
Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
The purpose of this study was to examine the impact of fructose consumption during the peri-pubertal period on the later onset of metabolic syndrome in rats. Additionally, the study sought to explore the potential of moderate-intensity physical training as a non-pharmacological approach to mitigate the fructose effects. Male rats, 30 days old, received 10% fructose, and were submitted to concomitant moderate-intensity training until 60 days old. They were divided into 4 groups: sedentary control (SC), sedentary fructose (SF), trained control (TC), and trained fructose (TF). Fructose consumption mainly affected adult animals, resulting in glucose intolerance, increased periepididymal fat, and increased total cholesterol, triglycerides, and insulin levels. Furthermore, these rats exhibited reduced sympathetic nerve activity. The moderate running led to a decrease in periepididymal and retroperitoneal adipose tissue, as well as a reduction in total cholesterol and improved glucose tolerance. Peri-pubertal rats that received combination treatments exhibited lower levels of glucose and insulin during an intraperitoneal glucose tolerance test. The results indicate that 10% fructose supplementation during peripuberty predisposes to metabolic syndrome in adulthood. However, simultaneous moderate-intensity exercise training attenuates these effects and induces positive changes in glycemic homeostasis, lipid metabolism, and autonomic nervous system activity.
Keywords:
aerobic exercise training
; metabolic programming
; DOHaD
; adolescence
1. Introduction
Metabolic disorders such as insulin resistance, hyperglycaemia, hypertension, dyslipidemia and visceral obesity, are the set of clinical markers of the Metabolic Syndrome (MetS) [1]. Increased consumption of highly processed foods, including those rich in fructose, has contributed to an epidemic incidence of MetS in the last decades [2], and now is a global problem [3]. Although present in fruits in small amounts, the large quantity of fructose in the diet comes from processed foods, largely, sweetened beverages as soft drinks [3]. The consumption of high fructose corn syrup (HFCS) is known for its role as a source of dietary fructose [4].
Consumption of sweetened beverages and foods has increased in all age groups, with adolescents as their main consumers [4,5,6]. Several studies have shown that stressors, mainly dietary factors, in the early stages of life, such as preconception, gestation, lactation and peri-pubertal period, can trigger the development of MetS in adult life [7,8]. This hypothesis is part of the DOHaD (Developmental Origins of Health and Disease) concept, which postulates that early insults can program for metabolic disorders in adult life [9,10,11]. Current investigations already show that peri-pubertal period is a susceptible period that lead to metabolic programming [12,13], which can be induced by increased ingestion of high caloric drinks and foods.
Ingestion of fructose has been shown to cause hyperglyacemia and hyperinsulinemia [14,15], insulin resistance [16,17,18,19], as well as cardiovascular disorders [20]. Fructose is first captured and metabolized by the liver, where it will act in the formation of lipogenic precursors, such as dihydroxyacetone phosphate (DHAP), and Acetyl-CoA [21], leading to hepatic fat deposition, dyslipidemia and in hepatic insulin resistance [17]. Moreover, animals supplemented with 10% fructose for two weeks exhibited hepatomegaly, moderate steatosis, disorganized liver histoarquitecture, altered triacylglycerol (TAG) synthesis and ectopic lipid accumulation [22].
The promising effects of physical exercise on the prevention and treatment of metabolic disorders have been verified by many studies that point out its importance in weight loss, fat reduction and in the improvement of insulin sensitivity [23,24]. The moderate-intensity training exercise has been showed to be protective against development of the MetS [25,26]. Evidences from both human and animal model studies suggests that physical exercise performed at different ages facilitates neuroplasticity of certain brain structures, and some of these protective effects may be related to an attenuation of risk factors associated with MetS [27]. Our laboratory recently demonstrated that the moderate intensity running when performed during adolescence is able to decrease the Walker-256 tumor growth and metastasis in adult male rats, due to improvement in glucose–insulin homeostasis and beneficial effects in biometric and metabolic markers [28,29].
The experimental model used in the present investigation is reported in the literature as effective in producing MetS signs, mainly in adult rats [16]. However, the long-lasting effect of peri-pubertal fructose intake concomitant with physical exercise on adult metabolism remains unexplored, and also the probably effect of metabolic programming in this model is original and deserves to be elucidated. Therefore, the present study aims to investigate: (I) the effects of fructose intake during peri-pubertal period in the metabolism in adulthood and, (II) if the short-term moderate-intensity training performed at peri-pubertal period, concomitant with fructose intake, could protect the metabolism against MetS development in adult life.
2. Results
2.1. Oximetry and treadmill parameters
Table 1 shows that the trained fructose (TF) rats demonstrated significant increases in maximal oxygen uptake (VO2max) and energy expenditure (EE) (P= 0.0007) compared to the trained control (TC) rats at the end of exercise protocol or at 60 days-old. The FWL (P= 0.018) was greater in the TF rats compared to the sedentary fructose (SF) rats, and also greater in the TC rats compared to correspondent sedentary (P= 0.0240). On the other hand, the respiratory quotient/coefficient (RQ) (P= 0.009) was lower in the TF rats compared to the SF rats. No differences were observed in oximetry and performance parameters at 120 days-old (Table 2), indicating homogeneity of the experimental groups even after 60 days of cessation of the exercise protocol.
2.2. Biometric parameters
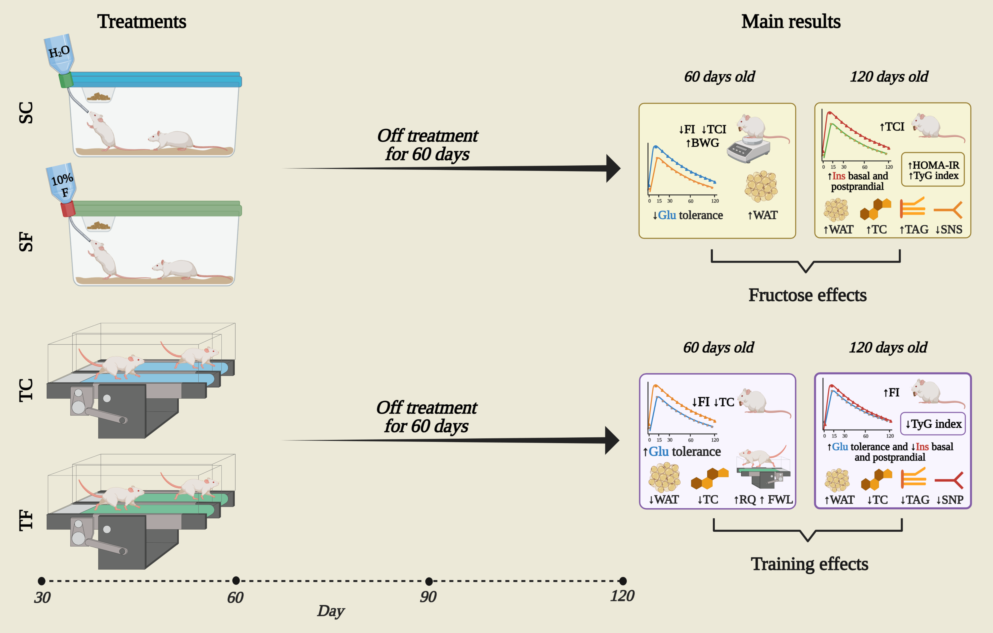
During 30 days of treatment with fructose and/or training, the rats of all groups exhibited different final workload (FI), body weight gain (BWG) and total caloric intake (TCI). The SF rats showed a significant decrease in the AUC of FI (P= 0.001, Figure 1A) compared to SC rats. The TF rats also showed a decrease in the AUC of FI (P= 0.001, Figure 1A) compared to the TC rats. Yet, immediately after the nutritional transition, from 60-120 days, the TC rats experienced a catch-up in chow consumption (P= 0.001, Figure 1A) compared to equivalent control rats.
The BWG (P= 0.05, Figure 1B) in rats that ingested fructose was opposite compared to control rats, from 30-60 days. We found no significance difference in the BWG pattern between the treatments and groups, from 60-120 days (Figure 1B). The AUC of TCI (P< 0.001, Figure 1C) after the first 30 experimental days was greater in TC and TF rats than sedentary counterparts. However, SF and TF rats ingested more calories (P=0.001, Figure 1C) than equivalent control non-supplemented rats from 60 to 120 days-old. The increase in FluI by fructose-supplemented peri-pubertal rats (supplementary material), could be explained, at least, the highest TCI presented in both experimental periods.
2.3. Tissues and plasma parameters
At 60 days-old, TC rats wasn’t found differences among all groups in fast glycaemia (Table 3). In contrast, SF rats showed an increase in fasting glycaemia (P= 0.02, Table 3) compared to SC rats at 120 days-old. SF rats showed a significantly increase in the AUC of glycaemia (P= 0.01, Figure 2A) during the ipGTT compared to their counterparts at 60 days-old. The training performed during peri-pubertal period was able to decrease the AUC of glycaemia of TF rats compared to SF rats at 60 (P= 0.0004, Figure 2A) and 120 days-old (P= 0.01, Figure 2B).
Moderate-intensity training and fructose intake affected fasting insulin levels in different manners. At 120 days-old, SF rats showed an increase in insulin levels (P= 0.0004, Table 3) compared to SC rats. Otherwise, the training decreased this parameter when TF rats was compared to SF (P= 0.004, Table 3). SF rats also showed an increase in the AUC of insulinemia (P< 0.0001, Figure 3) during the ipGTT compared to SC rats. The training reduced significantly the AUC of insulinemia (P< 0.0001, Figure 3) in TF rats compared to sedentary equivalents.
