
Submitted:
19 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
The erythropoietin-producing hepatocellular carcinoma (Eph) receptors are the largest receptor tyrosine kinases family. EphB4 is essential for cell adhesion and motility during embryogenesis. Pathologically, EphB4 is overexpressed and contributes to poor prognosis in various tumors. Therefore, sensitive monoclonal antibodies (mAbs) should be developed to predict the prognosis for multiple tumors with high EphB4 expression, including breast and gastric cancers. This study aimed to develop highly sensitive and specific anti-EphB4 mAbs for several applications using the Cell-Based Immunization and Screening (CBIS) method. EphB4-overexpressed Chinese hamster ovary (CHO)-K1 (CHO/EphB4) cells were immunized into mice, and we established an anti-EphB4 mAb (clone B4Mab-7), which is applicable for flow cytometry, western blotting, and immunohistochemistry. B4Mab-7 reacted with endogenous EphB4-positive breast cancer cell lines, MCF-7 and MDA-MB-468, but did not react with EphB4-knockout MCF-7 (BINDS-52) in flow cytometry. Dissociation constant (KD) values were determined to be 2.9 × 10‑9 M, 1.3 × 10‑9 M, and 3.3 × 10‑9 M by flow cytometric analysis for CHO/EphB4, MCF-7, and MDA-MB-468 cells, respectively. B4Mab-7 detected the EphB4 protein bands from breast cancer cells in western blotting, and stained breast cancer tissues immunohistochemistry. Altogether, B4Mab-7 demonstrated high sensitivity and specificity against EphB4 in various applications.
Keywords:
EphB4
; monoclonal antibody
; Cell-Based Immunization and Screening
; immunohistochemistry
1. Introduction
Breast cancer is a malignant tumor with overwhelmingly high incidence in women. In 2020, there were estimated 2,261,419 new breast cancer cases (11.7% of all cancer cases) and estimated 684,996 deaths (6.9% of all cancer deaths) worldwide [1]. In most developed countries, breast cancer fatality rates have been declining in recent years, owing to improved treatment strategies and earlier detection techniques [2,3,4]. Breast cancer treatment currently includes surgery, radiation therapy, chemotherapy, and molecular targeted therapy [5,6,7].
Molecular targeted therapy depends on the subtype of breast cancer. Breast cancers are categorized into clinicopathological surrogate intrinsic subtypes defined by immunohistochemistry (IHC) testing of estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor 2 (HER2), and Ki-67 status [8,9,10]. Breast cancer subtypes can be largely classified into hormone receptor (ER or PR)-positive luminal A, luminal B, HER2-positive, and triple-negative breast cancer (TNBC) [8,9,10].
The erythropoietin-producing hepatocellular carcinoma (Eph) receptors belong to the largest family of receptor tyrosine kinases in mammals [11]. Eph receptors are physiologically associated with angiogenesis, cell migration, vascularization, axon guidance, and neurogenesis [12]. The Eph family was first discovered in human cancer cell lines in 1987 [13], and is now known to have two classes of receptors consisting of nine EphA members and five EphB members in humans [14]. The extracellular region of the Eph receptor consists of a ligand binding domain, followed by a cysteine-rich domain (including the sushi and epidermal growth factor-like domains) and two fibronectin domains [15,16]. The intracellular side of the Eph receptor consists of a transmembrane region, a tyrosine kinase domain, a sterile alpha motif, and a PDZ domain. The EphA receptor binds promiscuously to ephrin A ligands (five types) and the EphB receptor binds promiscuously to ephrin B ligands (three types), but there may be cross-talk between the groups [17,18,19,20,21,22,23]. In contrast to other Eph receptors, EphB4 shows clear specificity for the ligand ephrinB2 and very weak binding affinity for ephrinB1 and ephrinB3 [16].
EphB4 and ephrinB2 are biologically essential for cell adhesion, cell migration, stabilization, branching, and remodeling during embryogenesis for cardiovascular formation [24,25,26,27,28,29,30]. Mice lacking either EphB4 (expressed in veins) or ephrinB2 (expressed in arteries) showed identical defects in the formation of capillary connections between the network of arteries and veins in the head and yolk sac, resulting in embryonic lethal [29,30,31,32]. EphB4 and ephrinB2 create a repulsive effect between them and act to prevent the fusion of the veins with the arteries in adults. Phosphorylation of ephrinB2 triggers angiogenesis, and downstream inhibition of phosphorylation-dependent or PDZ domain-dependent signaling of the ephrinB2 prevents endothelial cell engraftment and prevents the appropriate association of endothelial cells with other endothelial cells and with the pericytes [25].
EphB4 is pathologically overexpressed in various malignant tumors, including breast cancer [33,34,35], lung cancer [36,37,38], gastric cancer [39], colorectal cancer [40,41], acute myeloid leukemia [42], cervical cancer [43], glioma [44], ovarian cancer [45,46], prostate cancer [47,48], thyroid cancer [49,50], bladder cancer [51,52], and contributes to poor prognosis. Mutations in the phosphorylation site of the EphB4 intracellular region in lung cancer promote tumor growth in vitro [53]. EphB4 gene amplification has been found and correlated with higher tumor stage in various tumors [34,48,54]. Therefore, mutations, gene amplification, and overexpression of EphB4 contribute to malignant progression. EphB4 is involved in cell proliferation, migration, differentiation, and angiogenesis in various cancers [55]. Several data have been reported on the forward and reverse signaling of EphB4 and ephrinB2 in tumors. EphB4 extracellular domain can induce angiogenic responses by stimulating ephrinB2 reverse signaling in cultured endothelial cells and promote tumor growth [56]. On the other hand, ephrinB2-independent effects of EphB4 have also been reported in tumors [42,57]. Several reports have shown that high expression of EphB4 in tumors causes activation of ligand-independent downstream signaling, and the effect of EphB4 on cancer malignancy in the absence of ephrinB2 has been examined [42,58]. EphrinB2 expression in breast and lung cancer cell lines with high expression of EphB4 is very low compared to normal cell lines [37,59]. Therefore, EphB4 may often contribute to tumor malignancy by ligand-independent effects in cancer patients.
In breast cancer in particular, EphB4 is expressed endogenously in numerous cell lines [33,34], and the EphB4-positive rate in immunohistochemical staining is high [60]. In breast cancer cell lines, the knockdown of EphB4 by small interfering RNA results in reduced cell viability, activation of caspase-8, and induction of apoptosis [34]. Furthermore, EphB4 knockdown in a mouse breast cancer xenograft model resulted in significantly smaller tumors, induced apoptosis, and reduced tumor vascular distribution [34]. Ding et al. reported that overexpression of EphB4 caused gain-of-function activity in HER2-positive breast cancer cell lines and resistance to lapatinib, a dual HER2/epidermal growth factor receptor inhibitor [61]. Comparison of EphB4 mRNA expression levels in four different clinicopathological surrogate intrinsic subtypes of breast cancer revealed that EphB4 transcription is significantly higher in HER2-positive breast cancer and TNBC than luminal breast cancer [62].
Highly sensitive and specific monoclonal antibodies (mAbs) are needed to facilitate and ensure the diagnosis of breast cancer subtype and to determine treatment strategies. EphB4 may be a useful diagnostic target for prognostication and therapeutic targets of breast cancers, requiring the development of sensitive and specific mAb. In this study, we report the establishment of sensitive and specific mAbs against EphB4 for flow cytometric, western blot, and immunohistochemical analyses.
2. Materials and methods
2.1. Plasmid preparation
Synthesized DNA encoding Human EphB4 (Accession No. NM_004444; catalog No: RC208559) was purchased from OriGene Technologies, Inc. (Rockville, MD, USA). The signal sequence was excluded and the EphB4 open reading frame with an N-terminal PA16 tag or an N-terminal MAP tag was subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) using an In-Fusion HD Cloning Kit (Takara Bio, Inc. Shiga, Japan); the recombinant expression vectors were named as pCAG/PA16-EphB4 and pCAG/MAP-EphB4 DNA with an N-terminal PA16 and with an N-terminal MAP tag, respectively. The amino acid sequences of each protein tag are as follows: PA16 tag, 16 amino acids (GLEGGVAMPGAEDDVV) and MAP tag [63], 12 amino acids (GDGMVPPGIEDK) [64]. The clone NZ-1 (rat IgG2a, lambda) was used for PA16 tag [65], and clone PMab-1 (rat IgG2a, kappa) was used for MAP tag [64].
2.2. Cell lines
Chinese hamster ovary (CHO)-K1, P3X63Ag8U.1 (P3U1), and MDA-MB-468 were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). MCF-7 was obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging and Cancer, Tohoku University (Miyagi, Japan). CHO-K1 cells that overexpress PA16-EphB4 (CHO/EphB4) and MAP-EphB4 (CHO/MAP-EphB4) were generated by transfection of pCAG/PA16-EphB4 and pCAG/MAP-EphB4 to CHO-K1 cells, respectively, using Lipofectamine LTX Reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA) by a Neon Transfection System (Thermo Fisher Scientific, Inc.). Stable transfectants were sorted using a cell sorter (SH800; Sony Corp., Tokyo, Japan) and cultivated in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA, USA). The MCF-7/EphB4-knockout (KO) cell line (BINDS-52) was generated by transfection using CRISPR/Cas9 plasmids (Assay ID: CRISPR710315_SGM) that target EphB4 (Thermo Fisher Scientific, Inc.). Plasmids were transfected, using Lipofectamine LTX Reagent (Thermo Fisher Scientific, Inc.) by a Neon Transfection System (Thermo Fisher Scientific, Inc.). Stable transfectants were sorted using a cell sorter (SH800). CHO-K1, P3U1, CHO/EphB4, CHO/MAP-EphB4, MCF-7, and BINDS-52 cells were cultured in a Roswell Park Memorial Institute (RPMI)-1640 medium (Nacalai Tesque, Inc., Kyoto, Japan). MDA-MB-468 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Nacalai Tesque, Inc.). All media were supplemented with 10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific Inc.), 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B (Nacalai Tesque, Inc.). All cells were cultured in a humidified incubator at 37°C, 5% CO2, and 95% air.
2.3. Hybridoma production
Female BALB/c mice (five weeks old) were purchased from CLEA Japan (Tokyo, Japan). The animals were housed under specific pathogen-free conditions. All animal experiments were conducted by relevant guidelines and regulations to minimize the pain and suffering of animals in the laboratory. Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001). Mice were monitored daily for health during the full five-week duration of the experiment. A reduction of more than 25% of total body weight was defined as a humane endpoint. For splenectomy, mice were euthanized by cervical dislocation and death was confirmed by respiratory arrest and cardiac arrest.
