
Submitted:
18 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Salinity is the most significant abiotic stress affecting crop yield worldwide, breeders are Salinity is the most important abiotic stress limiting crop production throughout the world, and breeders are encouraged to test novel genotypes for their resistance to this selection agent. However, getting a sufficient number of plants for the evaluation can be a lengthy and time-consuming procedure that might be shortened by applying in vitro techniques. An experiment was conducted under controlled saline circumstances using a tissue culture technique to investigate the influence of salt on several physic-biochemical parameters. Micro-shoots of Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin rootstocks were re-cultured on MS media supplemented with NaCl from 25, up to 150 mM. Following the experiments, all of the tested rootstocks' morphological and biochemical characteristics, stomata behavior, and element accumulation were measured. As a result, adding NaCl from 25 to 150 mM to MS media caused a reduction in all morphogenetic parameters such as shoot number, shoot length, leaves number, and survival percentage was measured compared with control. Photosynthesis pigment, RWC, K, and Ca ions were decreased by increasing NaCl from 100 to 150 mM. On the contrary, proline, Cl, and Na were increased. Cleopatra mandarin and Trifoliate orange are more tolerant to salt stress than Volkamariana lemon and sour orange citrus.
Keywords:
Citrus rootstocks
; tissue culture
; biotechnology
; NaCl
; EDX
; Stomata
1. Introduction
Salinity has developed into a significant hazard to agriculture, reducing agricultural output all around the world. According to estimates, the global consequences of soil salinization could result in up to 50% less land by 2050 [1]. Crops are threatened by both ion toxicity and high salinity, which makes it more difficult for roots to absorb water and puts them under stress similar to that brought on by drought (osmotic impact). Salinity has an impact on photosynthesis, protein synthesis, energy metabolism, and lipid metabolism, which inhibits growth, causes wilting, or even causes mortality [2,3]. Some plants evolve defense mechanisms that either prevent salt from entering their cells or allow it to exist there. There are differences across species in how plants respond to the salt environment and how well their responses work [4]. Proline and glycine betaine, amino acids, inorganic ions, soluble sugars, and other compatible substances are present in high quantities in halophyte plants, which likely aid in osmotic adjustment, but compatible solute concentrations that build in glycophytes are not as high [5]. As a result, screening salt-tolerant rootstocks gives an alternative source to improve the breeding program and to better understand the pathways involved in the salt tolerance mechanism. Citrus is a popular name for a genus of flowering plants in the Rutaceae family. All citrus species have 2n = 18, and their karyotypic form and size are remarkably similar. Furthermore, the Rutaceae family is a huge, mostly tropical and subtropical family with 150–162 genera and 1500–2096 species [6,7]. At any time, the citrus industry’s major breeding aims include improving fruit quality and minimizing biotic and abiotic stresses [8]. Tissue culture has emerged as a relevant tool for stress evaluation in recent decades since it allows for the management of huge populations in a small space while allowing for more stringent environmental control [9]. It was demonstrated that tissue culture is a reliable tool to determine whether a genotype is tolerant to salinity or not since plants of the same genotype responded in a similar way to salinity in both experiments. Henceforth, in vitro evaluations can be employed to test genotypes at a very early stage and use very little time and space. Isozymes markers provide a convenient method for cultivar and rootstock identification. In the current study, we looked at four citrus rootstocks to examine some physiological and biochemical factors, as well as the biochemical genetic diversity in the leaves, to better understand the mechanisms underlying citrus rootstocks’ ability to withstand salinity.
2. Materials and Methods
This study was carried out in the tissue culture laboratory of the Horticulture Department, Faculty of Agriculture, Al-Azhar University, Nasr City, Cairo, Egypt, for three successive years 2019, 2020, and 2021.
2.1. Initiation and Stock
Fruits from four citrus rootstocks, including Cleopatra mandarin, trifoliate orange, Volkamariana lemon, and Sour orange, were sterilized by dipping them in tap water with soap for 30 minutes to remove dust. They were then dipped in 99.6% ethanol for 1/4 minute before being burned on the flame for 5 seconds under laminar flow conditions. Then, the fruits were placed on paper for sterilization. Finally, seeds were removed from the bulbs of fruits and, finally, seeds were prepared for culturing on MS media free of plant growth regulators.
2.1.1. Proliferation of Citrus Rootstocks
After germinating seeds of all tested rootstocks, soot tips or micro-cuttings were cultivated on MS media complemented with BA at 1.0 and NAA at 0.01 mg/L to develop explants of all tested rootstocks. The explants were taken from seed germination above, and the cultures were incubated for four weeks at 27 C using cool white fluorescent lamps at an intensity of 3000 Lux for 16 h daily). Each treatment was repeated three times.
2.1.2. Mass-Production of Citrus Rootstocks
Axillary shoots from various citrus rootstocks were transferred to MS basal medium supplemented with 30 g/L sucrose and both BA and KIN at 1.0 mg/L, and the cultures were incubated in a growth chamber at 27 °C using cool white fluorescent lamps with an intensity of 3000 Lux for 16 hours daily for four weeks. Each treatment was repeated three times to achieve mass production of all tested rootstocks.
2.2. Stress Treatments
The goal of this study was to see how several citrus rootstocks, such as Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin, responded in vitro to increasing salt stress in the culture media. Each treatment was performed three times with three plantlets in each duplicate, and uniformly developed explants from the multiplication medium were selected and transferred to MS media supplemented with sucrose, 30 g/l, BA at 1.0, NAA at 0.01, and with varying concentrations of sodium chloride (NaCl) (0.0, 25, 50, 75, 100, and 150 mM).
2.2.1. Data Collection and Morphological and Biochemical Characteristics
2.2.1.1. Determination of Morphological Characteristics
The growth of axillary shoots under salt stress was evaluated after thirty days by measuring the increase in shoot number, shoot length (cm), leaf number, shoot fresh weight, and survival percentage. The following equation by
Survival (%) = number of explants alive at end of period X100
2.2.1.2. Determination of Photosynthetic Pigments
According to [10], the chlorophyll a, b, total chlorophyll, and carotenoid contents of the leaves were measured at the end of the experiment period (after 30 days). Chlorophyll a, b, and total Chlorophyll and carotenoids were measured after thirty days using the method of abrading about 0.2 g of fresh tissue of plant leaf in a mortar with 15 mL of acetone 80% and, after filtering, its absorption was measured by spectrophotometer UV-Vis model 715 Jenway at 470, 663, and 646 mm. For calibrating the device, we used acetone at 80%. The concentrations of pigments were calculated using the following formula:
where Chl, a Chl, b, T, Chl, and car represent the concentrations of chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids. The measurements of photosynthetic pigment content were based on fresh weight.
Chl, a = (12.25 A 663.2 - 2.79 A 646.8)
Chl, b = (21.21 A 646.8 – 5.1 A 663.2)
2.2.1.3. Determination Proline Content
To assess proline content, [11] used a quick colorimetric approach. Warming 1.25 g of ninhydrin in 30 mT of glacial acetic acid and 20 mL of 6 M phosphoric acid with agitation until dissolved yielded acid-ninhydrin. The reagent is stable for 24 hours when kept cold (at 4 °C) (7 procedures). After being homogenized in 10 ml of 3 percent aqueous sulfosalicylic acid, the homogenate was filtered through Whatman # 2 filter paper. 2) In a test tube, two milliliters of the filtrate were mixed with two milliliters of acldninhdrin and two milliliters of glacial acetic acid for one hour at 100 °C before being stopped in an ice bath. 3) 4 mL of toluene was used to extract the reaction mixture, which was aggressively agitated for 15-20 seconds using a test tube stirrer. Toluene-containing chromophore was aspirated from the aqueous phase, warmed to room temperature, and absorbance was measured at 520 nm with toluene as a blank. 5) The proline concentration was measured on a fresh weight basis using a standard curve as follows:
{(ag proline /ml X ml toluene) /115.5ixg /Izmole}/{(g sample)/5}- = moles proline/g of fresh weight material
2.2.1.4. Leaf Relative Water Content
This work was carried out to study the effect of NaCl on the relative water content of four tested citrus rootstocks. Leaf relative water content (RWC) was calculated based on the [12] method. Two leaves were randomly chosen from the middle parts of the plants in each repetition. First, leaves were separated from the stems and their fresh masses (FM) were calculated. To measure the saturation mass (TM), they were placed into the distilled water in closed containers for 24 hours under the air condition of 22° C to reach their greatest amount of saturation mass, and then, they were weighed. Then the leaves were placed inside the electrical oven for 48 hours under the condition of 80° C, and the dry mass of the leaves (DM) was obtained (DM). All measurements were taken with scales with 0.001g accuracy and entered into the following formula:
where (FW) denotes fresh weight, (DW) denotes dry weight, and (TW) denotes turgid weight.
Relative water content (%)= FW-DW/TW-DWX100
2.2.1.5. Ions Accumulation
This inquiry was carried out to study the effect of MS media complemented with NaCl at different concentrations: 0.0, 25, 50, 75, 100, and 150 mM on ion accumulation in the leaves of four tested citrus rootstocks. EDX was used to estimate the accumulation content of some elements such as Na, Cl, K, Ca, P, and some microelements in the leaves of some citrus rootstocks. The samples were examined under X-ray microanalysis (Module Oxford 6587 INCAx-sight) attached to JEOL JSM-5500 LV scanning electron microscopy at 20KV at the Regional Center of Mycology and Biotechnology, Cairo, Egypt.
2.2.2. Stomata Behavior
This experiment was carried out to study the effect of MS media complemented with NaCl at 0.0, 25, 50, 75, 100, 125, and 150 mM on stomata characteristics such as the number of stomata, inside length of stomata µm, inside width of stomata µm, outside length of stomata µm and outside width of stomata µm of four tested grape rootstocks. The samples of the low surface of a leaf in some citrus rootstocks were coated by a gold sputter, coater (SPI-Module). For each imprint, the number of stomata/mm2 (average number of 3 fields) and stomata dimensions (10 stomata randomly selected per field) of four tested grape rootstocks were measured. Finally, the samples were examined by scanning electron microscopy (JEOL JSM-5500 LV) using high vacuum mode at the Regional Center of Mycology and Biotechnology, Cairo, Egypt. [13].
2.3. Statistical Procedure
Analysis of variance (ANOVA) was done using two ways of ANOVA Co-stat software, with three replicates for each treatment, in a randomized design. According to [14], Duncan’s test with a probability of 0.05 was used to show significant differences between treatments.
3. Results
3.1. Evaluation of Growth Characteristics
The effects of NaCl treatments on morphological traits measured in the four tested citrus rootstocks are depicted in Figure 1. It was clear that shoot number, shoot length (cm), leaf number, and survival percentage were decreased due to the increase in NaCl from 25 to 150 mM through direct treatment (shock treatment) compared with control. In contrast, increasing salt stress induced by NaCl from 25 up to 150 mM led to an increase in the death percentage of all tested rootstocks. A significant difference between the four tested citrus rootstocks was noticed regarding the effect of NaCl at different concentrations through shock treatment on previous morphological characteristics, where trifoliate orange rootstocks recorded the highest value compared with other citrus rootstocks.
