
Submitted:
16 July 2023
Posted:
18 July 2023
You are already at the latest version
Abstract
Spent mushroom compost (SMC) substrates are commonly used as growth media for greenhouse crops and horticulture production. This study aimed to investigate the responses of physiochemical soil properties, enzyme activities, and microbial community compositions to different cultivation durations and SMC soil treatments on tomatoes (Solanum lycopersicum L.) grown in plastic greenhouses on the Loess Plateau, China. The experiment included treatments: Two control treatments of non-planting SMC substrates (Sub CK) and continuous mono-cropping soil (Soil CK), and SMC substrate and the surrounding soil after planting at 1, 3, and 7 years. The results revealed that the SMC substrates had higher contents of major nutrients (e.g., total N and total P) than the surrounding soil treatments. The physicochemical soil properties and soil enzyme activities of the SMC substrates were significantly decreased with longer cultivation duration. Microbial alpha-diversity was higher in the SMC substrates regardless of cultivation duration than the control treatment (soil CK and substrate CK). Interestingly, following several years of tomato cultivation, the compositions of bacterial communities had more similarities with fungal communities in both the SMC substrates and surrounding soils. It was observed that many beneficial microbes, such as bacteria of the Deinococcus-Thermus, Halanaerobiaeota, and Nitrospirae phyla, and the fungi of the Basidiomycota, Mortierellomycota, and Chytridiomycota phyla were enriched in the SMC substrates. The pathogenic bacterial genus Sphingomonas and fungal genus Fusarium were abundant in the Soil CK treatment, while the potentially beneficial bacterial genera Saccharimonadales, Gaiella, Bacillus, and fungal genera Thermomyces, Kernia, and Mortierella were abundant in both the SMC substrate and surrounding soil. This study demonstrated that agro-based SMC substrates were a suitable growth media for a new grooved cultivation system.
Keywords:
spent mushroom compost (SMC) substrate
; cultivation years
; physicochemical properties
; enzyme activities
; microbial community
1. Introduction
With rapidly intensifying climate change and an increasing global population, food security is a serious global scale issue [1]. The frequent occurrence of extreme weather events and imbalanced food supply patterns exacerbate the challenges that impact economic and social development worldwide [2,3]. To narrow the gap between food supplies and conventional open-field production, greenhouse technologies have been employed for the cultivation of crops and horticulture plants that can improve the production of off-season products [4]. Greenhouse cultivation has the capacity to finely control temperatures and microclimates to facilitate the year-round growth of horticulture plants to compensate for climatic instabilities in the field [5]. However, greenhouse technologies are a significant energy-intensive agricultural industry [6]. In the pursuit of high crop yields, surplus inputs of fossil-based mineral fertilizers can have deleterious effects on soil, leading to environmental contamination in greenhouse cultivation systems [7]. Hong et al. [8] suggested that excessive nutrient accumulation in plant root zones and downward leaching via irrigation result in soil and groundwater contamination in greenhouse cultivation systems. Furthermore, increased soil salinity, decreased crop yields, and continuous cropping obstacles (CCO) result in bottleneck issues for the development of the vegetable industry in greenhouses [9,10].
In seeking an alternative system to replace conventional greenhouse cultivation, researchers have invested more effort into potential alternatives. The application of agro-based spent mushroom compost substrates has been shown to be a beneficial alternative approach for the cultivation of crops and horticultural plants in greenhouse systems [11]. Spent mushroom compost (SMC) substrates are an effective cultivation media for vegetable production [12], which can reduce nitrogen loss in greenhouse systems [13]. The spent compost substrate consisted primarily of residual fungal mycelium, disintegrated lignocellulosic biomass, various nutrients, as well as organic matter and enzymes. Further, it had a low bulk density, loose texture, good air permeability, and nutrient retention. Thus, the SMC can be employed to bioremediate contaminated soil, and enhance its health [14] by improving the physical structure of the soil and ecological environment for soil microorganisms [15]. Furthermore, the SMC also contains a high organic matter content, phosphorus, and potassium, as well as trace elements that are required for plant assimilation and utilization [16]. Owing to its low level of toxic elements, strong absorption capacity, and enhanced soil aeration and water retention capacities, SMC has been utilized as a soil amendment that contributes to improving soil quality, agronomic efficiency, and environmental safety [15,17]. In addition, it contains several biologically active compounds such as therapeutically valuable polysaccharides and antibacterial peptides. These, together with its antibacterial properties protect plants against pathogens; thus, minimizing the incidence of plant diseases [18,19].
Under conventional greenhouse conditions, horticultural systems are subject to reduced soil organic matter, declining soil quality and fertility, and decreased microbial biomass [20,21,22]. Further, increased soil salinity, nutrient leaching, and soil compaction can result in continuous cropping obstacles (CCOs), which inhibit the prospects for increased production in greenhouse systems [8,10,23]. Although the use of SMC substrates as a biofertilizer, soil amendment, and for the bioremediation of pollution have attracted much attention [15], its role as a growth media for the cultivation of crops and horticultural plants in greenhouses is not well understood [11]. Unal [12] suggested that the application of SMC as a cultivation media exhibited positive impacts on the quality of tomato seedlings, plant growth, available organic matter, and nutrient uptake of plants [24,25]. Thus, there is a need to determine exactly how SMC influences the growth of tomato plants in greenhouses, inclusive of soil microbial communities and enzyme activities under different cultivation durations.
For this study, we investigated the impacts of SMC under varying cultivation timelines on physicochemical soil properties, soil enzyme activities, and the composition of microbial communities. We hypothesized that under greenhouse conditions: a) the application of agro-based SMC as growth media enhances the contents of major nutrients, enzyme activities, and microbial alpha-diversity in contrast to continuous mono-cropping soil; b) soil physicochemical properties and enzyme activities decrease with longer cultivation timelines; c) the composition of bacterial communities are more similar than fungal communities between the growth substrate and surrounding soil along with longer cultivation timelines; d) abundant beneficial microbes can be enriched by the SMC substrate and surrounding soil, which contribute to soil amelioration and improved plant growth.
2. Materials and Methods
2.1. Experimental design and soil sampling
This study was conducted in Hongtong County, Shanxi Province, on the Loess Plateau, in Northwestern China (35°87′84″N, 111°28′91″E). The annual temperature is 12.7℃, with 2079.1 hours of sunlight, 441.5 mm of precipitation, and 210 frost-free days. In May 2013, our group began to employ the SMC substrate as growth media to cultivate tomatoes (Solanum lycopersicum L.) in a plastic greenhouse. This compost consisted of spent mushroom compost cylinders of oyster mushroom (45%~47%), cow manure (45%~47%), and fragmented residues of tomato plants (6%~10%). The SMC substrates were completely dried under sunlight and used as the growth media in a groove model system (Figures S1 and S2). Thus, various SMC substrate model cultivation durations were continually observed in the testing area. A non-planted substrate was also sampled prior to the experiment, which served as a control (Sub CK). Further, a continuous mono-cropping soil under greenhouse conditions was considered as another control treatment (Soil CK), to assess the effects of the SMC substrate amendment. We selected three different greenhouse tomato cultivation durations as the subjects of this study: Substrate after planting 1 year (Sub 1yr); surrounding soil of 1-yr substrate (Soil 1yr); substrate after planting 3 years (Sub 3yr); surrounding soil of 3-yr substrate (Soil 3 yr); substrate after planting 7 years (Sub 7yr); surrounding soil of 7-yr substrate (Soil 7 yr).
Each experimental plot was established in an independent greenhouse with an area of ~0.1 ha. All experimental plots received the same management based on the standard recommended practices for growing tomato plants in greenhouses. The physicochemical soil properties and soil microbe characteristics, as well as the surrounding soil (situated at the bottom and sides of the substrate) were quantified between all treatments. Five soil cores were extracted from each experimental plot (substrates and surrounding soil, respectively) using a soil-drilling sampler (10 cm inner diameter) on October 30, 2020. Five cores from the same plots were combined to form four mixed samples, for a total of sixteen soil samples and sixteen substrate samples. All samples were sifted through a 2 mm mesh sieve, after which each sample was divided in half. One half was stored in a 50 mL centrifuge tube with liquid N, and immediately transported to the laboratory pending DNA extraction. The other half was used for the quantification of physicochemical soil properties and soil enzyme activities.
2.2. Measurement of soil physicochemical properties and enzyme activities
The soil pH was determined from the supernatant of 0.01 M CaCl2 soil slurries (1:1 (w/v)) after 10 min of vigorous shaking and soil particle settling [26]. The soil organic matter (SOM), total phosphorus (P), and total nitrogen (N) were determined via chromic acid titration, as well as the MADAC and Conway methods, respectively [27,28,29]. The soil organic carbon (C) was measured using an Isoprime isotope ratio mass spectrometer with a Eurovector elemental analyser [30]. Further, the soil alkaline phosphatase (ALP) enzyme activities were determined using a photometric technique [31]. The soil protease (Pro) and β-glucosidase (β-Glu) enzyme activities were calculated using a solid phase enzyme linked immunosorbent assay (ELISA) method with a protease ELISA Kit and β-Glucosidase ELISA Kit in soil infused biological fluids [32].
2.3. DNA extraction, PCR, and amplicon sequencing
The total microbial genomic DNA were extracted from 0.5 g of a soil sample using the OMEGA Soil DNA Kit (M5635-02) (Omega Bio-Tek, Norcross, GA, USA) according to the manufacturer’s instructions [33]. Using a NanoDrop NC-2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis, the quantity and quality, respectively, of extracted DNA were assessed. The PCR amplification of the bacterial 16S rRNA gene V3-V4 region was performed using the forward primer 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and the reverse primer 806R (5’-GGACTACHVGGGTWTCTAAT-3’). The fungal internal transcribed spacer (ITS) gene V1 region was determined using the forward primer (5’-GGAAGTAAAAGTCGTAACAAGG-3’) and the reverse primer (5’-GCTGCGTTCTTCATCGATGC-3’). The PCR components contained 5 µL of buffer (5×), 0.25 µL of Fast pfu DNA polymerase (5 U/µL), 2 µL of 2.5 mM dNTPs, 1 µL (10 µM) of each forward and reverse primers, 1 µL of DNA template, and 14.75 µL of ddH2O. The thermal cycling program proceeded as follows: initial denaturation at 98℃ for 5 min, followed by 25 cycles comprised of denaturation at 98℃ for 30 s, annealing at 55℃ for 30 s, and extension at 72℃ for 45 s, with a final extension of 5 min at 72℃. The PCR amplicons were purified using Vazyme VAHTSTM DNA Clean Beads (Vazyme, Nanjing, China) and quantified with a Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA, USA). The purified amplicons were pooled in equal amounts and paired-end sequenced (2×500 bp) using the Illumina MiSeq platform with MiSeq Reagent Kit v3 (Shanghai Personal Biotechnology Co., Ltd, Shanghai, China).
2.4. Data processing and bioinformatics analyses
Microbiome bioinformatics were run in QIIME2 2019.4 (http://docs.qiime2.org/2019.4/tutorials) according to the standardized protocols [34]. Briefly, raw FASTQ data were demultiplexed using the demux plugin followed by primer cutting with the cutadapt plugin [35], which were then quality filtered, denoised, and merged, with chimera removed using the DADA2 plugin [36]. The remaining high-quality amplicon sequence variants (ASVs) were submitted to the SRA (Sequence Read Archive) at the National Center for Biotechnology Information (NCBI) under accession number SUB12292980 (biosample SAMN31710080) for 16S bacteria sequences and number SUB12293105 (biosample SAMN31710019) for ITS fungi sequences. Non-singleton ASVs data were aligned with mafft [37] and used to construct a phylogeny with fasttree2 [38]. Alpha-diversity metrics (Chao1 [39], Shannon [40]), beta-diversity metrics (weighted UniFrac [41], unweighted UniFrac [42], and Bray-Curtis dissimilarity) were estimated using the diversity plugin. Taxonomy was assigned to ASVs using the classify-sklearn naïve Bayes taxonomy classifier of the feature-classifier plugin [43] against the SILVA_132 database for bacteria, and UNITE_8.0 database for fungi [44].
