Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
The zoonotic Anisakidae nematodes Anisakis sp., Pseudoterranova sp., Contracaecum sp. have been found in Baltic Sea fish species. In particular, there has been an alarming increase in the dispersion of Contracaecum sp. over the last decade, due to the growing number of grey seal (Halichoerus grypus) in the area. This marine mammal is also the final host in the Pseudoterranova sp. life cycle. The aim of our study was to summarize the presence of Anisakidae nematodes in the salmon (Salmo salar) and other commercially important fish species (cod, herring and sprat) from Polish marine waters. This was evaluated by parasitological inspection of the internal organs and body cavities of 89 individual salmon and the results were compared with infection levels in cod, herring and sprat. The presence of Pseudoterranova sp. is reported for the first time here in the muscle tissue of salmon from the Baltic Sea. In addition, we highlight the elevated levels of cod and sprat infection with Contracaecum sp. over the last decade. The presence of Anisakidae nematodes in commercially important fish species may have implications for human health since these parasites pose a risk of anisakidosis.
Keywords:
Anisakidae
; salmon
; zoonotic
; southern Baltic
1. Introduction
The sustained growth in overall production and consumption have been observed in the global fishery and aquaculture sectors [1]. Although seafood is part of a healthy diet, in particular cases its consumption may cause health problems [2,3]. Presence of parasites in wild fish is rather rule then exception. Most of the endoparasites present in visceral organs of fish are removed during the fish processing. However, there is a group of endoparasites able to migrate into edible parts of the fish (like muscle tissue, liver). These parasites may be a source of zoonosis (zoonotic diseases), therefore diseases that are transmitted from animal species to humans [4]. Zoonotic Anisakidae nematodes of the genera Anisakis, Pseudoterranova and Contracaecum have been found in a variety of Baltic Sea fish species, also these of consumers interest. The list of the most important Baltic Sea fish species intended for human consumption contains of: herring Clupea harengus, cod Gadus morhua, sprat Sprattus sprattus, salmon Salmo salar, flatfishes like flounder Platichthys flesus. Numerous studies are describing the presence of Anisakis simplex (called also herring worm or whale worm) in herring [5,6,7]. This parasite has been noted also in cod [8]; stickleback Gasterosteus aculeatus [9] and flatfishes: flounder [9], dab Limanda limanda [10], plaice Pleuronectes platessa [10] or turbot Psetta maxima [10]. However, during last decade the most abundant nematode in these fish species in the Baltic Sea is Contracaecum osculatum so colled liver worm [8,11,12,13], reported also in sprat [14], great sandeel [15] and salmon [16] Pseudoterranova decipiens (named also codworm or seal worm) the rarest among listed nematode species, was found in cod [8] and shorthorn sculpin [17] from the Baltic Sea.
The life cycles of these nematodes are very similar, with marine mammals playing the role of final hosts [18,19], but there is clear host specificity: particular species of marine mammal are responsible for closing the life cycle of a particular parasite species. Thus, the final hosts for A. simplex are cetaceans, in the Baltic Sea represented by the harbor porpoise, Phocoena phocoena [20]; for C. osculatum, the final host is the grey seal, Halichoerus grypus [21]; and for P. decipiens, both the harbor seal, Phoca vitulina [22] and the grey seal [23] can adopt this role. In the final host, larval stage L4 transforms to the dioecious mature stage, which is capable of producing fertilized eggs. Parasite eggs are transferred into the water environment via the feces of their final hosts. Transformation to the larval stages L1 and L2 (or even L3) occurs within the egg [24]. Crustaceans (intermediate hosts) take up L2 (or L3) larvae and transformation from L2 to L3 can take place in these intermediates. Such infected crustaceans are eaten by pelagic fish (e.g. sprat, herring), which in turn are eaten by predatory fish (e.g. cod, salmon). Marine mammals represent the last link in the trophic chain: they feed on infected fish and thus become the definitive, i.e. final, host and subsequently the life cycle is complete [24,25,26].
It is important to emphasize that Pseudoterranova spp. are characterized by the demersal life cycle. Eggs settle to the benthos, complete embryonation and hatch [27]. Then second or third-stage larvae (L2 or L3) emerge from the eggs and attach to the substratum, where intake by infaunal and epifaunal benthic copepods (harpacticoids and cyclopoids) may occur [28]. Various benthic macroinvertebrates species have been found infected with P. decipiens [27]. Contrary Anisakis spp. are considered to follow a pelagic life cycle, involving pelagic invertebrate and vertebrate intermediate hosts [25,29]. Eggs embryonate in the seawater [9]. [30] found larvae surrounded by sheaths with lateral extensions, which may be an adaptation to pelagic conditions. Larvae were able to swim and used mainly pelagic crustacean hosts, such as copepods and euphausiids. According to [24] in case of C. osculatum the first two moults occur in the egg, the next and last two moults occur in the final host, the seal. Therefore no moult occurs in the crustacean or fish intermediate hosts and larvae are free-swimming [24].
In the life cycle of these Anisakidae nematodes, fish play the role of paratenic hosts in which larvae do not change their larval stage, but migrate to the fish organs and muscle tissue, where they wait until the host fish is eaten by marine mammals. Organ specificity has been described in particular species of nematode for a particular species of fish. Each of the three listed parasite species have different organ preferences, which may vary depending on the host species. A. simplex is often found in herring in the body cavity and on the surfaces of the viscera [31]; in cod it is mostly recorded in muscle [32]; in salmon it has been found in the muscles and viscera [33]. P. decipiens prefers muscle tissue in both cod [32] and salmon [34]. C. osculatum is found predominantly in the liver of cod [11] and also salmon [16,35]. Fish accumulate these nematodes throughout their lives.
The European Food Safety Authority (EFSA) recommends that all wild-caught fish intended to be eaten raw or almost raw must be considered at risk of containing any viable parasite of concern to human health [36]. Thus, for example, the consumption of raw or unprocessed fish containing live Anisakidae nematodes can lead in humans to anisakidosis or anisakiosis in case of intake of Anisakis spp. larvae. The symptoms of acute anisakidosis are non-specific: nausea, diarrhea, vomiting and intense abdominal pain [37,38]. Adequate treatment of infected fish kills the parasites [39], but some allergenic proteins of anisakid nematodes are thermostable [40,41]. Therefore, even dead parasites can be dangerous and can cause allergic symptoms. Accordingly, the level of infection with Anisakidae nematodes should be monitored, at least in commercially important fish species.
