
Submitted:
13 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
Fine aroma cocoa (Theobroma cacao) is one of Ecuador's most iconic export products, representing 63% of world production; however, few advances have been made in fermentation processes that greatly influence the development of chocolate´s organoleptic characteristics. The study of starter cultures has been investigated in other countries, which seek to improve organoleptic properties or decrease fermentation time. The aim of this study was to analyze the effect of a native microbial consortium based on two yeasts (Torulaspora delbrueckii and Hanseniaspora uvarum), a lactic acid bacterium (LAB) (Lactobacillus plantarum) and an acetic acid bacterium (AAB) (Acetobacter ghanensis) inoculated at the beginning of the fermentative process, on physical and biochemical variables, microbial population dynamics, and percentage of almond fermentation. The started culture caused changes in sugar and acid content, which in turn generated temperature and pH changes in the dough, resulting in the dynamics of yeast, AAB, and mesophilic microorganisms remaining higher than those of the control throughout the fermentation process; a decrease of filamentous fungi that affect the flavor and quality of beans due to the production of acetic acid or secondary metabolites from yeasts and LAB; and optimized fermentation percentage by 24% higher than spontaneous fermentation in only 96 hours.
Keywords:
Cocoa Bean Fermentation
; Yeast
; Lactic Acid Bacteria
; Acetic Acid Bacteria
; Initiator Microbial Consortium
1. Introduction
Cocoa (Theobroma cacao) is one of the main traditional Ecuadorian export prod-ucts. According to the National Institute of Statistics and Censuses (INEC), the cocoa sector (dedicated to cocoa) contributes approximately 5% of the economically active population (EAP) and 15% of the rural EAP, becoming the fundamental basis of the economy familiar from the Ecuadorian coast, the foothills of the Andes mountains and the Ecuadorian Amazon. Ecuador occupies the third place of cocoa almond producers worldwide, thus representing 7% of the total world production and leads the production of the variety of fine aroma cocoa with a participation of 63% worldwide. The European Union and the United States are the main export destinations [1]. For this reason, it is essential that the harvest and post-harvest (fermentation and drying) be carried out in the best and most efficient way, generating a quality final product that could be marketed at a better price.
One of the two most important steps to develop the desired chocolate flavors in a cocoa bean is fermentation and drying, in this process the harvested beans can immediately undergo natural fermentation during which microbial action on the mucilaginous pulp produces ethanol and acids, as well as releasing heat. The fermentation of the cocoa bean generally takes 5 to 10 days and is initiated by microorganisms native to the area, which occur naturally in the fermentation sites, inhabiting the surfaces of the pods and the soil, where yeasts have been found, acetic acid bacteria (AAB), lactic acid bacteria (LAB), Bacillus sp. (which interact with each other triggering many physical changes and chemical reactions that promote promising biochemical characteristics in beans) and filamentous fungi (which generate mycotoxins such as ochratoxin and citrinin) that decrease the quality of the cocoa bean [2,3].
In cocoa bean fermentation, two bacterial species: Lactobacillus fermentum and Acetobacter pasteurianus can be considered good candidates to make starter cultures, specifically Lb. fermentum that has characteristics such as heterofermentative metabolism, fructose growth capacity, citrate conversion, mannitol production and acid heat, and ethanol tolerance, characteristics that are desired for starter strains in cocoa fermentation, as well as Bacillus species, which have shown a positive effect on cocoa fermentation [4].
Given the importance of cocoa in Ecuador, quality standards ranging from Ecuadorian INEN standards to European and American standards must be met. The places and countries of destination need the cocoa supply to be sustainable, uniform and of a quality always framed in their standards. This can be achieved by having a correct fermentation process of the cocoa beans [5]. Therefore, this study has the purpose of providing additional information about the use of starter cultures of microorganisms based on autochthonous species of the area, inoculated at the beginning of fermentation, in order to improve this process, either by reducing the fermentation time, increasing the concentrations of organic acids, sugars, ethanol, polyphenol content and possibly decreasing the appearance of filamentous fungi, thus obtaining an improvement in flavor and aroma properties.
To improve the quality of fermentation, recent studies have incorporated starter cultures of microorganisms that could optimize the cocoa fermentation process, for which they should be composed of at least one strain of each group of microorganisms (yeast, LAB and AAB). However, it is necessary to select the appropriate LAB strain to avoid lactic acid overproduction and its negative impact on cocoa bean quality [4]. The aim of this study was to evaluate a native microbial cocktail as a starter culture in the fermentation of the cocoa bean Nacional variety on physical and biochemical variables, the microbial population dynamics of yeast, LAB, AAB, filamentous fungi, and mesophilic microorganisms throughout the fermentation process of almond fermentation.
2. Materials and Methods
2.1. Inoculum Preparation
In a previous study, microbiological, biochemical, and molecular analyses identified the microbiota diversity from fine cocoa bean varieties Criollo (named Nacional) and Trinitario CCN-51 during commercial fermentation [6]. Pure isolates of Torulaspora delbrueckii, Hanseniaspora uvarum (yeasts), Lactobacillus plantarum (lactic acid bacterium - LAB), and Acetobacter ghanensis (acetic acid bacterium - AAB) obtained from the fermentation of cocoa beans in the COFINA company were collected. These were multiplied in different liquid media (without agar): NYDA for yeast, MRS (Man Rogosa Agar) for LAB, and YGC (Glucose Yeast Extract) for AAB, which were autoclaved (121°C, 15 psi for 20 minutes), inoculated in a laminar flow chamber and incubated for 72 hours with orbital shaking at 100 rpm, obtaining final concentrations of 1x108 CFU/ml.
2.2. Geographical Location of the Study
The study was conducted at the production and fermentation facilities of COFINA/República del Cacao, which is located in the province of Los Ríos, canton Vinces, Antonio Sotomayor parish, at an altitude of 17 m.a.s.l., 1°36 '00” S; 79°42'29" W. This area has a humid-tropical climate, annual rainfall of 1,000 to 2,000 mm, temperatures ranging from 24 to 30 °C and relative humidity of 98% and is classified as Tropical Humid Megathermal [7].
2.3. Inoculation and Beans Sampling during Fermentation
At the COFINA/República del Cacao facilities, Nacional cocoa beans were ferment-ed in jute sacks, placed on wooden pallets, both for the treatment inoculated with T. dulbruecki, H. uvarum (yeasts), L. plantarum (LAB) and A. ghanensis (AAB) (Figure 1A), as the non-inoculated treatment (natural fermentation). Every 24 hours, the temperature of the fermenting mass was registered and samples of 150 grams were collected from hour cero (beginning of the fermentation process) to hour 96 (end of the fermentation process). Samples were taken from different areas of the fermenting bean biomass. Each sample was transferred to a plastic bag, coded, and stored at -20°C for chemical and biochemical analysis. Assays were conducted twice.
2.4. Temperature, pH, and Water Content
The fermentation temperature of the cocoa mass was monitored using an IFC 400 digital sensor (MadgeTech, Inc.). The chemical parameters of pH were taken according to the methodologies described by [8], for which 15 g samples of cocoa beans were taken and 135 ml of distilled water was added, to which it was shaken in a vortex for 20 seconds and the supernatant of the sample was measured. The water content (percent of humidity) was determined based on international standards ISO 2291-1972 (drying at 103oC for 16 hours) for the inoculated and non-inoculated bean sample
2.5. Quantification of Population Dynamics
The population dynamics of microorganisms present during fermentation was determined by mixing 20g of beans from the initial 150g cocoa sample with 180 ml of sterile distilled water in sterile plastic bags (Ziploc). Each sample was homogenized for 30 seconds at high speed. Serial dilutions of 10 of the homogenized mixtures were made, of which dilutions 3 and 4 were spread in triplicate dishes with different medium. Microbial enumeration for filamentous fungi and yeasts was performed on malt extract agar and rose bengal agar with chloramphenicol; for LAB on MRS plus agar; for AAB on GYEA agar and for mesophilic microorganisms on NYDA agar. All dishes were incubated at 28 ºC for 1 to 4 days. Yeast, filamentous fungi, AAB, and mesophilic microorganism agar plates were incubated under aerobic conditions, while LAB plates were incubated under anaerobic conditions. After incubation, a colony count was performed.
2.6. Biochemical Characteristics
Samples of 150 g of cocoa beans of the Nacional variety were stored at -80ºC, lyophilized, and later homogenized to analyze by high-performance liquid chromatography (HPLC) (Agilent Technology 1260 Waldbronn, Germany). Changes in the concentration of sugars (sucrose, fructose, and galactose) and organic acids (citric, acetic, lactic, malic, and oxalic acids) were detected by using the methodologies described by [9,10], with modifications.
2.6.1. Sugar Concentration
To measure sugar content, extracts prepared from 500 mg of each sample were made, which were dissolved in 50 ml of MilliQ water and stirred by ultrasound for 20 minutes; 100 milliliters of this solution (pH 13.5 with 0.3 M NaOH), which were mixed with 100 µl of a 0.5 M methanol solution. The samples were incubated in a water bath for 2 hours at a temperature of 70ºC and then the pH was adjusted with a 0.3 M HCl solution. The resulting solution was evaporated to dryness and its residues were mixed with chloroform and shaken vigorously. Chloroform layers were removed and the aqueous residues were eluted in an Eclipse Plus C18 HPLC equipment 250 x 4.6 mm, 5 µm, with a mobile phase of 0.1 M phosphate buffer and acetonitrile in a ratio of 83:17 (v/v,%) at a flow rate of 1ml/min, 20 µl injection volume, 35 ºC and detected by the UV equipment /VIS Detector at a wavelength of 245 nm [10].