3.2. Evaluation of Photosynthesis Pigment
The effects of salt stress generated by NaCl by direct (shock treatment) on chlorophyll content as chlorophyll “a” chlorophyll “b,” total chlorophyll, and carotenoids were shown in Figure 2. A severe reduction in photosynthesis pigment as chlorophyll “a”, chlorophyll “b”, and total chlorophyll carotenoids was achieved when shoots of all tested citrus rootstocks were cultured on a high level of NaCl from 100 up to 150 mM in comparison with the control and lowest concentrations of NaCl. All of the citrus rootstocks evaluated showed substantial variations. Trifoliate orange rootstock had the highest levels of chlorophyll and carotenoids, followed by Cleopatra mandarin, Volkamariana Limon, and sour orange citrus rootstocks.
3.3. Effect of NaCl on RWC
Figure 3 shows the impact of salt stress generated by NaCl by direct (shock treatment) using various doses of sodium chloride (0.0, 25, 50, 75, 100, 125, and 150 mM) on relative water content (RWC) in leaves of certain citrus rootstocks. Reduction in RWC was accomplished when shoots of four tested citrus rootstocks were cultured on MS media supplied with any level of NaCl, whether using different concentrations of sodium chloride by direct (shock treatment) at 25 up to 150 mM in comparison to control. When compared with other rootstocks, the highest reduction in RWC was obtained when shoots of Sour Orang and Volkamariana lemon were cultivated on a high level of NaCl at 150 Mm in comparison with other rootstocks. There were significant variations amongst all tested rootstocks when they were cultured on MS media complemented with 125 and 150 mM. The Cleopatra mandarin rootstock had the highest RWC values, followed by the Trifoliate orange rootstock compared to the other two citrus rootstocks.
3.4. Effect of NaCl on Proline Content
The results in Figure 4 show the effect of direct (shock treatment) salt stress caused by NaCl at different doses of 0.0, 25, 50, 75, 100, 125, and 150 mM on the proline content of four citrus rootstocks. When compared to the control, increasing salinity from 25 to 150 mM resulted in a significant increase in proline concentration in leaves for all the citrus rootstocks studied. There were substantial variations between all treatments of all citrus rootstocks evaluated, with MS medium enriched with 150 mM recording the highest accumulation proline concentration, followed by 125 mM, 100 mM, 75, 50, and both 25 and without sodium chloride. The accumulation of proline content differed significantly between all tested citrus rootstocks concerning salt stress, with the Cleopatra citrus rootstock recording the highest accumulation of proline content, followed in descending order by Trifoliate orange, Volkamariana lemon, and Sour orange citrus rootstocks.
3.5. Effect of NaCl on Ion Content
The establishment of ion homeostasis is required for plants to survive under salt-stress circumstances. The data given in Figure 5 demonstrated the influence of NaCl-induced salt stress on the accumulation amounts of various elements such as Na, Mg, Cl, K, Ca, P, P; Na/K ratio and some micro-elements in the leaves of all investigated citrus rootstocks. When shoots of tested citrus rootstocks were cultivated on MS medium enriched with NaCl at varying doses of 25, 50, 75, 100, 125, and 150 mM, the accumulation of K, Ca, Mg, P, and certain micro-elements was significantly reduced in contrast to control. When the micro-shoots of four examined citrus rootstocks were cultivated on high levels of NaCl 125 and 150 mM, the least amounts of K, Ca, Mg, P, and some micro-elements were accumulated in comparison to control and the lowest value of sodium chloride. In contrast, the accumulation contents of Na, and Na/K ratio increased with increasing NaCl from 25 to 150 mM in comparison to the control. As compared to the control and other treatments, the largest element accumulation of Na, Cl, and Na/K ratio was obtained when the micro-plantlets were cultivated on an MS medium supplemented with high levels of sodium chloride (125 and 150 mM). In this regard, the Cleopatra mandarin citrus rootstock recorded the highest accumulation of K in the leaf petiole and the lowest accumulation values of Na and Cl by increasing the NaCl level in the culturing medium, followed by Trifoliate orange, Volkamariana lemon, and Sour orange citrus rootstock.
3.6. Effect of NaCl on the Stomata Status
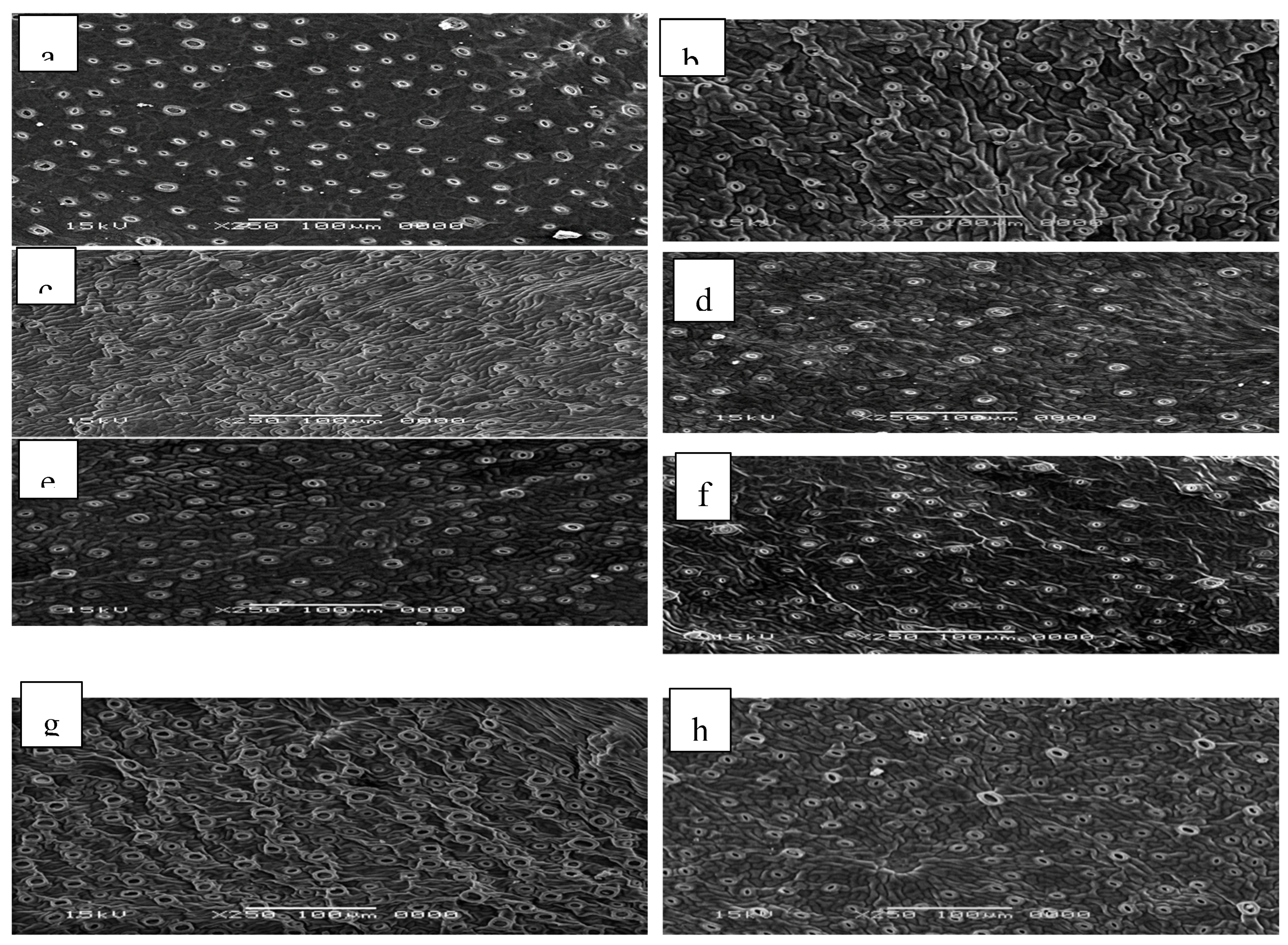
All stomata characteristics, including the number of open stomata, inner length of stomata (m), inside width of stomata (m), outside length of cell guard (m), and outside width of cell guard (m), were negatively influenced by adding NaCl from 25 to 150 mM to the growth medium (shock treatment technique) were data presented in Figure 6. All of the above-mentioned parameters of the four tested citrus rootstocks appeared to be strongly impacted by the citrus rootstocks, salt level, and their interaction, where all parameters of all tested citrus rootstocks were gradually lowered by raising the NaCl level in the growing medium. The magnitude of the decrease, however, differed from rootstock to rootstock. Re-cultivating the micro-shoots of all tested rootstocks on MS medium supplemented with 150 mM NaCl resulted in a reduction in all parameters of all tested citrus rootstocks, with Cleopatra mandarin rootstock exhibiting the highest percentage of reduction in the number of open stomata, inside length of stomata (m), outside length of cell guard (m), and outside width of cell guard (m), followed in descending order by Trifoliate orange and those of Volkamariana lemon and Sour orang.
Figure 6.
Effect of MS media supplemented with NaCl at different concentrations on leaf stomata behavior of some citrus rootstocks growing in vitro. a, b control and 150 mM NaCl respectively of volkamariana lemon rootstock; c, d (control and 150 mM NaCl respectively) of sour orange rootstock; e, f (control and 150 mM NaCl respectively) of trifoliate orange rootstocks and g, h (control and 150 mM NaCl respectively) of Cleopatra mandarin rootstocks.
Figure 6.
Effect of MS media supplemented with NaCl at different concentrations on leaf stomata behavior of some citrus rootstocks growing in vitro. a, b control and 150 mM NaCl respectively of volkamariana lemon rootstock; c, d (control and 150 mM NaCl respectively) of sour orange rootstock; e, f (control and 150 mM NaCl respectively) of trifoliate orange rootstocks and g, h (control and 150 mM NaCl respectively) of Cleopatra mandarin rootstocks.
3.7. Principal Components Analysis (PCA)
PCA was clarified first two components by 10 variables where the first and second components were (92.7-5.6%), (91.2 - 7.2%), (89.9-9.1%) and (89.4-8.1%) Cleopatra mandarin, Sour orange, Trifoliate orange, and Volkamariana lemon), respectively (Table 1) and (Figure 7). Table 8 provides a summary of the individual values, variance %, cumulative percentage, and component loading. The effect of MS media supplemented with different concentrations of NaCl is mostly located right side of the plot and has a strong positive correlation with the first component while the (Cleopatra mandarin) is recorded on the upper right side of the plot.