To develop mAbs against EphB4, we used CBIS method [66,67,68,69,70,71,72,73,74,75]. Briefly, two BALB/c mice were injected intraperitoneally with a mixture of CHO/MAP-EphB4 cells (1 × 108/mouse) with Imject Alum (Thermo Fisher Scientific, Inc.) as adjuvant only at the first immunization; the second immunization was performed one week after the first, and the third to fifth immunizations were performed at the same interval. Two days after the fifth immunization as boost immunization, mice were sacrificed and splenocytes were collected. The harvested spleen cells were subsequently fused with P3U1 cells using Polyethylene Glycol (PEG) 1500 (Roche Diagnostics, Indianapolis, IN, USA), and the hybridomas were grown in an RPMI-1640 medium supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific Inc.) on ten 96 well plates. Culture supernatants were screened by flow cytometry as described below.
2.4. Flow cytometry
Cultured cells were collected after brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (EDTA, Nacalai Tesque, Inc.). They were once washed with 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) and treated with primary mAbs for 30 min at 4°C. Fluorescein isothiocyanate conjugated anti-EphB4 mAb (clone: 04) was purchased from Abcam (Cambridge, MA, USA). After washing cells twice with 0.1% BSA in PBS, the cells were then treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:1000; Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 min at 4°C. After three washes, cells were diluted with 100 μL and shaken before measurement. Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp.). The collected data were analyzed with FlowJo software (FlowJo, OR, USA). 0.1% BSA in PBS was used to dilute the antibodies.
2.5. Determination of dissociation constant (KD) by flow cytometry
CHO/EphB4, MCF-7, and MDA-MB-468 cells were suspended in 100 μL of B4Mab-7 (0.006-50 μg/mL) serially diluted 1/2-fold, and exposed for 30 min, and the cells were washed twice with 0.1% BSA in PBS. Then, 50 μL of Alexa Fluor 488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology, Inc.) was added and exposed for 30 minutes. After washing twice, the cells were transferred to a 96-well plate for measurement. Fluorescence data were collected using the SA3800 Cell Analyzer. The collected data were analyzed with FlowJo software. The dissociation constant (KD) was calculated by fitting binding isotherms to built-in, one-site binding models in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA, USA). 0.1% BSA in PBS was used to dilute the antibodies.
2.6. Western blot analysis
Cell lysates were extracted from cultured cells using 1% Triton X-100, and cell remnants were removed by centrifugation. The cell lysates (10 μg) were boiled in sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc.). These proteins were then electrophoresed on 5%–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corporation) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA, Darmstadt, Germany). After blocking with 4% skim milk (Nacalai Tesque, Inc.) for 1 h, membranes were incubated with 1 μg/mL of B4Mab-7 or 1 μg/mL of an anti-isocitrate dehydrogenase 1 (IDH1) mAb, RcMab-1 [76], for 30 min. Then, the membranes were incubated with peroxidase-conjugated anti-mouse immunoglobulins (diluted 1:2000; Agilent Technologies, Inc., Santa Clara, CA, USA), or anti-rat immunoglobulins (diluted 1:10,000; Sigma-Aldrich Corp., St. Louis, MO, USA) for 30 min, respectively. Finally, the protein bands were visualized using ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) and Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan). All Western blot procedures were performed at room temperature, and all primary and secondary antibodies were diluted with 4% skim milk.
2.7. Immunohistochemical analysis
A formalin-fixed paraffin-embedded (FFPE) breast cancer tissue microarray (Cat. No.: T8235721-5, Lot. No.: B904111) containing 63 cases was purchased from BioChain Institute Inc. (Newark, CA, USA). The antigen retrieval was performed using a direct autoclave treatment at 121°C for 20 min in EnVision FLEX Target Retrieval Solution High pH (Agilent Technologies, Inc.) Peroxidase blocking was performed by treatment with 3.0% H2O2 for 10 min. Blocking was then conducted using SuperBlock Blocking Buffer (Thermo Fisher Scientific, Inc.). Sections were then incubated with 10 μg/mL of B4Mab-7 (diluted by SuperBlock Blocking Buffer) or buffer control (SuperBlock Blocking Buffer) for 1 h at room temperature, followed by treatment with an Envision+ kit (Agilent Technologies, Inc.) for 30 min. Antigen–antibody complexes were visualized using 3,3′-diaminobenzidine tetrahydrochloride solution (Agilent Technologies, Inc.) for 2 min, and sections were then counterstained with hematoxylin (FUJIFILM Wako Pure Chemical Corporation) for 5 min. The intensity of staining was evaluated as 0, 1+, 2+, or 3+. Hematoxylin and eosin (H&E) staining (FUJIFILM Wako Pure Chemical Corporation) was performed using consecutive tissue sections at room temperature for 5 and 10 min, respectively.
3. Results
3.1. Establishment of anti-EphB4 mAbs
To establish anti-EphB4 mAbs, we employed the Cell-Based Immunization and Screening (CBIS) method, using stable transfectants for both immunizations and flow cytometric screening (Figure 1). Two BALB/c mice were immunized with CHO/MAP-EphB4 cells. Hybridomas were then seeded onto 96 well plates, and culture supernatants positive for CHO/EphB4 and negative for CHO-K1 were selected by flow cytometry. The positive count was 54/956 wells (5.65%), which were cloned by limiting dilution. After several additional screenings, an anti-EphB4 mAb, B4Mab-7 (IgG1, kappa), was finally established. Subsequently, the specificity and usefulness of B4Mab-7 were evaluated by flow cytometry, western blotting, and immunohistochemical analyses.
3.2. Flow cytometric analyses
We next examined the B4Mab-7 reactivity against breast cancer cell lines in flow cytometry. B4Mab-7 reacted with CHO/EphB4 cells in a dose-dependent manner (Figure 2A), whereas it did not react with parental CHO-K1 cells (Figure 2B). B4Mab-7 also showed a concentration-dependent reactivity to breast cancer cell lines, MCF-7 and MDA-MB-468 (Figure 2C,D). These results indicated that B4Mab-7 recognizes not only exogenous EphB4 but also endogenous EphB4 in breast cancer cell lines.
To confirm the specificity of B4Mab-7 against MCF-7, we established EphB4-knockout MCF-7 (BINDS-52) by a CRISPR/Cas9 system, and performed flow cytometry using B4Mab-7 against BINDS-52. B4Mab-7 showed no reaction against BINDS-52 cells (Figure 2E). We also confirmed that EphB4 was completely knocked out in BINDS-52 cells by commercially available mAb (clone 04). (Supplementary Figure 1) These results suggest that B4Mab-7 is specific for EphB4 of MCF-7.
3.3. Determination of the binding affinity of B4Mab-7
To investigate the binding ability of B4Mab-7, the apparent dissociation constant (KD) of B4Mab-7 for CHO/EphB4, MCF-7, and MDA-M468 was evaluated using flow cytometry. The KD values of B4Mab-7 for CHO/EphB4, MCF-7, and MDA-MB-468 were 2.9 × 10-9 M, 1.3 × 10-9 M, and 3.3 × 10-9 M (Figure 3), respectively, indicating that B4Mab-7 possesses a high binding affinity for CHO/EphB4, MCF-7, and MDA-M468 cells.
3.4. Western blot analysis
Then, we examined whether B4Mab-7 is also useful in western blot analysis. B4Mab-7 detected strong signals of 100~130-kDa in CHO/EphB4, MCF-7, and MDA-MB-468 cells, but not in CHO-K1 cells (Figure 4A). These results suggest that B4Mab-7 specifically and sensitively recognizes endogenous EphB4 in breast cancer cell lines on western blot analysis.
3.5. Immunohistochemical analysis against breast cancers
We next investigated whether B4Mab-7 applies to immunohistochemical analysis against formalin-fixed paraffin-embedded (FFPE) breast cancer sections [33,34,35]. As depicted in Figure 5A, D, G, J, M, P, B4Mab-7 strongly stained the plasma membrane of breast cancer cells, and weak staining was also observed in the cytoplasm. B4Mab-7 stained the membranes of cancer cells in 41 of 63 (65.1%) breast cancers (Figure 5A,D: score 3+, Figure 5G,J: score 2+, Figure 5M, P: score 0; Scores were divided by the intensity of membrane staining). Treatment with control buffer (Figure 5B, E, H, K, N, Q) and hematoxylin and eosin (H&E) staining (Figure 5C,F,I,L,O,R) were performed on consecutive breast cancer tissues.
B4Mab-7 also stained venous or lymphatic endothelial cells (Supplementary Figure 2), although it did not stain stromal tissues. These results indicate that B4Mab-7 is useful for immunohistochemical analysis of FFPE tumor sections. All staining results of B4Mab-7 for breast cancers are shown in Table 1. B4Mab-7 could stain invasive ductal carcinomas, invasive lobular carcinoma, adenocarcinoma, fibroadenoma, and medullary carcinoma (Table 2).
4. Discussion
In this study, we successfully developed a highly sensitive and specific anti-EphB4 mAb, B4Mab-7, which can be adapted to any experimental method using the CBIS method (Figure 1). The CBIS method does not need purified proteins for immunization and screening, and could target the three-dimensional structure of transmembrane proteins.
Many anti-EphB4 mAbs are now commercially available as shown in Supplementary Table 1. As listed, anti-EphB4 mAbs are usually applicable for only one or two experimental techniques. Therefore, B4Mab-7 is the first anti-EphB4 mAb, which is useful for flow cytometry, western blotting, and immunohistochemistry. B4Mab-7 was developed using the CBIS method, whereas almost all mAbs were developed using immunization of recombinant proteins or peptides; therefore, the CBIS method using EphB4-overexpressed cell lines, as in this study, might be the best way to obtain useful anti-EphB4 mAbs. The CBIS method could be advantageous for the development of mAbs to the other Eph receptors.
Although many mAbs against anti-Eph receptors, such as anti-EphA2 mAbs [77,78,79,80,81,82,83,84,85,86,87,88,89,90,91], anti-EphA3 mAbs [92,93,94,95,96,97,98,99], anti-EphA10 mAbs [100], anti-EphB2 mAbs [101], anti-EphB4 mAbs [102,103] have been developed, almost all mAbs have not been used clinically [104], excluding only a few anti-Eph receptor mAbs [89,90,95]. 1C1 was a promising anti-EphA2 mAb that showed agonist activity and caused phosphorylation and internalization of EphA2 [87]. Since 1C1 itself showed no antitumor effect, MEDI-547, an antibody-drug conjugate (ADC) composed of the cytotoxic drug auristatin (toxin) linked to 1C1, was created [87]. MEDI-547 inhibited tumor growth and metastasis in vivo. Although MEDI-547 was moved to phase I clinical study, it was terminated due to serious side effects such as treatment-related bleeding, coagulation events, and elevated liver enzymes at the starting dose in 3/6 patients [90]. IIIA4/KB004/Ifabotuzumab is an anti-EphA3 mAb currently in development for the treatment of glioblastoma multiforme [99]. The phase I study of IIIA4/KB004/Ifabotuzumab has been completed and the safety has been confirmed [105]. Interestingly, the immunogen for IIIA4 uses LK63 human pre-B acute lymphoblastic leukemia cells [106], and it is the only anti-EphA3 mAb that is adaptable to flow cytometry, western blotting, and immunohistochemistry like B4Mab-7 [106]. This fact supports the suitability of immunization using cell lines for the development of useful anti-Eph receptor mAbs.
