
Submitted:
12 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
The saphenous vein is the conduit of choice for bypass grafting. Unfortunately, the hemodynamic stress associated with the arterial environment of the bypass vein graft leads to the development of intimal hyperplasia (IH), an excessive cellular growth and collagen deposition that results in restenosis and secondary graft occlusion. Hydrogen sulfide (H2S) is a ubiquitous redox-modifying gasotransmitter that inhibits IH. H2S is produced via the reverse trans-sulfuration pathway by 3 enzymes: cystathionine γ-lyase (CSE), cystathionine β-synthase (CBS), and 3-mercaptopyruvate sulfurtransferase (3-MST). However, the expression and regulation of these enzymes in the human vasculature remains unclear. Here, we investigated the expression of CSE, CBS and 3-MST in segments of native human saphenous vein and large arteries. Furthermore, we evaluated the regulation of these enzymes in vein segments cultured under static, venous (7 mmHg pressure) or arterial (100 mmHg pressure) pressure. CSE was expressed in the media, neointima and intima of the vessels and was negatively regulated by arterial shear stress. Adenoviral-mediated CSE overexpression or RNA interference-mediated CSE knock-down revealed that CSE inhibited primary human VSMC migration but not proliferation. We propose that high shear stress in arteriovenous bypass grafts inhibits CSE expression in both the media and endothelium, which may contribute to increased VSMC migration in the context of IH.
Keywords:
Hydrogen sulfide (H2S)
; Cystathionine gamma-Lyase
; intimal hyperplasia
; venous bypass
1. Introduction
Arterio-venous bypass surgery is one of the main approaches for revascularization of chronic limb-threatening ischemia (CLTI) patients. However, veins are not designed to support arterial pressure and undergo significant vascular remodeling to adapt to the arterial environment. This remodeling is accompanied by the development of intimal hyperplasia (IH), i.e., the formation of a collagen-rich neointima layer between the media and the innermost layer (intima/endothelium) of the vein. IH is due to a cascade of cellular events leading to the differentiation, proliferation, and migration of vascular smooth muscle cells (VSMC) from the vessel wall into the intima [1]. This excessive cell growth and collagen deposition eventually lead to reduced blood flow (restenosis) or occlusion of the bypass. 30 to 50% of the saphenous grafts fail 1–18 months after the implantation [2].
Hydrogen sulfide (H2S) contributes to the homeostasis of a wide range of systems, including the cardiovascular systems [3]. Notably, endogenous H2S bioavailability is attenuated in patients with CLTI and in patients with diabetes-related vascular inflammation [4]. Circulating H2S is also reduced in humans suffering from vascular occlusive disease [5,6], and patients undergoing surgical revascularization with lower H2S production capacity have higher postoperative mortality rates [7].
H2S is produced in mammalian cells through the reverse transulfuration pathway by two pyridoxal 5ʹ-phosphate dependent enzymes, cystathionine γ-lyase (CSE) and cystathionine β-synthase (CBS), and by a combination of two additional enzymes, 3-mercaptopyruvate sulfurtransferase (3-MST) and cysteine aminotransferase (CAT). Mice lacking Cse display increased IH in a model of carotid artery ligation [8,9]. On the contrary, Cse overexpression decreases IH formation in a murine model of vein graft by carotid-interposition cuff technique [10]. In addition, we and others demonstrated that systemic treatment using diverse H2S donors inhibit IH in vivo in various models in rats [11], rabbits [12] and mice [8,9,13]. We also showed that several H2S donors inhibit IH ex vivo in human vein segments [9,13,14]. The study of Cse-/- mice supports that CSE expression in endothelial cells (EC) is the main source of endogenous H2S production in vessels [15,16,17,18]. However, CSE expression has also been found in VMSC, and may contribute to VSMC proliferation and migration, vascular remodeling and IH [8,19]. CBS is also found in the cardiovascular system, but its role and distribution in vessels is unclear [3]. Other reports suggest a key role of 3-MST in H2S production in the vascular endothelium [20]. It was recently demonstrated that Cse expression is negatively regulated by shear stress in vitro [21]. This is in line with a previous study showing that only disturbed flow regions show discernible CSE protein expression after carotid artery ligation in the mouse [22]. However, the expression of CSE in human vessels remains poorly characterized. In this study, we studied the expression and regulation of CSE, CBS and 3MST in segments from healthy human saphenous vein and artery. We observed that CSE is expressed both in the endothelium and media of large vessels, while CBS expression is detectable only in the media, and 3-MST expression is mainly restricted to the endothelium of small vessels. Our data confirms that CSE expression in vein is negatively regulated by shear stress and, as a result, upregulated in absence of flow and downregulated in vein segments placed under arterial perfusion. We further confirm that CSE is involved in primary human VSMC migration, but not proliferation.
2. Materials and Methods
For details on materials and reagents please see the Supplementary Table S1 and S2.
2.1. Human vessels culture
Healthy human artery segments were obtained from patients undergoing vascular reconstruction as part of our biobank. 9 artery segments were used in this study. Healthy human saphenous vein segments were surplus segments of non-varicose veins from donors who underwent lower limb bypass surgery. Static vein culture was performed as described [13,14,23]. Briefly, segments of great saphenous vein were cut in 5 mm segments randomly distributed between conditions. One segment (D0) was immediately preserved in formalin or flash frozen in liquid nitrogen and the others were maintained in culture for 7 days in RPMI-1640 Glutamax I supplemented with 10 % FBS and 1% antibiotic solution (10,000 U/mL penicillin G, 10,000 U/mL streptomycin sulphate) in cell culture incubator at 37°C, 5% CO2 and 21% O2.
Pulsatile vein culture using an ex vivo vein perfusion system was performed as previously described [24,25,26,27]. Upon harvest, veins were stored at 4 °C in a RPMI-1640 Glutamax medium, supplemented with 12.5% fetal calf serum (Gibco). Within 1 h after the surgery, the segment with an external diameter of 2.5–4 mm were divided in two equal parts. One part was fixed in either formalin for immunohistochemistry or rapidly frozen in liquid nitrogen for molecular analyses. A second part was perfused in the EVPS for 7 days to a pulsatile biphasic flow of 60 pulses/min under either low (LP = 7 mmHg; systolic/diastolic pressure = 8 ± 1/6 ± 1 mmHg) or high perfusion pressure (HP = 100 mmHg; systolic/diastolic pressure = 120 ± 5/90 ± 5 mmHg). Upon completion of the perfusion, the 5 mm proximal and distal ends, which attached the vein to the equipment, were discarded. A central, 5 mm-thick ring was cut from the remaining segment and fixed in formalin for morphometry. The remaining fragments were frozen and reduced into powder for RT-PCR and Western blot analysis. The veins segments in the EVPS were maintained in RPMI-1640, supplemented with Glutamax, 12.5% fetal calf serum (Gibco), 8% 70 kDa dextran (Sigma), and 1% antibiotic-antimycotic solution (10,000 U/ml penicillin G, 10 mg/ml streptomycin sulphate, 25 mg/ml amphotericin B, and 0.5 μg/ml gentamycin). This medium was changed every 2 days. In this study, eight veins obtained from randomly selected patients who underwent lower limb bypass surgery for critical ischemia were used.
2.2. Cell culture
Human VSMCs were prepared from human saphenous vein segments as previously described [28,29]. Vein explants were plated on the dry surface of a cell culture plate coated with 1% Gelatine type B (Sigma-Aldrich). Explants were maintained in RPMI, 10% FBS medium in a cell culture incubator at 37°C, 5% CO2, 5% O2 environment. 9 different veins/patients were used in this study to generate VSMC.
2.3. siRNA-mediated knock-down and adenoviral-mediated overexpression
CSE knockdown was performed using human siRNA targeting CTH (Ambion-Life Technologies, ID: s3710 and s3712). The control siRNA (siCtrl) was the AllStars Negative Control siRNA (Qiagen, SI03650318). VSMC grown at 70% confluence were transfected overnight with 30 nM siRNA using lipofectamin RNAiMax (Invitrogen, 13778-075). After washing, cells were maintained in full media for 48h prior to assessment.
CSE overexpression was achieved using a replication-deficient recombinant adenoviral (DE1/E3) vector. Adenoviral infection was achieved overnight in complete medium using AdCTH (kindly provided by James R. Mitchell [30], produced and purified by Vector Biolabs, Philadelphia, PA, U.S.A.), or the negative control virus Ad-eGFP (Vector Biolabs, Cat. No: 1060). After washing, cells were maintained in full media for 48h prior to assessment.
2.4. Histology
After 7 days in culture, or immediately upon vein isolation (D0), human vessel segments were fixed in buffered formalin, embedded in paraffin, cut into 5 µm sections, and stained with VGEL as previously described [14]. Slides were scanned using a ZEISS Axioscan 7 Microscope Slide Scanner. Polychrome Herovici staining was performed on paraffin sections as described [31]. Young collagen was stained blue, while mature collagen was pink. Cytoplasm was counterstained yellow. Hematoxylin was used to counterstain nuclei blue to black. For intimal and medial thickness, 96 (4 measurements/photos and 4 photos per cross section on six cross sections) measurements were performed [26,27]. Two independent researchers blinded to the conditions did the morphometric measurements using the Olympus Stream Start 2.3 software (Olympus, Switzerland) [13,14,23].
CSE, CBS and 3-MST immunohistochemistry were performed on paraffin sections [32]. After rehydration and antigen retrieval (TRIS-EDTA buffer, pH 9, 1 min in an electric pressure cooker autocuiser Instant Pot duo 60 under high pressure), immunostaining was performed on human vein or artery sections using the EnVision®+ Dual Link System-HRP (DAB+) according to manufacturer’s instructions. Slides were further counterstained with hematoxylin. The positive immunostaining area was quantified using the Fiji (ImageJ 1.53t) software and normalized to the total area of the tissue by two independent observers blinded to the conditions.
