
Submitted:
13 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
Idiopathic scoliosis (IS) is an unexplained dysplasia of the spine that can occur at any age, with idiopathic scoliosis making up the largest proportion of the total population worldwide, approximately 2-3%. Scoliosis is not just a cosmetic defect, but the development of the spinal deformity can lead to cardiovascular and respiratory lesions, often resulting in serious health problems. Treatment of the condition usually involves major surgery, which is both a physical and financial burden for patients. In order to clarify the aetiology of IS and provide a theoretical basis for new diagnostic and treatment methods IS, it is important to use different animal models for experiments. Zebrafish is an emerging model animal with a short reproductive cycle, minimal breeding expenses, and other beneficial traits including in vitro fertilisation, in vitro development, and embryo transparency. As a result, genetic alteration and observation are simpler than with traditional model animals. This study examines the history of animal models for IS research, focuses on the benefits and drawbacks of zebrafish as an IS model and the advances it provides to IS research, and anticipates zebrafish application prospects in IS research.
Keywords:
adolescent idiopathic scoliosis
; animal model
; zebrafish
; cilia
; cerebrospinal fluid
1. Concept of scoliosis
Scoliosis is a spinal deformity disorder characterised by lateral spine bending, sagittal physiological bending, and vertebral rotation [1]. Scoliosis is classified into three types based on its aetiology: idiopathic scoliosis (IS), which refers to various unexplained spinal dysplasias, congenital scoliosis (CS), which is a lateral curvature of the spine that occurs before birth, and neuromuscular scoliosis (NS), a type of spinal curvature resulting from disease or injury affecting the nerves, muscles or other neuromuscular systems [2]. Of these, the most common is IS, and this article will mainly disscuss scolioses caused by genetic factors, specifically IS.
Adolescent IS (AIS) has a prevalence ranging from 0.47% to 5.2% [3] and is thought to afflict 2-3% of the global population [4]. Pathological symptoms of this spinal condition, which primarily affects children and adolescents, include spinal S-shaped deformity, rib swelling, and chest curvature [5]. AIS not only impacts the appearance of human body flaws, but also the incorrect growth of the spine can create serious problems, such as cardiovascular and respiratory disorders, which often lead to severe impairment and greatly reduce the quality of life of patients. Currently, the primary method of treating AIS is follow-up observation, and when the deformity becomes severe, surgery is used to heal big wounds. This method places a significant financial and physical load on patients and their families. According to statistics, female AIS patients are more than male. According to survey data from various populations, the ratio between the two ranges from 1.5: 1 to 3: 1 [4]. A great deal of research is currently being carried out on scoliosis, especially IS, regarding its pathogenesis, pathophysiological changes, and the apparent link of the disease to genetic and environmental factors. The precise pathogenesis, however, remains unclear, making it unable to offer a useful guide for the disease's early prevention and therapy.
2. Research progress on the pathogenesis of scoliosis
The pathogenesis of scoliosis has been studied for more than a century. Early researchers thought that spinal deformity was primarily caused by biomechanical factors, such as imbalanced spinal muscle activity [6], incorrect posture [6,7], or excessive rib growth [8], and not by the development of the spine itself, but rather by other tissues, organs, and even personal behaviour. This theory does not apply to many AIS patients, but it can be used to explain a small percentage of scoliosis instances. In 1959, Thillard discovered that chickens would develop scoliosis if their pineal glands were removed [9]. Based on this, the theory that melatonin is the cause of scoliosis is proposed. This experiment was replicated with bipedal mice after the 1990s, and identical results were observed [10,11]. However, in other experimental species such hamsters, rabbits, and primates, alterations in melatonin did not promote scoliosis [10,12]. Most crucially, pineal damage in humans did not result in the development of scoliosis [13].
Simultaneously, based on various scoliosis hypotheses embraced by themselves, combined with pathological features and gene functions, the researchers selected some genes, such as connective tissue protein genes [14], bone formation and bone metabolism genes [15], pineal gland hormone genes [13], and puberty and growth genes [16]. These genes were examined and validated in animals such as mice and chickens, but no conclusive pathogenic gene responsible for scoliosis was identified in these experiments.
With the confirmation of the heritability of AIS [17] and the introduction of genome wide association study (GWAS), researchers have carried out many GWAS studies on scoliosis in different ethnic groups, and mapped some loci related to AIS susceptibility in chromosomes 12p, 18q and 19p13 [18,19,20]. Genes such as LBX1, TPH1, SER1 and matrilin-1 (MATN1) were involved [21,22,23,24], and LBX1 was verified in zebrafish and mice.
Afterwards, Hayes et al. created a ptk7a zebrafish model to investigate the gene's role in Wnt signaling and observed IS-like phenotypes in zygotic homozygotes, which prompted them to examine human samples for pathogenic variants in PTK7 [25]. Based on this finding, researchers carried out further studies using zebrafish models and proposed a hypothesis for the pathogenic mechanism of IS, that is, abnormal cilia development disrupts the normal flow of cerebrospinal fluid, thereby trigger IS [26]. A subsequent study substantiated the involvement of PTK7 in a sizable cohort of Chinese IS patients [27].
3. Animal models for IS research
A good animal model can help people perform in-depth study on the mechanisms of human diseases and reach more trustworthy results. The selection of animal models has always been a challenge in IS research. Unlike other diseases, IS is related to the stress of the spine when humans walk upright, hence its animal model must simulate these spinal force characteristics. The choice of animal models for IS has changed from quadrupeds to 'natural' bipeds, to 'artificial' bipeds, and then to fish. In the early days of scoliosis research, researchers used four-legged animals such as calves [28], goats [29], rabbits [30], and rats [31] for explorations. The axial load and dorsal shear stress on the spine of quadruped animals, on the other hand, are significantly different from those of upright humans. In subsequent IS studies, quadruped mice were also directly used as models [32], but this is entirely based on cost and efficiency considerations, and it was difficult to propose convincing hypotheses and theories in these studies, so quadrupeds are not suitable models for IS research.
IS bipedal animal models are now split into two categories: 'natural' bipedal animals and 'artificial' bipedal animals: ' natural ' bipedal animals include chickens and primates. As previously stated, the melatonin theory is based on a chicken experimental model. However, compared to non-avian vertebrates, chickens' spines have a completely different anatomical structure [33]. Only 2 intervertebral spaces (6-7 and 7-8 vertebrae) have clearly visible intervertebral discs, which are movable parts, and the spine only has 8 thoracic vertebrae and no fused lumbar vertebrae [34]. Although chickens are bipedal, their thoracic and lumbar vertebrae are still primarily horizontal, unlike how the human spine is affected by gravity [35]. Additionally, this makes it challenging for other bipedal animals, like primates [12] and rabbits [10], to reproduce the results reported in chickens.
Among the 'natural' bipedal species, primates are phylogenetically closest to humans, and most primates walk upright or semi-upright. Their spines are also anatomically similar to those of humans. These characteristics are very suitable for IS research. However, primates cannot be used extensively in the investigation of IS mechanism due to their expensive cost and lengthy generation cycle.
To study IS using bipeds, scientists have created "artificial" bipedal animal models. Cut off the forefoot of quadruped animals, such as bipedal rats and mice, then arrange food on a high spot to create this form of animal model [11]. The scientific significance of this model is demonstrated by the fact that bipedal rats and mice in several experimental models displayed a higher incidence of scoliosis than the matching quadruped animal model [11]. It should be pointed out that if the relevant tissues and cells of a species have developed a special molecular regulatory mechanism to adapt to an upright spine during long-term evolution (which is very likely), then even if quadrupeds are forcibly reversed to bipedalism, thus changing the load direction of the spine from horizontal to vertical, there will still be a substantial difference in the adaptation mechanism between the models and the target species.
In order to find an animal model more suitable for studying the mechanism of IS, the researchers compared the stress patterns of the human and fish spines. The spine of a teleost fish is not loaded horizontally due to buoyancy [33]. When swimming ahead, it is more likely to experience longitudinal water pressure, which is extremely comparable to the vertical stress features of the human spine. Interestingly, scoliosis is a prevalent ailment in bony fish, unlike in terrestrial mammals [33,36]. Further study has revealed that humans and fish share many common causes of vertebral system defects, including genetic factors, dysregulation of physiological processes ( e.g. calcium regulation ), developmental abnormalities ( e.g. fusion of vertebrae ) and infection by pathogens ( viruses or parasites ), demonstrating the homology of spinal deformities, and suggesting the viability of employing fish as a model for human scoliosis research [33]. Zebrafish, as the most extensively used model fish, is unquestionably the chosen fish IS model.
4. The advantages and disadvantages of zebrafish as a scoliosis research model
Although each lineage of teleosts and mammals has acquired unique traits or lost specific skeletal elements during long-term evolution, the verterate skeleton is a highly conserved organ system, and some distinctive features of the skeleton are related to early fish embryos and adults, making small teleost fish such as zebrafish an excellent animal model for studying human skeletal diseases [37]. There are three clear advantages to using zebrafish for genetic research: 1. The fish is small, easy to feeding, and the breeding cost is low; 2. Through in vitro fertilisation and reproduction, both fertilised eggs and embryos have optical transparency, which is convenient for observation and application of gene editing and other research techniques; 3. Artificial fertilisation is simple to operate, and the number of offspring is large, which is very conducive to the construction and screening of pure lines. Because of these benefits, zebrafish are becoming increasingly popular in medical research. The benefits of zebrafish animal models for IS study go beyond that.
The spine architecture and vertebral structure of zebrafish are quite similar to those of humans from a physiological standpoint. The notochord is the spine's centrosome, and the spinal cord travels through the neural arch [38]. The vertebrae are arranged uniformly from front to back, with bone bows on the dorsal and ventral sides [36]. Furthermore, the precaudal vertebrae of the zebrafish are associated with the rib segments, exhibiting the same natural lordosis as bipedal species such as humans [36]. Although important developmental changes, such as connections to fins and the Weberian apparatus for cranial linkage, must be considered when modelling the establishment of the spine [39]. But the basic structure of the zebrafish and human spines is well conserved [36]. The Weberian apparatus is a chain of tiny bones called Weberian ossicles that connect the swim bladder to the inner ear. This mechanism enhances the fish's ability to hear by amplifying sounds and sensing vibrations in the water [40].
From the perspective of molecular genetics, human and zebrafish coding genes are highly related, with approximately 71% of human protein-coding genes having one or more zebrafish orthologues and 47% having a one-to-one relationship [41]. The basic skeletal components, development, and essential developmental genes of the skeleton are conserved across a wide range of vertebrates, from teleosts to mammals [42,43]. Although developmental networks may differ between species, the core functions of these networks are often conserved, even across phyla. This has made it possible to study gene function between distantly related vertebrates such as teleost fish and mammals, and it is why the zebrafish can be used as a model to study human skeletal diseases [37].
