.1) and SnapGene. Conserved protein domains were identified by conserved domain database (CDD) search on the NCBI website [60,61,62,63].
2.4. Verification of Virus Presence by RT-PCR
To screen both isolates, NW-FVA 1581 and NW-FVA 1706, for the presence of DfPV1 and DfFV1 by reverse transcriptase PCR (RT-PCR), cDNA was synthesized by using 100 U Maxima H Minus Reverse Transcriptase (ThermoFisher Scientific) with random primers according to the manufacturer’s instructions from dsRNA extracted from mycelium and from VLPs. Presence of DfPV1 was verified by amplification of a 274 bp fragment of the RdRp gene encoded on segment 1 by using primer pair number 1 and 7 (Table S1) and by amplification of a 498 bp fragment from the ORF 1 gene of DfFV1 with primer pair number 8 and 9 (Table S1).
3. Results
3.1. NW-FVA 1581 and NW-FVA 1706 were Determined as D. fraxini and Harbor dsRNAs
The isolates NW-FVA 1581 and NW-FVA 1706 were determined as D. fraxini by the evaluation of the ITS, the TEF1-α and TUB genes. The phylogenetic analysis is displayed in Figure S1. On CMS, both isolates grew at the same speed covering the medium of a 6.5 mm petri dish within 21 days. While the mycelium of isolate NW-FVA 1581 developed aerial and colored hyphae, the mycelium of NW-FVA 1706 produced fewer aerial hyphae without extended pigmentation (Figure 1).
3.2. NW-FVA 1581 is Infected by a Novel Fusagravirus and NW-FVA 1706 Additionally by a Novel Partitivirus
Extraction of dsRNA from mycelium of NW-FVA 1581 and NW-FVA 1706 suggested the presence of a virus in both isolates (Figure S2). Next Generation Sequencing and completion by SPAT showed the presence of three segments of 1,755 bp (dsRNA 1), 1,588 bp (dsRNA 2) and 1,233 bp (dsRNA 3) in isolate NW-FVA 1706 and in both isolates bands of 8,720 bp in NW-FVA 1706 and 8,644 bp in NW-FVA 1581.
The ORF of dsRNA 1 (1755 bp) is flanked by 66 nts at the 5’-NTR and 69 nts at the 3’-NTR. It encodes the protein P1 which consists of 539 amino acids (aa) and a calculated MW of 62.06 kDa (Figure 2C). The viral RdRp of Botryospheria dothidea partitivirus 2 (BdPV2, acc. ID: WFJ08489.1) showed the highest similarity to P1 (80.45 % identical aa, E-value 0.0). Further in silico analysis by CDD search showed RdRp motifs (acc. ID: pfam00680) in the RT_like superfamily (acc. ID: cl02808) being present from position 46 to 492 (E-value 1.41e-85). The ML analysis showed that P1 clusters in the genus Gammapartitivirus together with BdPV2 and BdPV3 and with Aspergillus ochraceous virus (AoV).
The ORF of dsRNA 2 encodes for protein P2, consisting of 435 aa with a calculated MW of 47.18 kDa flanked by 84 nts at the 5’-NTR and 199 nts at the 3’-NTR (Figure 2C). It showed highest similarity to the capsid protein of Penicillium brevicompactum partitivirus 1 (PbPV1, 65.97 % identical aa, E-value 0.0, acc. ID: AYP71817.1) found in sea cucumber Holothuria poli [64]. Due to the BLASTp search, ORF 2 is hypothetically coding for the capsid subunits of the virus particle.
Segment 3 (dsRNA 3) codes for protein P3, consisting of 272 aa and with a calculated size of 30.75 kDa. The ORF is flanked by a 181 nts 5’-NTR and a 233 nts 3’-NTR (Figure 2C). Highest identity was found with the corresponding protein deduced from the unverified sequence of segment 3 of BdPV2 (acc. ID: MZ044012.1) with 70.97 % identical aa. Analysis by BLASTp revealed additional five sequences which produced significant alignments. The highest similarity of P3 was shared with an unknown protein from Aspergillus fumigatus partitivirus 1 (AfPV1, 34.86 % identical aa, E-value 1e-25, acc. ID: CAA7351346.1) and the lowest similarity with a hypothetical protein of Trichoderma harzianum partitivirus 3 (ThPV3, 25.86 % identical aa, E-value 4e-17, acc. ID: WGH72996.1). Even though AoV clusters in the same branch when comparing the aa sequences of the RdRps, it only shows 32.6 % identical aa when comparing P3 (acc. ID: AYP71820.1). For the other virus, BdPV3, which also clusters in the same branch, no third segment was reported.
The heptamer 5´-CGCAAAA-3´ of the extreme 5´-termini and the trimer of the extreme 3´-termini (5´-TCC-3´) are identical in all segments. In segment 3, a 36 nts insertion starts at position 14 which separates a conserved stretch (Figure 2E).
In total, the genome of DfPV1 consists of 4,576 bp. Due to the high similarity to gammapartitiviruses, we denominate the tri-segmented virus as Diplodia fraxini partitivirus 1 (DfPV1). The complete sequences were deposited in GenBank (acc. ID: OR199886 - OR199888).
The complete sequences of the 8,720 bp isolated from NW-FVA 1706 and the 8644 bp isolated from NW-FVA 1581 share 90.36 % identical nts and have a GC content of 51 %. Due to their high similarity, a detailed description will be given to the 8720 bp dsRNA. The dsRNA harbors two discontinuous open reading frames, which may be connected by a -1 slippery heptamer sequence 5’-4645GGAAAAC4651-3’.
The 5´ proximal ORF (ORF 1) starts at position 538, is terminated at position 4,599 and codes for a hypothetical protein (P1) consisting of 1,354 aa with a calculated MW of 150.98 kDa (Figure 2A). A BLASTp search revealed highest similarity (41.96 % identical aa, E-value 0.0) to the hypothetical protein of Macrophomia phaseolina fusagravirus 2 (MpFV2, acc. ID: QK002083.1).
The 3´ proximal ORF (ORF 2) starts at position 5109 and terminates at position 8621. It encodes a protein (P2) of 1,345 aa with a calculated MW of 151.59 kDa (Figure 2A) and showed highest similarity (40.58 % identical aa, E-value 0.0) to the RdRp of Macrophomina phaseolina fusagravirus 3 (MpFV3, acc. ID: QKO02086.1). Conserved motifs of the RT-like superfamily (acc. ID: cl02808) were detected by CDD search between position 513 and 791 (E-value 2.02e-12) suggesting that ORF 2 is coding for the viral RdRp. No Phytoreo_S7 domain was detected by CDD search.
To further analyze the taxonomic position of the two strains of DfFV1, a ML tree was constructed based on the aa sequences of both putative viral RdRps. Both viruses cluster together and build a clade with fusagraviruses found in Macrophomina phaseolina. Therefore, we denominated the viruses as Diplodia fraxini fusagravirus 1a and as Diplodia fraxini fusagravirus 1b. Both sequences were deposited at GenBank (acc. ID: OR224544 and OR228587).
3.3. Single and Double Virus Infections Result in Different Ultra-Structures and dsRNA Patterns
To further analyze the viral structure of DfFV1a/b, VLPs and PSs were purified from 21 d old mycelium from both fungal isolates and subjected to electron microscopy. In isolate NW-FVA 1706, isometric particles with an average size of around 25 nm were detected (Figure 3B). In fungal isolate NW-FVA 1581, exclusively cauliflower-like structures were detected (Figure 3A, left). The same cauliflower-like structures were observed when a protocol for polysome enrichment was applied for mycelium of NW-FVA 1581 (Figure 3A, right), and no structures were detected for NW-FVA 1706 when the same protocol was applied (not shown).