2.5. Western blotting
Vessels were flash-frozen in liquid nitrogen, grinded to power and resuspended in SDS lysis buffer (62.5 mM TRIS pH6,8, 5% SDS, 10 mM EDTA). Protein concentration was determined by DC protein assay (Bio-Rad Laboratories, Reinach, Switzerland). 10 to 20 µg of protein were loaded per well. Primary cells were washed once with ice-cold PBS and directly lysed with Laemmli buffer as previously described [14,28]. Lysates were resolved by SDS-PAGE and transferred to a PVDF membrane (Immobilon-P, Millipore AG, Switzerland). Immunoblot analyses were performed as previously described [28] using a CSE antibodies described in Supplementary Table S1. Membranes were revealed by enhanced chemiluminescence (Immobilon, Millipore) using the Azure 280 device (Azure Biosystems) and analyzed using the Fiji (ImageJ 1.53t) software. Protein abundance was normalized to total protein using Pierce™ Reversible Protein Stain Kit for PVDF Membranes (cat 24585; Thermo Fisher Scientific).
2.6. Reverse transcription and quantitative polymerase chain reaction (RT-qPCR)
Flash frozen vessels powder was homogenized in Tripure Isolation Reagent (Roche, Switzerland), and total RNA was extracted according to the manufacturer's instructions. After RNA Reverse transcription (Prime Script RT reagent, Takara), cDNA levels were measured by qPCR Fast SYBR™ Green Master Mix (Ref: 4385618, Applied Biosystems, ThermoFischer Scientific AG, Switzerland) in a Quant Studio 5 Real-Time PCR System (Applied Biosystems, ThermoFischer Scientific AG, Switzerland), using the following primers: MPST Fw: 5’-CCGAGACGGCATTGAACCT-3’; MPST Rv:5’- CCTGGCTCAGGAAGTCTGTG-3’; CTH Fw: 5’-CCAGCACTCGGGTTTTGAAT-3’; CTH Rv: 5’- TACTTAGCCCCATCCAGTGC-3’; CBS Fw: 5’-GCAGATCCAGTACCACAGCA-3’; CBS Rv: 5’-GCCACGAAGTTCAGCAAGTC-3’.
2.7. Lead acetate (CSE activity assay)
Flash frozen vessels powder was homogenized in passive lysis buffer (Promega) and protein content was determined using a BCA protein assay (Thermofischer). Using a 96 well plate, 300mg of proteins were diluted into 100µl of PBS supplemented with 10mM Cysteine and 1mM pyridoxal phosphate as substrate and cofactor for CSE. The plate was then covered with Whatman paper impregnated with 20mM lead acetate and incubated at 37°C for 5h. Lead sulfide precipitate on the Whatman paper were scanned using a high-resolution scanner (HP) and quantified using Fiji (ImageJ 1.53t) software.
2.8. H2S and persulfidation measurement
Free H2S was measured in cells using the SF7-AM fluorescent probe [33] (Sigma-Aldrich). The probe was dissolved in anhydrous DMF at 5 mM and used at 5 μM in serum-free RPMI. Live-cell image acquisition was performed using a Nikon Ti2 spinning disk confocal microscope. Global protein persulfidation was assessed on VSMC grown on glass coverslips as previously described [9]. Cells were incubated for 20 minutes with 1mM 4-Chloro-7-nitrobenzofurazan (NBF-Cl, Sigma-Aldrich) diluted in PBS. Then, cells were washed with PBS and fixed for 10 minutes in ice-cold methanol. Coverslips were rehydrated in PBS and incubated with 1mM NBF-Cl for 1h at 37°C. Cells were further incubated at 37°C for 1h in Daz2-Cy5.5 solution. Finally, coverslips were washed 3 times in methanol and 2 times in PBS, mounted in Vectashield mounting medium with DAPI, and visualized with a 90i Nikon fluorescence microscope. Persulfidation was measured as the ratio of Daz2-Cy5.5 over NBF-Cl signal per cell by two independent experimenter blinded to the conditions using the Fiji (ImageJ 1.53t) software.
2.9. BrdU assay
VSMCs were grown at 80% confluence (5·103 cells per well) on glass coverslips in a 24-well plate and starved overnight in serum-free medium. Then, VSMC were either treated or not (ctrl) with the drug of choice for 24 hours in full medium (RPMI 10% FBS) in presence of 10µM BrdU. All conditions were tested in parallel. All cells were fixed in ice-cold methanol 100% after 24 hours of incubation and immunostained for BrdU. Images were acquired using a Nikon Eclipse 90i microscope. BrdU-positive nuclei and total DAPI-positive nuclei were automatically detected using the Fiji (ImageJ 1.53t) software [29].
2.10. Wound healing assay
VSMCs were grown at confluence (104 cells per well) in a 12-well plate and starved overnight in serum-free medium. Then, a scratch wound was created using a sterile p200 pipette tip and medium was changed to full medium (RPMI 10% FBS) in presence of 0.5µg/mL mitomycin C to block proliferation. Repopulation of the wounded areas was recorded by phase-contrast microscopy in a Nikon Ti2-E live cell microscope. All conditions were tested in parallel. The area of the denuded area was measured automatically using the macro Wound_healing_size_tool_updated.ijm [34] in the Fiji (ImageJ 1.53t) software. Data were expressed as a percentage of the wound closure. Morphometric measurement of cell geometry was performed manually by two independent experimenters blinded to the conditions using the shape descriptors in measurements in Fiji (ImageJ 1.53t) software
2.11. Statistical analyses
All experiments adhered to the ARRIVE guidelines and followed strict randomization. All experiments and data analysis were conducted in a blind manner using coded tags rather than actual group name. All experiments were analyzed using GraphPad Prism 9. Normal distribution of the data was assessed using Kolmogorov-Smirnov tests. All data with normal distribution were analyzed by unpaired bilateral Student’s t-tests or Mixed-effects model (REML) followed by post-hoc t-tests with the appropriate correction for multiple comparisons when comparing more than 2 groups. For non-normal distributed data, Kruskal-Wallis non-parametric ranking tests were performed, followed by Dunn's multiple comparisons test to calculate adjusted p values. Unless otherwise specified, p-values are reported according to the APA 7th edition and New England Journal of Medicine statistical guidelines. *p<.033, **p<.002, ***p<0.001.
2.12. Ethics Statement
Human vein and artery segments were obtained from donors who underwent vascular surgery at the Lausanne University Hospital. Written, informed consent was obtained from all donors. The study protocols for organ collection and use were reviewed and approved by the Lausanne University Hospital (CHUV) and the Cantonal Human Research Ethics Committee (http://www.cer-vd.ch/, no IRB number, Protocol Number 170/02), and are in accordance with the principles outlined in the Declaration of Helsinki of 1975, as revised in 1983 for the use of human tissues.
3. Results
3.1. CSE is expressed in the media and intima of human saphenous vein and artery segments
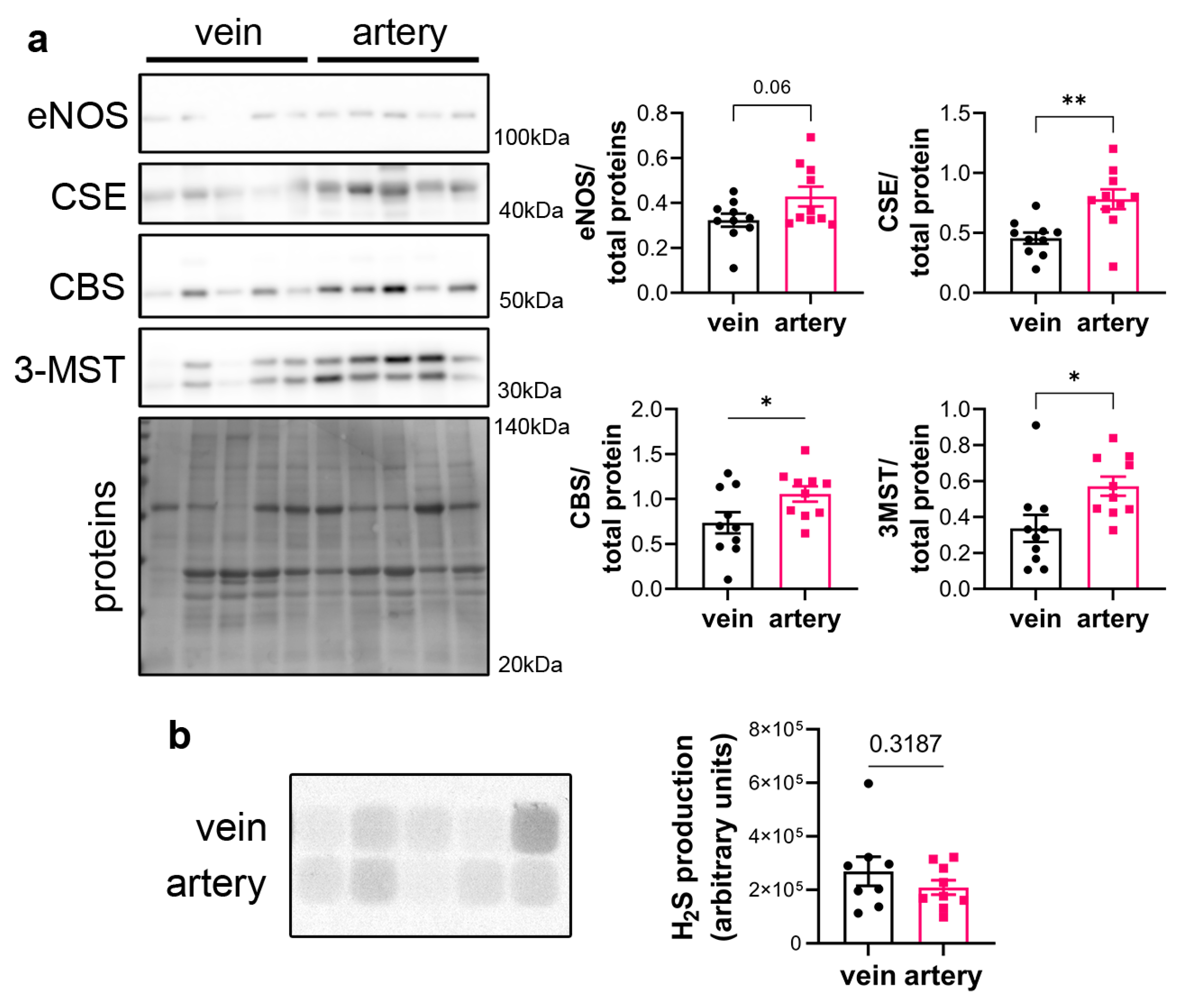
CSE protein was assessed in segments of healthy human vein and artery. Artery segments expressed higher levels of CSE, eNOS, CBS and 3MST than vein segments as measured by WB analysis (Figure 1a). However, this did not translate in higher H2S production capacity as assessed by led acetate assay (Figure 1b).