For decades, a range of fish species in the Baltic Sea, including cod (Gadus morhua), sprat (Sprattus sprattus), herring (Clupea harengus), salmon (Salmo salar), flatfishes have been crucial to Polish fisheries. These fish were mainly intended for human consumption. The National Marine Fisheries Research Institute (NMFRI) has collected data regularly since the 1980s on the level of herring infection with Anisakidae parasites in the body cavity; data on cod-liver infection with these nematodes are also available from 2011. In contrast, information on sprat infection with Anisakids is scarce. Similarly, there is a lack of knowledge about the level of salmon infection with these zoonotic nematodes. Therefore, the aim of our study was to summarize current data on the presence of zoonotic Anisakidae nematodes in the salmon (S. salar) and other commercially important fish species (cod, herring and sprat) from Polish marine waters.
2. Materials and Methods
2.1. Sampling of Salmon and Parasitological Inspection
Salmon were caught in March 2012 in the Polish waters of the southern Baltic Sea (ICES 39G7 and 39G8) during commercial cruises and were gutted on board. Sex of the fish was determined. Basic measurements (length and gutted weight) were conducted and scales were collected for age determination. To evaluate the condition of the fish Clark’s condition factor (KC) was calculated as follows: KC = 100 * GW / L3, where GW is gutted weight and L is the total length of the individual fish. In total, the body cavities of 89 fish were visually inspected for the presence of Anisakidae nematodes. Descriptions of nematode anatomo-morphological features as described [42,43,44] were used for preliminary identification of the larva found. Parasitological descriptors were calculated according to the definitions given by [45].
2.2. Identification of Nematode
The single nematode larva found was also identified using molecular methods (ITS-1 rDNA). Genomic DNA was isolated using the Sherlock AX DNA purification kit (A&A Biotechnology) by a modified method that employed mechanical lysis (i.e. mincing tissue with a surgical blade). For ITS-1 analysis, a polymerase chain reaction (PCR) was performed using NC5 (forward) 5’-GTA GGT GAA CCT GCG GAA GGA TCA TT-3’ and NC13R (reverse) 5’-GCT GCG TTC TTC ATC GAT 3’ primers [46,47]. The reaction mixture consisted of 25 μl 2x PCR Master Mix Plus High (A&A Biotechnology), 0.2 μl each primer (concentration 100 μM) and 5 μl DNA template, supplemented with deionized water up to 50 μl. The PCR conditions were as follows: 2 min at 94°C (initial denaturation) followed by 35 cycles of denaturation at 94°C for 30 s, annealing of primers at 58°C for 30 s, strand elongation at 72°C for 45 s and a final extension step of 5 min at 72°C. PCR products were sequenced directly using standard procedures and the amplification primers. Sequences were analysed using GeneStudio TM Professional (GeneStudio, Inc., USA) and confirmed by a BLAST search of GenBank. The sequence obtained was deposited in GenBank with the accession number given in the Results.
3. Results
3.1. Salmon
The salmon investigated in this study were between 43 and 102 cm long, but the majority of the fish (63.92 %) belonged to the length class 70-79 cm. Among analysed fish 66.29% were females (66 - 96 cm in lenght) and 33.71% males (43 - 102 cm in lenght). Length-weight relationfish was similar for males and females (Figure 1.). Clark’s condition factor (KC) was similar for both sexes: females (min = 0.754; max = 1.128; mean = 0.934) and males (min = 0.780; max = 1.131; mean = 0.944). Regarding age, two-year-old fish predominated (71.91%), one-year-old fish constitutes 15% of analysed individuals, three-year-old 8% and four-year-old 2%.
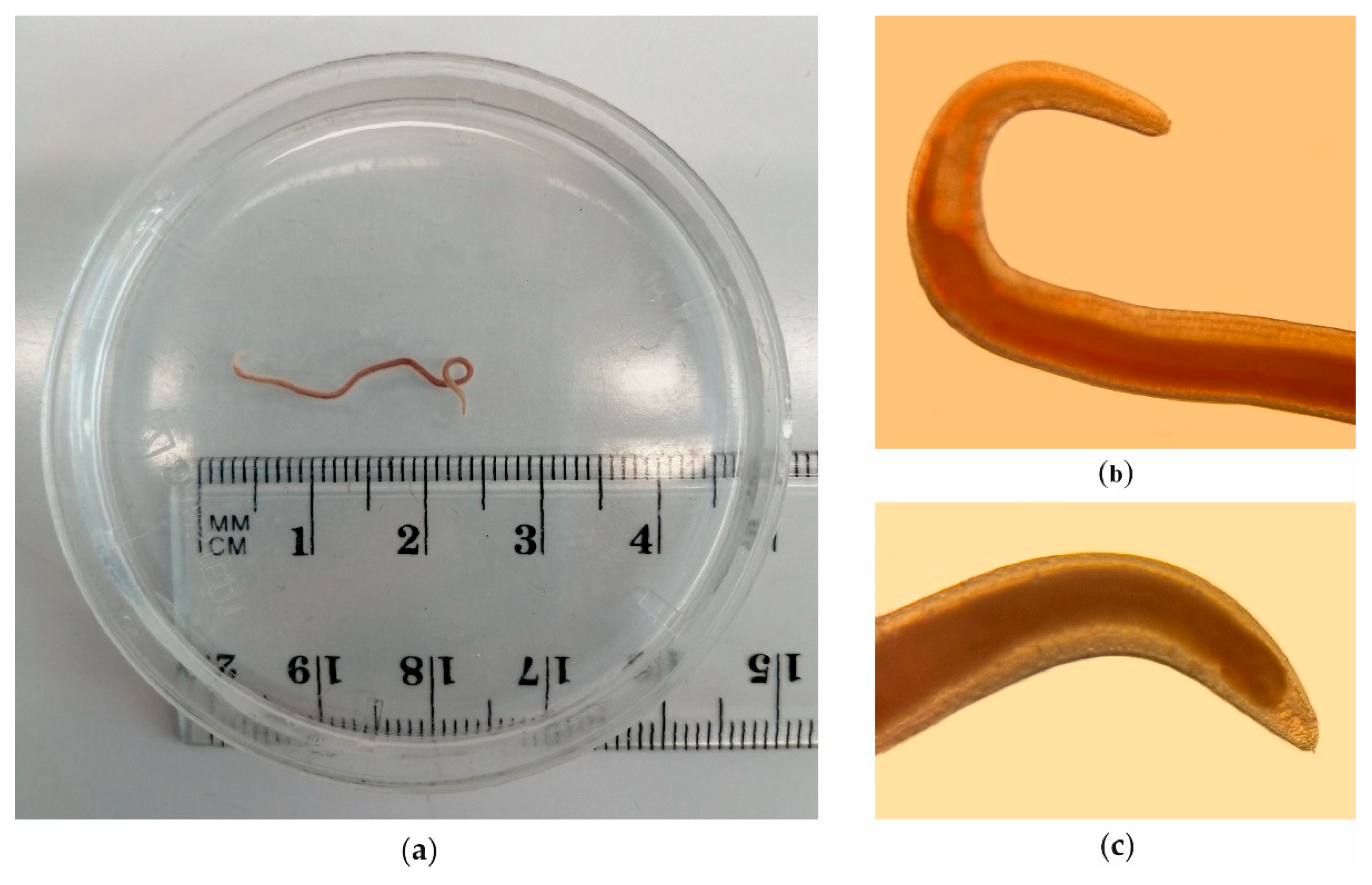
During visual inspection of the body cavity of a single salmon, one Anisakidae nematode larva, attached to muscle tissue, was detected. The larva was identified based on anatomo-morphological features, and the prevalence of infection was 1.1% (intensity of infection = 1). Pseudoterranova sp. from Baltic salmon has been presented on Figure 2.