Table 4 shows that HOMA-IR was increased in adult SF rats compared to SC (P= 0.002) and TF rats (P= 0.02). The TyG index also showed a significantly increase in SF compared to SC rats (P< 0.0001, Table 4). However, the training performed by TF rats contributed to reduce the TyG index compared to the rats that receive fructose supplementation, only (P= 0.0003, Table 4).
At 60 day of treatment, TC rats showed a significant decrease in the retroperitoneal fat pad (P= 0.002; Table 3) compared to SC rats. No changes in the liver weight were observed in all experimental groups (Table 3). At 120 days-old, SF rats exhibited significantly increased in the periepididymal and retroperitoneal fat pads (P= 0.001 and P= 0.03, respectively, Table 4) compared to the equivalent control. Curiously, the TC rats also exhibited an increased in the periepididymal fat pad (P= 0.01; Table 4) compared to SC rats.
TC levels (P= 0.001, Table 3) in rats treated with fructose and training was reduced when compared to sedentary counterparts at 60 days-old. However, at 120 days-old, the SF rats showed an increase the TC and TAGs levels (P= 0.0001 and P< 0.0001, respectively, Table 4) compared to SC rats. TF rats had a reduction in these parameters, compared to SF rats (P= 0.001 and P= 0.0003, respectively, Table 4).
2.4. Parasympathetic and sympathetic electrical nerve activity
3. Discussion
We found that chronic intake of 10% fructose in drinking water during peri-pubertal period programming male rats to the MetS-related phenotype at adulthood. Besides, for the first time, we demonstrated that moderate physical training applied during peri-pubertal period, simultaneously with fructose consumption, was able to preventing several MetS components, such as BWG, dyslipidemia, insulin resistance, hyperglycaemia and hyperinsulinemia during ipGTT and increased parasympathetic nerve activity in adult Wistar rats.
In this study, we obtained substantial data that early fructose supplementation is capable of induce the onset of MetS, such as impaired blood glucose-excursion following ipGTT in 60 days-old sedentary rats. Consistent with previous findings [30], we also showed a reduction in FI and an increase in BWG and adiposity in these animals, a phenomenon unrelated to the TCI, but sufficient to disturb glucose tolerance at end of the 30 days of treatment. No alteration in fasting glycaemia was detected, in agreement with previous findings that introduced fructose in 21 days-old male rats [31].
At 60 days-old, trained animals exhibited different chronic adaptations. The moderate physical training was able to improve functional and metabolic capacity in control rats, as measured by FWL and RQ. Interestingly, the VO2max, EE, glucose tolerance and total cholesterol levels improves only in rats that were treated with fructose and training during peri-pubertal period. Fructose-rich diet was shown to not impair the ability to perform aerobic trainings in a similar study [32]. It has been shown that fructose co-ingestion increases carbohydrate availability and post-exercise glycogen replacement rates, maximizing endurance performance in athletes during prolonged moderate- to high-intensity exercise, as well as for acute post-exercise recovery [33]. Furthermore, combined treatments in this study may have enhanced the energetic demand and lipolysis, preventing weight gain and dyslipidemia in young rats, in accord to previous data [34].
High levels of plasma insulin concentration has been associated to increased insulin secretion and/or decreased insulin clearance [35]. Our results showed that the high fructose consumption during peri-pubertal period promoted an increase in fasting glycaemia and insulinemia in sedentary adult rats compared to equivalent control. It is important to highlight that the hyperinsulinemia during ipGTT persisted even 60 days after the interruption of fructose consumption, despite normal blood glucose during in the same test. Consistent with previous findings, our data also showed that the administration of fructose induces insulin resistance in adult rats, according the HOMA IR and/or TYG index [15,36,37]. TYG index making a relation between glycaemia and lipid profile, being strongly associated with diabetes, nonalcoholic fatty liver disease and metabolic syndrome [38,39].
Fructose consumption in 5 month-old rats increases the risk of cardiovascular disease by alterations in fat liver and blood lipid parameters [17]. Cardiovascular disease, visceral adiposity and type 2 diabetes is also present in young and adult humans [40,41,42]. In our study, with only 30 days of fructose intake, we detected an increase in TCI, excessive TC and TAGs levels, and elevated periepididymal and retroperitoneal adiposity in sedentary adult rats that received fructose during peri-pubertal period, compared to equivalent control. It was recently reported that the ingestion of 20% fructose elevates circulating levels of fibroblast growth factor 21 (FGF21) in C57BL/6J mice, causing resistance to FGF21 and, subsequently, an increase of lipid droplets in the liver and the size of adipose cells in white adipose tissue (WAT) [43].
Although weight gain remained unchanged in adult rats, our data suggest the high potential of fructose to alters lipid metabolism and promote adipogenesis-induced hyperinsulinemia [44]. Similar studies in literature that used 10% fructose in rats showed no increase in body weight [1,2]. Harrel et al. [45] showed that the fructose consumption by adolescents increased fat stores without the BWG. Contrasting BWG outcome was observed after consumption of 20% fructose [4]. We did not evaluate the hepatic lipogenesis, which also seems to contribute to a large production in TAGs levels [36,42,46]. The livers were weighed, but no statistical difference was found.
Nutritional insults during critical developmental periods are known for their plastic long-term impact on metabolism and described on literature as being responsible for several adult health outcomes [9,11,13,47]. On the other hand, interventions using physical exercise were reported to attenuate their disturbing effects [48,49,50]. Accordingly, the present data revealed that adolescent rats that received fructose and training concomitantly, presented a reduction in vagal electrical activity compared to the sedentary counterpart, and this mechanism could be explained, at least, a better metabolic profile in these animals. A high activity of the parasympathetic nervous system has been related to metabolic disorders such as obesity [51], hyperinsulinemia and insulin resistance [7].
We also showed that fructose treatment programmed a large reduction in sympathetic nervous system activity in trained and non-trained adult rats. It has been shown that sympathetic nervous system activity is decreased in obese animals and humans [52,53]. Our observation is in agreement of “MONA LISA” (Most Obesities kNown Are Low In Sympathetic Activity) hypothesis, proposed by Bray, which says that the resistance to leptin, related with the obese state, is linked to a decreased local sympathetic activity [52,54,55].
American teenager consumes approximately 72.8 g/day of fructose [45]. Although the difficulty in developing experimental models that present all the characteristics of humans MetS, in the present study we used a dose equivalent to a diet containing 48–57% fructose [14] that also mimics fructose consumption by human adolescents [4], we suggest that 10% fructose ingestion by pubescent rats in this investigation was associated with may result in a long-last programming composed for an onset of MetS-related disorders in adult life [16,36,56,57,58].
Several studies have demonstrated an effectiveness of the physical exercise on prevention of MetS risk factors [22,56,59] and a direct correlation between moderate exercise and improved insulin sensitivity in young and adult individuals [60,61], even after cessation of training [26,28]. Our laboratory has already demonstrated that the short-term physical training when applied in adolescence, protects the organism from the MetS caused by a high-fat diet (HFD) in adolescence [25,62]. Frantz and colleagues [2,22] reinforce the beneficial effects of physical exercise as an important regulator of fructose dyslipidemia, especially in plasma TAGs levels reduction, corroborating with our results in TF animals.
Consistent with previous findings on the protective ability of moderate-intensity physical exercise on glucose-insulin homeostasis, lipid metabolism and autonomic parameters [25,34,63], our results showed a significant lower insulin, TC and TAGs fast levels, as well as in HOMA-IR and TYG index in adult rats that were supplemented with fructose and submitted to the moderate physical protocol during peri-pubertal period, compared to sedentary group. Likewise, after 6 weeks of a moderate exercise program, Dupas and coworkers found reductions in BW, fat stores and improved glucose-insulin metabolism in fructose-fed rats. For the respective authors, the results were associated with exercise-triggered improvement in muscle insulin sensitivity [37].
Our data also showed that moderate-intensity training performed at peri-pubertal period could reduce the parasympathetic nerve activity in adult rats treated with fructose. This result corroborates with previous results that related beneficial effects for vagus nerve activity in HFD-fed exercised animals [25,62]. Nevertheless, it is notable that the effect of moderate-intensity training was observed in the present study, even when it was interrupted after 60 days before the experiments, suggesting some physiological adaptations and/or plasticity of the young organism, as it was carried in a sensitive window to programming health or disease [7] in order to prevent components of MetS. When moderate physical training have been started early, the effects of discontinuation are not so deleterious [25,26,28,29].
This work predominantly focuses on a non-pharmacological strategy to prevent or attenuate several compounds of fructose-induced MetS in male Wistar rats. Overall, the main results reveal the ability of a young trained organism to recover, at least in part, to its previous metabolic pattern, as observed in supplemented and non-supplemented rats. Despite some limitations, we suggest that the expression of some proteins should be investigated to identify other mechanisms, in addition to glucose-insulin homeostasis and ANS activity experiments performed, the way an organism responds to fructose supplementation, as well as the potential beneficial effects of moderate-intensity training on programming metabolism especially in fructose-supplemented rats. This needs to be considered over the long term when thinking about translational research.
4. Materials and Methods
4.1. Animals and experimental design
The handling of animals and experimental procedures were in accordance with the ARRIVE guidelines for experiments involving animals [64] and approved by the Ethics Committee on Animal Use of Maringa State University – CEUA/UEM (protocol number 5669210917). Male Wistar rats were obtained at 25 days-old. They were kept in appropriate cages (5 rats per cage) under controlled temperature conditions (22±2°C), and a light/dark cycle of 12 h (07:00 a.m. to 07:00 p.m.), with ad libitum access to water and a standard diet (Nuvital®, Curitiba, PR, Brazil).