2.6.2. Organic Acids Concentration
Organic acid extracts were prepared with 5 g of each lyophilized cocoa sample, dissolved in 60 ml of MilliQ water, and shaken for 10 min. The homogeneous mixture was centrifuged at 5000 rpm for 30 min at 5 ºC, the pellet was washed twice with 20 ml of MilliQ water. The supernatants were centrifuged again for 15 min and filtered through a 0.45 um Millex Millipore membrane. The organic acid extracts were removed with the Eclipse Plus C18 250 x 4.6 mm 5 µm HPLC equipment with a mobile phase of 50 mM KH2PO4 (monopotassium phosphate) (pH=2.8 with phosphoric acid) at a flow rate of 0.7 ml/min, with an injection volume of 10 µl at 20 ºC and detected by the 1260 Infinity II Multiple Wavelength Detector (Agilent 1260 II Array Diode Detector - G7117C) at 210 nm wavelength. The concentration of organic acids was determined by comparison with standard curves, constructed from stock solutions of commercial acids [10].
2.7. Analysis of Data
Data was collected and analyzed by the HPLC equipment software. The concentration of organic acids and sugars was determined by comparing them with standard curves constructed from standard solutions. Data were expressed in milligrams per gram of cocoa dry matter (mg/g dm). Data from two experiments were pooled for analysis.
2.8. Total Polyphenol Content
The total content of polyphenols (TPC) during fermentation was determined by spectrophotometry using the methodology described by [11], with modifications. The 100 µL methanolic extract (from 2 gr of the initial cocoa sample dissolved in 40 ml of methanol) was mixed with 50 µL of Folin-Ciocalteu`s reagent, 750 µL of distilled water and 100 µL of a Na₂CO₃ solution. Reactions were stored in the dark at room temperature. The absorbance was measured after one hour at a wavelength of 760 nm, based on a calibration curve of gallic acid as a reference. The TPC was expressed in milligrams of gallic acid per gram of cocoa dry matter (mg GAE/g dm).
2.9. Fermentation Percentage
One hundred cocoa beans were sampled from different areas of the bag, both from the inoculated treatment and from the natural fermentation, and were cut in half and quickly evaluated to avoid oxidative processes. All beans were taken into account, well fermented which had well-defined internal grooves, which had a brown color (light or dark), without considering partially fermented, slaty, with white spots, moldy, over fermented or unfermented grains [12,13].
2.10. Experimental Design
Two factors were evaluated: (i) type of inoculation (inoculated with T. delbrueckii, H. uvarum; LAB: L. plantarum; AAB: A. ghanensis and not inoculated or natural fermentation) and (ii) fermentation time (0. 24, 48, 72, 96 hours). The experimental unit was a jute bag with 60 kg of cocoa beans and a total of 4 bags were used. The experiment was arranged under a completely randomized design (DCA), in a divided plot (2x5) with 2 repetitions, whose mathematical model is as follows:
Y_ijk=μ+P_i+S_(k(i))+T_j+〖(PT)〗_ij+e_ijk
Where:
Y_ijk = Random variable.
μ = Overall mean.
P_i = Effect of the ith type of inoculant.
S_(k(i)) = Error for the type of inoculant.
T_j = Effect of the i-th hour of fermentation.
〖(PT)〗_ij = Effect of the interaction Type of inoculant x Fermentation time.
e_ijk = Error for fermentation time.
2.11. Statistic Analysis
The variables of temperature, pH of the fermented dough, population dynamics, and total content of polyphenols, sugars, and organic acids were characterized by descriptive statistics (mean and standard deviation). To compare the variables between treatments, an analysis of variance was performed, and comparison tests of means by using the LSD Fisher test at 5%, for treatments, times, and interactions. All analyzes were done using the INFOSTAT software.
3. Results
3.1. Variation of Temperature, pH and Water Content during Fermentation
The treatment with spontaneous fermentation and the treatment with a microbial cocktail started showed an increase in temperature from 0 hours. With spontaneous fermentation, the temperature increased from 27.4 to 44.5°C while with starter culture increased from 24.7°C to 45.4°C, after 96 h of fermentation. Regarding the pH, it increased over time from 3.7 to 4.75 in spontaneous fermentation and from 3.55 to 4.65 in the inoculated treatment (Figure 2).
3.2. Quantification of Population Dynamics during Fermentation
Figure 3 shows that despite the fact that the yeast population was similar at the beginning of the process, both in spontaneous fermentation and in the treatment with a starter culture, in natural fermentation (control) a decrease was observed from 0 hours (3.2x107 CFU/ml) until 96 hours (7.7x16x104 CFU/ml), meanwhile, in the inoculated treatment this decrease was not so marked, observing a reduction in the population from 5.7x107 CFU/ml to 6.5x105 CFU/ml perhaps because the cocktail was composed of two yeasts.
In the treatment with the microbial cocktail, a greater LAB population was observed from 0 to 72 hours (7.9x107 CFU/ml to 2.3x107 CFU/ml), but then decreased to 4.9x106 CFU/ml at 96 hours, while the opposite occurred in the control, that is, a decrease in the population from 0 to 72 hours (4.6x107 CFU/ml to 4.7x106 CFU/ml), to then increase at 96 hours to 2.6x107 CFU/ml (Figure 3).
In the case of AAB, both the control with spontaneous fermentation and the treatment with starter culture showed a decrease in their populations from 0 to 96 hours, from 3.9x107 CFU/ml to 3.1x104 CFU/ml and from 7.1x107 CFU/ml to 1.5x106 CFU/ml, respectively (Figure 3).
Regarding filamentous fungi, it was observed that in the control the population in-creased from 0 to 48 hours from 1.3x105 CFU/ml to 7.7x105 CFU/ml, to later decrease until 72 hours to 8.1x104 CFU/ml, and increase again at 96 (1.8x105 CFU/ml). For its part, in the treatment inoculated with the microbial cocktail, the populations increased from 0 to 24 hours (1.3x105 CFU/ml to 6.4x105 CFU/ml), then they decreased until the end of fermentation (96 hours) to 1.5x103 CFU/ml (Figure 3).
No effect was observed in the treatment with the microbial cocktail on the popula-tion of mesophilic microorganisms, presenting populations between 5.1x107 CFU/ml and 1.1x107 CFU/ml throughout the fermentation process (Figure 3).
3.3. Variation in Concentrations of Sugars, Organic Acids, and Polyphenols
The analyzes of sugars and ethanol, organic acids, and shell-mucilage polyphenols during the cocoa shell-mucilage bean fermentation are shown in Figure 4. After opening the pods were 47.1 ± 5.6 mg g−1 dry matter (DM) for glucose, 31.3 ± 8.4 mg g−1 DM for fructose, 18.8 ± 2.9 mg g−1 DM for sucrose and 13.8 ± 1.6 mg g−1 MS for ethanol.
Although there were no significant differences for the treatment per hour interaction, a higher sucrose consumption was observed when it was inoculated with the starter culture, obtaining final values of 8.48 mg/g ms compared to the control (Figure 4).
The glucose content decreased substantially during the fermentation process in the control, starting with a concentration of 4.72 mg/g ms, reaching complete consumption at 48 hours, while the treatment with the microbial cocktail began with a concentration of 5.33 mg/g ms and decreased to zero after 72 hours (Figure 4).
The fructose content decreased from 0 to 72 hours, from 31.3 to 19.4 mg/g dm and from 39.6 to 31.5 mg/g dm respectively, for both the control and the treatment with the starter culture, from that moment on in both cases an increase was observed up to 20.0 mg/g ms in the control and 32.1 mg/g ms in the treatment (Figure 4).
No significant differences were found for the treatment*hour interaction in ethanol content, but an effect of treatments and hour was found separately. An increase was observed both in the control and in the inoculated treatment with values of 0.575 mg/g ms and 0.599 mg/g ms respectively, at 96 hours (Figure 4).
Despite the fact that no significant differences were found in the lactic acid content for the interaction treatment per hour, the highest content had the control at 72 and 96 hours (15.3 and 15.3 mg/g m/s, respectively), while that treatment with starter culture was 12.93 and 12.53 mg/g ms at 48 and 96 hours, respectively.
The transformation of malic acid to lactic acid and CO2 is conducted by some LAB such as Lb. plantarum [14]. There were also no significant differences for the interaction treatment per hour, in the content of acetic acid, but it was observed that both the treatment and the control showed two peaks with greater production of this acid, at 48 and 96 hours, with values of 1.23 and 2.77 mg/g ms for the control and 2.35 and 3.30 mg/g ms for the treatment, respectively (Figure 4). The oxalic acid content presented a higher concentration in the inoculated treatment at 96 hours (11.7 mg/g ms) compared to the control, which despite having its highest value at 72 hours (14.0 mg/ g ms) subsequently decreased at 96 hours (8.6 mg/g ms). For malic acid, the inoculated treatment obtained a concentration of 4.6 mg/g ms at 96 hours, while the control, as with oxalic acid, its highest concentration was found at 72 hours (3.5 mg/g ms), to then decrease to 2.6 mg/g more at 96 hours (Figure 5).
No significant effect was found for the interaction treatment per time on the polyphenol content, but a peak could be observed at 48 hours (17.2 mg AG/g dm) in the control, however at the end of the fermentation process (96 hours), the treatment with the starter microbial cocktail showed higher content of polyphenols (16.9 mg AG/g dm), compared to 11.2 mg AG/g dm of control (Figure 6). Our values are much lower than those reported by [15], who reported that for Nacional cacao there was 44.29 mg/g on the first day of fermentation, decreasing to 36.78mg/g on the last day. This lower content may be due to the harvest time, and the age of the plant, however, low polyphenol contents generate chocolates with less astringency and bitterness [16,17].