3.2. Molecular Identification
ITS-1 amplification and sequencing identified the larva as Pseudoterranova sp. The sequence has been deposited in GenBank with ID OQ732699. This is the first report of Pseudoterranova sp. in the muscle tissue of salmon from the Baltic Sea.
4. Discussion
Parasites, including A. simplex, C. osculatum and P. decipiens, found in fishery products represent a possible public health risk [48]. All the above species of nematode have been detected in commercially important fish species, after harvesting for human consumption in Polish marine waters.
In this report, we describe a single example of a salmon that was infected with an Anisakidae larva attached to the muscle tissue. It is the first report of Pseudoterranova sp. in the muscle tissue of a salmon from the Baltic Sea. Attachment of the parasite to the muscle tissue in the body cavity indicates post-mortem migration of the larva. Presented studies were conducted in 2012, when the dispersion of Anisakidae nematodes in the southern Baltic Sea waters was not as widespread as today. Repeating the study in more recently caught salmon, with a focus on the presence of Anisakidae parasites not only in the body cavity, but also in the muscle tissue, would be justified, due to the high level of infection observed recently in other fish species caught in Polish marine waters. Salmon are diadromous fish, which spawn in the rivers, where they spend the first years of their life. They then undertake a migration to their feeding areas, which in the Baltic are located in the southern part of the sea, eastwards of Bornholm, where seals colonies occur. In the sea, salmon prey intensively on smaller fish, mostly sprat. It is at this time that salmon are exposed to infection with Anisakidae nematodes. Analysis of the livers of 120 salmon caught in Polish marine waters in 2020 revealed that 13% of them were infected with Anisakids, i.e. C. osculatum and Pseudoterranova sp. [16]. It was the first attempt to describe the level of liver infection in salmon caught in Polish marine waters (southern Baltic Sea).
The reports describing the infection of cod in Polish marine waters with Anisakidae nematodes date back to the 1970s. In 1974–75 the average prevalence of cod infection with C. osculatum was 28%, with intensities of 1–24, while the presence of A. simplex was recorded in the liver of cod in this area for the first time [49]. The studies conducted by [50] from 1988 to 1993 also indicate that nematodes were mainly found in the liver, predominately C. osculatum (the prevalence of infection ranged between 1.8 and 3.2% in cod sampled from several Polish fishing grounds), but also A. simplex. [51] investigated cod in the period 1987 to 1994 and revealed the presence of all three zoonotic Anisakids. The vast majority were C. osculatum (98% of larvae), but A. simplex and P. decipiens were also found (in 1 of 3036 fish examined). Overall, 4% of the cod sampled were infected with Anisakidae. [52] recorded the presence of A. simplex and C. osculatum in the liver of cod sampled in the Gulf of Gdansk and the Pomeranian Bay (the Western Baltic) between 2001–2003: the prevalence of infection with A. simplex larvae in cod ranged from 1.9% in the Gulf of Gdansk to 8.9% in the Pomeranian Bay, while with C. osculatum the prevalence was 9.8% and 2.5%, respectively. Studies on the presence of Anisakidae nematodes in cod liver conducted in 2011 revealed the presence of Contracaecum sp. (95% of collected nematodes), Anisakis sp. and Pseudoterranova sp.; the prevalence of cod infected with Anisakidae was 11% [11]. Recent results are alarming, indicating that the level of cod infection with C. osculatum extends throughout the entire southern Baltic, reaching 90% prevalence in ICES Subdivision 26 for length classes of cod 35 cm and greater, and showing an increasing trend for the period 1987–2019 [53].
In the case of herring from the Baltic Sea, A. simplex is the most common nematode parasite recorded. Studies conducted by [5] in 1970s indicated that more than 90% of herring caught in May in the Pomeranian Gulf (western part of Polish marine waters) was infected with this nematode. The highest prevalence of herring infection with A. simplex in the Polish EEZ, estimated using a generalized linear model, was in the period 1997-1999 (up to 97% in fish at length of 30 cm) [6]. The estimated year effect decreased markedly during the period 2013–2019, however, although the highest prevalence of infection was still >26% in herring caught in the first and second quarters of 2019 in coastal waters of ICES Subdivision 24 [53]. It is worth to emphasise, that there are a few herring stocks in the Baltic Sea and for decades the scientists stated that only western herring migrating out off the Baltic Sea might get infected with A. simplex larvae [5]. It was believed that herring can not get infected with that parasite in the Baltic Sea due to the lack of Euphasiacean intermediate hosts, lack of cetacean final hosts responsible for dispersion of the eggs of parasites in the water, too low salinity [5]. The results obtained by [54] indicated that infected fish represented a mixed group of western and central Baltic herring. Experiments conducted by [9] revealed that intermediate hosts are not indispensable in the life cycle of A. simplex. Morover, nowadays due to the presence of harbour porpoise and appearance of other visiting cetacean species in the western area of the Baltic Sea it is likely that fish might get infected with A. simplex also in these sea waters. Such possibility has been already suggested by [55], despide of the fact, that so far there was no evidence that A. simplex can complete the life cycle in the Baltic Sea.