Because Eph receptors are expressed in normal tissues and have important physiological functions, targeting them with conventional antibody drugs has been difficult. Therefore, recently, naked antibodies have been applied to chimeric antigen receptor-T cell therapy [107,108], ADC [87,88,99,109], bispecific T-cell engager [100,110], bispecific antibody [111,112], and liposomes [113], to target Eph receptors. Bispecific antibodies against EphA2/EphA3 reduced the clonogenicity of recurrent glioblastoma in vitro and the tumorigenic potential of xenograft recurrent glioblastoma in vivo [112].
In conclusion, we established an anti-EphB4 mAb, B4Mab-7, which is applicable for flow cytometry, western blotting, and immunohistochemistry. In the future study, the heavy chain subclass of B4Mab-7 should be modified from IgG1 to IgG2a to enhance effector function (antibody-dependent cellular cytotoxicity and complement-dependent cytotoxicity) and its antitumor effect on breast cancer in vitro and in vivo should be investigated.
Supplementary Materials
Supplementary Figure S1, Flow cytometry using anti-EphB4 mAbs (clone: 04). Supplementary Figure S2, Immunohistochemical analysis by B4Mab-7 for endothelial cells. Supplementary Table S1, Commercially available anti-EphB4 mAbs.
Author Contributions
R.N. performed the experiments. M.K.K. and Y.K. designed the experiments. R.N., H.S. and Y.K. analyzed the data. R.N., H.S. and Y.K. wrote the manuscript. All authors read and approved the final manuscript and agreed to be accountable for all aspects of the research in ensuring that the accuracy or integrity of any part of the work is appropriately investigated and resolved.
Funding
This research was supported by Grant-in-Aid for the Japan Society for the Promotion of Science (JSPS) Fellows grant no. 22J20977 (to R.N.), JST SPRING, Grant Number JPMJSP2114 (R.N.), and also supported by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP23ama121008 (Y.K.) and JP23am0401013 (Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Allemani, C.; Weir, H.K.; Carreira, H.; Harewood, R.; Spika, D.; Wang, X.S.; Bannon, F.; Ahn, J.V.; Johnson, C.J.; Bonaventure, A.; et al. Global surveillance of cancer survival 1995-2009: analysis of individual data for 25,676,887 patients from 279 population-based registries in 67 countries (CONCORD-2). Lancet 2015, 385, 977–1010. [Google Scholar] [CrossRef] [PubMed]
- Massat, N.J.; Dibden, A.; Parmar, D.; Cuzick, J.; Sasieni, P.D.; Duffy, S.W. Impact of Screening on Breast Cancer Mortality: The UK Program 20 Years On. Cancer Epidemiol Biomarkers Prev 2016, 25, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Bleyer, A.; Welch, H.G. Effect of three decades of screening mammography on breast-cancer incidence. N Engl J Med 2012, 367, 1998–2005. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- McDonald, E.S.; Clark, A.S.; Tchou, J.; Zhang, P.; Freedman, G.M. Clinical Diagnosis and Management of Breast Cancer. J Nucl Med 2016, 57 Suppl 1, 9S–16S. [Google Scholar] [CrossRef]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm Nanotechnol 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thurlimann, B.; Senn, H.J.; Panel, m. Personalizing the treatment of women with early breast cancer: highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann Oncol 2013, 24, 2206–2223. [Google Scholar] [CrossRef]
- Prat, A.; Pineda, E.; Adamo, B.; Galvan, P.; Fernandez, A.; Gaba, L.; Diez, M.; Viladot, M.; Arance, A.; Munoz, M. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast 2015, 24 Suppl 2, S26–35. [Google Scholar] [CrossRef]
- Mohammed, A.A. The clinical behavior of different molecular subtypes of breast cancer. Cancer Treat Res Commun 2021, 29, 100469. [Google Scholar] [CrossRef]
- Tuzi, N.L.; Gullick, W.J. eph, the largest known family of putative growth factor receptors. Br J Cancer 1994, 69, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Lisabeth, E.M.; Falivelli, G.; Pasquale, E.B. Eph receptor signaling and ephrins. Cold Spring Harb Perspect Biol 2013, 5. [Google Scholar] [CrossRef]
- Hirai, H.; Maru, Y.; Hagiwara, K.; Nishida, J.; Takaku, F. A novel putative tyrosine kinase receptor encoded by the eph gene. Science 1987, 238, 1717–1720. [Google Scholar] [CrossRef] [PubMed]
- Darling, T.K.; Lamb, T.J. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front Immunol 2019, 10, 1473. [Google Scholar] [CrossRef] [PubMed]
- Himanen, J.P.; Rajashankar, K.R.; Lackmann, M.; Cowan, C.A.; Henkemeyer, M.; Nikolov, D.B. Crystal structure of an Eph receptor-ephrin complex. Nature 2001, 414, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Kania, A.; Klein, R. Mechanisms of ephrin-Eph signalling in development, physiology and disease. Nat Rev Mol Cell Biol 2016, 17, 240–256. [Google Scholar] [CrossRef]
- Pasquale, E.B. Eph-ephrin promiscuity is now crystal clear. Nat Neurosci 2004, 7, 417–418. [Google Scholar] [CrossRef]
- Himanen, J.P.; Chumley, M.J.; Lackmann, M.; Li, C.; Barton, W.A.; Jeffrey, P.D.; Vearing, C.; Geleick, D.; Feldheim, D.A.; Boyd, A.W.; et al. Repelling class discrimination: ephrin-A5 binds to and activates EphB2 receptor signaling. Nat Neurosci 2004, 7, 501–509. [Google Scholar] [CrossRef]
- Forse, G.J.; Uson, M.L.; Nasertorabi, F.; Kolatkar, A.; Lamberto, I.; Pasquale, E.B.; Kuhn, P. Distinctive Structure of the EphA3/Ephrin-A5 Complex Reveals a Dual Mode of Eph Receptor Interaction for Ephrin-A5. PLoS One 2015, 10, e0127081. [Google Scholar] [CrossRef]
- North, H.A.; Zhao, X.; Kolk, S.M.; Clifford, M.A.; Ziskind, D.M.; Donoghue, M.J. Promotion of proliferation in the developing cerebral cortex by EphA4 forward signaling. Development 2009, 136, 2467–2476. [Google Scholar] [CrossRef]
- Murai, K.K.; Pasquale, E.B. 'Eph'ective signaling: forward, reverse and crosstalk. J Cell Sci 2003, 116, 2823–2832. [Google Scholar] [CrossRef]
- Depaepe, V.; Suarez-Gonzalez, N.; Dufour, A.; Passante, L.; Gorski, J.A.; Jones, K.R.; Ledent, C.; Vanderhaeghen, P. Ephrin signalling controls brain size by regulating apoptosis of neural progenitors. Nature 2005, 435, 1244–1250. [Google Scholar] [CrossRef]
- Mignon, L.; Vourc'h, P.; Romero-Ramos, M.; Osztermann, P.; Young, H.E.; Lucas, P.A.; Chesselet, M.F. Transplantation of multipotent cells extracted from adult skeletal muscles into the subventricular zone of adult rats. J Comp Neurol 2005, 491, 96–108. [Google Scholar] [CrossRef]
- Jensen, P.L. Eph receptors and ephrins. Stem Cells 2000, 18, 63–64. [Google Scholar] [CrossRef]
- Salvucci, O.; Tosato, G. Essential roles of EphB receptors and EphrinB ligands in endothelial cell function and angiogenesis. Adv Cancer Res 2012, 114, 21–57. [Google Scholar] [CrossRef]
- Cheng, N.; Brantley, D.M.; Chen, J. The ephrins and Eph receptors in angiogenesis. Cytokine Growth Factor Rev 2002, 13, 75–85. [Google Scholar] [CrossRef]
- Makinen, T.; Adams, R.H.; Bailey, J.; Lu, Q.; Ziemiecki, A.; Alitalo, K.; Klein, R.; Wilkinson, G.A. PDZ interaction site in ephrinB2 is required for the remodeling of lymphatic vasculature. Genes Dev 2005, 19, 397–410. [Google Scholar] [CrossRef]
- Zhang, G.; Brady, J.; Liang, W.C.; Wu, Y.; Henkemeyer, M.; Yan, M. EphB4 forward signalling regulates lymphatic valve development. Nat Commun 2015, 6, 6625. [Google Scholar] [CrossRef]
- Gerety, S.S.; Wang, H.U.; Chen, Z.F.; Anderson, D.J. Symmetrical mutant phenotypes of the receptor EphB4 and its specific transmembrane ligand ephrin-B2 in cardiovascular development. Mol Cell 1999, 4, 403–414. [Google Scholar] [CrossRef]