The protein pattern of VLPs and is shown in Figure 3C. In extractions of VLPs of NW FVA 1581, a band in the size of about 90 kDa is present (lane 2). Note, that polysomal structures were purified when the protocol for VLP purification was applied. Protein sequencing of the band observed in the particle preparation resulted in peptides corresponding to the P1 sequence (Figure S3). Polysome extraction revealed a slightly smaller band with the MW of about 80 kDa (lane 4). In addition, a typical band pattern for polysomes is visible below 40 kDa. When extracting VLPs from NW-FVA 1706, two bands with sizes between 55 and 40 kDa are visible, but no band in the range of 80 to 90 kDa (lane 1). Peptides corresponding to P2 of DfPV1 were obtained by protein sequencing (Figure S4). After polysome extraction, no proteins were detected (lane 3).
RNA extraction from VLPs purified from NW-FVA 1581 showed exclusively a band of about 9,000 bp (Figure 4A, lane 2) which corresponds with the segment size of 8,644 bp, four bands corresponding to the sizes of the segments of DfFV1 and DfPV1 were visible at around 9,000 bp and at 1,500 to 2,000 bp, when RNA was extracted from VLPs, purified from NW-FVA 1706 (Fig 4A, lane 1).
A band of about 9,000 bp according to the size of DfFV1b was detected when RNA was extracted from NW-FVA 1581 PSs (Figure 4B, lane 2). In addition, bands in the range from 1,000 to 2,000 bp were detected which we addressed to host rRNA and mRNA. Neither a viral band nor host RNA was present when RNA was derived from PSs from NW-FVA 1706 (Figure 4B; lane 1).
3.4. Single and Double Infections were Verified by RT-PCR from Mycelium and VLPs
To verify the presence of the viruses DfPV1 and DfFV1 in the two fungal isolates, NW-FVA 1581 and NW-NVA 1706, RT-PCR with species specific primers was conducted using dsRNA extracted from fungal tissue and RNA extracted from VLPs. In isolate NW-FVA 1706, both viruses were detected irrespective of the template used, while in isolate NW-FVA 1581 only DfFV1b was detected (Figure 5).
4. Discussion
The two strains, NW-FVA 1581 and NW-FVA 1706, originated from different forest stands of Northern Germany. Although they were preliminary classified as Botryosphaeria stevensii/Diplodia mutila sl. [5], we could unequivocally address them to D. fraxini in our study. In NW-FVA 1581, we detected a single virus infection, in NW-FVA 1706 a double infection and both isolates showed a different phenotype. The single infected NW-FVA 1581 produced numerous aerial hyphae which turned brownish after several days of incubation, while the mycelium of NW-FVA 1706, which harbors the double infection, stayed whitish with reduced aerial hyphae production. Similarly, Alves et al. [65] reported several phenotypes from strains of D. fraxini. However, no data about the virome of these isolates are available. Therefore, the morphotype-development cannot be addressed to the genotype or virome of the two strains.
In NW-FVA 1706, we detected four dsRNAs and addressed the three smaller bands after sequencing to monocistronic segments with sizes of 1,755 bp (segment 1), 1,588 bp (segment 2) and 1,233 bp (segment 3). Protein 1, which is encoded on segment 1, shows RdRp motifs and is closest related to the respective protein of BdPV2. The putative capsid protein, which is encoded on segment 2, is closest related to the respective segment of PbPV1. The rules of the ICTV for the establishment of new species of partitiviruses claim 90 % or less sequence identity for the RdRp and 80 % or less for the capsid protein. According to the rules, the three dsRNAs found in NW-FVA 1706 are segments of a new tri-partite member of the genus Gammapartitivirus within the Partitiviridae family and therefore we denominated it as Diplodia fraxini partitivirus 1 (DfPV1). The ascomycete host D. fraxini and the lack of a 3’ poly(A) additionally correspond with the ICTV demarcation criteria for gammmapartitiviruses. Members of this genus are usually described to be bipartite, however species with a tripartite genome are listed as definite species of the Partitiviridae family and the genome of the putative member Ustilaginoidea virens gammapartitivirus 1 (UvPV-1) is even divided into four segments. According to the 9th report on subviral agents by the ICTV [66], satellite-like nucleic acids are defined to be distinct from their helper virus and are either coding for no or for a non-structural protein. As it was shown for an additional segment (dsRNA 4) for the Fusarium solani alternavirus 1 (FsAV1) [67], the third segment of DfPV1 also has extended NTRs and is therefore distinct from segment 1 and 2. The criterium of encoding either no protein or a non-structural protein is fulfilled since protein patterns of VLPs did not show any band with the expected size of about 30 kDa. Several related proteins to P3 were found by BLASTp which are the corresponding proteins of other tripartite gammapartitiviruses. There is no obvious pattern identifiable which can be linked to the RdRp similarity and the pre- or absence of the third segment regarding to their taxonomical relationship. Within the clade, BdPV2 and BdPV3 are more closely related to DfPV1 than AoV, even though no third segment was described for BdPV3. Due to their close relationship regarding the RdRp and their similar P3, BdPV2 and DfFV1 may have a common ancestor. Additionally, both fungal hosts are members of the Botryosphaeria genus which supports this hypothesis. In contrast, the genus Aspergillus belongs to the class Eurotiomyces and is not related to the genus Botryosphaeria which is accommodated in the class Dothideomycetes. Therefore, the third segment of AoV may have been acquired independently by horizontal gene transfer (HGT) as it was speculated by Wang et al. [68] for a papain-like protease domain on dsRNA 2 of Sclerotinia sclerotiorum megabirnavirus 1 (SsMBV1), for a Phytoreo_S7 domain in non-phytoreoviruses by Liu et al. [24] and from Lutz et al. [67] for segment 4 of FsAV1.
The sequences of the two bands of about 9,000 bp which were detected in both isolates, NW-FVA 1581 and NW-FVA 1706, are putative strains of the same virus since they share 90.36 % identical nts. In the 5´-NTR of NW-FVA 1581, a deletion of 79 nts was detected and verified by RT-PCR. No Phytoreo_S7 domain was detected by CDD search for neither of them.
The putative RdRp showed highest similarity to the MpFV3 (40.58 % identical aa) and clusters within a distinct clade. Due to sequence characteristics as genome size, the putative coding strategy by a -1 frameshift and lengths of the 5´- and 3´-NTRs, we classify the two viruses found in the fungal isolates, NW-FVA 1706 and NW-FVA 1581, as strains of a new member of the proposed family “Fusagraviridae” which was suggested by Wang et al. [14] and denominate them as Diplodia fraxini fusagravirus 1a (NW-FVA 1706) and as Diplodia fraxini fusagravirus 1b (NW-FVA 1581).
