
Submitted:
17 July 2023
Posted:
18 July 2023
You are already at the latest version
Abstract
We previously showed that mice with knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding the PGC-1a protein and nuclear factor erythroid 2 like 2 (NFE2L2) gene, showed some features of the age-related macular degeneration (AMD) phenotype. To further explore the mechanism behind the involvement of PGC-1a in AMD pathogenesis we used young (3-month) and old (12-month) mice with knockout in the PPARGC1A gene and age-matched wild-type (WT) animals. An immunohistochemical analysis showed age-dependent different expression of markers of oxidative stress defense, senescence and autophagy in the retinal pigment epithelium of KO animals as compared with their WT counterparts. Multivariate inference testing showed that senescence and autophagy proteins had the greatest impact on the discrimination between KO and WT 3-month animals, but proteins of antioxidant defense also contributed to that discrimination. A bioinformatic analysis showed that PGC-1a might coordinate the interplay between genes encoding proteins involved in antioxidant defense, senescence and autophagy in the aging retina. These data support importance of PGC-1a in AMD pathogenesis and confirm the utility of mice with PGC-1a knockout as an animal model to study AMD pathogenesis.
Keywords:
age-related macular degeneration
; aging retina
; peroxisome proliferator-activated receptor gamma coactivator 1-alpha
; PGC-1 AMD
; cellular senescence
; autophagy
; oxidative stress
Introduction
Age-related macular degeneration (AMD) is an eye disease affecting aging populations that may lead to legal blindness and sight loss with aging and oxidative stress consistently reported as AMD risk factors [1]. AMD affects the retinal pigment epithelium (RPE) cells with subsequent degeneration of photoreceptors, resulting in visual disturbances and ultimately – vision loss. AMD can be clinically divided into two distinct forms: dry and wet. The former is incurable but targeting the vascular endothelial growth factor A (VEGFA) with is antibodies and inhibitors of their receptors, may stop the progression of wet AMD and protect against sight loss [2]. Effective treatment of AMD is impeded by a poor knowledge of the disease pathogenesis, which is, at least in part, underlined by a restricted accessibility of the human retina for research and limited adequacy of cellular and animal models to mimic human AMD. We established an animal AMD model with mice carrying mutations in the nuclear factor erythroid 2 like 2 (NFE2L2) and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) genes (dKO mice), whose phenotype resembled dry AMD [3]. These transgenic animals showed a significant age-dependent RPE degeneration, an increase in the oxidative stress and endoplasmic reticulum markers, 4-HNE (4-hydroxynonenal) and GRP78 (glucose-regulated protein 78) and damaged mitochondria. Mice bearing mutations in the NFE2L2 and PPARGC1A genes displayed changes in protein ubiquitination and autophagy markers. These mice showed morphological defects in photoreceptors that were associated with vision loss. Therefore, the expression of NFE2L2 and PPARGC1A genes may play a role in AMD pathogenesis. In a subsequent work we showed that NFE2L2/PPARGC1A dKO mice showed some alterations in inflammatory pathways that are important in AMD pathogenesis [3]. We also showed that the dKO mice displayed an alternated expression of epithelial-mesenchymal transition (EMT) transcription factors [4]. Also, an increased immunoreactivity of senescence markers p16, basic helix-loop-helix family member E40 (DEC1) and high mobility group box 1 (HMGB1) suggesting that EMT and senescence pathways might intersect in the retinas of dKO mice, contributing to an AMD-like phenotype. Both EMT and cellular senescence may be stimulated by oxidative stress, which may be potentiated by the absence of NFE2L2 and PGC-1α genes, important in antioxidant defense [5].
NFE2L2 and PPARGC1A may be involved in AMD pathogenesis in several pathways and their combined effects may be related to the mechanism of the action of any single gene. Golestaneh and her coworkers showed that PGC-1α was important in AMD pathogenesis, mainly through its involvement in mitochondrial homeostasis, autophagy and metabolic pathways as well as an interaction with the anti-aging hormone, Klotho [6-9]. They used mice with knockout in the PPARGC1A gene, RPE from native AMD eyes and human RPE cells derived from induced pluripotent stem cells obtained from skin of AMD donors. They showed that PPARGCA1(+/–) mice expressing lower levels of PGC-1α than their WT counterparts displayed an AMD-like phenotype when administrated with high-fat diet, a consistently reported AMD risk factor [7]. Also, Saint-Geniez et al. using PPARGC1A KO mice showed an import role of PGC-1α in retinal angiogenesis, a key event in the pathogenesis of wet AMD [10]. They also showed that induction of PGC-1α promoted EMT and metabolism in RPE cells and protected them against oxidative damage [11, 12].
PGC-1α is a co-activator of nuclear receptors and transcription factors involved in the regulation of many aspects of cellular homeostasis, including the reaction to oxidative stress and aging [13, 14]. PGC-1α belongs to the peroxisome proliferator-activated receptor gamma coactivator-1 (PGC-1) family, which consists of PGC-1α, PGC-1β and PRC (PGC-1-related coactivator) [15]. Apart from oxidative stress, PGC-1α is involved in the regulation of cellular senescence and autophagy, processes that play an important role in AMD pathogenesis [16-18]. On the other hand, modification of autophagy inducers may regulate PGC-1α activity [19]. Generally, PGC-1α is expressed at relatively high levels in the mouse retina, so mice are a good subject to study PGC-1α influences [20]. Aging is per definition a crucial AMD risk factor and PGC-1α plays an important role in aging, mainly through its involvement in mitochondrial energy metabolism [13]. This justifies studies on the role of PGC-1a in the aging retina in AMD pathogenesis.
Inspired by these and our own results we further explored the role of PGC-1α in the pathways important in AMD pathogenesis: antioxidant defense, senescence and autophagy in young and old mice [11]. In the present work, we verified that PGC-1α played a role in the interplay between oxidative stress, senescence and autophagy in the aging retina. To do so, we immunohistochemically determined the expression of genes involved in oxidative stress defense, senescence and autophagy in transgenic mice carrying knock out in the PPARGC1A gene and WT animals. In addition, we performed a bioinformatic analysis of interactions between the genes, whose expression we determined.
Materials and Methods
Reagents
Tris-based antigen unmasking solution was purchased in Vector Laboratories Inc. (Burlingame, CA, USA), DAPI (4′,6-diamidino-2-phenylindole dihydrochlorided) was obtained from Sigma-Aldrich (St. Louis, MO, USA). Primary and secondary antibodies and their suppliers were listed in further text. Mowiol mounting media was made in our laboratory.
Animals
All animal experiments were performed according to the protocols that agreed with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and approved by the Project Authorization Board of Regional Administrative Agency for Southern Finland (ESAVI/8893/04.10.07/2014). The animals were maintained in a 12/12 h light-dark cycle at constant temperature 22 ± 1 °C and had a free access to drinking water and standard pellet chow.
Transgenic mice with constitutive knockout in the PPARGC1A gene (PGC-1a KO mice) were originally provided by Dr. Bruce Spiegelman of Dana Faber Cancer Institute, Philadelphia, PA, USA and generated from the C57BL/6J strain as described elsewhere [21]. The animals were genotyped as described previously [22]. The 3- or 12-month-old mice were sacrificed, and their eyes were collected immediately, enucleated and fixed in 2% paraformaldehyde for 2 h.
Immunohistochemistry
Gene expression in the mice RPE was evaluated by immunohistochemistry. We studied the expression of the following genes coding for proteins involved in the cellular response to oxidative stress: APEX1 (apurinic/apyrimidinic endodeoxyribonuclease 1, APE1), OGG1 (8-oxoguanine DNA glycosylase), PDIA2 (protein disulfide isomerase family A member 2), TXN (thioredoxin), SOD1 (superoxide dismutase 1) and H2AX (H2A.X variant histone, H2AX phosphorylated at Ser139). The genes encoding proteins involved in cellular senescence were: TP53 (tumor protein p53), HMGB1 (high mobility group box 1), CDKN1A (cyclin dependent kinase inhibitor 1A, p21), CDKN2A (cyclin dependent kinase inhibitor 2A, p16INK4a, p16) and BHLHE40 (basic helix-loop-helix family member E40, DEC1). The genes involved in macroautophagy and mitophagy were BECN1 (BECLIN1), LAMP2 (lysosomal associated membrane protein 2), MAP1LC3B (microtubule associated protein 1 light chain 3 beta, LC3B), SQSTM1 (sequestosome 1, p62), UBB (ubiquitin B), PINK1 (PTEN induced kinase 1) and PRKN (parkin RBR E3 ubiquitin protein ligase, PRK8).