Many genes and pathways function in the zebrafish vertebrae and surrounding tissues, and are conserved across species. For example, the Hedgehog (Hh) signaling pathway is essential for the development of the spine and surrounding tissues, and one of the most studied genes in this pathway, sonic hedgehog (Shh), is conserved in all vertebrates [44]. The Wnt signaling pathway, which is involved in some key developmental processes such as the formation of the body axis, vertebrae, somites, and neural crest, has many components that are conserved from zebrafish to humans [45]. The aforementioned ptk7 mutation is associated with CS and IS via dysregulation of the Wnt signaling pathway [25].
It should be mentioned that throughout its evolutionary history, teleost fish like zebrafish have undergone a fish-specific genome-wide doubling event. Zebrafish possesses more copies of several coding genes than humans and mice do, despite the fact that some of the genes obtained via doubling were quickly lost [46]. For instance, both zebrafish orthologs runx2a and runx2b share similarities with the human RUNX2 gene [47]. Although this feature of zebrafish increases the functional redundancy of the genome, it also increases the difficulty of the operation, as several homologous genes must be knocked out or silenced at the same time when testing gene function. In the case of non-lethal gene knockouts, multiple copies mean that more experimental procedures are required to obtain samples with complete inactivation of the target gene for subsequent research, and may also raise the issue of potential functional differences between different copies. This is clearly a disadvantage for the study of human disease mechanisms using zebrafish as a model. The retention of homologous genes in zebrafish, however, frequently enables the subtle functional examination of such genes [47], without the need to apply a number of genetic approaches to accomplish conditional knockout as in mice. Therefore, the fact that zebrafish have multiple copies of genes also has positive aspects that can be exploited in research.
5. Two uses of zebrafish model in IS research
The purpose or use of zebrafish models in IS research can be divided into two categories: functional verification, in which zebrafish is subjected to gene editing experiments such as CRISPR/Cas9 to verify candidate pathogenic genes obtained from human IS samples; and mechanism exploration, in which zebrafish with scoliosis phenotype is directly used as the research object to explore the mechanism of IS occurrence.
In the first category of studies, candidate genes can be identified through three main approaches: genome-wide or whole-exome association analysis of large groups of disease and control individuals; locating gene loci from families with scoliosis phenotype in some individuals; or identifying genes with abnormal sequences from a single patient sample (Table 1). The choice of approach depends largely on the availability of suitable sample groups. In fact, the difficulty of obtaining familial data and the ease of obtaining random groups determined the high proportion of GWAS research. Due to the higher prevalence of idiopathic scoliosis in females and the fact that women are prone to more severe deformities, most of these studies have intentionally selected severely curved female individuals as the disease group to locate candidate genes. GWAS are commonly used, with female individuals with severe spinal curvature often selected as the disease group to identify candidate genes. Studies based on a single scoliosis case tend to introduce a large number of false positive candidate genes, so only those that have been validated by animal models are likely to be accepted and published. Therefore, this type of research has the lowest proportion. Validation in model organisms is essential for human-derived genes to be of value for future research. For instance, the candidate genes GPR126 [50], POC5 [51], and LBX1 [52], identified through multiple population association analyses, have been validated for their association with IS through zebrafish knockout experiments. Other genes, such as BNC2 [53], PAX1 [54], MAPK7 [55], TTLL11 [56] and PTK7 [57], have also been confirmed through gene knockout experiments in model animals, mainly using zebrafish [25,51,52,53,54,56,58] (Table 2).
However, identifying and verifying candidate genes is only the first step towards understanding the pathogenesis of IS. Firstly, mutations in these genes affect only a small proportion of individuals with IS. For instance, even genes such as GPR126, which have been identified in several ethnic groups by GWAS, can only explain a limited number of cases. Secondly, proposing one or more hypotheses that can reasonably connect these genes, which may not be related to existing human knowledge, is a challenge for researchers' creativity. Finally, the GWAS study of IS may have reached its bottleneck period, as researchers have compared different races, increased the sample size to almost 100,000 individuals, and still failed to identify further explanatory clues. As a result, the proportion of such research in exploring the mechanism of IS has gradually declined over the past five years.
The second type of research involves obtaining zebrafish with a scoliosis phenotype, which can be achieved mainly in two ways: serendipitous discovery and screening for induced mutations. It has occasionally been discovered in investigations with different goals that knocking down particular genes in zebrafish can lead to IS. For instance, scientists studying the human developmental disorder cilia disease discovered that zebrafish with the cc2d2a gene mutation displayed scoliosis phenotype, and scientists studying primary ciliary motor dysfunction discovered that zebrafish with the c21orf59, ccdc40, ccdc151, and dyx1c1 mutations also displayed juvenile or late-onset scoliosis, similar to AIS [85,86,87,88] (Table 2).

Table 2.
Model biological verification for diseases with spinal deformity.
| Related diseases | Gene | Animal model validation | Year | Reference | |
|---|---|---|---|---|---|
| Reissner Fiber | AIS | scospondin | zebrafish | 2018, 2020 | [89,90,91] |
| dnah10 | zebrafish | 2022 | [92] | ||
| spine curvature | camel | zebrafish | 2021 | [93] | |
| vangl2 | zebrafish | 2022 | [94] | ||
| IS | scospondin | zebrafish | 2020 | [95] | |
| katnb1 | zebrafish | 2022 | [96] | ||
| ccdc57 | zebrafish | 2023 | [97] | ||
| cilia | AIS | rpgrip1l | zebrafish | 2019 | [98] |
| cfap298 | zebrafish | 2021 | [99] | ||
| dnah10 | zebrafish | 2022 | [100] | ||
| ccdc57 | zebrafish | 2023 | [101] | ||
| IS | ttll11 | zebrafish | 2021 | [102] | |
| katnb1 | zebrafish | 2022 | [103] | ||
| ccdc57 | zebrafish | 2023 | [104] | ||
| CS, IS | ptk7 | zebrafish | 2014, 2016 | [26,105] | |
| IS, CSF stasis | kif6 | mouse, zebrafish | 2014, 2018 | [106,107] | |
| PCD | ccdc40 | zebrafish | 2011 | [108] | |
| dyx1c1 | mouse, zebrafish | 2013 | [109] | ||
| ccdc151 | mouse, zebrafish | 2014 | [110] | ||
| zmynd10 | medaka, zebrafish | 2017, 2018 | [111,112] | ||
| ciliopathies | c21orf59 | zebrafish | 2013 | [113] | |
| cep290 | zebrafish | 2019, 2022 | [114,115] | ||
| ciliopathy syndromes | bbs-5 | C. elegans, mouse, zebrafish | 2022 | [116] | |
| nphp-4 | |||||
| others | AIS | gpr126 | zebrafish | 2013 | [117] |
| mouse | 2019 | [118] | |||
| bnc2 | zebrafish | 2015 | [119] | ||
| mapk7 | zebrafish | 2018 | [120] | ||
| lbx1 | mouse, zebrafish | 2016, 2022, 2023 | [121,122,123] | ||
| uncx | zebrafish | 2023 | [124] | ||
| CS | Foxo4 | rat | 2019 | [125] | |
| IS | poc5 | zebrafish | 2015 | [126] | |
| kif7 | zebrafish | 2021 | [127] | ||
| ppp2r3b | zebrafish | 2023 | [128] | ||
| PCD | zmynd10 | zebrafish | 2018 | [129] | |
| kif3b | |||||
| uts2ra | |||||
| thoracic aortic aneurysm | col1a2 | zebrafish | 2022 | [130] | |
| col5a1 | |||||
| col5a2 | |||||
| fbn1 | |||||
| spine curvature | pkd2l1 | zebrafish | 2018 | [131] | |
| urp1 | zebrafish | 2022, 2023 | [132,133] | ||
| urp2 |
AIS: adolescent idiopathic scoliosis, IS: Idiopathic scoliosis, CS: congenital scoliosis, PCD: primary ciliary dyskinesia.
Furthermore, a random mutant zebrafish population can also be created to screen individuals with a heritable scoliosis trait. In 2020, Sun et al. obtained a mutant zebrafish line with congenital scoliosis-like vertebral malformations through ENU mutagenesis screening. [134]. They also discovered and validated the related gene dstyk in the ENU-induced family, and its protein product DSTYK is found in the late endosomal / lysosomal compartment and contributes in lysosome production in mammalian cells. This gene's knockout enhances mTORC1-dependent TFEB nuclear translocation, which inhibits notochord vacuole and lysosomal production, resulting in vacuole deficits in notochord cells during notochord development and vertebral malformations in dstyk mutant zebrafish. In turn, inhibiting mTORC1 activity can repair the dstyk mutant's abnormalities in notochord vacuole biosynthesis and scoliosis[134]. In the same year, Bagwell et al. conducted a study demonstrating that mutations in dstyk cause notochord vacuole fragmentation and a severe congenital scoliosis-like phenotype in zebrafish [135].
The second type of study can only provide possible evidence for the molecular mechanism of spinal dysplasia and does not exhibit a direct correlation between IS and actual cases. Therefore, combining the candidate genes of AIS patients obtained from the first type of research with the results of the second type of research can provide potential hypotheses. Through innovative approaches and using zebrafish as a model, these hypotheses can be fully tested and confirmed, which may be the most practical approach to discover the underlying mechanism of IS. In the following sections, we will describe in detail the research involving the PTK7 gene, which hypothesizes that cilia damage and abnormal cerebrospinal fluid flow contribute to IS, as well as other related studies using this combination of techniques.
6. The relationship between cilia and IS was studied by zebrafish model
Researchers generated maternal-zygotic and zygotic mutant zebrafish strains of protein tyrosine kinase-7 (ptk7). The former had congenital scoliosis, while the latter had an idiopathic scoliosis phenotype that was highly comparable to that of humans [25,57] . Su et al. also confirmed the involvement of PTK7 in IS [27]. Cilia function is intimately connected to the classical β-catenin and planar cell polarity (PCP) Wnt signalling pathways, which Ptk7 is a key regulator of. Organelles called cilia can be seen sticking out from the surface of eukaryotic cells. They can be classified as primary (non-moving) cilia, and moving cilia based on how they move. The movement of single-cell organisms or the fluid of metazoan animals, such as cerebrospinal fluid (CSF), can be facilitated by the action of moving cilia [136,137,138]. It is worth mentioning that scoliosis phenotypes have been seen in several investigations of cilia-related genes [139,140,141,142], and scoliosis patients have CSF flow abnormalities. CSF is a watery fluid found in the brain and spinal cord that is necessary for central nervous system homeostasis. Based on the preceding evidence, the researchers hypothesised that the lack of zebrafish ptk7 produced cilia malfunction, which resulted in CSF flow abnormalities and scoliosis. To test this idea, researchers used the transcription factor Foxj1 [143], which is the major enhancer of human motor cilia development, to create Tg (foxj1a:: ptk7) zebrafish. Ptk7 malfunction in the motor cilia lineage resulted in brain motor cilia development problems and aberrant CSF flow, according to the findings. And foxj1 overexpression can reverse scoliosis caused by ptk7 deletion while also allowing normal development of motor cilia and CSF flow [144]. The researchers also created zebrafish strains with conditional knockouts of other genes essential for the development of motor cilia, like c21or f59, and mutant will also exhibit the same phenotypes as ptk7 mutant zebrafish in order to further demonstrate that the abnormal development of motor cilia can cause scoliosis [144].
