
Submitted:
17 July 2023
Posted:
18 July 2023
You are already at the latest version
Abstract
The key enzyme, 1-hydroxy-2-methyl-2-(E)-butenyl-4-pyrophosphate reductase HDR, responsible for the final step in the MEP pathway, was successfully cloned from the petals of Narcissus tazetta var. chinensis, herein referred to as NtHDR (accession number OQ739816). NtHDR exhibits a full-length coding sequence spanning 1380 base pairs, encoding a total of 460 amino acids. Through phylogenetic tree analysis, it has been determined that NtHDR shared the closest evolutionary relationship with monocotyledons, specifically Asparagus officinalis and Zingiber officinale. Utilizing quantitative real-time PCR analysis, it was observed that NtHDR exhibited significant expression in both petals and corona, with expression levels varying throughout the flowering process of Narcissus. Specifically, in petals, NtHDR expression demonstrated a pattern of initial increase followed by subsequent decrease, while in corona, it consistently increased. Subsequent subcellular localization experiments indicated that NtHDR was localized within the chloroplasts. To investigate the functional impact of NtHDR, stable transformation was performed, where NtHDR was introduced into Nicotiana benthamiana. The resulting transgenic N. benthamiana flowers were analyzed using solid-phase microextraction coupled with gas chromatography–mass spectrometry (SPME-GC-MS) to characterize their volatile components. The analysis revealed the presence of the monoterpene compound linalool, as well as the phenylpropanoids benzyl alcohol and phenylethanol, within the floral fragrance components of the transgenic N.benthamiana plants. However, these compounds were absent in the floral fragrance components of wild-type tobacco plants, thus highlighting the impact of NtHDR transformation on the floral scent profile.
Keywords:
Chinese Narcissus ‘Jinzhanyintai’
; floral scent
; HDR
; GC-MS
; transgenic Nicotiana benthamiana
1. Introduction
Floral fragrance consists of a diverse range of volatile organic compounds (VOCs) [1]. In defense against pathogens, parasites, and herbivores, as well as for attracting pollinators, plants release a vast amounts of phytogenic VOCs [2]. Terpenes represent a pivotal class of plant volatiles, and their metabolic pathways have been extensively elucidated. The precursors of terpenoids, including Isopentenyl pyrophosphate (IPP) and dimethylallyl diphosphate (DMAPP), are synthesized through two distinct routes: the cytoplasmic mevalonate pathway and the plastid-localized methylerythritol 4-phosphate pathway (MEP). Subsequently, terpenoids are generated via the catalytic actions of various terpene synthases [3,4,5,6]. Notably, the MEP pathway predominantly governs the biosynthesis of monoterpenes [7], which undergo a seven-step enzymatic reaction, where 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate reductase (HDR) serves as the final step of the reaction catalase. This last catalytic step precisely regulates the production of IPP and DMAPP mixtures [8,9,10]. Variations in HDR expression have been demonstrated to correlate significantly with alterations in terpene metabolite levels. Studies involving Solanum lycopersicum and Arabidopsis thaliana seeds demonstrated that increased HDR expression led to a corresponding increase in tetra-terpenoid carotenoid content [11]. Furthermore, anatomical investigations of HDR-silenced Nicotiana tabacum leaves have revealed the structural disarray in fenestrated tissue accompanied by a reduction in plastid numbers [12]. In the study of metabolic engineering for the valuable terpenoid tanshinone [13], HDR overexpression was proved to indirectly or directly facilitate the accumulation of these terpenoids. In summary, HDR genes can exert indispensable regulatory roles in plant growth and development, with their overexpression resulting in substantial elevation of indirect precursor levels. This can contribute to the upregulation of biosynthesis pathways for numerous downstream products within the MEP pathway [14].
Chinese Narcissus (Narcissus tazetta var. chinensis M.Roem.), a renowned traditional flower in China, has been cultivated for millennia and is esteemed for its lush green leaves, graceful blossoms, enchanting fragrance, and exceptional ornamental value [15]. Among its significant cultivars, 'Jingzhanyintai' holds a special place. Extensive previous investigations [16,17,18,19,20,21,22] have revealed that the primary components of the floral fragrance volatiles in Chinese Narcissus 'Jingzhanyintai' encompass terpenoids and phenylpropanoids, including monoterpene ocimene, linalool, methyl benzoate, and ethyl benzoate. In recent years, although research on key genes involved in the metabolic pathway of Chinese Narcissus floral fragrance has become an increasing concentration [17,23,24,25,26,27,28], studies that specifically aim to address the HDR remain unreported. To address this knowledge gap, we conducted a comprehensive study where the HDR gene was successfully cloned from the petals of Chinese Narcissus 'Jingzhanyintai' using RT-PCR, leveraging transcriptome data. Subsequent subcellular localization experiments provided insights into its specific function. The expression pattern of the NtHDR was thoroughly analyzed through quantitative real-time PCR analysis. Furthermore, to achieve functional validation, the NtHDR was applied into the generation of Nicotiana benthamiana via transformation, coupled with the employment of the solid-phase microextraction coupled with gas chromatography–mass spectrometry (SPME-GC-MS) technique. This integrated approach established a solid foundation for delving deeper into the floral metabolism and molecular regulatory mechanisms underlying the captivating flower fragrance of Chinese Narcissus.
2. Materials and Methods
2.1. Plant Materials
The plant material utilized in this study comprised Chinese Narcissus cultivar 'Jinzhanyintai' which was purchased from Zhangzhou, Fujian, China. The plants were cultivated within a controlled artificial climate chamber, maintaining a temperature of 20°C for 14 h of light exposure, as well as a temperature of 16°C for 8 h of darkness, alongside a humidity level of 70%. Figure 1 visually presents the varying stages of ‘Jinzhanyintai’ flowering. At different time points, the petals, corona, leaves, bulbs, and roots were promptly subjected to snap-freezing in liquid nitrogen and subsequently stored in a -80°C refrigerator for subsequent analysis.
N. benthamiana was grown in a light incubator at 22°C for 22 h of light and 2 h of dark at 50–70% humidity for transient transformation assays.
2.2. Total RNA Extraction and cDNA Synthesis
The total RNA extraction from 'Jinzhanyintai' petals, corona, bulbs, roots, and leaves was performed through the application of the Quick RNA Isolation Kit (0416-50GX, Huayueyang, Beijing, China), according to the manufacturer’s instructions. The purity and concentration of the obtained RNA samples were subsequently assessed using NanoDrop2000. For cDNA synthesis, a final reaction volume of 20 µL from total RNA (1 μg) was employed, with the utilization of Oligo(dT)15 primers, and AMV Reverse Transcriptase (A3500, Promega, Madison, WI, USA) according to manufacturer’s specifications.
