
Submitted:
04 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Ranunculus asiaticus L. is an ornamental geophyte, mainly propagated by rehydrated tuberous roots. Vernalization before planting is a common practice to overcome the natural dormancy of tuberous roots, however little is known about the mechanisms underlying the plant response to low temperature. We investigated the influence of two hybrids, MBO and MDR, and three preparation procedures of tuberous roots, only rehydration (control, C), and rehydration plus vernalization at 3.5 °C for 2 weeks (V2) and for 4 weeks (V4), on plant growth, leaf photosynthesis, flowering and metabolism in plants grown in pot in unheated greenhouse. We reported the responses observed in the aerial part previously in this journal (Fusco et al., 2023). In this paper, we show changes in carbohydrate, amino acids, polyphenols and protein levels throughout the growing cycle, in the different plant stages: pre-planting, vegetative phase and flowering.
Keywords:
geophytes
; tuberous roots
; cold requirement
; GABA
; BCAAs
; polyphenols.
1. Introduction
Ranunculus asiaticus L. (Family Ranunculaceae) is a perennial geophyte cultivated for cut flower and flowering potted plant production, which is gaining attention for gardening and landscape design [1]. Cultivation of R. asiaticus has been growing in the recent years, also thank to the advance of knowledge on vegetative propagation through the underground storage organs (namely tuberous roots), and the development of new hybrids. As several spring flowering geophytes, R. asiaticus exhibits a summer rest period and requires a warm-cold-warm sequence to complete its life cycle [1]. In fact, in the cool and wet winter period plants are in a vegetative phase, while in the dry and hot summer, they require a quiescence phase [2]. In Mediterranean climate, the first autumn rainfalls rehydrate the tissues of dry dormant tuberous roots of spontaneous ranunculus making them sprouting and developing leaf rosettes [3]. Flowering lasts from February to May, when the aerial part wilts and the roots enter in dormancy [1]. Unlike other geophytes, in R. asiaticus the shoot apical meristem of dormant tuberous roots is inactive, while its activity restarts under mild temperatures, after rehydration. Tuberous roots may survive to long quiescence, tolerating prolonged storage, so that ranunculus is recognised as a “resurrection geophyte” species [4,5]. During root growth, cortical cells size increases, and cell walls get rich in pectin materials, while starch granules and protein bodies accumulate for the next growth phase of plants, upon awakening from dormancy [4,5].
Plant cultivation of R. asiaticus for commercial production can start either via seeds or via tuberous roots, harvested at the end of the growing period and prepared as propagation material through dehydration to less than 15% of moisture content. Planting tuberous roots anticipates flowering and increases flower stem yield compared to sowing [1]. However, flowering is influenced by the thermal history of tuberous roots before and after planting, and the photoperiod experienced during the growth, as plants exhibit a low temperature requirement (5-10/12-25 °C night/day, optimum day 16 °C) and a quantitative photoperiodic response to long day [3]. Cold treatments of tuberous roots (vernalization) hasten sprouting, leaf rosette formation, and flowering, and increase the number of flowers per plant compared to untreated roots (4), breaking the summer dormancy, similarly to what occurs in nature [5]. Glier and Caruso [6] demonstrated that low temperature is pivotal to induce the starch breakdown, increasing the availability of free sugars like sucrose [7], down-regulates the biosynthesis of abscisic acid (ABA), and promotes that of active forms of gibberellic acid (GA) [8]. In the quantitative (or facultative) response, the request for a certain condition (i.e., long day for R. asiaticus) as inductive stimulus is not strictly required for flowering, and the occurrence of this condition accelerates or enlarges flowering, which however is still possible in different conditions [9].
In Lilium pumilum the vernalization ends bulb dormancy and activates starch breakdown and accumulation of free sugars (e.g., sucrose) [7]. This process could allow both cryoprotection to the developing leaf rosette, and an energy source for the following bolting process [6]. Indeed, GA has an antagonist role against ABA in the regulation of bulb dormancy [8]. The exposure to low temperatures allows the overcoming of dormancy by repressing and/or downregulating the genes involved in the ABA and jasmonic acid (JA) biosynthetic pathways, thus relieving the antagonistic effect of ABA and JA on GA signalling and promoting sprouting [10]. According to literature data, tuberous roots of R. asiaticus undergo a fast starch hydrolysis upon vernalization, promoting not only the synthesis and accumulation of sucrose, but also of other primary metabolites acting as osmolytes and antioxidants, like proline, alanine and γ-aminobutyic acid (GABA) [11,12]. In addition, minor amino acids, and particularly branched chain amino acids (BCAAs) whose synthesis increases when hexoses increase under stress [13], may exert a scavenging function against reactive oxygen species (ROS), or an anaplerotic function by supplying intermediates to the mitochondrial electron transport chain, supporting plant metabolism and flowering [12,14,15].
In R. asiaticus, flowering and tuberization are antagonistic processes, and both undergo a crosstalk with thermo- and photo- period. Short day and vernalization promotes the meristematic activity, increasing the percentage of sprouting buds and the number of leaves and flowers [1]. Conversely, long day anticipates flowering of already formed buds, hence reducing the flowers yield and quality, while promoting the enlargement of tuberous roots [1,9]. Flowering earliness and flower yield and characteristics (i.e. stem length and bud size) vary widely with the plant genotype, the size of tuberous roots and the procedure of storing and preparation before planting, as well as the plant growing conditions [1]. Vernalization of tuberous roots is a common practice to schedule cut flower production of R. asiaticus, as in many ornamental geophytes [2]. However, it is known that the response to low temperature is genotype-dependent and changes among the hybrids, and data on the effects of cold treatments on the primary and secondary metabolites and the mechanisms underlying the plant response to vernalization and, more in general, about plant physiology and metabolism in this species, are quite scarce.
As reported in Carillo et al. [16], in geophytes the tuberous roots act as storage organs, and in particular they synthetise starch. Sometimes in the roots this sugar is in combination with other sugars-based polymers such as glucomannan and fructans that can help or replace starch [17] to overcome prolonged adverse environmental conditions.
Due to the importance of their role as a 'reservoir', it is important to gain new knowledge not only on growth but also on the metabolism changes of root tubers, focusing on the source-sink relationships between storage organs and aerial part of the plant from the rest period until sprouting and flowering to assess whether the newly formed young roots and reserves are going to be formed or reconstituted [18].
We carried out a series of experiments aiming at elucidating the influence of temperature [11,12] and photoperiod [16,19,20], and their interaction, on plant physiology and metabolism, and flowering process of R. asiaticus. In the present experiment, we investigated the influence of three preparation procedures of tuberous roots, including only rehydration and two times of exposure to cold temperature after rehydration, on plant growth, and metabolic profile in two hybrids with different flowering earliness grown in pot in an unheated glasshouse. We reported results on the responses observed in the aerial part in a previous article in this journal [12]. In the present paper, we show data on the growth and metabolism of tuberous roots that demonstrate a different response of the two hybrids depending on the genotype and sensitivity in two different vernalization procedures. This in-depth analysis in both storage organ and leaves contributes at unveiling the importance of root-shoot relationship in determining differences in plant behaviour and sensitivity to vernalization of tuberous roots among different genotypes, also useful in production scheduling of R. asiaticus L.
2. Results
2.1. Plant Growth and Flowering
Detailed results about growth, physiological and metabolic responses to vernalization procedures in the aerial part are reported in Fusco et al., 2023 [12]. Briefly, the number of leaves of plants from only rehydrated (C) tuberous roots and the leaf dry matter did not differ significantly in the two hybrids (23.2 leaves and 3.21 g per plant, on average, respectively), while the total leaf area was 37% greater in MDR than in MBO (883.6 vs. 646.5 cm2 per plant). Hybrid and root preparation interacted on plant leaf area, which decreased under the V4 vernalization only in MBO (−82%), and after both the procedures in MDR (−34% in V2 and −72% in V4), compared to Control. Accordingly, the leaf dry weight was reduced by the longer vernalization only in MBO (−90% than in Control) and both the treatments in MDR (−17% in V2 and −72% in V4) (Figure 1).
