Submitted:
18 July 2023
Posted:
19 July 2023
You are already at the latest version
Abstract
Early weaning of piglets caused excessive concentration of free radicals, which would oxidize and damage the intestinal tract of piglets, resulting in dyspepsia, low growth, diarrhea, microbiota disorders and intestinal infections. This research studied on effect of different levels of taurine in diets on intestinal oxidative damage in early weaned piglets. A total of 48 weaned piglets (6.71 ± 0.73 kg) of 28-day-old crossbred "Duroc × Landrace × Yorkshire" were assigned to 4 groups with 12 replicates per group and 1 pig per replicate: piglets were fed a basal diet with 0.0% (CON), 0.2% taurine (L-Tau), 0.3% taurine (M-Tau), or 0.4% taurine (H-Tau). The animal experiment lasted for 30 days. With the increase of dietary taurine dosage, the activity of antioxidant enzymes including superoxide dismutase(SOD),glutathione peroxidase(GPX),catalase(CAT) and peroxidase(POD) were up-regulated (Linear, P < 0.05), the mRNA abundance of antioxidant enzymes including SOD1, SOD2, GPX1 and heme oxygenase-1(HO-1)were up-regulated (P < 0.05), glutathione (GSH) concentrations was up-regulated (Linear, P < 0.05) , nitric oxide (NO), malondialdehyde (MDA) and oxidized glutathione (GSSG) concentrations were down-regulated (Linear, P < 0.05);The villus height (VH) and villus height/ crypt depth (VH/CD) of jejunum and ileum were increased (Linear, P < 0.05); The expression of tight junction proteins including zonula occludens protein 1(ZO-1), occludin (OCLN) and closure protein 1(CLDN-1) in the jejunum mucosa were up-regulated (Linear, P < 0.05), the mRNA abundance of ZO-1, OCLN and CLDN-1 were up-regulated (P < 0.05),and the D-lactose (D-LA) content in blood was down-regulated (Linear, P < 0.05); the colonization abundance of Firmicutes was increased (Linear, P < 0.05), while Proteobacteria and Spirochaetota was decreased (Linear, P < 0.05); The diarrhea index decreased (Linear, P < 0.05), while the feed conversion rate increased (Linear, P < 0.05). Collectively, supplementation of 0.3% and 0.4% taurine in feed can significantly alleviate intestinal oxidative damage by enhancing antioxidant capacity of piglets, and improve feed utilization of piglets after repairing intestinal damage.
Keywords:
taurine
; weaned piglets
; antioxidant capacity
; Intestine Healthy
; microorganism
; growth performance
; Nrf2/ARE signaling pathway
1. Introduction
At present, in order to improve the reproductive rate of sows and the survival rate of piglets, the pig breeding industry normally adopts early weaning isolation technology to shorten the traditional suckling period of approximately 2 months to 15-28 days [1]. However, this technique will cause piglets to bear nutritional, psychological and environmental pressure in advance [2], causing piglets stress response [3]. Studies have found that excessive concentration of free radicals produced by the body during weaning stress will destroy the redox balance [4], which will oxidize and damage the intestinal morphological structure and mucosal barrier of piglets. The damaged intestinal morphological structure will reduce the digestion and absorption ability of food, and the damaged intestinal mucosal barrier will easily lead to intestinal allergy and the activation of certain neuroregulatory pathways. Viruses and bacteria can invade and colonize the intestine, causing intestinal inflammation, flora imbalance, dyspepsia, diarrhea, slow growth and a series of intestinal diseases [5].
The addition of excessive doses of zinc and copper in feed is a common international practice to prevent and alleviate intestinal diseases of piglets, although it can significantly improve the intestinal health of piglets, but the zinc and copper not absorbed by the animal body will seriously pollute the water and soil. This approach completely violates the conservation, green and environmental protection development concept advocated by the International Health Organization, so in recent years, many countries have begun to introduce regulations to limit the dosage of zinc and copper in feed, calling for exploring new ways to deal with intestinal diseases from greener and safer aspects such as enzyme preparations, acidifiers and amino acids.