After five days of adaptation, at 30 days-old, the animals were divided into four groups: Sedentary control (SC), that received water and standard rat chow ad libitum during all the period; Sedentary fructose (SF), that received 10% fructose in the drinking water from the 30 to 60 days-old and standard rat chow ad libitum; Trained control (TC), that performed moderate-intensity training from the 30 to 60 days-old and received water and standard rat chow ad libitum, and Trained fructose (TF), that received 10% fructose in the drinking water and performed moderate-intensity training at the same time from the 30 to 60 days-old. In order to evaluate the effect of the treatment (fructose and physical training) on metabolism immediately after the treatment period, data was collected in 60 days-old rats. To evaluate the potential role of fructose in programming, data was collected in 120 day-old rats.
4.2. Preparation of fructose drinking water
The fructose used in this protocol was D-fructose >99% (Labsynth®, São Paulo/SP, Brazil). The fructose drink was prepared each two days and based on the formula weight/volume (w/v). For the preparation of 10% of fructose [16], 10 g of fructose was diluted in 100 mL of filtered water.
4.3. Effort test and physical training
All rats performed an effort test (ET) before (initial: 30 days-old), in the final of treatment and physical training (middle: 60 days-old), and in the final of the experimental period (final: 120 days-old), to determine their individual oximetry [maximal oxygen uptake (VO2max), energy expenditure (EE) and respiratory quotient/coefficient (RQ)] and treadmill [final workload (FWL)] parameters. Incremental ET were performed every 15 days to adjust the training load. The test utilized a gas analyzer coupled to a treadmill for rodents (Panlab, Harvard Apparatus®, LE405 76-0195 O2/CO2, Cornellà, Barcelona, Spain). The ET began with a warm up (5 min, 10 cm/s, 0º of inclination), after which the velocity was increased by 9 cm/s every 3 min until exhaustion of the animal to obtain VO2max and FWL, using Metabolism software, version 2.2.02. The decision to use 3 min at each stage was previously described [65], who reported that oxygen consumption stabilized after approximately 3 min at each stage of exercise after a change in workload. At the end of the treadmill line, a stainless steel grid emitted electrical stimuli (0.2 mA in < 1 s) to keep the animal in motion, as previously reported [66]. The animal’s inability to maintain the pace was considered to be a sign of exhaustion [67].
Moderate-intensity training was performed with running on a treadmill (Panlab, Harvard Apparatus®, LE8710R 76-0308, rat 5-lanes). Previous adaptation was performed in 5 sessions with durations of 10, 12, 14, 16 and 18 minutes and an intensity of 16 cm/s. Two days of rest were established to apply the ET. The prescription was based on effort test (ET) and was performed by the individual value of the FWL corresponding to 55% and 65% of VO2max to optimize the fat metabolism zone [68]. The treadmill training protocol was performed for 44 minutes a day (9 am to 10 am), 3 days a week in a 4-week macrocycle. The sessions were distributed with 2 minutes of warming up and cooling down at 20 cm/s and 40 minutes of continuous running at a moderate-intensity training (~ 55% to 65% FWL of the ET) [29]. Rats that reached the same FWL trained at the same time. The training protocols were completed at 60 days.
In the supplementary material we provide additional tables referring to the training periodization (A), treadmill adaptation (B), effort test (C), and training sessions (D).
4.4. Biometric parameters
Food (FI) and fluid (FluI) intake and body weight gain (BWG) were measured weekly from 30 to 120 days-old. The FI and FluiI were calculated by subtracting the measured amount provided to the remaining amounts in the cage, and divided by the number of days and the number of rats in the box [4]. The total caloric intake (TCI) was calculated based on the amount of FI and FluI and the corresponding constants [4,69]. The area under the curve (AUC) was calculated for the FI, BWG and TCI.
4.5. Tissues and plasma parameters
At 60 and 120 days-old, the animals from all groups were weighed and decapitated after a 12-h overnight fast, and blood samples were collected and centrifuged (10,000 rpm for 5 min) to obtain plasma which was used to measure specific parameters. Glucose, total cholesterol (TC) and triglycerides (TAGs) levels in plasma were measured by an enzymatic method using a specific colorimetric commercial kit (Gold Analisa®, Belo Horizonte, Brazil) and quantified by spectrophotometry (BIO200FL, Bio Plus®, São Paulo, Brazil).
Insulin was measured by radioimmunoassay (RIA) [36] with a gamma counter through a gamma particle emission counter (Wizard Automatic Gamma Counter, TM-2470, PerkinElmer®, Shelton, CT, United States), using standard rat insulin, anti-rat insulin (Sigma-Aldrich®, St Louis, MO, United States) and recombinant human insulin ([125I]-Insulin (h)] (PerkinElmer®, Shelton, CT, United States). Insulin resistance markers were evaluated by homeostasis model assessment-index (HOMA-IR) [70] and triglyceride and glucose index (TyG) [39].
Retroperitoneal and periepididymal fat pads were removed and weighted to provide an estimate of the fat accumulation. The liver was dissected and weighed. Each of the fat pad stores, and liver values were correlated with the BW of each rat and were calculated as g/100kg of BW [7]. The samples were weighed on a precision scale (Model BD-600; Instrutherm, São Paulo, Brazil).
4.6. Intraperitoneal glucose tolerance test (ipGTT)
The ipGTT was performed at 60 and 120 days-old. Food was withdrawn 12 h before the test, and free access to water was allowed. Rats were weighed and had their tail tips cut for measurement of blood glucose levels at baseline (time = 0). Afterwards, glucose solution 50% (w/v) was administered by I.P. injection at a dose of 2 g kg–1 body weight and blood samples were collected at 15, 30, 60 and 120 min to determinate the glucose and insulin levels. The AUC was calculated from the values for each rat. Anesthesia was not used during the ipGTT test [28].
The glucose was measured by the enzymatic method using a commercial colorimetric kit (Gold Analisa®, Belo Horizonte, Brazil) and quantified by spectrophotometry (BIO200FL, Bio Plus®, São Paulo, Brazil). The insulin levels were measured by RIA [71]. Standard human insulin, anti-rat insulin antibody (Sigma Aldrich®, St. Louis, MO, United States) and recombinant human insulin labeled Iodo125 (PerkinElmer®, Shelton, CT, United States) were used. The intra- and interassay coefficients of variation were 9.8% and 12.2%, respectively, for insulin. The insulin level of the detection limit was 0.006 ng/mL, and measurements were performed in a single assay.
4.7. Parasympathetic and sympathetic electrical nerve activity
To verify the autonomic nervous system activity (ANS) at 120 days-old, a longitudinal surgical incision was made on the anterior cervical region of the animals. The left vagus superior branch was isolated and placed over a silver electrode inside a faraday cage [25]. After 12 min of vagus nerve electrical recordings, a laparotomy was performed to isolate the branch of the sympathetic nerve that is located in the splanchnic region; this branch originates in the lumbar plexus at the level of L2 and extends to the retroperitoneal adipose tissue. The neural signal output was acquired using the Insight interface (Insight®, Ribeirão Preto, SP, Brazil) for 12 min, from which 20 recorded frames of 5 s from each animal were randomly chosen for spike counting. Spikes >0 mV were considered. The average number of spikes was used as the nerve firing rate.
4.8. Statistical analysis
Data were expressed as the mean ± SEM and were subjected a two-way ANOVA considering fructose (F), training (T) and interaction (I; fructose vs training) as factors. Tukey’s post hoc test was used for comparison between groups (SC, TC, SF and TF). Repeated measures ANOVA with Sidak’s multiple comparisons test was performed to analyze the food intake, weight gain, total caloric intake, blood glucose and insulin levels measured at different time points. P < 0.05 was considered statistically significant. GraphPad Prism version 9.1.0 for Windows (GraphPad Software, La Jolla, CA, USA) was used for statistical analyses and developing graphs.
5. Conclusions
In conclusion, our results showed that the 10% fructose intake during peri-pubertal period remarkably programs adult offspring to MetS, and concomitantly, the moderate-intensity training confers the ability to prevent autonomic and metabolic dysfunctions in male rats, protecting the organism against the deleterious effects of physical inactivity and fructose-induced MetS.
Supplementary Materials
Supporting information can be downloaded from Supplementary Files.
Author Contributions
Conceptualization and methodology design, S.S., V.M. and P.M.; formal analysis, S.S. and V.M.; performing experiments and collecting data S.S., V.M., M.P., L.S, L.T., F.F., T.R., C.F., C.M., A.M. and D.A.; writing—original draft preparation, S.S. and V.M.; writing—review and editing, S.S. and V.M.; supervision, P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Brazilian National Council for Scientific and Technological Development (CNPq), and Coordination for the Improvement of Higher Education Personnel (CAPES).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee on Animal Use of Maringá State University – CEUA/UEM (protocol number 5669210917).