In Control plants, the flowering earliness (in days after sowing, DAS) was similar in the hybrids (110.2±5.3 DAS in MBO C and 92.5±3.2 DAS in MDR C). Compared to only rehydration, vernalization for 2 weeks (V2) did not change the time for flowering in MBO (118.5±4.0 DAS) and reduced it in MDR (65.5±5.6 DAS), while that for 4 weeks (V4) anticipated flowering in both the hybrids (84.7±11.2 DAS in MBO and 34.3±8.9 DAS in MDR). The cut flower characteristics (stem length and diameter, and dry weight) did not differ in the hybrids (58,5 cm, 5,26 mm, and 3,76 g, respectively) while they were influenced by the root preparation. Compared to Control, both the vernalization procedures determined thinner stems and reduced the dry weight (−42% and −66% in MBO, and −60% and −74% in MDR, in V2 and V4 respectively), and also the stem height in MDR (−17% and −33% after 2 and 4 weeks of vernalization).
The dry weight of tuberous roots was similar in plants of MBO and MDR (Figure 1). The vernalization treatment lasting 2 weeks determined a reduction in dry matter accumulation in root, which revealed to be significant compared to Control only in MDR, while the procedure of 4 weeks caused a significant effect in both the hybrids.
2.2. Tuberous roots’ Metabolic Profile
The hybrid (H), vernalization (V) and “Hybrid × Vernalization procedure” (H × V), significantly affected the metabolites in two hybrids of Ranunculus asiaticus L., MDR and MBO, in the three different growth stages, pre-germination, vegetative and flowering phase. In particular, the content of glucose was significantly affected by H in pre-germinative and flowering phases. In the pre-germinative phase and flowering phases, it was higher in MDR (+19,3% and + 113%, respectively) than in MBO (Table 1). This parameter was significantly affected by H x V in all growth phases. In the vegetative phase, the glucose content underwent a drastic decrease in MDR V4 (- 51%) and in MBO V2 (-54%) compared to respective controls (Table 1); while at flowering it was affected both by H x V and V. In fact, as showed in Table 1, glucose increased in MBO V2 (+54,5%) while decreased in both hybrids MDR MBO under V4 (-47,8% and -92,9%, respectively), compared to respective control.
H x V and V influenced the fructose content in the vegetative stage; while at flowering this sugar was affected only by H x V. Specifically, in vegetative phase the content of this sugar significantly increased only in MDR V4 compared to the other two MDR treatments (+70% on average). In MBO, the length of vernalization procedure (V) determined contrasting effects decreasing fructose of 66% in MBO V2, while increasing it of 239% in MBO V4 compared to control (Table 1). On the contrary, at flowering, there was a decrease of fructose content in MBO V4 (-96%) compared to control.
In the pre-germinative phase the sucrose content was slightly higher in MDR than in MBO (on avg. 8.63 mg g−1 DW and 8.28 mg g−1 DW, respectively). Instead, the vegetative and flowering phases were significantly affected by H x V and only vegetative phase also by V. Particularly, in the former phase, the sucrose content increased in both MDR V4 and MBO V4 (+55% and +271%, respectively) while decreased of 77% only in MBO V2, compared to respective controls (Table 1). At flowering, only MBO V4 significantly decreased (-96%) compared to control.
Starch was affected by H in pre-germinative and flowering stage. In fact, in both these growth stages in fact, this parameter was lower in MDR (on avg. 39.55 and 38.42 mg g−1 DW, respectively) than in MBO (on avg. 52.86 and 53.75 mg g−1 DW). The starch content was also affected by H x V in all growth stages (Table 1).
The polyphenols content was affected by H, V and H x V. In particular, in the vegetative and flowering stages, it was higher in MDR (on avg. 12.64 and 10.13 µg mg−1 DW, respectively) than in MBO (on avg. 8.01 and 6.01 µg mg−1 DW, respectively). In the vegetative phase, polyphenols increased in MDR V2 (+15%) while decreased in MDR V4 (-43%) compared to control. At flowering polyphenols increased in MDR V2 and V4 (+119% and 99,8%, respectively) but decreased in MBO V2 and V4 (-56% and -77%, respectively), compared to their respective controls (Table 1).
The soluble proteins content was affected by H x V at vegetative and flowering stage. At vegetative stage, itr was higher in MDR (+66%) than in MBO (Table 2). Specifically, MDR V4 and MBO V2 increased (+46% and +95%, respectively) compared to respective controls. At flowering proteins content in MDR increased from 5.85 µmol g−1 (control) to 19.04 and 31.94 µmol g−1 (MDR V2 and V4, respectively). As showed in Table 2, this parameter decreased in MBO from 29.51 µmol g−1 (control) to 20.97 and 14.68 µmol g−1 (MBO V2 and V4, respectively).
Amino acids were mostly affected by V and H x V and less by H. At pre-germinative stage, the total amino acids content was strongly influenced by V; in fact, they increased in both hybrids under V4 treatment (+147% and +205%, respectively), compared to controls (Table 2). The total amino acids content decreased during the vegetative stage under the two vernalization procedures (V2 and V4) both in MDR (-65% and -71%, respectively) and MBO (-23% and -61%, respectively) compared to respective controls. At flowering there was a strong decrease (-95%) of the total amino acids content but only in MBO V4 (-95%) compared to control (Table 2).
Alanine, GABA, glutamate, proline and serine were the amino acids undergoing the main changes in the vernalized MDR hybrid. In fact, these amino acids increased both in MDR V2 (+387%, +57%, +49%, +78% and +44%, respectively) and MDR V4 (+806%, 1247%, 162%, +103% and +171%, respectively) compared to respective controls. Asparagine, glutamine, glycine, monoethanolamine (MEA), ornithine and threonine increased only in MDR V4 of 166%, 54%, 235%, 91%, 158% and 129%, respectively, compared to controls. While these amino acids decreased in vegetative and at flowering stage. Specifically, in both this growth stage GABA and glutamate showed a decreased in MDR V2 (-67% and -62%, respectively) and in MDR V4 (-88% and -35%, respectively) in vegetative stage; while at flowering GABA decreased of 74% in MDR V2 and of 58% in MDR V2, and glutamate of 61% in MDR V2 and V4, compared to controls (Table 2).
In the vegetative stage asparagine, serine and threonine, decreased in MDR V2 of 74%, 46% and 83%, respectively, and in MDR V4 of 77%, 81% and 29%, respectively, compared to controls. Alanine, glutamine, glycine, MEA, ornithine and proline showed a decrease only in MDR V4 of 68%, 61%, 57%, 57%, 30% and 58%, respectively, compared to their controls. On the contrary, proline increased (+81%) in MDR V4 at flowering stage (Table 2). In the flowering stage, aspartate, glycine and MEA decreased in MDR V2 of 40%, 41%, and 58% compared to respective controls; while in MDR V4 ornithine decreased of 49% compared to control. Regarding the hybrid MBO, during the pre-germinative stage, alanine, asparagine, GABA, glutamate, glycine, MEA, ornithine, proline and serine increased in MBO V2 (+237%, +56%, +348%, +79%, +200%, +120%, +74% and +90%, respectively) and also in MBO V4 (+404%, +173%, +889%, +94%, +590%, +226%, +192% and +161%, respectively) compared to controls (Table 2). Also, glutamine and proline increased only in MBO V4 of 151% and 104% compared to controls. In addition, proline increased in MBO V4 of 80% in the vegetative stage; and also, at flowering in MBO V2 of 188%, compared to respective controls. Alanine, GABA, ornithine and threonine showed an increase of their content also at flowering stage in MBO V2 (+197%, +214%, +108% and +51%, respectively) compared to controls (Table 2). Except for proline that remained unchanged (MBO V2) or increased (MBO V4, +104%), in vegetative stage, alanine, MEA, ornithine, serine and threonine decreased in MBO V2 of 48%, 62%, 46%, 61% and 83%, respectively, and in MBO V4 of 29%, 71%, 51%, 53% and 81%, respectively, compared to controls; while aspartate, GABA and glutamate showed a decrease only in MBO V2 (-45%, -34% and -40%, respectively) and asparagine, glutamine and glycine only in MBO V4 (-69%, -59% and -44%, respectively), compared to their controls. At flowering, except for glycine that decreased in both MBO V2 and V4 (-48% and -82%, respectively), alanine, asparagine, aspartate, GABA, glutamine, glutamate and serine decreased only in MBO V2 of 87%, 99%, 98%, 93%, 99%, 90% and 96%, compared to respective controls.