Tissues sections were deparaffinized using xylene, rehydrated, washed and pretreated with Tris-based antigen unmasking solution for 7.5 min at 90 °C. The sections were encircled with a PAP pen and quenched with 0.1 M glycine in PBS for 10 min prior to a 0.1% Triton-X wash for 10 min before proceeding with blocking for 30 min. For single staining, quenched slides were incubated with 20% goat serum for 30 min before adding a primary antibody and incubated 24 h at 4 °C. Then, the slides were washed with TBS and incubated with the secondary antibody for 3 h at room temperature. For double staining (PINK1/PRKN), the first primary antibody (PINK1) was added followed by its secondary antibody and then the second primary antibody (PRKN) was added, followed by its secondary antibody. The secondary antibodies were goat anti-rabbit Alexa Fluor 488 and 594 (A11034/A11037) and goat anti-mouse Alexa Fluor 594 (A11032) (Thermo Fisher Scientific, Waltham, MA, USA) diluted at 1:500 in TBS.
Data acquiring
The slides were mounted using the Mowiol mounting media and stored in the dark at room temperature. The stained sections were examined with a confocal microscope (Carl Zeiss AX10 Imager A2, Carl Zeiss Microscopy, Jena, Germany) using a 63× (NA:1.42. Plan Apochromat) oil (Zeiss Immersol™) immersion objective. At least nine repetitive images were taken from each section for all markers. Images were color enhanced using Adobe photoshop for visual representation. All the captured images were converted into 8-bit and processed using ImageJ v1.53t (https://imagej.nih.gov/ij/). The background was subtracted using a default rolling ball radius method. Regions of interest (ROI) were drawn over RPE cell layer followed by colocalization analysis using 2 channel spots colocalization analyzer ComDet v0.5.5 (https://imagej.net/plugins/spots-colocalization-comdet), an ImageJ plugin. The total number of puncta from each color (channel) was calculated and the corresponding correlations of colocalization were measured. The mean grey value was used to quantify optical changes seen in images.
Data analysis
Six (3 KO and 3 WT) or 8 (4 KO and 4 WT) mice aged 3 months (young) or 12 months (old) respectively, were used in this study. Data were presented as the BCA bootstrap-boosted means + SD or median with interquartile range (lower quartile [25%] to upper quartile [75%]), depending on data distribution. Data normality and variance homogeneity of the acquired data were verified using Shapiro–Wilk’s or Levene’s test, respectively. The data complying with the assumptions of normal distribution and homogeneity of variances were analyzed with Student’s t-test and one-way or two-way ANOVA (either block or main component model or the model with interactions, both in the version with replicates), while for the remaining data with a Mann-Whitney rank sum U test and Kruskal-Wallis test. In order to assess the significance of differences between particular samples, we used the post-hoc multiple comparisons tests (the least significant difference test [LSD] or the Bonferroni’s correction for multiple comparisons). The resampling bootstrap technique (10,000 iterations) was used to minimize the risk that the revealed differences were observed by pure chance. Statistical analyses were performed using Statistica v.13 (Dell Inc., Tulsa, OH, USA), GraphPad Prism for Windows ver. 9.5 (GraphPad Software, Boston, MA, USA), Resampling Stats Add-In for Excel v.4 (The Institute for Statistics Education, An Elder Research Company, Arlington, VA, USA) and R Package Software v. 4.4, in which we used the algorithm for data resampling in various employed analyses, written by one of the authors (J.K.).
Multivariate inference testing was accomplished in three alternative approaches. First, we used linear discriminant analysis (LDA) to verify which proteins in three sets (antioxidant defense, senescence and autophagy) contributed mostly to the best discrimination between KO and WT mice. Second, we employed MANOVA and T2 Hotelling test to verify whether the overall sets of proteins studied differed between KO and WT animals. Third, we transformed real values in each variable into normal scores according to the van der Waerden formula [23]. Next, we calculated cumulative scores (CSs) by summing up the relevant normal scores across any given set of proteins (antioxidant defense, senescence or autophagy), or alternatively across all variables. The resultant CSs were further used for the inference analyses.
In bioinformatic analysis, gene expression data were normalized with interquartile range method. Principal component analysis (PCA) was performed with FactoMineR and factoextra packages in the R 4.2.2 for Windows software (R Development Core Team, Vienna, Austria). Interaction networks were constructed using the StringApp for Cytoscape, an open source bioinformatic software (version 3.9.1). A score exceeding 0.4 was set as a threshold for considering an interaction.
Results
Firstly, we validated the main experimental method we used, immunohistochemistry, to verify the lack of PGC-1α in the KO animals. Although a background signal was relatively strong, we confirmed the lack of the expression of the PPARGCA1 gene in the PGC-1α KO mice (results not shown). Moreover, we validated immunohistochemistry as a suitable method in our experimental conditions.
Antioxidant defense
Figure 1 presents results of immunohistochemical determination of the expression of markers of antioxidant defense in young (3-month) and old (12-month) mice with knockout in the gene encoding PGC-1α. APE1, OGG1 and SOD1 increased in 3-month KO animals, whereas H2AX (pSer139) and TDX – decreased. No changes were observed in PDIA2. 12-month KO mice displayed higher expression of APE1 and H2AX (pSer239), while OGG1, PDIA2, TDX and SOD1 decreased as compared with their WT counterparts.
As the results of the expression of all markers were influenced by two variables, the expression of the PGCRCA1 gene (KO vs. WT) and animal age (3 vs. 12-month), we investigated the interaction between these two factors in their effect on the markers expression. We observed that the expression of APE1, PDIA2, TDX were affected by both age and the presence/absence of PGC-1α (p < 0.05).
Senescence
Three-month KO animals displayed increased expression of p16, p53, DEC1 and HMGB1, while p21 did not change (Figure 2). In 12-month mice, DEC1 and HMGB1 increased in KO as compared with WT animals. No changes were observed in the expression of p16, p21 and p53 markers in KO and WT animals.
The expressions of p16, p53 and DEC1 we affected by both time and the presence/absence of PGC-1α and these factors interacted (p < 0.001).
Autophagy
Ubiquitin and BECN1 showed an increased expression in 3-month KO animals as compared with their WT counterparts, while p62, LAMP-2 and LC3B were not changed (Figure 3). 12-month KO mice showed an increased expression of all six autophagic markers as compared with the WT animals. The expressions of p62, LAMP2 and the PINK1/PRKN ratio were affected by both age and PGC-1a and there was an interaction between them (p < 0.05 for all factors).
Multivariate interference testing
To determine contribution of a specific group of proteins (antioxidant defense, senescence and autophagy) and a specific protein within each group to the discrimination between PGC-1α KO and WT cells a multivariate interference testing was performed [24].
We observed a stronger discrimination between PGC-1a KO and WT mice for 12-month than 3-month animals as we noted a greater number of markers contributing to differences between the KO and WT in old than in young animals and these differences were greater in the former than the latter (Table 1).
Bioinformatic analysis
To assess the role of PGC-1α and aging in mutual interactions between genes and proteins of antioxidant defense, senescence and autophagy, a bioinformatic analysis was performed.
Although our work was far from typical high-throughput analysis, the number of analyzed genes (17), and variants of age (2) and PGC-1α expression (2) made it necessary to reduce dimensions, which can be done by principal component analysis (PCA) [25]. Figure 4 presents PCA with dimensional reduction on PGC-1α expression (A, B) and age (C, D) as well as data without dimensional reduction. It can be seen that PGC-1α expression moderately separated the data (A and B), but when the separation was done by age, the data were strongly split into two non-overlapping clusters (C and D), which was confirmed by PCA for all data (E).
To assess the contribution of specific genes to the total gene expression, a PCA correlation plot was made (Figure 5). The genes whose expression most significantly contributed to the total expression were BECN1 (autophagy), HMGB1 (senescence), PDIA2 (antioxidant defense) and CDKN2A (senescence). The contribution of specific genes strongly depended on age and PGC-1α expression.