Sequence data analyses were primarily performed using the QIIME2 and R packages (v3.2.0). The ASV-level alpha-diversity indices (Chao1 and Shannon) were calculated using the ASV table in QIIME2 and visualized as box plots. Beta-diversity analysis was performed to investigate the structural variations in microbial communities using Bray-Curtis metrics [45] and UniFrac distance metrics [41]. These were visualized using principal coordinate analysis (PCoA) and an unweighted pair-group method with arithmetic means (UPGMA) hierarchical clustering [46]. The significance of the differentiation of microbiota structures between groups was assessed by PERMANOVA (Permutational multivariate analysis of variance) [47], ANOSIM (Analysis of similarities) [48], and Permdisp [49] using QIIME2. The taxonomy compositions and abundances were visualized using MEGAN [50] and GraPhlAn [51]. A Venn diagram was created for visualization using the R package “VennDiagram” [52]. The relative abundances of taxa were statistically compared between samples or treatments by MetagenomeSeq, and visualized as Manhattan plots [53]. LEfSe (Linear discriminant analysis effect size) was performed to investigate differentially abundant taxa across groups using the default parameters [54]. Random forest analysis was performed to discriminate the samples from different groups using QIIME2 [55,56].
2.5. Statistical analysis
Data analysis was performed using SPSS statistics 19.0 software (SPSS Inc.). Analysis of variance was performed to assess the impacts of soil physicochemical properties, enzyme activities, microbial community diversity, and composition on the different treatments. The differences between means between the various treatments were calculated via the Tukey-HSD test at a probability level of 0.5 Spearman’s rank correlations were used to correlate the physicochemical soil properties, enzyme activities, and soil microbial communities.
3. Results
3.1. Physicochemical soil properties
Significant influences of the cultivation duration were observed for the physicochemical properties of the SMC substrate and surrounding soil (Table 1). The organic matter, pH, and organic C content were significantly higher in the Sub 1yr treatment than the Sub 3yr and Sub 7yr treatments; however, there were no differences seen between Sub 1yr the Sub CK treatments. With increased cultivation duration, the organic matter, total N, and organic C content were significant decreased, with the lowest values being observed in the Sub 7yr treatment. The total P and pH were higher in the Sub 1yr treatment than the Sub 3- and 7yr treatments, which had no differences between them. For the surrounding soil, the Soil 3yr treatment possessed higher total P than the other treatments. The organic matter and organic C content were both highest under the Soil CK treatment and decreased with the cultivation duration, but showed no significant difference among them. Soil 1yr treatment exhibited the highest pH over the other treatments, which did not differ among themselves. Moreover, the total P, total N, organic matter, and organic C content were significantly higher for the substrate than in the surrounding soil with the cultivation duration (Table 1).
3.2. Soil enzyme activities
The enzyme activities were significantly different between the substrates and surrounding soils (Table 2). Soil alkaline phosphatases (ALP), protease (Pro), and β-Glucosidase (β-Glu) enzyme activities were highest in the substrate for the Sub 1yr treatment and significantly decreased with the cultivation duration. For the surrounding soil, the soil ALP enzyme activities were higher in the Soil 3yr treatment and lower under the Soil 7yr treatment, whereas they did not differ for the soil CK. The Soil Pro enzyme activity was highest under the Soil 1yr treatment, but decreased significantly with longer cultivation durations. The soil β-Glu enzyme activity was highest in the Soil 7yr treatment than the other treatments. Further, the Soil Pro enzyme activities were significantly decreased with longer cultivation durations between the substrate and surrounding soil (Table 2). With longer cultivation durations, the soil β-Glu enzyme activities were significantly decreased in the substrates and conversely for the surrounding soil. The soil ALP enzyme activities had a higher peak value in the Sub 1yr and the Soil 3yr treatments.
3.3. Microbial alpha- and beta-diversity
The microbial alpha-diversity Chao1 and Shannon indices were significantly different between the substrate and surrounding soil under respective cultivation durations (Figure 1). In particular, the soil microbial (fungal and bacterial) α-diversity of the Shannon index were significantly different between the Sub CK and Sub 7yr treatments for the substrate (p = 0.021 and p = 0.0011, respectively). For the substrate, the bacterial α-diversity Chao1 and Shannon indices were significantly different between the Sub CK and Sub 3yr treatments (Figure 1a).
The bacterial and fungal beta-diversity differed between the substrate and surrounding soil under the respective cultivation durations (Figure 2). The bacterial beta-diversity in the Sub CK and Soil CK treatments were significantly different from the other treatments. Following 1-, 3-, and 7- years of cultivation, the bacterial structural compositions of the substrates and in the surrounding soil exhibited a similar trend. The bacterial structural composition under the Sub 7yr treatment was quite similar to the other treatments (Figure 2a). The Sub 1-, 3-, and 7- year treatments differed in terms of their fungal community compositions in the substrates, and obviously differed from the Sub CK treatment (Figure 2b). Interestingly, the fungal structural compositions were not significantly different between Sub 7yr and Soil 3yr treatments.
3.4. Microbial community composition
For the substrates, the bacterial taxa of the top-10 most abundant phylum strongly fluctuated with the cultivation duration (Table 3, Figure S3). The Sub CK treatment possessed a higher abundance of Chloroflexi, Bacteroidetes, and Deinococcus-Thermus phyla than did the other treatments and had a decreased trend with longer cultivation durations. In contrast, the abundances of Proteobacteria, Actinobacteria, Firmicutes, and Gemmatimonadetes phyla showed an increased trend with the cultivation duration, especially the Acidobacteria phyla. Further, the predominant fungal taxa of the top-5 most abundant phylum were significantly different between treatments. The abundance of the Ascomycota phylum was higher under the Sub 1yr treatment, while the Mortierellomycota phyla was higher under the Sub 3yr treatment. However, the Sub 7-year treatment exhibited a higher abundance of Olpidiomycota phylum than did the other treatments.
For the surrounding soil, the abundances of Proteobacteria, Firmicutes, Chloroflexi, Cyanobacteria, Bacteroidetes, and Patescibacteria bacterial phyla varied strongly between treatments (Table 3, Figure S3). The Soil 7yr treatment had a higher abundance of Firmicutes, the Soil 3-yr treatment showed higher abundances of Chloroflexi and Patescibacteria phyla, and the Soil CK treatment exhibited higher abundances of Cyanobacteria and Bacteroidetes phyla. The dominant soil fungal taxa of Ascomycota, Mortierellomycota, and Olpidiomycota phyla were significantly different between treatments. The Soil 7yr treatment showed a higher abundance of Mortierellomycota, Soil 1yr treatment had a higher abundance of Olpidiomycota, and the Soil CK treatment had a higher abundance of Ascomycota phylum compared to the other treatments.
The heat map and clustering analysis of bacterial and fungal community compositions of the top-20 most abundant genera (Figure 3) revealed that the four dominant bacterial genera (AKYG1722 (of Chloroflexi phyla), A4b (of Chloroflexi phyla), Chryseolinea (of Bacteroidetes phyla), and SBR1031 (of Chloroflexi phyla)) (Figure 3a), and two fungal genera (Melanocarpus and Sodiomyces of the Ascomycota phyla) had significantly higher abundances under the Sub CK treatment (Figure 3b). The Sub 1yr treatment showed higher abundances of bacterial genera (KD4-96 (Chloroflexi phyla), Saccharimonadales (Patescibacteria phyla), 67-14 (Actinobacteria phyla)), and fungal genera (Melanocarpus, Ascobolus, Thermomyces, and Iodophanus (Ascomycota phyla)). The Sub 3yr treatment had higher abundances of bacterial genera (Saccharimonadales (Patescibacteria phyla), Nocardioides (Actinobacteria phyla), Subgroup_6 (Acidobacteria phyla), and Gitt-GS-136 (Chloroflexi phyla)) (Figure 3a), and fungal genera (Kernia (Ascomycota phyla), Mortierella, and Acaulium (Ascomycota phyla)). The dominant bacterial genera (67-14 (Actinobacteria phyla), Nocardioides (Actinobacteria phyla), S085 (Chloroflexi phyla), and Gaiella (Actinobacteria phyla)), and fungal genera (Remersonia, Aspergillus, Sodiomyces, Lophotrichus, and Fusarium (Ascomycota phyla)) had significantly higher abundances under the Sub 7yr treatment (Figure 3b). Furthermore, the Soil CK treatment revealed higher abundances of the dominant bacterial genera (Dongia and Sphingomonas (Proteobacteria phyla)), and fungal genera (Myceliophthora and Acremonium) of the Ascomycota phyla (Figure 3). The Soil 1yr treatment had a higher abundance of bacterial Sphingomonas (Proteobacteria phyla), and two fungal genera (Mycothermus and Pseudallescheria) of the Ascomycota phyla. Two dominant bacterial genera (JG30-KF-CM45 and JG30-KF-CM66) belonging to the Chloroflexi phyla, and fungal genera (Remersonia and Kernia) (Ascomycota phyla) were higher abundance under the Soil 3yr treatment. The Soil 7yr treatment had higher abundances of bacterial genera (Paenisporosarcina (Firmicutes phyla), Bacillus (Firmicutes phyla), Gaiella (Actinobacteria phyla), and Dongia (Proteobacteria phyla)) (Figure 3a), and fungal genera (Aphanoascus, Stachybotrys, Acremonium, Pseudallescheria, Aspergillus, and Mortierella) of the Ascomycota phyla (Figure 3b).
3.5. Biomarker taxa analyses
Random Forests Based Importance Analysis of the top-10 most abundant phyla (Figure 4), revealed that bacterial Deinococcus-Thermus and Bacteroidetes phyla, and fungal Basidiomycota phyla were identified as the biomarker taxa of the Sub CK treatment with obviously higher relative abundances in the samples. Bacterial Dependentiae and Patescibacteria phyla, and fungal Ascomycota and Rozellomycota phyla were biomarker taxa of the Sub 1yr treatment. Bacterial Patescibacteria, Dependentiae and Nitrospirae phyla, and fungal Mortierellomycota, Rozellomycota, and Mucoromycota phyla were biomarker taxa of the Sub 3yr treatment. The biomarker taxa of the Sub 7yr treatment were the bacterial Halanaerobiaeota phyla and fungal Chytridiomycota and Olpidiomycota phyla. Moreover, for the surrounding soil, biomarker taxa of the bacterial Verrucomicrobia and Armatimonadetes phyla and fungal Ascomycota phylum were more highly abundant under the Soil CK treatment. Bacterial Firmicutes, Dependentiae, Nitrospirae, Rokubacteria, and Armatimonadetes phyla, and fungal Chytridiomycota and Olpidiomycota phyla were biomarker taxa of the Soil 1yr treatment. Bacterial Rokubacteria, Nitrospirae, and Dependentiae phyla, and fungal Aphelidiomycota and Calcarisporiellomycota phyla were biomarker taxa of the Soil 3yr treatment. Bacterial Firmicutes phyla and fungal Blastocladiomycota and Mortierellomycota phyla were biomarker taxa in the Soil 7yr treatment.