Taurine (Tau) is a semi-essential β-amino acid and is the most abundant free amino acid in animal tissues [6], accounting for 25%, 50%, 53% and 19% of the total free amino acids in liver, kidney, muscle and brain tissues, respectively [7]. The significance of its high concentration is not to provide raw materials for protein synthesis, nor to provide energy for body metabolism, but to play its rich biological functions as a body regulator, and antioxidant is one of its main functions. It has been found that Tau can avoid the mistranslation of proteins related to the electron transport chain by strengthening the precise positioning of codons and anti-codons, so as to maintain the structural and functional integrity of the electron transport chain more efficiently, thus avoiding the leakage of electrons during the transport process and reducing the generation of reactive oxygen species (ROS) [8,9]. Tau can act on the Keap1-Nrf2/ARE antioxidant signaling pathway, promote NF-E2-related factor 2 (Nrf2) to enter the nucleus and bind to the small Maf protein (Maf) in the nucleus into heterodimer, and then act on the antioxidant reaction element (ARE) on the nucleic acid sequence to induce the expression of antioxidant oxidase genes [10]. In addition to acting as a pH regulator in the glutathione redox system [11], exogenous supplementation of Tau can also increase the amount of glutathione synthesis in the body [12], thereby improving the antioxidant capacity of the body.
Because Tau has a clear antioxidant effect, this study is of great significance to explore the antioxidant effect of Tau on the alleviation of intestinal oxidative damage in weaned piglets through three main aspects: intestinal morphology, mucosal barrier and microbial flora: On the one hand, expand the new understanding of the application value of Tau in livestock and poultry nutrition and medical circles, promote the development of related research, and further provide theoretical guidance for the application of Tau in livestock and poultry breeding industry; On the other hand, it provides a more efficient, green and safe solution to the intestinal oxidative damage induced by early weaning in livestock and poultry breeding, thereby improving the health of weaned piglets and improving the economic benefits of the farm.
2. Materials and Methods
2.1. Experimental Material
Tau (A3820, 99.0%), pharmaceutical grade, purchased from Shanghai Yuanju Biotechnology Co., Ltd. Weaned piglets purchased from Chongqing Nongdianshan Agricultural Science and Technology Development Co., Ltd.
2.2. Experimental Design and Sample Collection
A total of 48 weaned piglets (6.71±0.73 Kg) of 28-day-old crossbred “ Duroc × Landrace × Yorkshire “ were assigned to 4 groups with 12 replicates per group and 1 pig per replicate. piglets were fed a basal diet with 0.0% (CON), 0.2% taurine (L-Tau), 0.3% taurine (M-Tau), and 0.4% taurine (H-Tau). The basic diet is designed according to the nutritional requirements of NRC (2012), and the composition and nutritional composition of feed materials for each group are shown in Table 1. Piglets were kept alone in a mechanically ventilated and temperature-controlled chamber (28±1.2 °C) with free feed and water intake during the experiment.
The feed was pre-fed for 3 days and the formal experiment lasted for 30 days, during which feed intake was recorded every day and the degree of diarrhea was scored, as shown in Table 2. After the experiment, the representative 4 piglets in the selected group underwent fasting blood collection and were killed after blood collection. The length and weight of jejunum and ileum were measured. The cecum contents were collected in a 10 mL sterilized centrifuge tube and quickly frozen in liquid nitrogen. Two pieces of intestinal tissue were taken from the middle part of jejunum and ileum with a length of 5 cm and placed into a 50 mL centrifuge tube containing tissue fixative. After rinsed with normal saline, 30 cm of jejunum and ileum mucosa were scraped and wrapped in sterile tin foil and frozen with liquid nitrogen.
2.3. Biochemical Index Determination
Biochemical assay kit (Nanjing Jiancheng Bioengineering Institute) was used to determine: Serum superoxide dismutase (SOD) activity, glutathione peroxidase (GPX) activity, catalase (CAT) activity, peroxidase (POD) activity, total antioxidant capacity (T-AOC), superoxide anion (O2•-) clearance rate, hydroxyl radical (•OH) clearance rate, glutathione (GSH), oxidized glutathione (GSSG), malondialdehyde (MDA), nitric oxide (NO) and D-lactose (D-LA) concentrations; Liver O2•- clearance rate, •OH clearance rate, POD activity and glutathione synthase (GSHS) activity index; POD activity, •OH clearance rate, O2•- clearance rate and GSHS activity of jejunum mucosa.