Sprat from Polish marine waters was investigated between 1987 and 1990 for the presence of Anisakidae nematodes (3511 individuals analysed), but parasites were not detected [14]. In 2009 and 2011 in the Gulf of Gdańsk [56] found 2 of 187 sprats to be infected with a single C. osculatum larva each (prevalence 1.1%, mean intensity 1.0, abundance -0.5). In 2016, 2 of 30 sprats from the central offshore area of Polish marine waters were infected with C. osculatum (prevalence of infection in that specific area was 6.67%, and 2.22% across Polish marine waters as a whole; mean intensity was 1.0) [14].
Pilot studies conducted in NMFRI in 2012 on the presence of Anisakidae nematodes in flounder from the Gulf of Gdańsk did not reveal the presence of nematodes. Similar results have been obtained during studies conducted in 2016 on flounders from western, central and eastern parts of the Polish marine waters. In 2019 Anisakidae nematode C. osculatum has been detected in the livers of flounder [57].
The dispersion of zoonotic Anisakidae nematodes among marine organisms of the Baltic Sea has been reported over the last decade [8,13,16] and is also observed in fish caught in Polish waters. It is in line with the growing population of the grey seal, which is the final host in the life cycle of C. osculatum [21] and P. decipiens [23], to over 40,000 individuals in 2020 for the whole Baltic Sea [58]. Studies on the presence of nematodes in Baltic Sea seals (H. grypus, P. vitulina and P. hispida) reported that each seal was infected with Anisakidae nematodes, with one grey seal showing an intensity of infection of up to 590 nematodes [59]. It is worth emphasizing that C. osculatum and P. decipiens females are very productive: one female P. decipiens produces on average about 60,000 eggs, while a female C. osculatum produces on average about 9,000 eggs [60]. If we multiply the number of grey seals by the number of nematodes in each seal and by the number of eggs that might be produced by each mature female parasite, we end up with a extremely high load of nematodes shed into the environment. The presence of Anisakidae parasites in the food web of the Baltic Sea is not surprising, therefore.
However, if in the Baltic Sea the grey seal is the definitive host for Contracaecum and Pseudoterranova genera, the question arises why dispersion of only one parasite species - C. osculatum is currently reported in the area. It might be explained by several factors. First of all the demersal life cycle of Pseudoterranova spp. may not be conducive to dispersion of that genera in the Baltic Sea, that may be related with the lack of particular invertebrate species or limited number of intermediate hosts in particular area, needed in the life cycle of that parasite [17]. Dispersion of the C. osculatum might be related with the fact that free-swimming larvae do not need intermediate hosts to complete the life cycle, that has been proven by [24]. Moreover, Pseudoterranova spp. prefer water with high salinity [17], because low salinity limits the survival of the sealworm larvae [61] and occurrence of its the intermediate hosts [17]. Contrary, C. osculatum has adapted to the prevailing even in brackish water conditions of the northern Baltic Sea - Bothnian Bay [62]. Due to the mentioned reasons, the range of Pseudoterranova occurrence is limited mainly by low salinity in the Eastern Baltic Sea [63].
In case of representatives of Anisakis genera in the Baltic Sea low number of cetaceans - indespensable final hosts and low water salinity are pointed out as the most important factors limiting the dispersion [64]. According to [65] distribution of this parasite is restricted to south-western areas of the Baltic Sea.
The main source of fish infection with Anisakidae nematodes is infected prey. Studies conducted at the NMFRI revealed that cod might become infected with A. simplex via Crangon crangon and with C. osculatum via Gammarus sp. [66]. Fish feed on invertebrates, and fish like sprat are important elements in the diet of cod [67,68] and salmon [16]. Therefore, sprat might be a source of infection for predatory fish.
Furthermore, sprat, herring, cod and salmon are consumed by humans, when the presence of zoonotic nematodes in a raw or inadequately prepared fish meal might lead to acute anisakidosis or an allergic response. Treatment to kill viable parasites in fishery products intended for human consumption is mandatory in many countries. Studies conducted at the NMFRI have shown that Anisakidae may survive freezing if it is performed incorrectly [69]; therefore, compliance with legal regulations regarding marine food treatment is crucial to avoid consumers suffering health problems.
In fact, most reported human case studies are related to the consumption of A. simplex larvae [12]. However, there are some examples of anisakidosis caused by P. decipiens [12,70,71]. Both nematode species prefer to localise to muscle tissue in their fish hosts, thus increasing the risk of infection with these nematodes. However, C. osculatum larvae, which prefer the liver of fish, also have zoonotic potential [12,72] and infections with this species have been recorded, possibly because fish liver might be consumed as well as muscle. In the past, most of the anisakidosis cases have been noted in countries where consumption of raw fish is common, therefore Asian countries especially Japan [12]. Recent reviews prepared by [73,74] show that allergic anisakidosis has been reported almost all over the world with the hot spot in northeastern Atlantic, where during nearly 40 years the average Anisakis spp. abundance increased more than 100 [64]. It is noteworthy that, although information on salmon infection with Anisakidae nematodes is scarce and the level of infection relatively low, there was a recently reported case of human anisakidosis in Poland caused by ingestion of viable A. simplex present in a meal of raw salmon [75]. Japanese cuisine is becoming more popular in many European countries, including Poland. With the increasing popularity of sushi and sashimi the incidence of cases of anisakidosis is expected to increase [12,76]. Morover, [77] drew attention to the issue that lack of awareness related with parasitic diseases is associated also with developed countries. It was suggested that crucial for prevention of seafood-borne parasitic diseases are raising awareness, supporting research and the training of all stakeholders [77].
Presence of the parasite has negative impact no only on the consumers, but above all on the fish host. Numerous A. simplex larvae reported in Atlantic cod were responsible for severe inflammatory reactions with tissue deformation, including marked cellular infiltration in the stomach wall and mucosa [78] and the term “stomach crater syndrome” was applied for this pathological reaction [12]. High number of A. simplex larvae causing so colled “red vent syndrome” (haemorrhages and inflammation around the vent) was reported in Atlantic salmon and sea trout returning to rivers in Scotland, England and Wales [79,80]. It was proven, that this nematode species provokes an inflammatory reaction in salmonids [81]. In case of P. decipiens encapsulation by host cells has been observed in case of cod muscle tissue [12]. However, its presence may affect the physiological state, health and survival of the host [12]. P. decipiens excretes substances that might be responsible for muscle contractility of fish, therefore reduce its swimming abilities and make easier the predation by marine mammals [12].