A capsidless [18,23] as well as encapsidated [15,17] nature was discussed for fusagraviruses. Our results suggest that the viral RNA of DfFV1 is protected by P1 of DfFV1 together with ribosomal proteins in a single infection. The protection of a viral genome by polysomes was also suggested for the capsidless narnaviruses which encode a RdRp on their (+)ssRNA genome [69]. In an ambigrammatic way, a function-less protein on the complementary (-)ssRNA is translated and is involved to encapsulate and protect both strands [70]. Wilkinson et al. [70] discussed this process to be performed by “frozen polysomes” which are unable to detach from the 3´-terminus, a mechanism which has to be reversible and may be performed by virus encoded proteins. The structures obtained by VLP and PS enrichments from DfFV1 infected NW-FVA 1581 may be based on those “frozen polysomes”. The protein encoded from ORF1 with its unknown function may serve as the factor to support the freezing process and the conversion of the fungal metabolism to prevent stalled polysomes from degradation by no-go decay [71]. A large ORF on the (+)ssRNA and (-)ssRNA is discussed to be necessary for an extended coverage of the complete viral genome [72]. Due to the absence of a large ORF at the complementary strand of DfFV1, we only could give an explanation for the viral (+)ssRNA encapsulation. Since we isolated dsRNA from PS enrichments, this mechanism seems unlikely to be involved. Other unknown factors must be responsible for the stabilization of these polysomal structures in case the encapsulation of the genome by ribosomes is based on the proposed mechanism. Whether the protein encoded on ORF 1 is involved in the switch of translation to freezing and back or whether a completely different mechanism is involved has to be investigated.
The ultra-structures recovered from the double infection of NW-FVA 1706 differ from that of NW-FVA 1581, although both cultures were grown under identical conditions. From mycelium of NW-FVA 1706, no polysomal structures could be isolated, neither in a VLP nor in a PS enrichment. When using the protocol for VLP purification, isometric particles were recovered containing dsRNA from both viruses, DfPV1 and DfFV1. Since no ORF 1-related putative capsid protein was detected, it is unlikely that the protein encoded on ORF 1 builds the viral shell for the fusagravirus genome as it was shown for Cryphonectria carpinicola fusagravirus 1 (CcFGV1) by Das et al. [15]. However, transcapsidation of the replicative from was described as a common feature of members of the Yadokaviridae family with a given distantly related partner [73]. We hypothesize, that the dsRNA of the novel fusagravirus DfFV1 was transcapsidated by partitiviral capsids since exclusively particles, with the typical size for partitivirueses of around 25 nm were detected.
Another co-infection of a fusagravirus and a partitivirus was described from Rosellinia necatrix [74], but no data of the ultra-structure and their RNA content are available yet. It will be interesting whether the fusagravirus genome is transcapsidated by the partitiviral capsid in this infection and whether the fusagraviruses are generally promiscuous and use for the protection of their genome either their own capsid, a capsid from a co-infecting virus or ribosomes together with P1 of DfFV1.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Cornelia Heinze; Funding acquisition, Gitta Jutta Langer and Cornelia Heinze; Investigation, Steffen Bien and Cornelia Heinze; Methodology, Steffen Bien; Project administration, Gitta Jutta Langer and Cornelia Heinze; Supervision, Gitta Jutta Langer and Cornelia Heinze; Writing—original draft, Steffen Bien, Gitta Jutta Langer and Cornelia Heinze; Writing—review & editing, Cornelia Heinze.
Funding
This project is financed by the Agency for Renewable Resources (FNR) in the program “Waldklimafonds“ [Forest and Climate Fund] (2219WK22A4 and 2219WK22G4) funded by the German Federal Ministry of Food and Agriculture and the GermanFederalMinistry for Environment, Nature Conservation and Nuclear Safety.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are openly available in this work and its supplementary material.
Acknowledgments
We thank Elke Woelken for electron microscopy and Birgit Hadeler for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- George, J.-P.; Sanders, T.G.M.; Timmermann, V.; Potočić, N.; Lang, M. European-wide forest monitoring substantiate the neccessity for a joint conservation strategy to rescue European ash species (Fraxinus spp.). Sci. Rep. 2022, 12, 4764. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The Main Species Associated with Cankers and Dieback of Fraxinus excelsior in North-Eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Przybyl, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. Forest Pathol 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Bakys, R.; Vasaitis, R.; Barklund, P.; Thomsen, I.M.; Stenlid, J. Occurrence and pathogenicity of fungi in necrotic and non-symptomatic shoots of declining common ash (Fraxinus excelsior) in Sweden. Eur J Forest Res 2009, 128, 51–60. [Google Scholar] [CrossRef]
- Langer, G. Collar Rots in Forests of Northwest Germany Affected by Ash Dieback. Baltic Forestry 2017, 23, 4–19. [Google Scholar]
- Alves, A.; Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Phillips, A.J.L. The complex of Diplodia species associated with Fraxinus and some other woody hosts in Italy and Portugal. Fungal Diversity 2014, 67, 143–156. [Google Scholar] [CrossRef]
- Ayllón, M.A.; Vainio, E.J. Mycoviruses as a part of the global virome: Diversity, evolutionary links and lifestyle. Adv. Virus Res. 2023, 115, 1–86. [Google Scholar] [CrossRef]
- HOLLINGS, M. Viruses Associated with A Die-Back Disease of Cultivated Mushroom. Nature 1962, 196, 962–965. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M.; Ictv, R.C. ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, J.; Yang, B.; Wang, Q.; Zhou, J.; Yu, W. Molecular Characterization of a Debilitation-Associated Partitivirus Infecting the Pathogenic Fungus Aspergillus flavus. Front. Microbiol. 2019, 10, 626. [Google Scholar] [CrossRef]
- Nerva, L.; Silvestri, A.; Ciuffo, M.; Palmano, S.; Varese, G.C.; Turina, M. Transmission of Penicillium aurantiogriseum partiti-like virus 1 to a new fungal host (Cryphonectria parasitica) confers higher resistance to salinity and reveals adaptive genomic changes. Environ. Microbiol. 2017, 19, 4480–4492. [Google Scholar] [CrossRef] [PubMed]
- Filippou, C.; Coutts, R.H.A.; Stevens, D.A.; Sabino, R.; Kotta-Loizou, I. Completion of the sequence of the Aspergillus fumigatus partitivirus 1 genome. Arch. Virol. 2020, 165, 1891–1894. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wang, X.; Hua, H.; Cao, W.; Zhou, T.; Zhao, C.; Wu, X. Full genome sequence of a new three-segment gammapartitivirus from the phytopathogenic fungus Alternaria tenuissima on cotton in China. Arch. Virol. 2021, 166, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Zhang, H.; Qiu, D.; Guo, L. Two Novel Relative Double-Stranded RNA Mycoviruses Infecting Fusarium poae Strain SX63. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Das, S.; Hisano, S.; Eusebio-Cope, A.; Kondo, H.; Suzuki, N. A Transfectable Fusagravirus from a Japanese Strain of Cryphonectria carpinicola with Spherical Particles. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Cornejo, C.; Hisano, S.; Bragança, H.; Suzuki, N.; Rigling, D. A New Double-Stranded RNA Mycovirus in Cryphonectria naterciae Is Able to Cross the Species Barrier and Is Deleterious to a New Host. J. Fungi (Basel) 2021, 7. [Google Scholar] [CrossRef]
- Sá Antunes, T.F.; Amaral, R.J.V.; Ventura, J.A.; Godinho, M.T.; Amaral, J.G.; Souza, F.O.; Zerbini, P.A.; Zerbini, F.M.; Fernandes, P.M.B. The dsRNA Virus Papaya Meleira Virus and an ssRNA Virus Are Associated with Papaya Sticky Disease. PLoS One 2016, 11, e0155240. [Google Scholar] [CrossRef]