Figure 1.
CSE is expressed in human artery and saphenous vein segments. a) Representative Western blot and quantitative assessment of eNOS, CSE, CBS and 3-MST from freshly isolated human arteries and saphenous veins. Data are mean±SEM of 10 arteries and 9 veins. b) Lead acetate assay in freshly isolated human arteries and saphenous veins. Data are mean±SEM of 9 arteries and 8 veins. *p<.033, **p<.002, as determined by bilateral paired t-test.
Figure 1.
CSE is expressed in human artery and saphenous vein segments. a) Representative Western blot and quantitative assessment of eNOS, CSE, CBS and 3-MST from freshly isolated human arteries and saphenous veins. Data are mean±SEM of 10 arteries and 9 veins. b) Lead acetate assay in freshly isolated human arteries and saphenous veins. Data are mean±SEM of 9 arteries and 8 veins. *p<.033, **p<.002, as determined by bilateral paired t-test.
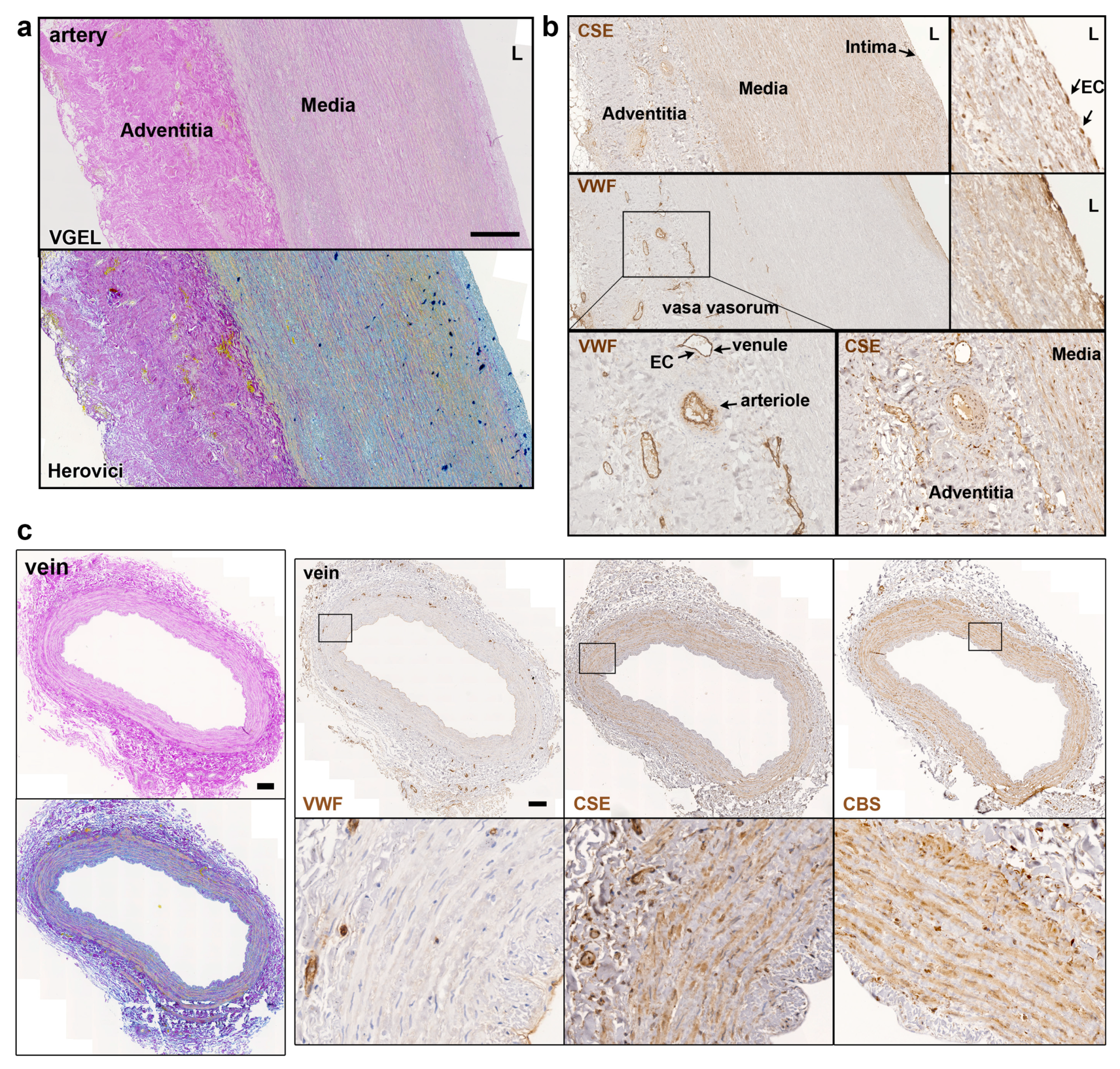
Further CSE immunohistology show that CSE was expressed both in the media and intima layers of the artery segments (Figure 2a, b). CSE was also expressed in the endothelium of the small vessels of the vasa vasorum of arteries, easily identified using a Von Willebrand factor (VWF) immunostaining of EC (Figure 2a, b). In human veins, CSE was mostly detectable in the media layer, but not so much in the endothelium (Figure 2c). CSE was also expressed in the endothelium of the small vessels of the vasa vasorum (Figure 2c). In veins, CBS was expressed mostly in the media layer (Figure 2c). In artery segments, 3-MST was detectable in the media, but was mostly expressed by the EC of the intima and smaller vessels of the vasa vasorum. Interestingly, some arteries featured benign intimal hyperplasia and 3-MST was not expressed in this neointima layer (Figure S1). In contrast, CSE was similarly expressed throughout the media and neointima layers. In the saphenous vein, 3MST was not expressed in the media layer, mainly detected in the vasa vasorum vessels of the media and adventitia layers, and seldom detected in the EC from the intima (Figure S1).
Figure 2.
CSE is expressed in the media and intima of human artery and saphenous vein. Representative, VGEL, Herovici, CSE, CBS and VWF staining as indicated in section of a human artery (a, b) and a saphenous vein (c). Images are representative of 8 arteries and 10 saphenous veins. Scale bar=100µm.
Figure 2.
CSE is expressed in the media and intima of human artery and saphenous vein. Representative, VGEL, Herovici, CSE, CBS and VWF staining as indicated in section of a human artery (a, b) and a saphenous vein (c). Images are representative of 8 arteries and 10 saphenous veins. Scale bar=100µm.
3.2. CSE expression is regulated by flow
To study the regulation of CSE expression in response to flow, we next investigated CSE expression in a model of static human vein culture. Static vein culture leads to formation of IH as previously described [9,13,14], as assessed by VGEL staining (Figure 3a). qPCR analysis revealed that CSE mRNA expression was increased 4-fold after 7 days in static culture (Figure 3b). CBS mRNA expression was also increased about 4-fold, while MPST mRNA expression increased about 2-fold (Figure 3b). WB analysis confirmed CSE 4-fold protein overexpression in static culture, whereas CBS protein levels were decreased by about 30%, and 3-MST was increased about 1.5-fold (Figure 3c). The increase in CSE expression was accompanied by an increase in H2S production, as measured by lead acetate (Figure S2). IHC analyses confirmed that CSE expression was increased in the media layer of human veins after static culture (Figure 3d). Of note, CSE expression seemed reduced in the endothelium after 7 days in static culture (Figure 3d). CBS expression was reduced and restricted to fewer cells expressing higher CBS levels in the media and CBS expression was not detectable in the endothelium after 7 days in static culture (Figure 3d). 3-MST expression was lost in the EC of the intima, still detectable in the EC of the vasa vasorum, as well as in the media layer (Figure S3a).
To selectively evaluate the role of pressure and shear stress on CSE expression, we further assessed CSE, 3-MST and CBS expression in human saphenous veins segments perfused under pulsatile low pressure (LP; venous regimen mean pressure=7mmHg) or high pressure (HP; arterial regimen mean pressure=100mHg). LP perfusion did not induce the development of IH (data not shown) as previously described [26,27], while arterial (HP) perfusion stimulated thinning of the media layer and the formation of IH, as assessed by VGEL and Herovici staining (Figure 4a). After 7 days in LP conditions, CTH and CBS mRNA expression remained unchanged, while MPST expression was reduced 4-fold (Figure 4b). After 7 days in HP condition, MPST was further reduced and CTH expression was reduced 3-fold, while CBS mRNA expression increased 2-fold (Figure 4c). We further looked at CSE and CBS protein expression in human vein segments perfused at high pressure (HP). Western blot analysis revealed that both CSE and CBS protein levels were reduced in those conditions compared to the native vein (Figure 4d). IHC staining confirmed that CSE expression was decreased by HP throughout the media and intima layers (Figure 4e). Similar to what we observed in static vein culture (Figure 3d), the CBS protein expression was reduced and restricted to fewer cells expressing higher CBS levels in the media. CBS was not detectable in the endothelium (Figure 4e). In contrast with static vein culture, 3-MST expression remained undetectable in the media layer. 3-MST was still detectable in the EC of the small vessels of the vasa vasorum (Figure S3b).