Negative impact of parasites on fish individuals may have consequences for the entire population. The negative effect of the presence of Anisakidae nematodes in commercially important fish stocks is most visible in the case of Baltic cod. Thus, a study conducted at the NMFRI [82] revealed that Anisakidae nematodes in the liver of cod negatively affects the condition of the fish, may increase natural mortality, and may be one of the reasons for the critical condition of the Baltic cod stocks.
5. Conclusions
The facts described above show that zoonotic Anisakidae nematodes are emerging pathogens for Baltic sea fish species and the humans that eat them. The level of infection with Anisakidae nematodes should be monitored, at least in commercially important fish species. It is also important to focus future studies on the presence of the parasites in edible parts of the salmon, due to the implications for the human health.
Author Contributions
Conceptualization, KN-A, MP and JP; methodology, KN-A; software, KN-A; investigation, KN-A; writing - original draft preparation, KN-A.; writing - review and editing, MP and JP; funding acquisition, KN-A. All authors have read and agreed to the published version of the manuscript.
Funding
Biological material collected as part of the EU Multiannual Programme for Fisheries Data Collection (EU DCF) was used for our research. This research was supported by the statutory funding of National Marine Fisheries Research Institute provided by the Ministry of Science and Higher Education in Poland (DOT 21-22-23 ParaSalmon).
Institutional Review Board Statement
The current study was carried out in strict accordance with the recommendations in the Polish ACT of 21 January 2005 of Animal Experiments (Dz. U. of. 2005 No 33, item 289) and the special permission is not required by local law for the carried-out experiments.
Informed Consent Statement
Not applicable.
Data Availability Statement
The presented in the current manuscript data are available from the corresponding author upon request. The sequences of the parasite is deposited in GenBank with ID OQ732699.
Acknowledgments
The authors would like to thank to the team of Technicians for support during ichthyological analysis and collecting the samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. The State of World Fisheries and Aquaculture; Food and Agriculture Organisation of the United Nations: Italy, 2014; p. 223. [Google Scholar]
- Deardorff, T.L.; Overstreet, R.M. Seafood-Transmitted Zoonoses in the United States: The Fishes, the Dishes, and the Worms. In Microbiology of Marine Food Products; Ward, D.R., Hackney, C., Eds.; Springer: Boston, MA, 1991. [Google Scholar] [CrossRef]
- Shamsi, S. Seafood-borne parasitic diseases in Australia: How much do we know about them? Microbiol. Aust. 2016, 37, 27–29. [Google Scholar] [CrossRef]
- Han, B.A.; Kramer, A.M.; Drake, J.M. Global patterns of zoonotic disease in mammals. Trends Parasitol. 2016, 32, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Grabda, J. The dynamics of the nematode larvae, Anisakis simplex (Rud.) invasion in the South Western Baltic herring (Clupea harengus L.). Acta Ichthyol. Piscat. 1974, 4, 3–21. [Google Scholar] [CrossRef]
- Podolska, M.; Horbowy, J. Infection of Baltic herring (Clupea harengus membras) with Anisakis simplex larvae, 1992–1999: a statistical analysis using generalized linear models. ICES J. Mar. Sci., 2003, 60, 85–93. [Google Scholar] [CrossRef]
- Lang, T.; Damm, U.; Weber, W.; Neudecker, T.; Kühlmorgen-Hille, G. Infestation of herring (Clupea harengus L.) with Anisakis sp. larvae in the western Baltic. Archiv für Fischereiwissenschaft 1990, 40(1/2), 101–117. [Google Scholar]
- Mehrdana, F.; Bahlool, Q.Z.M.; Skov, J.; Marana, M.H.; Sindberg, D.; Mundeling, M.; Overgaard, B.C.; Korbut R.; Strøm, S.B.; Kania, P.W.; Buchmann, K. Occurrence of zoonotic nematodes Pseudoterranova decipiens, Contracaecum osculatum and Anisakis simplex in cod (Gadus morhua) from the Baltic Sea. Vet. Parasitol. 2014, 205, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Køie, M. Experimental infection of copepods and sticklebacks Gasterosteus aculeatus with small ensheathed and large third-stage larvae of Anisakis simplex (Nematoda, Ascaridoidea, Anisakidae). Parasitol. Res. 2001, 87, 32–36. [Google Scholar] [CrossRef]
- Lüthen, K. Mitteilung zum Auftreten von Anisakis sp. Larvae (Nematoda, Ascaridida) bei Plattfischen aus Gebieten der mittleren Ostsee. Wiss Z Padagog Hochschule Lisselotte Hermann Güstrow. 1989, 1, 83–87. [Google Scholar]
- Nadolna, K.; Podolska, M. Anisakid larvae in the liver of cod (Gadus morhua) L. from the southern Baltic Sea. J. Helminthol. 2014, 88, 237–246. [Google Scholar] [CrossRef]
- Buchmann, K.; Mehrdana, F. Effects of anisakid nematodes Anisakis simplex (s.l.), Pseudoterranova decipiens (s.l.) and Contracaecum osculatum (s.l.) on fish and consumer health. Food and Waterborne Parasiol. 2016, 4, 13–22. [Google Scholar] [CrossRef]
- Zuo, S.; Kania, P.W.; Mehrdana, F.; Marana, M.H.; Buchmann, K. Contracaecum osculatum and other anisakid nematodes in grey seals and cod in the Baltic Sea: molecular and ecological links. J. Helminthol. 2018, 92, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Nadolna-Ałtyn, K.; Szostakowska, B.; Podolska, M. Sprat (Sprattus sprattus) as a possible source of invasion of marine predators with Contracaecum osculatum in the Southern Baltic Sea. Russ. J. Mar. Biol. 2018, 44, 471–476. [Google Scholar] [CrossRef]
- Nadolna-Ałtyn, K.; Podolska, M.; Szostakowska, B. Great sandeel (Hyperoplus lanceolatus) as a putative transmitter of parasite Contracaecum osculatum (Nematoda: Anisakidae). Parasitol. Res. 2017, 116, 1931–1936. [Google Scholar] [CrossRef]
- Nadolna-Ałtyn, K.; Pawlak, J.; Podolska, M.; Lejk, A. Contracaecum osculatum and Pseudoterranova sp. in the liver of salmon (Salmo salar) from Polish marine waters. Fish. Aquatic. Life. 2023, 31, 44–53. [Google Scholar] [CrossRef]