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
We thank Ms. Maroly Pinto and Ms. Marli Licero for helping care for the rats in the animal facility. Thanks also to Luiz D. C. Lima for helping with the training periodization.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amaral, F.; Lima, N.E.A.; Ornelas, E.; Simardi, L.; Fonseca, F.L.A.; Maifrino, L.B.M. Effect of Different Exercise Intensities on the Pancreas of Animals with Metabolic Syndrome. Diabetes, Metab. Syndr. Obes. Targets Ther. 2015, 8, 115–120. [Google Scholar] [CrossRef]
- Frantz, E.D.C.; Prodel, E.; Braz, I.D.; Giori, I.G.; Bargut, T.C.L.; Magliano, D.C.; da Nobrega, A.C.L. Modulation of the Renin-Angiotensin System in White Adipose Tissue and Skeletal Muscle: Focus on Exercise Training. Clin. Sci. 2018, 132, 1487–1507. [Google Scholar] [CrossRef] [PubMed]
- Bargut, T.C.L.; Santos, L.P.; Machado, D.G.L.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Eicosapentaenoic Acid (EPA) vs. Docosahexaenoic Acid (DHA): Effects in Epididymal White Adipose Tissue of Mice Fed a High-Fructose Diet. Prostaglandins Leukot. Essent. Fat. Acids 2017, 123, 14–24. [Google Scholar] [CrossRef]
- Mamikutty, N.; Thent, Z.C.; Sapri, S.R.; Sahruddin, N.N.; Mohd Yusof, M.R.; Haji Suhaimi, F. The Establishment of Metabolic Syndrome Model by Induction of Fructose Drinking Water in Male Wistar Rats. Biomed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lebda, M.A.; Tohamy, H.G.; El-Sayed, Y.S. Long-Term Soft Drink and Aspartame Intake Induces Hepatic Damage via Dysregulation of Adipocytokines and Alteration of the Lipid Profile and Antioxidant Status. Nutr. Res. 2017, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- White, A.H.; James, S.A.; Paulson, S.W.; Beebe, L.A. Sugar Sweetened Beverage Consumption among Adults with Children in the Home. Front. Nutr. 2018, 5, 1–7. [Google Scholar] [CrossRef]
- de Oliveira, J.C.; Lisboa, P.C.; de Moura, E.G.; Barella, L.F.; Miranda, R.A.; Malta, A.; da Silva Franco, C.C.; da Silva Ribeiro, T.A.; Torrezan, R.; Gravena, C.; et al. Poor Pubertal Protein Nutrition Disturbs Glucose-Induced Insulin Secretion Process in Pancreatic Islets and Programs Rats in Adulthood to Increase Fat Accumulation. J. Endocrinol. 2013, 216, 195–206. [Google Scholar] [CrossRef]
- Trombini, A.B.; Franco, C.C.S.; Miranda, R.A.; de Oliveira, J.C.; Barella, L.F.; Prates, K. V.; de Souza, A.A.; Pavanello, A.; Malta, A.; Almeida, D.L.; et al. Early Treatment with Metformin Induces Resistance against Tumor Growth in Adult Rats. Cancer Biol. Ther. 2015, 16, 958–964. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Beedle, A.S. Early Life Events and Their Consequences for Later Disease: A Life History and Evolutionary Perspective. Am. J. Hum. Biol. 2007, 19, 1–19. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef]
- Barker, D.J.P. The Developmental Origins of Chronic Adult Disease. Acta Paediatr. 2007, 93, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, C.A.; Erthal, R.P.; Ogo, F.M.; Peres, M.N.C.; Vieira, H.R.; Conejo, C.; Tófolo, L.P.; Francisco, F.A.; Silveira, S. da S.; Malta, A.; et al. A High Fat Diet during Adolescence in Male Rats Negatively Programs Reproductive and Metabolic Function Which Is Partially Ameliorated by Exercise. Front. Physiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.R.O.; Ribeiro, M.V.G.; Peres, M.N.C.; Piovan, S.; Gonçalves, G.D.; Saavedra, L.P.J.; Martins, J.N. de L.; Junior, M.D.F.; Cavalcante, K.V.N.; Lopes, G. kian G.; et al. Protein Restriction in the Peri-Pubertal Period Induces Autonomic Dysfunction and Cardiac and Vascular Structural Changes in Adult Rats. Front. Physiol. 2022, 13, 663. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; McNeill, J.H. Fructose-Induced Hypertension in Rats Is Concentration- and Duration-Dependent. J. Pharmacol. Toxicol. Methods 1995, 33, 101–107. [Google Scholar] [CrossRef]
- Maiztegui, B.; Morelli, M.I.; Raschia, M.A.; Del Zotto, H.; Gagliardino, J.J. Islet Adaptive Changes to Fructose-Induced Insulin Resistance: β-Cell Mass, Glucokinase, Glucose Metabolism, and Insulin Secretion. J. Endocrinol. 2009, 200, 139–149. [Google Scholar] [CrossRef]
- de Moura, R.F.; Ribeiro, C.; de Oliveira, J.A.; Stevanato, E.; de Mello, M.A.R. Metabolic Syndrome Signs in Wistar Rats Submitted to Different High-Fructose Ingestion Protocols. Br. J. Nutr. 2009, 101, 1178–1184. [Google Scholar] [CrossRef]
- Sadowska, J.; Bruszkowska, M. Comparing the Effects of Sucrose and High-Fructose Corn Syrup on Lipid Metabolism and the Risk of Cardiovascular Disease in Male Rats. Acta Sci. Pol. Technol. Aliment. 2017, 16, 231–240. [Google Scholar] [CrossRef]
- Castro, M.C.; Francini, F.; Schinella, G.; Caldiz, C.I.; Zubiría, M.G.; Gagliardino, J.J.; Massa, M.L. Apocynin Administration Prevents the Changes Induced by a Fructose-Rich Diet on Rat Liver Metabolism and the Antioxidant System. Clin. Sci. 2012, 123, 681–692. [Google Scholar] [CrossRef]
- Francini, F.; Castro, M.C.; Gagliardino, J.J.; Massa, M.L. Regulation of Liver Glucokinase Activity in Rats with Fructose-Induced Insulin Resistance and Impaired Glucose and Lipid Metabolism. Can. J. Physiol. Pharmacol. 2009, 87, 702–710. [Google Scholar] [CrossRef]
- Kindlovits, R.; Bertoldi, J.M.C.R.J.; Rocha, H.N.M.; Bento-Bernardes, T.; Gomes, J.L.P.; de Oliveira, E.M.; Muniz, I.C.; Pereira, J.F.; Fernandes-Santos, C.; Rocha, N.G.; et al. Molecular Mechanisms Underlying Fructose-Induced Cardiovascular Disease: Exercise, Metabolic Pathways and MicroRNAs. Exp. Physiol. 2021, 106, 1224–1234. [Google Scholar] [CrossRef]
- Steenson, S.; Umpleby, A.M.; Lovegrove, J.A.; Jackson, K.G.; Fielding, B.A. Role of the Enterocyte in Fructose-induced Hypertriglyceridaemia. Nutrients 2017, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Frantz, E.D.C.; Medeiros, R.F.; Giori, I.G.; Lima, J.B.S.; Bento-Bernardes, T.; Gaique, T.G.; Fernandes-Santos, C.; Fernandes, T.; Oliveira, E.M.; Vieira, C.P.; et al. Exercise Training Modulates the Hepatic Renin–Angiotensin System in Fructose-Fed Rats. Exp. Physiol. 2017, 102, 1208–1220. [Google Scholar] [CrossRef] [PubMed]
- Soares de Alencar Mota, C.; Ribeiro, C.; de Araújo, G.G.; de Araújo, M.B.; de Barros Manchado-Gobatto, F.; Voltarelli, F.A.; de Oliveira, C.A.M.; Luciano, E.; de Mello, M.A.R. Exercise Training in the Aerobic/Anaerobic Metabolic Transition Prevents Glucose Intolerance in Alloxan-Treated Rats. BMC Endocr. Disord. 2008, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mostarda, C.; Rogow, A.; Moraes Silva, I.C.; De La Fuente, R.N.; Jorge, L.; Rodrigues, B.; Heeren, M. V.; Caldini, E.G.; De Angelis, K.; Irigoyen, M.C. Benefits of Exercise Training in Diabetic Rats Persist after Three Weeks of Detraining. Auton. Neurosci. Basic Clin. 2009, 145, 11–16. [Google Scholar] [CrossRef]