3.3. CSE regulates human VSMC migration
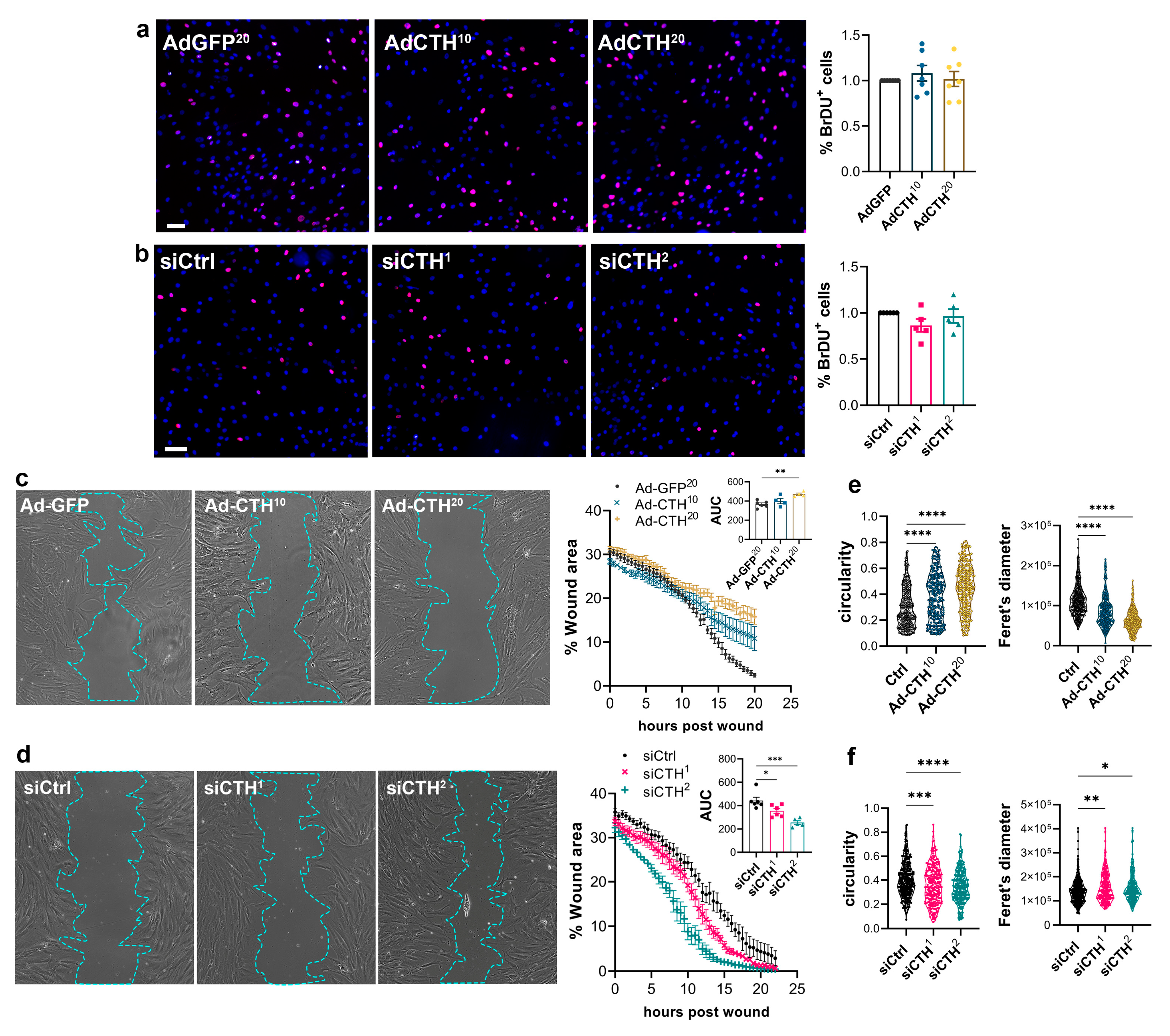
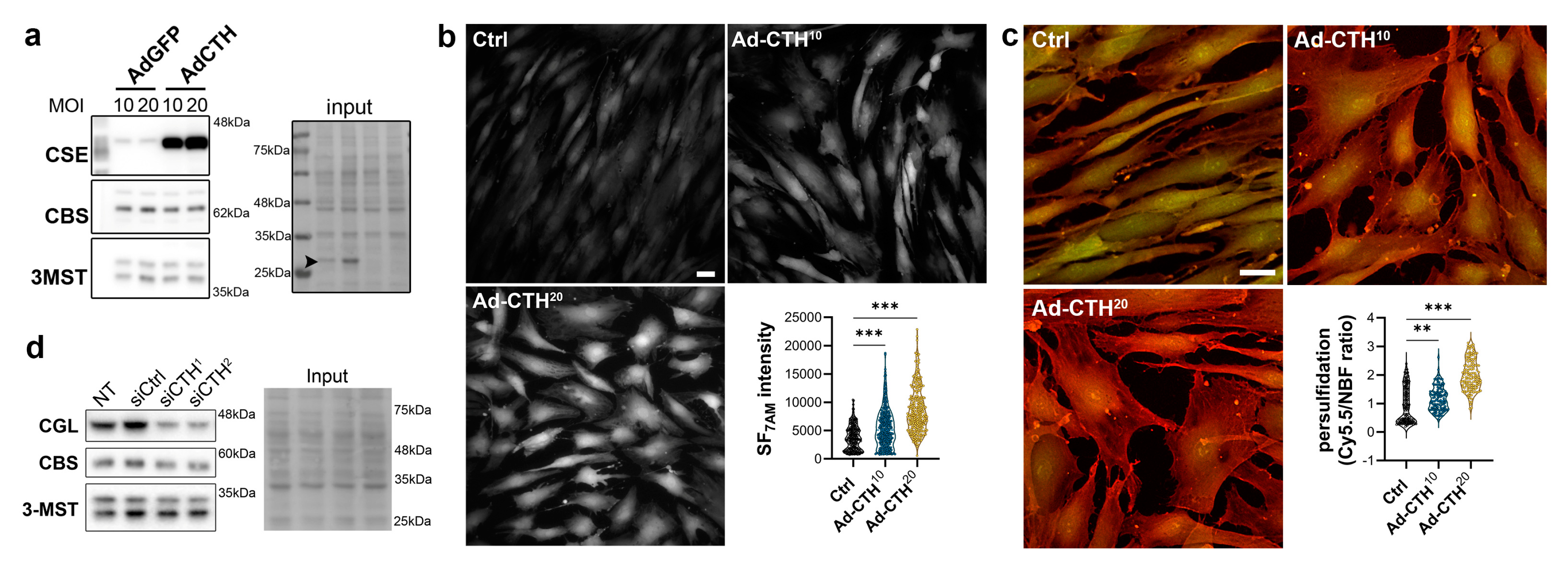
CSE expression in the media of large human vein was decreased under pulsatile arterial high pressure (Figure 3 and Figure 4). We then tested the role of CSE in VSMC proliferation and migration in primary human VSMC derived from great saphenous vein segments. Adenoviral-mediated CSE overexpression increased CSE protein expression in primary VSMC but did no impact CBS and 3MST expression (Figure 5a). As expected, CSE overexpression increased H2S production (SF7AM; Figure 5b) and protein persulfidation (Figure 5c). In contrast, siRNA-mediated CSE silencing reduced CSE protein level by 60-70% (siCTH1=0.37±0.09, p=.008; siCTH2=0.45±0.05, p=.02 by mixed effect model with Šídák's multiple comparisons test), without impacting CBS or 3MST protein levels (Figure 5d).
CSE overexpression or knock-down had no impact on VSMC proliferation as assessed by BrdU incorporation (Figure 6a-b). In contrast, CSE overexpression slowed-down migration in a wound healing assay (Figure 6c), whereas CSE knock-down accelerated wound healing (Figure 6d). Of note, CSE overexpression also increased the circularity and reduced the feret diameter of the VSMC, whereas CSE knock-down reduced circularity and increased the feret diameter, indicating that CSE modulation had an impact on the geometry of the cells associated with migration (Figure 6 e, f). CSE overexpression caused the most prominent changes, characterized by reduced spindle shape morphology due to a more static phenotype. In contrast, CSE knocked-down cells were more mobile and more elongated (decreased circularity and increased feret diameter).
Figure 6.
CSE inhibits VSMC migration, but not proliferation in vitro. a) VSMC proliferation (BrdU incorporation) in VSMC infected with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. b) VSMC proliferation (BrdU incorporation) in VSMC transfected with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH 1 and 2), as indicated. a-b) Data are % of BrdU positive nuclei (pink) over DAPI positive nuclei. Scale bar: 50 µm. Data shown as mean ± SEM of 5 to 6 independent experiments. No statistical differences as determined by repeated measures one-way ANOVA with Dunnett's multiple comparisons tests. c) VSMC migration (wound healing) in VSMC infected with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. d) VSMC migration (wound healing) in VSMC transfected with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH 1 and 2), as indicated. c-d) Bright field images of VSMC 12 h post wound. Scale bar: 100 µm. Data are mean ± SEM of the percentage of wound closure in 5 to 6 independent experiments. Insets show area under the curve (AUC) of wound healing. *p<.033, **p<.002, ***p<.001, report adjusted p-values as determined by one-way ANOVA followed by Dunnett's multiple comparisons tests. e-f) Cell morphology during wound healing is expressed as the circularity index and the Feret’s diameter shown as violin plots of individual cells across 5 independent experiments. *p<.033, **p<.002, ***p<.001 report adjusted p-values as determined by Kruskal-Wallis non-parametric ranking followed by Dunn's multiple comparisons tests.
Figure 6.
CSE inhibits VSMC migration, but not proliferation in vitro. a) VSMC proliferation (BrdU incorporation) in VSMC infected with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. b) VSMC proliferation (BrdU incorporation) in VSMC transfected with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH 1 and 2), as indicated. a-b) Data are % of BrdU positive nuclei (pink) over DAPI positive nuclei. Scale bar: 50 µm. Data shown as mean ± SEM of 5 to 6 independent experiments. No statistical differences as determined by repeated measures one-way ANOVA with Dunnett's multiple comparisons tests. c) VSMC migration (wound healing) in VSMC infected with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. d) VSMC migration (wound healing) in VSMC transfected with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH 1 and 2), as indicated. c-d) Bright field images of VSMC 12 h post wound. Scale bar: 100 µm. Data are mean ± SEM of the percentage of wound closure in 5 to 6 independent experiments. Insets show area under the curve (AUC) of wound healing. *p<.033, **p<.002, ***p<.001, report adjusted p-values as determined by one-way ANOVA followed by Dunnett's multiple comparisons tests. e-f) Cell morphology during wound healing is expressed as the circularity index and the Feret’s diameter shown as violin plots of individual cells across 5 independent experiments. *p<.033, **p<.002, ***p<.001 report adjusted p-values as determined by Kruskal-Wallis non-parametric ranking followed by Dunn's multiple comparisons tests.