2.3. Cloning and Expression Vector Construction of NtHDR
Using the transcriptome sequencing database as a reference, a homology blast analysis was conducted employing the HDR protein sequence from the model plant Arabidopsis thaliana. Subsequently, specific primers were designed following careful screening of the HDR homologous gene sequence (Table 1). Gene coding sequence (CDS) amplification was conducted with applying PrimeSTAR® HS (Premix) (Takara, Kusatsu, Japan). The PCR amplification system and protocol were as follows: PrimeSTAR® HS (Premix, Kusatsu, Japan) 25 µL, 4 µL cDNA template, 2 µL of a 10 μmol·L-1 mixture of upstream and downstream primers and supplemented with ddH2O to a total volume of 50 µL. The amplification procedure was as follows: 95°C for 3 min; 95°C for 15 s, 56℃ for 15 s, 72°C for 2 min (35 cycles); 72°C for 5 min; and 4°C +∞. Following PCR, the agarose gel containing the target fragment was excised and purified according to the instructions of the DNA Recovery Kit (DP209, Tiangen, Beijing, China). The purified fragment was then ligated into pCE2 TA/Blunt-Zero Vector (Vazyme, Nanjing, China) and transformed into Escherichia coli DH5α receptor state (CB101, Tiangen, Beijing, China).Positive monoclonal colonies were selected and sent for sequencing to Genewiz. Subsequently, NtHDR was ligated to the pBI121 expression vector through the In-Fusion Cloning, with XmaI and XbaI selected as the enzyme cleavage sites. The enzyme cleavage system and timing were operated according to the instructions provided by BioLab. The homologous amplification primers applied are detailed in Table 2. The PCR amplification system and protocol remained consistent with the aforementioned steps, with the amplification template being the plasmid obtained after correct sequencing in the previous step. The PCR products were recovered by agarose gel purification. The homologous recombination system was prepared following the guidelines outlined in the ClonExpress® II One Step Cloning Kit (Vazyme, Nanjing, China): 200 ng of the digested pBI121 vector, 60 ng of PCR product, 4 µL of 5×CEII Buffer, 2 µL of ExnaseII, and 20 µL of ddH2O were mixed and incubated at 37°C for 30 min. The resulting mixture was then transferred into E. coli DH5α receptor state. Positive monoclonal colonies were selected and submitted to Genewiz for sequencing.
2.4. NtHDR Bioinformatics Analysis
The structural domain identification of the proteins was analyzed through adopting the NCBI Conserved Domain Search online tool (https://www.ncbi.nlm.nih.gov/). The amino acid composition, protein molecular weight, theoretical isoelectric point (pI), and stability were predicted through utilizing the online software ProtParam tool (http://web.expasy.org/protparam/). The hydrophobic property and charge distribution of the protein were analyzed via the ProtScale tool (https://web.expasy.org/protscale/). To analyze the transmembrane domain of the NtTPS protein, the TMHMM server 2.0 software (http://www.cbs.dtu.dk/services/TMHMM) was employed. The presence of a signal peptide in NtTPS was predicted with the SignalP4.1 Server (http://www.cbs.dtu.dk/services/SignalP/). The tertiary protein structure predictions were conducted through the application of SWISS-MODEL (https://swissmodel.expasy.org/) platforms, respectively. The sequence similarity of comparison was conducted to analyze the NtHDR sequence results using BLAST from NCBI [32]. The Neighbor-Joining (NJ) method was employed to construct the phylogenetic tree using MEGA 7.0 software with 1000 bootstrap replicates [33,34].
2.5. Fluorescence Quantification of NtHDR in ‘Jinzhanyintai’
Specific amplification primers were designed through applying Primer Premier 6 (Table 1), with primers for reference gene (Actin) selected based on previous studies [35]. Moreover, following the manufacturer’s recommendation, a LightCycler® 480 fluorescent qPCR instrument and TB Green Premix ExTaqII were utilized for conducting the experiments and the qRT-PCR assays, respectively. Furthermore, each experiment was performed with three technical replicates. The relative expression of the NtHDR gene was calculated using the 2-△△Ct method[36]. Correlation analysis between the relative expression of the NtHDR gene in various periods of Narcissus flowering and in diverse plant parts was performed with SPSS 25.
2.6. Subcellular Localization Assay of NtHDR Proteins in Tobacco
Subcellular localization studies of NtHDR were conducted through the agrobacterium-mediated transient expression technique. The plant overexpression vector plasmid pBI121-NtHDR-GFP was transferred into GV3101 agrobacterium tumefaciens competent cells (Shanghai Weidi Biotechnology, Shanghai, China) via heat stimulation. The resulting single colonies were cultured for 60 h and subjected to colony PCR for identification. Positive single colonies were selected and transferred to 20 mL of LB liquid medium containing 50 mg/L kanamycin and 25 mg/L rifampicin, followed by being incubated at 28℃ with shaking at 200 rpm until the OD600 value reached approximately 0.8 [37]. The bacteria were collected, suspended in a formulation containing 10 mM MES-KOH, 10 mM MgCl2, and 150 µM acetosyringone, and left for 2–3 h. The suspension was injected into N. benthamiana leaves and incubated under low light conditions for 48 h. Subsequently, chloroplast autofluorescence was observed and imaged with a LSM 880 (ZEISS) laser confocal microscope, with an excitation wavelength of 633 nm and an emission wavelength of 684 nm. Additionally, GFP fluorescence was observed and imaged with an excitation wavelength of 488 nm and an emission wavelength of 533 nm [38,39] (Zhang et al., 2022; Liu, 2017). The plant expression vector plasmid pBI121-GFP was utilized as a positive control and treated as pBI121-NtHDR-GFP.
2.7. Agrobacterium-Mediated Transformation and Identification of N. benthamiana
The plant overexpression vector plasmid pBI121-NtHDR was transferred into GV3101 agrobacterium tumefaciens competent cells through heat stimulation, followed by colony PCR to confirm successful transformation. The subsequent culture procedures were the same as described in Section 4.6. The bacteria were collected, suspended in the MS liquid medium, and allowed to incubate for 2–3 h.
Under aseptic conditions in the ultra-clean table, healthy leaves from well-cultured sterile N. benthamiana seedlings were carefully excised into 0.8 cm × 0.8 cm sized leaves (avoiding the veins as much as possible) using sterilized knives. These leaves were then placed onto a culture medium containing MS+6-BA 2.0 mg/L+NAA 0.2 mg/L and kept in the dark for a period of 2 d. Pre-cultured N. benthamiana leaves were subsequently immersed in a prepared agrobacterium solution for approximately 10 min, during which the solution was continuously agitated to ensure optimal contact with the leaves. After completion, excess bacterial solution was removed using sterile filter paper, and the leaves were rinsed 2–3 times with sterile water. Subsequently, the leaves were placed on sterile filter paper to dry. A new pre-culture medium was prepared by placing three layers of sterile filter paper in a flat position, and the pre-treated leaves were positioned backside up on the filter paper layers. This setup was maintained in the dark for an additional 2 d. Once adventitious buds started to grow, the leaves were transferred to a normal light condition and the culture medium was changed approximately once every 10 d. The adventitious buds presenting excellent growth were excised and transferred to a differentiation culture medium containing MS+NAA 0.2 mg/L + Kan 100 mg/L + Tim 300 mg/L. For rooting, the adventitious buds were cultured in a 1/2MS+Kan 100 mg/L + Tim 300 mg/L medium until they developed 2–3 leaves and reached sufficient height. Once the root system became adequately grown, the cap of the culture bottle was loosened, and sterile water was added for a period of 3–4 d under low light conditions. Prior to transplanting the seedlings into small plastic pots, the medium on the roots was carefully cleaned, and the seedlings were covered with transparent glass or plastic containers for a period of 10 d after transplantation. This step aimed to maintain a high ambient humidity, allowing the seedlings to gradually adapt to the new environment.