- Tófolo, L.P.; da Silva Ribeiro, T.A.; Malta, A.; Miranda, R.A.; Gomes, R.M.; de Oliveira, J.C.; Abdennebi-Najar, L.; de Almeida, D.L.; Trombini, A.B.; da Silva Franco, C.C.; et al. Short-Term Moderate Exercise Provides Long-Lasting Protective Effects against Metabolic Dysfunction in Rats Fed a High-Fat Diet. Eur. J. Nutr. 2014, 54, 1353–1362. [Google Scholar] [CrossRef]
- Lehnen, A.M.; Leguisamo, N.M.; Pinto, G.H.; Markoski, M.M.; de Angelis, K.; Machado, U.F.; Schaan, B. The Beneficial Effects of Exercise in Rodents Are Preserved after Detraining: A Phenomenon Unrelated to GLUT4 Expression. Cardiovasc. Diabetol. 2010, 9, 1–8. [Google Scholar] [CrossRef]
- Hötting, K.; Röder, B. Beneficial Effects of Physical Exercise on Neuroplasticity and Cognition. Neurosci. Biobehav. Rev. 2013, 37, 2243–2257. [Google Scholar] [CrossRef]
- Moreira, V.M.; Franco, C.C. da S.; Prates, K.V.; Gomes, R.M.; de Moraes, A.M.P.; Ribeiro, T.A.; Martins, I.P.; Previate, C.; Pavanello, A.; Matiusso, C.C.I.; et al. Aerobic Exercise Training Attenuates Tumor Growth and Reduces Insulin Secretion in Walker 256 Tumor-Bearing Rats. Front. Physiol. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Moreira, V.M.; Almeida, D.; da Silva Franco, C.C.; Gomes, R.M.; Palma-Rigo, K.; Prates, K.V.; Tófolo, L.P.; Malta, A.; Francisco, F.A.; Pavanello, A.; et al. Moderate Exercise Training since Adolescence Reduces Walker 256 Tumour Growth in Adult Rats. J. Physiol. 2019, 597, 3905–3925. [Google Scholar] [CrossRef]
- Sulis, P.M.; Motta, K.; Barbosa, A.M.; Besen, M.H.; Da Silva, J.S.; Nunes, E.A.; Rafacho, A. Impact of Fish Oil Supplementation and Interruption of Fructose Ingestion on Glucose and Lipid Homeostasis of Rats Drinking Different Concentrations of Fructose. Biomed Res. Int. 2017, 2017, 1–16. [Google Scholar] [CrossRef]
- Vasiljević, A.; Bursać, B.; Djordjevic, A.; Milutinović, D.V.; Nikolić, M.; Matić, G.; Veličković, N. Hepatic Inflammation Induced by High-Fructose Diet Is Associated with Altered 11βHSD1 Expression in the Liver of Wistar Rats. Eur. J. Nutr. 2014, 53, 1393–1402. [Google Scholar] [CrossRef]
- Ghezzi, A.C.; Cambri, L.T.; Ribeiro, C.; Botezelli, J.D.; Mello, M.A. Impact of Early Fructose Intake on Metabolic Profile and Aerobic Capacity of Rats. Lipids Health Dis. 2011, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.J.; Gonzalez, J.T.; van Loon, L.J.C. Fructose Co-Ingestion to Increase Carbohydrate Availability in Athletes. J. Physiol. 2019, 597, 3549–3560. [Google Scholar] [CrossRef] [PubMed]
- Moraes-Silva, I.C.; Mostarda, C.; Dias Moreira, E.; Silva, K.A.S.; dos Santos, F.; de Angelis, K.; de Moura Azevedo Farah, V.; Irigoyen, M.C. Preventive Role of Exercise Training in Autonomic, Hemodynamic, and Metabolic Parameters in Rats under High Risk of Metabolic Syndrome Development. J. Appl. Physiol. 2013, 114, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Vettorazzi, J.F.; Kurauti, M.A.; Soares, G.M.; Borck, P.C.; Ferreira, S.M.; Branco, R.C.S.; Michelone, L.D.S.L.; Boschero, A.C.; Junior, J.M.C.; Carneiro, E.M. Bile Acid TUDCA Improves Insulin Clearance by Increasing the Expression of Insulin-Degrading Enzyme in the Liver of Obese Mice. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Dupas, J.; Feray, A.; Goanvec, C.; Guernec, A.; Samson, N.; Bougaran, P.; Guerrero, F.; Mansourati, J. Metabolic Syndrome and Hypertension Resulting from Fructose Enriched Diet in Wistar Rats. Biomed Res. Int. 2017. [Google Scholar] [CrossRef]
- Dupas, J.; Feray, A.; Guernec, A.; Pengam, M.; Inizan, M.; Guerrero, F.; Mansourati, J.; Goanvec, C. Effect of Personalized Moderate Exercise Training on Wistar Rats Fed with a Fructose Enriched Water. Nutr. Metab. (Lond). 2018, 15, 1–12. [Google Scholar] [CrossRef]
- Dengel, D.R.; Pratley, R.E.; Hagberg, J.M.; Rogus, E.M.; Goldberg, A.P. Distinct Effects of Aerobic Exercise Training and Weight Loss on Glucose Homeostasis in Obese Sedentary Men. J. Appl. Physiol. 1996, 81, 318–325. [Google Scholar] [CrossRef]
- Vasques, A.C.J.; Novaes, F.S.; de Oliveira, M. da S.; Matos Souza, J.R.; Yamanaka, A.; Pareja, J.C.; Tambascia, M.A.; Saad, M.J.A.; Geloneze, B. TyG Index Performs Better than HOMA in a Brazilian Population: A Hyperglycemic Clamp Validated Study. Diabetes Res. Clin. Pract. 2011, 93, e98–e100. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Yoffe, P.; Hills, N.; Lustig, R.H. The Relationship of Sugar to Population-Level Diabetes Prevalence: An Econometric Analysis of Repeated Cross-Sectional Data. PLoS One 2013, 8, e57873. [Google Scholar] [CrossRef]
- Page, K.A.; Chan, O.; Arora, J.; Belfort-Deaguiar, R.; Dzuira, J.; Roehmholdt, B.; Cline, G.W.; Naik, S.; Sinha, R.; Constable, R.T.; et al. Effects of Fructose vs Glucose on Regional Cerebral Blood Flow in Brain Regions Involved with Appetite and Reward Pathways. JAMA - J. Am. Med. Assoc. 2013, 309, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming Fructose-Sweetened, Not Glucose-Sweetened, Beverages Increases Visceral Adiposity and Lipids and Decreases Insulin Sensitivity in Overweight/Obese Humans. J. Clin. Invest. 2009, 119, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tian, Y.; Zuo, Y.; Zhang, X.; Gao, Y.; Wang, P.; Sun, L.; Zhang, H.; Liang, H. Nicotinamide Riboside Ameliorates High-Fructose-Induced Lipid Metabolism Disorder in Mice via Improving FGF21 Resistance in the Liver and White Adipose Tissue. Food Funct. 2022, 13, 12400–12411. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Díazcouder, A.; Romero-Nava, R.; Carbó, R.; Sánchez-Lozada, L.G.; Sánchez-Muñoz, F. High Fructose Intake and Adipogenesis. Int. J. Mol. Sci. 2019, 20, 2787. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.S.; Burgado, J.; Kelly, S.D.; Johnson, Z.P.; Neigh, G.N. High-Fructose Diet during Periadolescent Development Increases Depressive-like Behavior and Remodels the Hypothalamic Transcriptome in Male Rats. Psychoneuroendocrinology 2015, 62, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Lustig, R.H. Fructose and Nonalcoholic Fatty Liver Disease. J Calif Dent Assoc 2016, 44, 613–617. [Google Scholar] [CrossRef]
- Munetsuna, E.; Yamada, H.; Yamazaki, M.; Ando, Y.; Mizuno, G.; Ota, T.; Hattori, Y.; Sadamoto, N.; Suzuki, K.; Ishikawa, H.; et al. Maternal Fructose Intake Disturbs Ovarian Estradiol Synthesis in Rats. Life Sci. 2018, 202, 117–123. [Google Scholar] [CrossRef]
- Almeida, D.L.; Moreira, V.M.; Cardoso, L.E.; Junior, M.D.F.; Pavanelo, A.; Ribeiro, T.A.; da Silva Franco, C.C.; Tófolo, L.P.; Peres, M.N.C.; Ribeiro, M.V.G.; et al. Lean in One Way, in Obesity Another: Effects of Moderate Exercise in Brown Adipose Tissue of Early Overfed Male Wistar Rats. Int. J. Obes. 2022, 46, 137–143. [Google Scholar] [CrossRef]