Later research has further supported the impact of cilia abnormalities on zebrafish axial development (Figure 1). For instance, the cilia's structure is influenced by the tubulin tyrosine ligase-like 11 (TTLL11), whose loss also causes scoliosis in zebrafish [145]. The deletion of CCDC57 protein, which has an effect on the assembly of cilia, also causes the zebrafish mutants displaying ventral curvature at the embryonic stage [146,147]. Li et al established a ccdc57 knockout zebrafish strain with frameshift mutations. The team's research results showed that the dysfunction of ccdc57 can disrupt the coordinated beating of multiple cilia bundles in brain ventricle, change the cilia beat pattern, and result in scoliosis in zebrafish [147]. Additionally, Wang et al performed exome sequencing analysis on 195 AIS patients in southern China and discovered the genes dnaaf1 and zmynd10 that are related to ciliary dynein assembly and that also exhibit scoliosis phenotypes in zebrafish when any one of them is knocked out [148].
How does CSF influence the development of the zebrafish body axis? The discovery of the cilia-CSF-scoliosis relationship is a significant advancement in understanding the mechanism of scoliosis formation. Urotensin-related peptide (Urp), which is present in CSF, has been linked to scoliosis in zebrafish, according to study from Zhang et al. Zebrafish adults who have Urp receptor mutations can develop severe scoliosis [149]. To investigate whether the urotensin signaling pathway is also involved in the regulation of spinal curvature in human scoliosis patients, bilateral paravertebral muscle tissue was collected from scoliosis patients during surgery. The researchers compared the expression of urotensin signaling pathway genes in paravertebral muscle tissue obtained from the convex and concave sides of the spinal curvature sites. It was found that the expression of UTS2R was significantly higher in convex muscle tissue than in concave muscle tissue [146].
Reissner fibre (RF) has received attention from other researchers (Figure 1). Most notochords contain RF, although its function is unknown. It is a protein filament released at the mesencephalic-diencephalic (M-D) junction by specialised ependymal cells. It is positioned in the centre of the notochord canal filled with CSF. It continues to the notochord canal's caudal end and decomposed here, the byproduct enters the CSF and blood [150]. Recent research has revealed that RF in zebrafish is in a very dynamic balance of secretion and breakdown [151]. The glycoprotein SCO-spondin, which is highly conserved in notochords, is the major composed of RF [152]. It is a big multi-domain extracellular matrix molecule that is primarily involved in cell adhesion and axon guidance [150]. Cantaut-Belarif et al. (2018) created a scospondin gene knockout line in zebrafish and discovered that RF was required for zebrafish body axis expansion. The cilia and CSF flow were normal in RF-deficient zebrafish, but the juveniles displayed an evident scoliosis phenotype [153]. Later, the researchers created a zebrafish strain producing scospondin with a missense mutation. In early embryos, this SCO-spondin protein can ensure the assembly and extension of RF, but as the embryo develops, RF increasingly disintegrates, and zebrafish embryos also exhibit the phenotypic of scoliosis [154]. The previously mentioned deletion of the ccdc57 not only results in abnormal cilia movement, but also causes significant thinning, discontinuity or deletion of RF in adult zebrafish mutants of ccdc57. The results of these studies once again demonstrated the importance of RF integrity for the formation of the zebrafish spine. Additionally, scientists have discovered that CSF-contacting neurons (CSF-cNs) in the spinal cord need RF to perceive the body axis' curvature [155].
7. Conclusions and perspectives
Zebrafish is a well-established model organism with several benefits. It is quite practical to observe and modify its embryos genetically. Imaging and bone staining both have established technologies. More importantly, its similarity to human spine pressure during exercise and conservation with human bone formation are of unique value for the research of IS. The relevance of the zebrafish model is exemplified by the fact that many potential pathogenic genes identified in human patients through GWAS analysis have been confirmed in zebrafish.
As a result, zebrafish have taken the position of other animals as the primary animal model for research on the pathophysiology of scoliosis. Recently, scientists have discovered a strong correlation between cilia-driven CSF flow and scoliosis using zebrafish as a model. They later discovered that the presence of urotensin in CSF fluid might influence how the spine bends dorsally and ventrally, and that RF breakdown can result in scoliosis. We now have a much better knowledge of how scoliosis develops because to these recent discoveries.
However, there are still numerous issues and connections that need to be researched in the study of the scoliosis mechanism. A large number of genes detected in human IS patients cannot be explained by CSF flow and RF abnormalities (Table 2). The SCO, which is a synthesizing organ for the RF, atrophies shortly after human birth, resulting in the cessation of SCO-spondin synthesis. Even in the zebrafish cerebrospinal fluid-RF model, there are still many aspects that require further investigation [156]. For instance, are there additional substances in cerebral fluid, besides urotensin, that influence the geometry of the body axis? What substances does the body utilise to control how the body axis is shaped? What part does RF play in the formation of the spine? How do RF and CSF-cNs work together to detect axis curvature? To fully explore and study all of these issues, researchers must fully utilise zebrafish and other animal models.
Author Contributions
Conceptualization, K.R., M.S. and X.-Q.X.; writing—original draft preparation, K.R.; writing—review and editing, M.S., X.-Q.X.; supervision, Y.C., Z.W and Y.W.; funding acquisition, M.S. and X.-Q.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant No. 31801055), the National Key R&D Program of China (Grant No. 2021YFD1200804).
Acknowledgments
We would like to thank the Wuhan Branch, Supercomputing Center, Chinese Academy of Sciences, for the support of data computing in this research work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trobisch, P.; Suess, O.; Schwab, F. Idiopathic Scoliosis. Deutsches Ärzteblatt international 2010. [Google Scholar] [CrossRef] [PubMed]
- Vialle, R.; Thévenin-Lemoine, C.; Mary, P. Neuromuscular Scoliosis. Orthopaedics & Traumatology: Surgery & Research 2013, 99, S124–S139. [Google Scholar] [CrossRef]
- Konieczny, M.R.; Senyurt, H.; Krauspe, R. Epidemiology of Adolescent Idiopathic Scoliosis. J Child Orthop 2013, 7, 3–9. [Google Scholar] [CrossRef]
- Fadzan, M.; Bettany-Saltikov, J. Etiological Theories of Adolescent Idiopathic Scoliosis: Past and Present. TOORTHJ 2017, 11, 1466–1489. [Google Scholar] [CrossRef]
- Latalski, M.; Danielewicz-Bromberek, A.; Fatyga, M.; Latalska, M.; Kröber, M.; Zwolak, P. Current Insights into the Aetiology of Adolescent Idiopathic Scoliosis. Arch Orthop Trauma Surg 2017, 137, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Huebert, H.T. Scoliosis. A Brief History. Manit Med Rev 1967, 47, 452–456. [Google Scholar] [PubMed]
- Yan, B.; Lu, X.; Qiu, Q.; Nie, G.; Huang, Y. Association Between Incorrect Posture and Adolescent Idiopathic Scoliosis Among Chinese Adolescents: Findings From a Large-Scale Population-Based Study. Frontiers in Pediatrics 2020, 8. [Google Scholar] [CrossRef]
- Gréalou, L.; Aubin, C.-É.; Labelle, H. Rib Cage Surgery for the Treatment of Scoliosis: A Biomechanical Study of Correction Mechanisms. J. Orthop. Res. 2002, 20, 1121–1128. [Google Scholar] [CrossRef]
- Thillard, M.J. Vertebral column deformities following epiphysectomy in the chick. C R Hebd Seances Acad Sci 1959, 248, 1238–1240. [Google Scholar]
- O’Kelly, C.; Wang, X.; Raso, J.; Moreau, M.; Mahood, J.; Zhao, J.; Bagnall, K. The Production of Scoliosis after Pinealectomy in Young Chickens, Rats, and Hamsters. Spine (Phila Pa 1976) 1999, 24, 35–43. [Google Scholar] [CrossRef]