- Lunneryd, S.G.; Boström, M.K.; Aspholm, P.E. Sealworm (Pseudoterranova decipiens) infection in grey seals (Halichoerus grypus), cod (Gadus morhua) and shorthorn sculpin (Myoxocephalus scorpius) in the Baltic Sea Parasito. Res. 2015, 114, 257–264. [Google Scholar] [CrossRef] [PubMed]
- McClelland, G.; Misra, R.K.; Martell, D.J. Larval anisakine nematodes in various fish species from Sable Island Bank and vicinity. Population biology of sealworm (Pseudoterranova decipiens) in relation to its invertebrate and seal hosts. Can. Bulletin Fish. Aquat. Sci. 1990, 222, 83–118. [Google Scholar]
- Klimpel, S.; Palm, H.W. Anisakid nematode (Ascaridoidea) life cycles and distribution: increasing zoonotic potential in the time of climate change. In Progress in Parasitology; Mehlhorn, H., Ed.; Parasitology Research Monographs; Springer: Berlin, Heidelberg, 2011; pp. 201–222. ISBN 978-3-642-21396-0. [Google Scholar]
- Herreras, M.V.; Balbuena, J.A.; Aznar, F.J.; Kaarstad, S.E.; Fernandez, M.; Raga, J.A. Population structure of Anisakis simplex (Nematoda) in harbor porpoises Phocoena phocoena off Denmark. J. Parasitol. 2004, 90, 933–938. [Google Scholar] [CrossRef]
- Fagerholm, H.P. Systematic position and delimitation of Ascaroid nematode parasites of the genus Contracaecum with a note on the superfamily Ascaridoidea; National Veterinary Institute: Helsinki, 1990. [Google Scholar]
- Aspholm, P.E.; Ugland, K.I.; Jødestøl, K.A.; Berland, B. Sealworm (Pseudoterranova decipiens) infection in common seals (Phoca vitulina) and potential intermediate fish hosts from the outer Oslofjord. Int. J. Parasitol. 1995, 25, 367–373. [Google Scholar] [CrossRef]
- Hauksson, E. The prevalence, abundance, and density of Pseudoterranova sp.(p) larvae in the flesh of cod (Gadus morhua) relative to proximity of grey seal (Halichoerus grypus) colonies on the coast off Drangar, Northwest Iceland. J. Mar. Biol. 2011, ID 235832, 1–8. [Google Scholar] [CrossRef]
- Køie, M.; Fagerholm, H.P. The life cycle of Contracaecum osculatum (Rudolphi, 1802) sensu stricto (Nematoda, Ascaridoidea, Anisakidae) in view of experimental infection. Parasitol. Res. 1995, 81, 481–489. [Google Scholar] [CrossRef]
- Klimpel, S.; Palm, H.W.; Rückert, S.; Piatkowski, U. The life cycle of Anisakis simplex in the Norwegian Deep (northern North Sea). Parasitol. Res. 2004, 94, 1–9. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Hederholm, R.; Schack, H.B.; Møller, L.N.; Storr-Paulsen, M.; Dzido, J.; Rokicki, J. Occurrence of anisakid nematodes in Atlantic cod (Gadus morhua) and Greenland cod (Gadus ogac) West Greenland. Acta Parasitol. 2010, 55, 81–89. [Google Scholar] [CrossRef]
- Martell, D.J.; McClelland, G. Transmission of Pseudoterranova decipiens (Nematoda: Ascaridoidea) via benthic macrofauna to sympatric flatfishes (Hippoglossoides platessoides, Pleuronectes ferrugineus, P. americanus) on Sable Island Bank, Canada. Mar. Biol 1995, 122, 129–135. [Google Scholar] [CrossRef]
- McClelland, G. Phocanema decipiens (Nematoda: Anisakidae): Experimental infections in marine copepods. Can. J. Zool. 1982, 60, 502–509. [Google Scholar] [CrossRef]
- Klimpel, S.; Kellermanns, E.; Palm, H.W. The role of pelagic swarm fish (Myctophidae: Teleostei) in the oceanic life cycle of Anisakis sibling species at the Mid-Atlantic Ridge, Central Atlantic. Parasitol. Res. 2008, 104, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Køie, M.; Berland, B.; Burt, M.D.B. Development to third-stage larvae occurs in the eggs of Anisakis simplex and Pseudoterranova decipiens (Nematoda, Ascaridoidea, Anisakidae). Can. J. Fish. Aquat. Sci. 1995, 52, 134–139. [Google Scholar] [CrossRef]
- Horbowy, J.; Podolska, M. Modelling infection of Baltic herring (Clupea harengus membras) by larval Anisakis simplex. ICES J. Mar. Sci. 2001, 58, 321–330. [Google Scholar] [CrossRef]
- Nadolna-Ałtyn, K.; Podolska, M.; Pawlak, J.; Szostakowska, B. Distribution of anisakid nematodes in the muscle tissue of cod (Gadus morhua) from the Norwegian Sea. Oceanologia. 2022, 64, 489–502. [Google Scholar] [CrossRef]
- Kent, A.J. Pert, C.C., Briers, R.A., Diele, K., Rueckert, S. Increasing intensities of Anisakis simplex third-stage larvae (L3) in Atlantic salmon of coastal waters of Scotland. Parasit. Vectors. 2020, 13, 62. [Google Scholar] [CrossRef]
- Wotten, R.; Yoon, G.H.; Bron, J.E. A Survey of Anisakid Nematodes in Scottish Wild Atlantic Salmon. FSAS Project S14008. Final Report 3rd February 2010.
- Setyawan, A.C.; Zuo, S.; Kania, P.W.; Buchmann, K. Endoparasitic helminths in Baltic salmon Salmo salar: ecological implications. Dis. Aquat. Org. 2019, 135, 193–199. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on risk assessment of parasites in fishery products. Panel on Biological Hazards (BIOHAZ). EFSA J. 2010, 8, 1543. [Google Scholar] [CrossRef]
- Hochberg, N.S.; Hamer, D.H.; Hughes, J.M.; Wilson, M.E. Anisakidosis: Perils of the Deep. Clin. Infect. Dis. 2010, 51(7), 806–812. [Google Scholar] [CrossRef] [PubMed]
- Ishikura, H.; Kikuchi, K.; Nagasawa, K.; Ooiwa, T.; Takamiya, H.; Sato, N.; Sugane, K. Anisakidae and anisakidosis. Prog. Clin. Parasitol. 1993, 3, 43–102. [Google Scholar] [CrossRef]
- Wharton, D.A.; Aalders, O. The response of Anisakis larvae to freezing. J. Helminthol. 2002, 76, 363–368. [Google Scholar] [CrossRef]
- Audicana, M.T.; Ansotegui, I.J.; de Corres, L.F.; Kennedy, M.W. Anisakis simplex: dangerous – dead and alive? Trends Parasitol. 2002, 18, 20–25. [Google Scholar] [CrossRef]
- Moneo, I.; Caballero, M.L.; Gonzalez-Munoz, M.; Rodriguez-Mahillo, A.I.; Rodriguez-Perez, R.; Silva, A. Isolation of a heat-resistant allergen from the fish parasite Anisakis simplex. Parasitol. Res. 2005, 96, 285–289. [Google Scholar] [CrossRef]
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia. 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Berland, B. Identification of larval nematodes from fish. Nematode problems in North Atlantic fish, Möller, H., Eds.; Report from a workshop in Kiel, 3–4 April 1989. International Council for the Exploration of the Sea, C.M./F6:16–22; pp. 16–22.