4. Discussion
Endogenous H2S production in mammals results from the oxidation of the sulfur-containing amino acids cysteine and homocysteine via the reverse ''trans-sulfuration'' pathway mainly via CSE and CBS, and 3-MST. Although the enzymes and pathways responsible for H2S production are well described, little is known about their regulation in pathophysiologic conditions. In this study, we investigated the expression and regulation of CSE, CBS and 3-MST in segments of human saphenous vein and artery. CBS was mainly detected in the media layer of human vessels. In contrast, 3-MST was mainly detected in the endothelium and in the EC of small vessels of the vasa vasorum in both arteries and veins. This suggests a more prominent role of 3-MST in EC of small caliber vessels and capillaries, which is consistent with previous studies suggesting a key role for 3-MST in EC [20]. 3-MST is also expressed in the media of arteries, but not the media of veins, indicating differential regulation of 3-MST in arterial and venous vessels. Rodent studies suggest that CSE is mainly expressed in EC in the cardiovascular system and that the endothelium is the main source of H2S in blood vessels [15,16,17,18,37]. That said, CSE has also been described in VSMC and proposed to be a functional, albeit minor source of H2S [8,19,38]. Here, CSE was expressed in the endothelium of large vessels, and in small vessels of the vasa vasorum in native human artery and vein. CSE was also abundant in the media layer.
We then investigated the regulation of the three enzymes in vein segments placed in ex vivo culture. CBS protein expression was reduced in vein segments placed in pathological culture conditions ex vivo, both in the absence of flow and in high pressure flow. This indicates that shear stress does not regulate CBS expression in VSMC. Interestingly, while CBS protein levels were lower, CBS mRNA levels were increased in ex vivo culture, suggesting a differential regulation of mRNA and protein expression and a possible effect of ex vivo culture on protein stability. However, histological analysis suggested a more complex regulation, as CBS was no longer detected in most cells, while it was overexpressed in a few cells present in the media and neointimal layer. Further studies are required to determine which cells overexpress CBS when most cells in the media and neointima appear to down-regulate CBS expression. This is of particular interest as adult VSMC are highly plastic cells [39], and the switch from a quiescent 'contractile' phenotype to a proliferative 'synthetic' phenotype plays a major role in the context of IH [40]. Recent VSMC lineage tracing studies in mice using in vivo cell fate tracing with SMC-specific genetic reporter tools suggest that a small subset of VSMCs expand after injury to form clonal patches of neointimal cells [41,42,43]. Further studies are required to elucidate the identity of this small subset of VSMCs in human tissue, and whether CBS is expressed or not in this subset in the context of IH.
3-MST mRNA and protein expression were severely reduced by ex vivo perfusion of vein segments, independent of flow and shear stress. This is not surprising as our model of ex vivo culture results in endothelial dysfunction and rapid loss of endothelial-specific markers [26,27,44]. However, static ex vivo culture tended to stimulate 3-MST mRNA and protein expression despite severe endothelial dysfunction. This suggests that 3-MST may be negatively regulated by shear stress in a similar way to CSE. 3-MST was largely undetectable in the media layer of veins, but it was detectable in the media of arteries, suggesting that 3-MST plays a role in arterial VSMC, but that high shear stress per se does not negatively regulate 3-MST expression. 3-MST was overexpressed in VSMC in veins in static condition and in cultured VSMC in vitro, suggesting that 3-MST could be involved in VSMC reprogramming in the context of IH. However, arterial perfusion prevented 3-MST expression, so it is unlikely that 3-MST play a main role in IH in vivo. In a recent study, 3-MST was found to be expressed in VSMC and cardiomyocytes, and 3-Mst-/- mice were protected against myocardial ischemia-reperfusion injury [45]. Further studies are required to better characterize the role and regulation of 3-MST in VSMC.
About CSE, our data confirm that CSE is negatively regulated by shear stress. Using our model of ex vivo vein perfusion, we observed that high pressure inhibits CSE expression both in the media and in the endothelium. In contrast, static ex vivo vein culture stimulates CSE expression. This is in line with previous evidence showing that CSE expression is downregulated by high shear stress and is predominantly found in regions of disturbed flow.[21,22]. However, we did not observe higher levels of CSE in native human vein segments compared to aortic segments, despite the high shear stress in arteries. In fact, CSE, CBS, and 3MST levels were higher in arteries than in veins. However, this could be due to a higher cell content in arterial tissue compared to venous tissue, which contains more connective tissue than arteries. Consistent with this hypothesis, higher CSE levels did not translate into higher CSE activity as measured by the lead acetate assay. Interestingly, this regulation by shear stress is not unique to EC and was also observed in VSMC. Further studies are needed to better understand the regulation of CSE expression in vein and artery, specifically in VSMC and EC.
Given that CSE is highly expressed in VSMC and downregulated by high-pressure perfusion in veins, we further investigated the role of CSE in human primary venous VSMC. We document that CSE is a specific modulator of VSMC function independent of EC or EC-derived H2S production, and that CSE controls the migration of primary human VSMC. This is in agreement with studies showing that VSMCs isolated from Cse-/- mice are more motile than their WT counterparts, and that blocking Cse activity with PAG in VSMC increases cell migration [8,19]. We and others reported that H2S donors inhibits VSMC proliferation [12,14,36]. Here, we did not observe a significant effect of CSE on cell proliferation. This contrasts with a previous study showing that Cse overexpression decreased proliferation and even induced VSMC apoptosis. This discrepancy probably results from the level of Cse overexpression, which was much higher in the previous study by Yang et al. [36]. In this study, the authors used arterial VSMC, whereas we used venous VSMC. Venous and arterial VSMC are functionally different so the venous versus arterial origin of VSMC may also contributes to the discrepancies in our results. Nevertheless, it would not be surprising if higher CSE expression and activity, leading to greater H2S production, resulted in cell apoptosis. Indeed, elevated levels of H2S are known to induce cell cycle arrest and apoptosis [46]. Note that evidence suggests that the threshold for the pro-apoptotic effect of CSE and H2S may differ depending on the cell type [8,19], which may reflect cell specific sulfur metabolism and H2S detoxifying capacities.
The mechanisms whereby H2S affect VSMC proliferation and/or migration are not fully understood. Cytotoxic CSE overexpression or exogenous H2S supplementation induces VSMC cycle arrest and apoptosis by stimulating ERK1/2, p38 MAPK, and p21 Cip [36]. Exogenous H2S donor treatment in VSMC has also been shown to inhibit the MAPK pathway, especially ERK1,2, and the mTOR pathways, which correlates with reduced VSMC proliferation and migration [13]. Here, no effects on ERK and mTOR signaling were observed upon CSE manipulation. This probably reflects the more subtle CSE variation in our experimental design compared with exogenous H2S supply or high Cse overexpression in previous studies. Rather than a physiological response to Cse-derived H2S, the effect of Cse overexpression on VSMC proliferation may be due to cell cycle arrest associated with a cytotoxic effect of H2S. In accordance with this hypothesis, it was previously shown that inhibition of ERK did not prevent the effect of NaHS on VSMC migration [8], while ERK inhibition was instrumental in the effect of Cse and H2S on VSMC proliferation [36]. Of note, several studies in EC also reported that Cse regulates cell migration, but not proliferation [30,47]. We propose that low levels of H2S affect VSMC migration without affecting their proliferation. Our main finding is that CSE variations reshape VSMC, suggesting an effect on the cytoskeleton and interaction with the ECM. This is consistent with our previous finding that H2S donors inhibit microtubule polymerization in VSMC [9], and the findings that CSE deficiency in mouse VSMC results in increased expressions of β1-integrin and increased migration [8]. In EC, it was recently shown that integrins are extensively sulfhydrated, and that β3 integrin S-sulfhydration promotes adhesion and is required for EC alignment with flow [48]. In this study, they further demonstrated that Cse deficiency in EC leads to overactivation of RhoA, a major hub regulating cell migration and adhesion. Interestingly, β3-integrin is also expressed in VMSC and β3-integrin signaling is instrumental for enhanced VSMC proliferation and migration in vascular disease [49,50]. We hypothesize that CSE-mediated integrin sulfhydration promotes VSMC adhesion, thereby limiting migration. Further studies are needed to determine the exact role and balance between β1- and β3-integrins and RhoA signaling in the regulation of cell migration by CSE in VSMC.
The fact that high pressure inhibits CSE expression in vein segments suggests that CSE downregulation occurs in vivo in arterio-venous bypass grafts, which contributes to VSMC migration and graft IH.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1: Antibodies, Supplementary Table S2: reagents Figure S1: 3MST is mainly expressed in endothelial cells in large artery and vein segments, Figure S2. Static vein culture promotes CSE-derived H2S production, Figure S3. Static vein culture promotes 3-MST expression in the media of saphenous veins, Figure S4. CSE modulation does not regulate P-ERK or P-S6RP levels.
Author Contributions
Conceptualization, F.A. and S.D.; methodology, F.A., S.D, M.L. and S.U.; validation, F.A., S.D.; formal analysis, S.Z., F.A., S.U. and M.L; investigation, S.Z., C.D., S.U. and M.L.; writing—original draft preparation, S.Z. and F.A.; writing—review and editing, F.A., C.D., S.Z., M.L. and S.D.; visualization, F.A.; supervision, F.A. and S.D.; project administration, F.A. and S.D.; funding acquisition, F.A. and S.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Swiss National Science Foundation to F.A and S.D., grant number FN-310030_176158, the Novartis Foundation to FA, the Union des Sociétés Suisses des Maladies Vasculaires to SD, and the Fondation pour la recherche en chirurgie thoracique et vasculaire to FA and SD. The APC was funded by the Swiss National Science Foundation.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, reviewed and approved by the Lausanne University hospital (CHUV) and the Cantonal Human Research Ethics Committee (http://www.cer-vd.ch/, no IRB number, Protocol Number 170/02).