- Almeida, D.L.; Fabricio, G.S.; Tófolo, L.P.; Ribeiro, T.A.; Matiusso, C.C.I.; Ribeiro, M.V.G.; Oliveira Ferreira, A.R.; Pavanello, A.; Malta, A.; Palma-Rigo, K.; et al. Early Postnatal Overnutrition Impairs VO2max Gains with Moderate Exercise and Increase Post-Exercise Muscle Damage in Adult Male Rats. J. Dev. Orig. Health Dis. 2022, 13, 406–410. [Google Scholar] [CrossRef]
- Ribeiro, T.A.; Tófolo, L.P.; Martins, I.P.; Pavanello, A.; De Oliveira, J.C.; Prates, K.V.; Miranda, R.A.; Da Silva Franco, C.C.; Gomes, R.M.; Francisco, F.A.; et al. Maternal Low Intensity Physical Exercise Prevents Obesity in Offspring Rats Exposed to Early Overnutrition. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Barella, L.F.; de Oliveira, J.C.; Branco, R.C.S.; Camargo, R.L.; Gomes, R.M.; Mendes, F.C.V.; Miranda, R.A.; Gravena, C.; Torrezan, R.; Grassiolli, S.; et al. Early Exposure to a High-Fat Diet Has More Drastic Consequences on Metabolism Compared with Exposure during Adulthood in Rats. Horm. Metab. Res. 2012, 44, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A.; York, D.A. The MONA LISA Hypothesis in the Time of Leptin. Recent Prog. Horm. Res. 1998, 53, 95–117. [Google Scholar]
- Messina, G.; de Luca, V.; Viggiano, A.; Ascione, A.; Iannaccone, T.; Chieffi, S.; Monda, M. Autonomic Nervous System in the Control of Energy Balance and Body Weight: Personal Contributions. Neurol. Res. Int. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Barateiro, A.; Mahú, I.; Domingos, A.I. Leptin Resistance and the Neuro-Adipose Connection. Front. Endocrinol. (Lausanne). 2017, 8, 1–4. [Google Scholar] [CrossRef]
- Bray, G.A. Obesity, a Disorder of Nutrient Partitioning: The MONA LISA Hypothesis. J. Nutr. 1991, 121, 1146–1162. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.; Dudfield, M. What Is the Relationship between Exercise and Metabolic Abnormalities? A Review of the Metabolic Syndrome. Sport. Med. 2004, 34, 371–418. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.J.; Ellacott, K.L.J.; King, V.L.; Hasty, A.H. Mouse Models of the Metabolic Syndrome. Dis. Model. Mech. 2010, 3, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Arias-Mutis, O.J.; Marrachelli, V.G.; Ruiz-Saurí, A.; Alberola, A.; Morales, J.M.; Such-Miquel, L.; Monleon, D.; Chorro, F.J.; Such, L.; Zarzoso, M. Development and Characterization of an Experimental Model of Diet-Induced Metabolic Syndrome in Rabbit. PLoS One 2017, 12, e0178315. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Rosset, R.; Petersen, O.; Gonzalez, J. Health Outcomes of a High Fructose Intake: The Importance of Physical Activity. J. Physiol. 2019, 597, 3561–3571. [Google Scholar] [CrossRef] [PubMed]
- Denke, M.A.; Pasternak, R.C. Defining and Treating the Metabolic Syndrome: A Primer from the Adult Treatment Panel III. Curr. Treat. Options Cardiovasc. Med. 2001, 3, 251–253. [Google Scholar] [CrossRef]
- Ross, R.; Dagnone, D.; Jones, P.J.H.; Smith, H.; Paddags, A.; Hudson, R.; Janssen, I. Reduction in Obesity and Related Comorbid Conditions after Diet-Induced Weight Loss or Exercise-Induced Weight Loss in Men: A Randomized, Controlled Trial. Ann. Intern. Med. 2000, 133, 92–103. [Google Scholar] [CrossRef]
- Gomes, R.M.; Tófolo, L.P.; Rinaldi, W.; Scomparin, D.X.; Grassiolli, S.; Barella, L.F.; de Oliveira, J.C.; Branco, R.C.S.; Agostinho, A.R.; da Silva Ribeiro, T.A.; et al. Moderate Exercise Restores Pancreatic Beta-Cell Function and Autonomic Nervous System Activity in Obese Rats Induced by High-Fat Diet. Cell. Physiol. Biochem. 2013, 32, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Dias, D.; Bernardes, N.; Stoyell-Conti, F.F.; Dos Santos, C.P.; De Araujo, A.A.; Llesuy, S.; Irigoyen, M.C.; De Angelis, K. Impact of Combined Exercise Training on the Development of Cardiometabolic and Neuroimmune Complications Induced by Fructose Consumption in Hypertensive Rats. PLoS One 2020, 15, e0233785. [Google Scholar] [CrossRef]
- du Sert, N.P.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting Animal Research: Explanation and Elaboration for the ARRIVE Guidelines 2.0. PLOS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef]
- Wisløff, U.; Helgerud, J.; Kemi, O.J.; Ellingsen, Ø. Intensity-Controlled Treadmill Running in Rats: VO2 Max and Cardiac Hypertrophy. Am. J. Physiol. - Hear. Circ. Physiol. 2001, 280, H1301–H131. [Google Scholar] [CrossRef]
- Jones, J.H. Resource Book for the Design of Animal Exercise Protocols. Am. J. Vet. Res. 2007, 68, 1–137. [Google Scholar] [CrossRef]
- Rodrigues, B.; Figueroa, D.M.; Mostarda, C.T.; Heeren, M. V.; Irigoyen, M.C.; De Angelis, K. Maximal Exercise Test Is a Useful Method for Physical Capacity and Oxygen Consumption Determination in Streptozotocin-Diabetic Rats. Cardiovasc. Diabetol. 2007, 6, 1–7. [Google Scholar] [CrossRef]
- Caponi, P.W.; Lehnen, A.M.; Pinto, G.H.; Borges, J.; Markoski, M.; Machado, U.F.; Schaan, B.D. Aerobic Exercise Training Induces Metabolic Benefits in Rats with Metabolic Syndrome Independent of Dietary Changes. Clinics 2013, 68, 1010–1017. [Google Scholar] [CrossRef]
- Abdulla, M.H.; Sattar, M.A.; Abdullah, N.A.; Khan, M.A.H.; Anand Swarup, K.R.L.; Johns, E.J. The Contribution of A1B-Adrenoceptor Subtype in the Renal Vasculature of Fructose-Fed Sprague-Dawley Rats. Eur. J. Nutr. 2011, 50, 251–260. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and Beta-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Scott, M.L.; Thornley, M.J.; Coombs, R.R.A. Comparison of Red-Cell Linked Anti-IgE and 125I-Labelled Anti-IgE in a Solid-Phase System for the Measurement of IgE Specific for Castor Bean Allergen. Int. Arch. Allergy Immunol. 1981, 64, 230–235. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Food intake, weight gain and total caloric intake. The values are presented as the mean ± SEM of 8-10 rats per group. Food intake (A), weight gain (B) and total caloric intake (C) from 30 to 120 days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze the evolution of food intake, weight gain and total caloric intake. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, training factor; and I, interaction between fructose and training factors.
Figure 1.
Food intake, weight gain and total caloric intake. The values are presented as the mean ± SEM of 8-10 rats per group. Food intake (A), weight gain (B) and total caloric intake (C) from 30 to 120 days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze the evolution of food intake, weight gain and total caloric intake. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, training factor; and I, interaction between fructose and training factors.
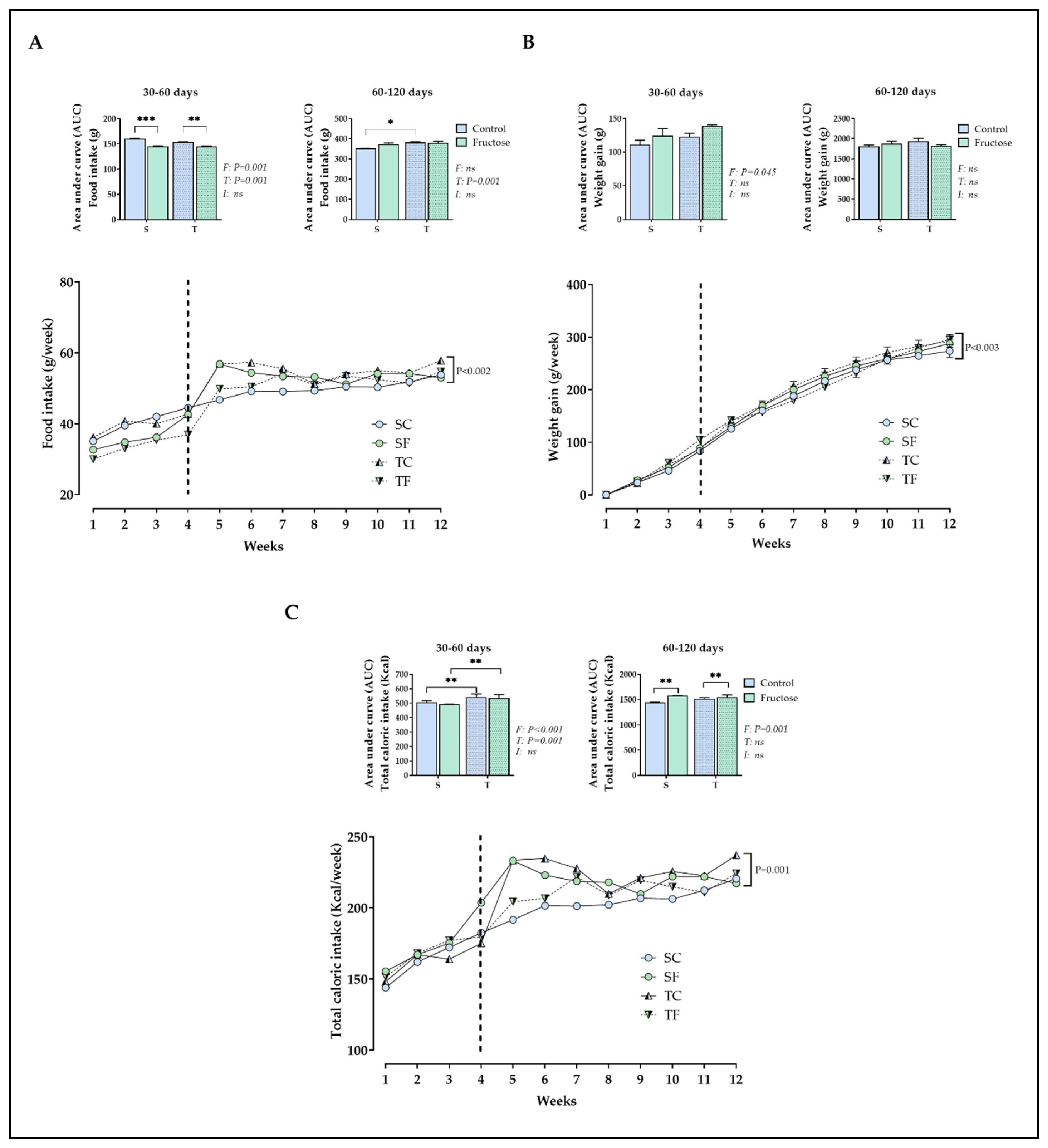
Figure 2.
Glucose during the ipGTT. The values are presented as the mean ± SEM of 6-8 rats per group. Glucose during the ipGTT from 60 (A) and 120 (B) days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze glycaemia after glucose injection. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
Figure 2.