- Spear, A.; Sisterson, M.S.; Yokomi, R.; Stenger, D.C. Plant-feeding insects harbor double-stranded RNA viruses encoding a novel proline-alanine rich protein and a polymerase distantly related to that of fungal viruses. Virology 2010, 404, 304–311. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, Y.; Zhao, H.; Ni, Y.; Liu, X.; Zhao, X.; Wang, G.; Xiao, X.; Liu, H. A novel double-stranded RNA mycovirus that infects Macrophomina phaseolina. Arch. Virol. 2019, 164, 2411–2416. [Google Scholar] [CrossRef]
- Brierley, I.; Pennell, S.; Gilbert, R.J.C. Viral RNA pseudoknots: versatile motifs in gene expression and replication. Nat. Rev. Microbiol. 2007, 5, 598–610. [Google Scholar] [CrossRef]
- Dreher, T.W.; Miller, W.A. Translational control in positive strand RNA plant viruses. Virology 2006, 344, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.L.; Atkins, J.F.; Gesteland, R.F. Programmed ribosomal frameshifting: much ado about knotting! Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 14177–14179. [Google Scholar] [CrossRef] [PubMed]
- Kozlakidis, Z.; Hacker, C.V.; Bradley, D.; Jamal, A.; Phoon, X.; Webber, J.; Brasier, C.M.; Buck, K.W.; Coutts, R.H.A. Molecular characterisation of two novel double-stranded RNA elements from Phlebiopsis gigantea. Virus Genes 2009, 39, 132–136. [Google Scholar] [CrossRef]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Peng, Y.; Yi, X.; Jiang, D. Evolutionary genomics of mycovirus-related dsRNA viruses reveals cross-family horizontal gene transfer and evolution of diverse viral lineages. BMC Evol. Biol. 2012, 12, 91. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Xiang, J.; Zhang, M.; Fu, M.; Yang, Z.; Hong, N.; Wang, G. Characterization of a novel double-stranded RNA mycovirus conferring hypovirulence from the phytopathogenic fungus Botryosphaeria dothidea. Virology 2016, 493, 75–85. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Cao, J.; Yin, X.; Guo, Y.; Guo, L.; Wu, H.; Zhang, M. Characterization of a Novel Mycovirus from the Phytopathogenic Fungus Botryosphaeria dothidea. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Ding, Z.; Zhou, T.; Guo, L.-Y. Characterization of a novel strain of Botryosphaeria dothidea chrysovirus 1 from the apple white rot pathogen Botryosphaeria dothidea. Arch. Virol. 2017, 162, 2097–2102. [Google Scholar] [CrossRef]
- Khan, H.A.; Telengech, P.; Kondo, H.; Bhatti, M.F.; Suzuki, N. Mycovirus Hunting Revealed the Presence of Diverse Viruses in a Single Isolate of the Phytopathogenic Fungus Diplodia seriata From Pakistan. Front. Cell. Infect. Microbiol. 2022, 12, 913619. [Google Scholar] [CrossRef]
- Leach, J.; Lang, B.R.; Yoder, O.C. Methods for Selection of Mutants and In Vitro Culture of Cochliobolus heterostrophus. Microbiology 1982, 128, 1719–1729. [Google Scholar] [CrossRef]
- Damm, U.; Mostert, L.; Crous, P.W.; Fourie, P.H. Novel Phaeoacremonium species associated with necrotic wood of Prunus trees. Persoonia 2008, 20, 87–102. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes--application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal and RNA Genes for Phylogenetics. In PCR protocols: A guide to methods and applications, [Nachdr.]; Innis, M.A., Ed.; Acad. Press: San Diego, Calif., 1994; ISBN 9780123721808. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Lutz, T.; Petersen, J.M.; Yanık, C.; de Oliveira, C.; Heinze, C. Processing of the capsid proteins of the Betachrysovirus Fusarium graminearum virus-China 9 (FgV-ch9). Virology 2021, 563, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Pang, X.D.; Zhu, H.J.; Da Gao, B.; Huang, W.K.; Zhou, Q. Molecular Characterization of a Trisegmented Mycovirus from the Plant Pathogenic Fungus Colletotrichum gloeosporioides. Viruses 2016, 8. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef]
- Li, W.; Cowley, A.; Uludag, M.; Gur, T.; McWilliam, H.; Squizzato, S.; Park, Y.M.; Buso, N.; Lopez, R. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Res. 2015, 43, W580–W584. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics 2004, 5, 113. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, W597–W600. [Google Scholar] [CrossRef] [PubMed]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Higashiura, T.; Katoh, Y.; Urayama, S.-I.; Hayashi, O.; Aihara, M.; Fukuhara, T.; Fuji, S.-I.; Kobayashi, T.; Hase, S.; Arie, T.; et al. Magnaporthe oryzae chrysovirus 1 strain D confers growth inhibition to the host fungus and exhibits multiform viral structural proteins. Virology 2019, 535, 241–254. [Google Scholar] [CrossRef]
- Urayama, S.-I.; Sakoda, H.; Takai, R.; Katoh, Y.; Minh Le, T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. A dsRNA mycovirus, Magnaporthe oryzae chrysovirus 1-B, suppresses vegetative growth and development of the rice blast fungus. Virology 2014, 448, 265–273. [Google Scholar] [CrossRef]
- Kashif, M.; Hyder, R.; de Vega Perez, D.; Hantula, J.; Vainio, E.J. Heterobasidion wood decay fungi host diverse and globally distributed viruses related to Helicobasidium mompa partitivirus V70. Virus Res. 2015, 195, 119–123. [Google Scholar] [CrossRef]
- Vainio, E.J.; Korhonen, K.; Tuomivirta, T.T.; Hantula, J. A novel putative partitivirus of the saprotrophic fungus Heterobasidion ecrustosum infects pathogenic species of the Heterobasidion annosum complex. Fungal Biol. 2010, 114, 955–965. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bryant, S.H. CD-Search: protein domain annotations on the fly. Nucleic Acids Res. 2004, 32, W327–W331. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI's conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: a Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [PubMed]
- Nerva, L.; Forgia, M.; Ciuffo, M.; Chitarra, W.; Chiapello, M.; Vallino, M.; Varese, G.C.; Turina, M. The mycovirome of a fungal collection from the sea cucumber Holothuria polii. Virus Res. 2019, 273, 197737. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.; Correia, A.; Phillips, A. Multi-gene genealogies and morphological data support Diplodia cupressi sp. nov., previously recognized as D. pinea f. sp. cupressi, as a distinct species. Fungal Diversity 2006, 23, 1–15. [Google Scholar]
- The Subviral Agents. Virus Taxonomy; Elsevier, 2012; pp. 1211–1219. [Google Scholar]
- Lutz, T.; Japić, E.; Bien, S.; Langer, G.J.; Heinze, C. Characterization of a novel alternavirus infecting the fungal pathogen Fusarium solani. Virus Res. 2022, 317, 198817. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.; Sun, X.; Cheng, J.; Fu, Y.; Liu, H.; Jiang, D.; Ghabrial, S.A.; Xie, J. Characterization of a Novel Megabirnavirus from Sclerotinia sclerotiorum Reveals Horizontal Gene Transfer from Single-Stranded RNA Virus to Double-Stranded RNA Virus. J. Virol. 2015, 89, 8567–8579. [Google Scholar] [CrossRef]
- Hillman, B.I.; Cai, G. The family narnaviridae: simplest of RNA viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar] [CrossRef]
- Wilkinson, M.; Yllanes, D.; Huber, G. Polysomally protected viruses. Phys. Biol. 2021, 18. [Google Scholar] [CrossRef]
- Matsuo, Y.; Ikeuchi, K.; Saeki, Y.; Iwasaki, S.; Schmidt, C.; Udagawa, T.; Sato, F.; Tsuchiya, H.; Becker, T.; Tanaka, K.; et al. Ubiquitination of stalled ribosome triggers ribosome-associated quality control. Nat. Commun. 2017, 8, 159. [Google Scholar] [CrossRef]
- Dinan, A.M.; Lukhovitskaya, N.I.; Olendraite, I.; Firth, A.E. A case for a negative-strand coding sequence in a group of positive-sense RNA viruses. Virus Evol. 2020, 6, veaa007. [Google Scholar] [CrossRef]
- Sato, Y.; Das, S.; Velasco, L.; Turina, M.; Osaki, H.; Kotta-Loizou, I.; Coutts, R.H.A.; Kondo, H.; Sabanadzovic, S.; Suzuki, N.; et al. ICTV Virus Taxonomy Profile: Yadokariviridae 2023. J. Gen. Virol. 2023, 104. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Kim, D.-H. Co-infection of a novel fusagravirus and a partitivirus in a Korean isolate of Rosellinia necatrix KACC40168. Virus Genes 2021, 57, 121–126. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Diplodia fraxini NW-FVA 1581 and NW-FVA 1706 growing on CMs for 21 days in the dark from the front and the back.