The minor amino acids content (including arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, tyrosine, tryptophan, and valine) was influenced by V and H x V. In fact, they increased in the pre-germination stage, and in particular in both MDR and MBO hybrids under V4 (+136% and +230%, respectively). Whereas, minor amino acids decreased in the vegetative stage in both hybrids and in both vernalization procedures, MDR V2 and V4 (-52%, -64%, respectively) and in MBO V2 and V4 (-79% and -68%, respectively) compared to controls; but they decreased only in MDR V2 (-33%) respect to control at flowering stage (Table 2).The branched chain amino acids (including isoleucine, leucine and valine; BCCAs) showed a variation at all growth stages. At the pre-germination phase, the BCAAs increased in both MDR V4 (+201%) and MBO V4 (291%) compared to controls. On the contrary, these amino acids showed a decrease at the vegetative stage in MDR V2 (-40%), MDR V4 (-64%), MBO V2 (-67%) and MBO V4 (-52%), and only in MBO V4 (-78%) at flowering stage, compared to respective controls (Table 2).
2.3. Principal Component Analysis (PCA)
A principal component analysis (Figure 2) was performed on all metabolites data obtained by the analyses of root tissues of the Ranunculus asiaticus L. MDR and MBO hybrids, undergoing three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the pre-planting, vegetative phase and flowering growth phases. The variables in the first four principal components (PCs) were highly correlated, with eigen values greater than 1, thus explaining for 83.3% of the total variance, with PC1, PC2, PC3 and PC4 accounting for 46.5%, 22.1%, 8.7% and 6.0%, respectively. PC1 was positively correlated to BCAAs, serine, alanine, glycine, GABA, threonine, glutamate, glutamine and proline. Whereas PC1 was negatively correlated to polyphenols and aspartate. PC2 was positively correlated to MEA and asparagine; while it was negatively correlated to starch, fructose, and sucrose. The two hybrids in the pre-germinative phases were well separated along PC1, with MDR V2 and V4 close to the X axis in the positive side of PC1, while the samples of the flowering phase were in the negative side of PC1 but for the most closed to axes intercept. The MDR samples in the pre-germinative phase were clustered in the negative side of PC2, while those of the vegetative stage, and in particular MDR C, were clustered in the positive side of PC2.
3. Discussion
In this experiment, plants of the hybrids MBO (early flowering) and MDR (medium earliness) of Ranunculus asiaticus L. were subjected to three preparation procedures of tuberous roots and grown in unheated glasshouse in the South of Italy, from September to March. In our environmental and cultural conditions, flowering of Control plants from only rehydrated roots started at the beginning of January and the time for flowering was similar in the hybrids, despite the different earliness expected, and flower stem characteristics were comparable, as previously observed on the same genotypes subjected to the same preparation [20].
Vernalization for 2 weeks at 3.5 °C (V2), anticipated flowering in MDR (-27 days compared to Control) and was ineffective in MBO, and that for 4 weeks (V4) promoted flowering in both the hybrids (26 days in MBO and 58 days in MDR) and reduced the quality of flower stems in both the hybrids, similarly to previous experiments on the same genotypes [16].
In Italy, ranunculus is mostly prepared through rehydration followed by vernalization, with planting from end of August - beginning of September, and harvest from the end of November to April. In different R. asiaticus hybrids, the exposure of tuberous roots to 3.5 °C for 2 weeks after rehydration (as in our V2 treatment), anticipated flowering [1]. However, in our experiment this procedure, which is the most common in breeding farms, was effective in MDR but ineffective or detrimental in MBO, confirming that cold requirements are hybrid-specific [1,21]. Vernalization reduced the fresh weight in both the hybrids and also the stem height in MDR, with stronger effect after 4 weeks of treatment. Ohkawa studied the influence of two vernalization procedures, 5 °C for 2 weeks and for 4 weeks, compared to only rehydration, in 2 cultivars of R. asiaticus, and found a positive relationship between flowering anticipation and the duration of cold treatment in one genotype, as we observed in MDR, and a negative influence of the longer treatment on flowering earliness and quality in the other [21].
The MDR and MBO tuberous roots metabolic profile under the three treatments and in the three different growth phases, showed a significant interaction between genotype and vernalization procedure in relation to the content of amino acids, soluble sugars and polyphenols, as previously reported by Fusco et al. [12] in leaves. The first and main change at the pre-germinative phase in the two hybrids, under V2 and V4 vernalization treatments, was the stronger increase in γ-aminobutyric acid (GABA), alanine and proline in MDR compared to MBO, even if the absolute values of these amino acids were higher in MBO than in MDR. Moreover, MDR, unlike MBO, did not show a further consistent increase of all other amino acids or metabolites in the root tissues previously vernalized. Indeed, the strong and fast increase of GABA, proline and alanine together is a clear symptom of stress for the plant tuberous root. Clearly, vernalization-related cold stress may cause dysregulation of pH, oxidative stress and metabolic disfunction [23] particularly in plants sensitive to cold stress as already proven for MDR [12]. The increase of GABA, alanine and proline in the pre-germinative phase may alleviate the cold stress effects by buffering the cytoplasmic acidosis and potentiating the antioxidant defense [15,22]. In fact, the synthesis of both alanine and GABA involve proton-consuming reactions thus regulating cytosolic pH. In particular, alanine is synthetized through decarboxylation of malate to pyruvate by malic enzyme activity and then transamination of pyruvate to alanine; while GABA is obtained through glutamate decarboxylation catalyzed by glutamate decarboxylase [15,24,25]. In its turn, proline is not only well known for its ability to act as osmolyte, but it may detoxify ROS, buffer cellular redox potential and protect cellular structures for oxidative stress [14]. In addition, also GABA is able to scavenge ROS (e.g., hydrogen peroxide, singlet oxygen and superoxide anion radicals) like or even better than proline at similar concentrations [26,27]. Molina-Rueda et al. [27] reported that GABA and proline are often synthetized and accumulated at the same time in response to environmental stresses to protect the plant tissues from oxidative stress. Finally, after the relief from cold stress, alanine, GABA and proline can be metabolized and/or converted to intermediates of Krebs cycle and used to produce alpha-keto acids and ATP, thus re-activating plant metabolism and accelerating flowering [16,28]. Probably the initial higher capacity to synthesize and accumulate alanine and GABA, in addition to branched chain amino acids (BCAAs), in control conditions preserved MBO from the cold stress symptoms determining its lower sensitivity to shorter term applied cold stress (V2 treatment). It has previously suggested [16] that BCAAs in ranunculus may work as ROS scavengers reducing the onset of oxidative damages, in addition to act as alternative electron donors for the mitochondrial electron transport chain.