Glucose during the ipGTT. The values are presented as the mean ± SEM of 6-8 rats per group. Glucose during the ipGTT from 60 (A) and 120 (B) days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze glycaemia after glucose injection. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
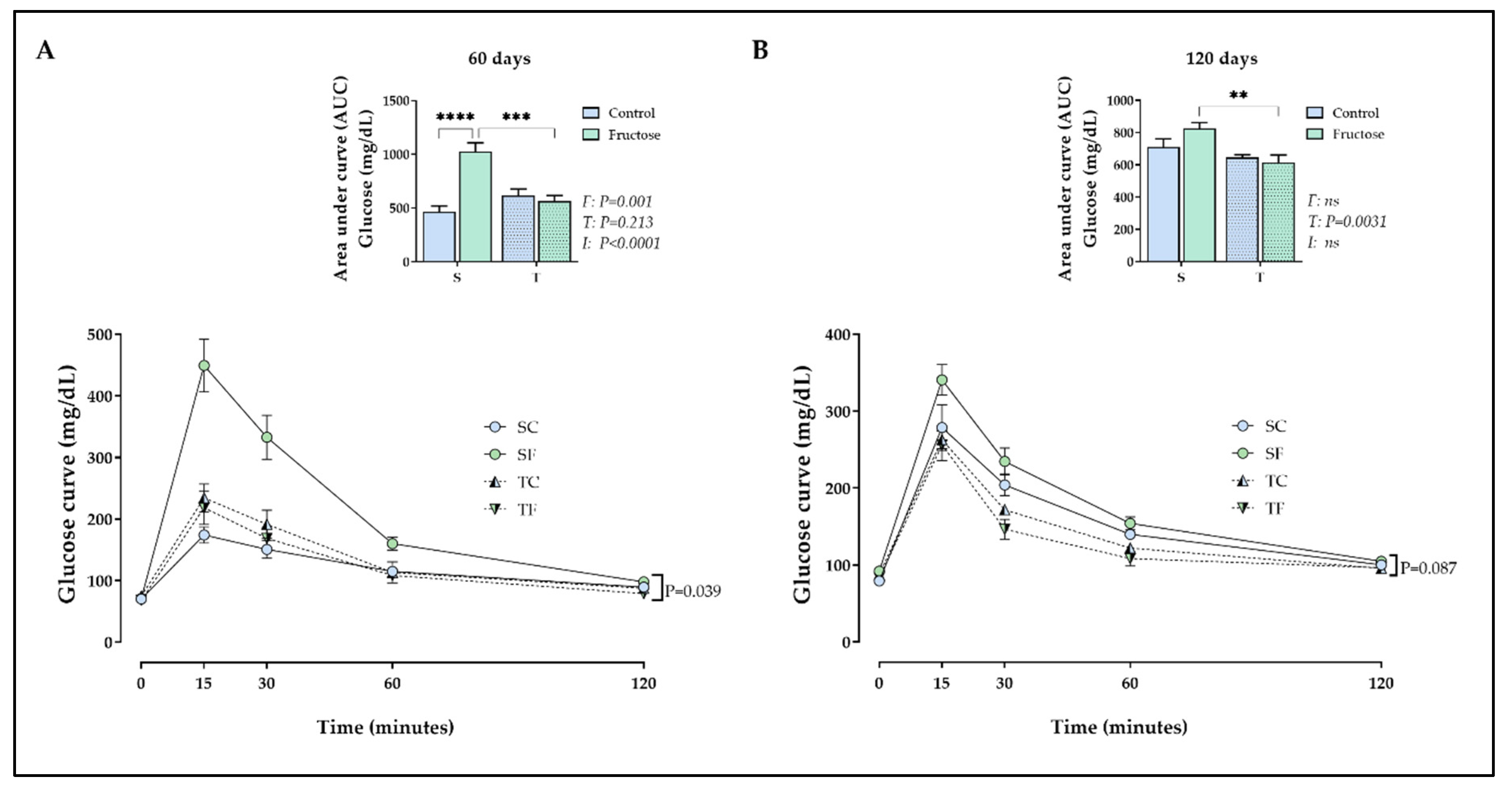
Figure 3.
Insulinemia during the ipGTT. The values are presented as the mean ± SEM 6-8 rats per group. Insulin during the ipGTT from 120 days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze insulinemia after glucose injection. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
Figure 3.
Insulinemia during the ipGTT. The values are presented as the mean ± SEM 6-8 rats per group. Insulin during the ipGTT from 120 days-old. Repeated measures ANOVA with Sidak’s post hoc test was performed to analyze insulinemia after glucose injection. Two-way ANOVA and Tukey’s post hoc test were used for AUCs comparison in each figure. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
Figure 4.
Parasympathetic and sympathetic electrical nerve activity. The values are presented as the mean ± SEM of 5-9 rats per group. Parasympathetic (A) and sympathetic (B) nerve activity at 120 days-old. Two-way ANOVA and Tukey’s post hoc test were used. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
Figure 4.
Parasympathetic and sympathetic electrical nerve activity. The values are presented as the mean ± SEM of 5-9 rats per group. Parasympathetic (A) and sympathetic (B) nerve activity at 120 days-old. Two-way ANOVA and Tukey’s post hoc test were used. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
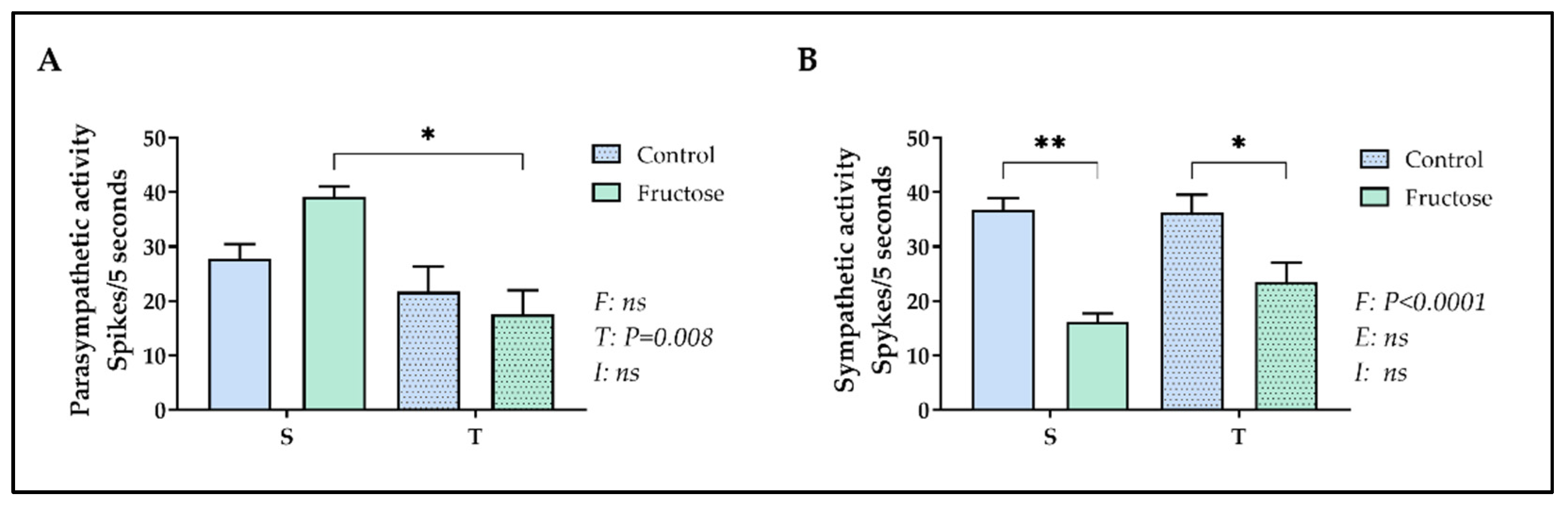

Table 1.
Oximetry and treadmill parameters for Wistar rats at 60 days-old.
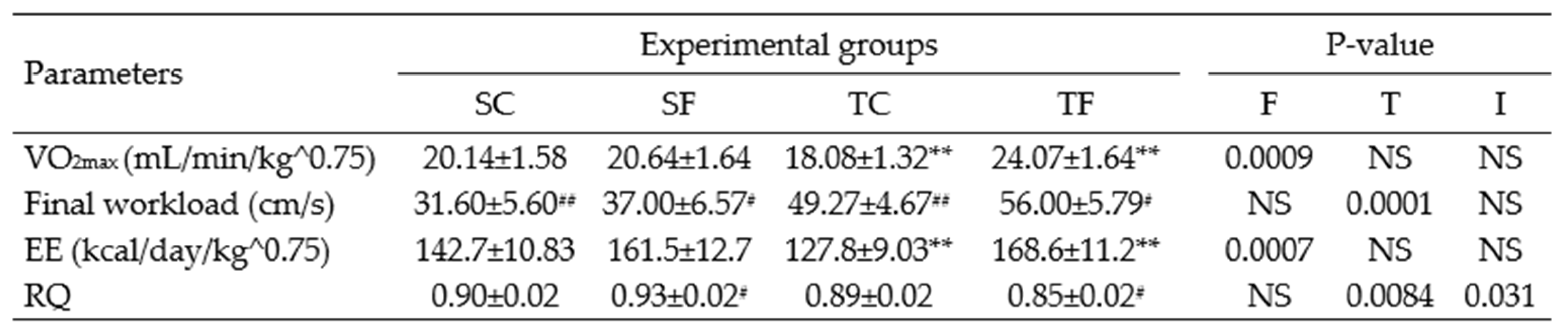
The data represent the mean ± SEM of 10-12 rats per group. Two-way ANOVA and by Tuckey’s post hoc test were used for factors and group comparisons. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors. *Represent difference between SC and SF; **Represent difference between TC and TF; #Represent difference between SF and TF; ##Represent difference between SC and TC.