Informed Consent Statement
Human vessels were obtained from donors who underwent surgery in the Lausanne University Hospital. Written, informed consent was obtained from all donors.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the Mouse Pathology Facility for their services in histology (https://www.unil.ch/mpf) and the Cellular Imaging Facility for their services in microscopy (https://cif.unil.ch).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Déglise S, Bechelli C, Allagnat F. Vascular Smooth Muscle Cells in Intimal Hyperplasia, an Update. Frontiers in Physiology (2023) 13. [CrossRef]
- Simpson EL, Kearns B, Stevenson MD, Cantrell AJ, Littlewood C, Michaels JA. Enhancements to Angioplasty for Peripheral Arterial Occlusive Disease: Systematic Review, Cost-Effectiveness Assessment and Expected Value of Information Analysis. Health Technol Assess (2014) 18(10):1-252. [CrossRef]
- Cirino G, Szabo C, Papapetropoulos A. Physiological Roles of Hydrogen Sulfide in Mammalian Cells, Tissues and Organs. Physiol Rev (2022). Epub 2022/04/19. [CrossRef]
- Jain SK BR, Rains JL, Bass PF, Levine SN, Reddy S, McVie R, Bocchini JA. Low Levels of Hydrogen Sulfide in the Blood of Diabetes Patients and Streptozotocin-Treated Rats Causes Vascular Inflammation? Antioxid Redox Signal (2010) 12:1333-7. [CrossRef]
- Islam KN, Polhemus DJ, Donnarumma E, Brewster LP, Lefer DJ. Hydrogen Sulfide Levels and Nuclear Factor-Erythroid 2-Related Factor 2 (Nrf2) Activity Are Attenuated in the Setting of Critical Limb Ischemia (Cli). J Am Heart Assoc (2015) 4(5). Epub 2015/05/16. 2015. [CrossRef]
- Beard RS, Jr., Bearden SE. Vascular Complications of Cystathionine Beta-Synthase Deficiency: Future Directions for Homocysteine-to-Hydrogen Sulfide Research. Am J Physiol Heart Circ Physiol (2011) 300(1):H13-26. [CrossRef]
- Longchamp A, MacArthur MR, Trocha K, Ganahl J, Mann CG, Kip P, et al. Plasma Hydrogen Sulfide Is Positively Associated with Post-Operative Survival in Patients Undergoing Surgical Revascularization. Front Cardiovasc Med (2021) 8:750926. Epub 2021/11/12. [CrossRef]
- Yang G, Li H, Tang G, Wu L, Zhao K, Cao Q, et al. Increased Neointimal Formation in Cystathionine Gamma-Lyase Deficient Mice: Role of Hydrogen Sulfide in Alpha5beta1-Integrin and Matrix Metalloproteinase-2 Expression in Smooth Muscle Cells. J Mol Cell Cardiol (2012) 52(3):677-88. [CrossRef]
- Macabrey D, Longchamp A, MacArthur MR, Lambelet M, Urfer S, Deglise S, et al. Sodium Thiosulfate Acts as a Hydrogen Sulfide Mimetic to Prevent Intimal Hyperplasia Via Inhibition of Tubulin Polymerisation. EBioMedicine (2022) 78:103954. Epub 20220322. [CrossRef]
- Trocha KM, Kip P, Tao M, MacArthur MR, Trevino-Villarreal JH, Longchamp A, et al. Short-Term Preoperative Protein Restriction Attenuates Vein Graft Disease Via Induction of Cystathionine Gamma-Lyase. Cardiovasc Res (2020) 116(2):416-28. Epub 2019/03/30. [CrossRef]
- Meng QH, Yang G, Yang W, Jiang B, Wu L, Wang R. Protective Effect of Hydrogen Sulfide on Balloon Injury-Induced Neointima Hyperplasia in Rat Carotid Arteries. Am J Pathol (2007) 170(4):1406-14. [CrossRef]
- Ma B, Liang G, Zhang F, Chen Y, Zhang H. Effect of Hydrogen Sulfide on Restenosis of Peripheral Arteries after Angioplasty. Mol Med Rep (2012) 5(6):1497-502. Epub 2012/04/04. [CrossRef]
- Macabrey D, Deslarzes-Dubuis C, Longchamp A, Lambelet M, Ozaki CK, Corpataux JM, et al. Hydrogen Sulphide Release Via the Angiotensin Converting Enzyme Inhibitor Zofenopril Prevents Intimal Hyperplasia in Human Vein Segments and in a Mouse Model of Carotid Artery Stenosis. Eur J Vasc Endovasc Surg (2022) 63(2):336-46. Epub 20211213. [CrossRef]
- Longchamp A, Kaur K, Macabrey D, Dubuis C, Corpataux JM, Deglise S, et al. Hydrogen Sulfide-Releasing Peptide Hydrogel Limits the Development of Intimal Hyperplasia in Human Vein Segments. Acta Biomater (2019) 97:374-84. Epub 20190726. [CrossRef]
- Yang G, Wu L, Jiang B, Yang W, Qi J, Cao K, et al. H2s as a Physiologic Vasorelaxant: Hypertension in Mice with Deletion of Cystathionine G-Lyase. Science (2008) 322:587-90. [CrossRef]
- Cheng Y, Ndisang JF, Tang G, Cao K, Wang R. Hydrogen Sulfide-Induced Relaxation of Resistance Mesenteric Artery Beds of Rats. Am J Physiol Heart Circ Physiol (2004) 287(5):H2316-23. Epub 2004/06/12. [CrossRef]
- Mustafa AK, Sikka G, Gazi SK, Steppan J, Jung SM, Bhunia AK, et al. Hydrogen Sulfide as Endothelium-Derived Hyperpolarizing Factor Sulfhydrates Potassium Channels. Circ Res (2011) 109(11):1259-68. [CrossRef]
- Kiesworo K, MacArthur MR, Kip P, Agius T, Macabrey D, Lambelet M, et al. Cystathionine-Gamma-Lyase Overexpression Modulates Oxidized Nicotinamide Adenine Dinucleotide Biosynthesis and Enhances Neovascularization. JVS Vasc Sci (2023) 4:100095. Epub 20230113. [CrossRef]
- Yang G, Wu L, Bryan S, Khaper N, Mani S, Wang R. Cystathionine Gamma-Lyase Deficiency and Overproliferation of Smooth Muscle Cells. Cardiovasc Res (2010) 86(3):487-95. [CrossRef]
- Shibuya N, Mikami Y, Kimura Y, Nagahara N, Kimura H. Vascular Endothelium Expresses 3-Mercaptopyruvate Sulfurtransferase and Produces Hydrogen Sulfide. J Biochem (2009) 146(5):623-6. Epub 2009/07/17. [CrossRef]
- Bibli SI, Hu J, Leisegang MS, Wittig J, Zukunft S, Kapasakalidi A, et al. Shear Stress Regulates Cystathionine Gamma Lyase Expression to Preserve Endothelial Redox Balance and Reduce Membrane Lipid Peroxidation. Redox Biol (2020) 28:101379. Epub 20191113. [CrossRef]
- Yuan S, Yurdagul A, Jr., Peretik JM, Alfaidi M, Al Yafeai Z, Pardue S, et al. Cystathionine Gamma-Lyase Modulates Flow-Dependent Vascular Remodeling. Arterioscler Thromb Vasc Biol (2018) 38(9):2126-36. Epub 2018/07/14. [CrossRef]
- Macabrey D, Joniova J, Gasser Q, Bechelli C, Longchamp A, Urfer S, et al. Sodium Thiosulfate, a Source of Hydrogen Sulfide, Stimulates Endothelial Cell Proliferation and Neovascularization. Front Cardiovasc Med (2022) 9:965965. Epub 20221003. [CrossRef]
- Saucy F, Probst H, Alonso F, Berard X, Deglise S, Dunoyer-Geindre S, et al. Ex Vivo Pulsatile Perfusion of Human Saphenous Veins Induces Intimal Hyperplasia and Increased Levels of the Plasminogen Activator Inhibitor 1. European surgical research Europaische chirurgische Forschung Recherches chirurgicales europeennes (2010) 45(1):50-9. [CrossRef]
- Berard X, Deglise S, Alonso F, Saucy F, Meda P, Bordenave L, et al. Role of Hemodynamic Forces in the Ex Vivo Arterialization of Human Saphenous Veins. J Vasc Surg (2013) 57(5):1371-82. [CrossRef]
- Longchamp A, Allagnat F, Berard X, Alonso F, Haefliger JA, Deglise S, et al. Procedure for Human Saphenous Veins Ex Vivo Perfusion and External Reinforcement. J Vis Exp (2014) (92):e52079. Epub 20141001. [CrossRef]