The building of the network of interactions on the String Cytoscape basis led to the conclusion that in 3-month animals PGC-1α knockout had the most pronounced effect on the expression of the BECN1 (autophagy, an increase) and H2AX (antioxidant defense, a decrease). In 12-month animals the most pronounced differences between knockout and WT animals were for the SQSTM1 (autophagy, an increase), HMGB1 (senescence, an increase), SOD1 and TXN (both antioxidant defense, a decrease). The interaction network showed that, as expected, the greatest number of interactions displayed the product of the TP53 gene, p53. On the other hand, the product of the PDIA2 gene directly interacted only with the TXN gene product, similarly to the unique interaction of the BHLHE40 (senescence) with TP53.
Finally, we included the PPARGC1A in the network of interactions built with String Cytoscape (Figure 6). PPARCG1A directly interacted with a substantial number of proteins and only TP53 had a greater number of interactions.
Discussion
We used slices of mouse eye sections and determined the expression of markers in situ in RPE cells. Studying the expression of markers in the RPE in the slices of the eye better reflects the reality than similar studies in the RPE-derived cell lines. In general, cells in the central part of the retina, including the macula, are quiescent and do not divide due to spatial constraints. They are not post-mitotic cells as they can reinitiate cell cycle in vitro, like ARPE-19 cells. Proliferation may affect the expression of genes and there is not a general rule establishing a specific relationship between proliferation and expression of all genes [26]. Therefore, the levels of the markers determined by immunohistochemistry may be more adequate to in vivo situation than immunocytochemistry. However, the question remains, what we determined – gene expression on the protein level or immunohistochemical protein abundance? Immunohistochemistry may give more appropriate information on gene expression in the RPE than determination of protein expression in proliferating RPE cells in vitro. The next question is why we did not apply a high-throughput analysis with the use e.g., microarrays dedicated to antioxidant defense, senescence and autophagy? The answer is the same – mRNA must have been isolated from primary cultures of RPE cells that are difficult to obtain in required quantity and purity and they do not reflect the processes occurring in situ, including gene expression.
H2AX is not a gene of antioxidant defense per se, as it does not code for antioxidant enzymes nor DNA repair protein. However, it may be considered as a marker of DNA damage and its repair, and if the damage is induced by oxidative stress, it is a marker of antioxidant defense as it codes for a variant of H2A histone, H2A.X, which, when phosphorylated at Ser186, colocalizes with DNA repair proteins at the sites of DNA double strand breaks [27].
Results of immunohistochemical studies supported by a deep statistical and bioinformatic analyses suggest that the expression of genes of antioxidant defense, senescence and autophagy depends on age and the presence/absence of the active PPARGC1 gene encoding the PGC-1a protein. Moreover, the interplay between these three categories of genes may occur and it can be coordinated by the PGC-1a protein and depend on age.
We consider as the most important results of our studies that aging may dramatically change the reaction of the retina on the presence and absence of PGC-1α. For example, the level of SOD1 increased in young KO mice, whereas it decreased in their old counterparts, when considered without interaction between aging and the presence or absence of PGC-1α. The main function of SOD1 is converting the radical superoxide into molecular oxygen and hydrogen peroxide, through redox reactions [28]. Therefore, we can interpolate that in young animals a lack of PGC-1α increased the expression of SOD1, but aging might attenuate that reaction, finally leading to a decreased level of SOD1 in old animals. Therefore, we can consider that the first reaction of the RPE cells to the lack of PGC-1α was the increase in the expression of SOD1 to recompensate its lack, but then during aging, it decreased. It is important to underline that it was a general picture of the expression of an antioxidant defense protein, not induced by oxidative stress as we expected that the situation may be different in the latter case.
Multivariate testing analysis showed that autophagy and senescence, in contrary to antioxidant defense, discriminated between 3-month WT and KO animals. The most significantly discriminating proteins in the autophagy group were BECLIN1 encoded by the BECN1 gene and ubiquitin encoded by the UBB gene. BECLIN1 is a component of the class III phosphatidylinositol 3-kinase (PI3K-III) complex, which is crucial in the assembly of autophagosomes [29]. Moreover, it may be important in the regulation of pro-life and pro-death functions of autophagy and its connections with apoptosis [30]. We presented arguments that autophagy might regulate RPE cells death in AMD [31]. Moreover, BECLIN1 is regulated by a cascade of ubiquitination events [32]. Reversible ubiquitination of essential autophagy inducers is an important mechanisms autophagy regulation [33]. Moreover, a crosstalk between autophagy and ubiquitin-proteasome system is crucial for the management of cellular waste, including all-trans retinal, drusen and lipofuscin, containing unfolded, damaged and no longer needed proteins in RPE cells [34]. Therefore, ubiquitin, as a cofactor for these processes, may be important in AMD pathogenesis.
The stepwise forward LDA in 3- and 12-month mice demonstrated that the groups of markers we chose distinguished between KO and WT animals.
Our bioinformatic analysis confirmed an interplay between some genes of antioxidant defense, senescence and autophagy. However, the expression of certain genes was negatively correlated with others. This confirms a complex relationship between the regulation of gene expression and phenotype. Our bioinformatic analysis also showed that p53 had the largest interacting network with other proteins that were studied in this work. It is not very surprised as p53 is involved in antioxidant defense, senescence and autophagy and plays important roles in these effects [35-37].
As we mentioned, the Golestaneh lab showed that PGC-1α was essential in mitochondrial homeostasis in RPE cells obtained from differentiation of iPSCs obtained from AMD donors [6]. In this work we investigated two variables that were directly related to mitochondrial quality control (mtQC): PINK1 and PRKN. These proteins, along with optineurin (OPT), belong to the main proteins of the mitophagy pathway [38]. In normal conditions, PINK1 is imported into and degraded within mitochondrial inside, but when a damage occurs, PINK1 is accumulated on the outer mitochondrial membrane and PRKN is recruited from the cytosol [39]. Therefore, the PINK1/PRKN ratio is a good indicator of damaged mitochondria and mitophagy. We did not observe any influence of PGC-1α on the level of PINK1/PRKN in young mice, but this ratio increased in old KO animals, indicating that the lack of PGC-1α might result in mitochondrial damage (Figure 5). Dysfunctional mitochondria and impaired mitophagy in RPE cells are important elements in AMD pathogenesis [40]. However, we investigated other proteins than PINK1 and PRKN that might be associated with mitochondrial maintenance in RPE cells as antioxidant defense and senescence interwind in mtQC and along with inflammation may contribute to neurodegeneration [41].
It was shown that different isoforms of vascular endothelial growth factor (VEGF), a molecule that is critical for choroidal neovascularization and wet AMD, are differentially expressed during mouse development and in its adulthood [42]. However, PGC-1α can be, at least in part, replaced by its analogue, PGC-1β, so apart from PGC-1α knockout, PGC-1β and PGC-1α/PGC-1β knockouts should also be used to completely asses the role of the PGC-1 family in AMD pathogenesis [43].
In summary, PGC-1α may be involved in the interplay between antioxidant defense, senescence and autophagy, important in AMD pathogenesis and aging may strongly modulate this interplay. Mice with knockout in the PPARGC1A gene may be suitable to investigate early and late signs of PGC-1α-related effects in AMD pathogenesis and therefore provide model for novel drug development.
Author contribution
Iswariyaraja Sridevi Gurubaran – data acquisition; review and editing; Cezary Watala – data analysis; review and editing; Joanna Kostanek – bioinformatic analysis, review and editing; Elzbieta Pawlowska – review and editing; Joanna Szczepanska – review and editing; Kai Kaarniranta – conceptualization, review and editing; Janusz Blasiak –conceptualization, writing (original draft) and editing.
Acknowledgements
This work was supported by National Science Centre, Poland grant number 2017/27/B/NZ3/00872 (JB), the Academy of Finland Grants (296840, 333302, GeneCellNano Flagship (KK) and Sigrig Juselius Foundation (KK). We thank Juha Hyttinen, Anne Seppänen and Elisa Toropainen for technical assistance.
References
- Lim LS, Mitchell P, Seddon JM, Holz FG, Wong TY. Age-related macular degeneration. Lancet 2012, 379, 1728–1738.
- Heier JS, Brown DM, Chong V, Korobelnik JF, Kaiser PK, Nguyen QD, et al. . Intravitreal aflibercept (VEGF trap-eye) in wet age-related macular degeneration. Ophthalmology 2012, 119, 2537–2548.
- Sridevi Gurubaran I, Heloterä H, Marry S, Koskela A, Hyttinen JMT, Paterno JJ, et al. (2021). Oxidative Stress and Mitochondrial Damage in Dry Age-Related Macular Degeneration Like NFE2L2/PGC-1α (-/-) Mouse Model Evoke Complement Component C5a Independent of C3. Biology (Basel), 10.