- Adams, R.H.; Wilkinson, G.A.; Weiss, C.; Diella, F.; Gale, N.W.; Deutsch, U.; Risau, W.; Klein, R. Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Genes Dev 1999, 13, 295–306. [Google Scholar] [CrossRef]
- Wang, H.U.; Chen, Z.F.; Anderson, D.J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef]
- Wang, Z.; Cohen, K.; Shao, Y.; Mole, P.; Dombkowski, D.; Scadden, D.T. Ephrin receptor, EphB4, regulates ES cell differentiation of primitive mammalian hemangioblasts, blood, cardiomyocytes, and blood vessels. Blood 2004, 103, 100–109. [Google Scholar] [CrossRef]
- de Muijnck, C.; van Gorkom, Y.; van Duijvenvoorde, M.; Eghtesadi, M.; Dekker-Ensink, G.; Bhairosingh, S.S.; Affinito, A.; Kuppen, P.J.K.; Vahrmeijer, A.L.; Sier, C.F.M. Evaluation of EphB4 as Target for Image-Guided Surgery of Breast Cancer. Pharmaceuticals (Basel) 2020, 13. [Google Scholar] [CrossRef]
- Kumar, S.R.; Singh, J.; Xia, G.; Krasnoperov, V.; Hassanieh, L.; Ley, E.J.; Scehnet, J.; Kumar, N.G.; Hawes, D.; Press, M.F.; et al. Receptor tyrosine kinase EphB4 is a survival factor in breast cancer. Am J Pathol 2006, 169, 279–293. [Google Scholar] [CrossRef]
- Brantley-Sieders, D.M.; Jiang, A.; Sarma, K.; Badu-Nkansah, A.; Walter, D.L.; Shyr, Y.; Chen, J. Eph/ephrin profiling in human breast cancer reveals significant associations between expression level and clinical outcome. PLoS One 2011, 6, e24426. [Google Scholar] [CrossRef]
- Zheng, M.F.; Ji, Y.; Wu, X.B.; Ye, S.G.; Chen, J.Y. EphB4 gene polymorphism and protein expression in non-small-cell lung cancer. Mol Med Rep 2012, 6, 405–408. [Google Scholar] [CrossRef]
- Ferguson, B.D.; Liu, R.; Rolle, C.E.; Tan, Y.H.; Krasnoperov, V.; Kanteti, R.; Tretiakova, M.S.; Cervantes, G.M.; Hasina, R.; Hseu, R.D.; et al. The EphB4 receptor tyrosine kinase promotes lung cancer growth: a potential novel therapeutic target. PLoS One 2013, 8, e67668. [Google Scholar] [CrossRef]
- Nanamiya, R.; Saito-Koyama, R.; Miki, Y.; Inoue, C.; Asavasupreechar, T.; Abe, J.; Sato, I.; Sasano, H. EphB4 as a Novel Target for the EGFR-Independent Suppressive Effects of Osimertinib on Cell Cycle Progression in Non-Small Cell Lung Cancer. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Yin, J.; Cui, Y.; Li, L.; Ji, J.; Jiang, W.G. Overexpression of EPHB4 Is Associated with Poor Survival of Patients with Gastric Cancer. Anticancer Res 2017, 37, 4489–4497. [Google Scholar] [CrossRef]
- Stammes, M.A.; Prevoo, H.A.; Ter Horst, M.C.; Groot, S.A.; Van de Velde, C.J.; Chan, A.B.; de Geus-Oei, L.F.; Kuppen, P.J.; Vahrmeijer, A.L.; Pasquale, E.B.; et al. Evaluation of EphA2 and EphB4 as Targets for Image-Guided Colorectal Cancer Surgery. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Lv, J.; Xia, Q.; Wang, J.; Shen, Q.; Zhang, J.; Zhou, X. EphB4 promotes the proliferation, invasion, and angiogenesis of human colorectal cancer. Exp Mol Pathol 2016, 100, 402–408. [Google Scholar] [CrossRef]
- Merchant, A.A.; Jorapur, A.; McManus, A.; Liu, R.; Krasnoperov, V.; Chaudhry, P.; Singh, M.; Harton, L.; Agajanian, M.; Kim, M.; et al. EPHB4 is a therapeutic target in AML and promotes leukemia cell survival via AKT. Blood Adv 2017, 1, 1635–1644. [Google Scholar] [CrossRef]
- Alam, S.M.; Fujimoto, J.; Jahan, I.; Sato, E.; Tamaya, T. Coexpression of EphB4 and ephrinB2 in tumor advancement of uterine cervical cancers. Gynecol Oncol 2009, 114, 84–88. [Google Scholar] [CrossRef]
- Tu, Y.; He, S.; Fu, J.; Li, G.; Xu, R.; Lu, H.; Deng, J. Expression of EphrinB2 and EphB4 in glioma tissues correlated to the progression of glioma and the prognosis of glioblastoma patients. Clin Transl Oncol 2012, 14, 214–220. [Google Scholar] [CrossRef]
- Kumar, S.R.; Masood, R.; Spannuth, W.A.; Singh, J.; Scehnet, J.; Kleiber, G.; Jennings, N.; Deavers, M.; Krasnoperov, V.; Dubeau, L.; et al. The receptor tyrosine kinase EphB4 is overexpressed in ovarian cancer, provides survival signals and predicts poor outcome. Br J Cancer 2007, 96, 1083–1091. [Google Scholar] [CrossRef]
- Alam, S.M.; Fujimoto, J.; Jahan, I.; Sato, E.; Tamaya, T. Coexpression of EphB4 and ephrinB2 in tumour advancement of ovarian cancers. Br J Cancer 2008, 98, 845–851. [Google Scholar] [CrossRef]
- Sagar, V.; Vatapalli, R.; Lysy, B.; Pamarthy, S.; Anker, J.F.; Rodriguez, Y.; Han, H.; Unno, K.; Stadler, W.M.; Catalona, W.J.; et al. EPHB4 inhibition activates ER stress to promote immunogenic cell death of prostate cancer cells. Cell Death Dis 2019, 10, 801. [Google Scholar] [CrossRef]
- Xia, G.; Kumar, S.R.; Masood, R.; Zhu, S.; Reddy, R.; Krasnoperov, V.; Quinn, D.I.; Henshall, S.M.; Sutherland, R.L.; Pinski, J.K.; et al. EphB4 expression and biological significance in prostate cancer. Cancer Res 2005, 65, 4623–4632. [Google Scholar] [CrossRef]
- Xuqing, W.; Lei, C.; Zhengfa, M.; Shengchun, D.; Xin, F.; Jianguo, Q.; Jianxin, Z. EphB4 is overexpressed in papillary thyroid carcinoma and promotes the migration of papillary thyroid cancer cells. Tumour Biol 2012, 33, 1419–1427. [Google Scholar] [CrossRef]
- Sharma, G.K.; Dhillon, V.K.; Masood, R.; Maceri, D.R. Overexpression of EphB4, EphrinB2, and epidermal growth factor receptor in papillary thyroid carcinoma: A pilot study. Head Neck 2015, 37, 964–969. [Google Scholar] [CrossRef]
- Xia, G.; Kumar, S.R.; Stein, J.P.; Singh, J.; Krasnoperov, V.; Zhu, S.; Hassanieh, L.; Smith, D.L.; Buscarini, M.; Broek, D.; et al. EphB4 receptor tyrosine kinase is expressed in bladder cancer and provides signals for cell survival. Oncogene 2006, 25, 769–780. [Google Scholar] [CrossRef]
- Li, X.; Choi, W.W.; Yan, R.; Yu, H.; Krasnoperov, V.; Kumar, S.R.; Schuckman, A.; Klumpp, D.J.; Pan, C.X.; Quinn, D.; et al. The differential expression of EphB2 and EphB4 receptor kinases in normal bladder and in transitional cell carcinoma of the bladder. PLoS One 2014, 9, e105326. [Google Scholar] [CrossRef]
- Ferguson, B.D.; Carol Tan, Y.H.; Kanteti, R.S.; Liu, R.; Gayed, M.J.; Vokes, E.E.; Ferguson, M.K.; John Iafrate, A.; Gill, P.S.; Salgia, R. Novel EPHB4 Receptor Tyrosine Kinase Mutations and Kinomic Pathway Analysis in Lung Cancer. Sci Rep 2015, 5, 10641. [Google Scholar] [CrossRef]
- Masood, R.; Kumar, S.R.; Sinha, U.K.; Crowe, D.L.; Krasnoperov, V.; Reddy, R.K.; Zozulya, S.; Singh, J.; Xia, G.; Broek, D.; et al. EphB4 provides survival advantage to squamous cell carcinoma of the head and neck. Int J Cancer 2006, 119, 1236–1248. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Zhang, Y. Targeting receptor tyrosine kinase EphB4 in cancer therapy. Semin Cancer Biol 2019, 56, 37–46. [Google Scholar] [CrossRef]
- Noren, N.K.; Lu, M.; Freeman, A.L.; Koolpe, M.; Pasquale, E.B. Interplay between EphB4 on tumor cells and vascular ephrin-B2 regulates tumor growth. Proc Natl Acad Sci U S A 2004, 101, 5583–5588. [Google Scholar] [CrossRef]
- Piffko, A.; Broggini, T.; Harms, C.; Adams, R.H.; Vajkoczy, P.; Czabanka, M. Ligand-Dependent and Ligand-Independent Effects of Ephrin-B2-EphB4 Signaling in Melanoma Metastatic Spine Disease. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Aslam, M.I.; Abraham, J.; Mansoor, A.; Druker, B.J.; Tyner, J.W.; Keller, C. PDGFRbeta reverses EphB4 signaling in alveolar rhabdomyosarcoma. Proc Natl Acad Sci U S A 2014, 111, 6383–6388. [Google Scholar] [CrossRef]
- Noren, N.K.; Foos, G.; Hauser, C.A.; Pasquale, E.B. The EphB4 receptor suppresses breast cancer cell tumorigenicity through an Abl-Crk pathway. Nat Cell Biol 2006, 8, 815–825. [Google Scholar] [CrossRef]
- Wu, Q.; Suo, Z.; Risberg, B.; Karlsson, M.G.; Villman, K.; Nesland, J.M. Expression of Ephb2 and Ephb4 in breast carcinoma. Pathol Oncol Res 2004, 10, 26–33. [Google Scholar] [CrossRef]