At pre-germinative stage, the content of amino acids, in particular asparagine, in the tuberous roots of MDR V4 and MBO V4 and also in MBO V2 increased. Their accumulation it is possibly due not only to the hydrolysis of root proteins, but also to the import of asparagine from the leaves. Asparagine, in fact, plays a key role in nitrogen transport because of its N:C ratio of 2:4, and can be rapidly deaminated producing aspartate and then oxaloacetic acid that can be used in the citric acid cycle or for gluconeogenesis. Carbon skeletons and ATP may be in fact necessary to produce polyphenols and MEA for continuing also in the vegetative phase to repair the oxidative damages caused by the vernalization procedure. Zhou et al. [29] found that tobacco plants subjected to 4 °C, similarly to Ranunculus, showed an increase in the content of various metabolites that fall into the class of polyphenols. This was considered a positive self-protection mechanism, able to increase the strength and rigidity of the cell wall, and the water transport in the vascular system, compromised by low temperature. However, this is a high energy consuming process which diverts intermediates and energy from growth, thus causing lower root and stem dry weight, thinner and shorter stems and early flowering. Also, the increase of MEA in MDR V4 and MBO V2 and V4, obtained for decarboxylation of serine, may play an important role for the synthesis and/or regeneration of phospholipids [16].
Differently from MDR, MBO started to accumulate free sugars, starch and proteins in tuberous roots already during vegetative stage, and particularly under V2 and V4 treatments, showing that in this hybrid the shorter vernalization activated the storage of surplus reserves even before flowering without affecting the stem dimension and/or the time of flowering. In MBO V4 at vegetative stage high amounts of fructose and sucrose were synthetised. Sucrose is a storage carbohydrate that can be rapidly mobilized according to metabolic needs and rapidly loaded into the translocation stream [30]. In fact, it was probably exported to leaves where it could anticipate flowering; in fact, the increase in sucrose is correlated with the flowering in several species [31]. Furthermore, this metabolite may also be a compatible compound able to stabilize cell membranes and maintain turgidity [32].
4. Materials and Methods
The experiment was carried out at the Department of Agriculture of the University of Naples (Portici, Italy—40°49′ N, 14°20′ E), from 18 September 2018 to 30 March 2019.
Dry tuberous roots of two hybrids of Ranunculus asiaticus L., MBO (early flowering) and MDR (medium earliness) (Biancheri Creazioni, Italy, https://www.bianchericreazioni.it/ accessed on 1 October 2022) were subjected to three preparation procedures:
- -
- only rehydration: exposure to 12 °C for 24 h in humid chamber (Control, C);
- -
- rehydration followed by vernalization at 3.5 °C for 2 weeks (V2);
- -
- rehydration followed by vernalization at 3.5 °C for 4 weeks (V4).
Tuberous roots of the most common size for each hybrid were used (3–4 cm for MBO and 4–5 cm for MDR). Plants were grown in pot, on a mixture of perlite and peat (70:30 in vol.). Irrigation was alternated with fertigation (4 pulses per week in total). In the nutrient solution Hoagland full strength, pH and electrical conductivity (EC) were kept at 5.5 and 1.7 dS/m, respectively, and monitored with a portable pH-EC sensor (HI9813 series, Hanna Instruments Intl.). The mean values of air temperature and relative humidity (day/night) recorded during the experiment were 23.7±5.0/12.30±4.1 °C and 58.5 ± 6.8/74.3 ± 16.9%, respectively (Mean Value ± Standard Deviation).
4.1. Plant Growth and Flowering
The number of leaves per plant was monitored weekly. Plant leaf area was estimated by non-destructive analysis of digital images of leaves with ImageJ software 1,50i version (Wayne Rasband National Institute of Health, USA), on 3 leaves per plant, on 5 plants per combination “Hybrid x Vernalization procedure”. The results obtained in fully grown plants at vegetative state (11 weeks after planting) are reported. The time for flowering was calculated as average of the number of days to obtain visible flower buds in 25 plants per treatment.
4.2. Metabolic Profile
4.2.1. Starch and Soluble Carbohydrate Analysis
Starch and soluble sugars were determined according to Fusco et al. [12] with some modifications. 10 mg of lyophilized tuberous roots were suspended twice in 140 µL ethanol 80% (v:v) and once in 70 µL ethanol 50% (v:v) at 80 °C in a thermomixer (Eppendorf ThermoMixer® C) for 20 minutes, and then centrifuged at 14000 rpm for 10 minutes at 4 °C. The clear supernatants, separated from pellets, were combined together and stored in 1.5 mL tubes at −20 °C until analysis of glucose, fructose and sucrose. The pellets of ethanolic extraction were extracted with 500 μL of 0.1 M KOH and heated at 90 °C for 2 h [33] cooled in ice and then acidified to pH 4.5 with acetic acid. An aliquot of acidified samples was mixed with sodium acetate 50 mM pH 4.8, α-amylase 2 U/mL and amyloglucosidase 20 U/mL and incubated at 37 °C for 18 h for enzymatic hydrolisis. The samples were centrifuged at 14000 rpm for 10 minutes at 4 °C, and the supernatant containing the glucose derived from starch hydrolysis was used for measurement. The content of soluble sugars in the ethanolic extracts, and the glucose derived by starch hydrolysis, were determined by an enzymatic assay coupled with reduction in pyridine nucleotides and the increase in absorbance at 340 nm was recorded using a Synergy HT spectrophotometer (BioTEK Instruments, Bad Friedrichshall, Germany). The content of sugars was expressed as mg g−1 DW.
4.2.2. Soluble Proteins, Free Amino Acid Analysis
Soluble proteins (mg g-1 DW) were extracted from 10 mg of lyophilized samples with a buffer containing 200 mM TRIS-HCl pH 7.5 and 500 mM MgCl2. Then the samples were vortexed and stored at 4 °C for 24 h. After 24h the samples were centrifuged at 14000 rpm for 10 minutes at 4 °C. The clear supernatants (10 µL) were added to 190 µL of Bio-Rad protein assay dye reagent diluted 1:5 (v:v) with bidistilled water [34]. The solutions were mixed and then absorbance at 595 nm was recorded on a microplate reader (Synergy HT, Bio-Tek, Germany). The soluble protein content in the samples was calculated by comparison with standard curves obtained using known concentrations of bovine serum albumin (BSA) as the reference standard as reported in Carillo et al. [35]. Free amino acids and proline were determined by the method described by Woodrow et al. [14]. Aliquots of about 10 mg of lyophilized samples were extracted in 1 mL ethanol:water (40:60 v:v) overnight at 4 °C. After 24h the samples were centrifuged at 14000 rpm at 4 °C for 10 minutes. Free amino acids were determined by high-performance liquid chromatography (HPLC) after precolumn derivatization with ophthaldialdehyde (OPA) according to the method described in Carillo et al. [36]. Proline was determined in the same ethanolic extract used for the amino acids determination by an acid ninhydrin colorimetric method according to Woodrow et al. [14].
4.2.3. Polyphenols Analysis
Polyphenols (mg GAE g−1 DW) were determinated according to Singleton et al. [37] with some modifications. 30 mg of lyophilized tuberous roots samples ware extracted in 700 μL of 60% methanol (v:v), and vortexed. Then the samples were centrifuged at 1000 rpm for 10 minutes at 25 °C. An aliquot of clear supernatant (35 μL) was mixed with 125 μL of the Folin Ciocalteu reagent diluted 1:4 (v:v) with bidistilled water and after mixing for 6 min at room temperature. After 6 minutes, 650 μL of 3% (v:v) sodium carbonate was added. After 90 min at room temperature, the absorbance at 760 nm was determined in a microplate reader (Synergy HT, BioTEK Instruments). Polyphenols concentration was determined against standard curves of gallic acid as described in Carillo et al. [35].
5. Statistical Analysis
The experiment was conducted on 25 plants per combination “Hybrid x Vernalization procedures”. Data were subjected to statistical analysis by using SigmaPlot 12.0 (SPSS Inc. Norman Nie Dale Bent, Hadlai "Tex" Hull, Chicago, IL) software package. The main effect of the categorical independent factors (i.e., hybrids and preparation procedures) and their interaction on the continuous dependent variables were analysed through the two-way ANOVA. In case of rejection of the null hypothesis, the Tukey’s HSD test was performed (p ≤0.05).