4. Discussion
Spent mushroom compost (SMC) substrates, which comprise a recycled and reutilized waste products in mushroom cultivation, have been recognized as an organic material source and soil amendment for greenhouse vegetable farming [11]. In this experiment, we found that SMC substrates had more abundant nutrients than continuous mono-cropping soil (soil CK) and the soil surrounding the substrate (Table 1). Particularly, in the greenhouse the total N and total P contents were significantly higher in the SMC substrate than in the surrounding soil [57,58]. These results reinforced that the application of SMC substrates can provide a suitable growth medium for horticultural crops [59], and can enhance the crop yields in greenhouse farming [11]. Meanwhile, the physicochemical soil properties of the SMC substrate were significantly decreased along with the duration of cultivation (Table 1). It was proved that nutrient reduction and the depletion of the SMC substrate was accompanied by plant growth with longer cultivation duration [60]. Nevertheless, the physicochemical soil properties were significantly higher in the SMC substrate than in the surrounding soil (Table 1). Lou et al. [61] proved that the SMC substrate applied to agricultural land enhanced the soil organic matter and nutrient contents, while it reduced the total N leaching. These results demonstrated that the SMC substrate was an efficient alternative growth medium for the cultivation of horticultural crops compared to typical greenhouse soil.
Enzyme activities are an indicator of soil quality and participate in nutrient cycling during plant growth [62], which can be utilized to assess plant growth medium in ecosystems in response to available nutrients and metabolic requirements [63]. Soil alkaline phosphatases [64], soil protease [65], and β-Glucosidase [66] are important enzymes for available P, N, and C cycling processes. The concentrations of these enzymes differ between substrates and surrounding soils under various cultivation durations (Table 2), which is mainly due to differences in physicochemical soil properties, such as soil organic matter, total N, and pH, and soil enzyme activities involving Glu, ALP, and Pro enzymes (Table 1, Figure 5). Sinsabaugh [67] reported that extracellular enzymes are often associated with the acquisition, transformation, and mineralization of C, P, and N in their growth medium. Among the older cultivated plants, the activities of C, N, and P cycling enzymes were reduced in the substrate medium (Table 2). It was elucidated that crop growth and harvesting can lead to decreased enzyme activities due to the reduction/shortage of nutrients year by year in the growth medium [68]. The β-Glucosidase activity was enhanced with longer cultivation duration, whereas the activities of protease and alkaline phosphatases reached their peak values under the Soil 1yr and Soil 3yr treatments compared to the continuous mono-cropping soil (Soil CK treatment), respectively (Table 2). These results may help to explain how enzyme activities were influenced by crop root systems via the alteration of root system architectures [69] and rhizosphere exudations [70] in the growth medium, as well as the exchange of nutrients between the SMC substrate and surrounding soil. Furthermore, enzyme activities primarily originated from root secretion and microorganisms [71], and were closely correlated with microbial communities and structures in the growth media ecosystem [72].
Spent mushroom compost (SMC) substrates, consisting of spent mushroom compost, manure residues, and other agricultural waste, can partially or completely substitute growth media for horticultural crop production in greenhouses [73]. Meng et al. [74] reported that SMC substrates were a better alternative than peat-based growth media for greenhouse tomato and pepper seedlings due to higher morphological growth and lower instances of Fusarium (of Ascomycota phylum) pathogen infections. It is also known that SMC substrates can be employed as an alternative source of organic matter for crop growth, which contribute to increased soil microbiological activities [75]. In this study, following multiple years of tomato cultivation, the SMC substrates still had higher microbial alpha-diversity than continuous monocropping soil (Soil CK) and non-planted SMC substrates (Sub CK) treatment (Figure 1). This result confirmed that crop root systems can augment microbial diversity through the formation of beneficial symbionts [76,77]. Undergoing several years of tomato cultivation, the compositions of bacterial and fungal communities showed similar trend in the growth medium (substrate and surrounding soil) (Figure 2). It was revealed that continuous cropping practices altered the microbial community structures and compositions in the rhizospheric soil [23,78], as well as in the spent mushroom substrate [79]. The continuous cropping system is a common practice in greenhouse vegetable farming, which has adverse effects on horticultural crop yields and quality due to pathogenic diseases [80].
Continuous cropping obstacles (CCO) induce declines in crop quality and yields and the exacerbation of diseases and pests [81], which may be correlated with the modification of soil enzyme activities and microorganism communities [82]. A reduction in beneficial microbes (i.e Bacillus and Trichoderma) and accumulation of fungal pathogens Fusarium occurred under various continuous cropping practices in greenhouse soil [83]. Fusarium wilt (FW) in strawberry was mainly caused by Fusarium oxysporum fungal pathogens in greenhouses [84]. In this study, the continuous cropping soil of tomatoes in greenhouses (Soil CK) exhibited a higher abundance of pathogenic bacteria of the Cyanobacteria phyla and fungi of the Ascomycota phyla(Table 3, Figure S3). Denikina et al. [85] reported that Cyanobacteria taxa were associated with the diseased endemic sponge Lubomirskia baicalensis in Lake Baikal. Challacombe et al. [86] revealed that Ascomycota fungi can exist as latent saprotrophs or pathogens within plant tissues based on genome and secretome analyses in arid ecosystems. In contrast, the SMC substrates had a higher abundance of bacteria (Deinococcus-Thermus, Patescibacteria, Dependentiae, Halanaerobiaeota, and Nitrospirae) (Table 3, Figure 4a) and fungi (Basidiomycota, Rozellomycota, Mortierellomycota, Mucoromycota, Chytridiomycota, and Olpidiomycota) (Table 3, Figure 4b). Deinococcus-Thermus bacteria are highly resistant against extreme environmental stress [87,88]. Tian et al. [89] demonstrated that the bacteria super-phyla Patescibacteria contains ultra-small cells, simple membrane structures and streamlined redundant and nonessential functions to avoid phage predation, and adapt to specific stressed environments. It was reported that the Halanaerobiaeota phyla was dominant in extremely haloalkaline environments due to its salt tolerance and anaerobic attributes [90], while Nitrospirae phyla were less prevalent and participated in C and N cycling between different ecosystems [91,92]. Further, as an important ectomycorrhizal fungi, the Basidiomycota phyla can undergo symbiosis with host plants, which become their C sources and habitats [93]. Both Rozellomycota and Chytridiomycota fungi belong to the Zoosporic phyla with motile spores, which typically play critical ecological roles in the recycling of energy and matter in food webs [94]. As fast growing saprotrophic fungi, the Mortierellomycota phyla are potentially influenced by soil temperature [95] and have important biological functions for the protection of plants against pathogens [96]. Interestingly, Mucoromycota fungi may be utilized as a biorefinery that employs fungi for its highly versatile metabolic system, which can generate several valuable bioproducts including of pigments, polyphosphates, ethanol, organic acids, enzymes, as well as low- and high-value lipids [97,98]. Therefore, it was revealed in this study that the greenhouse SMC substrate model was more enriched with several beneficial microbes than the continuous cropping field model.
Compared with open-field cultivation, plastic-greenhouse cultivation is a popular agricultural production platform on a global scale [9]. With the advantages of a prolonged growing season with a stable hospitable environment, plastic-greenhouse cultivation is preferred in many cases for the production of high value vegetables and other crops [99,100]. However, long-term intensive plastic-greenhouse cultivation can easily give rise to continuous cropping obstacles (CCO), which lead to deleterious changes in the composition of soil microbial communities [9]. Interesting, this study suggested that the long-term intensive cropping of tomatoes with the SMC substrate growth medium (1-, 3-, and 7-years cultivation) in a plastic-greenhouse had more similar bacterial beta-diversity than the non-planted SMC substrates (Sub CK) (Figure 2). It was demonstrated that long-term greenhouse vegetable cultivation with the SMC substrate growth medium altered the structures of soil microbial communities, as reported by Liu et al. [101]. Furthermore, the soil fungi beta-diversity of surrounding soil between the different cultivation durations (1-, 3-, and 7-years cultivation) was relatively consistent, in contrast to continuous mono-cropping soil (Soil CK) (Figure 2b). The root systems [102] and root exudates [103] of vegetable crops play critical roles in the compositional restructuring of soil microbial communities in the rhizosphere and surrounding soil environments, particularly under continuous mono-cropping [104]. Unal [11] suggested that SMC substrates were a viable alternative growth media for tomato seedling production in plastic greenhouses. Thus, SMC substrate cultivation might be a feasible strategy for the improved large scale production of horticultural vegetables and crops in greenhouses worldwide [73].
Meanwhile, we found that the dominant taxa Dongia and Sphingomonas (Proteobacteria phyla) were highly abundant in the continuous mono-cropping soil (Soil CK) (Figure 3a). Pathogenic bacteria in the Sphingomonas genus commonly cause brown spot disease on yellow Spanish melon fruits [105], and rase Panax ginseng with rusty root disease that seriously affects its production [106]. Yet, several beneficial bacterial genera (i.e. Saccharimonadales, Nocardioides, Gaiella) were significantly dominant in the SMC substrate growth media for tomato cultivation in the greenhouse (Figure 3a). Saccharimonadales (Patescibacteria phylum) was influenced by soil sugar concentrations, the high abundance of which can enhance soil alkaline phosphatase activities [107]. Nocardioides (Actinobacteria phylum) was responsible for the degradation of vinyl chloride (VC), which is carcinogenic to humans [108]. Zhao et al. [109] reported that Nocardioides and Gaiella were important beneficial bacteria for the suppression of Fusarium wilt in a long-term tomato monoculture soil. Even the Bacillus genus of important beneficial bacteria exhibited a significantly higher abundance in the surrounding soil of the 7-year cultivation (Figure 3a) [83]. Fungi were enriched in the SMC substrate and surrounding soil environment, which included several potentially beneficial fungal taxa of Thermomyces that secrete glycoside hydrolase and proteases [110] and coprophilous fungi of Kernia genus [111]. Further, the biodegradation fungi of the Kernia and Mortierella genera [112], the core fungus of Remersonia genus involved in humification processes [113], bioremediation microbiomes of contaminated Aspergillus genus environments [114], and alkaliphilic fungus of Sodiomyces genus were present [115], (Figure 3b). In contrast, the Ascobolus [116] and Fusarium [117] taxa of potential pathogens were significantly decreased during long-term cultivation with the SMC growth medium under greenhouse conditions.
5. Conclusions
The application of agro-based spent mushroom compost (SMC) substrates using a groove model system in greenhouses is a novel approach for the cultivation of horticultural plants. It was concluded that as an alternative growth media, SMC substrates could enhance the concentrations of major nutrients, such as total N and total P. In conjunction with longer cultivation durations, the physicochemical soil properties and soil enzyme activities were significantly reduced. However, the microbial alpha-diversity was considerably higher in the growth media of the groove model regardless of cultivation duration, in contrast to the continuous monocropping soil (Soil CK) and non-planted SMC substrate (Sub CK) treatment. The composition of bacterial communities had more similarities than fungal communities in both the SMC growth substrate and surrounding soil environment along with longer duration cultivation. Compared with the greenhouse continuous monocropping soil (Soil CK), the SMC substrate recruited more beneficial microbes, such as bacteria (Deinococcus-Thermus, Halanaerobiaeota, and Nitrospirae phyla) and fungi (Basidiomycota, Mortierellomycota, and Chytridiomycota phyla). Several potential beneficial bacterial genera (Saccharimonadales, Gaiella, Bacillus) and fungal genera (Thermomyces, Kernia, and Mortierella) were enriched in both the SMC substrate and surrounding soil. In general, the application of agro-based SMC substrates is recommended as a suitable growth media in the groove model for horticulture plant production under greenhouse conditions in North-western China.