3.4. Fermentation Percentage
After 96 hours of fermentation, the percentage of fermented beans in the treatment with spontaneous fermentation was 57.3%, while the treatment inoculated with the starter culture obtained 77.0%, which is 19.7% more fermented beans (Figure 7).
When carrying out the cut test of the cocoa beans after their fermentation process, the treatment inoculated with the starter culture showed 24% more fermented beans (cotyledons with deep and brown striations and/or cotyledons with no striations, deep and slightly purple in color) (Figure 1C,D), 5% fewer purple beans, 15% fewer slaty beans (unfermented beans with a gray-black or greenish color inside) and no moldy beans (Figure 1E), compared with natural fermented cocoa beans (Table 1), according to the Ecuadorian Technical Regulations for cocoa beans NTE INEN 176 for Arriba Superior cocoa (CBAS) [12].
4. Discussion
4.1. Variation of Temperature, pH and Water Content during Fermentation
Cocoa fermentation at a pH of approximately 5.0 shows cocoa with the highest potential for specific aromas [18]. The pH values found in this study coincide with those previously reported [19], which found initial values of 3.7-3.9 that progressively increased to 4.2-4.9 both in natural fermentations and inoculated with microorganisms. The assimilation of citric acid by microorganisms causes the pH to increase [20]. The degree and time of acidification of the cotyledon during the fermentation process [21] together with changes in temperature, generated by a succession of various microorganisms (yeasts, LAB, AAB, and Bacillus sp.), create an optimal environment inside the cocoa bean for enzymes such as endoprotease, carboxypeptidase, invertase and glycosidases to catalyze reactions that generate flavor precursors substances [22,23].
4.2. Quantification of Population Dynamics during Fermentation
The yeasts showed their maximum population growth between 0 and 24 hours for both the inoculated treatment and the control (Figure 3), values that exceeded those reported by [15], who indicate values between 6.69 and 5.67 units. logarithmic between 0 and 96 hours. Other authors point out that the highest population peak (1x107 - 1x108 CFU/g) of these microorganisms occurs between the first 12 to 36 hours, and then drops to around 1x103 CFU/g. This great difference is due to the fact that as the fermentation process progresses, the sources of nutrients for these micro-organisms are depleted, in addition, high temperatures are generated, a fact that produces some acids that inhibit their multiplication [24,25]. Other authors observed that, by inoculating starter populations of yeasts, LAB, and BAA, populations of 1x1010 CFU/g were obtained at 0 hours to subsequently reduced the population by 50% at 24 hours, and 1x104 CFU/g at 96 hours [26], corroborating the tendencies found in this study.
The trends in the LAB population of this study (Figure 3) were similar to those observed in another study [26], both for natural fermentation and for the treatment inoculated with the microbial cocktail, despite the fact that in this study [26], the treatment with LAB began with low densities of 1x106 CFU/g, which after 50 hours began to decrease progressively, while in the control an increase was observed after 96 hours. Other workers have shown that when a microbial consortium is inoculated, an increase in the population is observed during the first 2 days (1x109 CFU/g) and then drops to 1x102-3 CFU/g after 72 hours, while in natural fermentation the population starts with 1x102-3 CFU/g and then in-creases to 1x107 CFU/g and then decreases from the fourth day [24,25]. This group of bacteria plays a very important role since they ferment the sugars present in the pulp to lactic acid, acetic acid, and ethanol, as well as metabolize citric acid, producing lactic acid, and acetic acid, among other functions [9].
For AAB, our results showed a similar trend to those obtained by [15], who used wooden boxes -another method commonly used for spontaneous fermentation- which is used for the daily turning of the bean in the fermentation process. Under these conditions they obtained populations greater than 1 x106 CFU/g during the entire process, observing a slight increase at 72 hours. This was perhaps due to the fact that from this moment the environmental conditions within the mass become favorable for the development of these microorganisms. In other studies, when using a starter culture based on yeast, LAB, and AAB, inoculated at time zero, it was observed an increase in the AAB populations up to 72 hours, to then decrease rapidly [25,26]. However, when inoculating with two species of Torulaspora delbrueckii yeasts a decrease in the AAB population from zero to 96 hours was observed to later increase its population [27]. Despite the fact that in this study AAB was also inoculated, its population did not thrive. This species does not have the best characteristics that allow it to dominate over AAB populations, such as the dependence on glycerol and mannitol or lactate as energy sources, characteristics that Acetobacter pasteurianus does show [28]. In addition, this drop in the population could be due to the lack of aeration in the bag fermentation system, a condition that hinders the growth of these microorganisms [26].
Fungi such as Aspergillus niger, Aspergillus carbonarius, and Aspergillus ochraceus produce certain mycotoxins that decrease the quality of the cocoa beans and possibly have adverse effects on the final consumer [29,30]. However, with the microbial cocktail inoculation the population of filamentous fungi decreased until the end of fermentation (Compare data of Figure 2 with Figure 1F). Species such as Lactobacillus sp. and Hanseniaspora sp., do not allow the development of filamentous fungi, because as a result of their metabolism, they generate acetic and lactic acid and other substances that affect the initial quantity of filamentous fungi that usually appear in the bean drying and storage stage [31]. Additionally, the starter cultures increase the organoleptic properties of the cocoa bean [29,30]. In practice, producers associate "good fermentation" with the brown color of the almond, the presence of cracks, brittle cotyledons, the almond separates easily from the shell as shown in Figure 1C,D.
No effect was observed on mesophilic microorganisms with started microbial cocktail inoculations (Figure 3). Although the participation of mesophilic microorganisms in cocoa fermentation has not been studied in detail, these microorganisms produce a range of chemical compounds such as 2, 3-butanediol, pyrazines, acetic acid, lactic acid, and short-chain fatty acids which can contribute to acidity and perhaps off-flavors in fermented cocoa beans [26,32]. According to the European Food Safety Authority, the presence of species of the genus Bacillus sp., in the different cocoa postharvest processes could be an indicator of poor hygiene conditions that should be avoided [33].
4.3. Variation in Concentrations of Sugars, Organic Acids, and Polyphenols
Glucose consumption decreased substantially to 15.1 ± 1.6 mg g−1 after the first 24 hours. A higher consumption of sucrose was observed when it was inoculated with the starter culture, obtaining final values of 8.48 mg/g ms compared to 11.24 mg/g ms of the control (Figure 4). The sugar concentration we obtained was lower than the values usually found, which usually amount to 100 mg g−1 [34]. We did not detect sucrose, which contributed in part to the low total sugar content. This was indicative of an advanced ripening of the fruit [35]. Another substrate found was 5.9 ± 3.8 mg g−1 (WM) of citric acid. The decrease in sucrose is due to enzymatic processes (action of invertase enzymes) of hydrolysis, producing reducing sugars, mainly glucose, and fructose, which are considered flavor and aroma precursors, involved during roasting in Maillard’s non-enzymatic browning reactions to generate some specific aromatic components [36]. In similar studies, where various microorganisms (AAB, LAB, and yeast) were inoculated, the sucrose content was close to zero during the first 96 hours of fermentation [37], while others such as did not obtain a higher consumption of this sugar when inoculating with Saccharomyces cerevisae var. Chevalieri, reporting initial values of 8mg/g and final values of 7mg/g [38]. In this investigation, the yeasts H. uvarum and T. delbrueckii were used because in previous studies was observed that they have a preference for consuming glucose [6], although they can also oxidize sucrose. In addition, H. uvarum is a species with high potential due to it can generate a greater amount of invertase enzyme, which would produce a greater increase in the amount of total solids at the end of cocoa bean fermentation [39].
Glucose is one of the main carbohydrate sources for yeasts. Consumption of 83% of this sugar at 120 hours has been reported in natural fermentations [40], Likewise, inoculations with different microorganisms to the cocoa bean fermentation process, report values with a high glucose consumption up to 72 hours, reaching values close to zero in the beans [38], facts that agree with this study. On the other hand, an increase in glucose concentration was observed that is due to sucrose hydrolyzation [39]. In our study, this increase in concentration could not be observed since yeast species were inoculated that, among other characteristics, efficiently consume or oxidize this sugar [40].
Glucose and fructose are important in the fermentation process since they contribute to the flavor, by reacting with peptides and free amino acids (tyrosine, alanine, phenyl-alanine, and leucine), generating chocolates with "sweet notes" [41]. Some strains of bacteria such as L. plantarum, used in this study, consume fructose and glucose, and like L. fermentum do produce a greater amount of lactic acid [42]. This may explain the low consumption of fructose in the treatment compared to natural fermentation since inoculating these species affected the balance of microorganisms and could cause a decrease in certain microorganisms such as L. fermentum that possess groups of genes involved in the uptake and consumption of fructose and/or sorbose [42,43].
The increase in ethanol concentration was very low compared to 6, 12, and 40 mg/g of ethanol, reported by [9,25,43], respectively. This is mainly due to the low glucose content that was present from the beginning of the process, this being the main source for the generation of ethanol, mainly by yeasts [20]. In addition, the increase in ethanol concentration could be produced by some strains of heterofermentative lactic acid bacteria, when LAB species and no yeast are inoculated [44].