2.4. ELISA Index Determination
The index of tight junction protein occludin (OCLN), closure protein-1 (CLDN-1) and zonula occludens protein 1 (ZO-1) were determined by ELISA kit (Quanzhou Risin Biotechnology Co., LTD.).
2.5. mRNA Abundance
Total RNA was extracted using a universal RNA lifting kit (Accurate Biology, AG21022), and then the concentration and purity of total RNA (A260/A280, A260/A230) were detected using a nucleic acid detector. Samples with qualified concentration and purity were reverse-transcribed into cDNA using a reverse transcription kit (Accurate Biology, AG11728), followed by PCR analysis using a fluorescent quantitative kit (Accurate Biology, AG11701) using the SYBR Green method. The expression abundance of Nrf2, heme oxygenase-1 (HO-1), SOD1, SOD2, GPX1 and Glutathione-S-Transferase (GST) genes were measured in liver samples with GAPDH as the internal reference gene. The expression abundance of ZO-1, OCLN and CLDN-1 genes were determined by jejunum mucosal samples. The obtained Ct values were normalized by 2-ΔΔCt method. The gene primer sequences of internal reference genes and target genes obtained from NCBI are shown in Table 3.
2.6. Observation of Intestinal Histomorphology
The fixed samples of jejunum and ileum were prepared and stained by HE. The villus height (VH) and crypt depth (CD) were measured.
2.7. Cecal Microbial 16S rDNA Sequencing Analysis
The cecal contents collected with aseptic centrifuge tubes were sent to -80℃ for frozen storage, and then sent to Hangzhou Lianchuan Biotechnology Co., LTD., for microbial isolation of cecal contents. 16S rDNA high-throughput sequencing was used to analyze cecal microflora.
2.8. Growth Performance Measurement
Average daily gain (ADG) = (final weight - initial weight)/days
Average daily feed intake (ADFI) = total feed intake/days
Feed conversion rate = average daily gain/ average daily feed intake
2.9. Statistic Analysis
Anova was performed on the data using the General Linear Model (GLM) program in SAS Statistical software (SAS 9.4). The differences between the groups were determined by Duncan analysis. The significance was set as P < 0.05.
3. Results
3.1. Antioxygenic Capacity
As shown in Table 4, compared with CON group, with the increase of Tau supplemental dose, the O2•- and •OH clearance rate in blood, liver and jejunum mucosa were significantly up-regulated (Linear, P < 0.05). SOD activity, GSH-PX activity, CAT activity, POD activity and T-AOC in blood were significantly up-regulated (Linear, P < 0.05), GSH concentration and GSH/GSSG were significantly up-regulated (P < 0.05), and NO, MDA and GSSG concentrations were significantly down-regulated (Linear, P < 0.05). GSHS activity in liver and jejunum mucosa was significantly up-regulated (Linear, P < 0.05). Figure 1 showed that compared with CON group, mRNA abundance of antioxidant enzyme genes HO-1, SOD1, SOD2, GPX1 and nuclear factor Nrf2 in H-Tau group was significantly up-regulated (P < 0.05). The mRNA abundance of SOD1, SOD2 and Nrf2 in M-Tau group was significantly up-regulated (P < 0.05). L-Tau group significantly up-regulated the mRNA abundance of SOD2 (P < 0.05); The mRNA abundance of enzyme gene GST in H-Tau, M-Tau and L-Tau groups was not significantly different from CON group (P < 0.05). In general, the improvement effect of H-Tau group on the antioxidant capacity of the body was greater than M-Tau group, while the L-Tau group had no improvement effect on the antioxidant capacity of the body.
3.2. Intestinal Morphology and Structure
As shown in Table 5, compared with CON group, with the increase of Tau supplemental dose, the VH and VH/CD of jejunum and ileum were significantly up-adjusted (Linear, P < 0.05), while the length, length index, weight and weight index of jejunum and ileum had no significant changes (P > 0.05). In general, the improvement of intestinal morphology and structure in H-Tau group was greater than that in M-Tau and L-Tau groups, and the improvement effect in L-Tau group was the worst. Figure 2 showed that compared with CON group, H-Tau and M-Tau groups had clearer inter-villous boundaries, fewer damaged microvilli in the villus and intervillus space, and more orderly and consistent arrangement of microvilli.
