Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
Soy isoflavones are considered important sources of bioactive compounds, but they are poorly absorbable, due to their large hydrophilic structures. Some biotransformation strategies have been used to convert the glycosidic form into aglycones, making them available for absorption. This study evaluated the potential of enzymatic and/or microbial fermentation bioprocesses in soymilk extract before and after gastrointestinal in vitro digestion. Commercial β-glucosidase and mix of commercial probiotics containing Lactobacillus acidophilus, L. casei, Lactococcus lactis, Bifidobacterium bifidum e B. lactis were used to biotransform soymilk extract. Isoflavone profile was quantified by HPLC-DAD, total phenolic content by Folin–Ciocalteu test, and antioxidant capacity by ORAC and FRAP. Soymilk enzymatically treated (ET) followed by microbial fermentation (ET+F) resulted in the conversion of glycosylated isoflavones (6-fold lower than control for daidzin and 2-fold for genistin) to aglycones (18-fold greater than control for dadzein and genistein), besides to increase the total phenolic content (3.48 for control and 4.48 mg/ml ET+F) and to improve antioxidant capacity represented by the ORAC (120 for control and 151 mg/ml ET+F) and by the FRAP (285 for control and 317 µl/ml for ET+F) before in vitro digestion. Further, the digested ET+F samples resulted a higher content of genistein (2-fold higher than control) also an increase in the total phenolic content (2.81 for control and 4.03 mg/ml for ET+F) and antioxidant capacity by ORAC were greater compared to undigested samples. In addition, the microbial fermentation process isolated also resulted in the positive effects, but de combination of ET followed by F presented a synergistic effect, suggesting the greater potential for both bioprocesses to contribute to functional and nutritional properties on fermented soy-based products.
Keywords:
isoflavones
; soymilk
; β-glucosidase
; probiotics
; gastrointestinal digestion in vitro
1. Introduction
Soy isoflavones are a class of phytoestrogens that provide health benefits for humans [1]. However, the isoflavones present in soy products are usually found in the glycosylated form (daidzin, genistin and glycitin), presenting low availability, that is poorly absorbable by gastrointestinal tract. For isoflavones to exert their health effects, the glycosides forms can be deconjugated by β-glucosidases produced by the intestinal microbiota, forming aglycone isoflavones (daidzein, genistein and glycitein) and their secondary metabolites, which can be absorbed in the intestinal epithelial cell lines [2].
In this context, some strategies for the biotransformation of soy products in association or not with the use of probiotics, bacterial food supplements known to improve the balance of the intestinal microbiota [3], have been investigated in order to increase the bioavailability of bioactive compounds of these products. Ninety-two strains of lactic acid bacteria and bifidobacteria were analyzed for their ability to produce aglycone forms from isoflavones, being strains of the Lactobacillus, Enterococcus, Lactococcus and Bifidobacterium genera were able to produce daidzein, genistein and glycitein [4]. In a mixed culture of four different bacterial strains (Enterococcus faecium, Lactobacillus mucosae, Finegoldia magna and Veillonella sp.) conversion of dadzein to equol was observed [5], a secondary metabolite that presents superior nutraceutical effect compared to soy isoflavones [6]. During soymilk fermentation, the species Lactobacillus paraplantarum, Weissella sp., and Enterococcus faecium were effective in reducing the glycoside content and increasing the aglycone content. In this same study, about 92-100% of the daidzin and 98-100% of the genistin present in the soymilk were converted into their corresponding aglycones [7].
In addition to fermentative bioprocesses, the use of enzymes, such as β-glucosidase, has also been shown to be efficient in the hydrolysis of glycosides that are poorly absorbed by the intestine. The β-glucosidases isolated from microbial sources (Aspergillus niger) and lima beans (Phaseolus lunatus) resulted in satisfactory conversion rates of daidzin and genistin in their respective aglycone forms [8].
In the present study, we used the commercial β-glucosidase enzyme to biotransform soymilk extract in association or not with the activation of a commercial probiotic mix containing Lactobacillus acidophilus, L. casei, Lactococcus lactis, Bifidobacterium bifidum and B. lactis strains followed by in vitro gastrointestinal digestion. The same probiotic mix was used in association with a supplement rich in isoflavones in a previous prospective study with 60 menopausal women, being able to promote the growth of bacteria associated with the metabolism of isoflavones, such as Faecalibacterium prausnitzii, Roseburia faecis, in addition to Alkaliphilus crotonatoxidans and Blautia coccoide considered equol producers [9]. Thus, our objective was to determine the effects of enzymatic and/or fermentative biotransformation on the bioconversion and bioaccessibility of soy isoflavones. In addition, we also evaluated the total phenolic content and antioxidant capacity of soymilk extracts.
2. Materials and Methods
2.1. Materials
The soybeans or Glycine max (IAC Foscarin 31 variety) were obtained at the Agronomic Institute of Campinas, Campinas, SP, Brazil. Probiotic mix (Simfort® Vitafor LOT: 10515) containing Lactobacillus acidophilus, L. casei, Lactococcus lactis, Bifidobacterium bifidum and B. lactis (109 CFU each strain in 2g). The isoflavone standards (daidzin, genistin, glycitin, daidzein, genistein and glycitein), β-Glucosidase, pepsin from porcine gastric mucosa, pancreatin from porcine pancreas, bile salts, gallic acid, tannic acid, sodium alginate, 2,2′-azobis(2-methylpropionamidine) dihydrochloride (AAPH), ρ-nitrophenyl-β-D-glucopyranoside (ρ-NPG), Trolox®, and fluorescein were purchased from Sigma-Aldrich (Steinheim, Germany). Folin Ciocalteu’s reagent was purchased from Dinamica (Sao Paulo, Brazil). Methanol (99.9%, HPLC 10020 Appl Microbiol Biotechnol (2020) 104:10019–10031 grade) was obtained from J.T. Baker® (Center Valley, PA, USA). All other chemicals were obtained through commercial suppliers.