For acetic acid, a higher content was found at 96 hours (3.03 mg/g ms), this is due to the action of microorganisms such as BAA that oxidize both lactic acid and ethanol produced by LAB and yeast, respectively, to produce this acid, a fact so essential for the death of the embryo and generate cocoa with a high level of acidity. The acetic acid content influences the populations of filamentous fungi, causing them to decrease (Figure 3) [22]. Oxalic and malic acids that are produced by yeasts and some bacteria would contribute to the flavor, giving it more refreshing tones, and improving the aroma [45], added to the fact that it has antimicrobial properties to control the deterioration of food quality caused by microorganisms [46].
44. Fermentation Percentage
The percent of fermented cocoa bean agree with other studies where inoculated 0.5 g of yeast (Saccharomyces cerevisiae) and 0.24 g of bacteria (Acetobacter aceti) for every 100 g of cocoa obtaining 72% of fermented cocoa beans [47], or with inoculations of two yeasts, one AAB and one LAC obtained 81% fermentation at 120 h, with a pH of 4.71 [48].
A short fermentation time of 4 days, instead of the traditional fermentation period (6 days), is sufficient to obtain cocoa with the potential to develop a flavor of quality [22]. In this sense, inoculation with our native starter culture would go to the drying stage, shortening the fermentation process (Figure 1E).
New research is underway with the aim of evaluating new consortia of native microorganisms to increase organoleptic characteristics or generate products with specific flavors and aromas for specific markets.
5. Conclusions
A native microbial cocktail as a starter culture changes in the content of sugars and organic acids generated changes in temperature and pH in the fermented beans mass, this resulted in the dynamics of microorganisms being affected, observing that the populations of yeasts, AAB and mesophilic microorganisms remain higher than those of the control with spontaneous microbiota throughout the process. The exception was for LAB, which decreased after 72 h, and filamentous fungi population that decrease after 24 h up to 96 h by 79% in the inoculated treatment, compared to spontaneous fermentation, due to the production of acids (acetic) or metabolites of yeasts and LAB. Eventually, the addition of the inoculum increased the fermentation percentage by around 24% compared to natural fermentation, allowing the high-flavor variety cocoa beans to be suitable for the drying stage in merely 96 hours, which allows the maintenance of reducing sugars and acids that possibly improve organoleptic characteristics of the final product.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, C.F.S. and V.Y.; methodology, C.F.S., V.Y., R.H., D.C.; formal analysis, C.F.S., V.Y.; investigation, C.F.S., V.Y., R.H., D.C.; resources, C.F.S., V.Y.; writing—original draft preparation, C.F.S., V.Y.; writing—review and editing, C.F.S.; supervision, C.F.S., V.Y.; project administration, V.Y.; funding acquisition, V.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research work was funded by Universidad de Las Américas – UDLA through research project AGR.VYM.18.04 and Universidad de las Fuerzas Armadas – ESPE (logistic support).
Data Availability Statement
Data and results can be found at the following links: https://drive.google.com/drive/folders/0AH3tnwwxr0PzUk9PVA (upon request).
Acknowledgments
We would like to thank to Ministerio del Ambiente, Ecuador, for authorization to genetic resources access through Contrato Marco Nro. MAE-DNB-CM-2018-0101.
Conflicts of Interest
The authors declare that they have no known competing financial interest or personal relationship that could have appeared to influence the work reported in this paper.
References
- Ficha Sectorial, Cacao y Chocolate, Subgerencia de análisis de productos y servicios, Corporación Financiera Nacional (CFN). 2022. Available online: https://www.cfn.fin.ec/wp-content/uploads/downloads/biblioteca/2022/fichas-sectoriales-2-trimestre/Ficha-Sectorial-Cacao.pdf.
- Fowler, M.; Coutel, F. Cocoa beans: From tree to factory. In Beckett’s Industrial Chocolate Manufacture and Use, 5th ed.; Beckett, S., Fowler, M., Ziegler, G., Eds.; John Wiley & Sons Ltd., 2017. [Google Scholar] [CrossRef]
- Ganeswari, I.; Khairul Bariah, S.; Amizi, M.; Sim, K. Effects of different fermentation approaches on the microbiological and physicochemical changes during cocoa bean fermentation. Int. Food Res. J. 2015, 1, 70–76. [Google Scholar]
- Figueroa, C.; Mota, J.F.; Hernández, Z.; González, O.; Cocolin, L.; Suárez, M. The challenges and perspectives of the selection of starter cultures for fermented cocoa beans. Int. J. Food Microbiol. 2019, 301, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, G. Evaluación de la influencia del proceso de beneficio del cacao (Theobroma cacao) CCN-51 de altura en su calidad final, mediante el análisis físico, físico-químico y sensorial. Universidad Central del Ecuador. 2018. Available online: http://www.dspace.uce.edu.ec/bitstream/25000/16624/1/T-UCE-0008-CQU-044.pdf.
- Yánez-Mendizábal, V.; Falconi, C.E.; Palacios, H. Identification of microbial population in two fine cocoa varieties during the commercial fermentation process. Food Microbiol. Saf. accepted.
- Dutan, S.; Amaya, J.; Fredy, N.; Palacios, B. Plan de desarrollo y ordenamiento territorial 2015-2020. Gobierno Autónomo Descentralizado del cantón Vinces. 2015. Available online: http://app.sni.gob.ec/sni-link/sni/PORTAL_SNI/data_sigad_plus/sigadplusdiagnostico/1260001030001_Diagnostico%20Territorial%20del%20Cant%C3%B3n%20Vinces%202015_2020_04-04-2016_15-59-59.pdf.
- Gálvez, S.; Loiseau, G.; Paredes, J. Study on the microflora and biochemistry of cocoa fermentation in the Dominican Republic. Int. J. Food Microbiol. 2007, 114, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.T.; Zhao, J.; Graham, F. Yeasts are essential for cocoa bean fermentation. Int. J. Food Microbiol. 2014, 174, 72–87. [Google Scholar] [CrossRef]
- Nour, V.; Trandafir, I.; Ionica, M. HPLC Organic Acid Analysis in Different Citrus Juices under Reversed Phase Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 44–48. [Google Scholar]
- Carrillo, L.; Londoño, J.; Gil, A. Comparison of polyphenol, methylxanthines and antioxidant activity in Theobroma cacao beans from different cocoa-growing areas in Colombia. Food Res. Int. 2014, 60, 273–280. [Google Scholar] [CrossRef]
- INEN, INEN. NORMA TÉCNICA ECUATORIANA NTE INEN 176 Quinta revisión. 2018. Available online: https://www.normalizacion.gob.ec/buzon/normas/nte_inen_176-5.pdf.
- Coexca. Instructivo control calidad/Coexca 2017. Available online: https://www.swisscontact.org/_Resources/Persistent/d/3/b/f/d3bfbb5a8d042f05cbf5533494e288f2c 52800b8 /Guia_de_buenas_practicas_de_poscosecha.pdf.
- Mayo, B.; Flórez, A. Lactic Acid Bacteria: Lactobacillus plantarum. In McNamara, Encyclopedia of Dairy Sciences, 3rd ed.; Paul, L.H., McSweeney, John, P., Eds.; Academic Press, 2022; pp. 206–217. [Google Scholar] [CrossRef]
- Muñoz, A.; Gómez, S. Análisis comparativo de la diversidad microbiana y la producción de compuestos bioquímicos de cacao (Theobroma cacao L.) Variedades nacional y trinitario ccn-51 durante la fermentación. Graduate Thesis, Universidad de las Américas, 2020. Available online: http://dspace.udla.edu.ec/handle/33000/12186.