- Ding, J.; Yao, Y.; Huang, G.; Wang, X.; Yi, J.; Zhang, N.; Liu, C.; Wang, K.; Zhang, Y.; Wang, M.; et al. Targeting the EphB4 receptor tyrosine kinase sensitizes HER2-positive breast cancer cells to Lapatinib. Cancer Lett 2020, 475, 53–64. [Google Scholar] [CrossRef]
- Zhu, M.; Tang, X.; Gong, Z.; Tang, W.; Zhang, Y. TAD1822-7 induces ROS-mediated apoptosis of HER2 positive breast cancer by decreasing E-cadherin in an EphB4 dependent manner. Life Sci 2021, 285, 119954. [Google Scholar] [CrossRef]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C(44)Mab-5. Biochem Biophys Rep 2018, 14, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016, 35, 293–299. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; Nogi, T.; Kato, Y.; Takagi, J. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014, 95, 240–247. [Google Scholar] [CrossRef]
- Nanamiya, R.; Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Hosono, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 101–106. [Google Scholar] [CrossRef]
- Tanaka, T.; Asano, T.; Sano, M.; Takei, J.; Hosono, H.; Nanamiya, R.; Nakamura, T.; Yanaka, M.; Harada, H.; Fukui, M.; et al. Development of Monoclonal Antibody PMab-269 Against California Sea Lion Podoplanin. Monoclon Antib Immunodiagn Immunother 2021, 40, 124–133. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Nanamiya, R.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Harada, H.; et al. Development of Anti-human T Cell Immunoreceptor with Ig and ITIM Domains (TIGIT) Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 71–75. [Google Scholar] [CrossRef]
- Sayama, Y.; Kaneko, M.K.; Takei, J.; Hosono, H.; Sano, M.; Asano, T.; Kato, Y. Establishment of a novel anti-TROP2 monoclonal antibody TrMab-29 for immunohistochemical analysis. Biochem Biophys Rep 2021, 25, 100902. [Google Scholar] [CrossRef]
- Sayama, Y.; Kaneko, M.K.; Kato, Y. Development and characterization of TrMab-6, a novel anti-TROP2 monoclonal antibody for antigen detection in breast cancer. Mol Med Rep 2021, 23. [Google Scholar] [CrossRef]
- Asano, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 3 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Sano, M.; Takei, J.; Asano, T.; Sayama, Y.; Hosono, H.; Kobayashi, A.; Konnai, S.; Kato, Y. Development and Characterization of Anti-Sheep Podoplanin Monoclonal Antibodies PMab-253 and PMab-260. Monoclon Antib Immunodiagn Immunother 2020, 39, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Kaneko, M.K.; Kato, Y. Establishment of C(20)Mab-11, a novel anti-CD20 monoclonal antibody, for the detection of B cells. Oncol Lett 2020, 20, 1961–1967. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Kaneko, M.K.; Kato, Y. Establishment of an Anti-CD20 Monoclonal Antibody (C(20)Mab-60) for Immunohistochemical Analyses. Monoclon Antib Immunodiagn Immunother 2020, 39, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Furusawa, Y.; Yamada, S.; Itai, S.; Takei, J.; Sano, M.; Kaneko, M.K. Establishment of a monoclonal antibody PMab-225 against alpaca podoplanin for immunohistochemical analyses. Biochem Biophys Rep 2019, 18, 100633. [Google Scholar] [CrossRef]
- Ikota, H.; Nobusawa, S.; Arai, H.; Kato, Y.; Ishizawa, K.; Hirose, T.; Yokoo, H. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015, 32, 237–244. [Google Scholar] [CrossRef]
- Carles-Kinch, K.; Kilpatrick, K.E.; Stewart, J.C.; Kinch, M.S. Antibody targeting of the EphA2 tyrosine kinase inhibits malignant cell behavior. Cancer Res 2002, 62, 2840–2847. [Google Scholar]
- Coffman, K.T.; Hu, M.; Carles-Kinch, K.; Tice, D.; Donacki, N.; Munyon, K.; Kifle, G.; Woods, R.; Langermann, S.; Kiener, P.A.; et al. Differential EphA2 epitope display on normal versus malignant cells. Cancer Res 2003, 63, 7907–7912. [Google Scholar]
- Landen, C.N., Jr.; Lu, C.; Han, L.Y.; Coffman, K.T.; Bruckheimer, E.; Halder, J.; Mangala, L.S.; Merritt, W.M.; Lin, Y.G.; Gao, C.; et al. Efficacy and antivascular effects of EphA2 reduction with an agonistic antibody in ovarian cancer. J Natl Cancer Inst 2006, 98, 1558–1570. [Google Scholar] [CrossRef]
- Merritt, W.M.; Kamat, A.A.; Hwang, J.Y.; Bottsford-Miller, J.; Lu, C.; Lin, Y.G.; Coffey, D.; Spannuth, W.A.; Nugent, E.; Han, L.Y.; et al. Clinical and biological impact of EphA2 overexpression and angiogenesis in endometrial cancer. Cancer Biol Ther 2010, 10, 1306–1314. [Google Scholar] [CrossRef]
- Gokmen-Polar, Y.; Toroni, R.A.; Hocevar, B.A.; Badve, S.; Zhao, Q.; Shen, C.; Bruckheimer, E.; Kinch, M.S.; Miller, K.D. Dual targeting of EphA2 and ER restores tamoxifen sensitivity in ER/EphA2-positive breast cancer. Breast Cancer Res Treat 2011, 127, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Ansuini, H.; Meola, A.; Gunes, Z.; Paradisi, V.; Pezzanera, M.; Acali, S.; Santini, C.; Luzzago, A.; Mori, F.; Lazzaro, D.; et al. Anti-EphA2 Antibodies with Distinct In Vitro Properties Have Equal In Vivo Efficacy in Pancreatic Cancer. J Oncol 2009, 2009, 951917. [Google Scholar] [CrossRef] [PubMed]
- Wesa, A.K.; Herrem, C.J.; Mandic, M.; Taylor, J.L.; Vasquez, C.; Kawabe, M.; Tatsumi, T.; Leibowitz, M.S.; Finke, J.H.; Bukowski, R.M.; et al. Enhancement in specific CD8+ T cell recognition of EphA2+ tumors in vitro and in vivo after treatment with ligand agonists. J Immunol 2008, 181, 7721–7727. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, J.; Sue, M.; Yamato, M.; Ichikawa, J.; Ishida, S.; Shibutani, T.; Kitamura, M.; Wada, T.; Agatsuma, T. Novel anti-EPHA2 antibody, DS-8895a for cancer treatment. Cancer Biol Ther 2016, 17, 1158–1167. [Google Scholar] [CrossRef]
- Bruckheimer, E.M.; Fazenbaker, C.A.; Gallagher, S.; Mulgrew, K.; Fuhrmann, S.; Coffman, K.T.; Walsh, W.; Ready, S.; Cook, K.; Damschroder, M.; et al. Antibody-dependent cell-mediated cytotoxicity effector-enhanced EphA2 agonist monoclonal antibody demonstrates potent activity against human tumors. Neoplasia 2009, 11, 509–517, 502 p following 517. [Google Scholar] [CrossRef]
- Charmsaz, S.; Beckett, K.; Smith, F.M.; Bruedigam, C.; Moore, A.S.; Al-Ejeh, F.; Lane, S.W.; Boyd, A.W. EphA2 Is a Therapy Target in EphA2-Positive Leukemias but Is Not Essential for Normal Hematopoiesis or Leukemia. PLoS One 2015, 10, e0130692. [Google Scholar] [CrossRef]
- Jackson, D.; Gooya, J.; Mao, S.; Kinneer, K.; Xu, L.; Camara, M.; Fazenbaker, C.; Fleming, R.; Swamynathan, S.; Meyer, D.; et al. A human antibody-drug conjugate targeting EphA2 inhibits tumor growth in vivo. Cancer Res 2008, 68, 9367–9374. [Google Scholar] [CrossRef]
- Lee, J.W.; Stone, R.L.; Lee, S.J.; Nam, E.J.; Roh, J.W.; Nick, A.M.; Han, H.D.; Shahzad, M.M.; Kim, H.S.; Mangala, L.S.; et al. EphA2 targeted chemotherapy using an antibody drug conjugate in endometrial carcinoma. Clin Cancer Res 2010, 16, 2562–2570. [Google Scholar] [CrossRef]
- Shitara, K.; Satoh, T.; Iwasa, S.; Yamaguchi, K.; Muro, K.; Komatsu, Y.; Nishina, T.; Esaki, T.; Hasegawa, J.; Kakurai, Y.; et al. Safety, tolerability, pharmacokinetics, and pharmacodynamics of the afucosylated, humanized anti-EPHA2 antibody DS-8895a: a first-in-human phase I dose escalation and dose expansion study in patients with advanced solid tumors. J Immunother Cancer 2019, 7, 219. [Google Scholar] [CrossRef]
- Annunziata, C.M.; Kohn, E.C.; LoRusso, P.; Houston, N.D.; Coleman, R.L.; Buzoianu, M.; Robbie, G.; Lechleider, R. Phase 1, open-label study of MEDI-547 in patients with relapsed or refractory solid tumors. Invest New Drugs 2013, 31, 77–84. [Google Scholar] [CrossRef]
- Sakamoto, A.; Kato, K.; Hasegawa, T.; Ikeda, S. An Agonistic Antibody to EPHA2 Exhibits Antitumor Effects on Human Melanoma Cells. Anticancer Res 2018, 38, 3273–3282. [Google Scholar] [CrossRef] [PubMed]