2.2. Methods
2.2.1. Processing of soymilk extracts
Preparation of soymilk extract
Soymilk extract were extracted from soybean according to previous studies [6,10]. Lyophilized soymilk extract was suspended in sterile water (200 mg/ml) and used as soymilk control (CT).
Enzymatic Biotransformation Process
Soymilk extract prepared and lyophilized was reconstituted in autoclaved water (200 mg/ml). The commercial β-Glucosidase (3.6 mg/ml of the extract) was incubated for 10 minutes at 50ºC, pH 6.5. After incubation, the samples were shaken for 45 minutes at 50ºC. Then, the reaction was stopped in an ice bath for 15 minutes and frozen for later lyophilization. These samples were called of enzymatically treated soymilk (ET).
Fermented Biotransformation Process
Commercial probiotic mix (Simfort® Vitafor LOT: 10515) containing Lactobacillus acidophilus, L. casei, Lactococcus lactis, Bifidobacterium bifidum and B. lactis (109 CFU each strain in 2g) was added to MRS Broth-0.05% cysteine (250mg Simfort in 25 ml of the culture medium) and incubated at 37ºC overnight in an anaerobic flask. Aliquot from the first incubation was added to a new inoculum (1st passage) and new passages were performed until fermentation was observed with the medium becoming cloudy. After activating the mix, 1% of the culture was inoculated in soymilk extract reconstituted and incubated at 37ºC, 50 RPM for 24 hours in an anaerobic flask. After incubation, samples were frozen and lyophilized. These samples were called fermented soymilk (F).
Enzymatic treatment followed by Fermented Bioprocess
Samples obtained after enzymatic treatment (ET) were submitted to an additional step of probiotic fermentation as described on the “Fermented Biotransformation Process” section. These samples were called enzymatically treated and fermented soymilk (ET+F). All reactions were performed in triplicate. As control (CT), soymilk (without microorganism and enzyme addition) was submitted to the same conditions of both enzymatic and fermented biotransformation.
2.2.2. In vitro digestion assay
Samples from each experimental condition (CT, ET, F and ET+F) were subjected to in vitro digestion assay according to Minekus et al. [11]. For that, samples were reconstituted in autoclaved water (200 mg/ml) and submitted to a simulation of digestion of the oral, stomach and small intestine stages, using their respective digestive fluids, as well as pH adjustment, temperature and well time in each compartment, as shown in Figure 1. Subsequently, the samples were lyophilized and frozen for later use.
2.2.3. Analysis of isoflavone metabolites
Aliquots of samples (CT, ET, F and ET+F) obtained before and after in vitro digestion were centrifuged for 10 min at 13000g and the fractions obtained were stored at 20°C for quantification of isoflavone metabolites. For this, the aliquots were mixed with 5 ml of methanol (80%) at 25°C for 2 minutes at 500 rpm. The mixture was centrifuged at 5000 rpm for 15 minutes and the supernatant was filtered through a 0.45μm membrane before being analyzed by High Performance Liquid Chromatography (HPLC). Analyzes were performed on a Dionex UltiMate 3000 Liquid Chromatograph (Germany), equipped with an RP18 XTerra® column (Waters, 5 μm, 4.6 × 150 mm) maintained at 30°C. Detection was performed using a UV/VIS detector (DAD-3000) and the mobile phases: A (water/formic acid, 99.9: 0.1 v/v) and B (methanol). Gradient elution was: 20% B (0-15 min), 20-80% B (15-75 min), 80-100% B (75-80 min), 100-20% B (80-90 min) and 20% B (90−95 min), with a flow rate of 0.5 ml min-1. Spectra were recorded at 190 and 480 nm, and chromatograms processed at 254nm. Isoflavonoids were identified by comparing their retention time and absorbance spectrum. The quantification of individual bioactive compounds was performed using standard calibration curves (daidzin, genistin, glycitin, daidzein, genistein and glycitein). Results were expressed as μg of each isoflavonoid compound/mL of sample.
2.2.4. Analysis of Total Phenolics
The total phenolic content (TPC) of the extracts was measured according to the Folin-Ciocalteu spectrophotometric method as described previously [10]. For that, CT, ET, F and ET+F samples were analyzed in triplicate and the results were calculated using a calibration curve constructed using gallic acid standard (between 300 and 16 μg/mL). The reaction was determined at 725 nm using a fluorimeter (DU 640, Beckman Coulter, Pasadena, United States). The results were expressed as mg of gallic acid equivalents (GAE)/ml of reconstituted sample.
2.2.5. Analysis of Antioxidant Capacity
The analysis of antioxidant capacity was determined by the ORAC and FRAP assays, as previously described [10]. For this purpose, Trolox was used as a standard in the construction of the calibration curve at a concentration of 1500–1.5 μmol/mL for ORAC and 1500–15 μmol/mL for FRAP, and the results were expressed in μmol Trolox equivalents (TE) /ml of reconstituted sample triplicate (CT, ET, F and ET+F). For the ORAC assay, results were calculated as the difference between the area under the fluorescein decay curve of the samples and the blank (net AUC), using regression equations between the net AUC and the concentration of the samples. Fluorescence was monitored using a fluorimeter (DU 640, Beckman Coulter, Pasadena, USA) at 37°C for 75 min with a 485 nm excitation filter and 520 nm emission filter. For the FRAP assay, the fluorimeter was set to 37°C and the absorbance of the mixture was measured at 595 nm in kinetic reading for 10 minutes (7 cycles of 88 seconds).
2.2.6. Statistical analysis
Results were expressed as the mean ± standard error of the mean (SEM) in triplicate. Statistically significant differences were determined using analysis of variance (ANOVA), followed by Tukey's post-hoc test for multiple comparisons using GraphPad Instat (GraphPad Software, Inc., La Jolla, CA, USA). Note that p<0.05 were considered significant for all analyses.