Figure 1.
Diplodia fraxini NW-FVA 1581 and NW-FVA 1706 growing on CMs for 21 days in the dark from the front and the back.
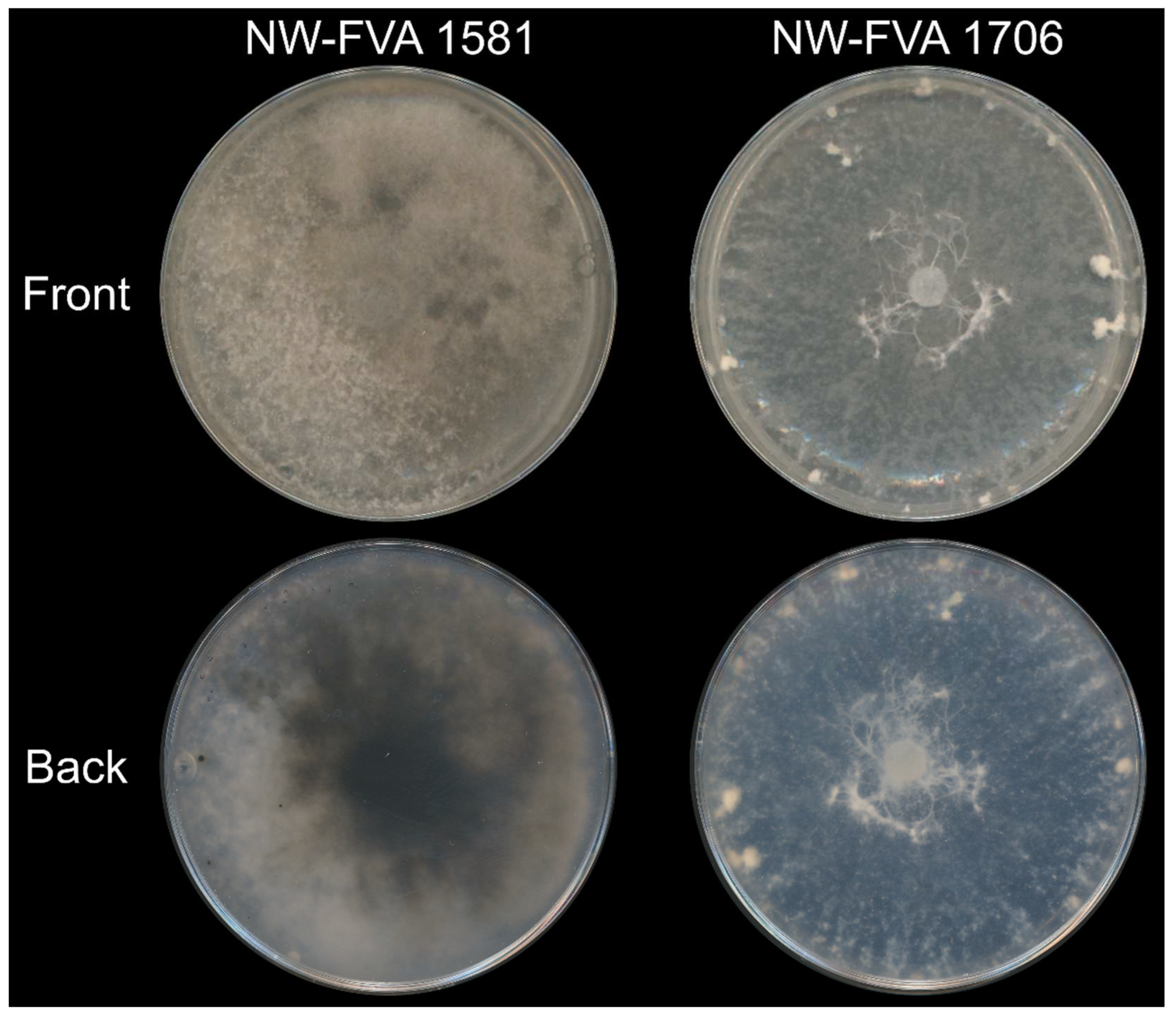
Figure 2.
Genome organization and phylogenetic analysis of DfFV1a/b and DfPV1. The dsRNA segments are displayed as horizontal lines with their respective UTRs at each terminus. ORFs are represented as boxes with start and stop codon positions indicated above and underneath the boxes. Note that the genome organization is not drawn to scale. Maximum-likelihood tree of DfFV1a/b and DfPV1 and selected viruses. 1000 bootstrap replicates were performed, their values are displayed at the nodes. The scale bar corresponds to the genetic distance. The colored dots indicate the novel viruses. A: Genome organization of DfFV1a. B Maximum-likelihood tree of DfFV1a/b. Additionally, the genetic distance is indicated above the interrupted lines. The names of viruses are abbreviated as follows: BcV1: Botrytis cinerea RNA virus 1; DfFV1a/b: Diplodia fraxini fusagravirus 1a/b; FgMV3: Fusarium graminearum dsRNA mycovirus-3; FpFV1: Fusarium poae fusagravirus 1; FpMV1: Fusarium poae mycovirus 1; FpV2/3: Fusarium poae dsRNA virus 2/3; MoCV1 B/D: Magnaporthe oryzae chrysovirus 1; MpFV1-5: Macrophomina phaseolina fusagravirus 1-5;MpV: Macrophomina phaseolina double-stranded RNA; MV-D: Monilinia virus D; PvMV961/962: Phomopsis viticola mycovirus 961/962; RnFV1: Rosellinia necatrix fusagravirus 1; SsFV1/2: Sclerotinia sclerotiorum fusagravirus 1/2; SsMV-L: Sclerotinia sclerotiorum dsRNA mycovirus-L; SsMV-L-WX1/2: Sclerotinia sclerotiorum dsRNA mycovirus-L-WX1/2; TaV1: Trichoderma asperellum dsRNA virus 1; ThV1: Trichoderma hamatum dsRNA virus 1; ThV2: Trichoderma harzianum dsRNA virus 2. C: Genome organization of DfPV1. D: Maximum-likelihood tree of DfPV1. Alpha(α)- and Gamma(γ)partitiviruses are indicated on the right. The names of viruses are abbreviated as follows: BdV2/3: Botryosphaeria dothidea partitivirus 2/3; DfPV1: Diplodia fraxini partitivirus 1; AoV: Aspergillus ochraceous virus; PdPV1: Penicillium digitatum partitivirus 1; PsV-S/F: Penicillium stoloniferum virus S/F; GaRV-MS: Gremmeniella abietina RNA virus MS1; CchPV1: Cordyceps chanhua partitivirus 1; OPV1: Ophiostoma partitivirus 1; DdV1/2: Discula destructiva virus 1/2; FsV1: Fusarium solani virus 1; HPV3/12: Heterobasidion partitivirus 3/12. E: Clipping of the alignments of NTRs of dsRNA segments of DfPV1. The 5’-terminus is displayed at the top and the 3’-terminus at the bottom.