- Blasiak J, Koskela A, Pawlowska E, Liukkonen M, Ruuth J, Toropainen E, et al. (2021). Epithelial-Mesenchymal Transition and Senescence in the Retinal Pigment Epithelium of NFE2L2/PGC-1α Double Knock-Out Mice. Int J Mol Sci, 22.
- Aquilano K, Baldelli S, Pagliei B, Cannata SM, Rotilio G, Ciriolo MR. p53 orchestrates the PGC-1α-mediated antioxidant response upon mild redox and metabolic imbalance. Antioxid Redox Signal 2013, 18, 386–399.
- Golestaneh N, Chu Y, Cheng SK, Cao H, Poliakov E, Berinstein DM. Repressed SIRT1/PGC-1α pathway and mitochondrial disintegration in iPSC-derived RPE disease model of age-related macular degeneration. J Transl Med 2016, 14, 344.
- Zhang M, Chu Y, Mowery J, Konkel B, Galli S, Theos AC, et al. (2018). Pgc-1α repression and high-fat diet induce age-related macular degeneration-like phenotypes in mice. Dis Model Mech, 11.
- Zhang M, Jiang N, Chu Y, Postnikova O, Varghese R, Horvath A, et al. . Dysregulated metabolic pathways in age-related macular degeneration. Sci Rep 2020, 10, 2464.
- Zhou S, Hum J, Taskintuna K, Olaya S, Steinman J, Ma J, et al. (2023). The Anti-Aging Hormone Klotho Promotes Retinal Pigment Epithelium Cell Viability and Metabolism by Activating the AMPK/PGC-1α Pathway. Antioxidants (Basel), 12.
- Saint-Geniez M, Jiang A, Abend S, Liu L, Sweigard H, Connor KM, et al. . PGC-1α regulates normal and pathological angiogenesis in the retina. Am J Pathol 2013, 182, 255–265.
- Rosales MAB, Shu DY, Iacovelli J, Saint-Geniez M (2019). Loss of PGC-1α in RPE induces mesenchymal transition and promotes retinal degeneration. Life Sci Alliance, 2.
- Satish S, Philipose H, Rosales MAB, Saint-Geniez M. Pharmaceutical Induction of PGC-1α Promotes Retinal Pigment Epithelial Cell Metabolism and Protects against Oxidative Damage. Oxid Med Cell Longev 2018, 2018, 9248640.
- Anderson R, Prolla T. PGC-1alpha in aging and anti-aging interventions. Biochim Biophys Acta 2009, 1790, 1059–1066.
- Wenz T. Regulation of mitochondrial biogenesis and PGC-1α under cellular stress. Mitochondrion 2013, 13, 134–142.
- Handschin C, Spiegelman BM. Peroxisome proliferator-activated receptor gamma coactivator 1 coactivators, energy homeostasis, and metabolism. Endocr Rev 2006, 27, 728–735.
- Blasiak J. Senescence in the pathogenesis of age-related macular degeneration. Cell Mol Life Sci 2020, 77, 789–805.
- Kaarniranta K, Blasiak J, Liton P, Boulton M, Klionsky DJ, Sinha D. Autophagy in age-related macular degeneration. Autophagy 2023, 19, 388–400.
- Salazar G, Cullen A, Huang J, Zhao Y, Serino A, Hilenski L, et al. . SQSTM1/p62 and PPARGC1A/PGC-1alpha at the interface of autophagy and vascular senescence. Autophagy 2020, 16, 1092–1110.
- Jäger S, Handschin C, St-Pierre J, Spiegelman BM. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc Natl Acad Sci U S A 2007, 104, 12017–12022.
- Egger A, Samardzija M, Sothilingam V, Tanimoto N, Lange C, Salatino S, et al. (2012). PGC-1α determines light damage susceptibility of the murine retina. PLoS One, 7:e31272.
- Handschin C, Choi CS, Chin S, Kim S, Kawamori D, Kurpad AJ, et al. . Abnormal glucose homeostasis in skeletal muscle-specific PGC-1alpha knockout mice reveals skeletal muscle-pancreatic beta cell crosstalk. J Clin Invest 2007, 117, 3463–3474.
- Felszeghy S, Viiri J, Paterno JJ, Hyttinen JMT, Koskela A, Chen M, et al. . Loss of NRF-2 and PGC-1alpha genes leads to retinal pigment epithelium damage resembling dry age-related macular degeneration. Redox Biol 2019, 20, 1–12.
- Landman B, Robertson A, Culver C (2005). Some New Exact van der Waerden Numbers.
- Brown CD, Ridder TD. Framework for multivariate selectivity analysis, part I: theoretical and practical merits. Appl Spectrosc 2005, 59, 787–803.
- Li Z, Safo SE, Long Q. Incorporating biological information in sparse principal component analysis with application to genomic data. BMC Bioinformatics 2017, 18, 332.
- Locard-Paulet M, Palasca O, Jensen LJ (2022). Identifying the genes impacted by cell proliferation in proteomics and transcriptomics studies. PLoS Comput Biol, 18:e1010604.
- Chowdhury D, Keogh M-C, Ishii H, Peterson CL, Buratowski S, Lieberman J. γ-H2AX Dephosphorylation by Protein Phosphatase 2A Facilitates DNA Double-Strand Break Repair. Molecular Cell 2005, 20, 801–809.
- Eleutherio ECA, Silva Magalhães RS, de Araújo Brasil A, Monteiro Neto JR, de Holanda Paranhos L. SOD1, more than just an antioxidant. Arch Biochem Biophys 2021, 697, 108701.
- McKnight NC, Zhenyu Y. Beclin 1, an Essential Component and Master Regulator of PI3K-III in Health and Disease. Curr Pathobiol Rep 2013, 1, 231–238.
- Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ 2011, 18, 571–580.
- Kaarniranta K, Tokarz P, Koskela A, Paterno J, Blasiak J. Autophagy regulates death of retinal pigment epithelium cells in age-related macular degeneration. Cell Biol Toxicol 2017, 33, 113–128.
- Boutouja F, Brinkmeier R, Mastalski T, El Magraoui F, Platta HW (2017). Regulation of the Tumor-Suppressor BECLIN 1 by Distinct Ubiquitination Cascades. Int J Mol Sci, 18.
- Klionsky DJ. Autophagy: from phenomenology to molecular understanding in less than a decade. Nat Rev Mol Cell Biol 2007, 8, 931–937.
- Blasiak J, Pawlowska E, Szczepanska J, Kaarniranta K (2019). Interplay between Autophagy and the Ubiquitin-Proteasome System and Its Role in the Pathogenesis of Age-Related Macular Degeneration. Int J Mol Sci, 20.
- Budanov AV. The role of tumor suppressor p53 in the antioxidant defense and metabolism. Subcell Biochem 2014, 85, 337–358.
- Rufini A, Tucci P, Celardo I, Melino G. Senescence and aging: the critical roles of p53. Oncogene 2013, 32, 5129–5143.
- White E (2016). Autophagy and p53. Cold Spring Harb Perspect Med, 6:a026120.
- Ge P, Dawson VL, Dawson TM. PINK1 and Parkin mitochondrial quality control: a source of regional vulnerability in Parkinson’s disease. Molecular Neurodegeneration 2020, 15, 20.
- Matsuda N, Sato S, Shiba K, Okatsu K, Saisho K, Gautier CA, et al. . PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J Cell Biol 2010, 189, 211–221.
- Kaarniranta K, Uusitalo H, Blasiak J, Felszeghy S, Kannan R, Kauppinen A, et al. . Mechanisms of mitochondrial dysfunction and their impact on age-related macular degeneration. Prog Retin Eye Res 2020, 79, 100858.
- Picca A, Calvani R, Coelho-Junior HJ, Landi F, Bernabei R, Marzetti E (2020). Mitochondrial Dysfunction, Oxidative Stress, and Neuroinflammation: Intertwined Roads to Neurodegeneration. Antioxidants (Basel), 9.
- Ng YS, Rohan R, Sunday ME, Demello DE, D'Amore PA. Differential expression of VEGF isoforms in mouse during development and in the adult. Dev Dyn 2001, 220, 112–121.
- Popov DV, Lysenko EA, Kuzmin IV, Vinogradova V, Grigoriev AI. Regulation of PGC-1α Isoform Expression in Skeletal Muscles. Acta Naturae 2015, 7, 48–59.