Transgenic N. benthamiana DNA was extracted following the protocol outlined in the DNA Extraction Kit (MF735-plus, Juhemei, Beijing, China) to ensure high-quality DNA samples Subsequently, to validate the expression of NtHDR at the protein level in the transgenic plants, petals were carefully selected from plants that had been identified as positive for DNA. Moreover, RNA extraction was performed with the RNA Extraction Kit, followed by reverse transcription to synthesize cDNA. The PCR identification process utilized specific primers, namely 35SF and NtHDR-R (Table 2), to amplify the target sequence.
2.8. Analysis of Volatile Components in Transgenic N. benthamiana Flowers
Entirely open flowers of transgenic and wild-type N. benthamiana were selected with three replicates performed for each assay). The samples were prepared by mixing approximately 0.2 g of randomly selected N. benthamiana flowers in a clean 10 mL round-bottomed headspace vial at room temperature (25±2°C). The vial was sealed using a blue Teflon silicone spacer, leaving about 2/3 of the space at the top. Analysis was performed using a 50/30 μm DVB/CAR/PDMS solid-phase microextraction head and HP-5MS (30 m × 250 μm × 0.25 μm) flexible quartz capillary column with a GC-MS model Agilent 7890A-5975C. The headspace solid-phase microextraction parameters were adjusted based on the work of Sun Miao [40] with appropriate modifications. The aging temperature and time of the extraction head were set at 250°C for 1 h. The extraction process was conducted at 50°C for 30 min, followed by a desorption time of 3 min. The injection port was of the auto injection type. Helium gas was used as the carrier gas, with a column flow rate of 1 mL/min in the pulsed non-split injection mode. The heating temperature of the injection port was maintained at 250°C. For MS analysis, the EI ionization mode was employed, with an electron energy of 70 eV. The ion source and the four-stage rod temperature were set at 230°C and 150°C, respectively. The full scan acquisition mode covered a scan range of 50–600 amu, and the EMV mode was set to the relative value. The equilibration time for the gas phase column chamber was 0.25 min. The ramp-up procedure was as follows: an initial temperature of 55°C held for 3 min, followed by a ramp-up rate of 3°C/min to 148°C for 1 min, then a ramp-up rate of 10°C/min to 210°C for 3 min, and finally a post-run temperature of 55°C for 1 min.
The identification of compounds involved a two-step approach: retention index method and qualitative analysis of volatiles in N. benthamiana flowers with the application of the NIST14 spectral library. In addition, manual spectral analysis was performed, taking into account the mass spectra and retention time indexes reported in relevant published literature. This comprehensive analysis facilitated the identification of specific volatile components.
3. Results
3.1. Characterization of NtHDR
The ORF of NtHDR spanned 1380 bp, encoding a protein consisting of 460 amino acid residues. The NtHDR can be classified under the Isoprenoid_Biosyn_C1 superfamily. The analysis with the ProtParam online tool revealed that the molecular weight of NtHDR was approximately 51.38kDa. The theoretical pI of the protein was calculated to be 5.58. Furthermore, the protein instability coefficient was determined to be 32.35, indicating its relatively unstable nature. The mean hydrophilicity (GRAVY) value of -0.372 suggested that NtHDR possessed a hydrophilic property (Figure 2C). Prediction analyses conducted through the application of Signa lP4.1 Server and TMHMM indicated the absence of signal peptide sites and transmembrane regions in the NtHDR protein, thereby categorizing it as a non-secreted and non-transmembrane protein (Figure 2A,B). To visualize the protein’s 3D structure, SWISS-MODEL was utilized, resulting in a model exhibiting approximately 81.21% similarity to the reference model (Figure 2D).
3.2. Phylogenetic Analysis and Sequence Alignment of NtHDR
The functional domain analysis revealed that NtHDR was attributed to the lytB-ispH superfamily of genes, with the encoded protein exhibiting a highly conserved region. Further analysis of NtHDR sequence homology indicated the highest amino acid sequence similarity with A. officinalis at 88.35%. Comparatively, Z. officinale, Elaeis guineensis, Musa acuminata subsp. malaccensis, and Ananas comosus presented 83.97%, 83.3%, 81.03%, and 80.99% gene similarity, respectively (Figure 3). These findings suggested a significant homology of HDR across various species, with strong conservation observed at the C-terminus of the encoded protein. Phylogenetic tree analysis, performed using MEGA7.0 (Figure 4), demonstrated the relationship between HDR genes of 'Jinzhanyintai' and other species. NtHDR was found to be closely associated with A. officinalis (XP_020241009.1), Z. officinale (XP_042395642.1), Dendrobium officinaleKimura (AGH62555.1), Oncidium flexuosum (ACJ83116.1), E. guineensis (XP_010909277.1), Phoenix dactylifera (XP_008799160.1), M. acuminata subsp. Malaccensis (XP_009412299.1), and A. comosus (XP_020104485.1), as well as other monocotyledons, forming a closely clustered branch. Notably, the closest relatives were observed to be A. officinalis and Z. officinale.
3.3. NtHDR Expression Specificity Analysis
Quantitative real-time fluorescence analysis revealed the expression pattern of NtHDR in various tissues, including petals, corona, leaves, bulbs, and roots, indicating certain variations among them (Figure 5A). Notably, NtHDR exhibited higher expression levels in petals and corona while it presented lower expressions in other tissues, with a ranking order of petals > corona > leaves > roots > bulbs. Moreover, examining the expression level of NtHDR in varying flowering stages revealed interesting trends. In petals, the expression level demonstrated an increasing and then decreasing trend during blooming (Figure 5B), while in the corona, the expression level presented a continuous increasing trend throughout the flowering process (Figure 5C). Overall, the expression pattern of NtHDR demonstrated a regular association with the flowering stage, and the pronounced increase in expression level observed in the corona suggested a potential involvement of NtHDR in the metabolism of floral terpenoids.
3.4. NtHDR Subcellular Localization Analysis
The pBI121-NtHDR-GFP plasmid-containing Agrobacterium GV3101 strain was infiltrated into N. benthamiana leaves, while the pBI121-GFP plasmid-containing Agrobacterium GV3101 strain served as a control. Subsequent observation by utilizing laser confocal microscopy revealed that the fusion protein of the target gene HDR and GFP generated green fluorescence exclusively on chloroplasts (Figure 6). In the control group, a distinct and bright green fluorescence signal of GFP was identified within the nucleus and cytoplasm, which indicated the localization of NtHDR on chloroplasts. This observation aligned with the knowledge that the MEP pathway, a metabolic pathway involved in floral metabolism, occurred within cellular chloroplasts, as predicted by the website and previous knowledge.
3.5. Stable Expression of NtHDR in N. benthamiana
A PCR-based screening method was employed to screen a total of 10 transgenic N. benthamiana plants overexpressing NtHDR, with wild-type plants serving as controls for subsequent investigations (Figure 7). Comprehensive observations, including plant size, leaf morphology, flower color, petal size, and florescence, were conducted on both NtHDR overexpressing N. benthamiana plants and wild-type plants. However, no significant differences were detected between the two groups. The breeding process of transgenic N. benthamiana plants is illustrated in Figure 8 below.