Figure 2.
Genome organization and phylogenetic analysis of DfFV1a/b and DfPV1. The dsRNA segments are displayed as horizontal lines with their respective UTRs at each terminus. ORFs are represented as boxes with start and stop codon positions indicated above and underneath the boxes. Note that the genome organization is not drawn to scale. Maximum-likelihood tree of DfFV1a/b and DfPV1 and selected viruses. 1000 bootstrap replicates were performed, their values are displayed at the nodes. The scale bar corresponds to the genetic distance. The colored dots indicate the novel viruses. A: Genome organization of DfFV1a. B Maximum-likelihood tree of DfFV1a/b. Additionally, the genetic distance is indicated above the interrupted lines. The names of viruses are abbreviated as follows: BcV1: Botrytis cinerea RNA virus 1; DfFV1a/b: Diplodia fraxini fusagravirus 1a/b; FgMV3: Fusarium graminearum dsRNA mycovirus-3; FpFV1: Fusarium poae fusagravirus 1; FpMV1: Fusarium poae mycovirus 1; FpV2/3: Fusarium poae dsRNA virus 2/3; MoCV1 B/D: Magnaporthe oryzae chrysovirus 1; MpFV1-5: Macrophomina phaseolina fusagravirus 1-5;MpV: Macrophomina phaseolina double-stranded RNA; MV-D: Monilinia virus D; PvMV961/962: Phomopsis viticola mycovirus 961/962; RnFV1: Rosellinia necatrix fusagravirus 1; SsFV1/2: Sclerotinia sclerotiorum fusagravirus 1/2; SsMV-L: Sclerotinia sclerotiorum dsRNA mycovirus-L; SsMV-L-WX1/2: Sclerotinia sclerotiorum dsRNA mycovirus-L-WX1/2; TaV1: Trichoderma asperellum dsRNA virus 1; ThV1: Trichoderma hamatum dsRNA virus 1; ThV2: Trichoderma harzianum dsRNA virus 2. C: Genome organization of DfPV1. D: Maximum-likelihood tree of DfPV1. Alpha(α)- and Gamma(γ)partitiviruses are indicated on the right. The names of viruses are abbreviated as follows: BdV2/3: Botryosphaeria dothidea partitivirus 2/3; DfPV1: Diplodia fraxini partitivirus 1; AoV: Aspergillus ochraceous virus; PdPV1: Penicillium digitatum partitivirus 1; PsV-S/F: Penicillium stoloniferum virus S/F; GaRV-MS: Gremmeniella abietina RNA virus MS1; CchPV1: Cordyceps chanhua partitivirus 1; OPV1: Ophiostoma partitivirus 1; DdV1/2: Discula destructiva virus 1/2; FsV1: Fusarium solani virus 1; HPV3/12: Heterobasidion partitivirus 3/12. E: Clipping of the alignments of NTRs of dsRNA segments of DfPV1. The 5’-terminus is displayed at the top and the 3’-terminus at the bottom.
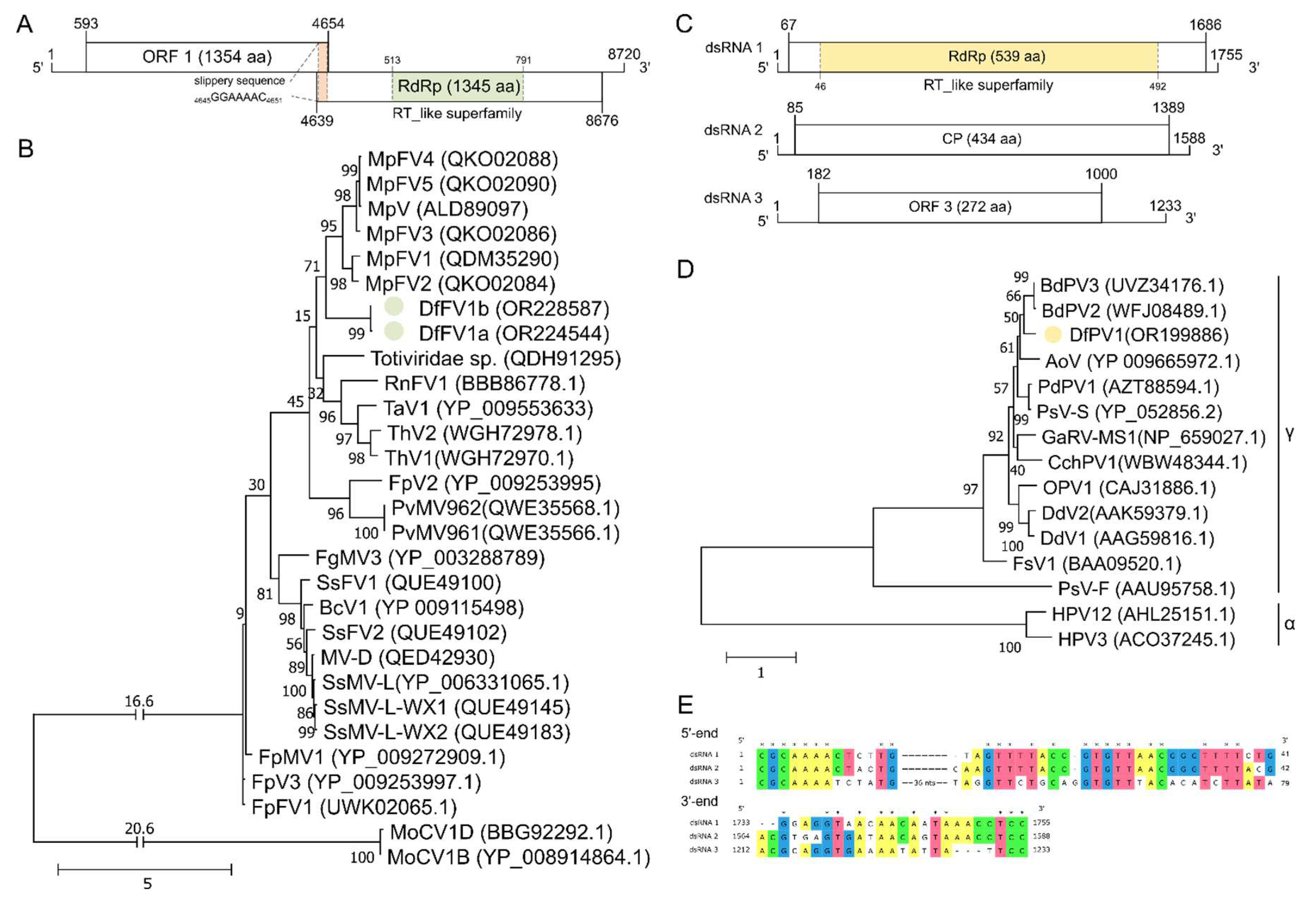
Figure 3.
Electron micrographs of VLPs and PSs examined by transmission electron microscopy contrasted with 2 % /w/v) uranyl acetate and SDS-PAGE (12.5 % w/v) stained with Coomassie Brilliant Blue. The black bar within the electron micrographs corresponds to 100 nm. A: Ultra-structures obtained from mycelium of NW-FVA 1581 using a protocol for VLPs-isolation (left) and protocol for PSs-isolation (right). B: Isometric particles obtained from mycelium of NW-FVA 1706 with a protocol for VLPs-isolation. C: Protein patterns of VLPs and PSs isolations. M, PageRuler Prestained Protein Ladder (Thermo Fisher Scientific). 1, VLPs isolated from NW-FVA 1706. 2, VLPs isolated from NW-FVA 1581. 3, PSs isolated from NW-FVA 1706. 4, PSs isolated from NW-FVA 1581.