3.3. Intestinal Mucosal Barrier
As shown in Table 6, compared with CON group, with the increase of Tau supplemental dose, blood D-LA content was significantly down-regulated (Linear, P < 0.05), while protein concentrations of CLDN-1, ZO-1 and OCLN in jejunum mucosa were significantly up-regulated (Linear and Quadratic, P < 0.05). Figure 3 showed that compared with CON group, mRNA abundance of ZO-1, OCLN and CLDN-1 proteins was significantly up-regulated in H-Tau group (P < 0.05), while mRNA abundance of OCLN protein was only significantly up-regulated in M-Tau and L-Tau groups (P < 0.05).
3.4. Cecum Microflora
As shown in Table 7, compared with CON group, there were no significant differences in observed_otus, shannon, simpson Chao1, pielou_e and goods_coverage indexes of H-Tau, M-Tau and L-Tau groups (P > 0.05). Compared with CON group, with the increase of Tau supplemental dose, the colonization abundance of Firmicutes was significantly increased (Linear, P < 0.05), while the colonization abundance of Proteobacteria and Spirochaetota was significantly decreased (Linear, P < 0.05). Compared with CON group, the Clostridium_sensu_stricto_1 and Turicibacter genera colonization abundance in the H-Tau, M-Tau and L-Tau groups were significantly upregulated with increasing Tau supplemental dose (Linear, P < 0.05). The colonization abundance of Terrisporter genus was significantly up-regulated (P < 0.05), while the colonization abundance of UCG-005 genus was significantly down-regulated (Linear, P < 0.05). Figure 4 shows 781 ASVs unique to CON group, 569 ASVs unique to L-Tau group, 575 ASVs unique to M-Tau group, and 441 ASVs unique to H-Tau group. CON, L-Tau and M-Tau groups share 78 ASV species, CON, L-Tau and H-Tau groups share 59 ASV species, CON, M-Tau and H-Tau groups share 59 ASV species, and L-Tau, M-Tau and H-Tau groups share 183 ASV species. CON group had the largest number of unique ASV species, and L-Tau, M-Tau and H-Tau combinations had the largest number of unique ASV species. Figure 5 shows that at the phylum level, the branch distances of L-Tau, M-Tau, H-Tau groups and CON groups (the shorter the distance, the more similar the microbial clustering) are greater than 0.06 units, while the branch distances of L-Tau, M-Tau and H-Tau groups are less than 0.04 units. Moreover, the branch distance between Tau group and CON group was positively correlated with Tau supplemental dose. As shown in Figure 6, the distance between L-Tau, M-Tau and H-Tau groups was relatively close, and there were many overlapping parts, indicating that the beta diversity of microorganisms in cecal contents was similar between Tau supplementation groups. CON group was far away from L-Tau, M-Tau and H-Tau groups, and there was almost no overlap, indicating that the addition of Tau in feed had a certain effect on the beta diversity of microorganisms in cecum contents.
3.5. Growth Performance
As shown in Table 8, compared with CON group, with the increase of Tau supplemental dose, the diarrhea index decreased significantly (Linear, P < 0.05), while the feed conversion rate increased significantly (Linear, P < 0.05).