4. Discussion
Salt stress is a crucial element in plants that harms plant development and metabolism. Salinity stress is caused by a variety of complicated and varied processes involving diverse metabolic pathways in various organs. This allows us to exclude genotypes at a very early stage while consuming very little time and space. Shoot length, leaf number, and shoot number all dropped significantly as salt increased (Figure 1). The results are in full agreement with those previously reported by many workers, [15]. It has also been reported that MS media supplemented with NaCl from 0.0 up to 300 mM harmed plant growth (leaf number, plant length, fresh weight, and dry weight of two citrus rootstocks (sour orange and Volkamer lemon) compared with control under in vitro conditions. In this study, increasing NaCl level from 150 up to 300 mM in the growth medium caused a severe reduction in the above growth parameters compared with control and other treatments. A similar study [16] found that adding NaCl from 50 up to 200 mM caused a reduction in the shoot fresh weight of four tested citrus rootstocks compared with the control. Likewise, the negative effects of salinity on growth parameters can be related to ionic imbalance, changed availability and absorption of other ions, ion buildup in leaf cell vacuoles, a decrease in photosynthetic rate, and decreased carbon fixation [17]. Otherwise, suppression of branch development has been seen as a whole-plant response to salt stress [18]. Furthermore, these deleterious effects of salt stress might be attributable to a decrease in both cell division and cell growth [19]. Plant growth inhibition under salt stress is also predominantly due to the osmotic impact, whereas toxicity caused by excessive salt buildup in plant cells becomes apparent later in the growth cycle[20]. Plants grown in salty conditions are stunted due to a decrease in cell elongation and cell division, which are controlled by several auxins, the synthesis of which is slowed by salinity [21,22]. Plant growth is a key factor in assessing the salt tolerance of various citrus rootstocks. Under saline circumstances, the citrus rootstocks Cleopatra mandarin and Trifoliate orange maintained the highest growth parameters. The decrease in biomass increased with salt, owing to disruptions in physiological and metabolic activity under saline circumstances [23,24], which may be related to a reduction in leaf area and the number of leaves (25) ; (25).Although chlorophyll is membrane-bound and dependent on membrane stability, it seldom survives intact under saltwater conditions [25]. Salinity has also been linked to a decrease in chlorophyll concentration e.g., [26]. However, researchers summarized the findings by indicating that the decline in chlorophyll may be attributed to a difference in its production between plant species due to variance in particular enzymes under saltwater circumstances [27,28]. Furthermore, another intriguing element of the current study was the shift in chlorophyll a/b ratio, which demonstrated that the loss in chlorophyll b was more severe than the drop in chlorophyll a, which altered the chlorophyll a/b ratio. These criteria can be used to distinguish between salt-tolerant and salt-sensitive citrus rootstocks. In this regard, these results are in agreement with the findings of [16] which reported that photosynthetic pigments such as chlorophyll a, chlorophyll b, and carotenoid have had a negative correlation with salinity concentration and concentration of sodium ions in the tissue of the leaves of four citrus rootstocks, namely Sour orange (Citrus aurantium L.), Poncirus (Poncirus trifoliate Raf.), Citromelo (Citrumelo), and Citrange (Citranges) by adding NaCl 50, 100 and 200 mM to MS media in comparison with control. It was clear that Citrange and Sour orange rootstock show better resistance to the damages, caused by salt stress than other citrus rootstocks. Change in chlorophyll content due to salinity is the most obvious biochemical response [29]. The amount of chlorophyll decreases due to salinity stress because the glutamate which is the prefabricate matter of chlorophyll and proline is spent on the production of proline [30]. Furthermore, decreased chlorophyll concentration could be due to the inhibitory effect of ions accumulating in chloroplasts, chlorophyll breakdown caused by oxidative stress caused by salt, activation of chlorophyllase enzyme by salinity ions, and its negative effect on protophyzine. Through increased salt, increasing salinity reduces chlorophyll production. It produces an increase in leaf temperature, and as a result, the stomata shut down owing to water constraint stress induced by salinity as well as abscisic acid formation in the root and its transfer to the stomata [31]. Carotenoids have antioxidant characteristics and are vital in scavenging ROS as well as functioning as light-harvesting pigments [32]. Changes in the lipid-protein ratio of pigment-protein complexes or increased chlorophyllase activity could explain the decrease in chlorophyll content in abiotic stress plants [33]. Furthermore, the decrease in chlorophyll concentrations is most likely owing to the inhibitory impact of accumulated ions of various salts on the production of the various chlorophyll components [34].
At moderate and high salinities, RWC was significantly reduced in all rootstocks studied. This reduction was especially noticeable for the sour orange rootstock compared to other tested rootstocks. RWC is a direct reflection of plant water status, and its decrease shows that salt caused a water deficit in plants. The results go in line with [16] who found that MS media supplemented with NaCl at (0, 50, 100, and 200 mM) affected on RWC of four citrus rootstocks, namely Sour orange (Citrus aurantium L.), Poncirus (Poncirus trifoliate Raf.), Citromelo (Citrumelo), and Citrange (Citranges). It was clear that the RWC of all tested citrus rootstocks was decreased by increasing NaCl from 50 up to 200 mM compared with the control. The lowest RWC was obtained when the micro-shoots of all tested cultivars were cultured on MS media complemented with NaCl at 200 mM compared with those control and other treatments. Also, [35]. They reported that the leaf of Seedless Red and Ghezel Uzum grape cultivars possessed the minimum RWC due to the increase of NaCl up to 150mM in the culturing medium. When the relative water content is between 70 and 100 percent, simple decreases in turgescence potential and pore closure are reversible. However, in the case of the relative water content of 30 to 70% (because of the optical block), the plant chloroplast can be regained by rewatering. The chloroplast membrane is irreversibly damaged when the relative water content is less than 30% [36]. An increase in soluble salts, which slow the absorption of water and nutrients, creating osmotic effects and toxicity, caused a detrimental effect on plant water relations [37,38]. The leaf area, dry weight of the leaf, amount of chlorophyll, and other growth rate indicators are primarily connected with relative water content [39,40]. Maintaining a high RWC in leaves is one way of coping with salt stress and keeping plant cells turgid [41]. In addition, changes in the leaf water content are the key signal that causes changes in the plant’s hormonal balance. Thus, despite the saltwater environment of the root, ABA content in the leaves does not grow when the leaf-water content does not fall significantly below the ideal, as in the saline-high humidity condition [35]. So, It may be used as an enzyme protectant, a free radical scavenger, a cytosolic pH buffer stabilizer for subcellular structures, and a cell redox balancer [42].
Plants accumulate suitable solutes such as proline and soluble carbohydrates, which are known to have osmoprotective properties when they are stressed by salt [43]. One of the most prevalent reactions of plants to changes in the external osmotic potential is the accumulation of metabolites that function as compatible solutes [44]. Proline protects plants against salt stress in addition to acting as a suitable osmolyte. In the current study, moderate and high salinity increased the free proline concentration in the leaves and roots of all rootstocks. This rise was especially noticeable for Cleopatra mandarin rootstocks. The results are in agreement with [45] who cleared that, the increment of sodium chloride levels (up to 5000 ppm) in culture medium leads to a significant accumulation of proline in leaves of citrus rootstocks. Moreover, Balady sour orange is considered the most citrus rootstock tolerant to salinity stress since it exhibited the highest survival percentage and proline accumulation followed by Spanish sour orange while Brazilian, Russian sour orange, Alemow, and Trifoliate orange were found to be moderate to salt tolerance. Also, [46] found that MS media supplemented with NaCl at 0.0, 60, and 120 mM affected proline accumulation in leaves of five Persian pistachio genotypes including Akbari, Ahmad-Aghaee, Italyayi, Badami, and Ghazvini cultivars. The proline content of all tested cultivars was significantly increased with increasing salinity levels from 60 mM up to 120 mM compared with the control. Several roles have been proposed for proline in salinity tolerance. One possibility is that it acts as a store of energy that can be rapidly broken down and used when the plant is relieved of stress. Another is that it acts as an osmolyte and reduces the osmotic potential of the cell, thus reducing toxic ion uptake. In this case, the latter is more likely, with the salt-tolerant plants not only producing more proline when stressed but also having (in most cases) no significant drop in the chlorophyll content. This indicates that the increase in proline is reducing the physiologically detrimental effects of the salt [47]. The glutamate ligase enzyme is activated in salt and drought stress to convert glutamine to proline [40]. Osmoregulation by cellulose solute buildup, such as proline, has been postulated as a feasible strategy for combating salt stress situations [48]. Furthermore, proline has been postulated to act as a molecular chaperone, stabilizing protein structure, and proline accumulation can provide a mechanism to buffer cytosolic pH and regulate cell redox state [49]. In addition, [50] suggested that proline may have a role in mitigating the harm induced by dehydration.
The Cl content of the leaves of all examined rootstocks rose considerably when exposed to salt. Also, with increasing salt concentration, the K+ level in the leaves of all tested rootstocks decreased significantly. It is widely assumed that higher K+ accumulation in plants leads to increased tolerance to rising Na+ [51]. It is generally recognized that K+ can play a vital role in plant growth and development, as well as osmotic correction and cell turgor maintenance [52]. Furthermore, it is the primary cation in plants that balances the negative charge of anions and plays an important role in the activation of enzymes involved in protein and carbohydrate metabolism, as well as in the regulation of stomata movement [53]. The findings are consistent with those previously reported by many researchers, who discovered that increasing the NaCl level in the growth medium resulted in increased Na and Cl accumulation and decreased Ca concentrations in the plant tissue of two rootstocks (sour orange and Volkamer lemon). And also, [54] illustrated that re-culturing the micro-shoots of rough lemon (Citrus jambhiri Lush.) rootstock on MS media complemented with NaCl at 0.0, 17, 35, 51, 68, 86, 103, 120, 137, and 154 mM caused an increase in Cl and Na while the K+/Na+ ratio was decreased. When micro-shoots of tested citrus rootstock were grown on MS media supplemented with a high level of NaCl at 154 mM, the highest values of Cl- and Na+, as well as the lowest K+/Na+ ratio, were obtained when compared to the control and other treatments. In addition, [55] illustrated that shoot tips (1–1.5 cm long) from the proliferation of “Allow” (Citrus macrophylla) citrus rootstock media were placed in each standard proliferation medium supplemented with 0 and 80 mM of NaCl to induce salinity stress. It was clear that salt stress caused an increase in the accumulation of Na and Cl compared with the control. Tus, or plant salt tolerance, is a direct measure of the intracellular K+/Na+ ratio. Plants appear to try to modify their cytosolic K+/Na+ ratio by either reducing Na+ accumulation or compensating for K+ loss [56]. The effect of K+ on osmotic potential balance is dependent on many factors, including water content and cell wall elasticity [57]. Moreover, potassium is important for protein synthesis and stimulates photosynthesis [58], and interaction between K and Na might represent a key factor in determining the salinity tolerance of plants. The uptake of K+ is affected by Na+ due to the chemical similarities between both ions. K+ is an essential nutrient for most terrestrial plants. Therefore, K+ transport systems involving good selectivity of K+ over Na+ are considered important salt-tolerant determinants [59]. In the soil or reduction of ion transport to the xylem could be an influential factor in diminishing the accumulation of those ions in leaves [60]. There is also a possibility that growth reduction is not entirely due to Cl-toxicity but may be partially due to Cl-induced NO-3 deficiency [61]. The inhibition of NO-3 uptake by Cl-Cl could be due to interactions between both ions at the sites for ion transport [62]. Also, a higher K+/Na+ ratio maintains cell homeostasis in the presence of stress [63]. Salt tolerance has been mainly associated with the ability of different cultivars, rootstocks, or their combinations to restrict Cl entry into shoots [64].