Figure 1.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of antioxidant defense in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of antioxidant defense: APE1 (apurinic/apyrimidinic endodeoxyribonuclease 1), OGG1 (8-oxoguanine DNA glycosylase), PDIA2 (protein disulfide isomerase family A member 2), TDX (thioredoxin), SOD1 (superoxide dismutase 1) and H2AX (H2A.X variant histone H2AX phosphorylated at Ser139) in RPE of wild-type (WT) and mice with the global knockout in the PPARGC1 gene encoding the PGC-1α protein (KO) aged 3 or 12 months. Puncta specific to the label of the primary antibody to each marker have the same color as its abbreviation displayed between image panels. DAPI was used to stain the nuclei of RPE cells (blue). Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars represent minimum and maximal values; n = 76-175; * – p < 0.05, ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
Figure 1.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of antioxidant defense in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of antioxidant defense: APE1 (apurinic/apyrimidinic endodeoxyribonuclease 1), OGG1 (8-oxoguanine DNA glycosylase), PDIA2 (protein disulfide isomerase family A member 2), TDX (thioredoxin), SOD1 (superoxide dismutase 1) and H2AX (H2A.X variant histone H2AX phosphorylated at Ser139) in RPE of wild-type (WT) and mice with the global knockout in the PPARGC1 gene encoding the PGC-1α protein (KO) aged 3 or 12 months. Puncta specific to the label of the primary antibody to each marker have the same color as its abbreviation displayed between image panels. DAPI was used to stain the nuclei of RPE cells (blue). Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars represent minimum and maximal values; n = 76-175; * – p < 0.05, ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
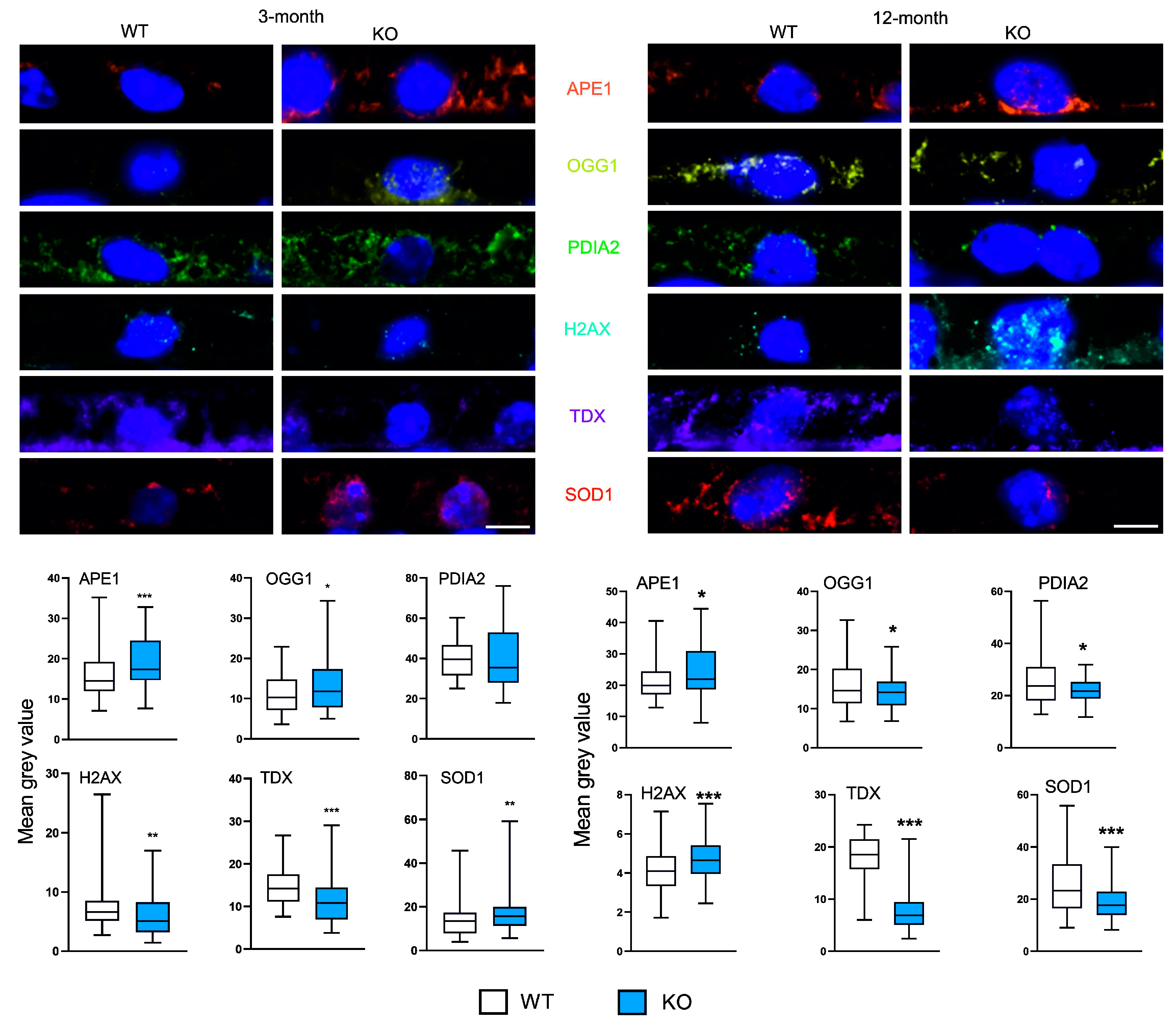
Figure 2.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of senescence in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of senescence: p16 (cyclin dependent kinase inhibitor 2A), p21 (cyclin dependent kinase inhibitor 1A), p53 (tumor protein p53), DEC1 (basic helix-loop-helix family member E40) and HMGB1 (high mobility group box 1) in RPE of wild-type (WT) and mice with the global knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha gene (PPARGC1A) encoding the PGC-1α protein (KO). Puncta specific to the label of the primary antibody to each marker have the same color as their abbreviations between images. DAPI was used to stain the nuclei of RPE cells (blue). Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars present minimum and maximal values; n = 51-105; ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
Figure 2.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of senescence in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of senescence: p16 (cyclin dependent kinase inhibitor 2A), p21 (cyclin dependent kinase inhibitor 1A), p53 (tumor protein p53), DEC1 (basic helix-loop-helix family member E40) and HMGB1 (high mobility group box 1) in RPE of wild-type (WT) and mice with the global knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha gene (PPARGC1A) encoding the PGC-1α protein (KO). Puncta specific to the label of the primary antibody to each marker have the same color as their abbreviations between images. DAPI was used to stain the nuclei of RPE cells (blue). Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars present minimum and maximal values; n = 51-105; ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