- Sharmistha, S.; Tanmay, S.; Runu, C.; Maksim, R.; Mohammad, A.S.; Muthu, T.; Kannan, R.R.R. Dark chocolate: An overview of its biological activity, processing, and fortification approaches. Curr. Res. Food Sci. 2022, 5, 1916–1943. [Google Scholar] [CrossRef]
- Colonges, K.; Seguine, E.; Saltos, A.; Davrieux, F.; Minier, J.; Jimenez, J.C.; Lahon, M.C.; Calderon, D.; Subia, C.; Sotomayor, I.; et al. Diversity and determinants of bitterness, astringency, and fat content in cultivated Nacional and native Amazonian cocoa accessions from Ecuador. Plant Genome 2022, 15, e20218. [Google Scholar] [CrossRef]
- Janek, K.; Niewienda, A.; Wöstemeyer, J.; Voigt, J. The cleavage specificity of the aspartic protease of cocoa beans involved in the generation of the cocoa-specific aroma precursors. Food Chem. 2016, 211, 320–328. [Google Scholar] [CrossRef]
- Cempaka, L.; Aliwarga L y Purwo, S. Dynamics of Cocoa Bean Pulp Degradation during Cocoa Bean Fermentation: Effects of Yeast Starter Culture Addition. J. Math. Fundam. Sci. 2014, 46, 14–25. [Google Scholar] [CrossRef]
- Nielsen, D. The microbiology of Ghanaian cocoa fermentations; Denmark: Department of Food Science, The Royal Veterinary and Agricultural University, Denmark, 2006. [Google Scholar]
- Voigt, J. Chocolate and Cocoa Aroma. In Chocolate in Health and Nutrition; Humana Press, 2013; pp. 89–93. [Google Scholar] [CrossRef]
- Santander, M.; Rodríguez, J.; Vaillant, F.; Escobar, S. An overview of the physical and biochemical transformation of cocoa seeds to beans and to chocolate: Flavor formation. Crit. Rev. Food Sci. Nutr. 2019, 60, 1593–1613. [Google Scholar] [CrossRef] [PubMed]
- Sârbu, I.; Csutak, O. The microbiology of cocoa fermentation. Caffeinated Cocoa Based Beverages 2019, 8, 423–446. [Google Scholar] [CrossRef]
- Sandhya, M.V.S.; Yallappa, B.S.; Varadaraj, M.C.; Puranaik, J.; Jaganmohan Rao, L.; Janardhan, P.; Murthy Pushpa, S. Inoculum of the starter consortia and interactive metabolic process in enhancing quality of cocoa bean (Theobroma cacao) fermentation. LWT Food Sci. Technol. 2016, 65, 731–738. [Google Scholar] [CrossRef]
- Saunshi, Y.P.; Sagar Sandhya, M.V.; Rastogi, N.K.; Murthy, P.S. Starter consortia for on-farm cocoa fermentation and their quality attributes. Prep. Biochem. Biotechnol. 2020, 50, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Schwan, R.F. Cocoa Fermentations Conducted with a Defined Microbial Cocktail Inoculum. Appl. Environ. Microbiol. 1998, 44, 1477–1483. [Google Scholar] [CrossRef]
- Visintin, S.; Ramos, L.; Batista, N.; Dolci, P.; Schwan, F.; Cocolin, L. Impact of Saccharomyces cerevisiae and Torulaspora delbrueckii starter cultures on cocoa beans fermentation. Int. J. Food Microbiol. 2017, 257, 31–40. [Google Scholar] [CrossRef]
- Illeghems, K.; Pelicaen, R.; De Vuyst, L.; Weckx, S. Assessment of the contribution of cocoa-derived strains of Acetobacter ghanensis and Acetobacter senegalensis to the cocoa bean fermentation process through a genomic approach. Food Microbiol. 2016, 58, 68–78. [Google Scholar] [CrossRef]
- Ramos, N.; Castro, A.; Juárez, J.; de la Cruz, O.; Rodríguez, N.; Blancas, J.; Navarro, A. Evaluación de ocratoxina a en Theobroma cacao l. “cacao blanco” durante el proceso de cosecha, fermentado, secado y almacenado. Rev. Soc. Química Perú. 2016, 82, 1810–634X. [Google Scholar]
- Ruggirello, M.; Nucera, D.; Cannoni, M.; Peraino, A.; Rosso, F.; Fontana, M.; Cocolin, L.; Dolci, P. Antifungal activity of yeasts and lactic acid bacteria isolated from cocoa bean fermentations. Food Res. Int. 2019, 115, 519–525. [Google Scholar] [CrossRef]
- Romanensa, E.; Freimüller, S.; Volland, A.; Stevens, M.; Krähenmann, U.; Isele, D.; Fischer, B.; Meile, L.; Schwenninger, S. Screening of lactic acid bacteria and yeast strains to select adapted antifungal co-cultures for cocoa bean fermentation. Int. J. Food Microbiol. 2019, 290, 262–272. [Google Scholar] [CrossRef]
- Schwan, R.; Wheals, A. The Microbiology of Cocoa Fermentation and its Role in Chocolate Quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Bastos, V.S.; Santos, M.F.S.; Gomes, L.P.; Leite, A.M.O.; Paschoalin, V.M.F.; Del Aguila, E.M. Analysis of the cocobiota and metabolites of Moniliophthora perniciosa-resistant (Theobroma cacao L.) beans during spontaneous fermentation in Southern Brazil. J. Sci Food Agric. 2018, 98, 4963–4970. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, S.L.; Loiseau, G.; Paredes, J.L.; Barel, M.; Guiraud, J.P. Study on the microflora and biochemistry of cocoa fermentation in the Dominican Republic. Int. J. Food Microbiol. 2007, 114, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Bickel Haase, T.; Schweiggert-Weisz, U.; Ortner, E.; Zorn, H.; Naumann, S. Aroma Properties of Cocoa Fruit Pulp from Different Origins. Molecules 2021, 26, 7618. [Google Scholar] [CrossRef]
- Santander, M.; Rodríguez, J.; Vaillant, F.; y Escobar, S. An overview of the physical and biochemical transformation of cocoa seeds to beans and to chocolate: Flavor formation. Crit. Rev. Food Sci. Nutr. 2019, 60, 1593–1613. [Google Scholar] [CrossRef]
- Papalexandratou, Z.; Dennis, S.; Nielsen, D.S. It's Gettin’ Hot in Here: Breeding Robust Yeast Starter Cultures for Cocoa Fermentation. Trends Microbiol. 2016, 24, 168–170. [Google Scholar] [CrossRef]
- Cempaka, L.; Rahmawati, E.A.; David, W.A. Sensory Profiles of Chocolate Drinks Made from Commercial Fermented Cocoa Powder and Unfermented Cocoa Beans. Curr. Res. Nutr. Food Sci. 2021, 9, 988–999. [Google Scholar] [CrossRef]
- Ooi, T.; Ting, A.; Siow, L. Influence of selected native yeast starter cultures on the antioxidant activities, fermentation index and total soluble solids of Malaysia cocoa beans: A simulation study. LWT Food Sci. Technol. 2020, 122, 1–8. [Google Scholar] [CrossRef]
- Ardhana, M.; Fleet, G. The microbial ecology of cocoa bean fermentations in Indonesia. Int. J. Food Microbiol. 2003, 86, 87–99. [Google Scholar] [CrossRef]
- Afoakwa, E.O.; Quao, J.; Takrama, J.; Budu, A.S.; Saalia, F.K. Chemical composition and physical quality characteristics of Ghanaian cocoa beans as affected by pulp pre-conditioning and fermentation. J. Food Sci. Technol. 2013, 50, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Weckx, S. The cocoa bean fermentation process: From ecosystem analysis to starter culture development. J. Appl. Microbiol. 2016, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Viesser, J.; Pereira, G.; de Carvalho, D.; Vandenberghe, L.; Azevedo, V.; Brenig, B.; de Carvalho, D.; Rogez, H.; Góes-Neto, A.; Soccol, C. Exploring the contribution of fructophilic lactic acid bacteria to cocoa beans fermentation: Isolation, selection and evaluation. Food Res. Int. 2020, 136. [Google Scholar] [CrossRef]
- Kresnowati, M.T.A.P.; Suryani, L.; Affifah, M. Improvement of cocoa beans fermentation by LAB starter addition. J. Med. Bioeng. 2013, 2, 274–278. [Google Scholar] [CrossRef]
- Puerari, C.; Teixeira Magalhães, K.; Freitas Schwan, R. New cocoa pulp-based kefir beverages: Microbiological, chemical composition and sensory analysis. Food Res. Int. 2012, 48, 634–640. [Google Scholar] [CrossRef]
- López, M.; Criollo, J.; Hernández, M.; Lozano, M. Physicochemical and microbiological dynamics of the fermentation of the CCN51 cocoa material in three maturity stages. Rev. Bras. Frutic. 2019, 41, 1–13. [Google Scholar] [CrossRef]
- El Salous, A.; Angulo, A.; Solís, L. Acceleration of cocoa fermentation through the action of bacteria (Acetobacter aceti) and yeast (Saccharomyces cerevisiae). Espirales Rev. Multidiscip. Investig. Científica 2019, 3, 1–20. [Google Scholar] [CrossRef]
- Fonseca, D.; López, D.; Ortíz, S.; Nuñez, C.; Lozano, D. Effect of Addition of a Specific Mixture of Yeast, Lactic and Acetic Bacteria in the Fermentation Process to Improve the Quality and Flavor of Cocoa Beans in Colombia. Pelita Perkeb. Coffee Cocoa Res. J. 2020, 36, 154–172. [Google Scholar] [CrossRef]
Figure 1.
Research flow of a native microbial cocktail as a starter culture in the fine aroma cocoa (Theobroma cacao) bean fermentation.
Figure 1.
Research flow of a native microbial cocktail as a starter culture in the fine aroma cocoa (Theobroma cacao) bean fermentation.