- Longchamp A, Alonso F, Dubuis C, Allagnat F, Berard X, Meda P, et al. The Use of External Mesh Reinforcement to Reduce Intimal Hyperplasia and Preserve the Structure of Human Saphenous Veins. Biomaterials (2014) 35(9):2588-99. Epub 20140113. [CrossRef]
- Allagnat F, Dubuis C, Lambelet M, Le Gal L, Alonso F, Corpataux JM, et al. Connexin37 Reduces Smooth Muscle Cell Proliferation and Intimal Hyperplasia in a Mouse Model of Carotid Artery Ligation. Cardiovasc Res (2017) 113(7):805-16. [CrossRef]
- Longchamp A, Kaur K, Macabrey D, Dubuis C, Corpataux JM, Deglise S, et al. Hydrogen Sulfide-Releasing Peptide Hydrogel Limits the Development of Intimal Hyperplasia in Human Vein Segments. Acta Biomater (2019). Epub 2019/07/29. [CrossRef]
- Longchamp A, Mirabella T, Arduini A, MacArthur MR, Das A, Trevino-Villarreal JH, et al. Amino Acid Restriction Triggers Angiogenesis Via Gcn2/Atf4 Regulation of Vegf and H2s Production. Cell (2018) 173(1):117-29 e14. Epub 2018/03/24. [CrossRef]
- Teuscher AC, Statzer C, Pantasis S, Bordoli MR, Ewald CY. Assessing Collagen Deposition During Aging in Mammalian Tissue and in Caenorhabditis Elegans. Methods Mol Biol (2019) 1944:169-88. Epub 2019/03/07. [CrossRef]
- Allagnat F, Haefliger JA, Lambelet M, Longchamp A, Berard X, Mazzolai L, et al. Nitric Oxide Deficit Drives Intimal Hyperplasia in Mouse Models of Hypertension. Eur J Vasc Endovasc Surg (2016) 51(5):733-42. Epub 20160319. [CrossRef]
- Lin VS, Lippert AR, Chang CJ. Cell-Trappable Fluorescent Probes for Endogenous Hydrogen Sulfide Signaling and Imaging H2o2-Dependent H2s Production. Proc Natl Acad Sci U S A (2013) 110(18):7131-5. Epub 2013/04/17. [CrossRef]
- Suarez-Arnedo A, Torres Figueroa F, Clavijo C, Arbelaez P, Cruz JC, Munoz-Camargo C. An Image J Plugin for the High Throughput Image Analysis of in Vitro Scratch Wound Healing Assays. PLoS One (2020) 15(7):e0232565. Epub 20200728. [CrossRef]
- Dubuis C, May L, Alonso F, Luca L, Mylonaki I, Meda P, et al. Atorvastatin-Loaded Hydrogel Affects the Smooth Muscle Cells of Human Veins. The Journal of pharmacology and experimental therapeutics (2013) 347(3):574-81. Epub 2013/09/28. [CrossRef]
- Yang G, Wu L, Wang R. Pro-Apoptotic Effect of Endogenous H2s on Human Aorta Smooth Muscle Cells. FASEB J (2006) 20(3):553-5. [CrossRef]
- Bibli SI, Hu J, Sigala F, Wittig I, Heidler J, Zukunft S, et al. Cystathionine Gamma Lyase Sulfhydrates the Rna Binding Protein Human Antigen R to Preserve Endothelial Cell Function and Delay Atherogenesis. Circulation (2019) 139(1):101-14. Epub 2018/07/05. [CrossRef]
- Bucci M, Papapetropoulos A, Vellecco V, Zhou Z, Zaid A, Giannogonas P, et al. Cgmp-Dependent Protein Kinase Contributes to Hydrogen Sulfide-Stimulated Vasorelaxation. PLoS One (2012) 7(12):e53319. [CrossRef]
- Owens GK, Kumar MS, Wamhoff BR. Molecular Regulation of Vascular Smooth Muscle Cell Differentiation in Development and Disease. Physiol Rev (2004) 84(3):767-801. Epub 2004/07/23. [CrossRef]
- Chakraborty R, Chatterjee P, Dave JM, Ostriker AC, Greif DM, Rzucidlo EM, et al. Targeting Smooth Muscle Cell Phenotypic Switching in Vascular Disease. JVS Vasc Sci (2021) 2:79-94. Epub 2021/10/08. [CrossRef]
- Chappell J, Harman JL, Narasimhan VM, Yu H, Foote K, Simons BD, et al. Extensive Proliferation of a Subset of Differentiated, yet Plastic, Medial Vascular Smooth Muscle Cells Contributes to Neointimal Formation in Mouse Injury and Atherosclerosis Models. Circ Res (2016) 119(12):1313-23. Epub 2016/09/30. [CrossRef]
- Worssam MD, Lambert J, Oc S, Taylor JC, Taylor AL, Dobnikar L, et al. Cellular Mechanisms of Oligoclonal Vascular Smooth Muscle Cell Expansion in Cardiovascular Disease. Cardiovasc Res (2022). Epub 2022/08/23. [CrossRef]
- Wang Y, Nanda V, Direnzo D, Ye J, Xiao S, Kojima Y, et al. Clonally Expanding Smooth Muscle Cells Promote Atherosclerosis by Escaping Efferocytosis and Activating the Complement Cascade. Proc Natl Acad Sci U S A (2020) 117(27):15818-26. Epub 2020/06/17. [CrossRef]
- Longchamp A, Allagnat F, Alonso F, Kuppler C, Dubuis C, Ozaki CK, et al. Connexin43 Inhibition Prevents Human Vein Grafts Intimal Hyperplasia. PLoS One (2015) 10(9):e0138847. Epub 20150923. [CrossRef]
- Peleli M, Bibli SI, Li Z, Chatzianastasiou A, Varela A, Katsouda A, et al. Cardiovascular Phenotype of Mice Lacking 3-Mercaptopyruvate Sulfurtransferase. Biochem Pharmacol (2020) 176:113833. Epub 2020/02/07. [CrossRef]
- Ngowi EE, Afzal A, Sarfraz M, Khattak S, Zaman SU, Khan NH, et al. Role of Hydrogen Sulfide Donors in Cancer Development and Progression. Int J Biol Sci (2021) 17(1):73-88. Epub 20210101. [CrossRef]
- Kiesworo K, MacArthur MR, Kip P, Agius T, Macabrey D, Lambelet M, et al. Cystathionine Gamma Lyase Overexpression Modulates Nad+ Biosynthesis and Enhances Neovascularization. JVS-Vascular Science (2023). [CrossRef]
- Bibli SI, Hu J, Looso M, Weigert A, Ratiu C, Wittig J, et al. Mapping the Endothelial Cell S-Sulfhydrome Highlights the Crucial Role of Integrin Sulfhydration in Vascular Function. Circulation (2021) 143(9):935-48. Epub 2020/12/15. [CrossRef]
- Misra A, Sheikh AQ, Kumar A, Luo J, Zhang J, Hinton RB, et al. Integrin Beta3 Inhibition Is a Therapeutic Strategy for Supravalvular Aortic Stenosis. J Exp Med (2016) 213(3):451-63. Epub 20160208. [CrossRef]
- Slepian MJ, Massia SP, Dehdashti B, Fritz A, Whitesell L. Beta3-Integrins Rather Than Beta1-Integrins Dominate Integrin-Matrix Interactions Involved in Postinjury Smooth Muscle Cell Migration. Circulation (1998) 97(18):1818-27. [CrossRef]
Figure 3.
Static culture significantly increases CSE expression in human vein segments. a) Representative histological VGEL staining and morphometric measurements of intima thickness, media thickness and intima over media ratio of freshly isolated human vein segments (D0) or after 7 days (D7) in static culture. Scale bar 100 µm. b-d) normalized CSE (CTH), CBS and 3-MST (MPST) mRNA (b) and protein expression as assessed by Western blotting (c) or immunohistochemistry (d) in freshly isolated human vein segments (D0) or after 7 days (D7) in static culture. Scale bar 100 µm. 5 to 8 different veins/patients. *p<.033, **p<.002, ***p<.001, as determined by bilateral paired t-test.
Figure 3.
Static culture significantly increases CSE expression in human vein segments. a) Representative histological VGEL staining and morphometric measurements of intima thickness, media thickness and intima over media ratio of freshly isolated human vein segments (D0) or after 7 days (D7) in static culture. Scale bar 100 µm. b-d) normalized CSE (CTH), CBS and 3-MST (MPST) mRNA (b) and protein expression as assessed by Western blotting (c) or immunohistochemistry (d) in freshly isolated human vein segments (D0) or after 7 days (D7) in static culture. Scale bar 100 µm. 5 to 8 different veins/patients. *p<.033, **p<.002, ***p<.001, as determined by bilateral paired t-test.
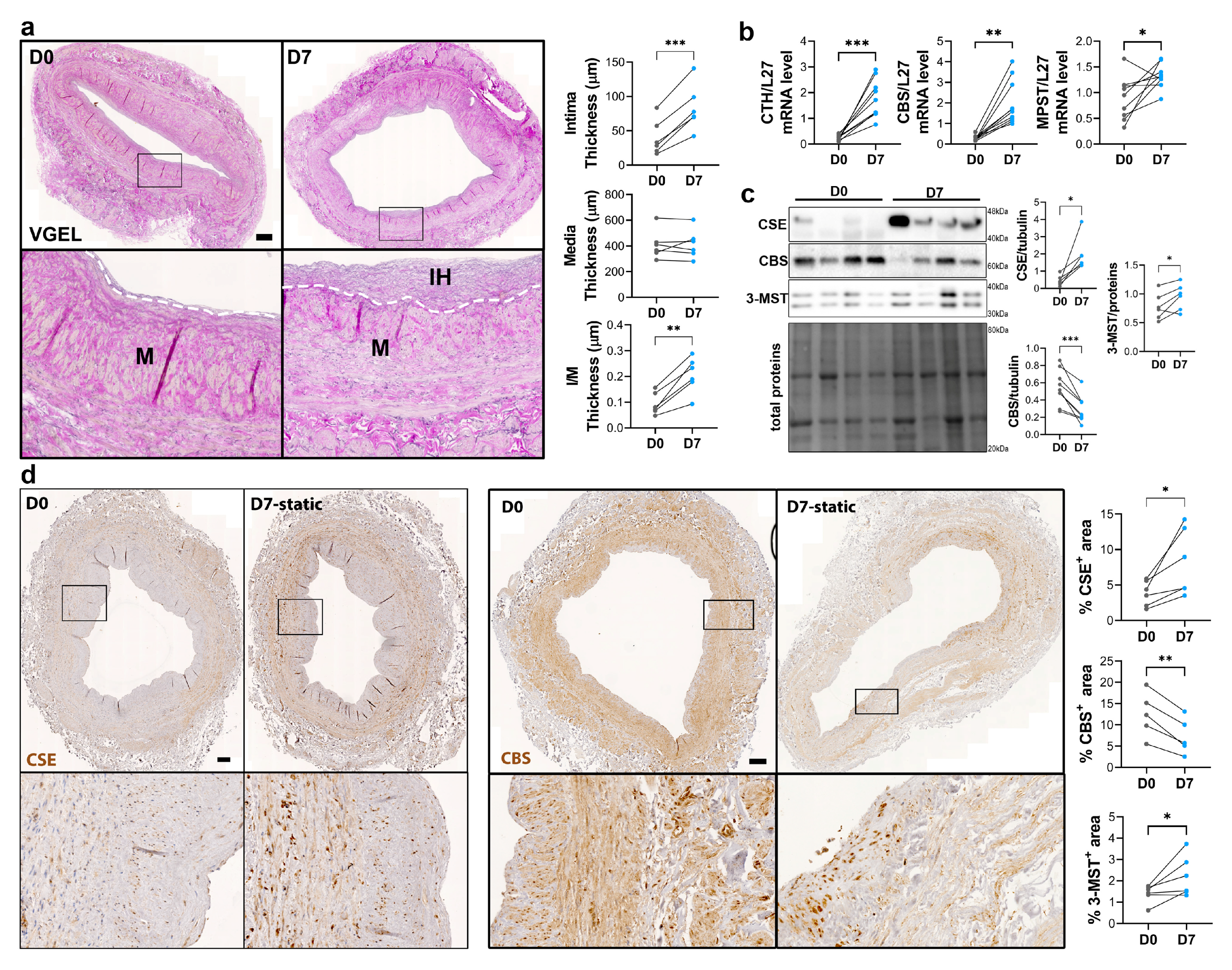
Figure 4.