4. Discussion
Early weaning is a necessary link in contemporary piglet breeding, and it is also an important means for farms to improve production benefit. However, after early weaning of piglets, the activity of antioxidant enzymes in the body is significantly decreased, resulting in increased concentrations of ROS and reactive nitrogen species (RNS), thus damaging proteins and lipids in the body and causing oxidative damage to tissues and organs [14-16]. Therefore, it is known that after early weaning of piglets, the body is in a peroxide state and there is an antioxidant demand. The results of this study showed that Tau could enhance its antioxidant activity by up-regulating the mRNA abundance of antioxidant oxidase genes, thereby improving the scavenging ability of ROS and RNS, and reducing the damage to the body. The reason for this result may be that Tau activates the Keap1-Nrf2/ARE antioxidant signaling pathway. Studies have shown that [17-19], hydrogen peroxide generates hypochlorous acid in the presence of chloride ions, and then hypochlorous acid reacts with Tau to produce nitrogen chloro-Tau, which can activate nuclear factor Nrf2 and make it enter the nucleus to interact with the antioxidant reaction element (ARE). The transcriptional levels of antioxidant enzyme genes HO-1, CAT, SOD and GSH-Px are up-regulated [20], which increases the antioxidant capacity of the body. Tau not only enhances the scavenging ability of ROS and RNS, but also reduces its production rate. It has been found [21,22] that Tau can enhance the effective synthesis rate of mitochondrial electron transport chain related proteins by enhancing the precise location of codons and anti-codons,such as NADH dehydrogenase subunit 5(ND5), NADH dehydrogenase subunit 6(ND6)and cytochrome b(Cytb),in order to maintain the structural and functional integrity of the electron transport chain, so as to avoid electron leakage during the transport process, and thus greatly reduce the generation of ROS. In addition, the results of this study also showed that Tau significantly up-regulated the blood GSH concentration and GSH/GSSG ratio, indicating that Tau increased the reducing capacity of glutathione and weakened its oxidation capacity, and indirectly reflected that Tau could up-regulate the antioxidant potential of glutathione. Some scholars have also found that Tau can maintain the REDOX ability of glutathione by regulating the pH in mitochondria [11]. At the same time, we also found that Tau up-regulated the activity of glutathione synthase in liver and jejunum mucosa, possibly because Tau and glutathione share the same synthetic raw material cysteine [12]. Exogenous Tau supplementation can reduce the endogenous synthesis of Tau by hepatocytes, thus limiting the participation of cysteine in the pathway of Tau synthesis. This will save a lot of cysteine raw materials for the synthesis of glutathione, and a lot of saved cysteine activates glutathione synthase as a substrate, and then synthesizes more glutathione, increasing the antioxidant capacity of the body.
Whether the nutrients in the feed can be fully digested and absorbed depends to a certain extent on whether the animal has a good intestinal morphological structure. Because of its long retention time of food and large surface area of absorption, the gut is the main supplier of nutrients to the body. Studies have shown [15] that oxidative stress caused by weaning can cause the destruction of jejunal villi accompanied by lamina propria edema, and significantly reduce the height and width of jejunal villi, and significantly increase the CD. These changes will seriously reduce the digestion and absorption function, and then seriously slow down the growth and development rate of piglets. In this study, Tau was found to significantly up-regulate VH and VH/CD in jejunum and ileum, which was consistent with previous studies [23]. The increase of VH can increase the surface area of nutrient absorption in the intestine, improve the absorption capacity of nutrients in the intestine of piglets, and further promote the growth and development of piglets. The small intestine epithelium is renewed every 2-5 days and is the most regenerative tissue in mammals [24]. The intestinal epithelium is composed of many repetitive crypt-villus units that undergo continuous cycle of renewal and repair [25]. The crypt was proliferative areas, and early progenitor cells produced by the intestinal stem cells (ISCs) at the bottom migrated to the crypt-villus axis and differentiated into epithelial cells [26], thereby repairing intestinal damage. However, the efficiency of ISCs in intestinal damage repair is closely related to the REDOX state. Excessive accumulation of ROS in mammals can inhibit the proliferation and differentiation of ISCs. Studies have found that the proliferation and differentiation process of ISCs is blocked under high ROS levels [27]. Further studies have confirmed that the activation of Keap1/Nrf2 signaling pathway significantly reduces the ROS content in the intestine, creating an appropriate ROS content for intestinal cells and enhancing the activity of ISCs [28]. In addition, Keap1/Nrf2 signaling may work synergistically with Notch signaling to determine the proliferation and differentiation of ISCs. Studies have found that the functional regulatory region of Notch1 gene is located at the proximal end of the Nrf2 promoter [29], which may be activated with the activation of Nrf2. It can negatively regulate the Notch downstream effector Math1, thus disrupting the Notch cascade and accelerating the proliferation and differentiation of mouse ISCs [30]. These results suggest that taurine can maintain the REDOX state required for the proliferation and differentiation of ISCs by activating the Keap1/Nrf2/ARE antioxidant pathway and increasing the abundance of glutathione, and may activate the functional regulatory region gene of Notch1 gene while activating Nrf2, thus interfering with Notch signaling pathway. The proliferation and differentiation ability of ISCs in piglets were enhanced, and the repair efficiency of intestinal damage was improved.