3. Results and Discussion
3.1. Profiling of bioactive isoflavones
The isoflavones profile of the enzymatic, fermented and enzymatic/fermented bioprocesses was evaluated by HPLC-DAD, as shown in Figure 2. Soymilk extract treated with β-glucosidase followed by fermentation with probiotics or only fermented soymilk resulted in the conversion of glycosylated isoflavones to aglycones before in vitro digestion, as evidenced by the lower daidzin content (from 30.78 of the soymilk control to 4.91 µg/ml of enzymatically treated/fermented soymilk; Figure 2A), genistin (from 28.86 of soymilk control to 11.50 µg/ml of fermented soymilk; Figure 2C) and glycitin (from 6.30 of soymilk control to 3.32 of fermented soymilk and 2.78 µg/ml of enzymatically treated/fermented soymilk; Figure 2E), and higher content of daidzein (from 2.42 of soymilk control to 26.41 of fermented soymilk and 44.82 µg/ml of enzymatically treated/fermented soymilk; Figure 2B) and genistein (from 2.03 of soymilk control to 9.52 of enzymatically treated soymilk; 20.94 of fermented soymilk and 37.23 µg/ml of enzymatically treated/fermented soymilk; Figure 2D) compared to soymilk control (no enzyme and/or probiotics).
The isolated enzymatic bioprocess was not enough to convert the glycosylated isoflavones into aglycones compared to the soymilk control, but it was able to significantly increase the genistein content by 4-fold compared to the soymilk control. In previous studies of our research group, the enzyme β-glucosidase was also used to ferment soymilk, resulting in the conversion of glycosidic isoflavones into aglicones, but the enzyme was used immobilized form and the enzymatic process was optimizated previously [12,13]. Moreover, the association of enzymatically treated and fermented soymilk resulted in an 18-fold increase for dadzein and genistein compared to soymilk control, indicating a synergistic effect between enzymatic and fermentative bioprocess, whereas only fermented soymilk resulted in an 11- and 10-fold increase for dadzein and genistein, respectively.
It is known that the stability and release of isoflavones can be altered by the gastrointestinal digestion process due to the acidic pH of the stomach and the chemical composition of the digestive fluids [14]. In this context, we investigated the bioaccessibility of compounds present in soymilk extract and their treatments by simulating in vitro gastrointestinal digestion at oral, gastric and intestinal levels. After in vitro digestion, a higher content of daidzein (from 5.13 of soymilk control to 15.0 µg/ml of fermented soymilk, 3-fold higher than the soymilk control; Figure 2B) and genistein (from 3.82 of soymilk control for 13.74 of fermented soymilk and 9.09 µg/ml of enzymatically treated and fermented soymilk, 3- and 2-fold higher than the soymilk control, respectively; Figure 2D) were observed compared to soymilk control.
The chromatograms of soymilk extracts obtained by HPLC-DAD are represented in Figure 3A-H, demonstrating that each form of isoflavone increased/decreased to different degrees depending on the treatment method employed before and after in vitro gastrointestinal digestion. In the soymilk control, the highest daidzin, genistin and glycitein peaks, and the lowest dadzein and genistein peaks were observed before in vitro digestion (Figure 3A). The same was not observed after in vitro digestion, in which we obtained peaks of genistin, glycitein and dadzein (Figure 3B). In enzymatically treated soymilk, genistin and glycitin peaks were observed, in addition to a lower daidzin and higher dadzein peak before and after in vitro digestion (Figure 3 C-D). In fermented soymilk, peaks of genistin and dadzein were observed before digestion, and peaks of genistin, glycitein and dadzein after in vitro digestion (Figure 3 E-F). In the enzymatically treated and fermented soymilk, peaks of glycosidic isoflavones were not observed, but peaks of genistein and daidzein appeared before in vitro digestion, and after digestion, only daidzein peak was observed (Figure 3 G-H). These data indicated that only enzymatically treated soymilk was not able to convert glycosidic isoflavones into aglycones, but the association of enzymatic and fermentative bioprocesses or only fermentative bioprocess with probiotics were efficient in converting isoflavones, considering the highest peaks of aglycone isoflavones observed. Moreover, the presence of aglycones after the in vitro digestion simulation indicates that these isoflavones would be available for absorption. In the previous study by Queirós et al. [15] , in which the enzymatic extraction of soy flour extracts using different enzymes (protease, tannase and cellulase) or their combination resulted in a significant increase in aglycones after in vitro gastrointestinal digestion, except that produced by the use of protease alone. Another study using kefir-fermented soymilk also resulted in increased aglycone isoflavone content after in vitro gastrointestinal digestion [16].
Considering β-glucosidase is an enzyme involved in the hydrolysis of the β-glycosidic bond present in the glycosidic isoflavone, releasing the bioactive forms of the aglycones [17,18], and as mentioned before, although enzymatically treated soymilk with commercial β-glucosidase was not able to efficiently convert the glycosidic forms into aglycones, the results obtained in this study may be associated with the β-glucosidase activity of the probiotic microorganisms used to carry out the soymilk fermentation process.
Previous studies have shown that the use of probiotic strains are involved in the ability to produce aglycone forms from glycosidic isoflavones. Previous studies of our research group have investigated some fermentative strategies of soymilk extract using different bacterial strains to ferment soymilk extract, such as Streptococcus thermophilus, Lactobacillus delbrueckii ssp. bulgaricus, Bifidobacterium animalis ssp. lactis and Lactobacillus acidophilus [6], B. animalis ssp. lactis, L. acidophilus, S. thermophilus, L. casei, L. delbrueckii ssp. Bulgaricus, L. rhamnosus e L. kefiri. [19], besides B. lactis and L. casei [10], resulting a significant conversion of glycosylated isoflavones to aglycones. All the studies mentioned, the combination of fermentative and enzymatic bioprocesses using the enzyme tannase or β-glucosidase resulted in a superior synergistic effect compared to bioprocesses alone. Other studies with soy-based foods used some bacterial cultures, such as Lactobacillus delbrueckii ssp. bulgaricus, L. acidophilus, L. plantarum, L. casei, L. fermentum, S. thermophilus and several species of Bifidobacterium were also responsible for increasing the content of aglycone isoflavones [20,21,22].