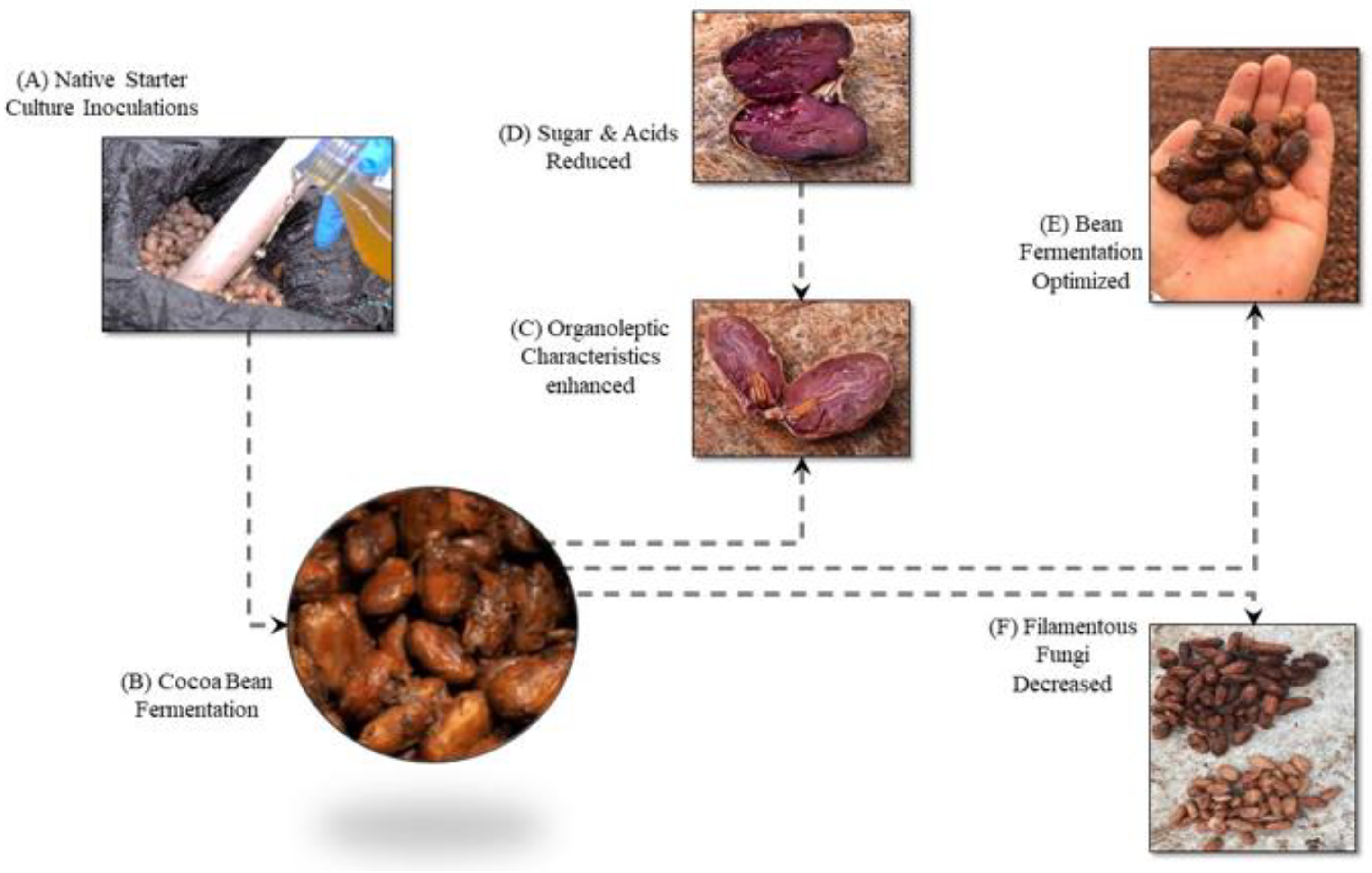
Figure 2.
Changes in temperature ( ) and pH ( ) during fermentation of national variety cocoa beans (a) by spontaneous fermentation (control) and (b) by effect of a microbial cocktail as a starter culture.
Figure 2.
Changes in temperature ( ) and pH ( ) during fermentation of national variety cocoa beans (a) by spontaneous fermentation (control) and (b) by effect of a microbial cocktail as a starter culture.
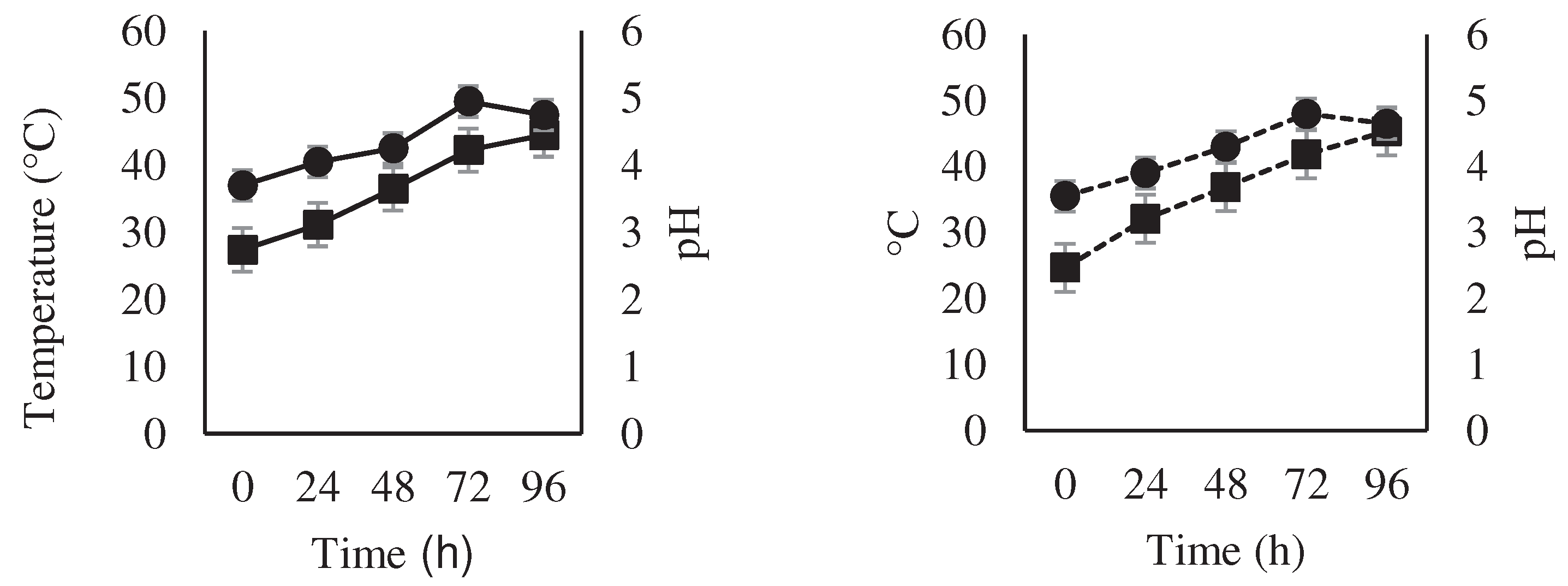
Figure 3.
Changes in the log10
population dynamics of yeasts,  ; BAL,
; BAL, ; BAA,
; BAA,  mesophilic
microorganisms,
mesophilic
microorganisms,  and filamentous
fungi
and filamentous
fungi  during the
fermentation of national variety cocoa beans (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a starter
culture.
during the
fermentation of national variety cocoa beans (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a starter
culture.
; BAL,; BAA, mesophilic
microorganisms, and filamentous
fungi during the
fermentation of national variety cocoa beans (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a starter
culture.
Figure 3.
Changes in the log10
population dynamics of yeasts, ; BAL,; BAA, mesophilic
microorganisms, and filamentous
fungi during the
fermentation of national variety cocoa beans (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a starter
culture.
; BAL,; BAA, mesophilic
microorganisms, and filamentous
fungi during the
fermentation of national variety cocoa beans (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a starter
culture.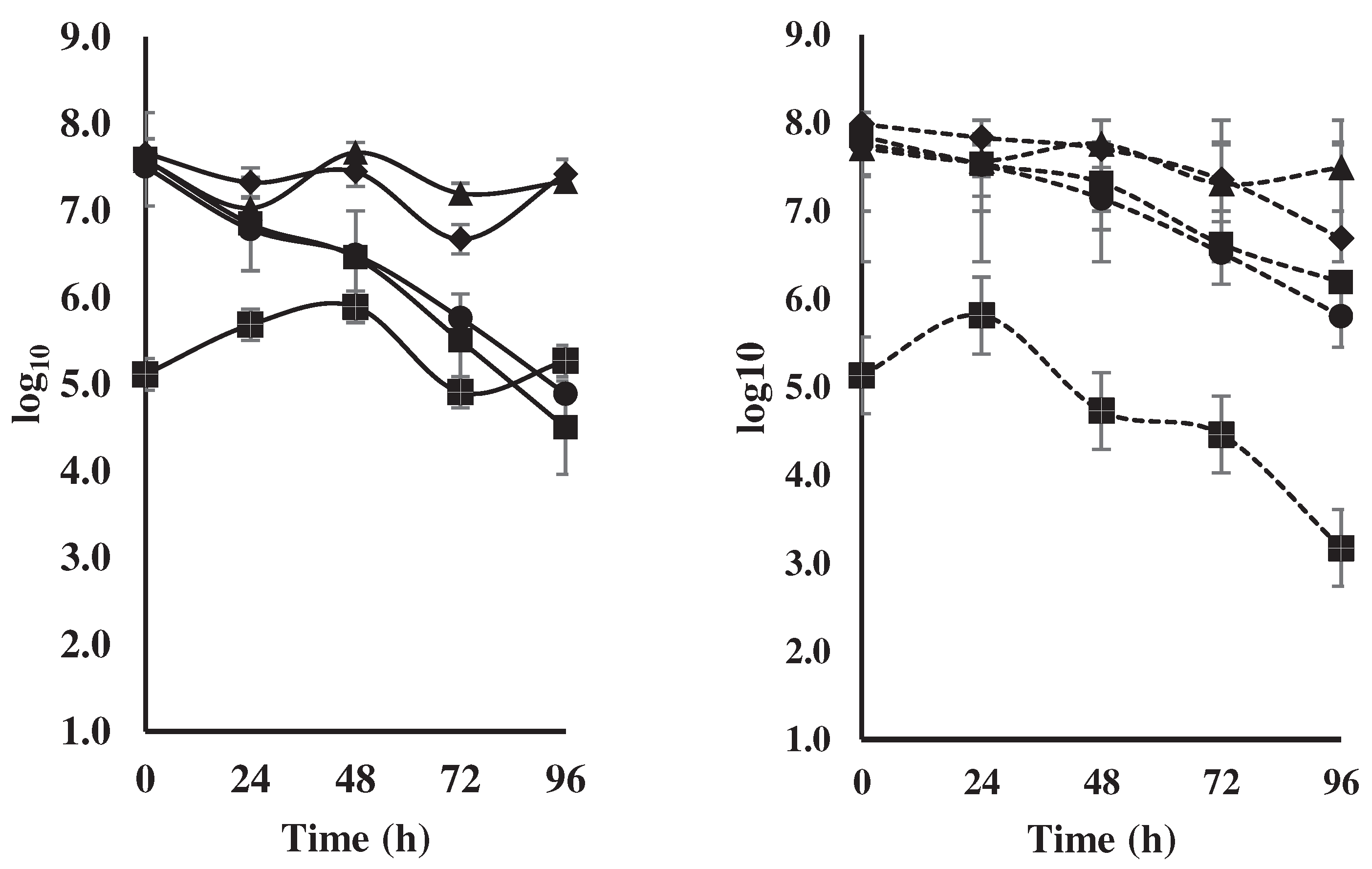
Figure 4.