Intestinal mucosal barrier can effectively prevent harmful substances from passing through the intestinal mucosa and entering the blood, thus avoiding the occurrence of enteroborne infections. Studies have found that oxidative stress can reduce the abundance of intestinal protein expression of OCLN, CLDN-1 and ZO-1 in piglets [31], and significantly increase the level of lipopolysaccharide in blood [32]. This indicates that oxidative stress can indeed increase the permeability of intestinal mucosa, damage the intestinal defense barrier of piglets, and cause endotoxins to enter the blood through the mucosa to further threaten the health of piglets. However, in this study, this phenomenon was alleviated to a certain extent, and it was observed that dietary Tau supplementation could significantly up-regulate the mRNA abundance of ZO-1, OCLN and CLDN-1 protein genes in jejunal mucosa and their expression levels in jejunal mucosa, and significantly reduce the content of D-LA in blood. This suggests that Tau can reduce intestinal mucosal permeability by restoring the tight connections between cells, thereby inhibiting enteric metabolites from crossing the intestinal defense barrier into the bloodstream, reducing the risk of enteric diseases in piglets. The mechanism of taurine promoting the intestinal barrier has not been fully established. Since taurine have the ability to promote intestinal antioxidant capacity as well as barrier function and the study found that taurine promoted the tight junction protein expression and activated the Nrf2 signaling pathway, we further hypothesized that taurine promoted intestinal barrier function via the Nrf2 signaling pathway. Previous studies found that inhibition or deletion of Nrf2 decreased the tight junction protein expression in cells or mice [33,34]. The effect of Nrf2 on intestinal barrier function is possibly indirectly promoted in many ways, which may be associated with the regulation of Nrf2-mediated inflammation, oxidative stress, T cell activation, and autophagy [35]. In addition, Nrf2 might directly regulate tight junction protein expression, and it has been reported that Nrf2 binds to the promoter of claudin 4, which has the potential to result in an increase in claudin 4 transcription [36]. Future research is needed to determine whether Nrf2, as a transcription factor, binds the promoter of other tight junction genes to promote their transcription.
The microbiota inhabits the intestines of piglets in a dynamic composition and is regulated by a variety of factors. The imbalance of intestinal flora caused by oxidative stress may be one of the main causes of various intestinal diseases. Studies have shown [37] that oxidative stress can significantly reduce the abundance of Lactobacillus and Faecalibacterium in the gut of piglets, while significantly increase the abundance of Clostridium cluster I. Lactobacillus is a beneficial bacterium that can resist the invasion and colonization of exogenous pathogenic microorganisms [38], Faecalibacterium is a resident anti-inflammatory probiotic in the gut of healthy animals, and Clostridium cluster Ⅰ is a conditional pathogenic bacterium [39]. The imbalance of intestinal microbiota can lead to a variety of intestinal diseases [40]. Therefore, the above study found that after oxidative stress of piglets, the abundance of beneficial bacteria in the intestine is down, and the abundance of harmful bacteria is up, which indicates that oxidative stress can cause a variety of intestinal diseases by changing the microbial flora in the intestine, and then adversely affect the digestion, absorption, growth and development of piglets. Interestingly, this appears to be reversed when Tau is added to the feed. According to the Venn diagram of ASV distribution between groups, the addition of Tau made 781 ASV microorganisms disappear from the cecum colonization, but 183 ASV microorganisms were introduced into the cecum colonization. The reason why Tau changes the microflora of cecum may be that it changes the microenvironment of cecum in some way. It is reported [41] that Tau, as a sulfite donor, can produce hydrogen sulfide in the intestine, and hydrogen sulfide is an important metabolic product in the intestine, which can have both beneficial and harmful effects on the intestinal environment. In this way, it showed certain selectivity to the microorganisms colonized in the cecum. Although Tau had no significant effect on alpha diversity of cecum microflora, it had some effect on Beta diversity. This shows that Tau does not change the total number of microflora species and their quantitative distribution in the cecum, but causes changes in the composition and proportion of microflora species.