3.2. Phenolic compounds and antioxidant activity of soymilk extracts
Soy isoflavones are considered an important class of antioxidant compounds, with the aglycone forms and their secondary metabolites having greater antioxidant potential compared to other isoflavonoids [23]. The role of antioxidant compounds presents in food or in the body is to delay, control and prevent oxidative processes that lead to food deterioration and the spread of degenerative diseases in the body. Therefore, measuring the antioxidant capacity of foods is essential not only to ensure the quality of functional foods but mainly to study the efficiency of food antioxidants in the prevention and treatment of diseases related to oxidative stress [24].
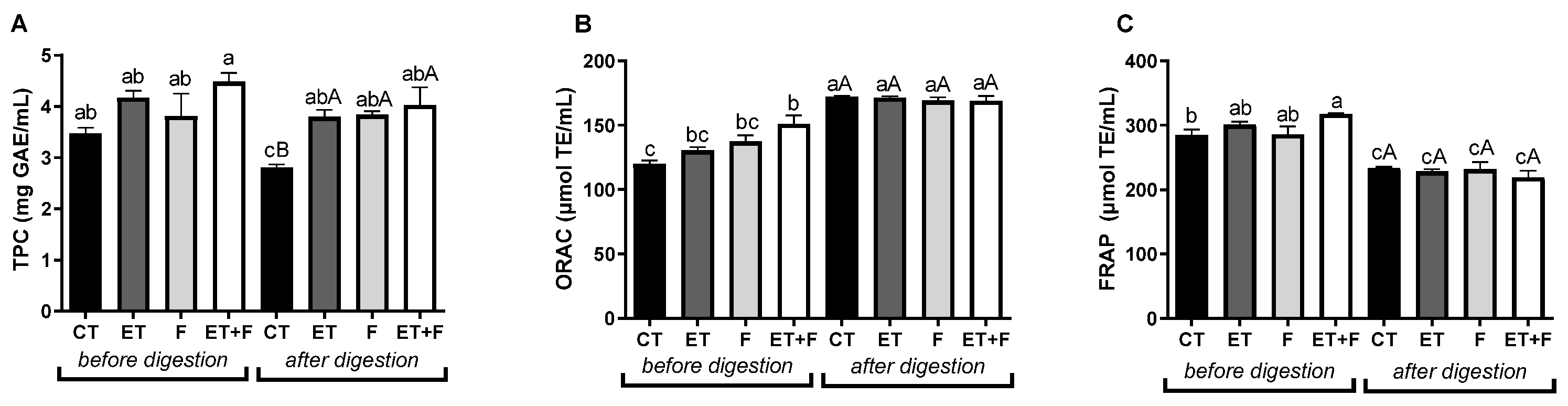
In this study, the total phenolic content using the Folin–Ciocalteu test, as well as the antioxidant capacity using the ORAC and FRAP assays were evaluated in enzymatic bioprocess with β-glucosidase and/or fermented with probiotics, as shown in Figure 4A-C. The results presented indicated that enzymatically treated soymilk followed by fermented soymilk had a higher content of total phenolics before in vitro digestion (from 3.48 of soymilk control and 4.48 mg/ml of enzymatically treated and fermented soymilk; Figure 4A). The samples that were submitted to the in vitro digestion assay resulted in an increase in the total phenolic content for all bioprocesses (from 2.81 for the soymilk control to 3.80 for the enzymatically treated soymilk; 3.84 for the fermented soymilk and 4, 03 mg/ml of enzymatically treated and fermented soymilk; Figure 4A). Moreover, we observed a reduction in total phenolic content for the soymilk control after the digestion process, which resulted in the highest significant values (p<0.05) for all soymilk extracts before and after digestion, as shown in Figure 4A.
Enzymatically treated soymilk with β-glucosidase followed by fermentation with probiotics also resulted in the highest antioxidant capacity represented by the ORAC assay (from 120 of the soymilk control to 151 mg/ml of the enzymatically treated and fermented soymilk; Figure 4B) and by the FRAP assay (from 285 of the soymilk control for 317 µl/ml of enzymatically treated and fermented soymilk; Figure 4C) compared to soymilk control before in vitro digestion. However, we did not observe significant changes between groups after the in vitro digestion assay, but we did observe an increase in antioxidant capacity by the ORAC assay and a reduction by the FRAP assay for all soymilk samples after digestion compared to the undigested samples (Figure 4B-C). On the other hand, in the study by Queirós et al. [15] the concentration of total phenolic compounds and the antioxidant capacity were higher after in vitro gastrointestinal digestion compared to undigested samples. Such differences can be explained by the different specificities of enzyme action, since different enzymes were used between the current studies and the study mentioned above, as well as variations in processing conditions, which may change the rate of absorption and metabolism of isoflavones and consequently, in their biological activities [25].
Additionally, the activation of the probiotic bacteria used in this study proved to be efficient not only in converting the glycosylated forms of isoflavones into aglycones, but also in improving the bioaccessibility of these aglycones and antioxidant capacity. Our data corroborate previous studies, in which fermented soy products showed greater antioxidant capacity than non-fermented ones [26,27].
Previous studies by our research group mentioned above demonstrated that the enzymatic and/or fermentative bioprocesses used associated the conversion of glycosidic isoflavones into aglycones with a higher content of total phenolics and the antioxidant capacity of soymilk extract [6,19] and soy flour extract [15]. In the first study, all bioprocesses tested resulted in a significant increase in total phenolic content and antioxidant capacity using the ORAC and DPPH assays, with the best results being observed in soymilk that were subjected to biotransformation with the tannase enzyme [6]. In the second study, the combination of microbial fermentation and tannase biotransformation also resulted in increased total phenolic content and antioxidant activity using the ORAC and FRAP assays [19]. The antioxidant capacity of soymilk using Lactobacillus fermentation; of enzymatic biotransformation with tannase and enzymatic biotransformation followed by fermentation was also evaluated previously, resulting in increased antioxidant capacity using ORAC and FRAP assays for all bioprocesses, with superior results being observed for enzymatic biotransformation followed by fermentation [10]. Finally, in the soybean flour extract, all the bioprocesses used resulted in an increase in the release of total phenolics and in the antioxidant capacity, measured by the scavenging of the ABTS+ radical, with the exception of the cellulase enzyme in isolation [15].