High pressure perfusion decreases CSE expression in human vein segments. a) Representative histological sections (left panels) stained for elastin (VGEL) and collagen (Herovici) and morphometric measurements (right panels) of intima thickness, media thickness and intima over media ratio of freshly isolated human vein segments (D0) or in veins exposed to pulsatile high pressure (D7-HP; mean=100 mmHg) perfusion for 7 days. Scale bar 50 µm. b-e) Normalized CSE, CBS and 3-MST (MPST) mRNA (b-c) and protein expression as assessed by Western blotting (d) or immunohistochemistry (e) in freshly isolated human vein segments (D0) or in veins exposed to pulsatile low pressure (D7-LP; mean=7 mmHg) or high pressure (D7-HP; mean=100 mmHg) perfusion for 7 days. Scale bar 100 µm. 4 to 5 different veins/patients. *p<.033, **p<.002, ***p<.001, as determined by bilateral paired t-test.
Figure 4.
High pressure perfusion decreases CSE expression in human vein segments. a) Representative histological sections (left panels) stained for elastin (VGEL) and collagen (Herovici) and morphometric measurements (right panels) of intima thickness, media thickness and intima over media ratio of freshly isolated human vein segments (D0) or in veins exposed to pulsatile high pressure (D7-HP; mean=100 mmHg) perfusion for 7 days. Scale bar 50 µm. b-e) Normalized CSE, CBS and 3-MST (MPST) mRNA (b-c) and protein expression as assessed by Western blotting (d) or immunohistochemistry (e) in freshly isolated human vein segments (D0) or in veins exposed to pulsatile low pressure (D7-LP; mean=7 mmHg) or high pressure (D7-HP; mean=100 mmHg) perfusion for 7 days. Scale bar 100 µm. 4 to 5 different veins/patients. *p<.033, **p<.002, ***p<.001, as determined by bilateral paired t-test.
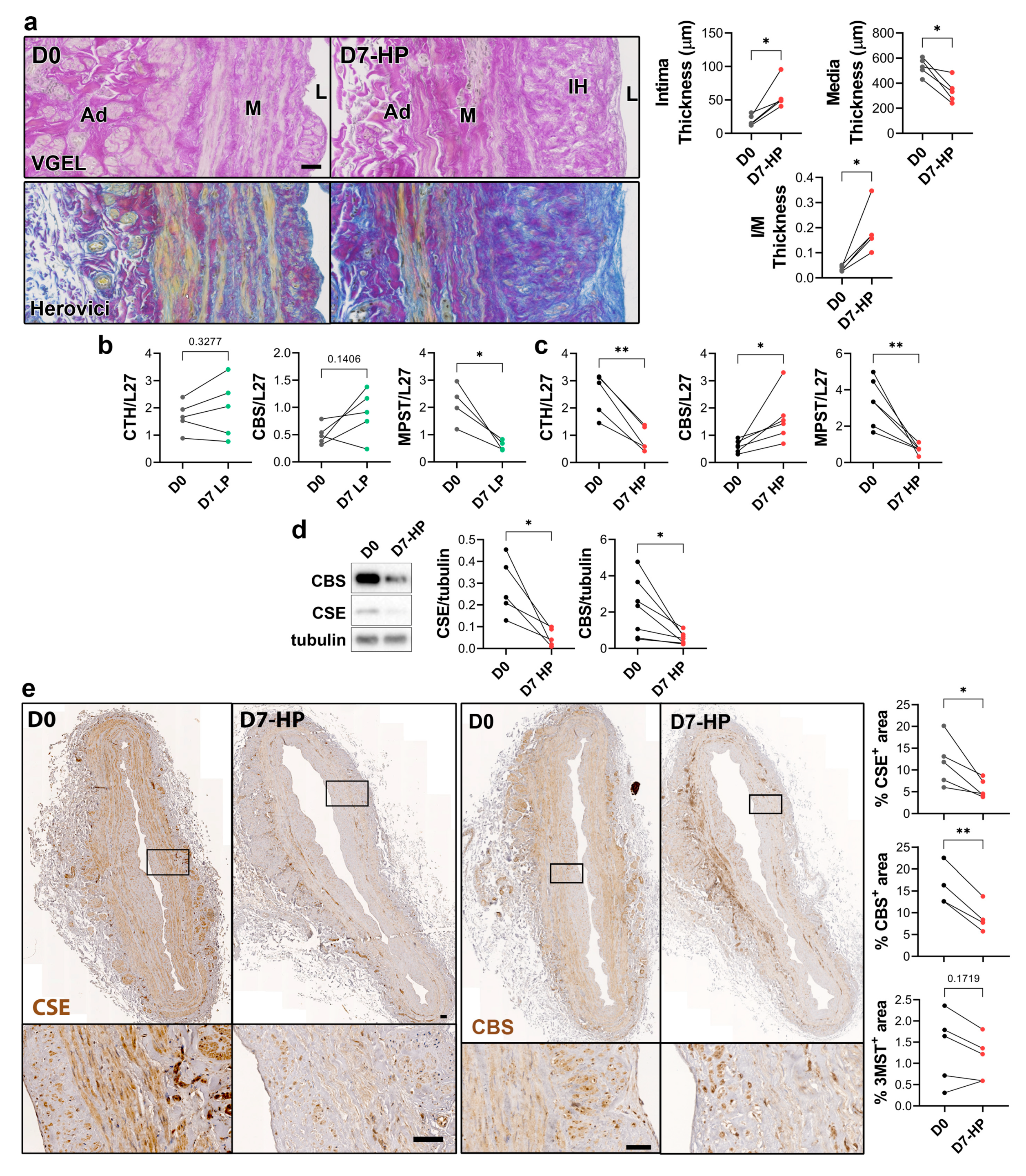
Figure 5.
CSE overexpression in human VSMC increases H2S production and protein persulfidation. a) CSE, CBS and 3-MST protein expression 48 hours post VSMC infection with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. Arrowhead in input indicate GFP expression upon Ad-GFP infection. Data are representative of 6 independent experiments. b) Live-cell imaging of H2S production using the SF7AM probe in VSMC infected, or not (Ctrl), with an adenovirus encoding CSE at MOI 10 or 20, as indicated. Images are representative of 5 independent experiments. Quantitative assessment (violin plots) of SF7AM fluorescent in individual cells across 5 experiments. c) In situ labelling of intracellular protein persulfidation assessed by DAz-2: Cy5.5 (red), normalized to NBF-adducts fluorescence (green), in VSMC infected, or not (Ctrl), with an adenovirus encoding CSE at MOI 10 or 20, as indicated. Data are representative of 5 independent experiments. Representative images of 5 independent experiments. Violin plots of DAz-2: Cy5.5 over NBF fluorescence in individual cells across 5 experiments. b-c) Scale bar 20 μm. **p<.002, ***p<.001 report adjusted p-values as determined by Kruskal-Wallis non-parametric ranking followed by Dunn's multiple comparisons tests. d) Western blot analyses of CSE, CBS and 3-MST expression 48 hours post VSMC transfection with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH1 and 2), as indicated. Data are representative of 6 independent experiments.
Figure 5.
CSE overexpression in human VSMC increases H2S production and protein persulfidation. a) CSE, CBS and 3-MST protein expression 48 hours post VSMC infection with an adenovirus encoding GFP (AdGFP) or CSE (AdCTH) at MOI 10 or 20, as indicated. Arrowhead in input indicate GFP expression upon Ad-GFP infection. Data are representative of 6 independent experiments. b) Live-cell imaging of H2S production using the SF7AM probe in VSMC infected, or not (Ctrl), with an adenovirus encoding CSE at MOI 10 or 20, as indicated. Images are representative of 5 independent experiments. Quantitative assessment (violin plots) of SF7AM fluorescent in individual cells across 5 experiments. c) In situ labelling of intracellular protein persulfidation assessed by DAz-2: Cy5.5 (red), normalized to NBF-adducts fluorescence (green), in VSMC infected, or not (Ctrl), with an adenovirus encoding CSE at MOI 10 or 20, as indicated. Data are representative of 5 independent experiments. Representative images of 5 independent experiments. Violin plots of DAz-2: Cy5.5 over NBF fluorescence in individual cells across 5 experiments. b-c) Scale bar 20 μm. **p<.002, ***p<.001 report adjusted p-values as determined by Kruskal-Wallis non-parametric ranking followed by Dunn's multiple comparisons tests. d) Western blot analyses of CSE, CBS and 3-MST expression 48 hours post VSMC transfection with a control siRNA (siCtrl) or two distinct CTH siRNA (siCTH1 and 2), as indicated. Data are representative of 6 independent experiments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