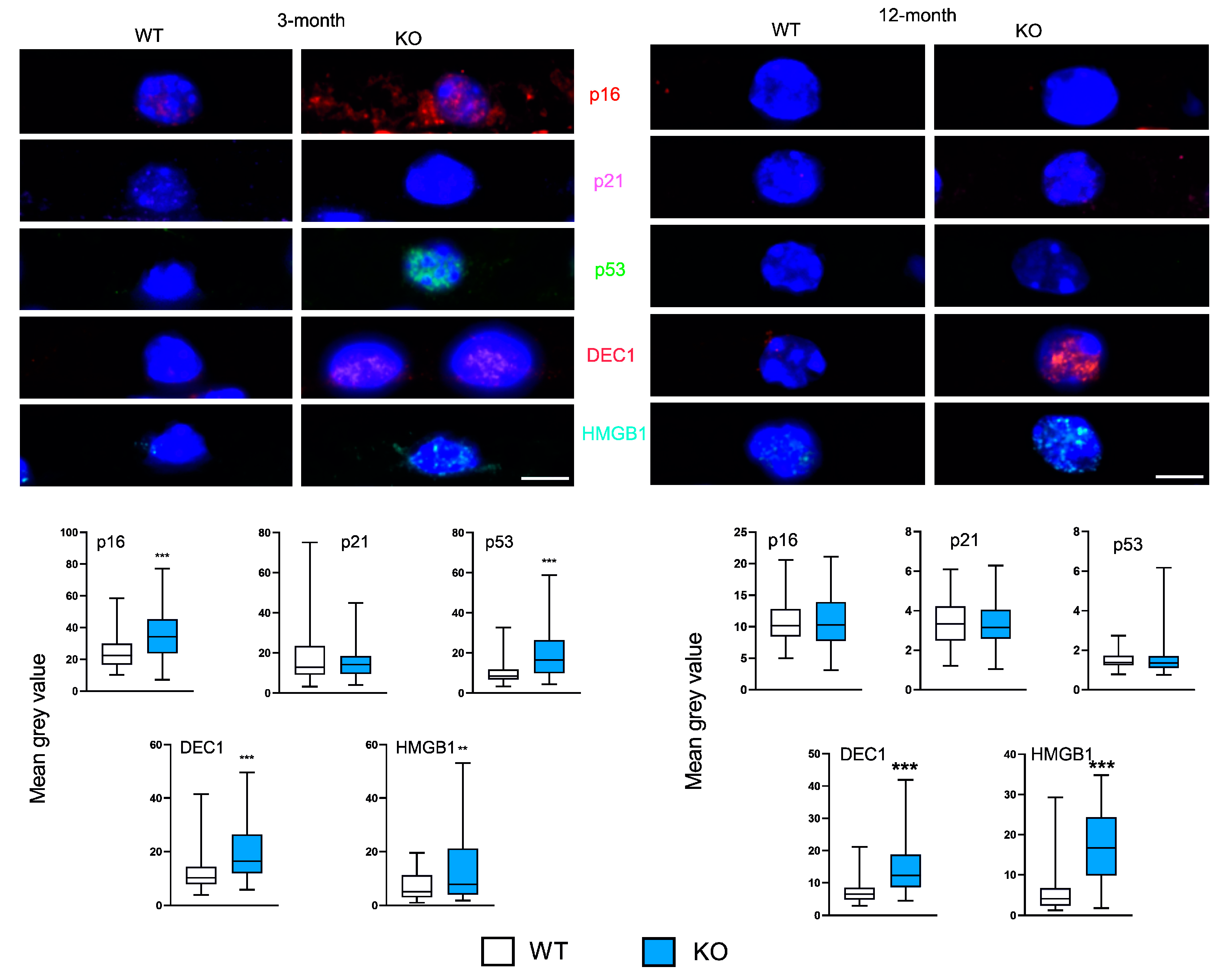
Figure 3.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of autophagy in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of autophagy: UBB (ubiquitin), p62 (sequestosome 1), BECN1 (BECLIN1), LAMP2 (lysosomal associated membrane protein 2), MAP1LC3B (microtubule associated protein 1 light chain 3 beta, LC3B), p62 (SQSTM1, sequestosome 1), PINK1 (PTEN induced kinase 1) and PRKN (parkin RBR E3 ubiquitin protein ligase) in RPE of wild-type (WT) and mice with the global knockout in the PPARGC1A gene encoding the PGC-1α protein (KO). Puncta specific to the label of the primary antibody to each marker have the same color as their abbreviations placed between image panels. DAPI was used to stain the nuclei of RPE cells (blue). Arrows in the PINK/PRKN image show clusters of colocalization of both markers. Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars present minimum and maximal values; n = 43-81; * – p < 0.05, ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
Figure 3.
Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) influences the expression of markers of autophagy in the retinal pigment epithelium (RPE) of young and old mice. Confocal microscopy images of the immunoreactivity of the markers of autophagy: UBB (ubiquitin), p62 (sequestosome 1), BECN1 (BECLIN1), LAMP2 (lysosomal associated membrane protein 2), MAP1LC3B (microtubule associated protein 1 light chain 3 beta, LC3B), p62 (SQSTM1, sequestosome 1), PINK1 (PTEN induced kinase 1) and PRKN (parkin RBR E3 ubiquitin protein ligase) in RPE of wild-type (WT) and mice with the global knockout in the PPARGC1A gene encoding the PGC-1α protein (KO). Puncta specific to the label of the primary antibody to each marker have the same color as their abbreviations placed between image panels. DAPI was used to stain the nuclei of RPE cells (blue). Arrows in the PINK/PRKN image show clusters of colocalization of both markers. Scale bar: 5 μm (upper panels). Mean grey value for the markers of antioxidant defense determined in WT and KO mice. Median ± lower and upper quartile, error bars present minimum and maximal values; n = 43-81; * – p < 0.05, ** – p < 0.01, *** – p < 0.001 as compared with WT animals (lower panels).
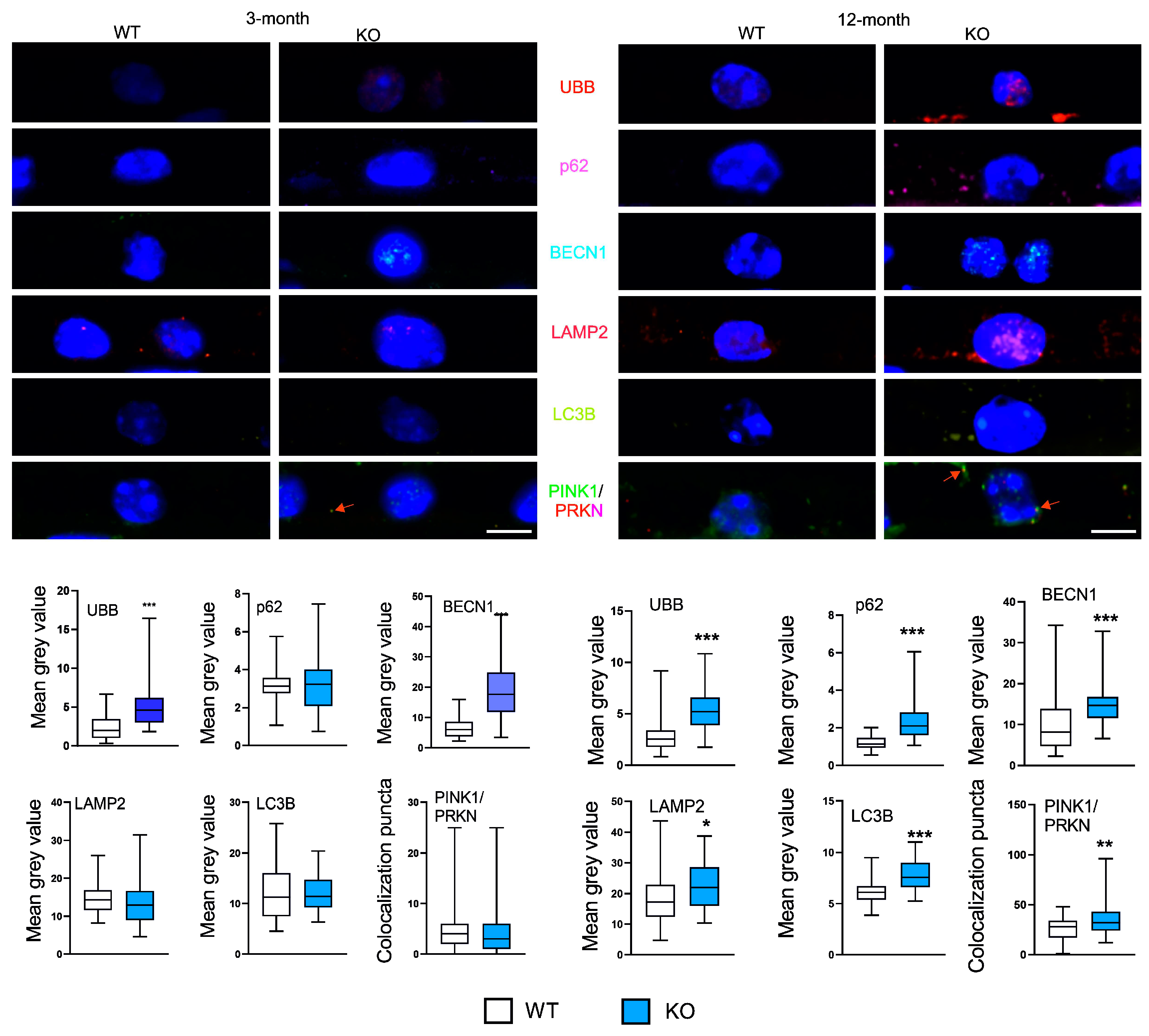
Figure 4.
Principal component analysis (PCA) of the expression of genes involved in antioxidant defense, senescence and autophagy in the aging retina of mice with knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding PGC-1α gene and wild-type animals. Each dot represents the expression of a single gene. PCA was performed for 3-month (A) and 12-month (B) mice, PGC-1α knockout (C) and wild-type (D) mice or data for all kinds of mice (E). PGC-1α is abbreviated to PGC1, its expression in WT animals is designated by 1 and its lack – by 0.