Plant leaves exhibit increased energy dissipation and partial deactivation of PSII reaction centers in conditions of limited CO2 availability, such as when stomata are closed, reducing the energy input to the electron transport chain and preventing overexcitation and photodamage [65]. PI quantifies the state of plant performance under stress by measuring the functioning of both photosystems I and II [66]. In this work, salt stress was found to diminish plant PI, which was linked to a decrease in active reaction centers, electron trapping, and electron transfer to the electron transport chain during photosynthesis [67]. Furthermore, when salinity rises, the functional antenna size expands, placing the photosynthetic system under constant excitation pressure, resulting in energy loss and reduced oxygen generation. The results are in agreement with [68] reported that salt caused a reduction in the stomata density, width, and length of stomata of six jojoba clones in comparison to the control. The reduction in stomata conductance and mesophyll might contribute to the inhibition of photosynthesis under salt stress, so salt-sensitive Aegiceras corniculatum photosynthesis rates were barely affected by salinity. The depression of carbon assimilation was attributed to a reduction in stomata opening [69,70] also revealed that increasing NaCl from 50 mM up to 150 mM caused a reduction in all parameters of stomata, such as stomata aperture, stomata density, and length of stomata of Populus euphratica. Increasing NaCl from 150 to 200 mM led to a severe reduction in the previous parameters of the stomata of Populus euphratica compared with the control. Stomata closure plays a major role in photosynthetic rate reduction in response to osmotic stress [71]. Stomata conductivity reduces with decreasing relative water content (RWC) owing to salt stress, and the actual quantity of CO2 uptake drops, resulting in a reduction in photosynthesis [72]. Furthermore, in extreme temperatures, stomata closure appears to be the first reaction to avoid cell dehydration and plant survival [73]. Many researchers have identified stomata closure as the primary cause of decreased photosynthetic rate under osmotic stress conditions[74]. Closing stomata in response to salt stress is a regulatory mechanism that reduces water loss from tissues. If stomata close and transpiration slows over an extended period, carbon dioxide uptake by leaf tissues diminishes, resulting in a loss in photosynthesis and plant [75] Stomata closure reduces water loss, especially in plants experiencing a water shortage due to high solute concentrations in the nutritional medium, which reduces transpiration and either prevents or minimizes harmful ion buildup. However, if CO2 cannot reach the plant tissues, stomata might close, reducing photosynthesis.
5. Conclusions
In conclusion, increased salinity levels in the growth medium led to plant growth reduction (leaf number, plant length, and shoot number) and biochemical characteristics (chlorophyll content and RWC) in all tested rootstocks after one month in culture. Complete death was observed at the highest salinity levels (150 mM). Increasing NaCl levels in the growth medium resulted in increased Na and Cl accumulation and decreased K and Ca concentrations in plant tissue for all rootstocks. Cleopatra mandarin and Trifoliate orange are more tolerant to salt stress than Volkamariana lemon and sour orange citrus.
Author Contributions
“Conceptualization, G.A.A; A.M.A; H.F.A; I.A.E; A.N. A; A.A.S; A.E.H, and A.O.A; methodology, G.A.A; A.M.A; H.F.A; I.A.E; A.N. A; A.A.S; A.E.H, and A.O.A;; software, G.A.A; A.M.A; H.F.A; I.A.E; A.N. A; A.A.S; A.E.H, and A.O.A; validation, G.A.A; A.M.A; H.F.A; I.A.E; A.N. A; A.A.S; A.E.H, and A.O.A; formal analysis, (H.F.A); investigation, (A.O.A); resources, (H.F.A) and (A.E.H) ; data curation, (H.F.A) and (A.E.H) ; writing—original draft preparation, (H.F.A) and (A.E.H) writing—review and editing, (G.A.A)and (A.M.A); visualization, G.A.A; A.M.A; H.F.A; I.A.E; A.N.A; A.A.S; A.E.H, and A.O.A; supervision, (A.M.A) and (G.A.A); project administration, (A.M.A) and (G.A.A); funding acquisition, (A.M.A) All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by Researchers Supporting Project number (RSP2023R334), King Saud University, Riyadh, Saudi Arabia.
Data Availability Statement
Not applicable.
Acknowledgments
The authors extend their appreciation to the Researchers Supporting Project number (RSP2023R334), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Ahmad, K.; Akhtar, M.A.; Maqbool, M.A. Salinity Stress in Crop Plants: Effects of Stress, Tolerance Mechanisms and Breeding Strategies for Improvement. 2017, 02, 17. [Google Scholar]
- Hamouda, M.M.; Badr, A.; Ahmed, H.I.S.; Saad-Allah, K.M. Growth, Physiological, and Molecular Responses of Three Phaeophyte Extracts on Salt-Stressed Pea (Pisum sativum L.) Seedlings. In Review. 2022. [Google Scholar]
- Galvan-Ampudia, C.S.; Testerink, C. Salt Stress Signals Shape the Plant Root. Current Opinion in Plant Biology 2011, 14, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in Halophytes. Annals of Botany 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Kubitzki, K. (Ed.) Flowering Plants, Eudicots: Sapindales, Cucurbitales, Myrtaceae; The Families and Genera of Vascular, Plants; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 978-3-642-14396-0. [Google Scholar]
- Guerra, M.; Pedrosa, A.; Silva, A.E.B.e; Cornélio, M.T.M.; Santos, K.; Soares Filho, W.d.S. Chromosome Number and Secondary Constriction Variation in 51 Accessions of a Citrus Germplasm Bank. Braz. J. Genet. 1997, 20, 489–496. [Google Scholar] [CrossRef]
- Cai, X.; Fu, J.; Guo, W. Mitochondrial Genome of Callus Protoplast Has a Role in Mesophyll Protoplast Regeneration in Citrus : Evidence From Transgenic GFP Somatic Homo-Fusion. Horticultural Plant Journal 2017, 3, 177–182. [Google Scholar] [CrossRef]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Citrus Plants Exude Proline and Phytohormones under Abiotic Stress Conditions. Plant Cell Rep 2017, 36, 1971–1984. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Elsevier, 1987; Vol. 148, pp. 350–382. ISBN 978-0-12-182048-0. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Garcı́a-Mata, C.; Lamattina, L. Nitric Oxide Induces Stomatal Closure and Enhances the Adaptive Plant Responses against Drought Stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef]
- Chen, S.; Theodorou, I.G.; Goode, A.E.; Gow, A.; Schwander, S.; Zhang, J. (Jim); Chung, K.F.; Tetley, T.D.; Shaffer, M.S.; Ryan, M.P.; et al. High-Resolution Analytical Electron Microscopy Reveals Cell Culture Media-Induced Changes to the Chemistry of Silver Nanowires. Environ. Sci. Technol. 2013, 47, 13813–13821. [Google Scholar] [CrossRef]
- Stern, R.D. CoStat-Statutical Software. California: CoHort Software (1989), Pp. 302, $76.00. Ex. Agric. 1991, 27, 87. [Google Scholar] [CrossRef]
- Ghaleb, W.S.; Sawwan, J.S.; Akash, M.W.; Al-Abdallat, A.M. In Vitro Response of Two Citrus Rootstocks to Salt Stress. International Journal of Fruit Science 2010, 10, 40–53. [Google Scholar] [CrossRef]
- Rostamian, L.; Chalavi, V.; Sadeghi, H. Effect of salt stress on physiological responses of four citrus rootstock plantlets under in vitro condition. Journal of Crops Improvement. 2019, 21, 447–458. [Google Scholar] [CrossRef]
- Prior, L.; Grieve, A.; Cullis, B. Sodium Chloride and Soil Texture Interactions in Irrigated Field Grown Sultana Grapevines. II. Plant Mineral Content, Growth and Physiology. Aust. J. Agric. Res. 1992, 43, 1067. [Google Scholar] [CrossRef]
- Mulholland, B.J.; Taylor, I.B.; Jackson, A.C.; Thompson, A.J. Can ABA Mediate Responses of Salinity Stressed Tomato. Environmental and Experimental Botany 2003, 50, 17–28. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. IJMS 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Munns, R. Comparative Physiology of Salt and Water Stress: Comparative Physiology of Salt and Water Stress. Plant, Cell & Environment 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Loreto, F.; Centritto, M.; Chartzoulakis, K. Photosynthetic Limitations in Olive Cultivars with Different Sensitivity to Salt Stress: Photosynthetic Limitations and Salt Stress in Olive Cultivars. Plant, Cell & Environment 2003, 26, 595–601. [Google Scholar] [CrossRef]
- Ndayiragije, A.; Lutts, S. Do Exogenous Polyamines Have an Impact on the Response of a Salt-Sensitive Rice Cultivar to NaCl? Journal of Plant Physiology 2006, 163, 506–516. [Google Scholar] [CrossRef]
- Craine, J.M. Reconciling Plant Strategy Theories of Grime and Tilman: Reconciling Plant Strategy Theories. Journal of Ecology 2005, 93, 1041–1052. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to Increasing the Salt Tolerance of Wheat and Other Cereals. Journal of Experimental Botany 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Shah, S.H. EFFECTS OF SALT STRESS ON MUSTARD AS AFFECTED BY GIBBERELLIC ACID APPLICATION. 10.
- Gu, R.; Fonseca, S.; Puskas, L.G.; Hackler, L.; Zvara, A.; Dudits, D.; Pais, M.S. Transcript Identification and Profiling during Salt Stress and Recovery of Populus Euphratica. Tree Physiology 2004, 24, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Kreps, J.A.; Wu, Y.; Chang, H.-S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome Changes for Arabidopsis in Response to Salt, Osmotic, and Cold Stress. Plant Physiology 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Keutgen, A.J.; Pawelzik, E. Modifications of Strawberry Fruit Antioxidant Pools and Fruit Quality under NaCl Stress. J. Agric. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, I.E.; Jeffrey, O. Does Financial Risks Has Effects on The Performance of Deposit Money Banks in Nigeria? Saudi J. Bus. Manag. Stud 2021, 6, 71–85. [Google Scholar] [CrossRef]
- Molazem, D.; Qurbanov, E.M.; Dunyamaliyev, S.A. Role of Proline, Na and Chlorophyll Content in Salt Tolerance of Corn (Zea Mays L.). Environ. Sci. 2010, 6. [Google Scholar]
- Garcia, M.; Charbaji, T. Effect of Sodium Chloride Salinity on Cation Equilibria in Grapevine. Journal of Plant Nutrition 1993, 16, 2225–2237. [Google Scholar] [CrossRef]
- Pascale, S.D.; Maggio, A.; Fogliano, V.; Ambrosino, P.; Ritieni, A. Irrigation with Saline Water Improves Carotenoids Content and Antioxidant Activity of Tomato. The Journal of Horticultural Science and Biotechnology 2001, 76, 447–453. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Mittra, B. Effects of Salt on Growth, Ion Accumulation, Photosynthesis and Leaf Anatomy of the Mangrove, Bruguiera Parviflora. Trees - Structure and Function 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Chutipaijit, S.; Cha-um, S.; Sompornpailin, K. Modulation of Proline and Anthocyanin Levels Improves Salt Tolerant in Indica Rice Seedlings. Journal of Biotechnology 2008, 136, S152. [Google Scholar] [CrossRef]
- Hatami, E.; Esna-Ashari, M.; Javadi, T. Effect of Salinity on Some Growth Characteristics and Concentration of Elements in Two Grape (Vitis Vinifera L.) Cultivars, ‘Rishbaba’ and ‘Sahebi.’ 4.