3.6. Determination and Analysis of Floral Volatiles of Transgenic N. benthamiana
Volatile compounds from the flowers of transgenic N. benthamiana plants and wild-type plants were collected using headspace solid-phase microextraction and subsequently analyzed using GC-MS. The analysis revealed discernible differences in the volatile components between the two plant groups(Table 2). Specifically, the transgenic N. benthamiana plants expressing NtHDR exhibited the release of linalool (RT=13.297), benzyl alcohol (RT=10.690), and phenethanol (RT=14.151) as evident in the chromatogram (Figure 9-B). Conversely, these three volatile components were absent in the wild-type plants (Figure 9-A).

Table 2.
Main volatile components in fresh flower of different strain N.benthamiana.
| Main volatile components | Retention time/min | Relative content/% | |||||
|---|---|---|---|---|---|---|---|
| A Wild-type N. benthamiana | B Transgenic N. benthamiana | ||||||
| CK1 | CK2 | CK3 | H1 | H2 | H3 | ||
| Benzyl alcohol | 10.690 | — | — | — | — | 1.288 | 1.687 |
| Phenylacetaldehyde | 10.832 | 1.682 | 1.001 | — | 2.652 | 4.247 | 1.105 |
| Linalool | 13.297 | — | — | — | 0.551 | 1.086 | 0.777 |
| Phenethyl alcohol | 14.151 | — | — | — | — | 1.812 | — |
| Decamethylcyclopentasiloxane | 15.963 | 10.766 | 10.488 | 7.715 | 16.885 | 12.974 | 11.461 |
| 2-Isobutyl-3-methoxypyrazine | 16.893 | 0.870 | — | — | 1.003 | 1.940 | — |
| Cinnamic aldehyde | 20.923 | 3.203 | — | 0.233 | 4.047 | — | 4.133 |
4. Discussion
In this study, we cloned an HDR enzyme gene from Chinese Narcissus 'Jinzhanyintai' and successfully introduced it into N. benthamiana plants to achieve stable transformation. The floral fragrance components of the transgenic plants were investigated using SPME-GC-MS. Our analysis revealed the presence of the monoterpene volatile linalool, as well as the phenylpropanoids benzyl alcohol and phenylethanol, in the NtHDR overexpression plants, whereas these components were undetectable in the wild-type plants. In addition, our findings align with previous research conducted by Pang Hongdong [29] who transferred the SAMT enzyme gene serving as a key component in the metabolic pathway of phenylpropanoids/phenylcyclic compounds in Rhamnus phyllanthus into N. tabacum. Pang Hongdong observed higher levels of hexenal, benzyl alcohol, and linalool, while methyl benzoate was not detected. The author hypothesized that the introduction of CpSAMT may have disrupted the synthesis pathway of methyl benzoate in N. benthamiana, thereby leading to more accumulation of benzyl alcohol. Hence, considering the outcomes of our study, we speculated that the production of linalool, benzyl alcohol, and phenylethanol may suggest an unknown association between the phenylpropanoid/phenylcyclic compound metabolic pathway and the terpene metabolic pathway. However, due to the incomplete understanding of the phenylpropanoid/phenylcyclic compound metabolic pathways, further investigations are necessary to elucidate the specific mechanisms underlying their interplay with terpene metabolic pathways. Furthermore, Zhang Tengxun [30] transferred the DXS enzyme gene, an essential component of the MEP pathway in Lilium brownii, into N. tabacum and observed significantly higher linalool content in the transgenic flowers compared to wild-type plants. Similarly, in our experiment, the introduction of the key enzyme gene HDR from the MEP pathway into N. benthamiana resulted in increased linalool content in the transgenic plants’ flowers. Chen Jindou [31] transferred the putative phenylpropane transcription factor CpODORANT into N. tabacum and suggested that the monoterpene ocimene was only present in the control and wild-type N. tabacum flowers, whereas it could not be detected in the flowers of transgenic CpODORANT N. tabacum. Conversely, a small amount of sesquiterpenes geraniene and germacrene D were produced. This observation may help explain the complexity of the floral aroma regulation mechanism in N. tabacum. Based on the aforementioned findings, we inferred that the overexpression of NtHDR in N. benthamiana plants led to increased accumulation of IPP and DMAPP, the floral precursor substances in the MEP pathway. Consequently, this upregulation affected the downstream formation of the monoterpene compound linalool, as well as the phenylpropanoid metabolic pathway in N. benthamiana, resulting in the production of benzyl alcohol and phenylethanol. Overall, our findings provide preliminary evidence supporting the significant regulatory role of the HDR gene in the floral metabolic pathway at the transgenic level.
5. Conclusions
In this study, HDR, a key enzyme gene located at the end of the MEP pathway, was cloned from the petals of Nar. tazetta var.chinensis 'Jinzhanyintai' and named NtHDR. NtHDR played a role in the floral biosynthesis process and was capable of promoting the synthesis of terpene volatiles.
Author Contributions
Conceptualization, Meng Hu, Tao Hu and Jun Ma; Data curation, Meng Hu, Na Zhang and Yun Deng; Formal analysis, Meng Hu and Na Zhang; Investigation, Meng Hu, Na Zhang, Bo Zhang and Hua Wang; Methodology, Meng Hu, Na Zhang, Yan Chang, Yun Deng, Tao Hu and Jun Ma; Resources, Meng Hu, Na Zhang, Yan Chang and Bo Zhang; Software, Meng Hu, Na Zhang and Ke Fan; Supervision, Meng Hu; Validation, Meng Hu and Na Zhang; Visualization, Meng Hu; Writing – original draft, Meng Hu and Na Zhang; Writing – review & editing, Meng Hu and Na Zhang.
Funding
This research was supported by ICBR Fundamental Research Funds Grant (NO. 1632020021 and NO. 1632022013).
Data Availability Statement
All data in this study could be found in the manuscript.