- Fagerholm, H.P. Parasites of fish in Finland. VI. Nematodes. Acta Acadamiae Aboensis, Series B 40, 1982; pp. 1–128. ISBN 9516488498.
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Zhu, X.; D’Amelio, S.; Paggi, L.; Gasser, R.B. Assessing sequence variation in the internal transcribed spacers of ribosomal DNA within and among members of the Contracaecum osculatum complex (Nematoda: Ascaridoidea: Anisakidae). Parasitol. Res. 2000, 86, 677–683. [Google Scholar] [CrossRef]
- Zhu, X.; D’Amelio, S.; Palm, H.W.; Paggi, L.; George-Nascimento, M.; Gasser, R.B. SSCP-based identification of members within the Pseudoterranova decipiens complex (Nematoda: Ascaridoidea: Anisakidae) using genetic markers in the internal transcribed spacers of ribosomal DNA. Parasitol. 2002, 124, 615–623. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on assessment of epidemiological data in relation to the health risks resulting from the presence of parasites in wild caught fish from fishing grounds in the Baltic Sea. Panel on Biological Hazards (BIOHAZ). EFSA J. 2011, 9, 2320. [Google Scholar] [CrossRef]
- Grabda, J. The occurence of Anisakidae nematode larvae in Baltic cod (Gadus morhua callarias L.) and the dynamics of their invasion. Acta Ichthyol. Piscat. 1976, 6, 4–22. [Google Scholar] [CrossRef]
- Myjak, P.; Szostakowska, B.; Wojciechowski, J.; Pietkiewicz, H.; Rokicki, J. Anisakidae larvae in cod from the southern Baltic Sea. Arch. Fish. Mar. Res. 1994, 42, 149–161. [Google Scholar]
- Szostakowska, B.; Myjak, P.; Wyszyński, M.; Pietkiewicz, H.; Rokicki, J. Prevalence of anisakin nematodes in fish from Southern Baltic Sea. Pol. J. Microbiol. 2005, 54, 41–45. [Google Scholar] [PubMed]
- Sobecka, E. Pasożyty dorsza atlantyckiego z podgatunków Gaadus morhua morhua L. i Gadus morhua callarias L. z wybranych rejonów Atlantyku i Bałtyku (in polish). Habilitation thesis no. 245, Agricultural Academy in Szczecin, 2007.
- ICES. 2020. Working Group on Pathology and Diseases of Marine Organisms (WGPDMO). ICES Scientific Reports. 2:53. 23 pp. [CrossRef]
- Podolska, M.; Horbowy, J.; Wyszyński, M. Discrimination of Baltic herring populations with respect to Anisakis simplex larvae infection. J. Fish. Biol. 2006, 68, 1241–1256. [Google Scholar] [CrossRef]
- Rolbiecki, L.; Janc, A.; Rokicki, J. Stickleback as a potential paratenic host in the Anisakis simplex life cycle in the Baltic Sea: results of experimental infection. Wiad. Parazytol. 2001, 47, 257–262. [Google Scholar]
- Skrzypczak, M.; Rolbiecki, L. Endoparasitic helminths of the European Sprat, Sprattus sprattus (Linnaeus, 1758) from the Gulf of Gdansk (the Southern Baltic Sea) with a checklist of its parasites, Russ. J. Mar. Biol. 2015, 41, 167–175. [Google Scholar] [CrossRef]
- Nadolna-Ałtyn, K.; Kucińki, M.; Mirny, Z.; Góra, A.; Pawlak, J. Anisakidae nematodes in the liver of European flounder (Platichthys flesus) from the southern Baltic Sea. The 21st International Conference on Diseases of Fish and Shellfish (EAFP XXI), Aberdeen, Scotland, 11-14.09.2023.
- HELCOM. Baltic grey seal censuses in 2020. 15th Meeting of HELCOM Expert Group on Marine Mammals. Online, 14-16 September 2021.