- Damani, Z.; Karimi, H.R.; Mirik, A.A.M.; Esmaelizadeh, M. Effect of Salinity and Drought Stresses on Growth and Eco Physiological Parameters in Carob Seedlings (Ceratonia Siliqua L.). 1.
- Yang, F.; Xiao, X.; Zhang, S.; Korpelainen, H.; Li, C. Salt Stress Responses in Populus Cathayana Rehder. Plant Science 2009, 176, 669–677. [Google Scholar] [CrossRef]
- Jiang, X.; Qi, W.; Xu, X.; Li, Y.; Liao, Y.; Wang, B. Higher Soil Salinity Causes More Physiological Stress in Female of Populus Cathayana Cuttings. Acta Ecologica Sinica 2014, 34, 225–231. [Google Scholar] [CrossRef]
- Azimzadeh, Z.; Hassani, A.; Abdollahi Mandoulakani, B.; Sepehr, E. Effects of salinity stress on some morpho-physiological characteristics, essential oil content, and ion relations of two oregano subspecies (Origanum vulgare ssp. vulgare & ssp. gracile). Iranian Journal of Medicinal and Aromatic Plants Research 2021, 37, 658–674. [Google Scholar]
- Emami, S.; Asghari, A.; Mohammaddoust Chamanabad, H.; Rasoulzadeh, A.; Ramzi, E. Evaluation of osmotic stress tolerance in durum wheat (Triticum durum L.) advanced lines. Environmental Stresses in Crop Sciences 2019, 12, 697–707. [Google Scholar] [CrossRef]
- Walker, R.R.; Blackmore, D.H.; Clingeleffer, P.R.; Tarr, C.R. Rootstock Effects on Salt Tolerance of Irrigated Field-Grown Grapevines (Vitis Vinifera L. Cv. Sultana). 3. Fresh Fruit Composition and Dried Grape Quality. Aust J Grape Wine Res 2007, 13, 130–141. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biologia plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Azza, C.-C.; Mosbah, A.B.; Maalej, M.; Gargouri, K.; Gargouri-Bouzid, R.; Drira, N. In Vitro Salinity Tolerance of Two Pistachio Rootstocks: Pistacia Vera L. and P. Atlantica Desf. Environmental and Experimental Botany 2010, 69, 302–312. [Google Scholar] [CrossRef]
- Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- El-Habashy, S. In Vitro Evaluation and Selection for Salinity Tolerance in Some Citrus Rootstock Seedlings. 2018, 11. [Google Scholar]
- Gutierrez-Partida, E.; Hempel, H.; Caicedo-Dávila, S.; Raoufi, M.; Peña-Camargo, F.; Grischek, M.; Gunder, R.; Diekmann, J.; Caprioglio, P.; Brinkmann, K.O.; et al. Large-Grain Double Cation Perovskites with 18 Μs Lifetime and High Luminescence Yield for Efficient Inverted Perovskite Solar Cells. ACS Energy Lett. 2021, 6, 1045–1054. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Yamada, S.; Yamaguchi, T.; Rueda-Puente, E.; Ávila-Serrano, N.; García-Hernández, J.L.; López-Aguilar, R.; Troyo-Diéguez, E.; Nieto-Garibay, A. Influence of Calcium Silicate on Growth, Physiological Parameters and Mineral Nutrition in Two Legume Species Under Salt Stress. J Agron Crop Sci 2007, 193, 413–421. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Enhancement of Tolerance of Abiotic Stress by Metabolic Engineering of Betaines and Other Compatible Solutes. Current Opinion in Plant Biology 2002, 5, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Javaid, T.; Farooq, M.A.; Akhtar, J.; Saqib, Z.A.; Anwar-ul-Haq, M. Silicon Nutrition Improves Growth of Salt-Stressed Wheat by Modulating Flows and Partitioning of Na+, Cl− and Mineral Ions. Plant Physiology and Biochemistry 2019, 141, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Rasel, M.; Tahjib-Ul-Arif, M.; Hossain, M.A.; Hassan, L.; Farzana, S.; Brestic, M. Screening of Salt-Tolerant Rice Landraces by Seedling Stage Phenotyping and Dissecting Biochemical Determinants of Tolerance Mechanism. J Plant Growth Regul 2021, 40, 1853–1868. [Google Scholar] [CrossRef]
- Mahmood-ur-Rahman; Munazza, I.; Qamar, S.; Bukhari, S.A.; Malik, K. Abiotic Stress Signaling in Rice Crop. In Advances in Rice Research for Abiotic Stress Tolerance. Elsevier, 2019; pp. 551–569. ISBN 978-0-12-814332-2. [Google Scholar]
- Wenji, L.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochemical and Biophysical Research Communications 2018, 495, 286–291. [Google Scholar] [CrossRef]
- El-Moneim, A.; Eman, A.A.; Abd-Allah AS, E.; Ebeed, S.S. EFFECT OF SOME ORGANIC AND BIOFERTILIZER TREATMENTS ON MINIMIZING MINERAL NITROGEN FERTILIZATION OF WASHINGTON NAVEL ORANGE TREES. Arab Universities Journal of Agricultural Sciences 2008, 16, 451–457. [Google Scholar] [CrossRef]
- Sharma, L.K.; Kaushal, M.; Bali, S.K.; Choudhary, O.P. Evaluation of Rough Lemon (Citrus Jambhiri Lush.) as Rootstock for Salinity Tolerance at Seedling Stage under in Vitro Conditions. Afr. J. Biotechnol. 2013, 12, 6267–6275. [Google Scholar] [CrossRef]
- Pérez-Jiménez, M.; Pérez-Tornero, O. In Vitro Plant Evaluation Trial: Reliability Test of Salinity Assays in Citrus Plants. Plants 2020, 9, 1352. [Google Scholar] [CrossRef]
- Showkat Ahmad, G.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in Understanding Salt Tolerance in Rice. Theor Appl Genet 2019, 132, 851–870. [Google Scholar] [CrossRef]
- Davis, L.; Sumner, M.; Stasolla, C.; Renault, S. Salinity-Induced Changes in the Root Development of a Northern Woody Species, Cornus Sericea. Botany 2014, 92, 597–606. [Google Scholar] [CrossRef]
- Weise, A.; Barker, L.; Kühn, C.; Lalonde, S.; Buschmann, H.; Frommer, W.B.; Ward, J.M. A New Subfamily of Sucrose Transporters, SUT4, with Low Affinity/High Capacity Localized in Enucleate Sieve Elements of Plants. Plant Cell 2000, 12, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Fariba, A.; Ehsanpour, A.A. Soluble Proteins, Proline, Carbohydrates and Na+/K+ Changes in Two Tomato (Lycopersicon Esculentum Mill.) Cultivars under in Vitro Salt Stress. American J. of Biochemistry and Biotechnology 2005, 1, 212–216. [Google Scholar] [CrossRef]
- Tester, M. Na+ Tolerance and Na+ Transport in Higher Plants. Annals of Botany 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Ashraf, M.; Khanum, A. Relationship Between Ion Accumulation and Growth in Two Spring Wheat Lines Differing in Salt Tolerance at Different Growth Stages. J Agron Crop Sci 1997, 178, 39–51. [Google Scholar] [CrossRef]
- Cram, W.J. Chloride Accumulation as a Homeostatic System: Set Points and Perturbations: THE PHYSIOLOGICAL SIGNIFICANCE OF INFLUX ISOTHERMS, TEMPERATURE EFFECTS AND THE INFLUENCE OF PLANT GROWTH SUBSTANCES. J Exp Bot 1983, 34, 1484–1502. [Google Scholar] [CrossRef]
- Edward, P.G.; Nelson, S.G.; Ambrose, B.; Martinez, R.; Soliz, D.; Pabendinskas, V.; Hultine, K. Comparison of Salinity Tolerance of Three Atriplex Spp. in Well-Watered and Drying Soils. Environmental and Experimental Botany 2012, 83, 62–72. [Google Scholar] [CrossRef]
- Haffter, P.; Granato, M.; Brand, M.; Mullins, M.C.; Hammerschmidt, M.; Kane, D.A.; Odenthal, J.; van Eeden, F.J.; Jiang, Y.J.; Heisenberg, C.P.; et al. The Identification of Genes with Unique and Essential Functions in the Development of the Zebrafish, Danio Rerio. Development 1996, 123, 1–36. [Google Scholar] [CrossRef]
- Ceusters, N.; Valcke, R.; Frans, M.; Claes, J.E.; Van den Ende, W.; Ceusters, J. Performance Index and PSII Connectivity Under Drought and Contrasting Light Regimes in the CAM Orchid Phalaenopsis. Front. Plant Sci. 2019, 10, 1012. [Google Scholar] [CrossRef]
- Stirbet, A.; Riznichenko, G.Y.; Rubin, A.B.; Govindjee. Modeling Chlorophyll a Fluorescence Transient: Relation to Photosynthesis. Biochemistry Moscow 2014, 79, 291–323. [Google Scholar] [CrossRef]
- Salim Akhter, M.; Noreen, S.; Mahmood, S.; Athar, H.-R.; Ashraf, M.; Abdullah Alsahli, A.; Ahmad, P. Influence of Salinity Stress on PSII in Barley (Hordeum Vulgare L.) Genotypes, Probed by Chlorophyll-a Fluorescence. Journal of King Saud University - Science 2021, 33, 101239. [Google Scholar] [CrossRef]
- El-Baz, E.; EL-Dengawy, E.; El-Shahat, S.; El-Hassan, E. STUDIES ON SOME MORPHOLOGICAL ASPECTS OF JOJOBA [Simmondsia Chinensis (LINK) SCHNEIDER] UNDER EGYPTION CONDITIONS. Journal of Plant Production 2009, 34, 10575–10568. [Google Scholar] [CrossRef]
- Adhikari, T.; Kumar, A. Phytoaccumulation and Tolerance of Riccinus Communis L. to Nickel. International Journal of Phytoremediation 2012, 14, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Amit Kumar, G.; Kumar, M. Prediction and Analysis of Quorum Sensing Peptides Based on Sequence Features. PLoS ONE 2015, 10, e0120066. [Google Scholar] [CrossRef]
- Sharam, S.; Abbasnia Zare, S.K. Interactive Effects of Salinity and Drought Stresses on the Growth Parameters and Nitrogen Content of Three Hedge Shrubs. Cogent Environmental Science 2019, 5, 1682106. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Tezara, W. Causes of Decreased Photosynthetic Rate and Metabolic Capacity in Water-Deficient Leaf Cells: A Critical Evaluation of Mechanisms and Integration of Processes. Annals of Botany 2009, 103, 561–579. [Google Scholar] [CrossRef] [PubMed]
- Abbaspour, H. Influence of Salt Stress on Growth, Pigments, Soluble Sugars and Ion Accumulation in Three Pistachio Cultivars. J. Med. Plants Res. 2012, 6, 2468–2473. [Google Scholar] [CrossRef]
- Morales, C.G.; Pino, M.T.; del Pozo, A. Phenological and Physiological Responses to Drought Stress and Subsequent Rehydration Cycles in Two Raspberry Cultivars. Scientia Horticulturae 2013, 162, 234–241. [Google Scholar] [CrossRef]
- Tardieu, F.; Parent, B.; Caldeira, C.F.; Welcker, C. Genetic and Physiological Controls of Growth under Water Deficit. Plant Physiology 2014, 164, 1628–1635. [Google Scholar] [CrossRef]
Figure 1.