Table 2.
Oximetry and treadmill parameters for Wistar rats at 120 days-old.
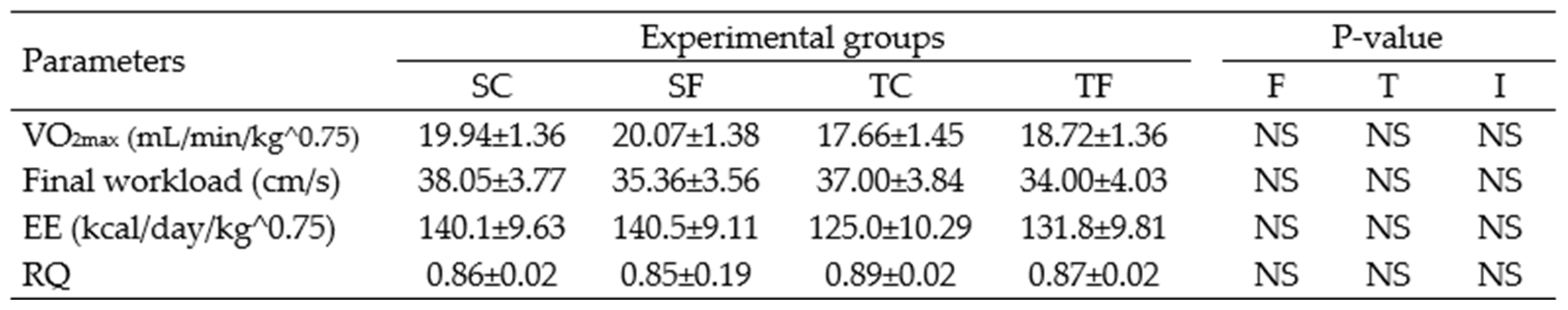
The data represent the mean ± SEM of 10-12 rats per group. Two-way ANOVA and by Tuckey’s post hoc test were used for factors and group comparisons. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors.
Table 3.
Tissues and biochemical parameters for Wistar rats at 60 days-old.
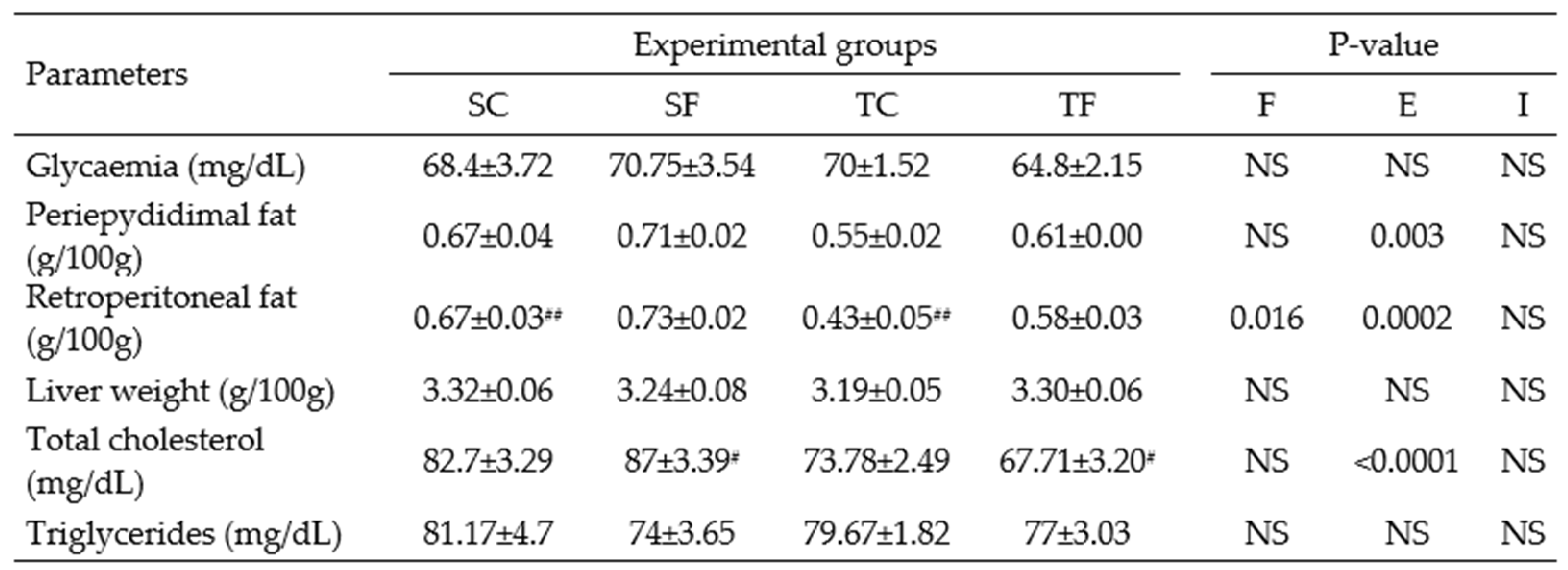
The data represent the mean ± SEM of 6-9 rats per group. Two-way ANOVA and by Tuckey’s post hoc test were used for factors and group comparisons. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors. *Represent difference between SC and SF; #Represent difference between SF and TF.
Table 4.
Tissues and biochemical parameters for Wistar rats at 120 days-old.
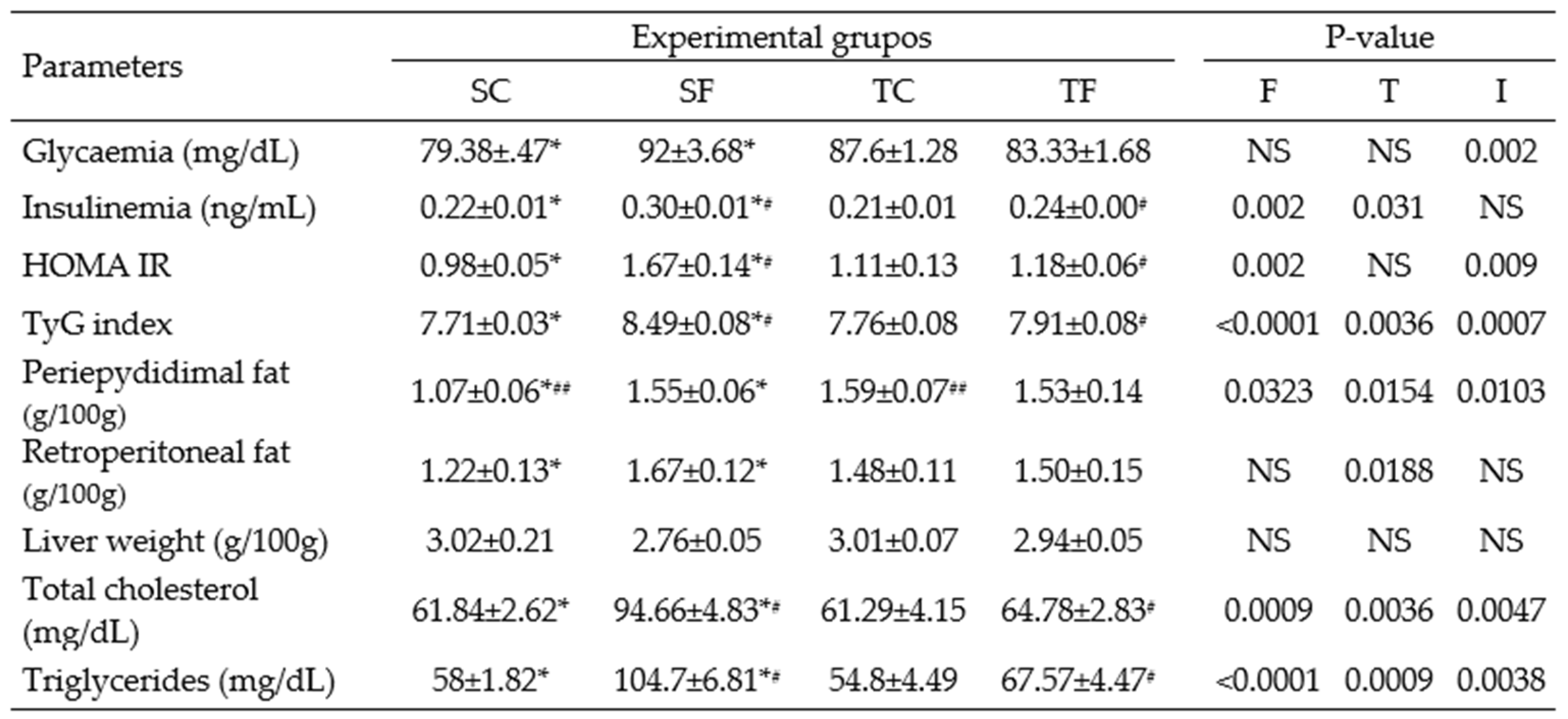
The data represent the mean ± SEM of 6-10 rats per group. Two-way ANOVA and by Tuckey’s post hoc test were used for factors and group comparisons. SC, sedentary rats subjected a normal diet; SF, sedentary rats subjected a fructose diet; TC, trained animals subjected to a normal diet; and TF, trained animals subjected to a fructose diet. F, fructose factor; T, trained factor; and I, interaction between fructose and training factors. *Represent difference between SC and SF; #Represent difference between SF and TF; ##Represent difference between SC and TC.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