4. Conclusions
In summary, the microbial fermentation processes as well as the enzymatic biotransformation followed by the microbial fermentation with probiotics proposed in this study were effective in the bioconversion of glycosidic isoflavones into aglycone forms in soymilk extract, which are biologically more active, resulting in a higher content of total phenolics and antioxidant capacity before and after in vitro gastrointestinal digestion. Moreover, the presence of aglycones after in vitro digestion indicates that these compounds would be bioaccessible for absorption. In this way, we can suggest that the fermented soymilk, and especially the enzymatically treated soymilk with β-glucosidase followed by the microbial fermentation with probiotics, has the potential to exert functional and nutritional properties on fermented soy-based products.
Author Contributions
Conceptualization, G.M and C.C. and P.B.; methodology, C.C.; A.G. and G.M.; software, C.C.; P.B..; validation, G.M., J.M. and A.G.; formal analysis, G.M.; investigation, C.C.; resources,G.M.; data curation, G.M; writing—original draft preparation, C.C.; writing—review and editing, M.M.; visualization, J.M.; supervision, G.M.; project administration, G.M. and J.M.; funding acquisition, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP, Brazil, (2019/10325-0).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ningtyas, D.W.; Hati, S.; Prakash, S. Bioconversion and Bioaccessibility of Isoflavones from Sogurt during in Vitro Digestion. Food Chem. 2021, 343, 128553. [CrossRef]
- Leonard, L.M.; Choi, M.S.; Cross, T.W.L. Maximizing the Estrogenic Potential of Soy Isoflavones Through the Gut Microbiome: Implication for Cardiometabolic Health in Postmenopausal Women. Nutrients 2022, 14. [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [CrossRef]
- Gaya, P.; Peirotén, Á.; Medina, M.; Landete, J.M. Isoflavone Metabolism by a Collection of Lactic Acid Bacteria and Bifidobacteria with Biotechnological Interest. Int. J. Food Sci. Nutr. 2016, 67, 117–124. [CrossRef]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and Characterisation of an Equol-Producing Mixed Microbial Culture from a Human Faecal Sample and Its Activity under Gastrointestinal Conditions. Arch. Microbiol. 2005, 183, 45–55. [CrossRef]
- de Queirós, L.D.; Macedo, J.A.; Macedo, G.A. A New Biotechnological Process to Enhance the Soymilk Bioactivity. Food Sci. Biotechnol. 2016, 25, 763–770. [CrossRef]
- Chun, J.; Jeong, W.J.; Kim, J.-S.; Lim, J.; Park, C.-S.; Kwon, D.Y.; Choi, I.; Kim, J.H. Hydrolysis of Isoflavone Glucosides in Soymilk Fermented with Single or Mixed Cultures of Lactobacillus Paraplantarum KM, Weissella Sp. 33, and Enterococcus Faecium 35 Isolated from Humans. J. Microbiol. Biotechnol. 2008, 18, 573–578.
- Fujita, A.; Alencar, S.M.; Park, Y.K. Conversion of Isoflavone Glucosides to Aglycones by Partially Purified β-Glucosidases from Microbial and Vegetable Sources. Appl. Biochem. Biotechnol. 2015, 176, 1659–1672. [CrossRef]
- Ribeiro, A.E.; Monteiro, N.E.S.; Moraes, A.V.G. De; Costa-Paiva, L.H.; Pedro, A.O. Can the Use of Probiotics in Association with Isoflavone Improve the Symptoms of Genitourinary Syndrome of Menopause? Results from a Randomized Controlled Trial. Menopause 2019, 26, 643–652. [CrossRef]
- Genova, V.M.; Fernandes, A.C.F.; Hiramatsu, É.; Queirós, L.D.; Macedo, J.A.; Macedo, G.A. Biotransformed Antioxidant Isoflavone Extracts Present High-Capacity to Attenuate the in Vitro Formation of Advanced Glycation End Products. Food Biotechnol. 2021, 35, 50–66. [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Static in Vitro Digestion Method Suitable for Food-an International Consensus. Food Funct. 2014, 5, 1113–1124. [CrossRef]