- Skrzypczak, M.; Rokicki, J.; Pawliczka, I.; Najda, K.; Dzido, J. Anisakids of seals found on the southern coast of Baltic Sea. Acta Parasitol. 2014, 59, 165–172. [Google Scholar] [CrossRef]
- Herreras, M.V.; Montero, F.E.; Marcogliese, D.J.; Raga, J.A.; Balbuena, J.A. Phenotypic tradeoffs between egg number and egg size in three parasitic anisakid nematodes. Oikos 2007, 116, 1737–1747. [Google Scholar] [CrossRef]
- Measures, L.N. Effect of temperature and salinity on development and survival of eggs and free-living larvae of sealworm (Pseudoterranova decipiens). Can J Fish Aquat Sci. 1996, 53, 2804–2807. [Google Scholar] [CrossRef]
- Valtonen, E.T.; Fagerholm, H.P.; Helle, E. Contracaecum osculatum (Nematoda: Anisakidae) in fish and seals in Bothnian Bay (northeastern Baltic Sea). Int. J. Parasitol. 1988, 18, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Gabel, M.; Theisen, S.; Palm, H.W.; Dähne, M.; Unger, P. Nematode Parasites in Baltic Sea Mammals, Grey Seal (Halichoerus grypus (Fabricius, 1791)) and Harbour Porpoise (Phocoena phocoena (L.)), from the German Coast. Acta Parasitol. 2021, 66, 26–3. [Google Scholar] [CrossRef]
- Fiorenza, E.A.; Wendt, C.A; Dobkowski, K.A; King, T.L.; Pappaionou, M.; Rabinowitz, P.; Samhouri, J.S.; Wood, C.L. It’s a wormy world: Meta-analysis reveals several decades of change in the global abundance of the parasitic nematodes Anisakis spp. and Pseudoterranova spp. in marine fishes and invertebrates. Glob. Change. Biol. 2020, 26, 2854–2866. [Google Scholar] [CrossRef]
- Unger, P.; Klimpel, S.; Lang, T.; Palm, H.W. Metazoan parasites from herring (Clupea harengus L.) as biological indicators in the Baltic Sea. Acta. Parasitol. 2014, 3, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Nadolna-Ałtyn, K.; Szostakowska, B.; Pachur, M.; Bańkowska, A.; Podolska, M. First evidence of the presence of Anisakis simplex in Crangon crangon and Contracaecum osculatum in Gammarus sp. by in situ examination of the stomach contents of cod (Gadus morhua) from the southern Baltic Sea. Parasitol. 2019, 146, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Pachur, M.E.; Horbowy, J. Food composition and prey selection of cod, Gadus morhua (Actinopterygii: Gadiformes: Gadidae), in the southern Baltic Sea. Acta Ichthyol. Piscat. 2013, 43, 109–118. [Google Scholar] [CrossRef]
- Haase, K.; Orio, A.; Pawlak, J.; Pachur, M.; Casini, M. Diet of dominant demersal fish species in the Baltic Sea: Is flounder stealing benthic food from cod? Mar. Ecol. Prog. Ser. 2020, 645, 159–170. [Google Scholar] [CrossRef]
- Podolska, M.; Pawlikowski, B.; Nadolna-Ałtyn, K.; Pawlak, J.; Komar-Szymczak, K.; Szostakowska, B. How effective is freezing at killing Anisakis simplex, Pseudoterranova krabbei, and P. decipiens larvae? An experimental evaluation of timetemperature conditions. Parasitol. Res. 2019, 118, 2139–2147. [Google Scholar] [CrossRef]
- Ludovisi, A.; Di Felice, G.; Carballeda-Sangiao, N.; Barletta, B.; Butteroni, C.; Corinti, S.; Marucci, G.; González-Muñoz, M.; Pozio, E.; Gómez-Morales, M.A. Allergenic activity of Pseudoterranova decipiens (Nematoda: Anisakidae) in BALB/c mice. Parasit. Vectors. 2017, 10, 290. [Google Scholar] [CrossRef]
- Nordholm, A.; Kurtzhals, J.A.L.; Karami, A.M.; Kania, P.W.; Buchmann, K. Nasal localization of a Pseudoterranova decipiens larva in a Danish patient with suspected allergic rhinitis. Helminthol. 2020, 94, E187. [Google Scholar] [CrossRef] [PubMed]
- Strøm, S.B.; Haarder, S.; Korbut, R.; Mejer, H.; Thamsborg, S.M.; Kania, P.W.; Buchmann, K. Third-stage nematode larvae of Contracaecum osculatum from Baltic cod (Gadus morhua) elicit eosinophilic granulomatous reactions when penetrating the stomach mucosa of pigs. Parasitol. Res. 2015, 114, 1217–1220. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, A.R.; Kiani, B.; Afshari, A.; Moghaddas, E.; Williams, M.; Shamsi, S. World-wide prevalence of Anisakis larvae in fish and its relationship to human allergic anisakiasis: a systematic review. Parasitol. Res. 2020, 119, 3585–3594. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, A.R.; Moghaddas, E.; Kiani, B.; Afshari, A.; Williams, M.; Shamsi, S. Anisakis allergy: unjustified social alarm versus healthy diet; commentary to the “Letter to the Editor” of Drs Daschner, Levsen, Cipriani, and del Hoyo, referencing to “World- wide prevalence of Anisakis larvae in fish and its relationship to human allergic anisakiasis: a systematic review. Parasitol. Res. 2020, 120, 1921–1923. [Google Scholar] [CrossRef]
- Kołodziejczyk, L.; Szostakowska, B.; Sobecka, E.; Szczucki, K.; Stankiewicz, K. First case of human anisakiasis in Poland. Parasitol Int. 2020, 76, 102073. [Google Scholar] [CrossRef]
- Takei, H.; Powell, S.Z. Intestinal anisakidosis (anisakiosis). Ann. Diagn. Pathol. 2007, 11, 350–352. [Google Scholar] [CrossRef]
- Shamsi, S. Seafood-Borne Parasitic Diseases: A “One-Health” Approach Is Needed. Fishes. 2019, 4, 1–11. [Google Scholar] [CrossRef]
- Levsen, A.; Berland, B. Anisakis species. In Fish Parasites, Pathobiology and Protection; Woo, P.T.K., Buchmann, K., Eds.; CAB International, 2012; pp. 298–309.
- Beck, M.; Evans, R.; Feist, S.W.; Stebbing, P.; Longshaw, M.; Harris, E. Anisakis simplex sensu lato associated with red vent syndrome in wild Atlantic salmon Salmo salar in England and Wales. Dis. Aquat. Org. 2008, 82, 61–65. [Google Scholar] [CrossRef]
- Noguera, P.; Collins, C.; Bruno, D.; Pert, C.; Turnbull, A.; McIntosh, A.; Lester, K.; Bricknell, I.; Wallace, S.; Cook, P. Red vent syndrome in wild Atlantic salmon Salmo salar in Scotland is associated with Anisakis simplex sensu stricto (Nematoda: Anisakidae). Dis. Aquat. Org. 2009, 87, 199–215. [Google Scholar] [CrossRef]
- Haarder, S.; Kania, P.W.; Bahlool, Q.M.; Buchmann, K. Expression of immune relevant genes in rainbow trout following exposure to live Anisakis simplex larvae. Exp Parasitol. 2013, 135, 564–569. [Google Scholar] [CrossRef]
- Horbowy, J.; Podolska, M.; Nadolna-Ałtyn, K. Increasing occurrence of Anisakid nematodes in the liver of cod (Gadus morhua) from the Baltic Sea: does infection affect the condition and mortality of fish? Fish. Res. 2016, 179, 98–103. [Google Scholar] [CrossRef]
Figure 1.
Length-weight relationfish of analysed salmons (males - blue dots, females - orange dots).
Figure 1.
Length-weight relationfish of analysed salmons (males - blue dots, females - orange dots).

Figure 2.
Pseudoterranova sp. from Baltic salmon (a) whole parasite; (b) anterior part (head); (c) posterior part (tail).
Figure 2.
Pseudoterranova sp. from Baltic salmon (a) whole parasite; (b) anterior part (head); (c) posterior part (tail).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