Changes in the sugar content (sucrose, ;fructose, ; glucose and ethanol in mg/g, during the fermentation of national variety cocoa almonds (a) by spontaneous fermentation (control) and (b) by the effect of a microbial cocktail as a starter culture.
Figure 4.
Changes in the sugar content (sucrose, ;fructose, ; glucose and ethanol in mg/g, during the fermentation of national variety cocoa almonds (a) by spontaneous fermentation (control) and (b) by the effect of a microbial cocktail as a starter culture.
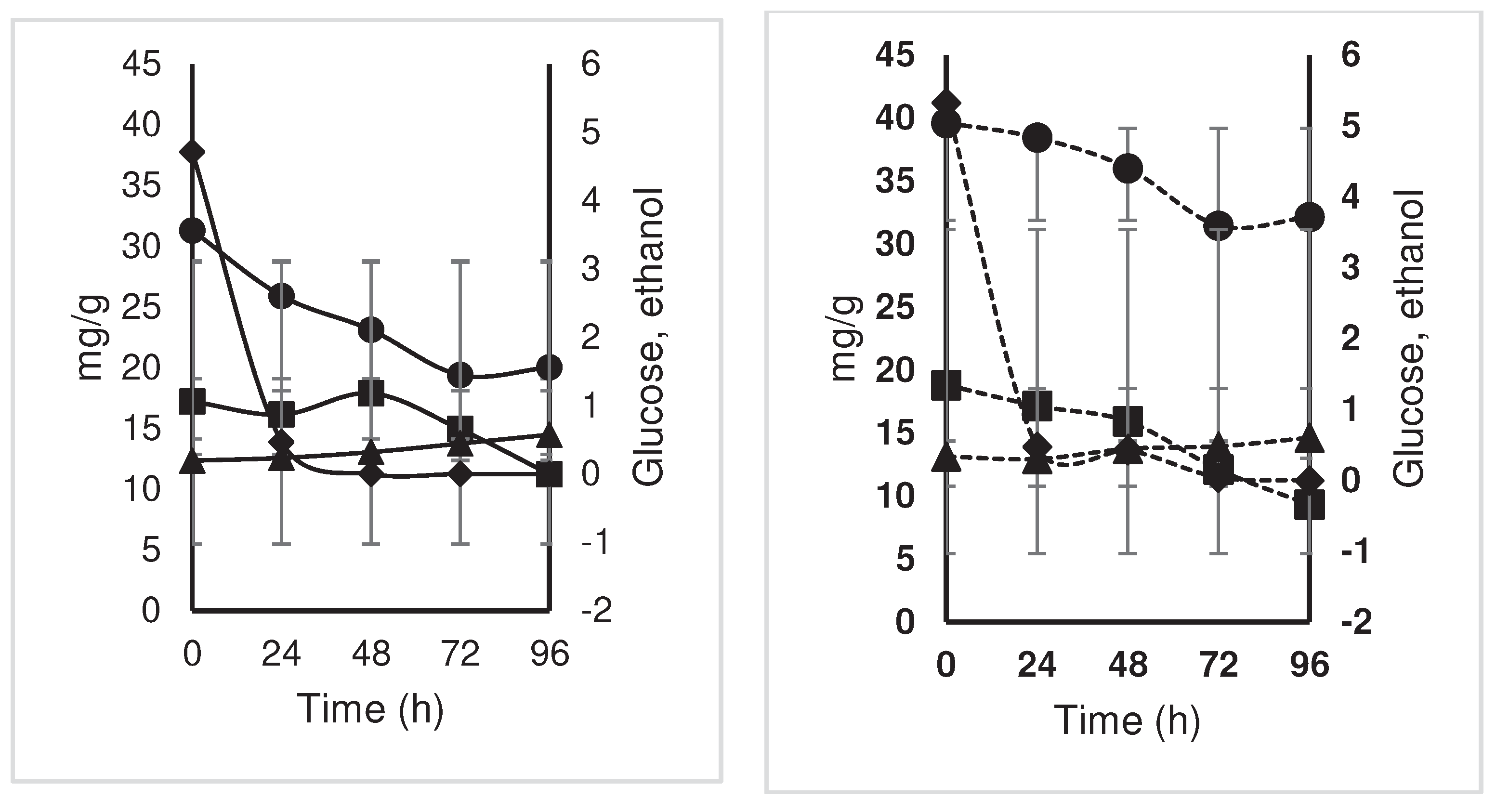
Figure 5.
Changes in acid content (lactic, ; acetic, ; citric,; oxalic,; and malic,) (mg/g) during the
fermentation of national variety cocoa almonds (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a
starter culture.
; acetic, ; citric,; oxalic,; and malic,) (mg/g) during the
fermentation of national variety cocoa almonds (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a
starter culture.
Figure 5.
Changes in acid content (lactic, ; acetic, ; citric,; oxalic,; and malic,) (mg/g) during the
fermentation of national variety cocoa almonds (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a
starter culture.
; acetic, ; citric,; oxalic,; and malic,) (mg/g) during the
fermentation of national variety cocoa almonds (a) by spontaneous
fermentation (control) and (b) by effect of a microbial cocktail as a
starter culture.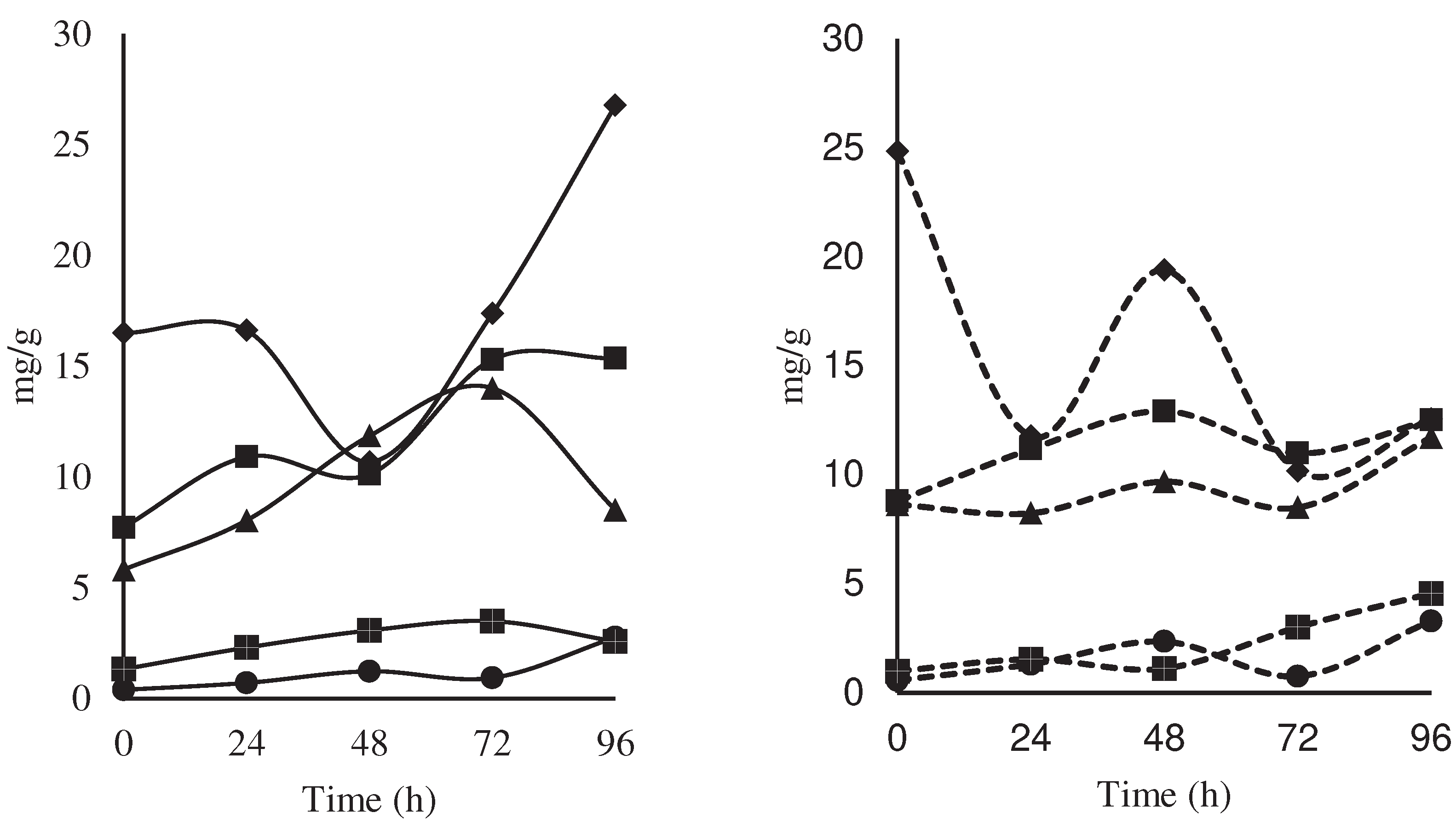
Figure 6.