Acknowledgments
We thank all the colleagues that helped with the development of different parts of this manuscript and the State Key Laboratory of Tree Genetics and Breeding for providing equipment and technical support for this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: Nature’s diversity and ingenuity. Science, 2006, 311, 808–811. [CrossRef]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science, 2001, 291, 2141–2144. [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J, 2011, 66, 212–229. 66. [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol, 2013, 198, 16–32. [CrossRef]
- Kitaoka, N.; Lu, X.; Yang, B.; Peters, R. J. The application of synthetic biology to elucidation of plant mono-, sesqui-, and diterpenoid metabolism. Mol. Plant, 2015, 8, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.T.; Cui, J.B.; Hu, H.L.; Xue, J.Y.; Yang, J.J.; Xu, J. Integrated four comparative-omics reveals the mechanism of the terpenoid biosynthesis in two different overwintering Cryptomeria fortunei phenotypes. Front. Plant Sci, 2021, 12, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Adam, P.; Hecht, S.; Eisenreich, W.; Kaiser, J.; Gräwert, T.; Arigoni, D.; Bacher, A.; Rohdich, F. Biosynthesis of terpenes: Studies on 1-hydroxy-2-methyl-2-(E)-butenyl 4-diphosphate reductase. Proc. Natl. Acad. Sci. U. S. A, 2002, 99, 12108–12113. [CrossRef]
- Altincicek, B.; Duin, E.C.; Reichenberg, A.; Hedderich, R.; Kollas, A.K.; Hintz, M.; Wagner, S.; Wiesner, J.; Beck, E.; Jomaa, H. LytB protein catalyzes the terminal step of the 2-C-methyl-D-erythritol-4-phosphate pathway of isoprenoid biosynthesis. FEBS Lett, 2002, 532, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Wolff, M.; Seemann, M.; Tse Sum Bui, B.; Frapart, Y.; Tritsch, D.; Garcia Estrabot, A.; Rodríguez-Concepción, M.; Boronat, A.; Marquet, A.; Rohmer, M. Isoprenoid biosynthesis via the methylerythritol phosphate pathway: The (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase (LytB/IspH) from Escherichia coli is a [4Fe-4S] protein. FEBS Lett, 2003, 541, 115–120. [CrossRef]
- Botella-Pavía, P.; Besumbes, Ó.; Phillips, M.A.; Carretero-Paulet, L.; Boronat, A.; Rodríguez-Concepción, M. Regulation of carotenoid biosynthesis in plants: Evidence for a key role of hydroxymethylbutenyl diphosphate reductase in controlling the supply of plastidial isoprenoid precursors. Plant J, 2004, 40, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Page, J.E.; Hause, G.; Raschke, M.; Gao, W.Y.; Schmidt, J.; Zenk, M.H.; Kutchan, T.M. Functional analysis of the final steps of the 1-deoxy-D-xylulose 5-phosphate (DXP) pathway to isoprenoids in plants using virus-induced gene silencing. Plant Physiol, 2004, 134, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.P.; Shi, R.J.; Tao, R.; Fang, Q.; Jiang, X.Y.; Ji, H.W.; Feng, L.; Huang, L.Q. Cloning, molecular characterization and functional analysis of 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphate reductase (HDR) gene for diterpenoid tanshinone biosynthesis in Salvia miltiorrhiza Bge. f. alba. Plant Physiol. Biochem, 2013, 70, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Wu, M.J.; Wang, X.; Lin, Y.; Cai Y.P.; Fan, H.H. Cloning and expression analysis of 1-hydroxy-2-methyl-2-(E)-butenyl-4-diphosphatereductase gene in Dendrobium officinale. Chinese Traditional and Herbal Drugs, 2015, 46, 405–411. (in Chinese).
- Jiang, Z.; Peng, Z. Chinese Narcissus; China Forestry Publishing House: Beijing, China, 2014; ISBN 978-75-0386-911-2. [Google Scholar]
- Gao, J. The study on mutagenic effects and mechanism of 60CoY-ray on Chinese narcissus. Chinese Academy of Forestry, Beijing, 2000.
- Peng, A.M. Analysis on volatile components of Narcissus tazetta var. chinensis and influence factors. Chinese Academy of Forestry, Beijing, 2010. (in Chinese).
- Dai, L.; Yang, L.; Guo, Y.; Peng, Q. Study on the chemical composition of the essential oil of Narcissus tazetta var. chinensis. Chromatography, 1990, 6, 377–380. (in Chinese). [Google Scholar]
- Huang, Q.Q.; Feng, J.Y. Study of aroma changes in daffodils by adsorption filament/chromatography/mass spectrometry. Chinese Journal of Analytical Chemistry, 2003, 31, 1408. (in Chinese). [Google Scholar]
- Huang, Q.Q.; Feng, J.Y. Study on the variation in narcissus aroma composition during blossoming. Journal of Instrumental Analysis, 2004, 23, 110–113. (in Chinese).
- Dou, Y.J.; Zhai, J.; Hou, F.M.; Leng, P.S.; Wang, W.F.; Hu, Z.H. Effect of Different Light Intensities on the Floral Aroma Emitted from Chinese Daffodil (Narcissus tazetta L. var. chinensis Roem). Acta Agriculturae Boreali-Occidentalis Sinica 2014, 23, 85–91. (in Chinese). [Google Scholar]
- Song, G.; Xiao, J.; Deng, C.; Zhang, X.; Hu, Y. Use of solid-phase microextraction as a sampling technique for the characterization of volatile compounds emitted from Chinese daffodil flowers. J. Anal. Chem, 2007, 62, 674–679. [Google Scholar] [CrossRef]
- Wu, Y. Study on Cloning of HDS gene and resistant transcription factor of Narcissus tazetta. Fujian Agriculture and Forestry University, Fuzhou, 2016. (in Chinese).
- Zhong, X. The study ofrelationship of B-glucosidase andvolatile aromatic components of Narcissus tazetta L. var. Chinensis Roem. Fujian Agriculture and Forestry University, Fuzhou, 2014. (in Chinese).
- Zheng, Q.B. Clone of the NtSAMT Gene in Narcissus tazetta L. Var. chinensis roem and transformation of Asarina procumbens. Northeast Agricultural University, Haerbin, 2016. (in Chinese).
- Li, K. Fragrance component analysis and genecloning in Narcissus tazetta. Fujian Agriculture and Forestry University, Fuzhou, 2014. (in Chinese).
- Ma, C. Cloning of DXPS, DXR and PAL related to floral fragrance from Narcissus tazetta L. var. chinensis Roem. Fujian Agriculture and Forestry University, Fuzhou, 2010. (in Chinese).
- Li, L. Analysis of genes related to the terpenoids synthesis pathway in Narcissus tazetta 'Yunxiang' based on Transcriptome data. Fujian Agriculture and Forestry University, Fuzhou, 2019. (in Chinese).
- Pang, H.D.; Xiang, L.; Zhao, K.G.; Li, X.; Yang, N.; Cheng, L.Q. Genetic transformation and functional characterization of Chimonanthus praecox SAMT gene in tobacco. Journal of Beijing Forestry University, 2014, 36, 117–122. (in Chinese). [Google Scholar]
- Zhang, T. Molecular cloning and functional analysis of the key enzyme LiDXS genes involved in monoterpene biosynthesis in lily. Beijing Forestry University, Beijing, 2017. (in Chinese).
- Chen, J.D. Founction research of odorant in chimonanthus praecox. Central China Agricultural University, Wuhan, 2016. (in Chinese).
- Altschup, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol, 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol, 1987, 4, 406–425. [CrossRef]
- Wang, G. Transcriptome analysis of flowers and functional studies of MYB genes offlavonoid biosynthetic pathway in Chinese narcissus (Narcissus tazetta var. chinensis). Fujian Agriculture and Forestry University, Fuzhou, 2018. (in Chinese).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method. Methods, 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Meng, N.; Liu, Y.L.; Dou, X.X.; Liu, H.L.; Li, F.Y. Transient gene expression in Phalaenopsis aphrodite petals via Agrobacterium tumefaciens infiltration. Acta Botanica Boreali-Occidentalia Sinica, 2018, 38, 1017–1023. (in Chinese). [Google Scholar]
- Zhang, C.G.; Ge, Y.Y.; Liu, H.H.; Zong, Y.X.; Wu, X.J.; Yang, L.C.; Li, H.G. Cloning and bioinformatics analysis of LtuHMGS in Liriodendron tulipifera. Journal of Central South University of Forestry & Technology, 2022, 42, 146–155. (in Chinese). [Google Scholar]
- Liu, G. Differential regulation of catechin metabolism in albino and normal tea (Camellia sinensis) leaves and tea linalool biosynthesis. Anhui Agricultural University, Hefei, 2017. (in Chinese).
- Sun, M. Cloning and functional analysis of TPS gene family in tree peony fragrance. Beijing Forestry University, Beijing, 2021. (in Chinese).
Figure 1.
Narcissus tazetta ‘Jinzhanyintai’ in various flowering stages. (I–V) represent the overall state, as well as tissue samples of petals and corona for each of the five distinct periods of ‘Jinzhanyintai’ flowering.