Supplementary Materials
Nucleotide sequence data[ The nucleotide data has been submitted and deposited to the SRA (Sequence Read Archive) at the National Center for Biotechnology Information (NCBI) under accession number SUB12292980 (biosample SAMN31710080) for 16S bacteria sequences and number SUB12293105 (biosample SAMN31710019) for ITS fungi sequences. The review links as below: https://dataview.ncbi.nlm.nih.gov/object/PRJNA901255?reviewer=9n9qumsq784i9klabfct48k0db.https://dataview.ncbi.nlm.nih.gov/object/PRJNA901255?reviewer=9n9qumsq784i9klabfct48k0db]: The nucleotide data has been submitted and deposited in the GenBank databases.
Author Contributions
C.G.H. and Y.J.N. Conceptualization; C.G.H., Y.J.C., X.L.H., M.K., M.M.T., Y.Y.F., X.L.L. and Q.L. Methodology, Data collection; C.G.H. and X.L.H. Data analysis, Writing-Original draft preparation; M.K. Supervision; Q.L. Software, Validation; Y.L.C. Writing-Reviewing and Editing.
Funding
This work was supported by Shanxi Province Scientific and Technological Achievements Transformation guidance special project of China [grant numbers: 201904D131038; 202104021301045].
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Smith, G. R.; Archer, R. Climate, population, food security: adapting and evolving in times of global change. International Journal of Sustainable Development and World Ecology 2020, 27, 419–423. [Google Scholar] [CrossRef]
- Abbade, E.B.; Dewes, H. Food insecurity worldwide derived from food supply patterns. Food Secur. 2014, 7, 109–120. [Google Scholar] [CrossRef]
- Schmitt, J.; Offermann, F.; Söder, M.; Frühauf, C.; Finger, R. Extreme weather events cause significant crop yield losses at the farm level in German agriculture. Food Policy 2022, 112, 102359. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, P.; Chang, Y.; Zhang, L.; Hao, Y.; Tang, S.; Xiong, X. The environmental performance of greenhouse versus open-field cherry production systems in China. Sustain. Prod. Consum. 2021, 28, 736–748. [Google Scholar] [CrossRef]
- Kumar, K.; Jha, M.K.; Tiwari, K.; Singh, A. Modeling and evaluation of greenhouse for floriculture in subtropics. Energy Build. 2010, 42, 1075–1083. [Google Scholar] [CrossRef]
- Iddio, E.; Wang, L.; Thomas, Y.; McMorrow, G.; Denzer, A. Energy efficient operation and modeling for greenhouses: A literature review. Renew. Sustain. Energy Rev. 2019, 117, 109480. [Google Scholar] [CrossRef]
- Yu, H.-Y.; Li, T.-X.; Zhang, X.-Z. Nutrient Budget and Soil Nutrient Status in Greenhouse System. Agric. Sci. China 2010, 9, 871–879. [Google Scholar] [CrossRef]
- Hong, E.-M.; Choi, J.-Y.; Nam, W.-H.; Kang, M.-S.; Jang, J.-R. Monitoring nutrient accumulation and leaching in plastic greenhouse cultivation. Agric. Water Manag. 2014, 146, 11–23. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.; Ren, X.; Chen, B.; Zhang, Y.; Shen, C.; Wang, F.; Wu, D. Long-Term Greenhouse Cucumber Production Alters Soil Bacterial Community Structure. J. Soil Sci. Plant Nutr. 2019, 20, 306–321. [Google Scholar] [CrossRef]
- Sun, H.; Wei, C.; Xu, W.; Yang, J.; Wang, X.; Qiu, Y. Characteristics of salt contents in soils under greenhouse conditions in China. Environ. Sci. Pollut. Res. 2018, 26, 3882–3892. [Google Scholar] [CrossRef]
- Polat, E.; Uzun, H. I.; Topcuoglu, B.; Oenal, K.; Onus, A. N.; Karaca, M. Effects of spent mushroom compost on quality and productivity of cucumber (Cucumis sativus L.) grown in greenhouses. African Journal of Biotechnology 2009, 8, 176–180. [Google Scholar]
- Unal, M. The utilization of spent mushroom compost applied at different rates in tomato (Lycopersicon esculentum Mill.) seedling production. Emirates Journal of Food and Agriculture 2015, 27, 692–697. [Google Scholar]
- Meng, L.; Liqiang, M.; Zhang, X.; Zhao, Y.; Chen, L.; Zhang, S. Influence of spent mushroom substrate and molasses amendment on nitrogen loss and humification in sewage sludge composting. Heliyon 2020, 6, e04988. [Google Scholar] [CrossRef]
- Najafi, B.; Ardabili, S.F.; Shamshirband, S.; Chau, K.-W. Spent mushroom compost (SMC) as a source for biogas production in Iran. Eng. Appl. Comput. Fluid Mech. 2019, 13, 967–982. [Google Scholar] [CrossRef]
- Leong, Y.K.; Ma, T.-W.; Chang, J.-S.; Yang, F.-C. Recent advances and future directions on the valorization of spent mushroom substrate (SMS): A review. Bioresour. Technol. 2021, 344, 126157. [Google Scholar] [CrossRef]
- Szulc, W.; Rutkowska, B.; Kusmirek, E.; Spychaj-Fabisiak, E.; Kowalczyk, A.; Debska, K. Yielding, Chemcial Composition and Nitrogen Use Efficiency Determined for White Cabbage (Brassica Oleracea L. Var. Capitata L.) Supplied Organo-mineral Fertilizers From Spent Mushroom Substrate. Journal of Elementology 2019, 24, 1063–1077. [Google Scholar]
- Marín-Benito, J.M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Impact of Spent Mushroom Substrates on the Fate of Pesticides in Soil, and Their Use for Preventing and/or Controlling Soil and Water Contamination: A Review. Toxics 2016, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.; Ferreira, I.C.F.R.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A Review on Antimicrobial Activity of Mushroom (Basidiomycetes) Extracts and Isolated Compounds. Planta Medica 2012, 78, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Ngan, N.M.; Riddech, N. Use of Spent Mushroom Substrate as an Inoculant Carrier and an Organic Fertilizer and Their Impacts on Roselle Growth (Hibiscus sabdariffa L.) and Soil Quality. Waste Biomass- Valorization 2020, 12, 3801–3811. [Google Scholar] [CrossRef]
- Ge, T.; Nie, S.; Wu, J.; Shen, J.; Xiao, H.; Tong, C.; Huang, D.; Hong, Y.; Iwasaki, K. Chemical properties, microbial biomass, and activity differ between soils of organic and conventional horticultural systems under greenhouse and open field management: a case study. J. Soils Sediments 2010, 11, 25–36. [Google Scholar] [CrossRef]
- Hao, Z.; Chen, B.; Li, X. Relationship between soil chemical properties and microbial metabolic patterns in intensive greenhouse tomato production systems. Arch. Agron. Soil Sci. 2019, 66, 1334–1343. [Google Scholar] [CrossRef]
- Tong, L.; Zhu, L.; Lv, Y.; Zhu, K.; Liu, X.; Zhao, R. Response of organic carbon fractions and microbial community composition of soil aggregates to long-term fertilizations in an intensive greenhouse system. J. Soils Sediments 2019, 20, 641–652. [Google Scholar] [CrossRef]
- Li, Y.; Chi, J.; Ao, J.; Gao, X.; Liu, X.; Sun, Y.; Zhu, W. Effects of Different Continuous Cropping Years on Bacterial Community and Diversity of Cucumber Rhizosphere Soil in Solar-Greenhouse. Curr. Microbiol. 2021, 78, 2380–2390. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Ahmad, T.; Gulfam, A.; Saleem, M. Growth and Flowering of Gerbera as Influenced by Various Horticultural Substrates. Pakistan Journal of Botany 2012, 44, 291–299. [Google Scholar]
- Gonani, Z.; Riahi, H.; Sharifi, K. IMPACT OF USING LEACHED SPENT MUSHROOM COMPOST AS A PARTIAL GROWING MEDIA FOR HORTICULTURAL PLANTS. J. Plant Nutr. 2011, 34, 337–344. [Google Scholar] [CrossRef]
- Thomas, G. Soil pH and soil acidity. Methods of Soil Analysis Part 3—Chemical Methods. 1996, 475–490.
- Cao, N.; Chen, X.; Cui, Z.; Zhang, F. Change in soil available phosphorus in relation to the phosphorus budget in China. Nutr. Cycl. Agroecosystems 2012, 94, 161–170. [Google Scholar] [CrossRef]
- Guo, W.; Chen, H.; Zhang, Q. Effects of biochar application on total nitrogen and alkali-hydrolyzable nitrogen content in the topsoil of the high-yield cropland in north China Plain. Ecology & Environmental Sciences 201. [Google Scholar]
- Ryan, J.; Estefan, G.; Rashid, A. Soil and plant analysis laboratory manual. ICARDA. 2007.