- Vearing, C.; Lee, F.T.; Wimmer-Kleikamp, S.; Spirkoska, V.; To, C.; Stylianou, C.; Spanevello, M.; Brechbiel, M.; Boyd, A.W.; Scott, A.M.; et al. Concurrent binding of anti-EphA3 antibody and ephrin-A5 amplifies EphA3 signaling and downstream responses: potential as EphA3-specific tumor-targeting reagents. Cancer Res 2005, 65, 6745–6754. [Google Scholar] [CrossRef] [PubMed]
- Charmsaz, S.; Al-Ejeh, F.; Yeadon, T.M.; Miller, K.J.; Smith, F.M.; Stringer, B.W.; Moore, A.S.; Lee, F.T.; Cooper, L.T.; Stylianou, C.; et al. EphA3 as a target for antibody immunotherapy in acute lymphoblastic leukemia. Leukemia 2017, 31, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Vail, M.E.; Murone, C.; Tan, A.; Hii, L.; Abebe, D.; Janes, P.W.; Lee, F.T.; Baer, M.; Palath, V.; Bebbington, C.; et al. Targeting EphA3 inhibits cancer growth by disrupting the tumor stromal microenvironment. Cancer Res 2014, 74, 4470–4481. [Google Scholar] [CrossRef] [PubMed]
- Swords, R.T.; Greenberg, P.L.; Wei, A.H.; Durrant, S.; Advani, A.S.; Hertzberg, M.S.; Jonas, B.A.; Lewis, I.D.; Rivera, G.; Gratzinger, D.; et al. KB004, a first in class monoclonal antibody targeting the receptor tyrosine kinase EphA3, in patients with advanced hematologic malignancies: Results from a phase 1 study. Leuk Res 2016, 50, 123–131. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, F.; Airoldi, I.; Di Carlo, E.; Marotta, P.; Falco, G.; Simeon, V.; Laurenzana, I.; Trino, S.; De Luca, L.; Todoerti, K.; et al. EphA3 targeting reduces in vitro adhesion and invasion and in vivo growth and angiogenesis of multiple myeloma cells. Cell Oncol (Dordr) 2017, 40, 483–496. [Google Scholar] [CrossRef]
- Day, B.W.; Stringer, B.W.; Al-Ejeh, F.; Ting, M.J.; Wilson, J.; Ensbey, K.S.; Jamieson, P.R.; Bruce, Z.C.; Lim, Y.C.; Offenhauser, C.; et al. EphA3 maintains tumorigenicity and is a therapeutic target in glioblastoma multiforme. Cancer Cell 2013, 23, 238–248. [Google Scholar] [CrossRef]
- Chu, L.; Wang, A.; Ni, L.; Yan, X.; Song, Y.; Zhao, M.; Sun, K.; Mu, H.; Liu, S.; Wu, Z.; et al. Nose-to-brain delivery of temozolomide-loaded PLGA nanoparticles functionalized with anti-EPHA3 for glioblastoma targeting. Drug Deliv 2018, 25, 1634–1641. [Google Scholar] [CrossRef]
- Offenhauser, C.; Al-Ejeh, F.; Puttick, S.; Ensbey, K.S.; Bruce, Z.C.; Jamieson, P.R.; Smith, F.M.; Stringer, B.W.; Carrington, B.; Fuchs, A.V.; et al. EphA3 Pay-Loaded Antibody Therapeutics for the Treatment of Glioblastoma. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Taki, S.; Kamada, H.; Inoue, M.; Nagano, K.; Mukai, Y.; Higashisaka, K.; Yoshioka, Y.; Tsutsumi, Y.; Tsunoda, S. A Novel Bispecific Antibody against Human CD3 and Ephrin Receptor A10 for Breast Cancer Therapy. Plos One 2015, 10. [Google Scholar] [CrossRef]
- Mao, W.; Luis, E.; Ross, S.; Silva, J.; Tan, C.; Crowley, C.; Chui, C.; Franz, G.; Senter, P.; Koeppen, H.; et al. EphB2 as a therapeutic antibody drug target for the treatment of colorectal cancer. Cancer Res 2004, 64, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Krasnoperov, V.; Kumar, S.R.; Ley, E.; Li, X.; Scehnet, J.; Liu, R.; Zozulya, S.; Gill, P.S. Novel EphB4 monoclonal antibodies modulate angiogenesis and inhibit tumor growth. Am J Pathol 2010, 176, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, S.A.; Douglas, E.L.; Mertens-Walker, I.; Lisle, J.E.; Maharaj, M.S.; Herington, A.C. Anti-tumour effects of antibodies targeting the extracellular cysteine-rich region of the receptor tyrosine kinase EphB4. Oncotarget 2015, 6, 7554–7569. [Google Scholar] [CrossRef] [PubMed]
- Janes, P.W.; Vail, M.E.; Gan, H.K.; Scott, A.M. Antibody Targeting of Eph Receptors in Cancer. Pharmaceuticals (Basel) 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Cher, L.; Inglis, P.; Lwin, Z.; Lau, E.; Ackermann, U.; Coombs, N.; Remen, K.; Guo, N.; Lee, S.T.; et al. Preliminary Findings of a Phase I Safety and Bioimaging Trial of Kb004 (Ifabotuzumab) in Patients with Glioblastoma. Neuro-Oncology 2019, 21, 6–6. [Google Scholar] [CrossRef]
- Boyd, A.W.; Ward, L.D.; Wicks, I.P.; Simpson, R.J.; Salvaris, E.; Wilks, A.; Welch, K.; Loudovaris, M.; Rockman, S.; Busmanis, I. Isolation and characterization of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line. J Biol Chem 1992, 267, 3262–3267. [Google Scholar] [CrossRef]
- Lin, Q.; Ba, T.; Ho, J.; Chen, D.; Cheng, Y.; Wang, L.; Xu, G.; Xu, L.; Zhou, Y.; Wei, Y.; et al. First-in-Human Trial of EphA2-Redirected CAR T-Cells in Patients With Recurrent Glioblastoma: A Preliminary Report of Three Cases at the Starting Dose. Front Oncol 2021, 11, 694941. [Google Scholar] [CrossRef]
- Cha, J.H.; Chan, L.C.; Wang, Y.N.; Chu, Y.Y.; Wang, C.H.; Lee, H.H.; Xia, W.; Shyu, W.C.; Liu, S.P.; Yao, J.; et al. Ephrin receptor A10 monoclonal antibodies and the derived chimeric antigen receptor T cells exert an antitumor response in mouse models of triple-negative breast cancer. J Biol Chem 2022, 298, 101817. [Google Scholar] [CrossRef]
- Sharma, P.; Roberts, C.; Herpai, D.; Fokt, I.D.; Priebe, W.; Debinski, W. Drug Conjugates for Targeting Eph Receptors in Glioblastoma. Pharmaceuticals (Basel) 2020, 13. [Google Scholar] [CrossRef]
- Hammond, S.A.; Lutterbuese, R.; Roff, S.; Lutterbuese, P.; Schlereth, B.; Bruckheimer, E.; Kinch, M.S.; Coats, S.; Baeuerle, P.A.; Kufer, P.; et al. Selective targeting and potent control of tumor growth using an EphA2/CD3-Bispecific single-chain antibody construct. Cancer Res 2007, 67, 3927–3935. [Google Scholar] [CrossRef]
- Janowicz, P.W.; Houston, Z.H.; Bunt, J.; Fletcher, N.L.; Bell, C.A.; Cowin, G.; Howard, C.B.; Taslima, D.; Westra van Holthe, N.; Prior, A.; et al. Understanding nanomedicine treatment in an aggressive spontaneous brain cancer model at the stage of early blood brain barrier disruption. Biomaterials 2022, 283, 121416. [Google Scholar] [CrossRef] [PubMed]
- Qazi, M.A.; Vora, P.; Venugopal, C.; Adams, J.; Singh, M.; Hu, A.; Gorelik, M.; Subapanditha, M.K.; Savage, N.; Yang, J.; et al. Cotargeting Ephrin Receptor Tyrosine Kinases A2 and A3 in Cancer Stem Cells Reduces Growth of Recurrent Glioblastoma. Cancer Res 2018, 78, 5023–5037. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, W.; Swindell, E.; Pien, C.; Luus, L.; Cain, J.; Pham, M.; Kandela, I.; Huang, Z.R.; Tipparaju, S.K.; Koshkaryev, A.; et al. Targeting EphA2 in Bladder Cancer Using a Novel Antibody-Directed Nanotherapeutic. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Production of anti-EphB4 mAbs. The procedure of Cell-Based Immunization and Screening (CBIS) method. (A) The EphB4-overexpressed cell lines were immunized into BALB/c mice by intraperitoneal injection. (B) Mouse spleens were harvested, fused with splenocytes and myeloma cells, and seeded into 96-well plates. (C) Hybridoma screening was performed using flow cytometry to select wells that did not react with the parental cell lines but with the EphB4-overexpressed cell lines. (D) Positive wells were cloned by limiting dilution.
Figure 1.
Production of anti-EphB4 mAbs. The procedure of Cell-Based Immunization and Screening (CBIS) method. (A) The EphB4-overexpressed cell lines were immunized into BALB/c mice by intraperitoneal injection. (B) Mouse spleens were harvested, fused with splenocytes and myeloma cells, and seeded into 96-well plates. (C) Hybridoma screening was performed using flow cytometry to select wells that did not react with the parental cell lines but with the EphB4-overexpressed cell lines. (D) Positive wells were cloned by limiting dilution.
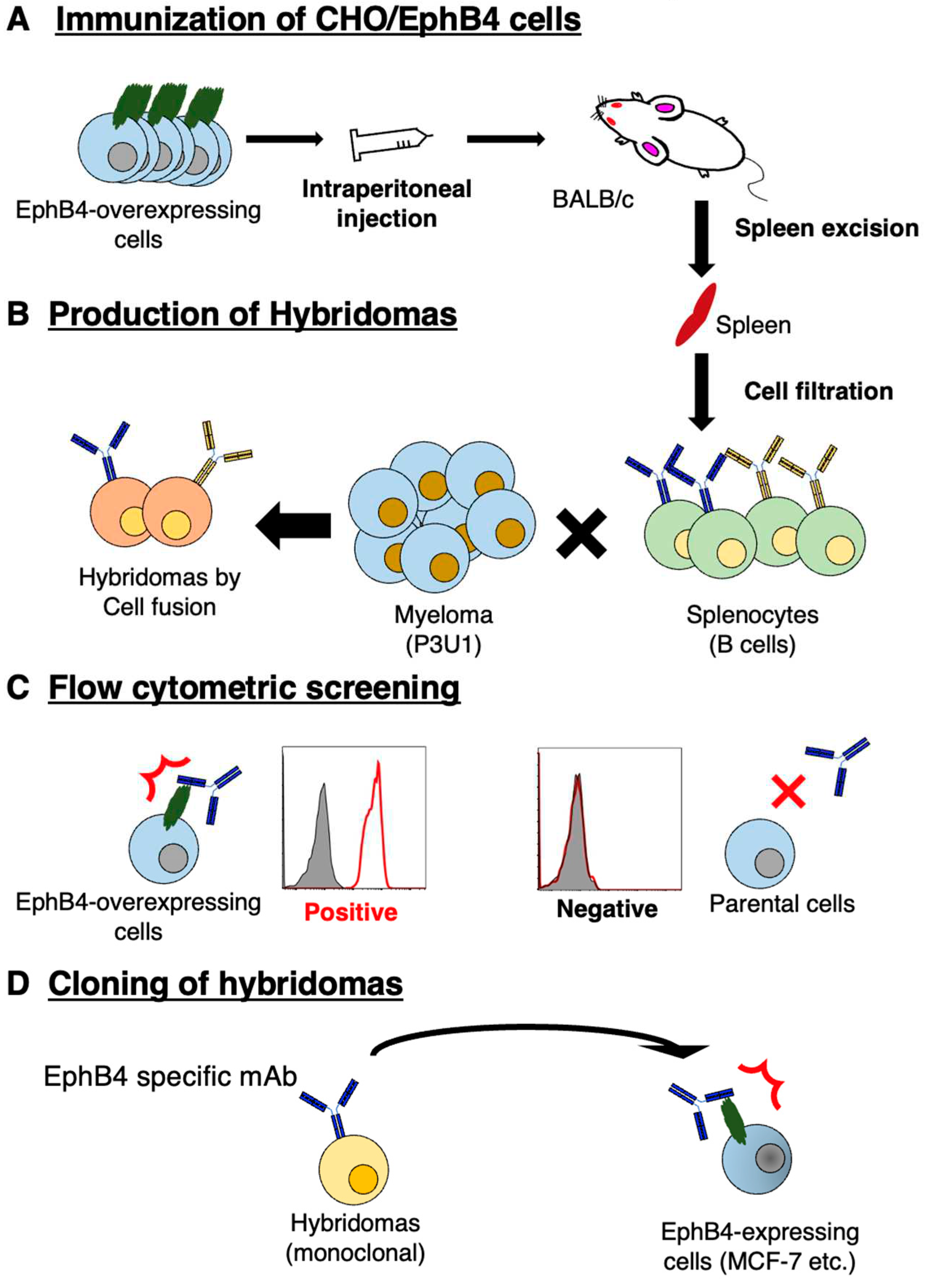
Figure 2.