- Machida, M.; Dubousset, J.; Yamada, T.; Kimura, J.; Saito, M.; Shiraishi, T.; Yamagishi, M. Experimental Scoliosis in Melatonin-Deficient C57BL/6J Mice without Pinealectomy. J Pineal Res 2006, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.M.C.; Wang, T.; Poon, A.M.S.; Carl, A.; Tranmer, B.; Hu, Y.; Luk, K.D.K.; Leong, J.C.Y. The Effect of Pinealectomy on Scoliosis Development in Young Nonhuman Primates. Spine (Phila Pa 1976) 2005, 30, 2009–2013. [Google Scholar] [CrossRef]
- Day, G.; McPhee, I.; Tuffley, J.; Tomlinson, F.; Chaseling, R.; Kellie, S.; Torode, I.; Sherwood, M.; Cutbush, K.; Geddes, A.; et al. Idiopathic Scoliosis and Pineal Lesions in Australian Children. J Orthop Surg (Hong Kong) 2007, 15, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Gorman, K.F.; Julien, C.; Moreau, A. The Genetic Epidemiology of Idiopathic Scoliosis. Eur Spine J 2012, 21, 1905–1919. [Google Scholar] [CrossRef]
- Nowak, R.; Szota, J.; Mazurek, U. Vitamin D Receptor Gene (VDR) Transcripts in Bone, Cartilage, Muscles and Blood and Microarray Analysis of Vitamin D Responsive Genes Expression in Paravertebral Muscles of Juvenile and Adolescent Idiopathic Scoliosis Patients. BMC Musculoskelet Disord 2012, 13, 259. [Google Scholar] [CrossRef]
- Zhao, D.; Qiu, G.; Wang, Y.; Zhang, J.; Shen, J.; Wu, Z. Association between Adolescent Idiopathic Scoliosis with Double Curve and Polymorphisms of Calmodulin1 Gene/Estrogen Receptor-α Gene. Orthop Surg 2009, 1, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Burwell, R.G.; Clark, E.M.; Dangerfield, P.H.; Moulton, A. Adolescent Idiopathic Scoliosis (AIS): A Multifactorial Cascade Concept for Pathogenesis and Embryonic Origin. Scoliosis Spinal Disord 2016, 11, 8. [Google Scholar] [CrossRef]
- Raggio, C.L.; Giampietro, P.F.; Dobrin, S.; Zhao, C.; Dorshorst, D.; Ghebranious, N.; Weber, J.L.; Blank, R.D. A Novel Locus for Adolescent Idiopathic Scoliosis on Chromosome 12p. J Orthop Res 2009, 27, 1366–1372. [Google Scholar] [CrossRef]
- Alden, K.J.; Marosy, B.; Nzegwu, N.; Justice, C.M.; Wilson, A.F.; Miller, N.H. Idiopathic Scoliosis: Identification of Candidate Regions on Chromosome 19p13. Spine (Phila Pa 1976) 2006, 31, 1815–1819. [Google Scholar] [CrossRef]
- Gurnett, C.A.; Alaee, F.; Bowcock, A.; Kruse, L.; Lenke, L.G.; Bridwell, K.H.; Kuklo, T.; Luhmann, S.J.; Dobbs, M.B. Genetic Linkage Localizes an Adolescent Idiopathic Scoliosis and Pectus Excavatum Gene to Chromosome 18 q. Spine (Phila Pa 1976) 2009, 34, E94–100. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Z.; Zhuang, Q.; Fei, Q.; Zhang, J.; Liu, Y.; Wang, Y.; Ding, Y.; Qiu, G. Association Study of Tryptophan Hydroxylase 1 and Arylalkylamine N-Acetyltransferase Polymorphisms with Adolescent Idiopathic Scoliosis in Han Chinese. Spine (Phila Pa 1976) 2008, 33, 2199–2203. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Qiu, Y.; Zhang, L.; Sun, Q.; Qiu, X.; He, Y. Association of Estrogen Receptor Gene Polymorphisms with Susceptibility to Adolescent Idiopathic Scoliosis. Spine (Phila Pa 1976) 2006, 31, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.S.; Tang, N.L.S.; Yeung, H.Y.; Lee, K.-M.; Hung, V.W.Y.; Ng, B.K.W.; Ma, S.L.; Kwok, R.H.K.; Qin, L.; Qiu, Y.; et al. Melatonin Receptor 1B (MTNR1B) Gene Polymorphism Is Associated With the Occurrence of Adolescent Idiopathic Scoliosis. Spine 2007, 32, 1748–1753. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Kou, I.; Takahashi, A.; Johnson, T.A.; Kono, K.; Kawakami, N.; Uno, K.; Ito, M.; Minami, S.; Yanagida, H.; et al. A Genome-Wide Association Study Identifies Common Variants near LBX1 Associated with Adolescent Idiopathic Scoliosis. Nat Genet 2011, 43, 1237–1240. [Google Scholar] [CrossRef]
- Hayes, M.; Gao, X.; Yu, L.X.; Paria, N.; Henkelman, R.M.; Wise, C.A.; Ciruna, B. Ptk7 Mutant Zebrafish Models of Congenital and Idiopathic Scoliosis Implicate Dysregulated Wnt Signalling in Disease. Nat Commun 2014, 5, 4777. [Google Scholar] [CrossRef]
- Hayes, M.; Gao, X.; Yu, L.X.; Paria, N.; Henkelman, R.M.; Wise, C.A.; Ciruna, B. Ptk7 Mutant Zebrafish Models of Congenital and Idiopathic Scoliosis Implicate Dysregulated Wnt Signalling in Disease. Nat Commun 2014, 5, 4777. [Google Scholar] [CrossRef]
- Su, Z.; Yang, Y.; Wang, S.; Zhao, S.; Zhao, H.; Li, X.; Niu, Y.; Qiu, G.; Wu, Z.; et al.; Deciphering Disorders Involving Scoliosis and COmorbidities (DISCO) Study Group The Mutational Landscape of PTK7 in Congenital Scoliosis and Adolescent Idiopathic Scoliosis. Genes 2021, 12, 1791. [Google Scholar] [CrossRef]
- Roshono, Y.; Kaneda, K.; Yamamoto, I. A Biomechanical Analysis of Zielke, Kaneda, and Cotrel-Dubousset Instrumentations in Thoracolumbar Scoliosis A Calf Spine Model. Spine 1991, 16, 1305–1311. [Google Scholar] [CrossRef]
- Braun, J.T.; Akyuz, E. Prediction of Curve Progression in a Goat Scoliosis Model. J Spinal Disord Tech 2005, 18, 272–276. [Google Scholar]
- Langenskiöld, A.; Michelsson, J.-E. EXPERIMENTAL PROGRESSIVE SCOLIOSIS IN THE RABBIT. The Journal of Bone and Joint Surgery. British volume 1961, 43-B, 116–120. [Google Scholar] [CrossRef]
- Kasuga, K. Experimental scoliosis in the rat spine induced by binding the spinous processes. Nihon Seikeigeka Gakkai Zasshi 1994, 68, 798–807. [Google Scholar]
- Bobyn, J.D.; Little, D.G.; Gray, R.; Schindeler, A. Animal Models of Scoliosis. J. Orthop. Res. 2015, 33, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Gorman, K.F.; Breden, F. Teleosts as Models for Human Vertebral Stability and Deformity. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2007, 145, 28–38. [Google Scholar] [CrossRef]
- Yoshihara, H.; Kawakami, N.; Matsuyama, Y.; Inoh, H.; Imagama, S.; Ishiguro, N. A Histomorphologic Study of Scoliosis in Pinealectomized Chickens. Spine (Phila Pa 1976) 2005, 30, 2244–2251. [Google Scholar] [CrossRef]
- Castelein, R.M.; van Dieën, J.H.; Smit, T.H. The Role of Dorsal Shear Forces in the Pathogenesis of Adolescent Idiopathic Scoliosis--a Hypothesis. Med Hypotheses 2005, 65, 501–508. [Google Scholar] [CrossRef]
- Boswell, C.W.; Ciruna, B. Understanding Idiopathic Scoliosis: A New Zebrafish School of Thought. Trends in Genetics 2017, 33, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Witten, P.E.; Harris, M.P.; Huysseune, A.; Winkler, C. Small Teleost Fish Provide New Insights into Human Skeletal Diseases. In Methods in Cell Biology; Elsevier, 2017; Volume 138, pp. 321–346. ISBN 978-0-12-803473-6. [Google Scholar]
- Fleming, A.; Keynes, R.; Tannahill, D. A Central Role for the Notochord in Vertebral Patterning. Development 2004, 131, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Bird, N.C.; Hernandez, L.P. Building an Evolutionary Innovation: Differential Growth in the Modified Vertebral Elements of the Zebrafish Weberian Apparatus. Zoology (Jena) 2009, 112, 97–112. [Google Scholar] [CrossRef]
- Grande, T.; Young, B. The Ontogeny and Homology of the Weberian Apparatus in the Zebrafish Danio Rerio (Ostariophysi: Cypriniformes). Zoological Journal of the Linnean Society 2004, 140, 241–254. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Harris, M.P.; Henke, K.; Hawkins, M.B.; Witten, P.E. Fish Is Fish: The Use of Experimental Model Species to Reveal Causes of Skeletal Diversity in Evolution and Disease. J. Appl. Ichthyol. 2014, 30, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Apschner, A.; Schulte-Merker, S.; Witten, P.E. Not All Bones Are Created Equal – Using Zebrafish and Other Teleost Species in Osteogenesis Research. In Methods in Cell Biology; Elsevier, 2011; Volume 105, pp. 239–255. ISBN 978-0-12-381320-6. [Google Scholar]
- Andoniadou, C.L.; Martinez-Barbera, J.P. Developmental Mechanisms Directing Early Anterior Forebrain Specification in Vertebrates. Cell. Mol. Life Sci. 2013, 70, 3739–3752. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/β-Catenin Signaling in Development and Disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, S.M.K.; Neuhauss, S.C.F. Whole-Genome Duplication in Teleost Fishes and Its Evolutionary Consequences. Mol Genet Genomics 2014, 289, 1045–1060. [Google Scholar] [CrossRef]
- Busse, B.; Galloway, J.L.; Gray, R.S.; Harris, M.P.; Kwon, R.Y. Zebrafish: An Emerging Model for Orthopedic Research. J. Orthop. Res. 2020, 38, 925–936. [Google Scholar] [CrossRef]