6. Conclusions
In conclusion, the two hybrids showed different responses to the root preparation procedure. The MBO hybrid, being able to synthetise higher constitutive amounts of protecting metabolites like GABA and alanine, better overcoming the negative effects of vernalization, at least when it was applied for shorter time (V2). On the contrary, the MDR hybrid resulted more sensitive to vernalization decreasing its capacity to export nutrients to leaves for the need to rapidly synthetise high energy cost organic osmolytes for comping with dysregulation of pH, oxidative stress and metabolic disfunction caused by cold stress. Moreover, the MDR need to continue synthetising protecting metabolites like polyphenols also in roots during the vegetative stage, contributing to subtract carbon intermediates and ATP from growth to contrast the oxidative effects due to vernalization and the consequent metabolic imbalance. Indeed, the new knowledge about the metabolic responses of the hybrids to vernalization may be directly translated in field and used for implementing the best hybrid-specific preparation procedures.
Author Contributions
R.P. and SDP proposed the experiment; G.C.M. carried out the plant cultivation; G.M.F., R.N. and L.P. performed the metabolic analyses. R.P., G.M.F. and P.C. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the PhD program in Agricultural and Agri-food Sciences University of Naples Federico II XXXII cycle and the University of Campania Luigi Vanvitelli (Grant No. VALERE: VAnviteLli pEr la RicErca).
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data are reported within the article.
Acknowledgments
The authors thank Biancheri Creazioni, Italy (https://www.bianchericreazioni.it/ accessed on 1 October 2022) and Mario Rabaglio for propagation material and technical assistance, and Raffaele Garramone (University of Naples Federico II) for his help in setting up the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ABA, abscisic acid; BCAAs, branched-chain amino acids; DW, dry weight; DAS, Days after sowing; GA, gibberellic acid; GABA, γ-aminobutyric acid; H, hybrid; HxV, ‘Hybrid x Vernalization procedure’; JA, jasmonic acid; MEA, monoethanolamine; PCs, principal components; ROS, reactive oxygen species; V, vernalization.
References
- Beruto, M.; Rabaglio, M.; Viglione, S.; Van Labeke, M.-C.; Dhooghe, E. Ranunculus. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 649–671. [Google Scholar] [CrossRef]
- Le Nard, M. and De Hertogh, A. In The physiology of flower bulbs, Chapter 4 in Bulb Growth and Development and Flowering; De Hertogh, A.A.; Le Nard, M., Eds.; Elsevier: Amsterdam, NL, USA, 1993.
- Meynet, J. Ranunculus. In The physiology of Flower Bulbs: A Comprehensive Treatise on the Physiology and Utilization of Ornamental Flowering Bulbous and Tuberous Plants; De Hertogh, A.A., Le Nard, M., Eds.; Elsevier: Amsterdam, NL, USA. 1993; pp. 603–610. [Google Scholar]
- Kamenetsky, R. Production of Flower Bulbs in Regions with Warm Climates; Okubo, H., Miller, W.B., Chastagner, G.A., Eds.; International Society for Horticultural Science: Niigata, Japan, 2005; Volume 2, pp. 59–66. [Google Scholar]
- Beruto, M.; Fibiani, M.; Rinino, S.; Scalzo, R.L.; Curir, P. Plant development of Ranunculus asiaticus L. tuberous roots is affected by different temperature and oxygen conditions during storage period. Isr. J. Plant Sci. 2009, 57, 377–388. [Google Scholar] [CrossRef]
- Glier, J.H.; Caruso, J.L. Low-temperature induction of starch degradation in roots of a biennial weed. Cryobiology 1973, 10, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Su, X.; Tian, Z.; Liu, Y.; Zhou, Y.; He, M. Transcriptome profiling provides insights into dormancy release during cold storage of Lilium pumilum. BMC genomics. 2018, 19(1), 1–17. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, H.; Wu, G.; Sun, J.; Hao, L.; Ge, X.; Yu, J.; Wang, W. Sequence variation and expression analysis of seed dormancy-and germination-associated ABA-and GA-related genes in rice cultivars. Frontiers in plant science. 2011, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.M. A review of plants’ flowering physiology: The control of floral induction by juvenility, temperature and photoperiod in annual and ornamental crops. Asian J. Agric. Food Sci. 2014, 2, 186–195. [Google Scholar]
- Zhao, L.; Li, S.; Yu, Q.; Zhang, C.; Wang, L.; Jiang, Y.; Wu, Z.; Pi, Z. Vernalization Promotes GA-Mediated Bolting Initiation via the Inhibition of ABA and JA Biosynthesis. Agronomy. 2023, 13(5), 1251. [Google Scholar] [CrossRef]
- Carillo, P.; Arena, C.; Modarelli, G.C.; De Pascale, S.; Paradiso, R. Photosynthesis in Ranunculus asiaticus L.: the influence of the hybrid and the preparation procedure of tuberous roots. Front. Plant Sci. 2019, 10, 241. [Google Scholar] [CrossRef]
- Fusco, G.M.; Carillo, P.; Nicastro, R.; Modarelli, G.C.; Arena, C.; De Pascale, S.; Paradiso, R. Vernalization Procedure of Tuberous Roots Affects Growth, Photosynthesis and Metabolic Profile of Ranunculus asiaticus L. Plants. 2023, 12(3), 425. [Google Scholar] [CrossRef]
- Fritz, C.; Palacios-Rojas, N.; Feil, R.; Stitt, M. Regulation of secondary metabolism by the carbon–nitrogen status in tobacco: nitrate inhibits large sectors of phenylpropanoid metabolism. The Plant Journal 2006, 46, 533–548. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D'Amelia, L.; Dell'Aversana, E.; Piccolella, S.; Fuggi, A. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Carillo, P. GABA shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Carillo, P.; Dell’Aversana, E.; Modarelli, G.C.; Fusco, G.M.; De Pascale, S.; Paradiso, R. Metabolic profile and performance responses of Ranunculus asiaticus L. hybrids as affected by light quality of photoperiodic lighting. Frontiers in Plant Science. 2020, 11, 597823. [Google Scholar] [CrossRef] [PubMed]