Figure 2. Flow cytometry using B4Mab-7. (A-E) CHO/EphB4, CHO-K1, MCF-7, MDA-MB-468, and BINDS-52 (EphB4-knockout MCF-7) were treated with 0.1 or 1 μg/mL of B4Mab-7, followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Filled, negative control.
Figure 2.
Figure 2. Flow cytometry using B4Mab-7. (A-E) CHO/EphB4, CHO-K1, MCF-7, MDA-MB-468, and BINDS-52 (EphB4-knockout MCF-7) were treated with 0.1 or 1 μg/mL of B4Mab-7, followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Filled, negative control.
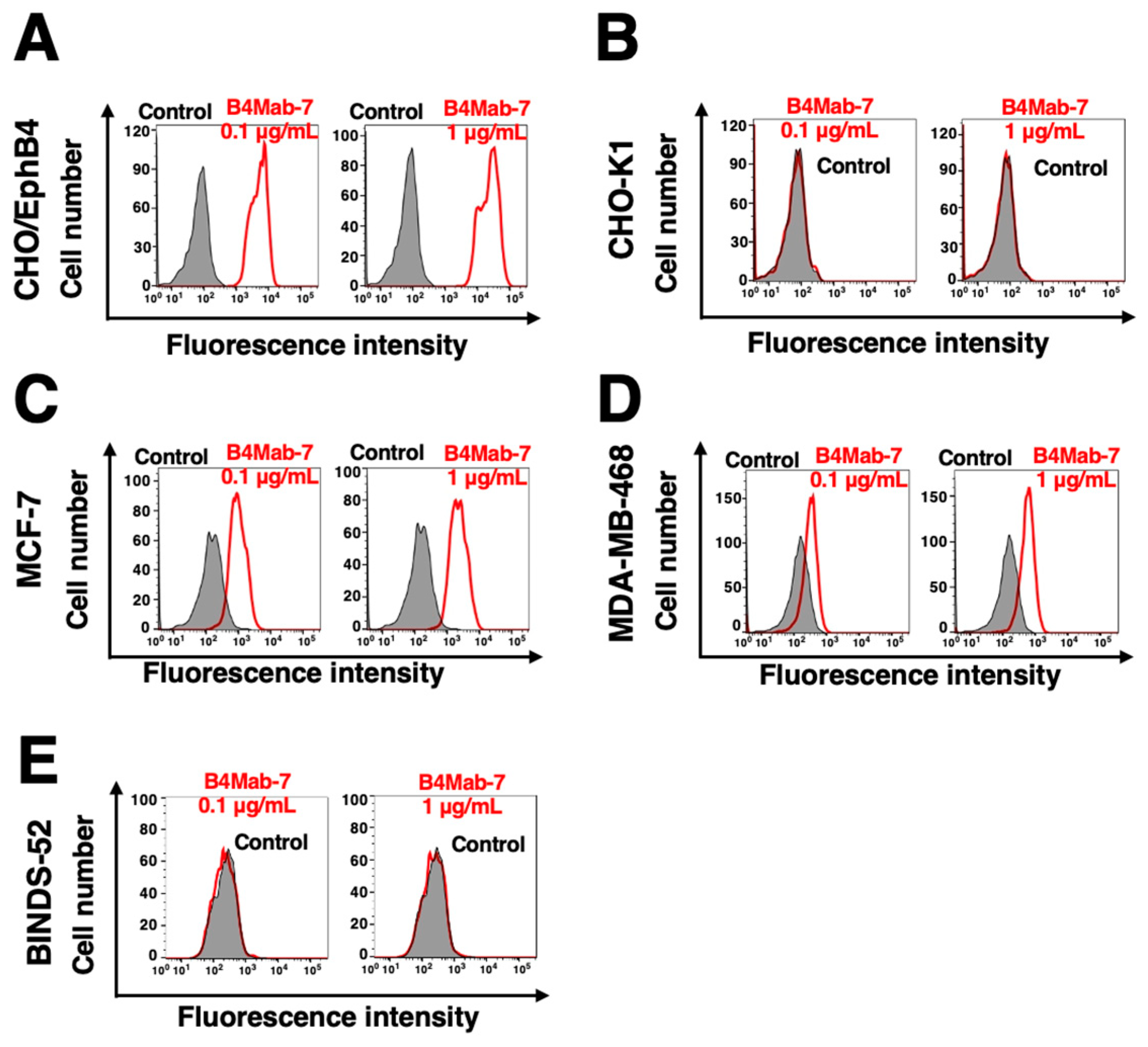
Figure 3.
Determination of the binding affinity of B4Mab-7. (A-C) CHO/EphB4, MCF-7, or MDA-MB-468 were suspended in 100 μL of serially diluted B4Mab-7 (6 ng/mL-50 μg/mL). Alexa Fluor 488-conjugated anti-mouse IgG was then added. Fluorescence data were collected using the SA3800 cell analyzer.
Figure 3.
Determination of the binding affinity of B4Mab-7. (A-C) CHO/EphB4, MCF-7, or MDA-MB-468 were suspended in 100 μL of serially diluted B4Mab-7 (6 ng/mL-50 μg/mL). Alexa Fluor 488-conjugated anti-mouse IgG was then added. Fluorescence data were collected using the SA3800 cell analyzer.
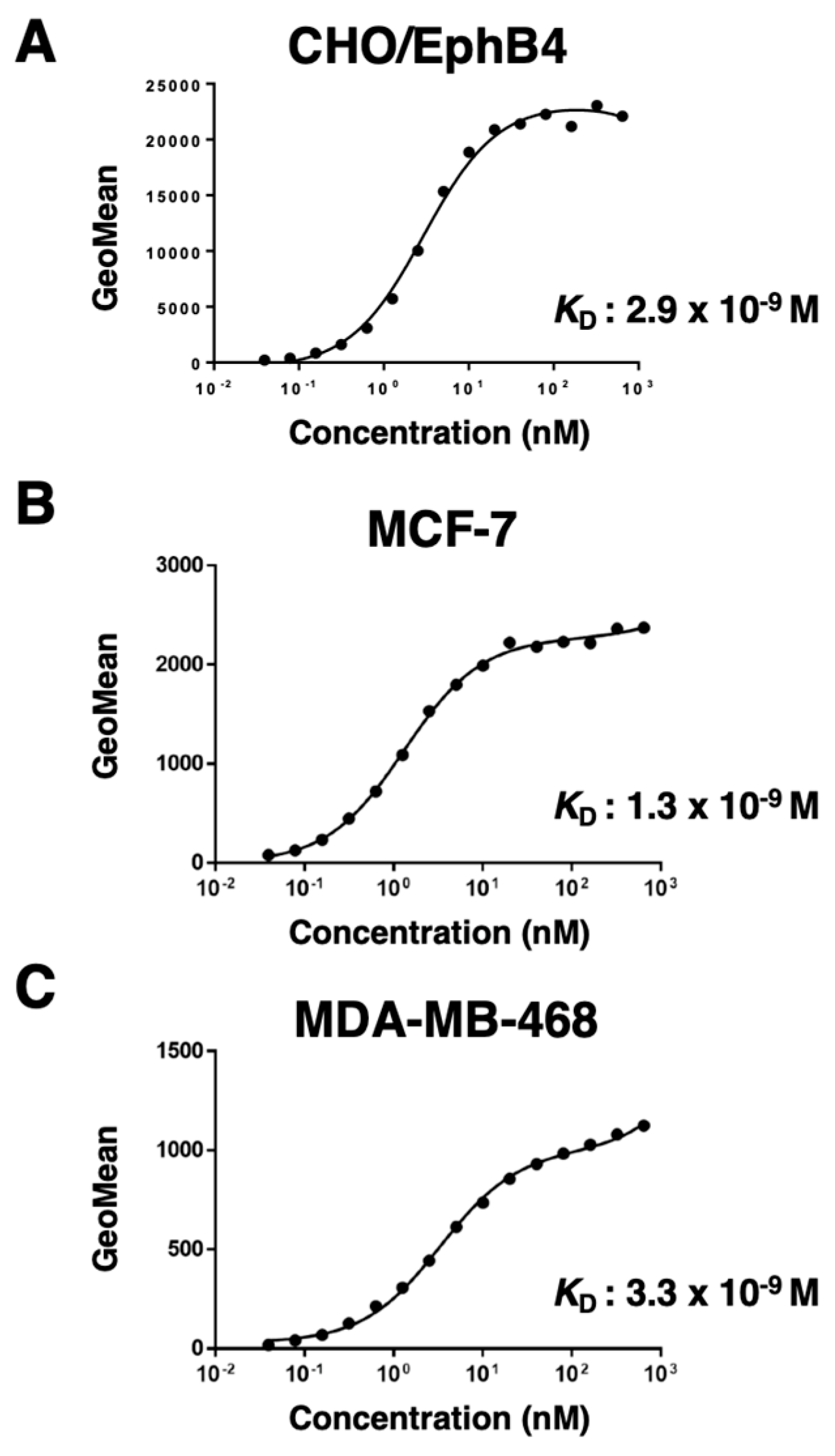
Figure 4.
Western blot analysis using B4Mab-7. Cell lysates (10 μg) were electrophoresed, and proteins were transferred onto polyvinylidene difluoride membranes. After blocking, membranes were incubated with 1 μg/mL of B4Mab-7 (A) or 1 μg/mL of an anti-IDH1 mAb (clone: RcMab-1) (B). The membranes were then incubated with peroxidase-conjugated anti-mouse immunoglobulins and rat immunoglobulins, respectively.
Figure 4.
Western blot analysis using B4Mab-7. Cell lysates (10 μg) were electrophoresed, and proteins were transferred onto polyvinylidene difluoride membranes. After blocking, membranes were incubated with 1 μg/mL of B4Mab-7 (A) or 1 μg/mL of an anti-IDH1 mAb (clone: RcMab-1) (B). The membranes were then incubated with peroxidase-conjugated anti-mouse immunoglobulins and rat immunoglobulins, respectively.
Figure 5.
Immunohistochemical analysis by B4Mab-7 for breast cancers. After antigen retrieval using EnVision FLEX Target Retrieval Solution High pH, tissue arrays were incubated with 10 μg/mL of primary B4Mab-7 for 1 hour at room temperature, followed by treatment with Envision+ kit for 30 minutes. The color was developed using 3,3-diaminobenzidine tetrahydrochloride for 2 minutes, and sections were then counterstained with hematoxylin. (A, D) The 3+ staining pattern (case 1), (B, E) The buffer control (case 1), (C, F) The H&E staining (case 1). (G, J) The 2+ staining pattern (case 2), (H, K) The buffer control (case 2), (I, L) The H&E staining (case 2), (M, P) The negative staining pattern (case 3), (N, Q) The buffer control (case 3), (O, R) The H&E staining (case 3). Scale bar = 100 μm. H&E, hematoxylin and eosin.