- Zhou, X.; Wu, H.; Li, G.; Chen, C. Short-term contributions of cover crop surface residue return to soil carbon and nitrogen contents in temperate Australia. Environ. Sci. Pollut. Res. 2016, 23, 23175–23183. [Google Scholar] [CrossRef]
- Li, W.-H.; Zhang, C.-B.; Jiang, H.-B.; Xin, G.-R.; Yang, Z.-Y. Changes in Soil Microbial Community Associated with Invasion of the Exotic Weed, Mikania micrantha H.B.K. Plant Soil 2006, 281, 309–324. [Google Scholar] [CrossRef]
- Castro-Jiménez, J.; Gonzalez, C. Immunoassay-based screening of polychlorinated biphenyls (PCB) in sediments: requirements for a new generation of test kits. J. Environ. Monit. 2011, 13, 894–900. [Google Scholar] [CrossRef]
- Davinic, M.; Fultz, L.M.; Acosta-Martinez, V.; Calderón, F.J.; Cox, S.B.; Dowd, S.E.; Allen, V.G.; Zak, J.C.; Moore-Kucera, J. Pyrosequencing and mid-infrared spectroscopy reveal distinct aggregate stratification of soil bacterial communities and organic matter composition. Soil Biol. Biochem. 2012, 46, 63–72. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J. R.; Dillon, M. R.; Bokulich, N. A.; Abnet, C. C.; Al-Ghalith, G. A.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2 (vol 37, pg 852, 2019). Nature Biotechnology 2019, 37, 1091. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet Journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic acids research 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Statist 1984, 11, 265–270. [Google Scholar]
- Shannon, C. E. A mathematical theory of communication. The Bell System Technical Journal 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Lozupone, C. A.; Hamady, M.; Kelley, S. T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Applied and environmental microbiology 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: a New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [PubMed]
- Brian, H.; McArdle, M.; Anderson, J. Fitting multivariate models to community data: a comment on distanceâbased redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Huson, D.H.; Mitra, S.; Ruscheweyh, H.-J.; Weber, N.; Schuster, S.C. Integrative analysis of environmental sequences using MEGAN4. Genome Res. 2011, 21, 1552–1560. [Google Scholar] [CrossRef]
- Asnicar, F.; Weingart, G.; Tickle, T.L.; Huttenhower, C.; Segata, N. Compact graphical representation of phylogenetic data and metadata with GraPhlAn. PeerJ 2015, 3, e1029. [Google Scholar] [CrossRef]
- Zaura, E.; Keijser, B.J.; Huse, S.M.; Crielaard, W. Defining the healthy "core microbiome" of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef]
- Zgadzaj, R.; Garrido-Oter, R.; Jensen, D. B.; Koprivova, A.; Schulze-Lefert, P.; Radutoiu, S. Root nodule symbiosis in Lotus japonicus drives the establishment of distinctive rhizosphere, root, and nodule bacterial communities. Proceedings of the National Academy of Sciences of the United States of America 2016, 113, E7996–E8005. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Li, J.; Yang, X.; Jiang, Y.; Xia, S.-T. Multinomial random forest. Pattern Recognit. 2021, 122, 108331. [Google Scholar] [CrossRef]
- Breiman. Random forests. Random forests. Mach Learn 2001, 45, 5–32. [Google Scholar]
- Hanafi, F.H.M.; Rezania, S.; Taib, S.M.; Din, M.F.M.; Yamauchi, M.; Sakamoto, M.; Hara, H.; Park, J.; Ebrahimi, S.S. Environmentally sustainable applications of agro-based spent mushroom substrate (SMS): an overview. J. Mater. Cycles Waste Manag. 2018, 20, 1383–1396. [Google Scholar] [CrossRef]
- Lou, Z.; Zhu, J.; Wang, Z.; Baig, S.A.; Fang, L.; Hu, B.; Xu, X. Release characteristics and control of nitrogen, phosphate, organic matter from spent mushroom compost amended soil in a column experiment. Process. Saf. Environ. Prot. 2015, 98, 417–423. [Google Scholar] [CrossRef]
- Gümüş, I.; Şeker, C. Effects of spent mushroom compost application on the physicochemical properties of a degraded soil. Solid Earth 2017, 8, 1153–1160. [Google Scholar] [CrossRef]
- Henry, A.; Chaves, N.F.; Kleinman, P.J.; Lynch, J.P. Will nutrient-efficient genotypes mine the soil? Effects of genetic differences in root architecture in common bean (Phaseolus vulgaris L.) on soil phosphorus depletion in a low-input agro-ecosystem in Central America. Field Crop. Res. 2010, 115, 67–78. [Google Scholar] [CrossRef]
- Lou, Z.; Sun, Y.; Bian, S.; Baig, S.A.; Hu, B.; Xu, X. Nutrient conservation during spent mushroom compost application using spent mushroom substrate derived biochar. Chemosphere 2017, 169, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A.; Lewu, F.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: a review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Nannipieri, P., Giagnoni, L., Landi, L., and Renella, G. (2011). Role of phosphatase enzymes in soil. In "Phosphorus in action", pp. 215-243. Springer.
- Vranova, V.; Rejsek, K.; Formanek, P. Proteolytic activity in soil: A review. Appl. Soil Ecol. 2013, 70, 23–32. [Google Scholar] [CrossRef]
- Utobo, E. B.; Tewari, L. Soil Enzymes as Bioindicators of Soil Ecosystme Status. Applied Ecology and Environmental Research 2015, 13, 147–169. [Google Scholar]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Luan, H.-A.; Gao, W.; Tang, J.-W.; Li, R.-N.; Li, M.-Y.; Zhang, H.-Z.; Chen, X.; Masiliunas, D.; Huang, S.-W. Aggregate-associated changes in nutrient properties, microbial community and functions in a greenhouse vegetable field based on an eight-year fertilization experiment of China. J. Integr. Agric. 2020, 19, 2530–2548. [Google Scholar] [CrossRef]
- Yahya, M.; Islam, E.U.; Rasul, M.; Farooq, I.; Mahreen, N.; Tawab, A.; Irfan, M.; Rajput, L.; Amin, I.; Yasmin, S. Differential Root Exudation and Architecture for Improved Growth of Wheat Mediated by Phosphate Solubilizing Bacteria. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Zhang, X.; Dippold, M.A.; Kuzyakov, Y.; Razavi, B.S. Spatial pattern of enzyme activities depends on root exudate composition. Soil Biol. Biochem. 2019, 133, 83–93. [Google Scholar] [CrossRef]
- Bilyera, N.; Zhang, X.; Duddek, P.; Fan, L.; Banfield, C.C.; Schlüter, S.; Carminati, A.; Kaestner, A.; Ahmed, M.A.; Kuzyakov, Y.; et al. Maize genotype-specific exudation strategies: An adaptive mechanism to increase microbial activity in the rhizosphere. Soil Biol. Biochem. 2021, 162, 108426. [Google Scholar] [CrossRef]
- Luo, S.-P.; He, B.-H.; Zeng, Q.-P.; Li, N.-J.; Yang, L. Effects of seasonal variation on soil microbial community structure and enzyme activity in a Masson pine forest in Southwest China. J. Mt. Sci. 2020, 17, 1398–1409. [Google Scholar] [CrossRef]
- Zied, D.C.; de Abreu, C.G.; Alves, L.d.S.; Prado, E.P.; Pardo-Gimenez, A.; de Melo, P.C.; Dias, E.S. Influence of the production environment on the cultivation of lettuce and arugula with spent mushroom substrate. J. Environ. Manag. 2021, 281, 111799. [Google Scholar] [CrossRef]
- Meng, X.; Dai, J.; Zhang, Y.; Wang, X.; Zhu, W.; Yuan, X.; Yuan, H.; Cui, Z. Composted biogas residue and spent mushroom substrate as a growth medium for tomato and pepper seedlings. J. Environ. Manag. 2018, 216, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, X. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar. Bioresour. Technol. 2014, 171, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yang, W.; Chen, Y.; Han, X.; Khan, S.; Gao, Z.; Yang, Z. Ridge-furrow and film-mulching sowing practices enhance enzyme activity and alter fungi communities. Agron. J. 2020, 112, 4775–4787. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Antunes, P.M.; Koch, A.; Khosla, K.; Klironomos, J.N.; Dunfield, K.E. The influence of tillage on the structure of rhizosphere and root-associated arbuscular mycorrhizal fungal communities. Pedobiologia 2011, 54, 235–241. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Li, S.; Zuo, Z.; Zhan, R.; He, R. Variations of rhizospheric soil microbial communities in response to continuous Andrographis paniculata cropping practices. Bot. Stud. 2020, 61, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-W.; Xu, M.; Cai, X.-Y.; Tian, F. Evaluation of soil microbial communities and enzyme activities in cucumber continuous cropping soil treated with spent mushroom (Flammulina velutipes) substrate. J. Soils Sediments 2021, 21, 2938–2951. [Google Scholar] [CrossRef]
- Sun, K.; Fu, L.; Song, Y.; Yuan, L.; Zhang, H.; Wen, D.; Yang, N.; Wang, X.; Yue, Y.; Li, X.; et al. Effects of continuous cucumber cropping on crop quality and soil fungal community. Environ. Monit. Assess. 2021, 193, 436. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, X.; Tian, X.; Yang, T.; Deng, R.; Huang, J. Effects of continuous cropping of Pinellia ternata (Thunb.) Breit. on soil physicochemical properties, enzyme activities, microbial communities and functional genes. Chem. Biol. Technol. Agric. 2021, 8, 43. [Google Scholar] [CrossRef]
- Gao, Z.; Hu, Y.; Han, M.; Xu, J.; Wang, X.; Liu, L.; Tang, Z.; Jiao, W.; Jin, R.; Liu, M.; Guan, Z.; Ma, Z. Effects of continuous cropping of sweet potatoes on the bacterial community structure in rhizospheric soil. BMC Microbiology 2021, 21, 102. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, X.; Huang, H.; Jing, J.; Zhao, H.; Wang, L.; Long, X.-E. Contrasting beneficial and pathogenic microbial communities across consecutive cropping fields of greenhouse strawberry. Appl. Microbiol. Biotechnol. 2018, 102, 5717–5729. [Google Scholar] [CrossRef]