- Bergen, D.J.M.; Kague, E.; Hammond, C.L. Zebrafish as an Emerging Model for Osteoporosis: A Primary Testing Platform for Screening New Osteo-Active Compounds. Front. Endocrinol. 2019, 10, 6. [Google Scholar] [CrossRef]
- Kwon, R.Y.; Watson, C.J.; Karasik, D. Using Zebrafish to Study Skeletal Genomics. Bone 2019, 126, 37–50. [Google Scholar] [CrossRef]
- Kou, I.; Takahashi, Y.; Johnson, T.A.; Takahashi, A.; Guo, L.; Dai, J.; Qiu, X.; Sharma, S.; Takimoto, A.; Ogura, Y.; et al. Genetic Variants in GPR126 Are Associated with Adolescent Idiopathic Scoliosis. Nat Genet 2013, 45, 676–679. [Google Scholar] [CrossRef]
- Patten, S.A.; Margaritte-Jeannin, P.; Bernard, J.-C.; Alix, E.; Labalme, A.; Besson, A.; Girard, S.L.; Fendri, K.; Fraisse, N.; Biot, B.; et al. Functional Variants of POC5 Identified in Patients with Idiopathic Scoliosis. J. Clin. Invest. 2015, 125, 1124–1128. [Google Scholar] [CrossRef]
- Guo, L.; Yamashita, H.; Kou, I.; Takimoto, A.; Meguro-Horike, M.; Horike, S.; Sakuma, T.; Miura, S.; Adachi, T.; Yamamoto, T.; et al. Functional Investigation of a Non-Coding Variant Associated with Adolescent Idiopathic Scoliosis in Zebrafish: Elevated Expression of the Ladybird Homeobox Gene Causes Body Axis Deformation. PLoS Genet 2016, 12, e1005802. [Google Scholar] [CrossRef]
- Ogura, Y.; Kou, I.; Miura, S.; Takahashi, A.; Xu, L.; Takeda, K.; Takahashi, Y.; Kono, K.; Kawakami, N.; Uno, K.; et al. A Functional SNP in BNC2 Is Associated with Adolescent Idiopathic Scoliosis. The American Journal of Human Genetics 2015, 97, 337–342. [Google Scholar] [CrossRef]
- Sharma, S.; Londono, D.; Eckalbar, W.L.; Gao, X.; Zhang, D.; Mauldin, K.; Kou, I.; Takahashi, A.; et al.; TSRHC Scoliosis Clinical Group; Japan Scoliosis Clinical Research Group A PAX1 Enhancer Locus Is Associated with Susceptibility to Idiopathic Scoliosis in Females. Nat Commun 2015, 6, 6452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Chen, C.; Xu, C.; Zhou, H.; Gao, B.; Su, D.; Liao, Z.; Li, Y.; Yang, S.; Su, P. Mutant MAPK7-Induced Idiopathic Scoliosis Is Linked to Impaired Osteogenesis. Cell Physiol Biochem 2018, 48, 880–890. [Google Scholar] [CrossRef]
- Mathieu, H.; Patten, S.A.; Aragon-Martin, J.A.; Ocaka, L.; Simpson, M.; Child, A.; Moldovan, F. Genetic Variant of TTLL11 Gene and Subsequent Ciliary Defects Are Associated with Idiopathic Scoliosis in a 5-Generation UK Family. Sci Rep 2021, 11, 11026. [Google Scholar] [CrossRef]
- Su, Z.; Yang, Y.; Wang, S.; Zhao, S.; Zhao, H.; Li, X.; Niu, Y.; Qiu, G.; Wu, Z.; et al.; Deciphering Disorders Involving Scoliosis and COmorbidities (DISCO) Study Group The Mutational Landscape of PTK7 in Congenital Scoliosis and Adolescent Idiopathic Scoliosis. Genes 2021, 12, 1791. [Google Scholar] [CrossRef]
- Bradley, E.C.; Cunningham, R.L.; Wilde, C.; Morgan, R.K.; Klug, E.A.; Letcher, S.M.; Schöneberg, T.; Monk, K.R.; Liebscher, I.; Petersen, S.C. In Vivo Identification of Small Molecules Mediating Gpr126/Adgrg6 Signaling during Schwann Cell Development. Ann. N.Y. Acad. Sci. 2019, 1456, 44–63. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Z.; Zhuang, Q.; Fei, Q.; Zhang, J.; Liu, Y.; Wang, Y.; Ding, Y.; Qiu, G. Association Study of Tryptophan Hydroxylase 1 and Arylalkylamine N-Acetyltransferase Polymorphisms With Adolescent Idiopathic Scoliosis in Han Chinese. Spine 2008, 33, 2199–2203. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Gao, X.; Londono, D.; Devroy, S.E.; Mauldin, K.N.; Frankel, J.T.; Brandon, J.M.; Zhang, D.; Li, Q.-Z.; Dobbs, M.B.; et al. Genome-Wide Association Studies of Adolescent Idiopathic Scoliosis Suggest Candidate Susceptibility Genes. Human Molecular Genetics 2011, 20, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Kou, I.; Takahashi, Y.; Johnson, T.A.; Takahashi, A.; Guo, L.; Dai, J.; Qiu, X.; Sharma, S.; Takimoto, A.; Ogura, Y.; et al. Genetic Variants in GPR126 Are Associated with Adolescent Idiopathic Scoliosis. Nat Genet 2013, 45, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Lv, F.; Zhu, Z.; Qian, B.; Wang, B.; Yu, Y.; Qiu, Y. Lack of Association between the CHL1 Gene and Adolescent Idiopathic Scoliosis Susceptibility in Han Chinese: A Case-Control Study. BMC Musculoskelet Disord 2014, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y.; Kou, I.; Miura, S.; Takahashi, A.; Xu, L.; Takeda, K.; Takahashi, Y.; Kono, K.; Kawakami, N.; Uno, K.; et al. A Functional SNP in BNC2 Is Associated with Adolescent Idiopathic Scoliosis. The American Journal of Human Genetics 2015, 97, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Tang, N.L.-S.; Xu, L.; Qin, X.; Mao, S.; Song, Y.; Liu, L.; Li, F.; Liu, P.; Yi, L.; et al. Genome-Wide Association Study Identifies New Susceptibility Loci for Adolescent Idiopathic Scoliosis in Chinese Girls. Nat Commun 2015, 6, 8355. [Google Scholar] [CrossRef] [PubMed]
- Chettier, R.; Nelson, L.; Ogilvie, J.W.; Albertsen, H.M.; Ward, K. Haplotypes at LBX1 Have Distinct Inheritance Patterns with Opposite Effects in Adolescent Idiopathic Scoliosis. PLoS ONE 2015, 10, e0117708. [Google Scholar] [CrossRef]
- Nada, D.; Julien, C.; Samuels, M.E.; Moreau, A. A Replication Study for Association of LBX1 Locus With Adolescent Idiopathic Scoliosis in French–Canadian Population. Spine 2018, 43, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kou, I.; Otomo, N.; Takeda, K.; Momozawa, Y.; Lu, H.-F.; Kubo, M.; Kamatani, Y.; Ogura, Y.; Takahashi, Y.; Nakajima, M.; et al. Genome-Wide Association Study Identifies 14 Previously Unreported Susceptibility Loci for Adolescent Idiopathic Scoliosis in Japanese. Nat Commun 2019, 10, 3685. [Google Scholar] [CrossRef]
- Xia, C.; Xu, L.; Xue, B.; Sheng, F.; Qiu, Y.; Zhu, Z. Rare Variant of HSPG2 Is Not Involved in the Development of Adolescent Idiopathic Scoliosis: Evidence from a Large-Scale Replication Study. BMC Musculoskelet Disord 2019, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Sheng, F.; Xia, C.; Xu, L.; Qin, X.; Tang, N.L.-S.; Qiu, Y.; Cheng, J.C.-Y.; Zhu, Z. New Evidence Supporting the Role of FBN1 in the Development of Adolescent Idiopathic Scoliosis. Spine 2019, 44, E225–E232. [Google Scholar] [CrossRef]
- Xu, L.; Sheng, F.; Xia, C.; Li, Y.; Feng, Z.; Qiu, Y.; Zhu, Z. Common Variant of POC5 Is Associated With the Susceptibility of Adolescent Idiopathic Scoliosis. Spine 2018, 43, E683–E688. [Google Scholar] [CrossRef]
- Mathieu, H.; Spataru, A.; Aragon-Martin, J.A.; Child, A.; Barchi, S.; Fortin, C.; Parent, S.; Moldovan, F. Prevalence of POC5 Coding Variants in French-Canadian and British AIS Cohort. Genes 2021, 12, 1032. [Google Scholar] [CrossRef]
- Terhune, E.A.; Wethey, C.I.; Cuevas, M.T.; Monley, A.M.; Baschal, E.E.; Bland, M.R.; Baschal, R.; Trahan, G.D.; Taylor, M.R.G.; Jones, K.L.; et al. Whole Exome Sequencing of 23 Multigeneration Idiopathic Scoliosis Families Reveals Enrichments in Cytoskeletal Variants, Suggests Highly Polygenic Disease. Genes 2021, 12, 922. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, S.; Liu, L.; Du, H.; Zhao, H.; Wang, S.; Niu, Y.; Li, X.; Qiu, G.; Wu, Z.; et al. Aberrant Interaction between Mutated ADAMTSL2 and LTBP4 Is Associated with Adolescent Idiopathic Scoliosis. Gene 2022, 814, 146126. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-R.; Chou, P.-H.; Huang, K.-J.; Ting, J.; Liu, C.-Y.; Chou, W.-H.; Lin, G.-H.; Chang, J.-G.; Ikegawa, S.; Wang, S.-T.; et al. Whole-Exome Sequencing Identifies Genetic Variants for Severe Adolescent Idiopathic Scoliosis in a Taiwanese Population. JPM 2022, 13, 32. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Z.; Shu, L.; Meng, Y.; Ma, J.; Gao, R.; Zhou, X. A Genetic Variant of the ROBO3 Gene Is Associated With Adolescent Idiopathic Scoliosis in the Chinese Population. Spine 2023, 48, E20–E24. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Londono, D.; Eckalbar, W.L.; Gao, X.; Zhang, D.; Mauldin, K.; Kou, I.; Takahashi, A.; Matsumoto, M.; Kamiya, N.; et al. A PAX1 Enhancer Locus Is Associated with Susceptibility to Idiopathic Scoliosis in Females. Nat Commun 2015, 6, 6452. [Google Scholar] [CrossRef]
- Einarsdottir, E.; Grauers, A.; Wang, J.; Jiao, H.; Escher, S.A.; Danielsson, A.; Simony, A.; Andersen, M.; Christensen, S.B.; Åkesson, K.; et al. CELSR2 Is a Candidate Susceptibility Gene in Idiopathic Scoliosis. PLoS ONE 2017, 12, e0189591. [Google Scholar] [CrossRef] [PubMed]