Figure 1.
Narcissus tazetta ‘Jinzhanyintai’ in various flowering stages. (I–V) represent the overall state, as well as tissue samples of petals and corona for each of the five distinct periods of ‘Jinzhanyintai’ flowering.

Figure 2.
(A) Transmembrane analysis; (B) Signal peptide analysis; (C) Hydrophilic and hydrophobic analysis; (D) Tertiary structure prediction.
Figure 2.
(A) Transmembrane analysis; (B) Signal peptide analysis; (C) Hydrophilic and hydrophobic analysis; (D) Tertiary structure prediction.
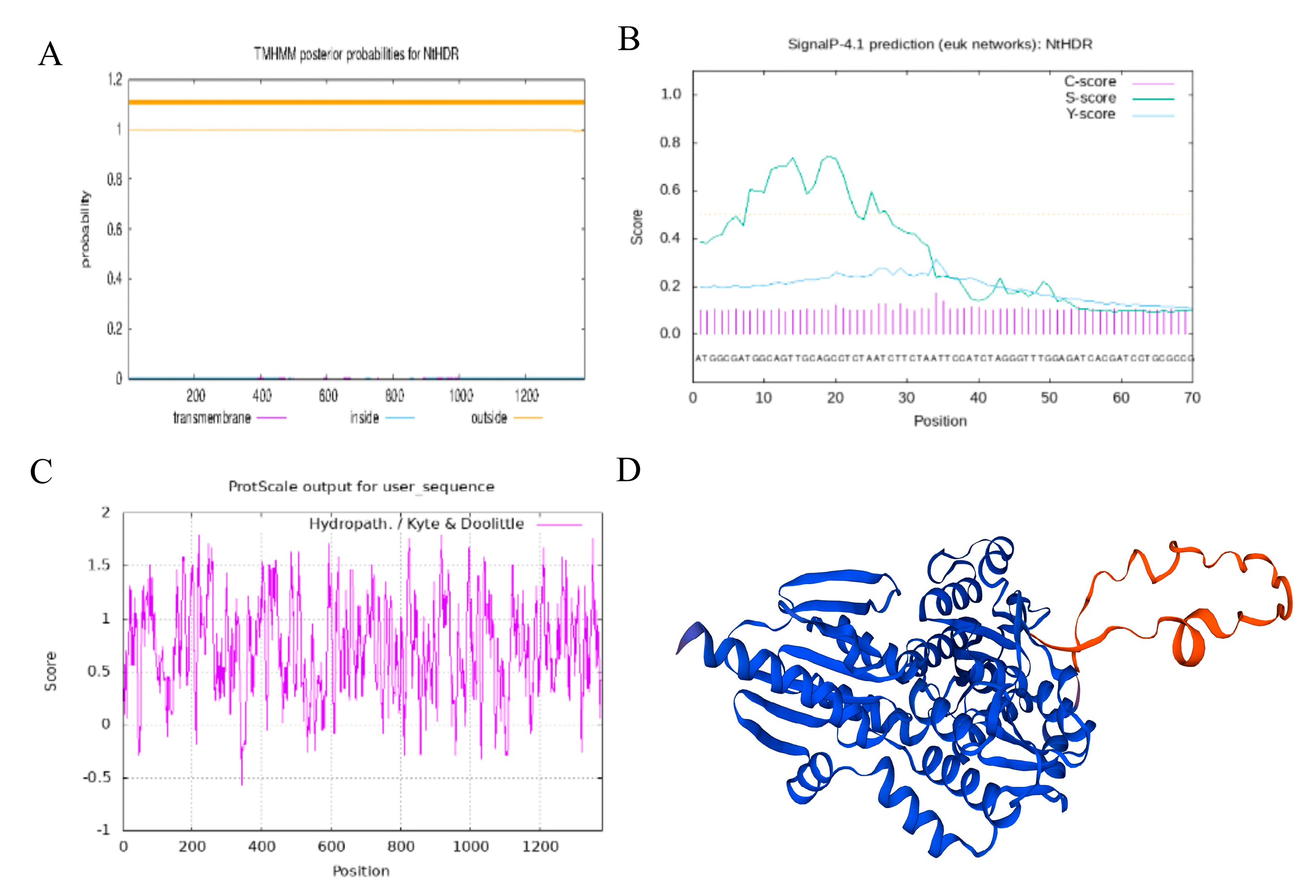
Figure 3.
Multiple alignments of HDR amino acid sequences of 'Jinzhanyintai' with those of other species.
Figure 3.
Multiple alignments of HDR amino acid sequences of 'Jinzhanyintai' with those of other species.

Figure 4.
Phylogenetic analysis of NtHDR protein selected from other plant species.
Figure 5.
Analysis of relative expression level of NtHDR. (A) Relative expression level of NtHDR in various tissues; (B) Relative expression level of NtHDR at different flowering stages of petals and corona; (C) Relative expression level of NtHDR at different flowering stages of corona; lowercase letters (a, b, and c) indicate significant differences observed between different treatments (p < 0.05).
Figure 5.
Analysis of relative expression level of NtHDR. (A) Relative expression level of NtHDR in various tissues; (B) Relative expression level of NtHDR at different flowering stages of petals and corona; (C) Relative expression level of NtHDR at different flowering stages of corona; lowercase letters (a, b, and c) indicate significant differences observed between different treatments (p < 0.05).
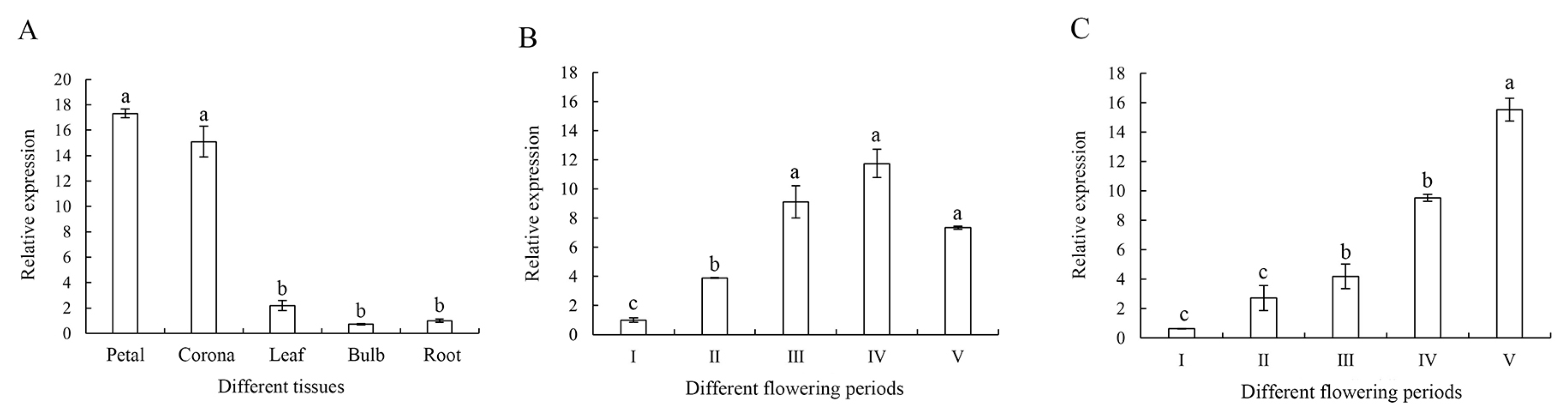
Figure 6.