- Dinler, H.; Benlioglu, S.; Benlioglu, K. Occurrence of Fusarium wilt caused by Fusarium oxysporum on strawberry transplants in Aydın Province in Turkey. Australas. Plant Dis. Notes 2016, 11, 10. [Google Scholar] [CrossRef]
- Denikina, N.N.; Dzyuba, E.V.; Bel’kova, N.L.; Khanaev, I.V.; Feranchuk, S.I.; Makarov, M.M.; Granin, N.G.; Belikov, S.I. The first case of disease of the sponge Lubomirskia baicalensis: Investigation of its microbiome. Biol. Bull. 2016, 43, 263–270. [Google Scholar] [CrossRef]
- Challacombe, J.F.; Hesse, C.N.; Bramer, L.M.; McCue, L.A.; Lipton, M.; Purvine, S.; Nicora, C.; Gallegos-Graves, L.V.; Porras-Alfaro, A.; Kuske, C.R. Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genom. 2019, 20, 976. [Google Scholar] [CrossRef] [PubMed]
- Mariano, C.; Mello, I.S.; Barros, B.M.; da Silva, G.F.; Terezo, A.J.; Soares, M.A. Mercury alters the rhizobacterial community in Brazilian wetlands and it can be bioremediated by the plant-bacteria association. Environ. Sci. Pollut. Res. 2020, 27, 13550–13564. [Google Scholar] [CrossRef] [PubMed]
- White, O.; Eisen, J. A.; Heidelberg, J. F.; Hickey, E. K.; Peterson, J. D.; Dodson, R. J. , et al. Genome Sequence of the Radioresistant Bacterium <i>Deinococcus radiodurans</i> R1. Genome Sequence of the Radioresistant Bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar]
- Tian, R.; Ning, D.; He, Z.; Zhang, P.; Spencer, S.J.; Gao, S.; Shi, W.; Wu, L.; Zhang, Y.; Yang, Y.; et al. Small and mighty: adaptation of superphylum Patescibacteria to groundwater environment drives their genome simplicity. Microbiome 2020, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, M.; Mu, T.; Miao, D.; Liu, J.; Xing, J. Composition and key-influencing factors of bacterial communities active in sulfur cycling of soda lake sediments. Arch. Microbiol. 2022, 204, 317. [Google Scholar] [CrossRef]
- Abujabhah, I.S.; Doyle, R.B.; Bound, S.A.; Bowman, J.P. Assessment of bacterial community composition, methanotrophic and nitrogen-cycling bacteria in three soils with different biochar application rates. J. Soils Sediments 2017, 18, 148–158. [Google Scholar] [CrossRef]
- Baker, B.J.; Sheik, C.S.; A Taylor, C.; Jain, S.; Bhasi, A.; Cavalcoli, J.D.; Dick, G.J. Community transcriptomic assembly reveals microbes that contribute to deep-sea carbon and nitrogen cycling. ISME J. 2013, 7, 1962–1973. [Google Scholar] [CrossRef]
- Luptáková, E.; Mihál, I. Dynamics of ectomycorrhizal mycobiota (Basidiomycota) communities on a former agricultural land (West Carpathians). Mycological Progress 2020, 19, 845–857. [Google Scholar] [CrossRef]
- Hurdeal, V.G.; Gentekaki, E.; Hyde, K.D.; Jeewon, R. Where are the basal fungi? Current status on diversity, ecology, evolution, and taxonomy. Biologia 2020, 76, 421–440. [Google Scholar] [CrossRef]
- Dahl, M.B.; Peršoh, D.; Jentsch, A.; Kreyling, J. Root-Associated Mycobiomes of Common Temperate Plants (Calluna vulgaris and Holcus lanatus) Are Strongly Affected by Winter Climate Conditions. Microb. Ecol. 2021, 82, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S. A.; Suzuki, N. Viruses of plant pathogenic fungi. Annual review of phytopathology 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Dzurendova, S.; Losada, C. B.; Dupuy-Galet, B. X.; Fjær, K.; Shapaval, V. Mucoromycota fungi as powerful cell factories for modern biorefinery. Applied Microbiology and Biotechnology 2022, 106, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Kosa, G.; Zimmermann, B.; Kohler, A.; Ekeberg, D.; Afseth, N.K.; Mounier, J.; Shapaval, V. High-throughput screening of Mucoromycota fungi for production of low- and high-value lipids. Biotechnol. Biofuels 2018, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Ghannem, A.; BEN Aissa, I.; Majdoub, R. Effects of regulated deficit irrigation applied at different growth stages of greenhouse grown tomato on substrate moisture, yield, fruit quality, and physiological traits. Environ. Sci. Pollut. Res. 2020, 28, 46553–46564. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, X.; Yang, Z.; Huang, Q.; Qiu, R. Characteristics of Plastic Greenhouse High-Temperature and High-Humidity Events and Their Impacts on Facility Tomatoes Growth. Front. Earth Sci. 2022, 10. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Ren, X.; Chen, B.; Shen, C.; Wang, F. Long-term greenhouse vegetable cultivation alters the community structures of soil ammonia oxidizers. J. Soils Sediments 2018, 19, 883–902. [Google Scholar] [CrossRef]
- Lareen, A.; Burton, F.; Schäfer, P. Plant root-microbe communication in shaping root microbiomes. Plant Molecular Biology 2016, 90, 575–587. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Zhao, Y.; Mao, X.; Zhang, M.; Yang, W.; Di, H.J.; Ma, L.; Liu, W.; Li, B. Response of soil microbial communities to continuously mono-cropped cucumber under greenhouse conditions in a calcareous soil of north China. J. Soils Sediments 2020, 20, 2446–2459. [Google Scholar] [CrossRef]
- Buonaurio, R.; Stravato, V.M.; Kosako, Y.; Fujiwara, N.; Naka, T.; Kobayashi, K.; Cappelli, C.; Yabuuchi, E. Sphingomonas melonis sp. nov., a novel pathogen that causes brown spots on yellow Spanish melon fruits. Int. J. Syst. Evol. Microbiol. 2002, 52, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Park, J.Y.; Balusamy, S.R.; Huo, Y.; Nong, L.K.; Le, H.T.; Yang, D.C.; Kim, D. Comprehensive Genome Analysis on the Novel Species Sphingomonas panacis DCY99T Reveals Insights into Iron Tolerance of Ginseng. Int. J. Mol. Sci. 2020, 21, 2019. [Google Scholar] [CrossRef]
- Wang, G.; Jin, Z.; Wang, X.; George, T.S.; Feng, G.; Zhang, L. Simulated root exudates stimulate the abundance of Saccharimonadales to improve the alkaline phosphatase activity in maize rhizosphere. Appl. Soil Ecol. 2021, 170, 104274. [Google Scholar] [CrossRef]
- Wilson, F.P.; Liu, X.; Mattes, T.E.; Cupples, A.M. Nocardioides, Sediminibacterium, Aquabacterium, Variovorax, and Pseudomonas linked to carbon uptake during aerobic vinyl chloride biodegradation. Environ. Sci. Pollut. Res. 2016, 23, 19062–19070. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, Y.; Dong, W.; Zhang, Y.; Zhang, G.; Sun, Z.; Yang, L. Vermicompost can suppress Fusarium oxysporum f. sp. lycopersici via generation of beneficial bacteria in a long-term tomato monoculture soil. Plant Soil 2019, 440, 491–505. [Google Scholar] [CrossRef]
- Shi, Z.; Gong, W.; Zhang, L.; Dai, L.; Chen, G.; Wang, L. Integrated Functional-Omics Analysis of Thermomyces lanuginosus Reveals its Potential for Simultaneous Production of Xylanase and Substituted Xylooligosaccharides. Appl. Biochem. Biotechnol. 2018, 187, 1515–1538. [Google Scholar] [CrossRef]
- Caretta, G.; Piontelli, E.; Savino, E.; Bulgheroni, A. Some coprophilous fungi from Kenya. Mycopathologia 1998, 142, 125–134. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhao, X.; Zhang, X.; Zhao, Q.; Wang, X.; Li, Y. Restructured fungal community diversity and biological interactions promote metolachlor biodegradation in soil microbial fuel cells. Chemosphere 2019, 221, 735–749. [Google Scholar] [CrossRef]
- Kong, Z.; Wang, M.; Shi, X.; Wang, X.; Zhang, X.; Chai, L.; Liu, D.; Shen, Q. The functions of potential intermediates and fungal communities involved in the humus formation of different materials at the thermophilic phase. Bioresour. Technol. 2022, 354, 127216. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Singh, M.; Joshi, D.; Singh, J.; Suyal, D.C.; Kumar, A.; Rajput, V.D.; et al. Beneficial microbiomes for bioremediation of diverse contaminated environments for environmental sustainability: present status and future challenges. Environ. Sci. Pollut. Res. 2021, 28, 24917–24939. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, M.V.; Ianutsevich, E.A.; Danilova, O.A.; Kamzolkina, O.V.; Tereshina, V.M. Lipids and soluble carbohydrates in the mycelium and ascomata of alkaliphilic fungus Sodiomyces alkalinus. Extremophiles 2019, 23, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Wu, Y.; Li, H.; Liu, Q.; Zhou, Z.; Chen, Q.; Zhang, N.; Xu, L. Soil Fungal Diversity and Community Composition in Response to Continuous Sweet Potato Cropping Practices. Phyton 2021, 90, 1247–1258. [Google Scholar] [CrossRef]
- Zafra, J.M.R.; García, M.d.C.; Marquina, J.T.; Llamas, D.P. Dispersal of Fusarium spp. by rainwater and pathogenicity on four plant species. Aerobiologia 2015, 32, 431–439. [Google Scholar] [CrossRef]
Figure 1.
Bacterial (a) and fungal (b) taxa α-diversity indices of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments. (* Significant difference at p ≤ 0.05 ** significant difference at p ≤ 0.01).
Figure 1.
Bacterial (a) and fungal (b) taxa α-diversity indices of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments. (* Significant difference at p ≤ 0.05 ** significant difference at p ≤ 0.01).
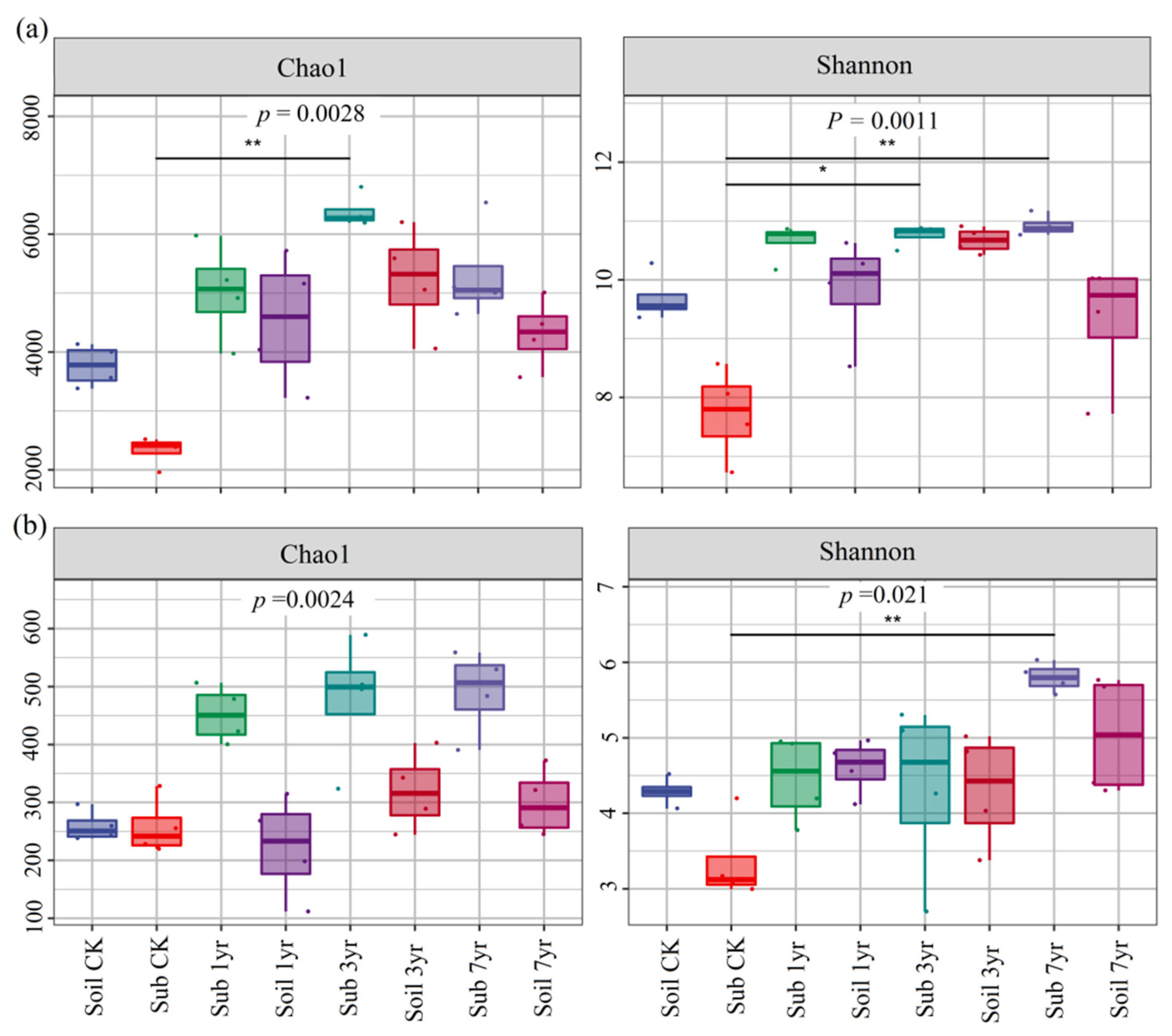
Figure 2.
Bacterial (a) and fungal (b) Beta-diversity of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
Figure 2.