Figure 3.
Electron micrographs of VLPs and PSs examined by transmission electron microscopy contrasted with 2 % /w/v) uranyl acetate and SDS-PAGE (12.5 % w/v) stained with Coomassie Brilliant Blue. The black bar within the electron micrographs corresponds to 100 nm. A: Ultra-structures obtained from mycelium of NW-FVA 1581 using a protocol for VLPs-isolation (left) and protocol for PSs-isolation (right). B: Isometric particles obtained from mycelium of NW-FVA 1706 with a protocol for VLPs-isolation. C: Protein patterns of VLPs and PSs isolations. M, PageRuler Prestained Protein Ladder (Thermo Fisher Scientific). 1, VLPs isolated from NW-FVA 1706. 2, VLPs isolated from NW-FVA 1581. 3, PSs isolated from NW-FVA 1706. 4, PSs isolated from NW-FVA 1581.
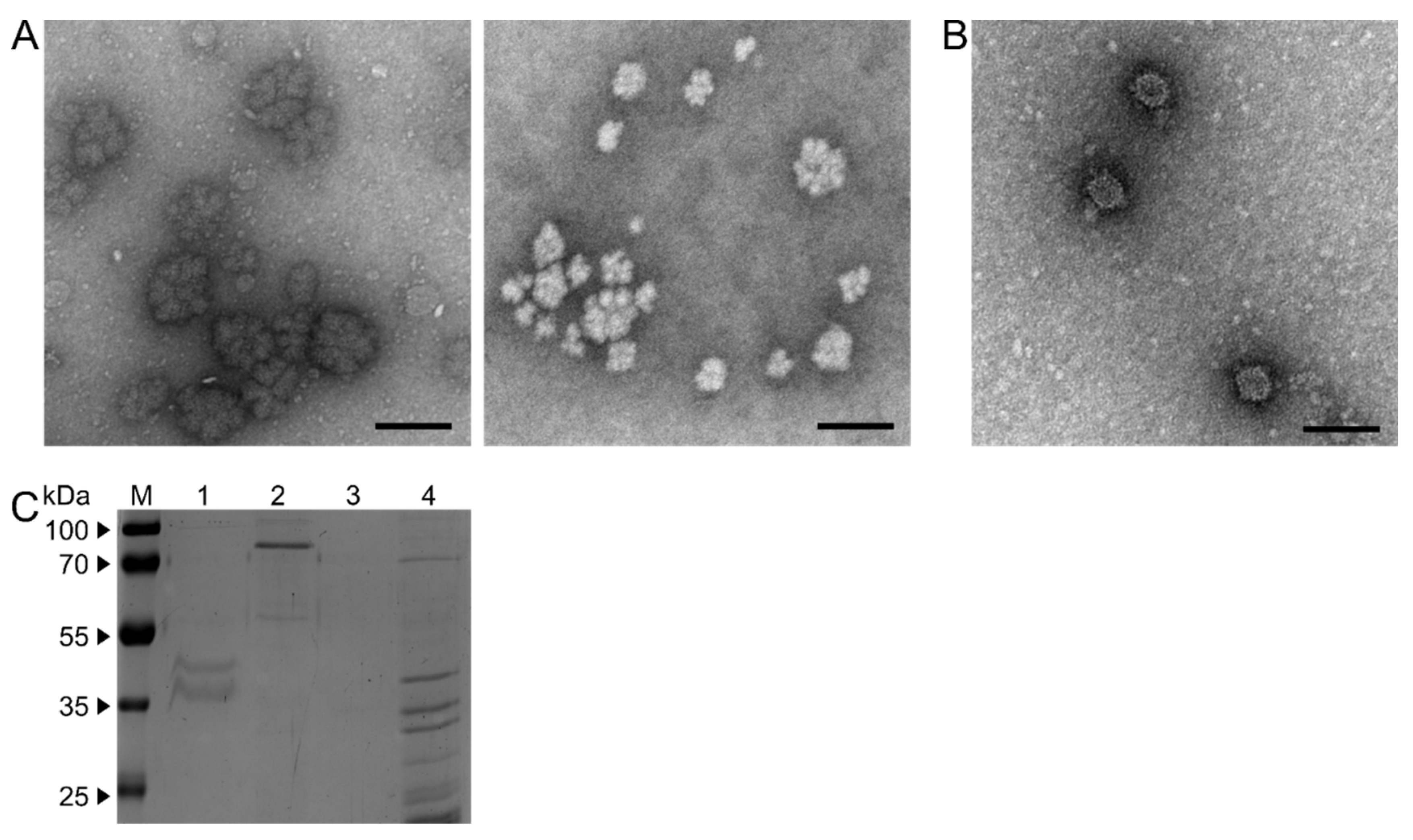
Figure 4.
Agarose gel electrophoresis (1 % w/v) of dsRNA extracted from VLPs and PSs isolated from mycelium of NW-FVA 1581 and NW-FVA-1706. M, GeneRuler 1 kb plus DNA ladder (Thermo Fisher Scientific). The sizes of the marker are given on the left. 1, RNA pattern isolated from NW-FVA 1706. 2, RNA pattern isolated from NW-FVA 1581 A: Viral dsRNA extracted from VLPs. B: Viral RNA extracted from PSs.
Figure 4.
Agarose gel electrophoresis (1 % w/v) of dsRNA extracted from VLPs and PSs isolated from mycelium of NW-FVA 1581 and NW-FVA-1706. M, GeneRuler 1 kb plus DNA ladder (Thermo Fisher Scientific). The sizes of the marker are given on the left. 1, RNA pattern isolated from NW-FVA 1706. 2, RNA pattern isolated from NW-FVA 1581 A: Viral dsRNA extracted from VLPs. B: Viral RNA extracted from PSs.