- Ranwala, A.P.; Miller, W.B. Analysis of nonstructural carbohydrates in storage organs of 30 ornamental geophytes by high performance anion exchange chromatography with pulsed amperometric detection. New Phytol. 2008, 180, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Kamenetsky, R. Production of Flower Bulbs in Regions with Warm Climates; International Society for Horticultural Science, Eds.: H. Okubo, W.B. Miller and G.A. Chastagner, Niigata, Japan, 2005; Volume 2, pp. 59–66. [CrossRef]
- Modarelli, G.C.; Arena, C.; Pesce, G.; Dell'Aversana, E.; Fusco, G.M.; Carillo, P.; De Pascale, S.; Paradiso, R. The role of light quality of photoperiodic lighting on photosynthesis, flowering and metabolic profiling in Ranunculus asiaticus L. Physiologia plantarum, 2020, 170(2), 187-201. [CrossRef]
- Carillo, P.; Modarelli, G.C.; Fusco, G.M.; Dell’Aversana, E.; Arena, C.; De Pascale, S.; Paradiso, R. Light spectral composition affects metabolic response and flowering in non-vernalized Ranunculus asiaticus L. Environmental and Experimental Botany. 2021, 192, 104649. [Google Scholar] [CrossRef]
- Ohkawa, K. Growth and Flowering of Ranunculus Asiaticus; Doss, R.P., Byther, R.S., Chastagner, G.A., Eds.; Acta Horticolturae: Seattle, WA, USA, 1986; Volume 1, pp. 165–172. [CrossRef]
- Bouche, N. & Fromm, H. GABA in plants: just a metabolite? Trends in plant science, 2004, 9(3), 110-115. [CrossRef]
- Eom, S.H.; Ahn, M.A.; Kim, E.; Lee, H.J.; Lee, J.H.; Wi, S.H.; Kim, S.K.; Lim, J.H., Hyun, T.K. Plant response to cold Stress: Cold stress changes antioxidant metabolism in heading type Kimchi cabbage (Brassica rapa L. ssp. Pekinensis). Antioxidants, 2022, 11(4), 700. [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; De Pascale, S.; Rouphael, Y. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy, 2019, 9(5), 249. [CrossRef]
- Van Oosten, M.J.; Dell’Aversana, E.; Ruggiero, A.; Cirillo, V.; Gibon, Y.; Woodrow, P.; Maggio, A.; Carillo, P. Omeprazole treatment enhances nitrogen use efficiency through increased nitrogen uptake and assimilation in corn. Frontiers in Plant Science, 2019, 10, 1507. [CrossRef]
- Liu, C.; Zhao, L.; Yu, G. The dominant glutamic acid metabolic flux to produce γ-amino butyric acid over proline in Nicotiana tabacum leaves under water stress relates to its significant role in antioxidant activity. Journal of Integrative Plant Biology, 2011, 53(8), 608-618. [CrossRef]
- Molina-Rueda, J.J.; Pascual, M.B.; Pissarra, J.; Gallardo, F. A putative role for γ-aminobutyric acid (GABA) in vascular development in pine seedlings. Planta, 2015, 241, 257–267. [Google Scholar] [CrossRef]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: accumulation of proline and glycine betaine. Functional Plant Biology, 2008, 35(5), 412-426. [CrossRef]
- Zhou, Z.; Xu, Z.; Feng, Q.; Yao, D.; Yu, J.; Wang, D.; Lv, S.; Liu, Y.; Zhou, N.; Zhong, M.E. Effect of pyrolysis condition on the adsorption mechanism of lead, cadmium and copper on tobacco stem biochar. Journal of Cleaner Production, 2018, 187, 996–1005. [Google Scholar] [CrossRef]
- Pollock, C.J. and Lloyd, E.J. The effect of low temperature upon starch, sucrose and fructan synthesis in leaves. Ann. Bot. 1987, 60: 231-235. [CrossRef]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, M.A.S.; Babar, M. Role of sugars, amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef]
- Jouve, L.; Hoffmann, L.; Hausman, J.F. Polyamine, carbohydrate, and proline content changes during salt stress exposure of aspen (Populus tremula L.): involvement of oxidation and osmoregulation metabolism. Plant Biol. 2004, 6: 74-80. [CrossRef]
- Salbitani, G.; Carillo, P.; Di Martino, C.; Bolinesi, F.; Mangoni, O.; Loreto, F.; Carfagna, S. Microalgae cross-fertilization: Shortterm effects of Galdieria phlegrea extract on growth, photosynthesis and enzyme activity of Chlorella sorokiniana cells. J. Appl. Phycol. 2022, 34, 1957–1966. [Google Scholar] [CrossRef]
- Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; De Pascale, S.; Rouphael, Y. Sensory and functional quality characterization of protected designation of origin ‘Piennolo del Vesuvio’ cherry tomato landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; Kyriacaou M.C.; Cardarelli, M.; Rouphael, Y. Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy, 2019, 9. [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R. M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
Figure 1.
Leaf and root dry weight (D.W.) and dry matter partitioning in Ranunculus asiaticus L. hybrids MBO and MDR obtained by three preparation procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), and grown in unheated glasshouse. Mean values ± Standard errors; n = 3. Re-elaborated from Fusco et al., 2023 [12].
Figure 1.
Leaf and root dry weight (D.W.) and dry matter partitioning in Ranunculus asiaticus L. hybrids MBO and MDR obtained by three preparation procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), and grown in unheated glasshouse. Mean values ± Standard errors; n = 3. Re-elaborated from Fusco et al., 2023 [12].
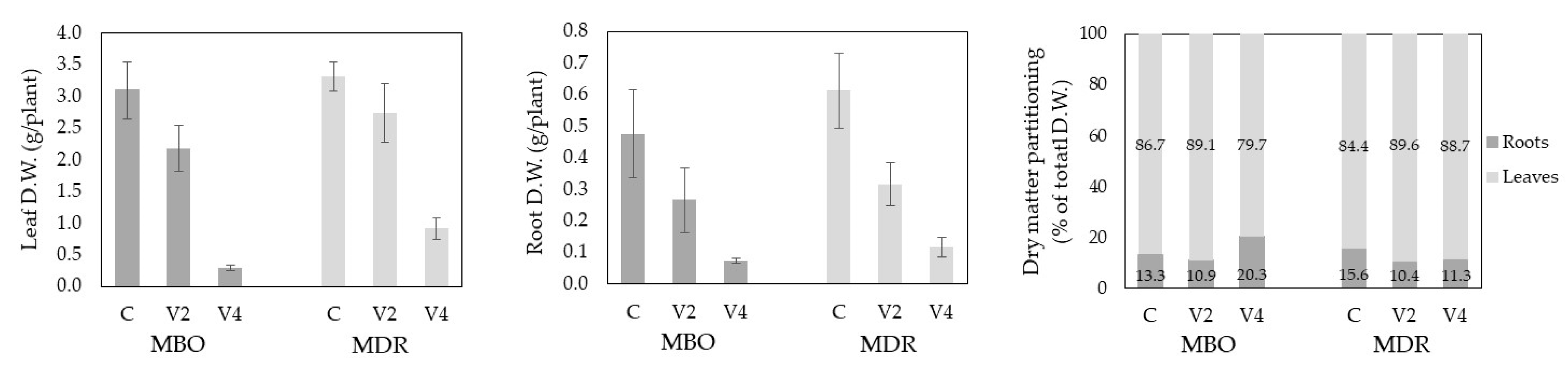
Figure 2.
Principal component loading plot and scores of principal component analysis (PCA) of carbohydrates, amino acids, polyphenols and soluble proteins in roots of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-germinative, vegetative phase and flowering).
Figure 2.
Principal component loading plot and scores of principal component analysis (PCA) of carbohydrates, amino acids, polyphenols and soluble proteins in roots of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-germinative, vegetative phase and flowering).