Confocal images of transiently expressed GFP fusions in leaves of N. benthamiana.
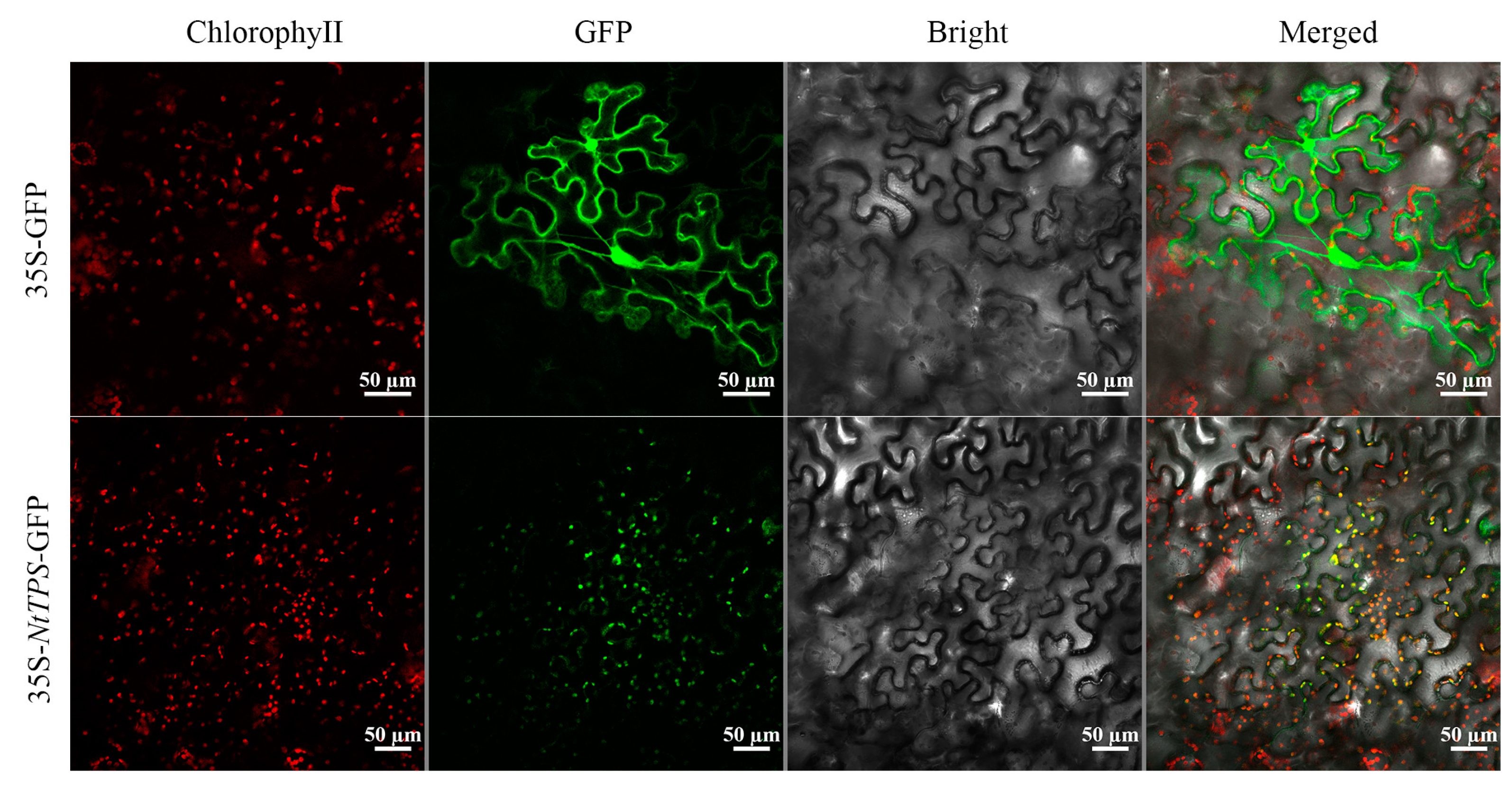
Figure 7.
PCR assay of transgenic N. benthamiana plants. M: DL2000 marker; 1: Wild-type N. benthamiana; 2–11: Transgenic N. benthamiana.
Figure 7.
PCR assay of transgenic N. benthamiana plants. M: DL2000 marker; 1: Wild-type N. benthamiana; 2–11: Transgenic N. benthamiana.
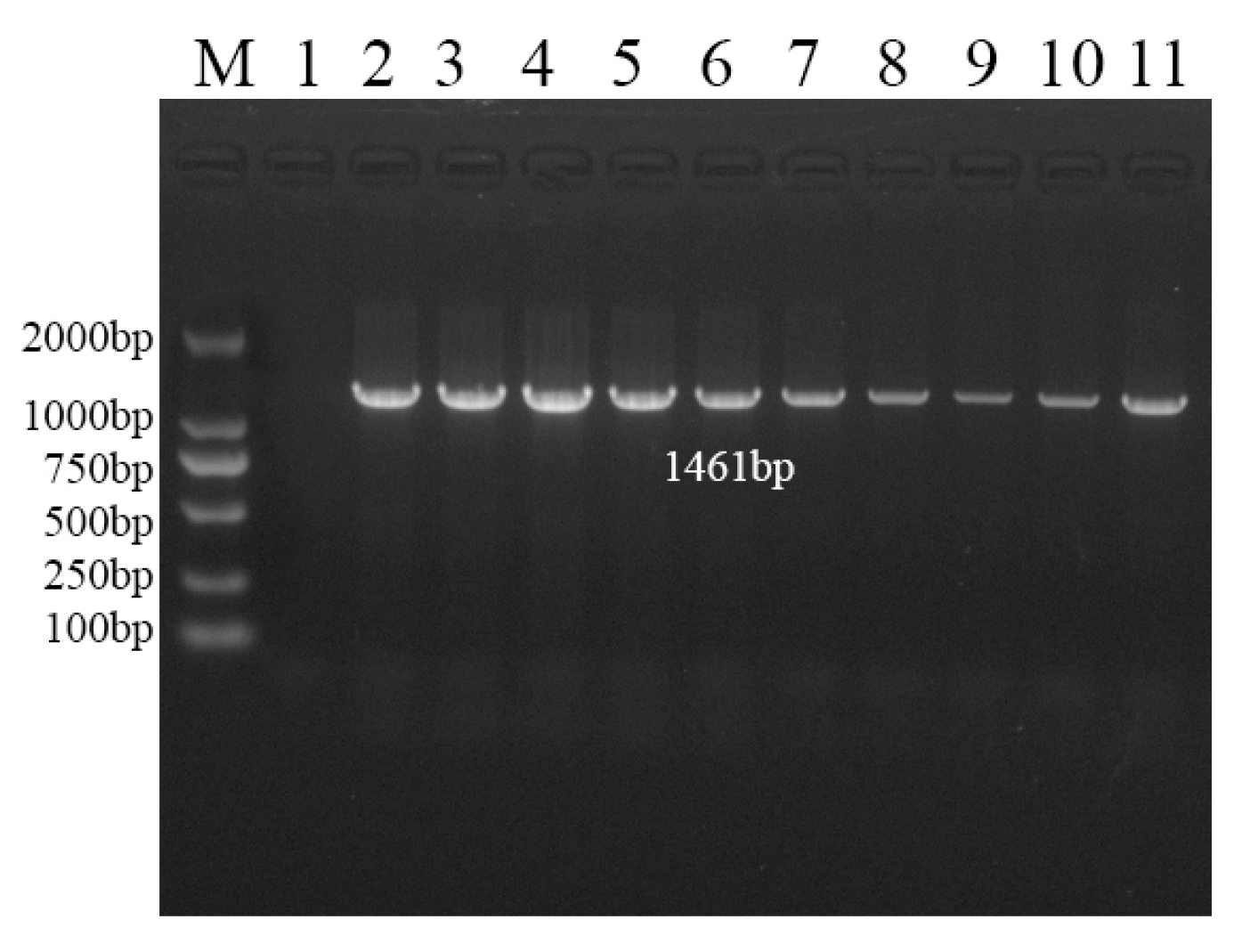
Figure 8.