Figure 4.
Principal component analysis (PCA) of the expression of genes involved in antioxidant defense, senescence and autophagy in the aging retina of mice with knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding PGC-1α gene and wild-type animals. Each dot represents the expression of a single gene. PCA was performed for 3-month (A) and 12-month (B) mice, PGC-1α knockout (C) and wild-type (D) mice or data for all kinds of mice (E). PGC-1α is abbreviated to PGC1, its expression in WT animals is designated by 1 and its lack – by 0.
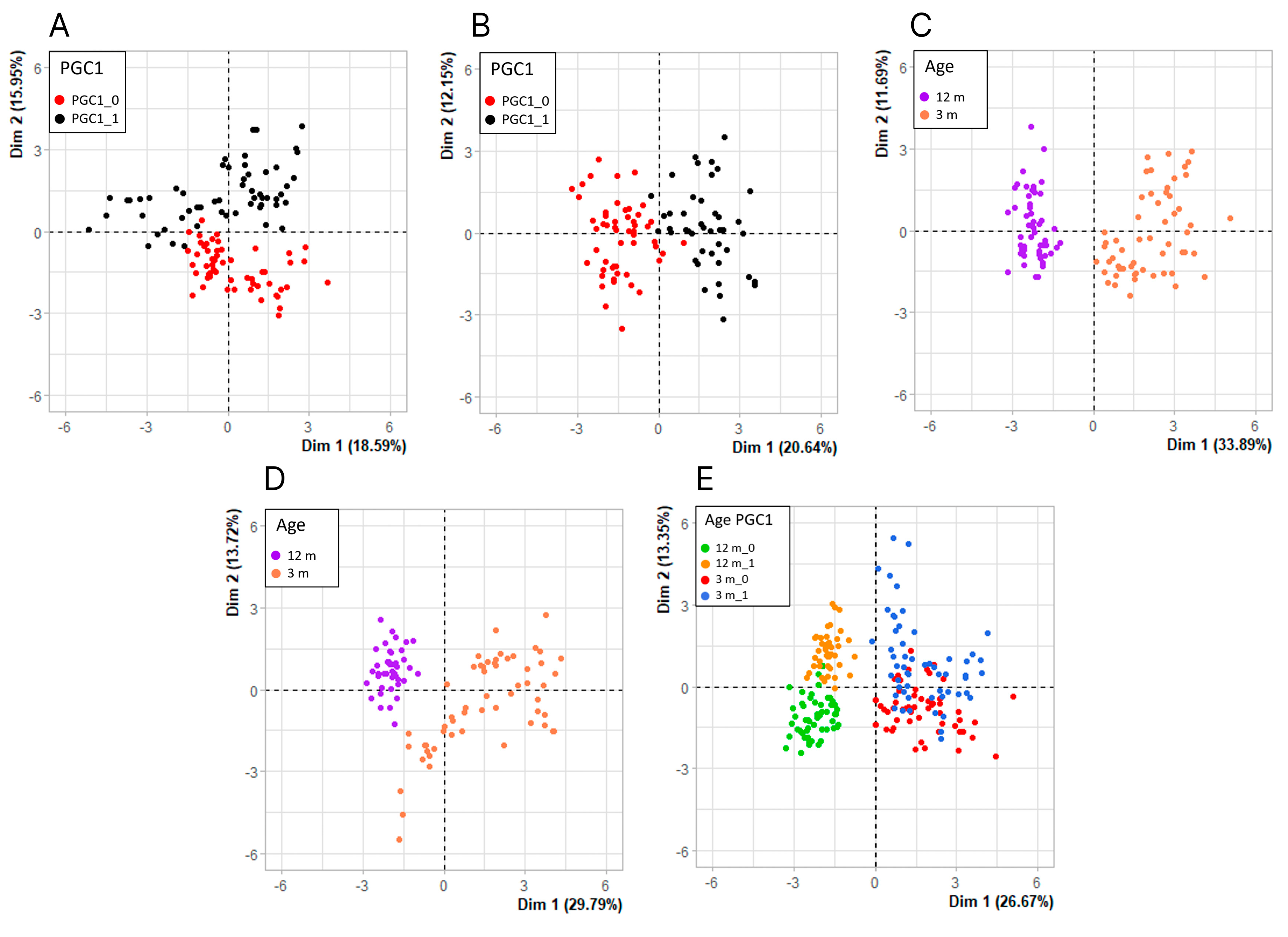
Figure 5.
The variable correlation plots of principal component analysis (PCA) on the first two component axes of the expression of genes involved in antioxidant defense, senescence and autophagy in the aging retina of mice with knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding PGC-1α gene and wild-type animals. The contribution of each gene expression level is represented by a color of a gradient scale (contrib). PCA was performed for 3-month (A) and 12-month (B) mice, PGC-1α knockout (C) and wild-type (D) mice or data for all kinds of mice (E).
Figure 5.
The variable correlation plots of principal component analysis (PCA) on the first two component axes of the expression of genes involved in antioxidant defense, senescence and autophagy in the aging retina of mice with knockout in the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding PGC-1α gene and wild-type animals. The contribution of each gene expression level is represented by a color of a gradient scale (contrib). PCA was performed for 3-month (A) and 12-month (B) mice, PGC-1α knockout (C) and wild-type (D) mice or data for all kinds of mice (E).
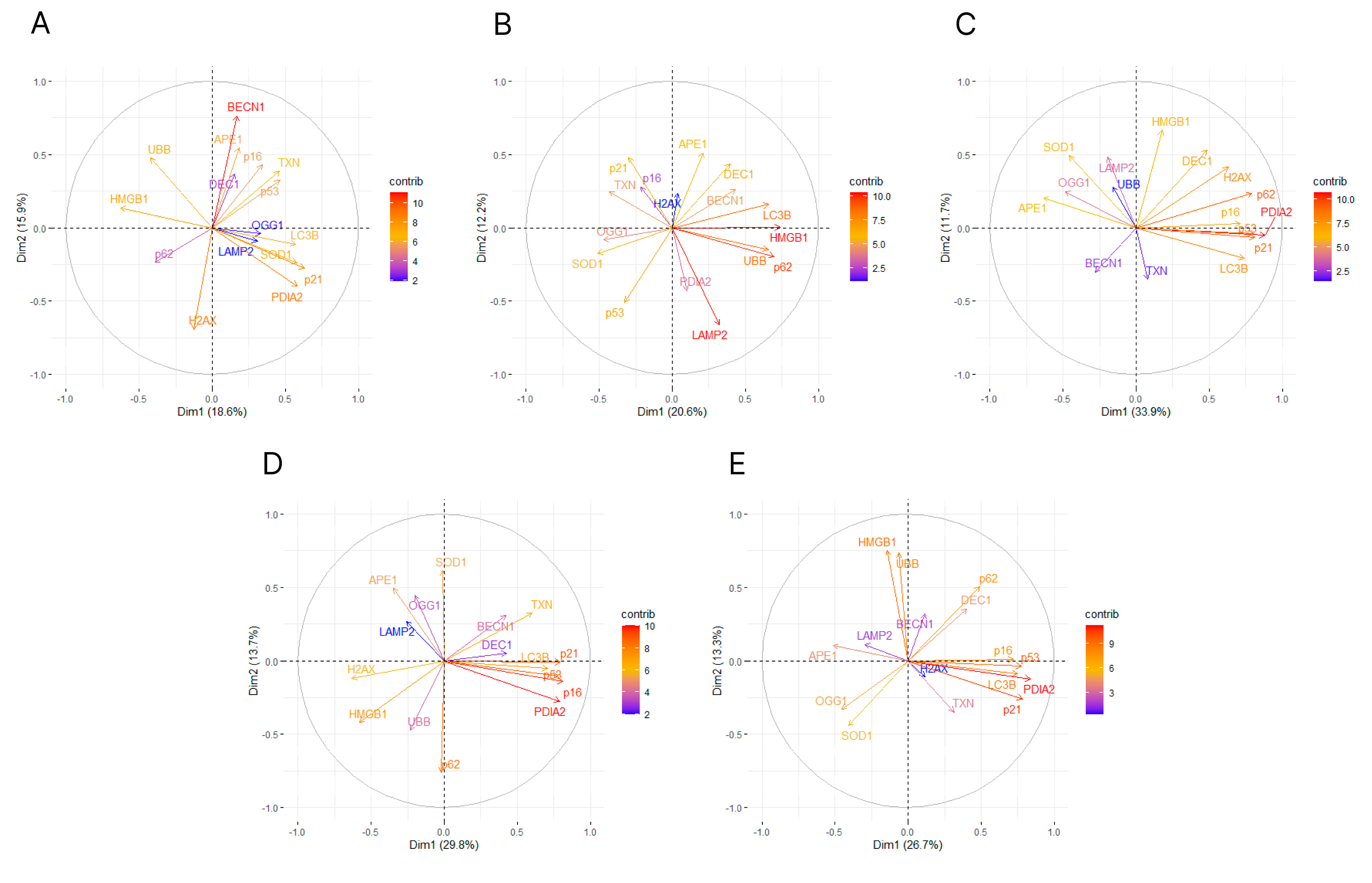
Figure 6.