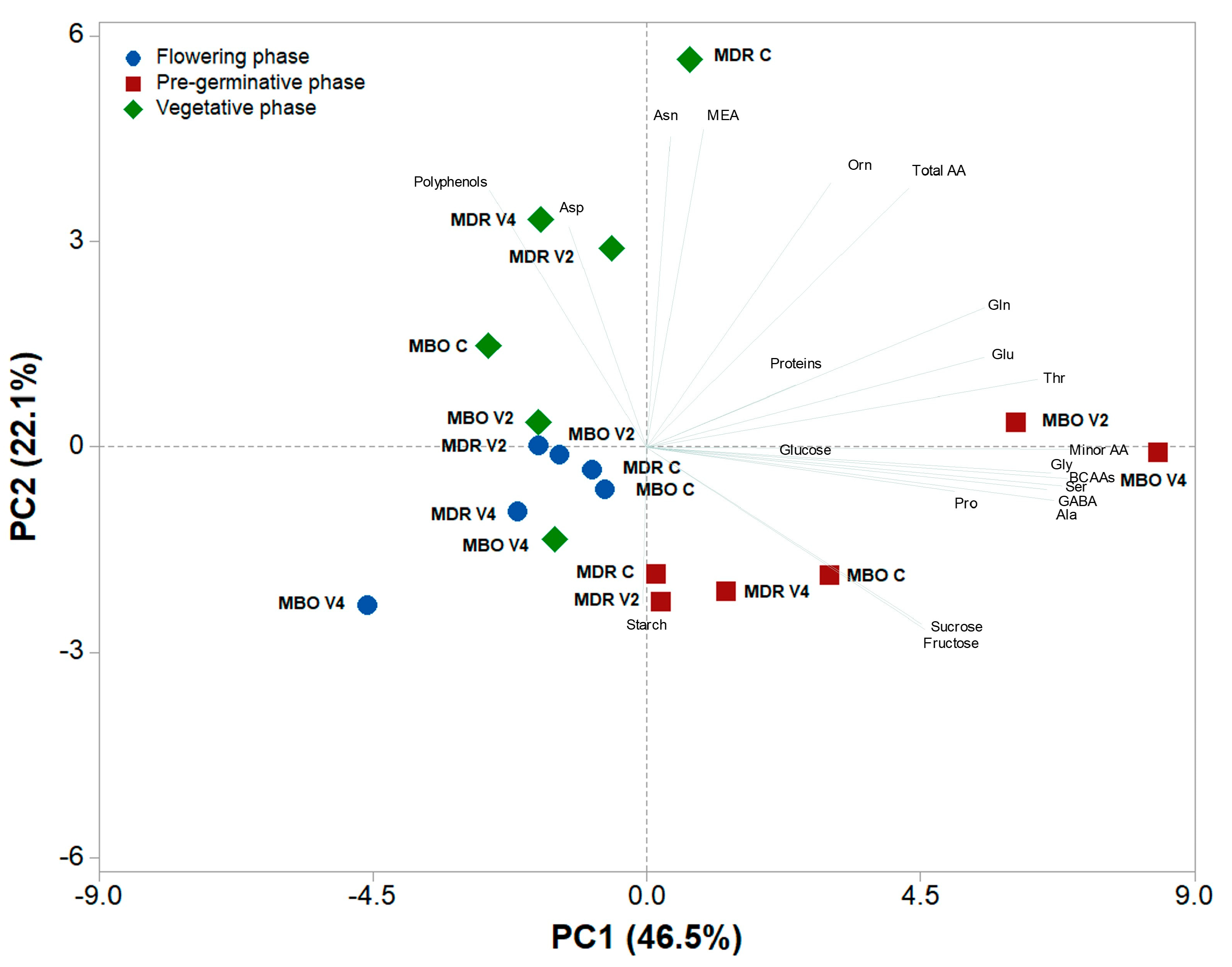

Table 1.
Polyphenols (in µg g−1 DW), glucose, fructose, sucrose and starch (in mg g−1 DW), in plants of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-planting, vegetative phase and flowering). ns, *, ** and ***; indicate non-significant or significant difference at p ≤ 0.05, p ≤ 0.01, p ≤ 0.001, respectively. Different lowercase or capital letters within each row, for specific vernalization procedure, indicate significant differences at p ≤ 0.05.
Table 1.
Polyphenols (in µg g−1 DW), glucose, fructose, sucrose and starch (in mg g−1 DW), in plants of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-planting, vegetative phase and flowering). ns, *, ** and ***; indicate non-significant or significant difference at p ≤ 0.05, p ≤ 0.01, p ≤ 0.001, respectively. Different lowercase or capital letters within each row, for specific vernalization procedure, indicate significant differences at p ≤ 0.05.
| MDR | MBO | H | V | H x V | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | ||||
| Pre-Germinative phase | |||||||||||
| Polyphenols | 4.05 ab | 3.30 a | 4.28 b | 3.88 | 3.67 ab | 3.61 a | 3.92 b | 3.73 | ns | * | ns |
| Glucose | 24.96 ab | 24.01 bc | 27.52 a | 25.5 A | 22.95 bd | 21.22 cd | 19.94 d | 21.37 B | *** | ns | ** |
| Fructose | 23.50 | 20.94 | 23.36 | 22.60 | 21.69 | 21.76 | 21.09 | 21.51 | ns | ns | ns |
| Sucrose | 8.78 | 8.35 | 8.76 | 8.63 A | 8.41 | 8.13 | 8.30 | 8.28 B | * | ns | ns |
| Starch | 41.05 a | 38.38 a | 39.21 a | 39.55 B | 54.14 bc | 59.82 b | 44.61 ac | 52.86 A | *** | ns | ** |
| Vegetative phase | |||||||||||
| Polyphenols | 13.97 a | 16.00 b | 7.94 cd | 12.64 A | 7.64 cd | 11.21 c | 5.16 d | 8.01 B | * | ** | ** |
| Glucose | 20.76 a | 21.47 a | 10.25 b | 17.49 | 18.97 a | 8.80 b | 22.47 a | 16.75 | ns | ns | *** |
| Fructose | 7.45 a | 6.17 a | 11.58 b | 8.40 | 6.62 a | 2.27 c | 22.50 d | 10.46 | ns | ** | *** |
| Sucrose | 2.86 a | 2.15 a | 4.43 b | 3.15 | 2.28 a | 0.52 c | 8.46 d | 3.76 | ns | ** | *** |
| Starch | 27.18 ab | 28.64 ab | 89.43 a | 48.41 | 27.42 ab | 20.95 b | 35.46 a | 27.94 | ns | ** | * |
| Flowering phase | |||||||||||
| Polyphenols | 5.85 a | 12.85 b | 11.69 b | 10.13 A | 10.82 b | 4.77 ac | 2.45 c | 6.01 | * | ns | *** |
| Glucose | 35.63 a | 31.04 a | 18.59 bc | 28.42 A | 15.26 c | 23.58 b | 1.08 d | 13.31 B | ** | * | *** |
| Fructose | 13.49 a | 13.89 a | 12.72 a | 13.36 | 10.63 a | 13.13 a | 0.42 b | 8.06 | ns | ns | *** |
| Sucrose | 5.79 a | 6.15 a | 4.78 a | 5.57 A | 2.12 b | 3.07 b | 0.09 c | 1.76 B | *** | ns | *** |
| Starch | 40.24 acd | 49.75 ab | 25.28 c | 38.42 B | 47.61 bd | 60.79 b | 52.83 b | 53.75 A | * | ns | * |
Table 2.
Soluble proteins (in mg g−1 DW) and free amino acids (in µmol g−1 DW) in plants of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-planting, vegetative phase and flowering). Amino acids (AA), γ-aminobutyric acid (GABA), monoethanolamine (MEA), branched-chain amino acids (BCAAs). ns, *, ** and ***; indicate non-significant or significant difference at p ≤ 0.05, p ≤ 0.01, p ≤ 0.001, respectively. Different lowercase or capital letters within each row, for specific vernalization procedure, indicate significant differences (p ≤ 0.05).
Table 2.
Soluble proteins (in mg g−1 DW) and free amino acids (in µmol g−1 DW) in plants of Ranunculus asiaticus L. hybrids MDR and MBO, obtained by three vernalization procedures of tuberous roots, only rehydration (Control, C), rehydration plus vernalization for 2 weeks (V2), rehydration plus vernalization for 4 weeks (V4), throughout the growing cycle, in relation to the different plant stages (pre-planting, vegetative phase and flowering). Amino acids (AA), γ-aminobutyric acid (GABA), monoethanolamine (MEA), branched-chain amino acids (BCAAs). ns, *, ** and ***; indicate non-significant or significant difference at p ≤ 0.05, p ≤ 0.01, p ≤ 0.001, respectively. Different lowercase or capital letters within each row, for specific vernalization procedure, indicate significant differences (p ≤ 0.05).