Cultivar of transgenic N. benthamiana with pBI121-NtHDR expression vector. The diameter of culture dish is 90 mm, and the diameter and height of culture bottle are 66 mm × 95 mm. (A) Preculturing of wild-type N. benthamiana; (B–C) Screening of wild-type N.benthamiana with kanamycin; (D) Differentiation culture of N. benthamiana; (E–F) Rooting culture; (G) Flowering of transgenic N. benthamiana; (H) Flowering of wild-type N. benthamiana.
Figure 8.
Cultivar of transgenic N. benthamiana with pBI121-NtHDR expression vector. The diameter of culture dish is 90 mm, and the diameter and height of culture bottle are 66 mm × 95 mm. (A) Preculturing of wild-type N. benthamiana; (B–C) Screening of wild-type N.benthamiana with kanamycin; (D) Differentiation culture of N. benthamiana; (E–F) Rooting culture; (G) Flowering of transgenic N. benthamiana; (H) Flowering of wild-type N. benthamiana.

Figure 9.
Total ion chromatograms of volatile compounds in fresh flower of different N. benthamiana strains. (A) Wild-type N. benthamiana; (B) Transgenic N. benthamiana with pBI121-NtHDR expression vector. The unlabeled spurious peak is the extractor head coating off.
Figure 9.
Total ion chromatograms of volatile compounds in fresh flower of different N. benthamiana strains. (A) Wild-type N. benthamiana; (B) Transgenic N. benthamiana with pBI121-NtHDR expression vector. The unlabeled spurious peak is the extractor head coating off.
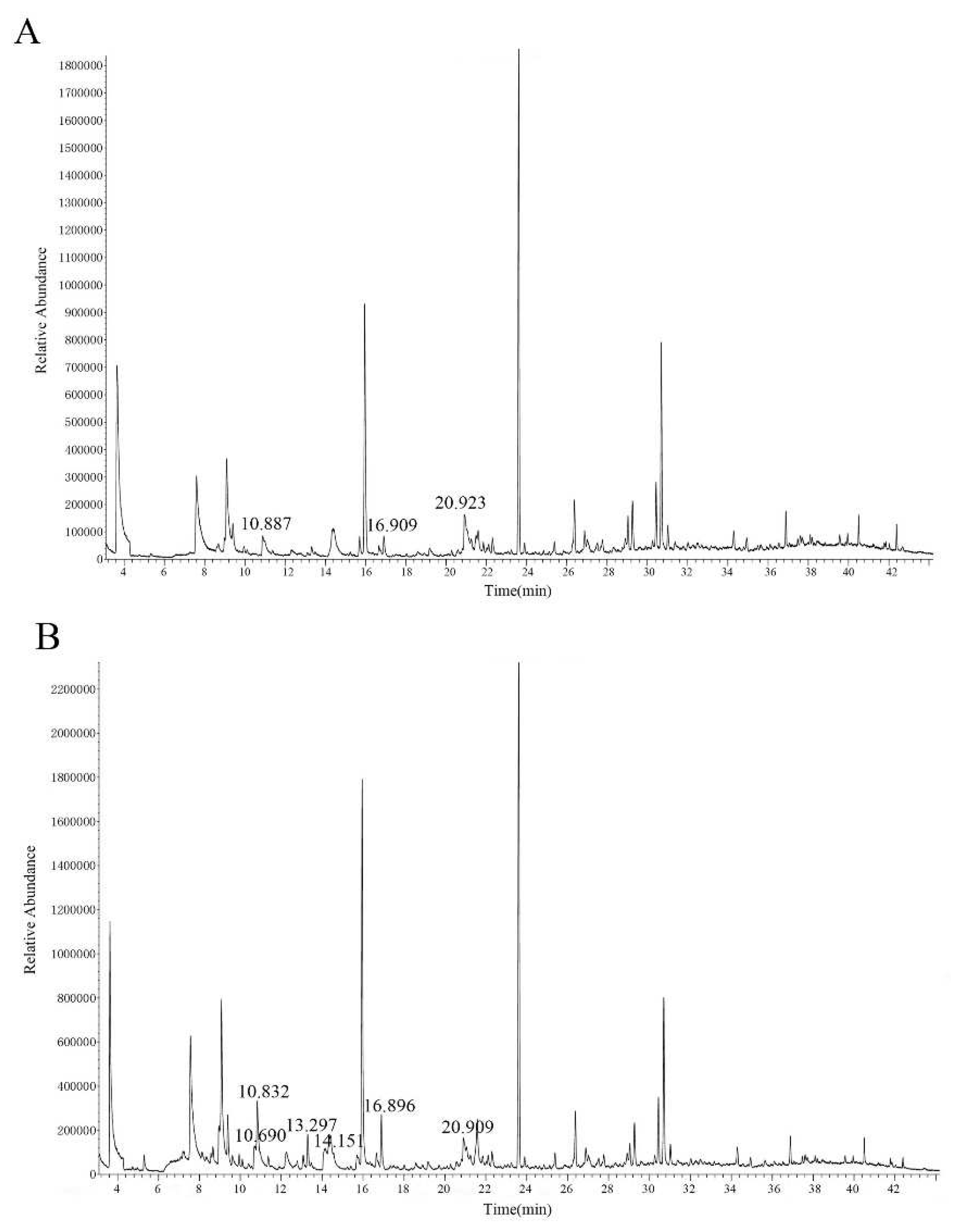
Table 1.
Primers sequences used for PCR.
| Primer use | Primer name | Sequences (5’→3’) |
|---|---|---|
| Cloning the CDS | NtHDR-F | atggcgatggcagttgcag |
| NtHDR-R | cgccaactgcatagattcttcc | |
| Subcellular localization analysis | pBI121-NtHDR-F | tggagagaacacgggggactatggcgatggcagttgcag |
| pBI121-NtHDR-R | ataagggactgaccacccgccaactgcatagattcttcc | |
| Stable transformation of N. benthamiana | pBI121-NtHDR-R2 | ataagggactgaccaccttacgccaactgcatagattcttcc |
| Real-time PCR | NtActin-F | tgcccagaagtgctattccag |
| NtActin-R | gttgacccaccactaagaacaatg | |
| qPCR -NtHDR-F | tgacgaggggcgataactacaatc | |
| qPCR -NtHDR-R | gactctgctagcttcaccgttac | |
| Identification of positive transgenic N. benthamiana | 35s-F | gacgcacaatcccactatcc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