Effect of MS media supplemented with NaCl at different concentrations on morphological characteristics of some citrus rootstocks growing in vitro.
Figure 1.
Effect of MS media supplemented with NaCl at different concentrations on morphological characteristics of some citrus rootstocks growing in vitro.
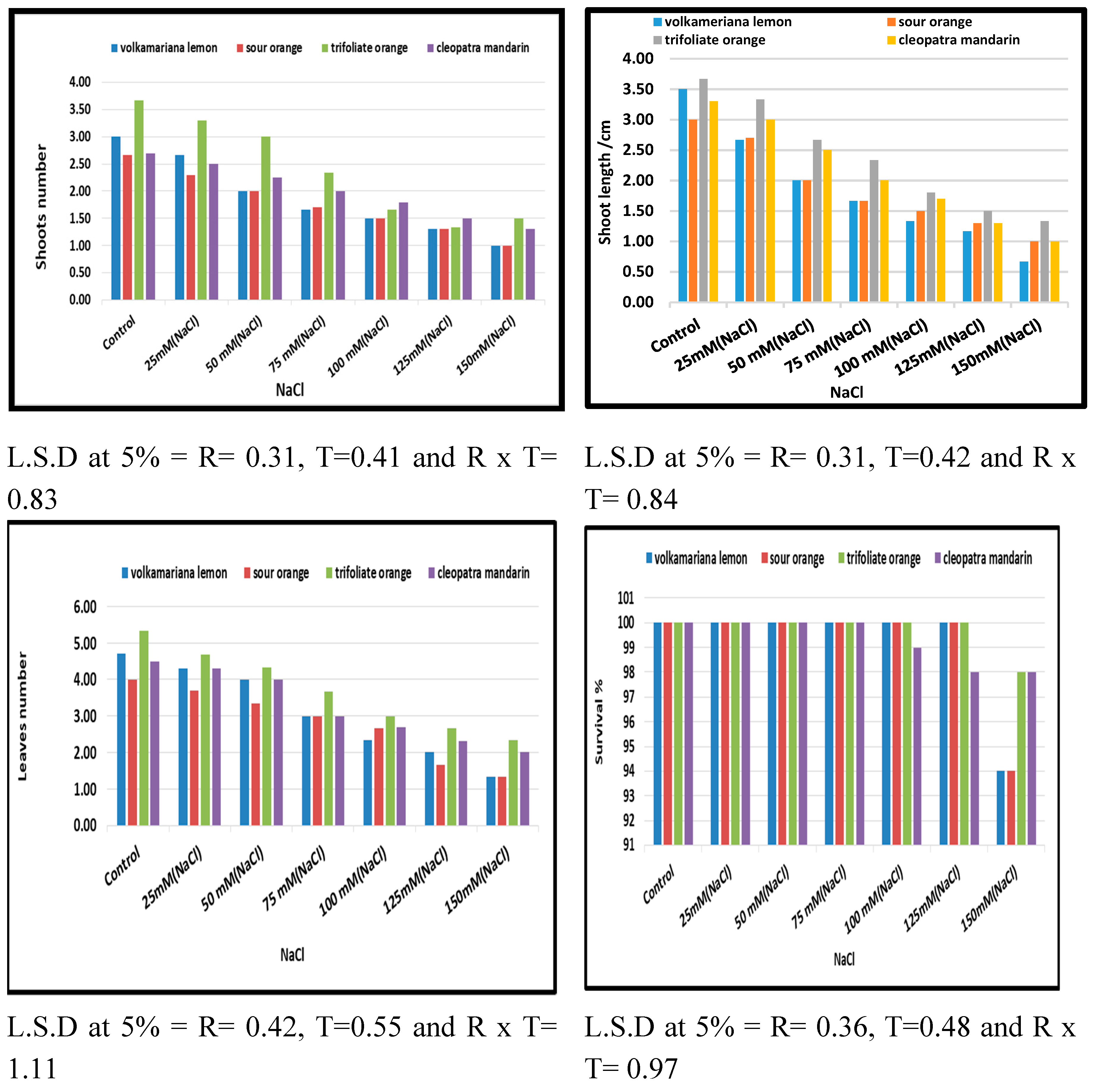
Figure 2.
Effect of MS media supplemented with NaCl at different concentrations on leaf biochemical characteristics of some citrus rootstocks grown in vitro.
Figure 2.
Effect of MS media supplemented with NaCl at different concentrations on leaf biochemical characteristics of some citrus rootstocks grown in vitro.
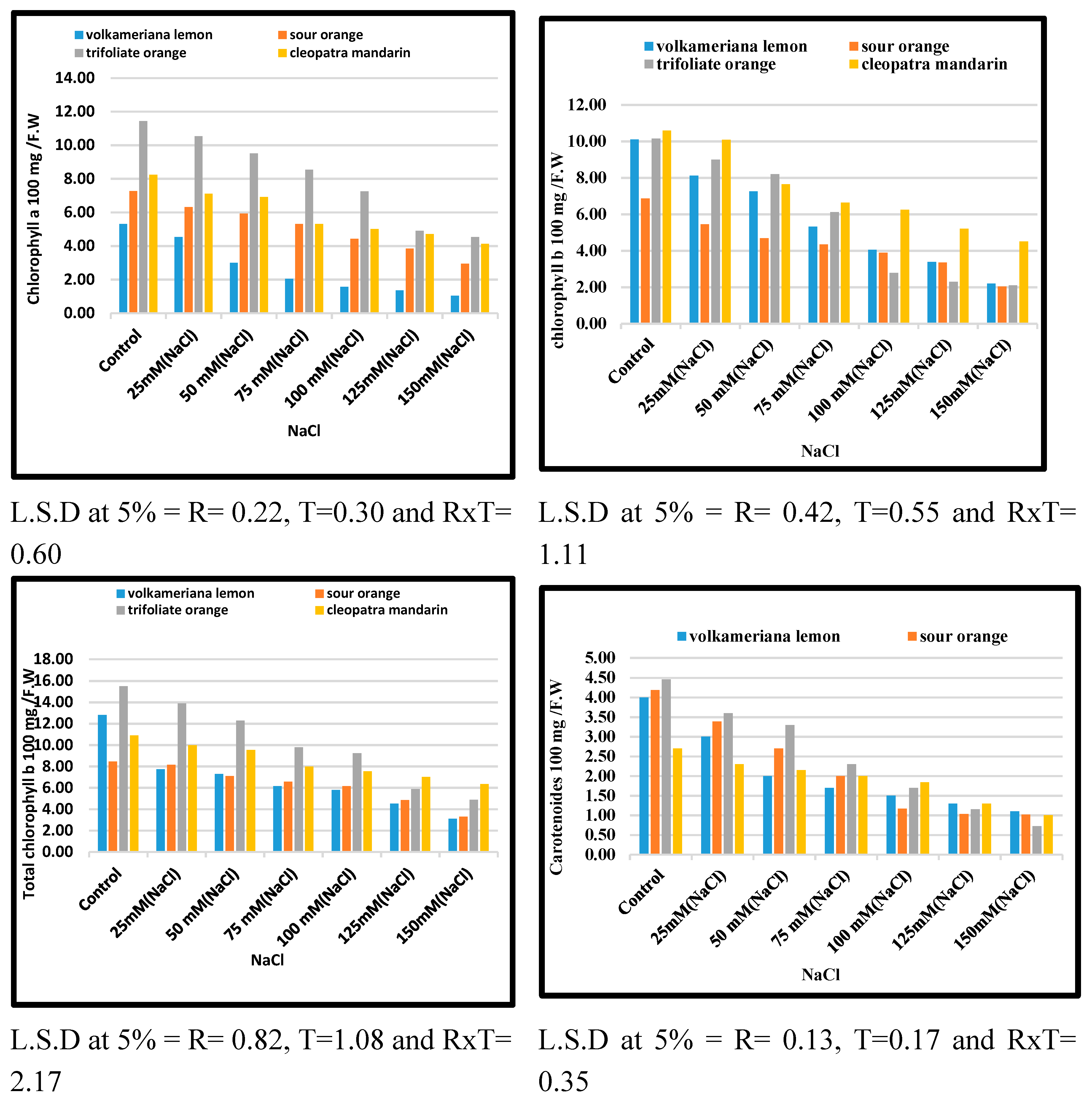
Figure 3.
Effect of MS media supplemented with NaCl at different concentrations on leaf-relative water contents of some citrus rootstocks growing in vitro.
Figure 3.
Effect of MS media supplemented with NaCl at different concentrations on leaf-relative water contents of some citrus rootstocks growing in vitro.
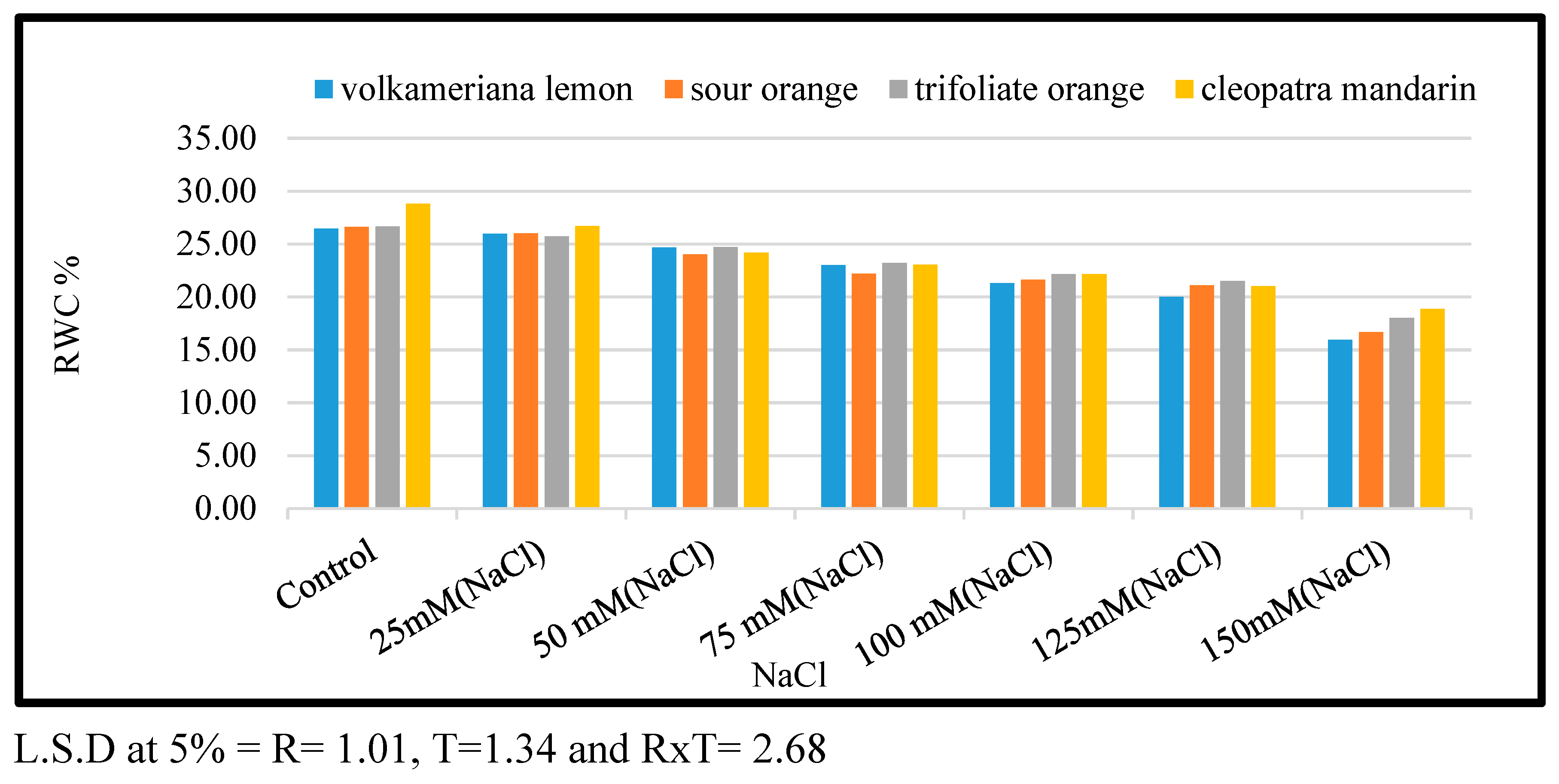
Figure 4.