| MDR | MBO | H | V | H x V | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V2 | V4 | Mean | C | V2 | V4 | Mean | ||||
| Pre-Germinative phase | |||||||||||
| Soluble proteins | 36.49 | 32.6 | 35.78 | 34.96 | 42.33 | 33.45 | 41.56 | 39.11 | ns | ns | ns |
| Alanine | 4.29 a | 20.9 b | 38.87 c | 21.35 | 8.73 a | 29.39 b | 43.99 c | 27.37 | ns | *** | ** |
| Asparagine | 59.72 ac | 65.39 ac | 156.8 b | 83.97 | 32.24 a | 50.37 c | 88.12 c | 56.91 | ns | *** | ** |
| Aspartate | 2.34 ab | 1.64 ac | 5.72 b | 3.23 | 2.39 bc | 0.63 d | 2.22 b | 1.75 | ns | ** | * |
| GABA | 2.42 a | 16.28 b | 32.59 c | 17.10 | 6.63 a | 29.72 c | 65.57 d | 33.97 | ns | *** | *** |
| Glutamine | 67.21 a | 39.05 a | 105.8 b | 70.69 | 51.56 a | 60.59 a | 129.5 b | 80.55 | ns | *** | *** |
| Glutamate | 20.92 a | 31.17 abd | 54.86 c | 35.65 | 23.95 a | 42.79 cd | 46.53 bc | 37.76 | ns | *** | * |
| Glycine | 2.18 a | 3.47 ab | 7.31 c | 4.32 | 1.6 a | 4.8 b | 11.05 d | 5.82 | ns | *** | *** |
| MEA | 3.43 a | 3.07 a | 6.55 b | 4.35 | 2.62 a | 5.76 b | 8.55 c | 5.64 | ns | *** | *** |
| Ornithine | 0.9 a | 1.18 ab | 2.32 c | 1.47 | 0.96 a | 1.67 b | 2.8 c | 1.81 | ns | *** | *** |
| Proline | 5.59 a | 9.97 b | 11.36 b | 8.97 | 6.77 a | 6.83 a | 13.84 d | 9.15 | ns | *** | *** |
| Serine | 5.03 a | 7.23 a | 13.63 b | 8.63 B | 7.44 a | 14.13 b | 19.43 c | 13.67 A | * | *** | *** |
| Threonine | 8.92 a | 7.13 a | 20.44 b | 12.16 | 6.77 a | 9.00 a | 19.46 b | 11.74 | ns | *** | *** |
| Total AA | 228.5 a | 258.2 a | 564 b | 350.20 | 196.4 a | 324.8 a | 598.6 b | 373.30 | ns | *** | *** |
| Minor AA | 45.61 a | 51.75 a | 107.7 b | 68.35 | 44.73 a | 69.14 a | 147.5 c | 87.12 | ns | * | *** |
| BCAAs | 17.68 a | 22.88 a | 53.25 b | 31.27 | 18.19 a | 30.74 ab | 71.18 c | 40.04 | ns | *** | ** |
| Vegetative phase | |||||||||||
| Soluble Proteins | 36.27 a | 38.44 a | 53.03 b | 45.58 A | 20.11 c | 39.21 a | 23.15 c | 27.49 B | * | ns | *** |
| Alanine | 4.81 ab | 5.77 a | 1.53 c | 4.04 | 4.20 bc | 2.20 c | 3.00 c | 3.13 | ns | * | *** |
| Asparagine | 687.1 a | 175.2 bc | 157.8 bc | 340.0 | 221 b | 208.8 b | 69.41 c | 166.4 | ns | ** | *** |
| Aspartate | 5.34 ab | 7.57 a | 5.96 ac | 6.29 | 5.99 ac | 3.28 d | 4.95 bc | 4.74 | ns | ns | * |
| GABA | 5.53 a | 1.81 b | 0.64 c | 2.66 | 2.83 d | 1.86 b | 2.62 d | 2.44 | ns | * | *** |
| Glutamine | 87.61 a | 67.04 ab | 34.04 c | 62.90 | 64.00 b | 56.4 b | 26.26 c | 48.89 | ns | ** | *** |
| Glutamate | 48.11 a | 18.13 bd | 31.23 c | 32.49 A | 24.89 bc | 14.95 d | 29.69 b | 23.18 B | * | * | *** |
| Glycine | 1.25 a | 1.2 a | 0.54 b | 1 B | 2.03 c | 1.95 c | 1.14 a | 1.71 A | ** | * | *** |
| MEA | 16.12 a | 15.9 a | 7.01 b | 13.01 A | 15.67 a | 5.93 b | 4.58 b | 8.73 B | * | * | *** |
| Ornithine | 2.9 a | 2.85 a | 2.01 b | 2.59 | 3.37 a | 1.81 b | 1.65 b | 2.28 | ns | ** | *** |
| Proline | 5.97 a | 5.84 a | 2.53 b | 4.78 | 3.35 b | 3.44 b | 6.04 a | 4.28 | ns | ns | *** |
| Serine | 4.83 a | 2.63 b | 0.93 b | 2.80 | 5.74 a | 2.21 b | 2.67 b | 3.54 | ns | *** | * |
| Threonine | 12.17 a | 2.12 b | 8.64 c | 7.64 | 9.71 ac | 1.66 d | 1.8 bd | 4.39 | ns | ** | *** |
| Total AA | 922.2 a | 325.4 bc | 267.7 cd | 505.10 | 410.6 b | 314.5 c | 158.9 d | 294.70 | ns | ** | *** |
| Minor AA | 40.43 a | 19.35 b | 14.77 bd | 24.85 | 47.77 c | 10.01 d | 15.12 bd | 24.30 | ns | *** | *** |
| BCAAs | 11.63 a | 6.93 b | 4.16 c | 7.57 | 13.04 a | 4.3 c | 6.25 bc | 7.86 | ns | *** | *** |
| Flowering phase | |||||||||||
| Soluble Proteins | 5.85 a | 19.04 b | 31.94 c | 18.94 | 29.51 c | 20.97 b | 14.68 b | 21.72 | ns | ns | *** |
| Alanine | 6.91 a | 3.42 a | 5.06 a | 5.13 | 4.04 a | 12.01 b | 0.51 c | 5.52 | ns | ns | *** |
| Asparagine | 108.3 ab | 80.17 bc | 120.4 a | 103 A | 76.37 bc | 56.57 c | 0.02 d | 44.32 B | ** | ns | *** |
| Aspartate | 6.09 a | 3.64 bc | 5.27 ab | 5.00 | 5.22 ac | 6.33 a | 0.08 d | 3.88 | ns | ns | *** |
| GABA | 8.76 a | 2.31 bc | 3.72 bc | 4.93 | 3.93 b | 12.36 d | 0.26 c | 5.52 | ns | ns | *** |
| Glutamine | 38.63 a | 36.47 a | 37.23 a | 37.44 | 81.61 b | 79.08 b | 0.07 c | 53.59 | ns | * | *** |
| Glutamate | 44.66 a | 17.2 b | 17.79 b | 26.55 | 29.58 c | 20.12 bc | 2.86 d | 17.52 | ns | *** | *** |
| Glycine | 1.79 a | 1.06 b | 2.44 a | 1.76 | 1.94 a | 1.00 b | 0.34 b | 1.09 | ns | ns | *** |
| MEA | 7.66 a | 3.22 b | 0.25 a | 5.63 A | 2.97 b | 3.36 b | 2.27 b | 2.87 B | ** | ns | * |
| Ornithine | 1.06 a | 1.02 a | 0.54 b | 0.87 | 0.87 ab | 1.81 c | 0.44 b | 1.04 | ns | ** | *** |
| Proline | 6.66 a | 3.63 ab | 12.07 c | 7.45 | 3.95 ab | 11.38 c | 2.20 b | 5.84 | ns | ns | *** |
| Serine | 3.44 a | 3.1 a | 3.74 a | 3.43 | 3.45 a | 3.91 a | 0.14 b | 2.50 | ns | ns | *** |
| Threonine | 2.83 a | 2.81 a | 3.97 ab | 3.20 | 2.91 a | 4.40 b | 0.08 b | 2.46 | ns | ns | *** |
| Total AA | 256 a | 173.9 a | 231.1 a | 220.30 | 233.5 a | 231.6 a | 12.03 b | 159 | ns | ns | * |
| Minor AA | 19.18 a | 15.82 ab | 12.9 bc | 15.97 | 16.72 a | 19.27 a | 2 ac | 12.66 | ns | ** | *** |
| BCAAs | 8.22 a | 6.02 a | 7.69 a | 7.31 A | 5.75 a | 6.83 a | 1.27 b | 4.62 B | * | ns | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