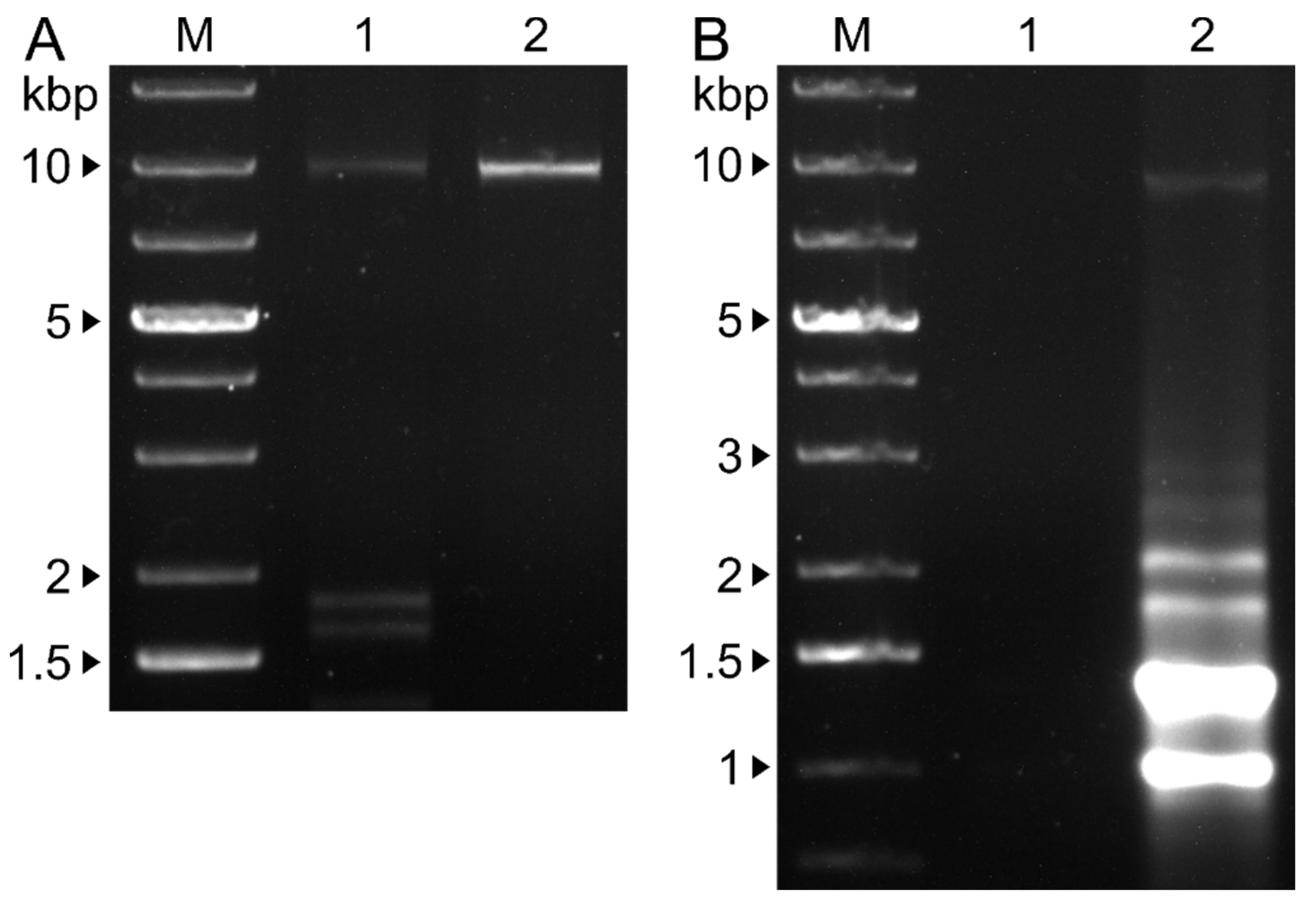
Figure 5.
Agarose gel electrophoresis (1% w/v) of RT-PCR products to detect DfPV1 and DfFV1 in NW-FVA 1581 and NW-FVA 1706 from RNA isolated from VLPs and from dsRNA extraction from mycelium. M, GeneRuler 1 kb plus DNA ladder (Thermo Fisher Scientific). The sizes of the marker are given on the left. When DfFV1 was detected, band sizes of 498 bp, and when DfPV1 was detected, band sizes of 274 bp were expected. The fungal isolates are indicated at the bottom. RNA was isolated from VLPs and used as template in lane 1,2,5,6. RNA was isolated from mycelium and used as template in lane 3,4,7,8. Lane 1: Detection of DfPV1. Lane 2: Detection of DfFV1. Lane 3: Detection of DfPV1. Lane 4: Detection of DfFV1. Lane 5: Detection of DfPV1. Lane 6: Detection of DfFV1. Lane 7: Detection of DfPV1. Lane 8: Detection of DfFV1. Lane 9: Water control RT-PCR for DfPV1. Lane 10: Water control RT-PCR for DfFV1.
Figure 5.
Agarose gel electrophoresis (1% w/v) of RT-PCR products to detect DfPV1 and DfFV1 in NW-FVA 1581 and NW-FVA 1706 from RNA isolated from VLPs and from dsRNA extraction from mycelium. M, GeneRuler 1 kb plus DNA ladder (Thermo Fisher Scientific). The sizes of the marker are given on the left. When DfFV1 was detected, band sizes of 498 bp, and when DfPV1 was detected, band sizes of 274 bp were expected. The fungal isolates are indicated at the bottom. RNA was isolated from VLPs and used as template in lane 1,2,5,6. RNA was isolated from mycelium and used as template in lane 3,4,7,8. Lane 1: Detection of DfPV1. Lane 2: Detection of DfFV1. Lane 3: Detection of DfPV1. Lane 4: Detection of DfFV1. Lane 5: Detection of DfPV1. Lane 6: Detection of DfFV1. Lane 7: Detection of DfPV1. Lane 8: Detection of DfFV1. Lane 9: Water control RT-PCR for DfPV1. Lane 10: Water control RT-PCR for DfFV1.
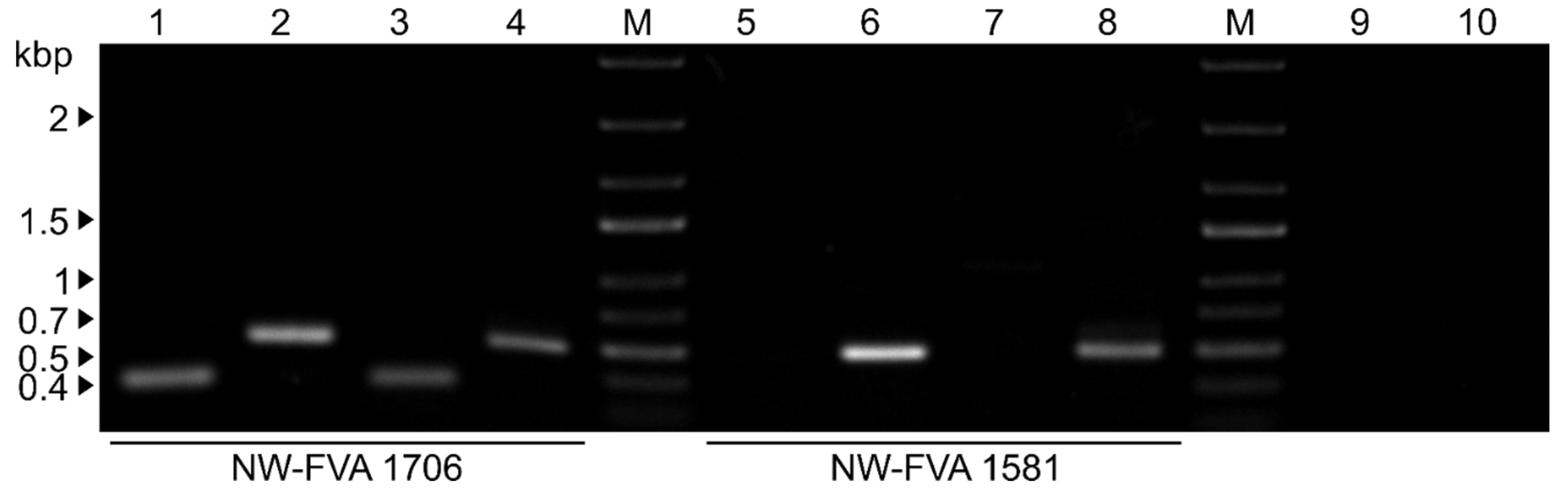

Table 1.
GenBank accession IDs of the sequenced isolates NW-FVA 1581 and NW-FVA 1706.
| Taxon | Isolate | Acc. ID ITS | Acc. ID TEF1-α | Acc. ID TUB |
|---|---|---|---|---|
| D. fraxini | NW-FVA 1581 | OR050980 | OR079892 | OR079888 |
| D. fraxini | NW-FVA 1706 | OR050981 | OR079893 | OR079889 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