Changes in the content of polyphenols (mg
AG/g dm) during the fermentation of national variety cocoa beans due to the
effect of a microbial cocktail as a starter culture ( ), compared to the
control by spontaneous fermentation ().
), compared to the
control by spontaneous fermentation ().
Figure 6.
Changes in the content of polyphenols (mg
AG/g dm) during the fermentation of national variety cocoa beans due to the
effect of a microbial cocktail as a starter culture ( ), compared to the
control by spontaneous fermentation ().
), compared to the
control by spontaneous fermentation ().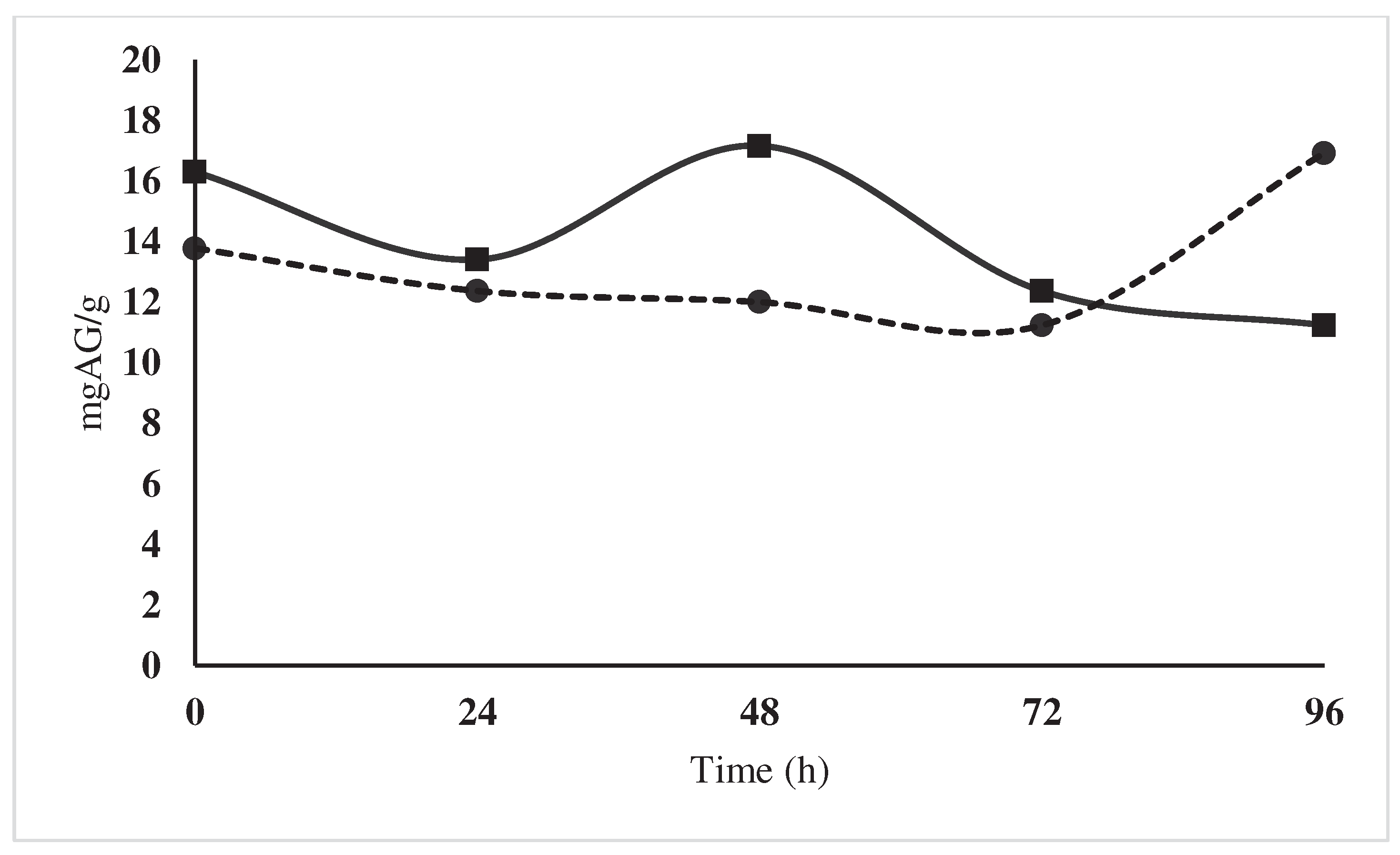
Figure 7.
Fermentation percentage of national variety cocoa beans (a) by spontaneous fermentation and (b) by effect of a microbial cocktail as a starter culture.
Figure 7.
Fermentation percentage of national variety cocoa beans (a) by spontaneous fermentation and (b) by effect of a microbial cocktail as a starter culture.
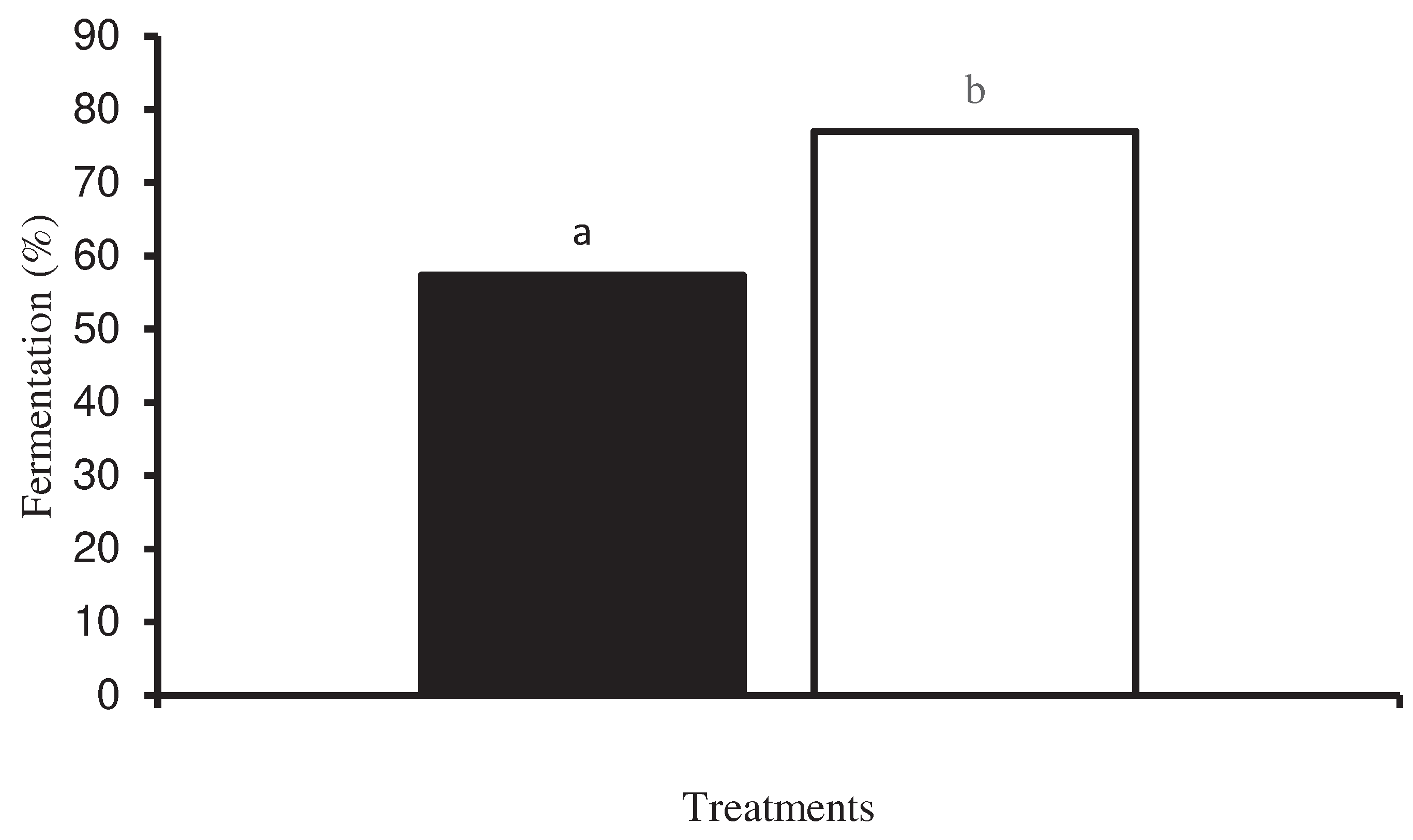

Table 1.
Quality of national variety cocoa beans fermented naturally or with the addition of a microbial starter culture, compared with quality requirements for cocoa beans Arriba Superior Time after processing (CBAS).
Table 1.
Quality of national variety cocoa beans fermented naturally or with the addition of a microbial starter culture, compared with quality requirements for cocoa beans Arriba Superior Time after processing (CBAS).
| Requirements | Unit | Natural Fermented |
Started Culture |
CBAS |
|---|---|---|---|---|
| Maximum humidity | % | 60,64 | 58 | 7 |
| 100 grain weight | g | 106,5 | 109,9 | 100-120 |
| Fermented grains, minimal | % | 57 | 77 | 53 |
| Violet grains, maximum | % | 43 | 20 | 25 |
| Slaty grains, maximum | % | 0 | 3 | 18 |
| Moldy grains, maximum | % | 0 | 0 | 4 |
| Totals (analysis in 100 grams) | % | 100 | 100 | 100 |
| Polyphenol content | mgAG/g | 11,23 | 16,92 | 25-40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