- de Ávila, A.R.A.; de Queirós, L.D.; Ueta, T.M.; Macedo, G.A.; Macedo, J.A. Exploring in Vitro Effects of Biotransformed Isoflavones Extracts: Antioxidant, Antiinflammatory, and Antilipogenic. J. Food Biochem. 2019, 43, 1–9. [CrossRef]
- de Ávila, A.R.A.; de Queirós, L.D.; Lopes, D.B.; Barin, C.G.; Ueta, T.M.; Ruiz, A.L.T.G.; Macedo, G.A.; Macedo, J.A. Enhanced Estrogenic Effects of Biotransformed Soy Extracts. J. Funct. Foods 2018, 48, 117–124. [CrossRef]
- Williams, C.F.; Walton, G.E.; Jiang, L.; Plummer, S.; Garaiova, I.; Gibson, G.R. Comparative Analysis of Intestinal Tract Models. Annu. Rev. Food Sci. Technol. 2015, 6, 329–350. [CrossRef]
- de Queirós, L.D.; Dias, F.F.G.; de Ávila, A.R.A.; Macedo, J.A.; Macedo, G.A.; Leite Nobrega de Moura Bell, J.M. Effects of Enzyme-Assisted Extraction on the Profile and Bioaccessibility of Isoflavones from Soybean Flour. Food Res. Int. 2021, 147. [CrossRef]
- da Silva Fernandes, M.; Sanches Lima, F.; Rodrigues, D.; Handa, C.; Guelfi, M.; Garcia, S.; Ida, E.I. Evaluation of the Isoflavone and Total Phenolic Contents of Kefir-Fermented Soymilk Storage and after the in Vitro Digestive System Simulation. Food Chem. 2017, 229, 373–380. [CrossRef]
- Baú, T.R.; Ida, E.I. Soymilk Processing with Higher Isoflavone Aglycone Content. Food Chem. 2015, 183, 161–168. [CrossRef]
- Nakai, S.; Fujita, M.; Kamei, Y. Health Promotion Effects of Soy Isoflavones. J. Nutr. Sci. Vitaminol. (Tokyo). 2020, 66, 502–507. [CrossRef]
- de Queirós, L.D.; de Ávila, A.R.A.; Botaro, A.V.; Chirotto, D.B.L.; Macedo, J.A.; Macedo, G.A. Combined Isoflavones Biotransformation Increases the Bioactive and Antioxidant Capacity of Soymilk. Appl. Microbiol. Biotechnol. 2020, 104, 10019–10031. [CrossRef]
- Tsangalis, D.; Ashton, J.F.; Mcgill, A.E.J.; Shah, N.P. Enzymic Transformation of Isoflavone Phytoestrogens in Soymilk by β-Glucosidase-Producing Bifidobacteria. J. Food Sci. 2002, 67, 3104–3113. [CrossRef]
- Donkor, O.N.; Shah, N.P. Production of β-Glucosidase and Hydrolysis of Isoflavone Phytoestrogens by Lactobacillus Acidophilus, Bifidobacterium Lactis, and Lactobacillus Casei in Soymilk. J. Food Sci. 2008, 73, 15–20. [CrossRef]
- Rekha, C.R.; Vijayalakshmi, G. Isoflavone Phytoestrogens in Soymilk Fermented with β-Glucosidase Producing Probiotic Lactic Acid Bacteria. Int. J. Food Sci. Nutr. 2011, 62, 111–120. [CrossRef]
- Hazra, B.; Biswas, S.; Mandal, N. Antioxidant and Free Radical Scavenging Activity of Spondias Pinnata. BMC Complement. Altern. Med. 2008, 8, 1–10. [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22. [CrossRef]
- Yu, X.; Meenu, M.; Xu, B.; Yu, H. Impact of Processing Technologies on Isoflavones, Phenolic Acids, and Antioxidant Capacities of Soymilk Prepared from 15 Soybean Varieties. Food Chem. 2021, 345, 128612. [CrossRef]
- Niyibituronsa, M.; Onyango, A.N.; Gaidashova, S.; Imathiu, S.; Boevre, M. De; Leenknecht, D.; Neirnck, E.; Saeger, S. De; Vermeir, P.; Raes, K. The Growth of Different Probiotic Microorganisms in Soymilk from Different Soybean Varieties and Their Effects on Anti-Oxidant Activity and Oligosaccharide Content. J. Food Res. 2019, 8, 41. [CrossRef]
- Król-Grzymała, A.; Amarowicz, R. Phenolic Compounds of Soybean Seeds from Two European Countries and Their Antioxidant Properties. Molecules 2020, 25, 1–11. [CrossRef]
Figure 1.
Methodological scheme of stages of gastrointestinal in vitro digestion of soymilk extracts during oral, gastric and intestinal phases.
Figure 1.
Methodological scheme of stages of gastrointestinal in vitro digestion of soymilk extracts during oral, gastric and intestinal phases.
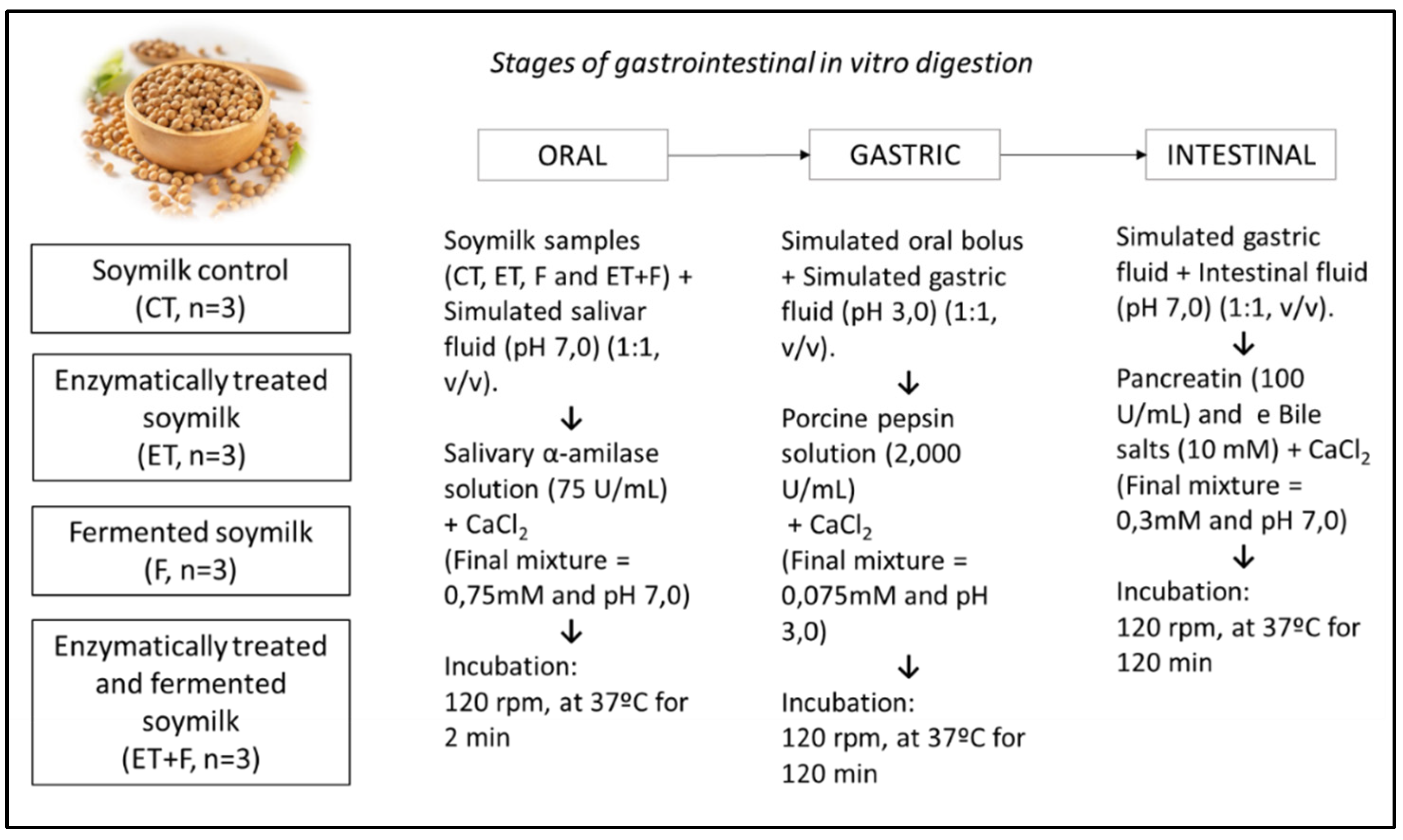
Figure 2.