Bacterial (a) and fungal (b) Beta-diversity of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
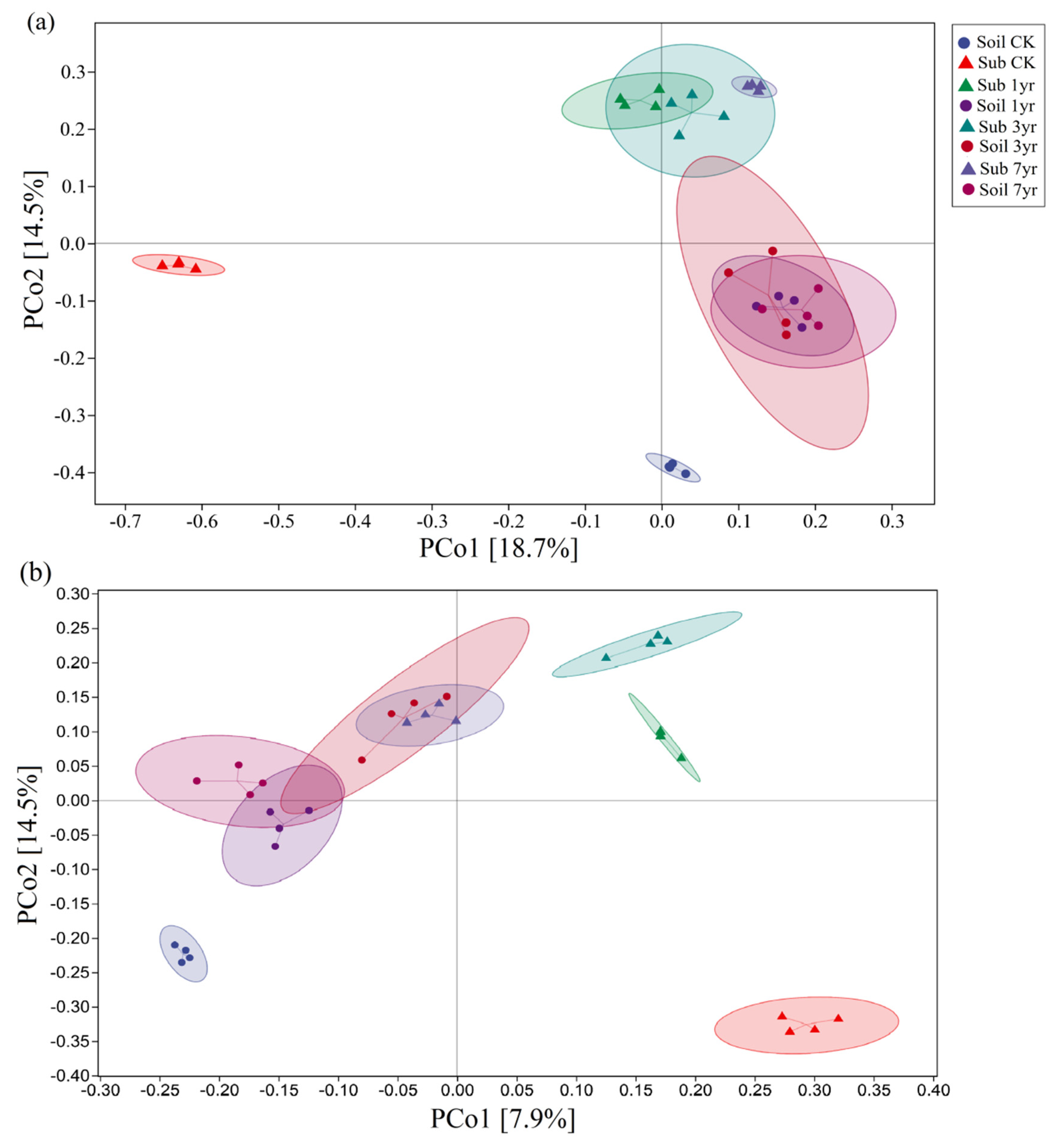
Figure 3.
Heat map and clustering analysis of bacterial (a) and fungal (b) community composition of the top-20 most abundant genera of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the treatments.
Figure 3.
Heat map and clustering analysis of bacterial (a) and fungal (b) community composition of the top-20 most abundant genera of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the treatments.
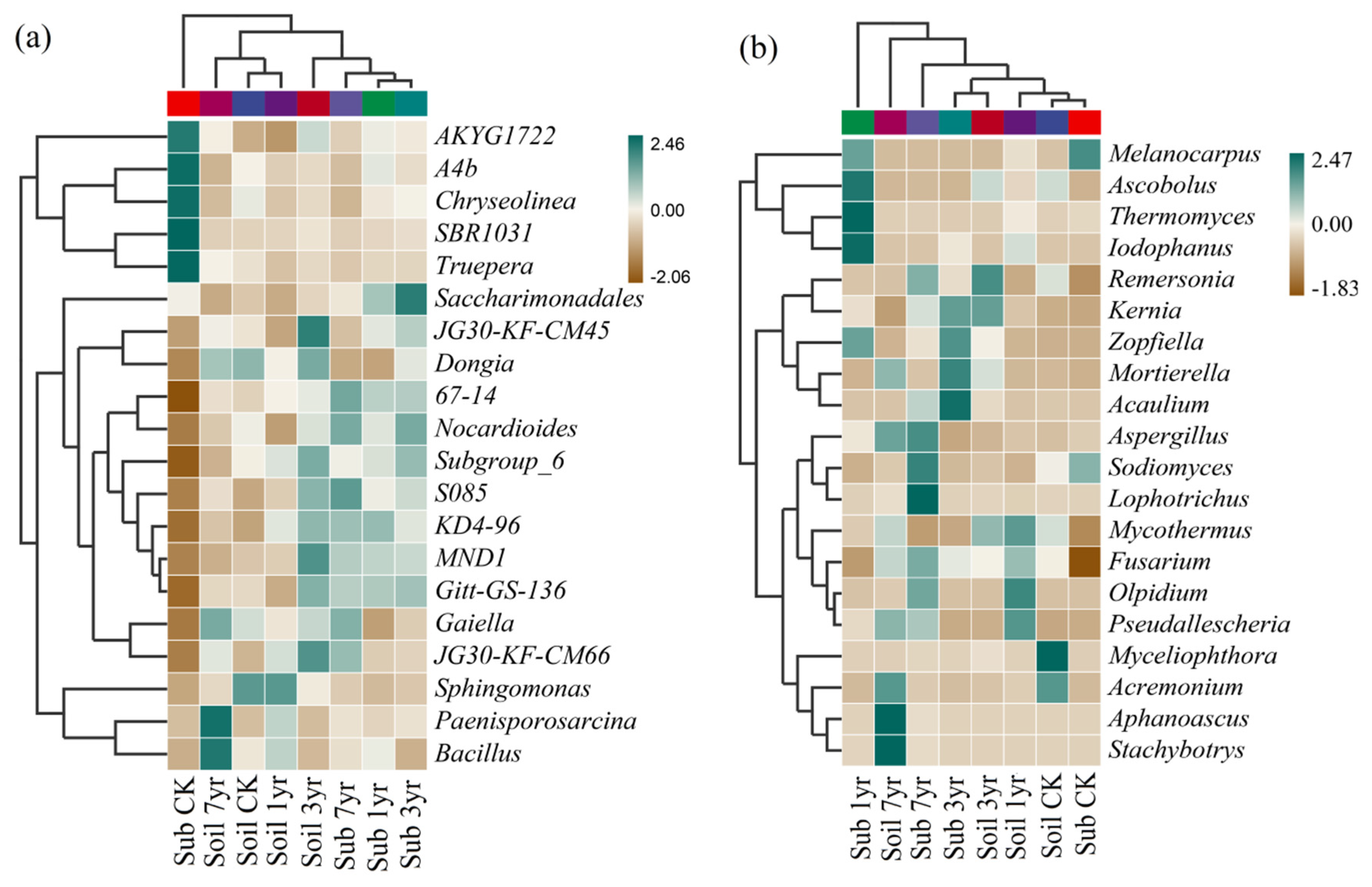
Figure 4.
Random forests-based importance analysis of bacterial (a) and fungal (b) community composition of the top-10 most abundant phyla of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
Figure 4.
Random forests-based importance analysis of bacterial (a) and fungal (b) community composition of the top-10 most abundant phyla of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
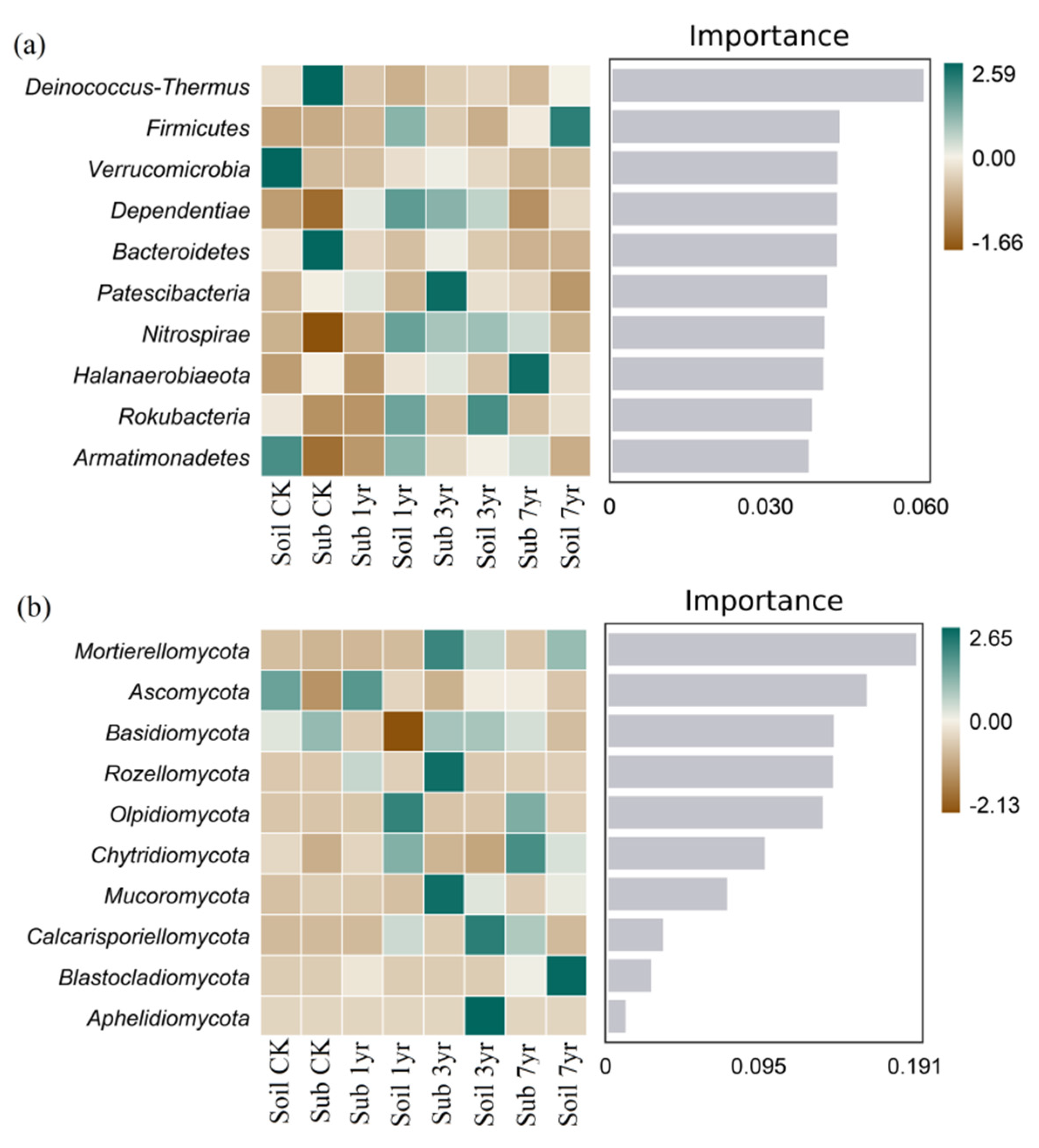
Figure 5.
Redundancy analysis (RDA) of soil properties and dominant microbial taxa of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the treatments. (Details: b1-b10 represent top 10 bacteria phyla, f1-f10 represents top 10 fungi phyla. b1, Deinococcus-Thermus; b2, Firmicutes; b3, Verrucomicrobia; b4, Dependentiae; b5, Bacteroidetes; b6, Patescibacteria; b7, Nitrospirae, b8, Halanaerobiaeota; b9, Rokubacteria; b10, Armatimonadetes; f1, Mortierellomycota; f2, Ascomycota; f3, Basidiomycota; f4, Rozellomycota; f5, Olpidiomycota; f6, Chytridiomycota; f7, Mucoromycota; f8, Calcarisporiellomycota; f9, Blastocladiomycota, f10, Aphelidiomycota.).