Effect of MS media supplemented with NaCl at different concentrations on leaf proline content of some citrus rootstocks growing in vitro.
Figure 4.
Effect of MS media supplemented with NaCl at different concentrations on leaf proline content of some citrus rootstocks growing in vitro.
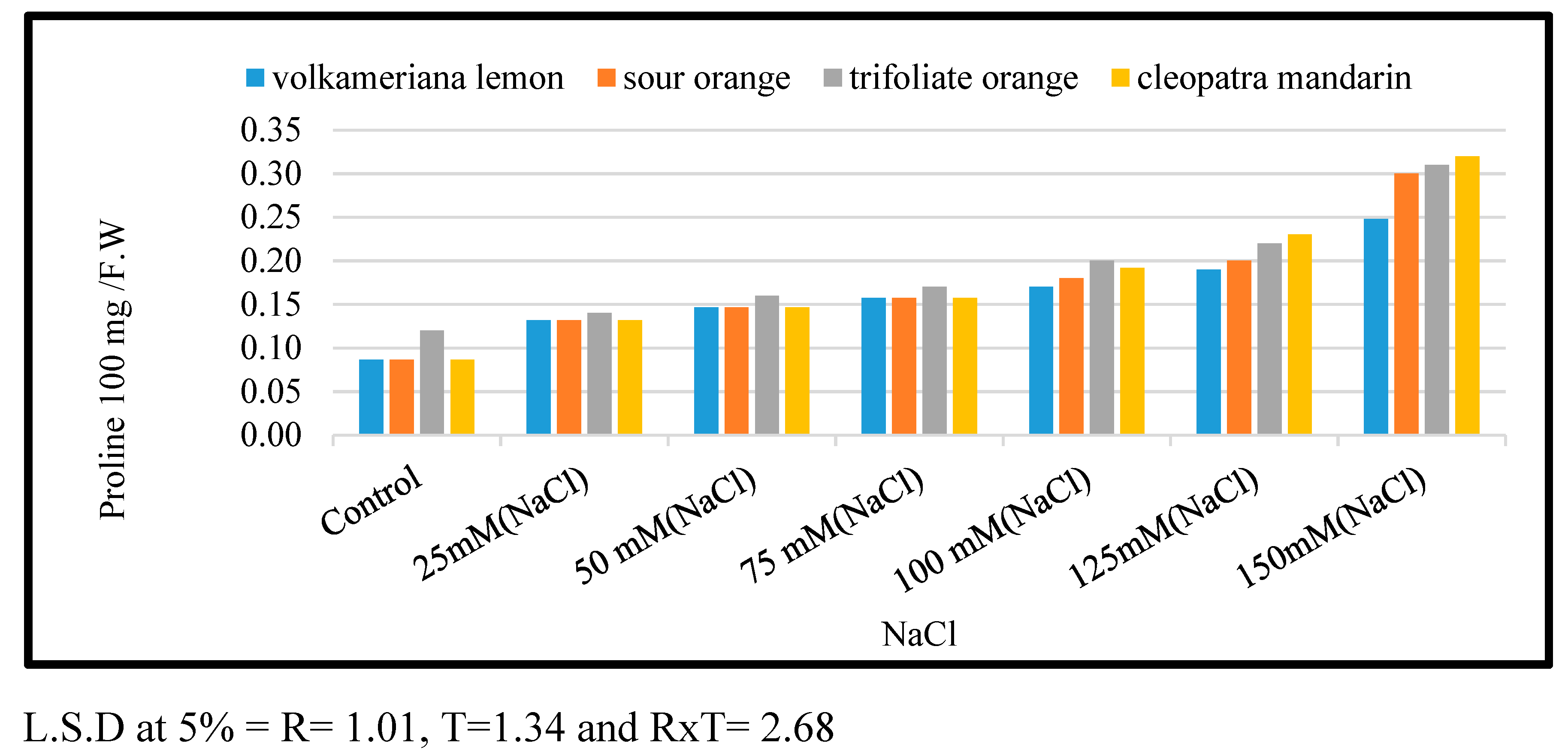
Figure 5.
Effect of MS media supplemented with NaCl at different concentrations on leaf biochemical characteristics of some citrus rootstocks growing in vitro. a, b control and 150 mM NaCl respectively of volkamariana lemon rootstock; c, d (control and 150 mM NaCl respectively) of sour orange rootstock; e, f (control and 150 mM NaCl respectively) of trifoliate orange rootstocks and g, h (control and 150 mM NaCl respectively) of Cleopatra mandarin rootstocks.
Figure 5.
Effect of MS media supplemented with NaCl at different concentrations on leaf biochemical characteristics of some citrus rootstocks growing in vitro. a, b control and 150 mM NaCl respectively of volkamariana lemon rootstock; c, d (control and 150 mM NaCl respectively) of sour orange rootstock; e, f (control and 150 mM NaCl respectively) of trifoliate orange rootstocks and g, h (control and 150 mM NaCl respectively) of Cleopatra mandarin rootstocks.
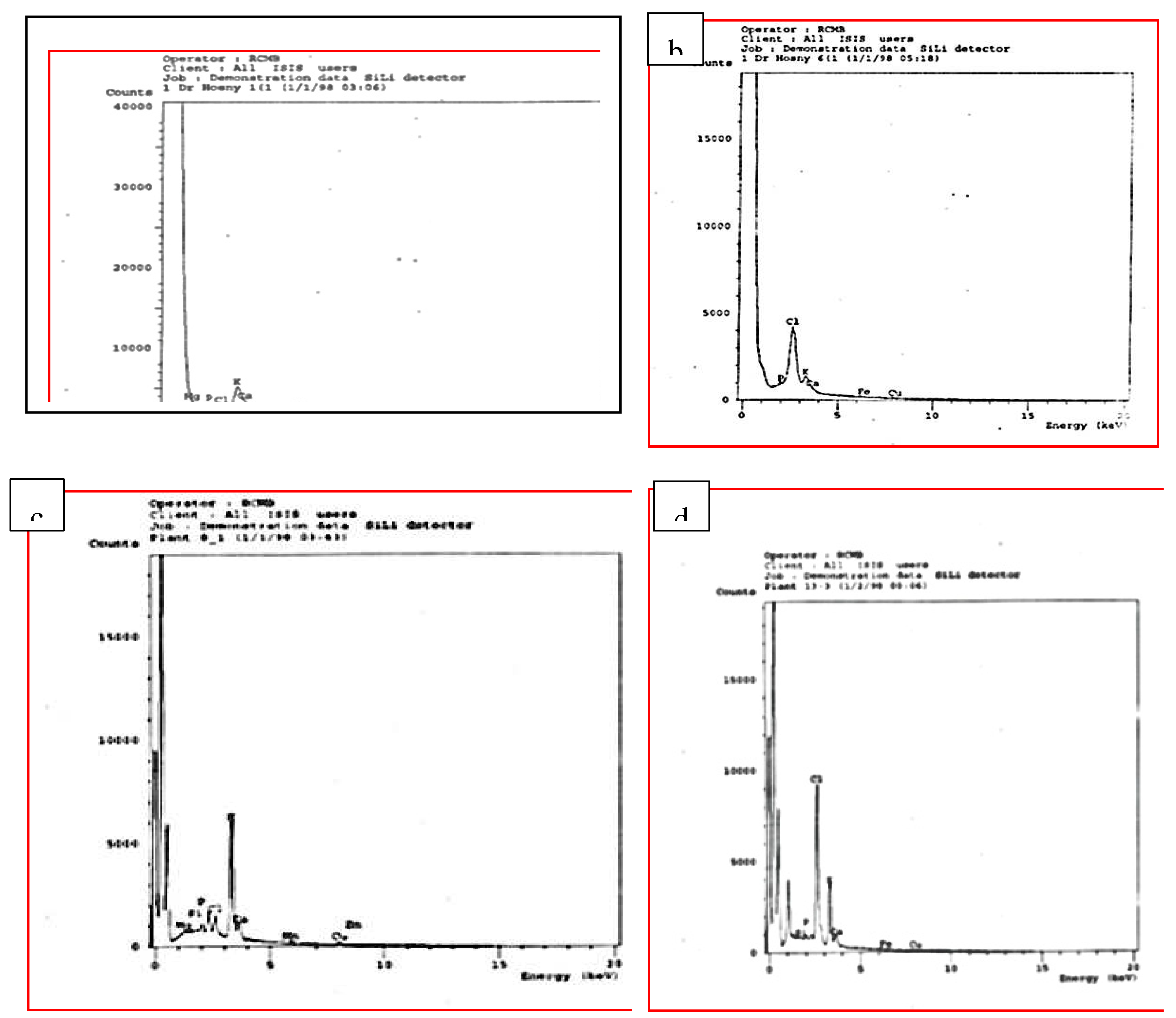
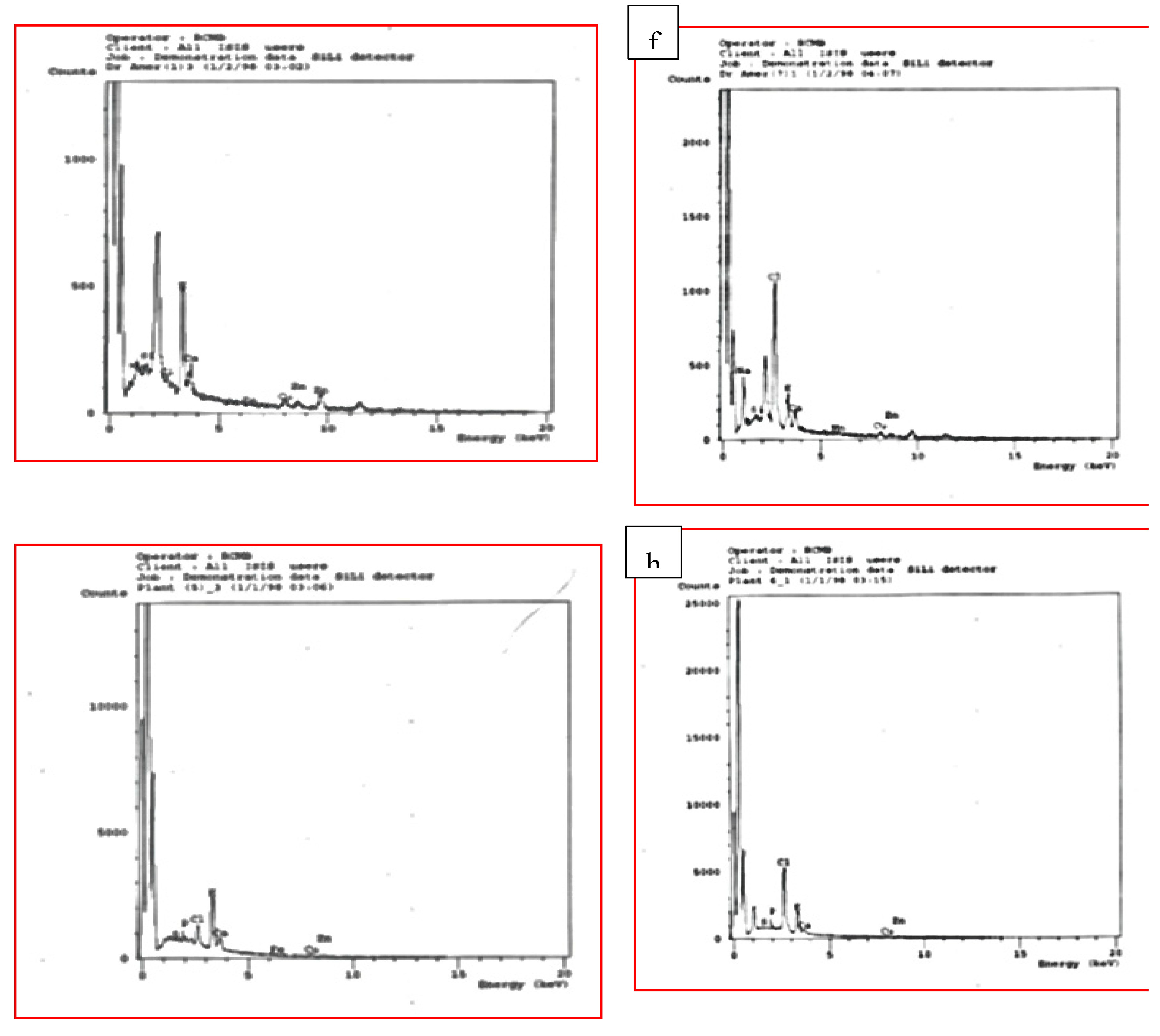
Figure 7.