Interaction network for the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding the PGC-1α protein in the mouse aging retina. The colors of the circles representing specific genes correspond to fold changes expressed by a logarithm of the ratio of the gene expression for 3-month and 12-month mice except PGC-1α represented by a yellow circle and abbreviated to PGC1. The network was built based on String Cytoscape. All abbreviations are defined in the main text.
Figure 6.
Interaction network for the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PPARGC1A) gene encoding the PGC-1α protein in the mouse aging retina. The colors of the circles representing specific genes correspond to fold changes expressed by a logarithm of the ratio of the gene expression for 3-month and 12-month mice except PGC-1α represented by a yellow circle and abbreviated to PGC1. The network was built based on String Cytoscape. All abbreviations are defined in the main text.
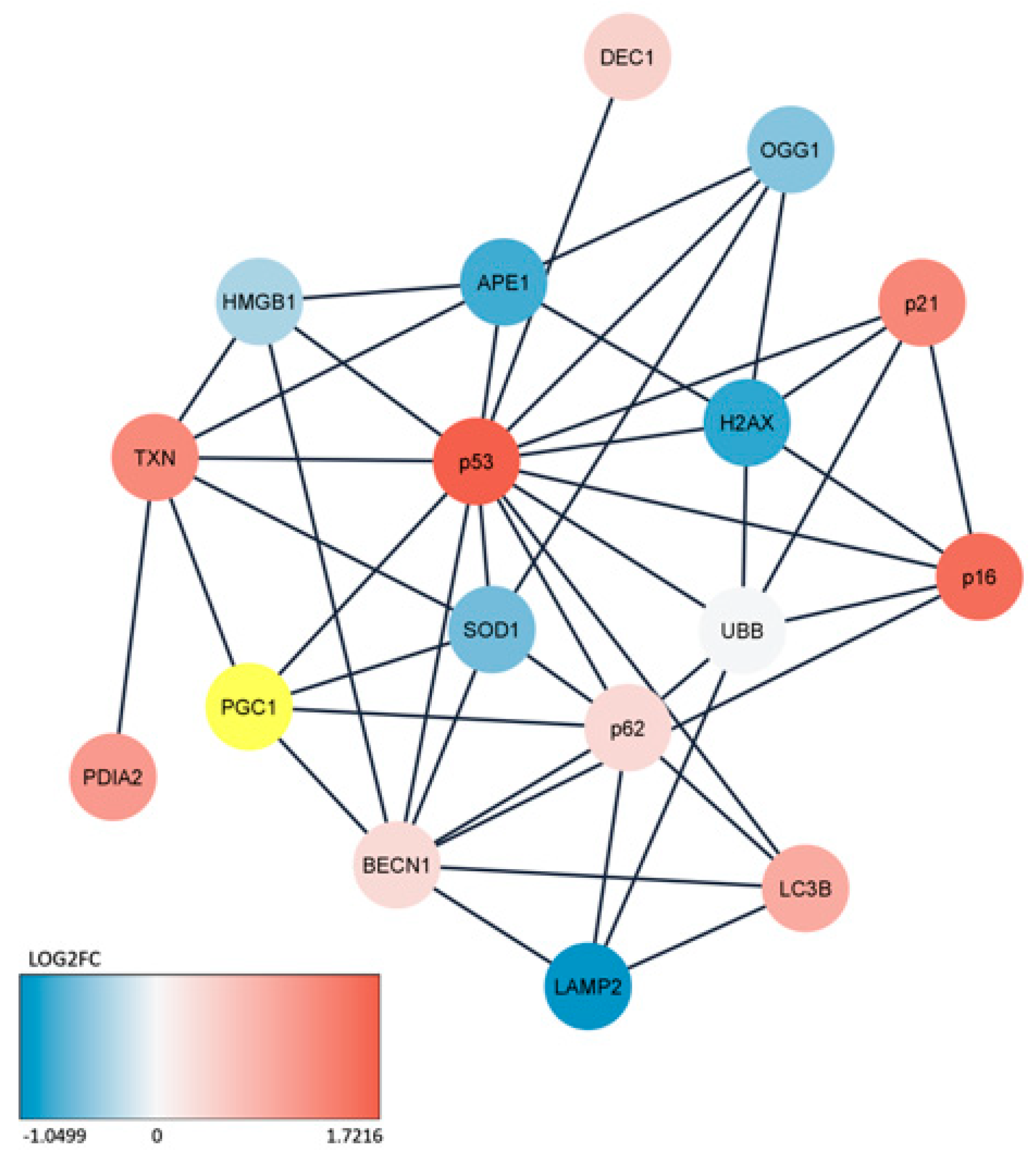

Table 1.
markers of antioxidant defense, senescence and autophagy most significantly contributing to the discrimination between gene expression in PGC-1α KO and WT animals.
Table 1.
markers of antioxidant defense, senescence and autophagy most significantly contributing to the discrimination between gene expression in PGC-1α KO and WT animals.
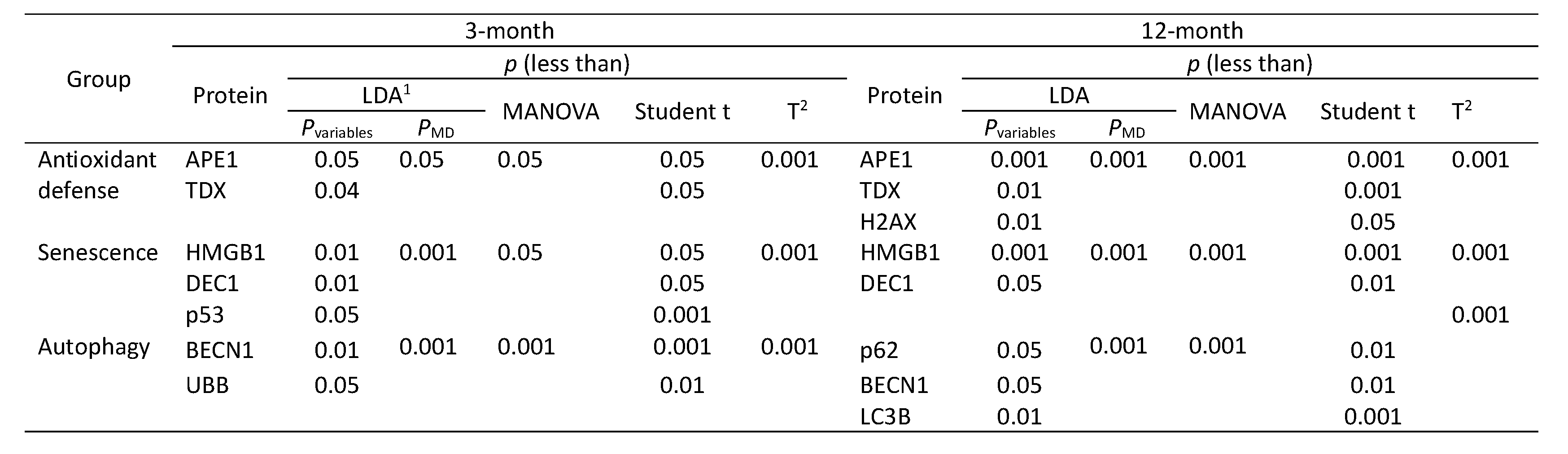 |
1 Abbreviations: LDA, linear discriminant analysis; Pvariables , significance of variables contributing to the best discrimination between KO and WT groups; PMD , significance of Mahalanobis’ distance; Student t, significance of differences between individual variables; MONOVA, refers to the Wilks’ significance of differences between the groups based on multivariate inference testing; T2, T Hotelling test, refers to the significance of multivariate comparison between two groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