Figure 5.
Redundancy analysis (RDA) of soil properties and dominant microbial taxa of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the treatments. (Details: b1-b10 represent top 10 bacteria phyla, f1-f10 represents top 10 fungi phyla. b1, Deinococcus-Thermus; b2, Firmicutes; b3, Verrucomicrobia; b4, Dependentiae; b5, Bacteroidetes; b6, Patescibacteria; b7, Nitrospirae, b8, Halanaerobiaeota; b9, Rokubacteria; b10, Armatimonadetes; f1, Mortierellomycota; f2, Ascomycota; f3, Basidiomycota; f4, Rozellomycota; f5, Olpidiomycota; f6, Chytridiomycota; f7, Mucoromycota; f8, Calcarisporiellomycota; f9, Blastocladiomycota, f10, Aphelidiomycota.).
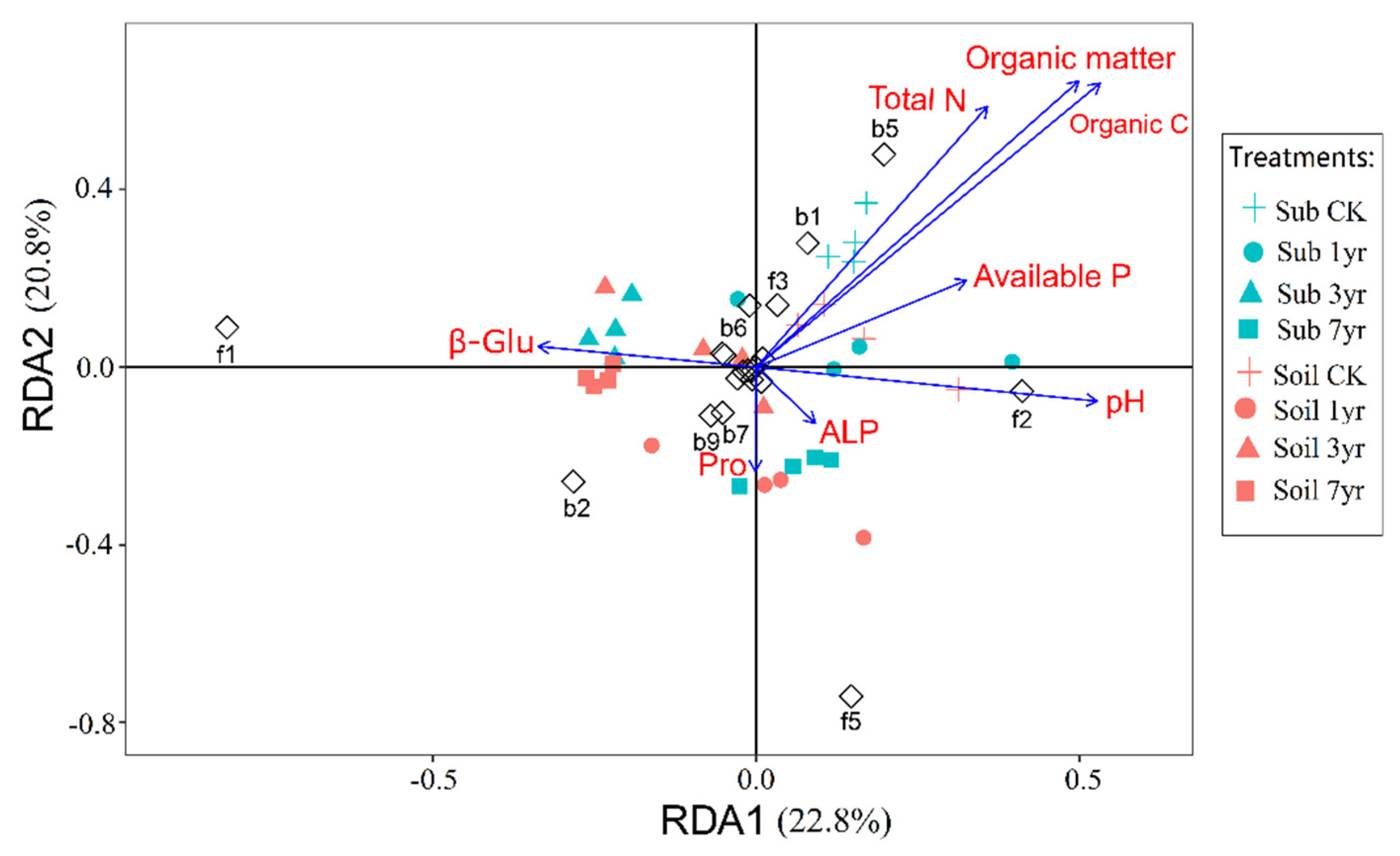

Table 1.
Physicochemical soil properties of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
Table 1.
Physicochemical soil properties of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
| Types | Treatments | Total P (g/kg) |
pHwater | Total N (g/kg) |
Organic C (g/kg) |
Organic matter (g/kg) |
|---|---|---|---|---|---|---|
| Substrates | Sub CK | 2.85±0.06 c | 7.81±0.07 a | 19.24±0.65 a | 38.87±7.83 a | 67.02±13.50 a |
| Sub 1yr | 4.07±0.02 a | 7.86±0.13 a | 16.92±0.88 a | 38.57±3.91 a | 66.50±6.75 a | |
| Sub 3yr | 3.11±0.04 bc | 7.35±0.05 b | 10.65±0.39 b | 28.60±4.80 b | 49.31±8.28 b | |
| Sub 7yr | 3.22±0.04 b | 7.27±0.05 b | 6.11±0.38 c | 16.70±1.43 c | 28.83±2.46 c | |
| Surrounding soil | Soil CK | 1.34±0.04 e | 7.45±0.06 b | 1.81±0.02 d | 33.97±0.46 ab | 58.57±0.80 ab |
| Soil 1yr | 1.32±0.01 e | 8.08±0.07 a | 1.29±0.07 d | 1.37±0.32 d | 2.37±0.55 d | |
| Soil 3yr | 1.74±0.08 d | 7.54±0.05 b | 2.27±0.09 d | 4.35±0.70 d | 7.50±1.21 d | |
| Soil 7yr | 0.74±0.04 f | 7.39±0.01 b | 1.73±0.06 d | 4.21±0.52 d | 7.25±0.90 d |
Data are means ± s.d. (n = 4), different letters means are significantly different at p ≤ 0.05 (Tukey HSD).
Table 2.
Enzyme activities of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
Table 2.
Enzyme activities of different cultivation duration of the substrate and the surrounding soil. See Table S1 for the abbreviations of the treatments.
| Types | Treatments | ALP (U/g) |
Pro (IU/g) |
β-Glu (U/g) |
|---|---|---|---|---|
| Substrates | Sub CK | 0.64±0.01 cd | 0.78±0.03 c | 0.37±0.01 d |
| Sub 1yr | 0.85±0.02 a | 1.61±0.07 a | 0.51±0.01 a | |
| Sub 3yr | 0.67±0.03 bcd | 1.44±0.06 b | 0.40±0.02 cd | |
| Sub 7yr | 0.71±0.05 bc | 0.77±0.05 c | 0.28±0.02 e | |
| Surrounding soil | Soil CK | 0.61±0.03 d | 0.42±0.02 d | 0.30±0.02 e |
| Soil 1yr | 0.74±0.03 b | 1.70±0.07 a | 0.45±0.01 b | |
| Soil 3yr | 0.92±0.02 a | 0.83±0.08 c | 0.44±0.01 bc | |
| Soil 7yr | 0.61±0.03 d | 0.49±0.03 d | 0.50±0.01 a |
Data are means ± s.d. (n = 4), different letters means are significantly different at p ≤ 0.05 (Tukey HSD).
Table 3.
Bacterial and fungal community composition of different cultivation duration of the substrate and the surrounding soil at the phyla level. See Table S1 for the abbreviations of the treatments.
Table 3.
Bacterial and fungal community composition of different cultivation duration of the substrate and the surrounding soil at the phyla level. See Table S1 for the abbreviations of the treatments.
| Taxon | Phyla | Substrate | Soil | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CK | 1yr | 3yr | 7yr | CK | 1yr | 3yr | 7yr | ||
| Bcteria | Acidobacteria | 0.008 c | 0.060 b | 0.070 ab | 0.069 ab | 0.056 bc | 0.090 a | 0.090 a | 0.042 bc |
| Actinobacteria | 0.166 c | 0.261 ab | 0.221 abc | 0.264 a | 0.187 abc | 0.155 c | 0.217 abc | 0.172 bc | |
| Bacteroidetes | 0.177 a | 0.045 b | 0.047 b | 0.020 cd | 0.046 b | 0.018 d | 0.026 c | 0.017 d | |
| Chloroflexi | 0.348 a | 0.149 bc | 0.132 cd | 0.118 de | 0.120 de | 0.102 ef | 0.193 ab | 0.092 f | |
| Cyanobacteria | 0.0002 e | 0.001 c | 0.0004 de | 0.003 b | 0.148 a | 0.003 b | 0.001 cd | 0.004 b | |
| Deinococcus-Thermus | 0.042 a | 0.004 bc | 0.004 bc | 0.002 c | 0.006 b | 0.0002 d | 0.004 bc | 0.009 b | |
| Firmicutes | 0.074 d | 0.107 bcd | 0.095 cd | 0.159 bc | 0.065 d | 0.200 ab | 0.059 d | 0.392 a | |
| Gemmatimonadetes | 0.010 c | 0.025 b | 0.030 b | 0.028 b | 0.070 a | 0.038 b | 0.063 a | 0.032 b | |
| Patescibacteria | 0.024 abc | 0.034 ab | 0.058 a | 0.019 bc | 0.013 cde | 0.008 de | 0.017 bcd | 0.006 e | |
| Proteobacteria | 0.140 c | 0.300 a | 0.319 a | 0.295 ab | 0.261 bc | 0.357 a | 0.296 ab | 0.215 c | |
| Others | 0.012 d | 0.015 d | 0.026 bc | 0.024 bc | 0.027 abc | 0.029 ab | 0.035 a | 0.018 cd | |
| Fungi | Ascomycota | 0.250 c | 0.727 a | 0.242 c | 0.465 abc | 0.593 ab | 0.349 bc | 0.367 bc | 0.327 bc |
| Mortierellomycota | 0.003 d | 0.006 d | 0.307 a | 0.027 bc | 0.012 cd | 0.012 cd | 0.0124 bc | 0.206 ab | |
| Mucoromycota | 0.0001 ab | 0.0001 ab | 0.001 a | 0.0001 ab | 0.000 b | 0.000 b | 0.0004 ab | 0.0004 ab | |
| Olpidiomycota | 0.000 c | 0.002 ab | 0.0001 bc | 0.108 a | 0.0003 c | 0.138 a | 0.0004 c | 0.007 bc | |
| Rozellomycota | 0.000 b | 0.002 a | 0.006 a | 0.0002 b | 0.000 b | 0.0002 b | 0.0001 b | 0.0003 b | |
| Others | 0.734 a | 0.256 b | 0.432 ab | 0.387 b | 0.386 b | 0.499 ab | 0.498 ab | 0.450 ab | |
Data are means ± s.d. (n = 4), different letters means are significantly different at p ≤ 0.05 in a line (Tukey HSD).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