Figure 5.
Immunohistochemical analysis by B4Mab-7 for breast cancers. After antigen retrieval using EnVision FLEX Target Retrieval Solution High pH, tissue arrays were incubated with 10 μg/mL of primary B4Mab-7 for 1 hour at room temperature, followed by treatment with Envision+ kit for 30 minutes. The color was developed using 3,3-diaminobenzidine tetrahydrochloride for 2 minutes, and sections were then counterstained with hematoxylin. (A, D) The 3+ staining pattern (case 1), (B, E) The buffer control (case 1), (C, F) The H&E staining (case 1). (G, J) The 2+ staining pattern (case 2), (H, K) The buffer control (case 2), (I, L) The H&E staining (case 2), (M, P) The negative staining pattern (case 3), (N, Q) The buffer control (case 3), (O, R) The H&E staining (case 3). Scale bar = 100 μm. H&E, hematoxylin and eosin.
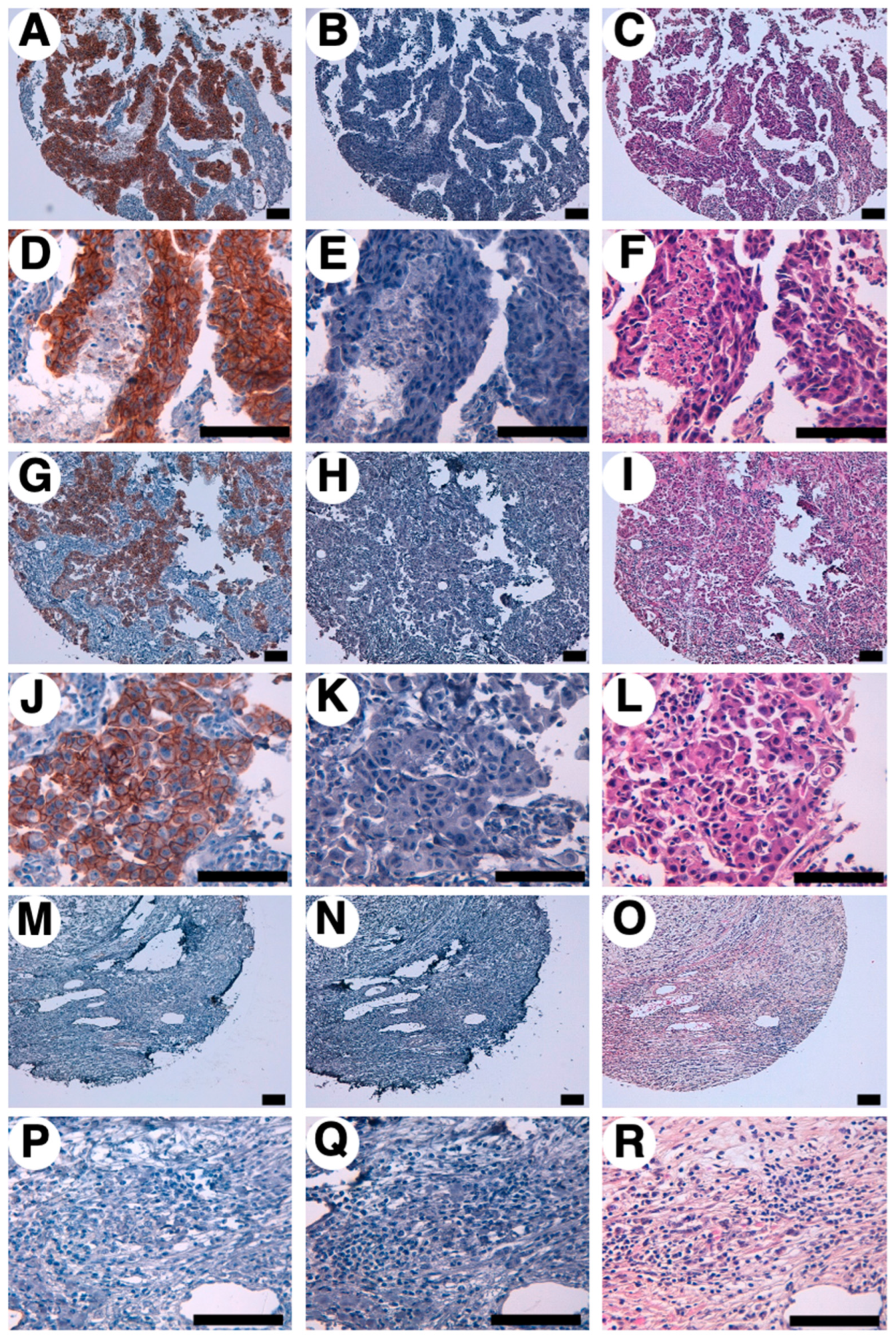

Table 1.
Results of EphB4 immunostaining in 63 patients with breast cancers.
| No. | Sex | Age | Organ | Pathology diagnosis | Differentiation | TNM | B4Mab-7 |
| 1 | F | 44 | Breast | Invasive ductal carcinoma | Moderately | T2N2M1 | 2+ |
| 2 | F | 58 | Breast | Medullary carcinoma | Moderately | T2N2M1 | 3+ |
| 3 | F | 40 | Breast | Invasive ductal carcinoma | Moderately | T2N1M0 | 1+ |
| 4 | F | 52 | Breast | Invasive ductal carcinoma | Moderately | T2N2M1 | 0 |
| 5 | F | 60 | Breast | Invasive ductal carcinoma | Moderately | T2N1M1 | 1+ |
| 6 | F | 57 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 7 | F | 48 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 8 | F | 66 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 0 |
| 9 | F | 58 | Breast | Adenocarcinoma | Moderately | T2N2M1 | 0 |
| 10 | F | 63 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 11 | F | 32 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 12 | F | 59 | Breast | Invasive lobular carcinoma | Well | T2N2M0 | 0 |
| 13 | F | 44 | Breast | Invasive lobular carcinoma | Well | T2N2M0 | 0 |
| 14 | F | 60 | Breast | Invasive lobular carcinoma | Moderately | T2N1M0 | 1+ |
| 15 | F | 44 | Breast | Invasive ductal carcinoma | Moderately | T2N2M0 | 3+ |
| 16 | F | 82 | Breast | Invasive ductal carcinoma | Moderately | T2N1M1 | 1+ |
| 17 | F | 58 | Breast | Adenocarcinoma | Moderately | T2N1M1 | 2+ |
| 18 | F | 57 | Breast | Invasive ductal carcinoma | Poorly | T3N3M0 | 1+ |
| 19 | F | 41 | Breast | Invasive ductal carcinoma | Moderately | T2N1M0 | 3+ |
| 20 | F | 44 | Breast | Invasive ductal carcinoma | Moderately | T2N2M0 | 0 |
| 21 | F | 78 | Breast | Invasive ductal carcinoma | Moderately | T2N1M0 | 1+ |
| 22 | F | 60 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 23 | F | / | Breast | Invasive ductal carcinoma | Moderately | T2N1M1 | 1+ |
| 24 | F | 46 | Breast | Invasive ductal carcinoma | Moderately | T2N3M1 | 0 |
| 25 | F | 41 | Breast | Invasive ductal carcinoma | Moderately | T2N2M0 | 0 |
| 26 | F | 59 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 1+ |
| 27 | F | 45 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 0 |
| 28 | F | 43 | Breast | Invasive ductal carcinoma | N/A | T2N1M1 | 0 |
| 29 | F | 26 | Breast | Fibroadenoma | N/A | T1N0M0 | 2+ |
| 30 | F | 40 | Breast | Invasive ductal carcinoma | N/A | T1N0M0 | 0 |
| 31 | F | 38 | Breast | Fibroadenoma | N/A | T2N0M0 | 1+ |
| 32 | F | 51 | Breast | Invasive ductal carcinoma | Moderately | T2N2M0 | 0 |
| 33 | F | 45 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 1+ |
| 34 | F | 45 | Breast | Invasive ductal carcinoma | Poorly | T2N1M0 | 2+ |
| 35 | F | 47 | Breast | Invasive ductal carcinoma | Moderately | T2N1M0 | 0 |
| 36 | F | 55 | Breast | Invasive ductal carcinoma | Moderately | T2N3M1 | 2+ |
| 37 | F | 58 | Breast | Invasive ductal carcinoma | Moderately | T3N3M0 | 1+ |
| 38 | F | 47 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 39 | F | 38 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 1+ |
| 40 | F | 40 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 2+ |
| 41 | F | 57 | Breast | Invasive ductal carcinoma | Poorly | T2N0M0 | 0 |
| 42 | F | 42 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 43 | F | 60 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 44 | F | 58 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 0 |
| 45 | F | 41 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 0 |
| 46 | F | 50 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 47 | F | 60 | Breast | Invasive ductal carcinoma | Moderately | T2N2M1 | 0 |
| 48 | F | 53 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 49 | F | 65 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 0 |
| 50 | F | 43 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 51 | F | 57 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 52 | F | 37 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 53 | F | 50 | Breast | Invasive ductal carcinoma | Moderately | T2N3M0 | 0 |
| 54 | F | 48 | Breast | Invasive ductal carcinoma | Poorly | T2N1M0 | 0 |
| 55 | F | 50 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 56 | F | 53 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 3+ |
| 57 | F | 49 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 58 | F | 65 | Breast | Invasive ductal carcinoma | Moderately | T2N1M0 | 0 |
| 59 | F | 43 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 2+ |
| 60 | F | 58 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 0 |
| 61 | F | 48 | Breast | Invasive ductal carcinoma | Moderately | T2N0M0 | 1+ |
| 62 | F | / | Breast | Invasive ductal carcinoma | Moderately | TxNxMx | 2+ |
| 63 | F | / | Breast | Invasive ductal carcinoma | Moderately | TxNxMx | 1+ |
Table 2.
Summary of B4Mab-7 immunostaining in 63 patients with breast cancers.
| Pathology diagnosis | No. of cases | B4Mab-7 immunostaining | No. of positive cases | |||
| 3+ | 2+ | 1+ | 0 | |||
| Invasive ductal carcinoma | 55 | 6 | 12 | 18 | 19 | 36/55 (65.4%) |
| Invasive lobular carcinoma | 3 | 0 | 0 | 1 | 2 | 1/3 (33.3%) |
| Adenocarcinoma | 2 | 0 | 1 | 0 | 1 | 1/2 (50.0%) |
| Fibroadenoma | 2 | 0 | 1 | 1 | 0 | 2/2 (100%) |
| Medullary carcinoma | 1 | 1 | 0 | 0 | 0 | 1/1 (100%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