- Terhune, E.A.; Cuevas, M.T.; Monley, A.M.; Wethey, C.I.; Chen, X.; Cattell, M.V.; Bayrak, M.N.; Bland, M.R.; Sutphin, B.; Trahan, G.D.; et al. Mutations in KIF7 Implicated in Idiopathic Scoliosis in Humans and Axial Curvatures in Zebrafish. Human Mutation 2021, 42, 392–407. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, Y.; Liu, S.; Song, X.; Yang, X.-Z.; Fan, Y.; Chen, W.; Akdemir, Z.C.; Yan, Z.; et al.; DISCO (Deciphering disorders Involving Scoliosis and COmorbidities) Study The Coexistence of Copy Number Variations (CNVs) and Single Nucleotide Polymorphisms (SNPs) at a Locus Can Result in Distorted Calculations of the Significance in Associating SNPs to Disease. Hum Genet 2018, 137, 553–567. [Google Scholar] [CrossRef]
- Hu, Z.; Xu, Y.; Li, J.; Zhu, Z.; Qiu, Y.; Liu, Z. Bioinformatics Analysis and Experimental Verification Identify Downregulation of COL27A1 in Poor Segmental Congenital Scoliosis. Computational and Mathematical Methods in Medicine 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Lu, C.; Yang, D.; Lei, C.; Wang, R.; Guo, T.; Luo, H. Identification of Two Novel DNAAF2 Variants in Two Consanguineous Families with Primary Ciliary Dyskinesia. PGPM 2021, Volume 14, 1415–1423. [Google Scholar] [CrossRef]
- Tan, Z.; Shek, H.T.; Chen, P.; Dong, Z.; Zhou, Y.; Yin, S.; Qiu, A.; Dong, L.; Gao, B.; To, M.K.T. Clinical Features and Molecular Characterization of Chinese Patients with FKBP10 Variants. Molec Gen & Gen Med 2023, 11, e2122. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, S.; Hu, W.; Jin, P. Analysis of a child with Marfan syndrome due to a novel variant of FBN1 gene. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2023, 40, 62–65. [Google Scholar] [CrossRef]
- Hendrickx, G.; Boudin, E.; Steenackers, E.; Collet, C.; Mortier, G.R.; Geneviève, D.; Van Hul, W. A Recessive Form of Craniodiaphyseal Dysplasia Caused by a Homozygous Missense Variant in SP7/Osterix. Bone 2023, 167, 116633. [Google Scholar] [CrossRef]
- Hjeij, R.; Onoufriadis, A.; Watson, C.M.; Slagle, C.E.; Klena, N.T.; Dougherty, G.W.; Kurkowiak, M.; Loges, N.T.; Diggle, C.P.; Morante, N.F.C.; et al. CCDC151 Mutations Cause Primary Ciliary Dyskinesia by Disruption of the Outer Dynein Arm Docking Complex Formation. Am J Hum Genet 2014, 95, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Tarkar, A.; Loges, N.T.; Slagle, C.E.; Francis, R.; Dougherty, G.W.; Tamayo, J.V.; Shook, B.; Cantino, M.; Schwartz, D.; Jahnke, C.; et al. DYX1C1 Is Required for Axonemal Dynein Assembly and Ciliary Motility. Nat Genet 2013, 45, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Becker-Heck, A.; Zohn, I.E.; Okabe, N.; Pollock, A.; Lenhart, K.B.; Sullivan-Brown, J.; McSheene, J.; Loges, N.T.; Olbrich, H.; Haeffner, K.; et al. The Coiled-Coil Domain Containing Protein CCDC40 Is Essential for Motile Cilia Function and Left-Right Axis Formation. Nat Genet 2011, 43, 79–84. [Google Scholar] [CrossRef]
- Austin-Tse, C.; Halbritter, J.; Zariwala, M.A.; Gilberti, R.M.; Gee, H.Y.; Hellman, N.; Pathak, N.; Liu, Y.; Panizzi, J.R.; Patel-King, R.S.; et al. Zebrafish Ciliopathy Screen Plus Human Mutational Analysis Identifies C21orf59 and CCDC65 Defects as Causing Primary Ciliary Dyskinesia. The American Journal of Human Genetics 2013, 93, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Cantaut-Belarif, Y.; Sternberg, J.R.; Thouvenin, O.; Wyart, C.; Bardet, P.-L. The Reissner Fiber in the Cerebrospinal Fluid Controls Morphogenesis of the Body Axis. Current Biology 2018, 28, 2479–2486. [Google Scholar] [CrossRef]
- Troutwine, B.R.; Gontarz, P.; Konjikusic, M.J.; Minowa, R.; Monstad-Rios, A.; Sepich, D.S.; Kwon, R.Y.; Solnica-Krezel, L.; Gray, R.S. The Reissner Fiber Is Highly Dynamic In Vivo and Controls Morphogenesis of the Spine. Current Biology 2020, 30, 2353–2362. [Google Scholar] [CrossRef]
- Rose, C.D.; Pompili, D.; Henke, K.; Van Gennip, J.L.M.; Meyer-Miner, A.; Rana, R.; Gobron, S.; Harris, M.P.; Nitz, M.; Ciruna, B. SCO-Spondin Defects and Neuroinflammation Are Conserved Mechanisms Driving Spinal Deformity across Genetic Models of Idiopathic Scoliosis. Current Biology 2020, 30, 2363–2373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Troutwine, B.R.; Zhang, H.; Gray, R.S. The Axonemal Dynein Heavy Chain 10 Gene Is Essential for Monocilia Motility and Spine Alignment in Zebrafish. Developmental Biology 2022, 482, 82–90. [Google Scholar] [CrossRef]
- Yang, S.; Emelyanov, A.; You, M.-S.; Sin, M.; Korzh, V. Camel Regulates Development of the Brain Ventricular System. Cell Tissue Res 2021, 383, 835–852. [Google Scholar] [CrossRef] [PubMed]
- Jussila, M.; Boswell, C.W.; Griffiths, N.W.; Pumputis, P.G.; Ciruna, B. Live Imaging and Conditional Disruption of Native PCP Activity Using Endogenously Tagged Zebrafish SfGFP-Vangl2. Nat Commun 2022, 13, 5598. [Google Scholar] [CrossRef]
- Lu, H.; Shagirova, A.; Goggi, J.L.; Yeo, H.L.; Roy, S. Reissner Fibre-Induced Urotensin Signalling from Cerebrospinal Fluid-Contacting Neurons Prevents Scoliosis of the Vertebrate Spine. Biology Open 2020, 9, bio052027. [Google Scholar] [CrossRef]
- Meyer-Miner, A.; Van Gennip, J.L.M.; Henke, K.; Harris, M.P.; Ciruna, B. Resolving Primary Pathomechanisms Driving Idiopathic-like Spinal Curvature Using a New Katnb1 Scoliosis Model. iScience 2022, 25, 105028. [Google Scholar] [CrossRef]
- Xie, H.; Kang, Y.; Liu, J.; Huang, M.; Dai, Z.; Shi, J.; Wang, S.; Li, L.; Li, Y.; Zheng, P.; et al. Ependymal Polarity Defects Coupled with Disorganized Ciliary Beating Drive Abnormal Cerebrospinal Fluid Flow and Spine Curvature in Zebrafish. PLoS Biol 2023, 21, e3002008. [Google Scholar] [CrossRef]
- Vesque, C.; Anselme, I.; Pezeron, G.; Cantaut-Belarif, Y.; Eschstruth, A.; Djebar, M.; Santos, D.L.; Ribeuz, H.L.; Jenett, A.; Khoury, H.; et al. Loss of the Reissner Fiber and Increased URP Neuropeptide Signaling Underlie Scoliosis in a Zebrafish Ciliopathy Mutant; Developmental Biology, 2019.
- Marie-Hardy, L.; Cantaut-Belarif, Y.; Pietton, R.; Slimani, L.; Pascal-Moussellard, H. The Orthopedic Characterization of Cfap298tm304 Mutants Validate Zebrafish to Faithfully Model Human AIS. Sci Rep 2021, 11, 7392. [Google Scholar] [CrossRef]
- Wang, Y.; Troutwine, B.R.; Zhang, H.; Gray, R.S. The Axonemal Dynein Heavy Chain 10 Gene Is Essential for Monocilia Motility and Spine Alignment in Zebrafish. Developmental Biology 2022, 482, 82–90. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Ou, Y.; Wu, J.; Li, H.; Wang, X.; Tang, L.; Dai, X.; Yang, C.; Wei, Z.; et al. Ccdc57 Is Required for Straightening the Body Axis by Regulating Ciliary Motility in the Brain Ventricle of Zebrafish. Journal of Genetics and Genomics 2023, 50, 253–263. [Google Scholar] [CrossRef]
- Mathieu, H.; Patten, S.A.; Aragon-Martin, J.A.; Ocaka, L.; Simpson, M.; Child, A.; Moldovan, F. Genetic Variant of TTLL11 Gene and Subsequent Ciliary Defects Are Associated with Idiopathic Scoliosis in a 5-Generation UK Family. Sci Rep 2021, 11, 11026. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Miner, A.; Van Gennip, J.L.M.; Henke, K.; Harris, M.P.; Ciruna, B. Resolving Primary Pathomechanisms Driving Idiopathic-like Spinal Curvature Using a New Katnb1 Scoliosis Model. iScience 2022, 25, 105028. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Kang, Y.; Liu, J.; Huang, M.; Dai, Z.; Shi, J.; Wang, S.; Li, L.; Li, Y.; Zheng, P.; et al. Ependymal Polarity Defects Coupled with Disorganized Ciliary Beating Drive Abnormal Cerebrospinal Fluid Flow and Spine Curvature in Zebrafish. PLoS Biol 2023, 21, e3002008. [Google Scholar] [CrossRef] [PubMed]
- Grimes, D.T.; Boswell, C.W.; Morante, N.F.C.; Henkelman, R.M.; Burdine, R.D.; Ciruna, B. Zebrafish Models of Idiopathic Scoliosis Link Cerebrospinal Fluid Flow Defects to Spine Curvature. Science 2016, 352, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Buchan, J.G.; Gray, R.S.; Gansner, J.M.; Alvarado, D.M.; Burgert, L.; Gitlin, J.D.; Gurnett, C.A.; Goldsmith, M.I. Kinesin Family Member 6 (Kif6) Is Necessary for Spine Development in Zebrafish: KIF6 in Zebrafish Spine Development. Dev. Dyn. 2014, 243, 1646–1657. [Google Scholar] [CrossRef]
- Konjikusic, M.J.; Yeetong, P.; Boswell, C.W.; Lee, C.; Roberson, E.C.; Ittiwut, R.; Suphapeetiporn, K.; Ciruna, B.; Gurnett, C.A.; Wallingford, J.B.; et al. Mutations in Kinesin Family Member 6 Reveal Specific Role in Ependymal Cell Ciliogenesis and Human Neurological Development. PLoS Genet 2018, 14, e1007817. [Google Scholar] [CrossRef]