Concentration of soymilk isoflavones before and after gastrointestinal in vitro digestion by LC-MS/MS. Abbreviations: CT, soymilk control; ET, enzymatically treated soymilk; F, fermented soymilk; ET+F, enzymatically treated and fermented soymilk. Values are presented as µg of sample/ml of extract and mean ± standard deviation (n = 3). a,b,c,d Different lowercase letters indicates a significant difference between bioprocesses methods before and after in vitro digestion. A,B Different capital letters indicates a significant difference between bioprocesses methods after in vitro digestion, as indicated by the Tukey’s Test (p < 0.05).
Figure 2.
Concentration of soymilk isoflavones before and after gastrointestinal in vitro digestion by LC-MS/MS. Abbreviations: CT, soymilk control; ET, enzymatically treated soymilk; F, fermented soymilk; ET+F, enzymatically treated and fermented soymilk. Values are presented as µg of sample/ml of extract and mean ± standard deviation (n = 3). a,b,c,d Different lowercase letters indicates a significant difference between bioprocesses methods before and after in vitro digestion. A,B Different capital letters indicates a significant difference between bioprocesses methods after in vitro digestion, as indicated by the Tukey’s Test (p < 0.05).
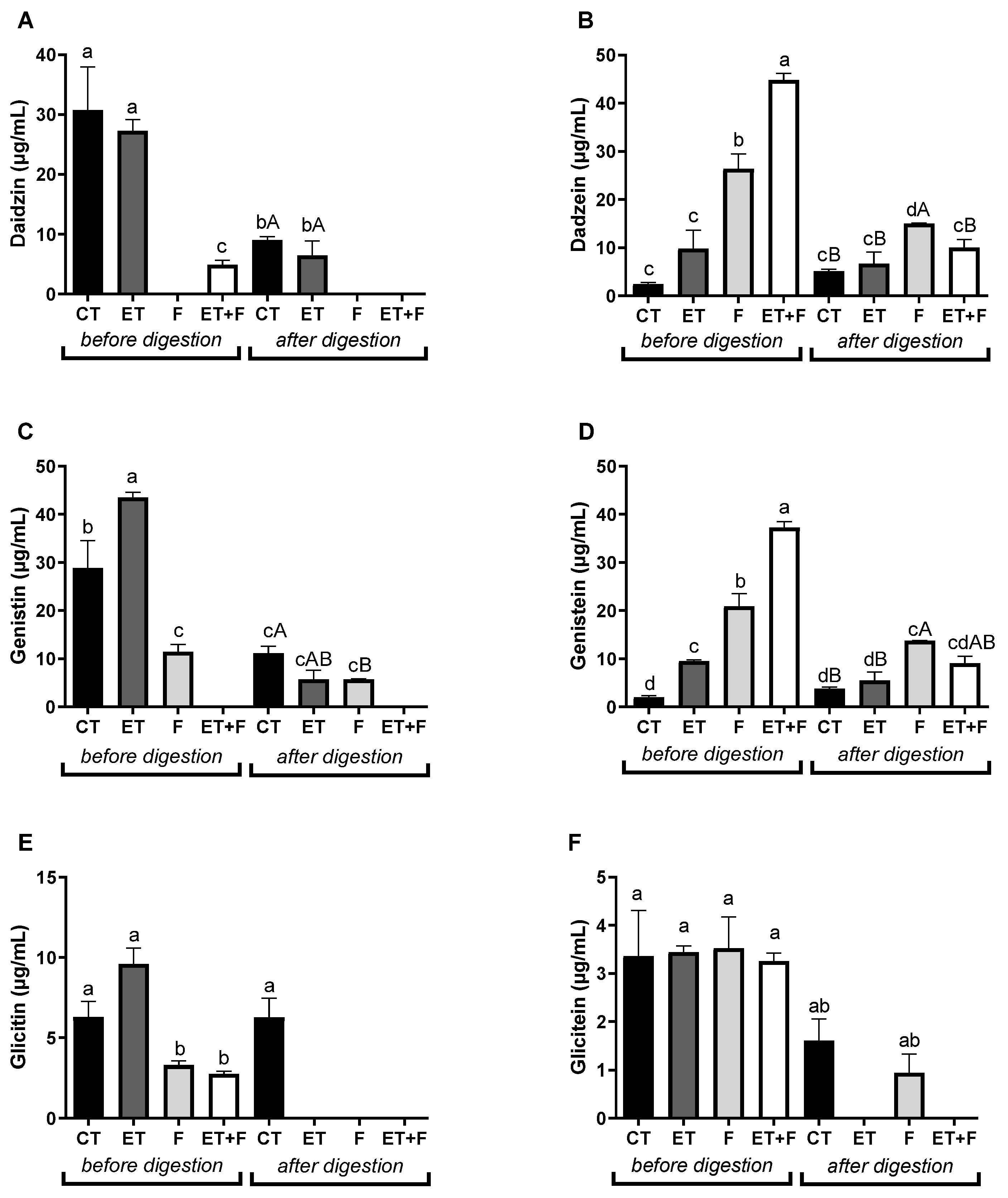
Figure 3.