Principal Components Analysis (PCA) Effect of MS media supplemented with NaCl at 0.0, 25, 50, 75, 100, 125, and 150 mM on micro-shoots of Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin.
Figure 7.
Principal Components Analysis (PCA) Effect of MS media supplemented with NaCl at 0.0, 25, 50, 75, 100, 125, and 150 mM on micro-shoots of Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin.
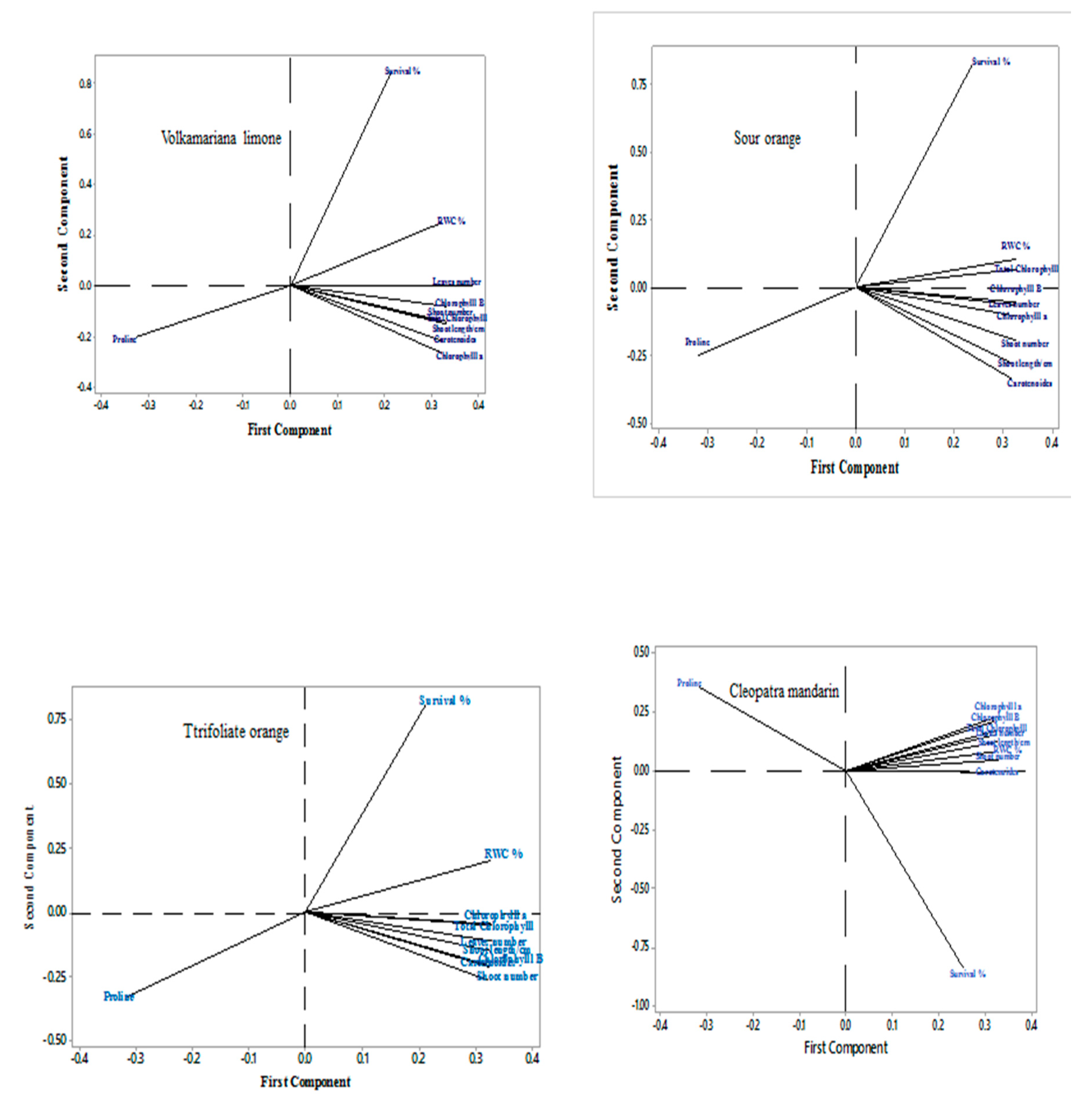

Table 1.
Results of principal component analysis (PCA) Effect of MS media supplemented with NaCl at 0.0, 25, 50, 75, 100, 125, and 150 mM on micro-shoots of Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin.
Table 1.
Results of principal component analysis (PCA) Effect of MS media supplemented with NaCl at 0.0, 25, 50, 75, 100, 125, and 150 mM on micro-shoots of Volkamariana lemon, sour orange, trifoliate orange, and Cleopatra mandarin.
| Cleopatra Mandarin | Sour Orange | ||||||
|---|---|---|---|---|---|---|---|
| Variable | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Eigenvalue | 9.2714 | 0.5646 | 0.075 | 9.1224 | 0.7171 | 0.0892 | |
| Variance | 92.7 | 5.6 | 0.007 | 91. 2 | 7.2 | 0.009 | |
| Cumulative | 92.7 | 98.4 | 0.991 | 91. 2 | 98.4 | 0.993 | |
| Components loadings | |||||||
| Shoot number | 0.328 | 0.046 | -0.100 | 0.326 | -0.195 | 0.112 | |
| Shoot length/cm | 0.327 | 0.125 | -0.017 | 0.318 | -0.279 | 0.34 | |
| Leaves number | 0.322 | 0.154 | 0.455 | 0.322 | -0.063 | -0.74 | |
| Survival % | 0.253 | -0.839 | 0.230 | 0.237 | 0.822 | 0.131 | |
| RWC % | 0.324 | 0.080 | -0.294 | 0.327 | 0.106 | 0.248 | |
| Chlorophyll a | 0.320 | 0.225 | 0.416 | 0.328 | -0.106 | -0.17 | |
| Chlorophyll B | 0.320 | 0.212 | -0.266 | 0.327 | -0.056 | 0.244 | |
| Total Chlorophyll | 0.324 | 0.168 | 0.314 | 0.328 | 0.065 | -0.351 | |
| Carotenoids | 0.321 | -0.005 | -0.521 | 0.316 | -0.335 | 0.175 | |
| Proline | -0.316 | 0.352 | 0.172 | 0.322 | -0.25 | -0.054 | |
| Trifoliate orange | volkamariana lemon | ||||||
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | ||
| Eigenvalue | 8.9933 | 0.9101 | 0.0494 | 8.9435 | 0.8147 | 0.1682 | |
| Variance | 89.9 | 9.1 | 0.005 | 89. 4 | 8. 1 | 0.017 | |
| Cumulative | 89.9 | 99 | 0.995 | 89. 4 | 97. 6 | 0.993 | |
| Components loadings | |||||||
| Shoot number | 0.322 | -0.264 | 0.18 | 0.329 | -0.153 | 0.106 | |
| Shoot length/cm | 0.328 | -0.152 | 0.014 | 0.331 | -0.142 | -0.13 | |
| Leaves number | 0.331 | -0.114 | 0.168 | 0.326 | 0.002 | 0.491 | |
| Survival % | 0.212 | 0.807 | 0.126 | 0.215 | 0.842 | -0.16 | |
| RWC % | 0.327 | 0.201 | 0.144 | 0.321 | 0.246 | 0.38 | |
| Chlorophyll a | 0.329 | -0.052 | -0.54 | 0.322 | -0.267 | 0.173 | |
| Chlorophyll B | 0.323 | -0.213 | 0.517 | 0.331 | -0.081 | 0.187 | |
| Total Chlorophyll | 0.33 | -4.60% | -0.586 | 0.318 | -0.142 | -0.604 | |
| Carotenoids | 0.326 | -0.208 | 0.035 | 0.324 | -0.22 | -0.274 | |
| Proline | -0.316 | -0.33 | -0.002 | -0.327 | -0.201 | 0.234 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