- Becker-Heck, A.; Zohn, I.E.; Okabe, N.; Pollock, A.; Lenhart, K.B.; Sullivan-Brown, J.; McSheene, J.; Loges, N.T.; Olbrich, H.; Haeffner, K.; et al. The Coiled-Coil Domain Containing Protein CCDC40 Is Essential for Motile Cilia Function and Left-Right Axis Formation. Nat Genet 2011, 43, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Tarkar, A.; Loges, N.T.; Slagle, C.E.; Francis, R.; Dougherty, G.W.; Tamayo, J.V.; Shook, B.; Cantino, M.; Schwartz, D.; et al.; UK10K DYX1C1 Is Required for Axonemal Dynein Assembly and Ciliary Motility. Nat Genet 2013, 45, 995–1003. [Google Scholar] [CrossRef]
- Hjeij, R.; Onoufriadis, A.; Watson, C.M.; Slagle, C.E.; Klena, N.T.; Dougherty, G.W.; Kurkowiak, M.; Loges, N.T.; Diggle, C.P.; Morante, N.F.C.; et al. CCDC151 Mutations Cause Primary Ciliary Dyskinesia by Disruption of the Outer Dynein Arm Docking Complex Formation. The American Journal of Human Genetics 2014, 95, 257–274. [Google Scholar] [CrossRef]
- Kobayashi, D.; Asano-Hoshino, A.; Nakakura, T.; Nishimaki, T.; Ansai, S.; Kinoshita, M.; Ogawa, M.; Hagiwara, H.; Yokoyama, T. Loss of Zinc Finger MYND-Type Containing 10 (Zmynd10) Affects Cilia Integrity and Axonemal Localization of Dynein Arms, Resulting in Ciliary Dysmotility, Polycystic Kidney and Scoliosis in Medaka (Oryzias Latipes). Developmental Biology 2017, 430, 69–79. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, S.; Chen, Z.; Chong, Y.L.; Xie, H.; Feng, D.; Wu, X.; Song, D.Z.; Roy, S.; Zhao, C. Cilia-Driven Cerebrospinal Fluid Flow Directs Expression of Urotensin Neuropeptides to Straighten the Vertebrate Body Axis. Nat Genet 2018, 50, 1666–1673. [Google Scholar] [CrossRef]
- Austin-Tse, C.; Halbritter, J.; Zariwala, M.A.; Gilberti, R.M.; Gee, H.Y.; Hellman, N.; Pathak, N.; Liu, Y.; Panizzi, J.R.; Patel-King, R.S.; et al. Zebrafish Ciliopathy Screen Plus Human Mutational Analysis Identifies C21orf59 and CCDC65 Defects as Causing Primary Ciliary Dyskinesia. The American Journal of Human Genetics 2013, 93, 672–686. [Google Scholar] [CrossRef]
- Lessieur, E.M.; Song, P.; Nivar, G.C.; Piccillo, E.M.; Fogerty, J.; Rozic, R.; Perkins, B.D. Ciliary Genes Arl13b, Ahi1 and Cc2d2a Differentially Modify Expression of Visual Acuity Phenotypes but Do Not Enhance Retinal Degeneration Due to Mutation of Cep290 in Zebrafish. PLoS ONE 2019, 14, e0213960. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Thomas, H.R.; Thompson, R.G.; Waldrep, S.C.; Fogerty, J.; Song, P.; Li, Z.; Ma, Y.; Santra, P.; Hoover, J.D.; et al. Variable Phenotypes and Penetrance between and within Different Zebrafish Ciliary Transition Zone Mutants. Disease Models & Mechanisms 2022, 15, dmm049568. [Google Scholar] [CrossRef]
- Bentley-Ford, M.R.; LaBonty, M.; Thomas, H.R.; Haycraft, C.J.; Scott, M.; LaFayette, C.; Croyle, M.J.; Andersen, R.S.; Parant, J.M.; Yoder, B.K. Evolutionarily Conserved Genetic Interactions between Nphp-4 and Bbs-5 Mutations Exacerbate Ciliopathy Phenotypes. Genetics 2022, 220, iyab209. [Google Scholar] [CrossRef] [PubMed]
- Kou, I.; Takahashi, Y.; Johnson, T.A.; Takahashi, A.; Guo, L.; Dai, J.; Qiu, X.; Sharma, S.; Takimoto, A.; Ogura, Y.; et al. Genetic Variants in GPR126 Are Associated with Adolescent Idiopathic Scoliosis. Nat Genet 2013, 45, 676–679. [Google Scholar] [CrossRef]
- Musa, G.; Cazorla-Vázquez, S.; Amerongen, M.J.; Stemmler, M.P.; Eckstein, M.; Hartmann, A.; Braun, T.; Brabletz, T.; Engel, F.B. Gpr126 (Adgrg6) Is Expressed in Cell Types Known to Be Exposed to Mechanical Stimuli. Ann. N.Y. Acad. Sci. 2019, 1456, 96–108. [Google Scholar] [CrossRef]
- Ogura, Y.; Kou, I.; Miura, S.; Takahashi, A.; Xu, L.; Takeda, K.; Takahashi, Y.; Kono, K.; Kawakami, N.; Uno, K.; et al. A Functional SNP in BNC2 Is Associated with Adolescent Idiopathic Scoliosis. The American Journal of Human Genetics 2015, 97, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Chen, C.; Xu, C.; Zhou, H.; Gao, B.; Su, D.; Liao, Z.; Li, Y.; Yang, S.; Su, P. Mutant MAPK7-Induced Idiopathic Scoliosis Is Linked to Impaired Osteogenesis. Cell Physiol Biochem 2018, 48, 880–890. [Google Scholar] [CrossRef]
- Guo, L.; Yamashita, H.; Kou, I.; Takimoto, A.; Meguro-Horike, M.; Horike, S.; Sakuma, T.; Miura, S.; Adachi, T.; Yamamoto, T.; et al. Functional Investigation of a Non-Coding Variant Associated with Adolescent Idiopathic Scoliosis in Zebrafish: Elevated Expression of the Ladybird Homeobox Gene Causes Body Axis Deformation. PLoS Genet 2016, 12, e1005802. [Google Scholar] [CrossRef]
- Decourtye, L.; McCallum-Loudeac, J.A.; Zellhuber-McMillan, S.; Young, E.; Sircombe, K.J.; Wilson, M.J. Characterization of a Novel Lbx1 Mouse Loss of Function Strain. Differentiation 2022, 123, 30–41. [Google Scholar] [CrossRef]
- Matsuhashi, Y.; Horiuchi, K.; Nakagawa, T.; Takahashi, Y.; Imabayashi, H.; Hosogane, N.; Watanabe, K.; Matsumoto, M.; Chiba, K. Abrogation of LBX1 in Skeletal Muscle Results in Hypoplastic Limbs and Progressive Kyphosis in Mice. Journal Orthopaedic Research 2023, 41, 884–890. [Google Scholar] [CrossRef]
- Yonezawa, Y.; Guo, L.; Kakinuma, H.; Otomo, N.; Yoshino, S.; Takeda, K.; Nakajima, M.; Shiraki, T.; Ogura, Y.; Takahashi, Y.; et al. Identification of a Functional Susceptibility Variant for Adolescent Idiopathic Scoliosis That Upregulates Early Growth Response 1 (EGR1)-Mediated UNCX Expression. J of Bone & Mineral Res 2023, 38, 144–153. [Google Scholar] [CrossRef]
- Chen, C.; Tan, H.; Bi, J.; Li, L.; Rong, T.; Lin, Y.; Sun, P.; Liang, J.; Jiao, Y.; Li, Z.; et al. LncRNA-SULT1C2A Regulates Foxo4 in Congenital Scoliosis by Targeting Rno-miR-466c-5p through PI3K-ATK Signalling. J Cell Mol Med 2019, 23, 4582–4591. [Google Scholar] [CrossRef]
- Patten, S.A.; Margaritte-Jeannin, P.; Bernard, J.-C.; Alix, E.; Labalme, A.; Besson, A.; Girard, S.L.; Fendri, K.; Fraisse, N.; Biot, B.; et al. Functional Variants of POC5 Identified in Patients with Idiopathic Scoliosis. J. Clin. Invest. 2015, 125, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Terhune, E.A.; Cuevas, M.T.; Monley, A.M.; Wethey, C.I.; Chen, X.; Cattell, M.V.; Bayrak, M.N.; Bland, M.R.; Sutphin, B.; Trahan, G.D.; et al. Mutations in KIF7 Implicated in Idiopathic Scoliosis in Humans and Axial Curvatures in Zebrafish. Human Mutation 2021, 42, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Seda, M.; Crespo, B.; Corcelli, M.; Osborn, D.P.; Jenkins, D. A CRISPR/Cas9-Generated Mutation in the Zebrafish Orthologue of PPP2R3B Causes Idiopathic Scoliosis. Sci Rep 2023, 13, 6783. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, S.; Chen, Z.; Chong, Y.L.; Xie, H.; Feng, D.; Wu, X.; Song, D.Z.; Roy, S.; Zhao, C. Cilia-Driven Cerebrospinal Fluid Flow Directs Expression of Urotensin Neuropeptides to Straighten the Vertebrate Body Axis. Nat Genet 2018, 50, 1666–1673. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, A.; Ziganshin, B.A.; Papanikolaou, D.; Zafar, M.A.; Nicoli, S.; Mukherjee, S.; Elefteriades, J.A. Phenotyping Zebrafish Mutant Models to Assess Candidate Genes Associated with Aortic Aneurysm. Genes 2022, 13, 123. [Google Scholar] [CrossRef]
- Sternberg, J.R.; Prendergast, A.E.; Brosse, L.; Cantaut-Belarif, Y.; Thouvenin, O.; Orts-Del’Immagine, A.; Castillo, L.; Djenoune, L.; Kurisu, S.; McDearmid, J.R.; et al. Pkd2l1 Is Required for Mechanoception in Cerebrospinal Fluid-Contacting Neurons and Maintenance of Spine Curvature. Nat Commun 2018, 9, 3804. [Google Scholar] [CrossRef]
- Bearce, E.A.; Irons, Z.H.; O’Hara-Smith, J.R.; Kuhns, C.J.; Fisher, S.I.; Crow, W.E.; Grimes, D.T. Urotensin II-Related Peptides, Urp1 and Urp2, Control Zebrafish Spine Morphology. eLife 2022, 11, e83883. [Google Scholar] [CrossRef]
- Gaillard, A.-L.; Mohamad, T.; Quan, F.B.; De Cian, A.; Mosimann, C.; Tostivint, H.; Pézeron, G. Urp1 and Urp2 Act Redundantly to Maintain Spine Shape in Zebrafish Larvae. Developmental Biology 2023, 496, 36–51. [Google Scholar] [CrossRef]
- Sun, X.; Zhou, Y.; Zhang, R.; Wang, Z.; Xu, M.; Zhang, D.; Huang, J.; Luo, F.; Li, F.; Ni, Z.; et al. Dstyk Mutation Leads to Congenital Scoliosis-like Vertebral Malformations in Zebrafish via Dysregulated MTORC1/TFEB Pathway. Nat Commun 2020, 11, 479. [Google Scholar] [CrossRef] [PubMed]