Chromatograms obtained by HPLC-DAD before and after gastrointestinal in vitro digestion by LC-MS/MS from soymilk control (CT; A and B); enzymatically treated soymilk (ET; C and D); fermented soymilk (F; E and F); enzymatically treated and fermented soymilk (ET+F; G and H). The numbers represent the respective isoflavones: daidzin (1), glycitin (2), genistin (3), glycitein (4), daidzein (5) and genistein (6).
Figure 3.
Chromatograms obtained by HPLC-DAD before and after gastrointestinal in vitro digestion by LC-MS/MS from soymilk control (CT; A and B); enzymatically treated soymilk (ET; C and D); fermented soymilk (F; E and F); enzymatically treated and fermented soymilk (ET+F; G and H). The numbers represent the respective isoflavones: daidzin (1), glycitin (2), genistin (3), glycitein (4), daidzein (5) and genistein (6).
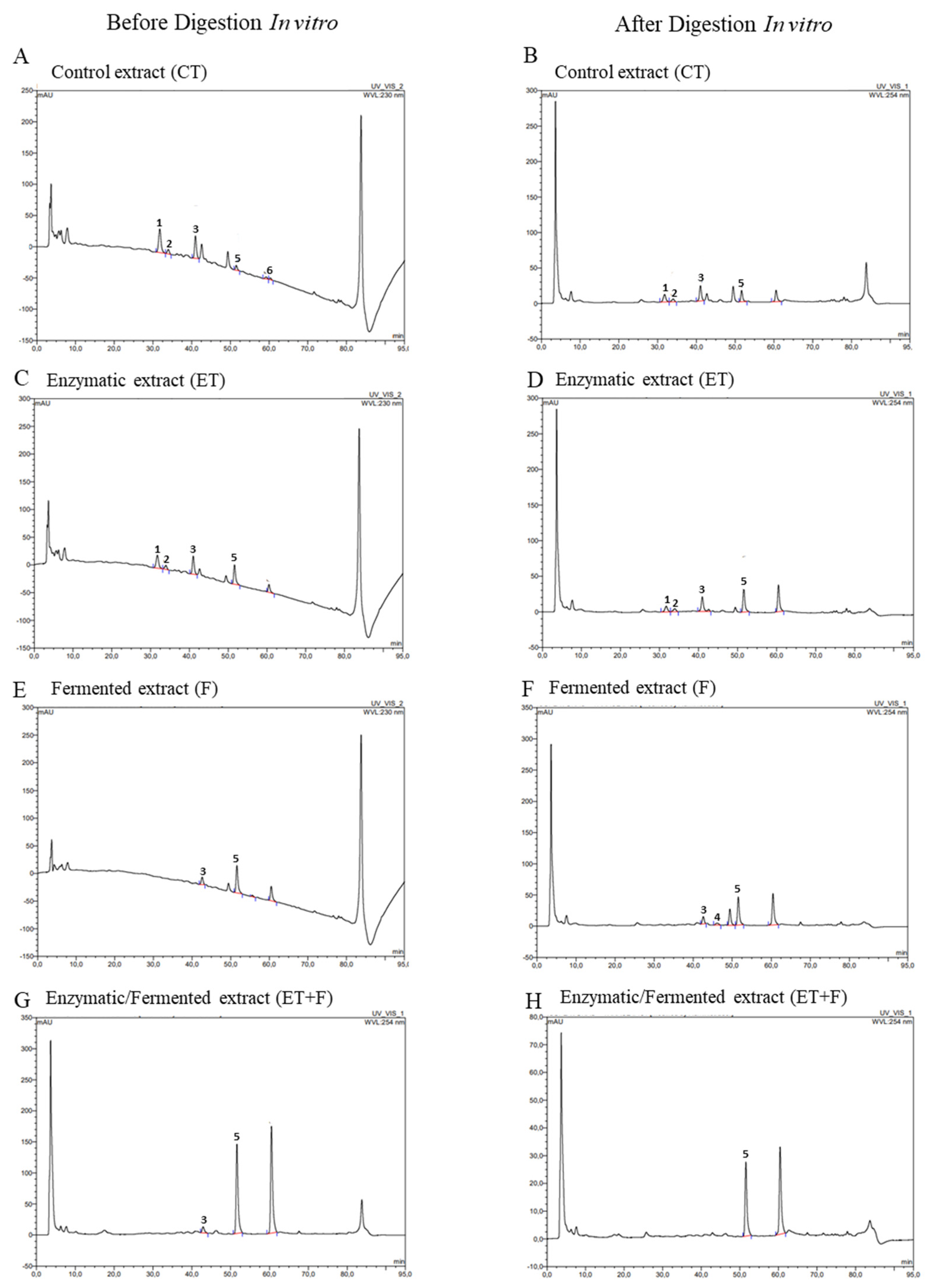
Figure 4.
Total phenolic content (A) and antioxidante capacity (B and C) before and after gastrointestinal in vitro digestion. Abbreviations: TPC, total phenolic content; GAE, acid gallic; TE, Trolox; CT, soymilk control; ET, enzymatically treated soymilk; F, fermented soymilk; ET+F, enzymatically treated and fermented soymilk. Values are presented as mg of sample/ml of extract (A) and µmol TE/ml (B and C) and mean ± standard deviation (n = 3). a,b,c Different lowercase letters indicates a significant difference between bioprocesses methods before and after in vitro digestion. A,B Different capital letters indicate a significant difference between bioprocesses methods after in vitro digestion, as indicated by the Tukey’s Test (p < 0.05).
Figure 4.
Total phenolic content (A) and antioxidante capacity (B and C) before and after gastrointestinal in vitro digestion. Abbreviations: TPC, total phenolic content; GAE, acid gallic; TE, Trolox; CT, soymilk control; ET, enzymatically treated soymilk; F, fermented soymilk; ET+F, enzymatically treated and fermented soymilk. Values are presented as mg of sample/ml of extract (A) and µmol TE/ml (B and C) and mean ± standard deviation (n = 3). a,b,c Different lowercase letters indicates a significant difference between bioprocesses methods before and after in vitro digestion. A,B Different capital letters indicate a significant difference between bioprocesses methods after in vitro digestion, as indicated by the Tukey’s Test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