- Bagwell, J.; Norman, J.; Ellis, K.; Peskin, B.; Hwang, J.; Ge, X.; Nguyen, S.V.; McMenamin, S.K.; Stainier, D.Y.; Bagnat, M. Notochord Vacuoles Absorb Compressive Bone Growth during Zebrafish Spine Formation. Elife 2020, 9, e51221. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Zhang, X.; Jia, S.; Yelick, P.C.; Zhao, C. Zebrafish as a Model for Human Ciliopathies. Journal of Genetics and Genomics 2016, 43, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Scholey, J.M. Assembly, Functions and Evolution of Archaella, Flagella and Cilia. Current Biology 2018, 28, R278–R292. [Google Scholar] [CrossRef]
- Klena, N.; Pigino, G. Structural Biology of Cilia and Intraflagellar Transport. Annu. Rev. Cell Dev. Biol. 2022, 38, annurev. [Google Scholar] [CrossRef]
- Van De Weghe, J.C.; Rusterholz, T.D.S.; Latour, B.; Grout, M.E.; Aldinger, K.A.; Shaheen, R.; Dempsey, J.C.; Maddirevula, S.; Cheng, Y.-H.H.; Phelps, I.G.; et al. Mutations in ARMC9, Which Encodes a Basal Body Protein, Cause Joubert Syndrome in Humans and Ciliopathy Phenotypes in Zebrafish. The American Journal of Human Genetics 2017, 101, 23–36. [Google Scholar] [CrossRef]
- Lessieur, E.M.; Song, P.; Nivar, G.C.; Piccillo, E.M.; Fogerty, J.; Rozic, R.; Perkins, B.D. Ciliary Genes Arl13b, Ahi1 and Cc2d2a Differentially Modify Expression of Visual Acuity Phenotypes but Do Not Enhance Retinal Degeneration Due to Mutation of Cep290 in Zebrafish. PLoS ONE 2019, 14, e0213960. [Google Scholar] [CrossRef]
- Bentley-Ford, M.R.; LaBonty, M.; Thomas, H.R.; Haycraft, C.J.; Scott, M.; LaFayette, C.; Croyle, M.J.; Andersen, R.S.; Parant, J.M.; Yoder, B.K. Evolutionarily Conserved Genetic Interactions between Nphp-4 and Bbs-5 Mutations Exacerbate Ciliopathy Phenotypes. Genetics 2022, 220, iyab209. [Google Scholar] [CrossRef]
- Masek, M.; Etard, C.; Hofmann, C.; Hülsmeier, A.J.; Zang, J.; Takamiya, M.; Gesemann, M.; Neuhauss, S.C.F.; Hornemann, T.; Strähle, U.; et al. Loss of the Bardet-Biedl Protein Bbs1 Alters Photoreceptor Outer Segment Protein and Lipid Composition. Nat Commun 2022, 13, 1282. [Google Scholar] [CrossRef]
- Yu, X.; Ng, C.P.; Habacher, H.; Roy, S. Foxj1 Transcription Factors Are Master Regulators of the Motile Ciliogenic Program. Nat Genet 2008, 40, 1445–1453. [Google Scholar] [CrossRef]
- Grimes, D.T.; Boswell, C.W.; Morante, N.F.C.; Henkelman, R.M.; Burdine, R.D.; Ciruna, B. Zebrafish Models of Idiopathic Scoliosis Link Cerebrospinal Fluid Flow Defects to Spine Curvature. Science 2016, 352, 1341–1344. [Google Scholar] [CrossRef]
- Mathieu, H.; Patten, S.A.; Aragon-Martin, J.A.; Ocaka, L.; Simpson, M.; Child, A.; Moldovan, F. Genetic Variant of TTLL11 Gene and Subsequent Ciliary Defects Are Associated with Idiopathic Scoliosis in a 5-Generation UK Family. Sci Rep 2021, 11, 11026. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Kang, Y.; Liu, J.; Huang, M.; Dai, Z.; Shi, J.; Wang, S.; Li, L.; Li, Y.; Zheng, P.; et al. Ependymal Polarity Defects Coupled with Disorganized Ciliary Beating Drive Abnormal Cerebrospinal Fluid Flow and Spine Curvature in Zebrafish. PLoS Biol 2023, 21, e3002008. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, J.; Ou, Y.; Wu, J.; Li, H.; Wang, X.; Tang, L.; Dai, X.; Yang, C.; Wei, Z.; et al. Ccdc57 Is Required for Straightening the Body Axis by Regulating Ciliary Motility in the Brain Ventricle of Zebrafish. Journal of Genetics and Genomics 2023, 50, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.; Yang, G.; Gao, Q.; Xiao, L.; Li, J.; Guo, C.; Troutwine, B.R.; Gray, R.S.; Xie, L.; et al. Coding Variants Coupled With Rapid Modeling in Zebrafish Implicate Dynein Genes, Dnaaf1 and Zmynd10, as Adolescent Idiopathic Scoliosis Candidate Genes. Front Cell Dev Biol 2020, 8, 582255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jia, S.; Chen, Z.; Chong, Y.L.; Xie, H.; Feng, D.; Wu, X.; Song, D.Z.; Roy, S.; Zhao, C. Cilia-Driven Cerebrospinal Fluid Flow Directs Expression of Urotensin Neuropeptides to Straighten the Vertebrate Body Axis. Nat Genet 2018, 50, 1666–1673. [Google Scholar] [CrossRef]
- Aboitiz, F.; Montiel, J.F. The Enigmatic Reissner’s Fiber and the Origin of Chordates. Front. Neuroanat. 2021, 15, 703835. [Google Scholar] [CrossRef]
- Troutwine, B.R.; Gontarz, P.; Konjikusic, M.J.; Minowa, R.; Monstad-Rios, A.; Sepich, D.S.; Kwon, R.Y.; Solnica-Krezel, L.; Gray, R.S. The Reissner Fiber Is Highly Dynamic In Vivo and Controls Morphogenesis of the Spine. Curr Biol 2020, 30, 2353–2362. [Google Scholar] [CrossRef]
- Gobron, S.; Creveaux, I.; Meiniel, R.; Didier, R.; Dastugue, B.; Meiniel, A. SCO-Spondin Is Evolutionarily Conserved in the Central Nervous System of the Chordate Phylum. Neuroscience 1999, 88, 655–664. [Google Scholar] [CrossRef]
- Cantaut-Belarif, Y.; Sternberg, J.R.; Thouvenin, O.; Wyart, C.; Bardet, P.-L. The Reissner Fiber in the Cerebrospinal Fluid Controls Morphogenesis of the Body Axis. Current Biology 2018, 28, 2479–2486. [Google Scholar] [CrossRef]
- Troutwine, B.R.; Gontarz, P.; Konjikusic, M.J.; Minowa, R.; Monstad-Rios, A.; Sepich, D.S.; Kwon, R.Y.; Solnica-Krezel, L.; Gray, R.S. The Reissner Fiber Is Highly Dynamic In Vivo and Controls Morphogenesis of the Spine. Current Biology 2020, 30, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Driever, W. Developmental Biology: Reissner’s Fiber and Straightening of the Body Axis. Current Biology 2018, 28, R833–R835. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Perdomo, A.; Carmona Calero, E.; H, P.-G.; Valenzuela, I.; P, P.-M.; C, O.-R.; González-Marrero, I.; A, T.-D.; R, F.-T. Ontogenic Development of the Human Subcommissural Organ. European Journal of Anatomy 2004, 8, 107–120. [Google Scholar]
Figure 1.
Schematic diagram of the structure of the zebrafish spinal cord.
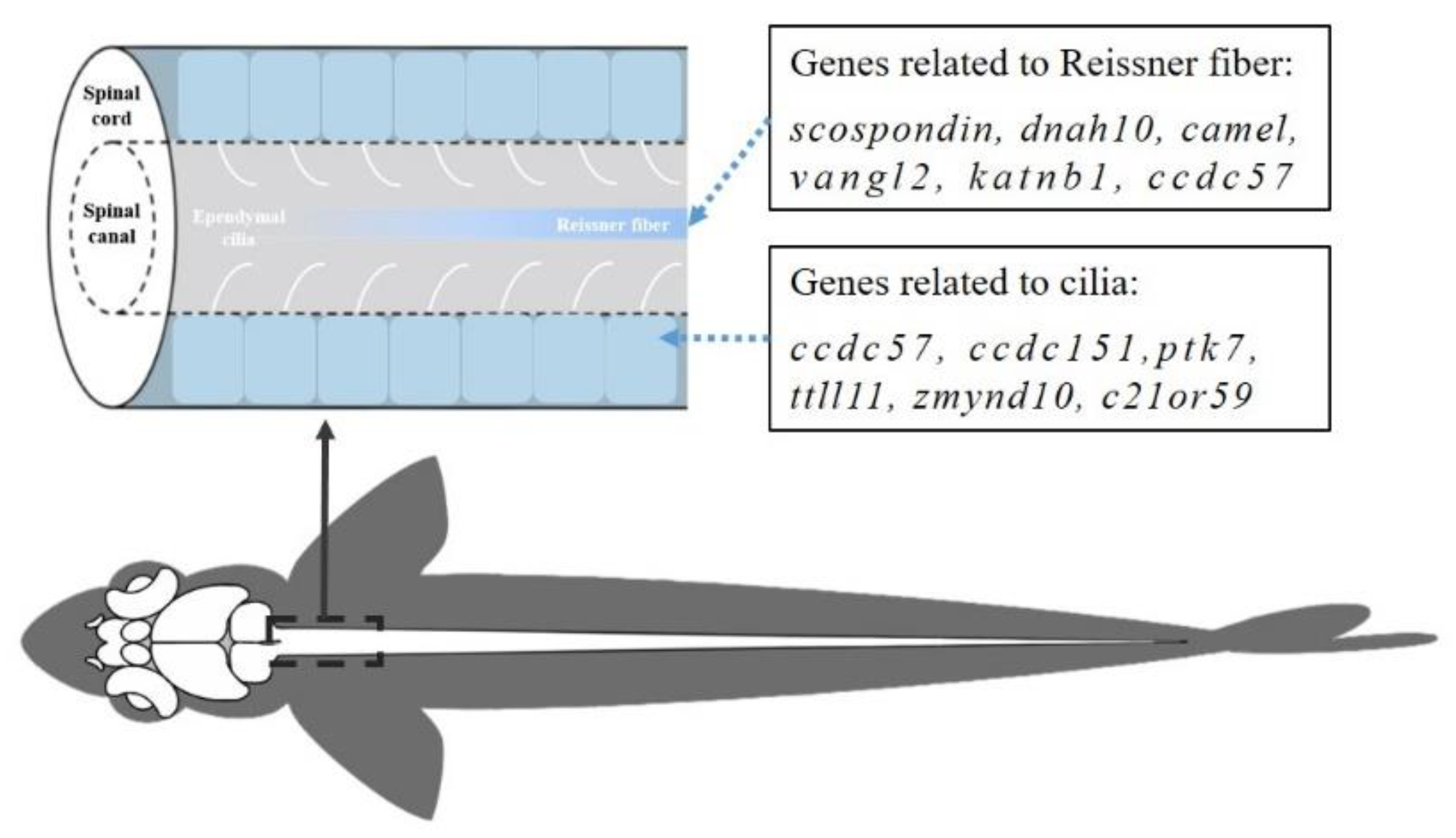
Table 1.
Human-derived candidate genes for spinal deformity.
| Related diseases | Gene | Sample source | Year | Reference |
|---|---|---|---|---|
| AIS | TPH1 | 103 AIS cases and 108 controls (Chinese Han population) | 2008 | [59] |
| DSCAM | 419 AIS families | 2011 | [60] | |
| CNTNAP2 | ||||
| GPR126 | 1,819 AIS cases and 25,939 controls | 2013 | [61] | |
| CHL1 | 500 AIS cases and 500 controls (10-18 years old females in a Han Chinese population) | 2014 | [62] | |
| BNC2 | 2,109 AIS cases and 11,140 controls | 2015 | [63] | |
| AJAP1 | 4,317 AIS cases and 6,016 controls (Chinese Han population) | 2015 | [64] | |
| PAX3 | ||||
| EPHA4 | ||||
| BCL-2 | ||||
| LBX1 | 620 AIS cases and 1,287 controls (female Caucasian) | 2015 | [65] | |
| 667 AIS cases and 901 controls (French-Canadian population) | 2018 | [66] | ||
| TBX1 | 79,211 individuals (Japanese population) | 2019 | [67] | |
| DSE | ||||
| FTO | ||||
| BOC | ||||
| HSPG2 | 1752 AIS cases and 1584 controls (China) | 2019 | [68] | |
| FBN1 and FBN2 | 952 AIS cases and 1499 controls | 2019 | [69] | |
| POC5 | 2432 AIS cases and 2292 controls (Chinese population) | 2018 | [70] | |
| French-Canadian and British families and sporadic cases | 2021 | [71] | ||
| COL8A2 | 23 Multigeneration Idiopathic Scoliosis Families | 2021 | [72] | |
| COL4A3 | ||||
| COL6A5 | ||||
| COL27A1 | ||||
| COL7A1 | ||||
| COL21A1 | ||||
| COL9A2 | ||||
| COL9A3 | ||||
| COL4A6 | ||||
| HSPG2 | ||||
| FBN2 | ||||
| ADAMTSL2 | 302 AIS cases and 818 controls | 2022 | [73] | |
| LTBP4 | ||||
| TTN | 11 AIS cases (Taiwan) | 2022 | [74] | |
| CLCN1 | ||||
| SOX8 | ||||
| ROBO3 | 135 AIS cases and 267 controls | 2023 | [75] | |
| IS | PAX1 | 3,102 individuals | 2015 | [76] |
| CELSR2 | 1739 IS cases and 1812 controls(Swedish-Danish) 3 IS cases in a multigenerational family |
2017 2020 |
[77] [78] |
|
| KIF7 | ||||
| CS | TBX6 | 161 CS cases and 166 controls (PUMCH in China, Oct. 2010 – Jun. 2014) | 2018 | [79] |
| PSCS | COL27A1 | GSE11854 expression dataset associated with somite formation in the GEO database | 2022 | [80] |
| PCD | DNAAF2 | patients from two consanguineous families | 2021 | [81] |
| OI | FKBP10 | Patients diagnosed with OI | 2023 | [82] |
| MFS | FBN1 | a child with Marfan syndrome | 2023 | [83] |
| CDD | SP7 | 2 CDD cases from a large consanguineous family | 2023 | [84] |
AIS: adolescent idiopathic scoliosis. IS: Idiopathic scoliosis, CS: congenital scoliosis, PSCS: poor segmental congenital scoliosis, PCD: primary ciliary dyskinesia, OI: Osteogenesis imperfecta, MFS: Marfan syndrome, CDD: Craniodiaphyseal dysplasia, PUMCH: Peking Union Medical College Hospital.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