The test results showed that at the phylum level, Tau significantly increased the proportion of Firmicutes, and significantly decreased the proportion of spirobacteroides and Proteobacteria. Although there was no significant effect on Bacteroides, the proportion of Bacteroides showed a significant downward trend with the increase of Tau dose. According to relevant reports, Firmicutes can improve energy utilization in the diet, and Bacteroides are common microorganisms that degrade polysaccharides [42], and the ratio of Firmicutes to Bacteroides is usually positively correlated with intestinal digestion and absorption [43,44]. It was also observed in this experiment that the feed utilization rate of piglets was significantly improved after the addition of Tau in the diet, which indicates that Tau may improve the production performance of animals by improving the intestinal flora. Proteobacteria are a group of highly adaptable and potentially pathogenic bacteria in the gastrointestinal tract, and the imbalance of intestinal flora in the colon is usually accompanied by the increase of Proteobacteria abundance [45]. Spirochaeta is mostly pathogenic bacteria or conditional pathogenic bacteria, which has a potential induction effect on piglet diarrhea [46]. The proportion of Proteobacteria and spirochaeta decreased significantly after the addition of Tau in the diet, which also indicates that the addition of Tau can limit the growth of harmful bacteria and is more conducive to the health of the body. At the generic level, Tau significantly increased the proportion of Clostridium_sensu_stricto_1, Terrisporobacter and Turicibacter. Clostridium_sensu_stricto_1, Terrisporobacter and Turicibacter all belong to the Firmicutes group of probiotics [47]. The study found that Clostridium_sensu_stricto_1 can utilize mucous derived sugars, such as glucose, and can also act as a fiber degrading bacterium that degrades cellulose and hemicellulose [48], thereby improving the animal’s utilization capacity of coarse fiber in feed. Terrisporter is a lactic acid-degrading bacterium. Under anaerobic conditions, Terrisporter can degrade lactic acid [49], thereby avoiding acidosis and intestinal damage caused by excessive lactic acid in the intestine. Therefore, the results of this experiment also indicate that Tau may improve the intestinal nutrient utilization capacity of piglets by increasing the proportion of beneficial bacteria, and then maintain intestinal health by improving the flora.
Intestinal health is closely related to the body’s digestion and absorption ability. The above results are sufficient to prove that the addition of Tau in feed can indeed alleviate the intestinal oxidative damage induced by early weaning of piglets. Meanwhile, we also observed that the addition of Tau in feed can significantly reduce the diarrhea index of piglets and improve the feed conversion rate. These results indicate that Tau can improve feed utilization ability of piglets by alleviating intestinal oxidative damage. In addition, we also speculate that this beneficial effect may be related to taurine improving the synthesis capacity of GSH, Because GSH can improve the body’s ability to absorb nutrients. Researches show that GSH can participate in the absorption of amino acids through the γ-glutamyl cycle, and the extracellular amino acids react with GSH under the action of γ-glutamul transpeptidase (γ-GT) to generate γ-glutamyl-amino-acids and cysteinyl-glycine into the cell. In the cytoplasm, γ-glutamyl amino acids are catalyzed by γ-glutamyl cyclotransferase to decompose into amino acids and 5-hydroxyproline. 5-hydroxyproline is hydrolyzed to glutamic acid under the catalysis of 5-hydroxyprolinase, and cysteine-glycine is hydrolyzed to cysteine and glycine by dipeptidase, and GSH can be synthesized in the cell [50]. GSH can also reduce Fe3+ to Fe2+, and Fe2+ is more easily absorbed by intestinal mucosa, thus promoting iron absorption [51]. Studies have shown that changes in the ratio of GSSG/GSH can directly or indirectly affect the activity of enzymes related to carbohydrate metabolism [52] and the activity of Na+ dependent D-glucose carriers, and the reduction of this ratio can improve the carrying capacity of glucose [53].
Although we also observed a decrease in feed intake after Tau supplementation, this was not a negative effect. The reason may be that with the increase of Tau dosage, intestinal oxidative damage can be repaired, the degree of diarrhea of piglets is significantly reduced, the digestion and absorption ability of feed is significantly improved, and the body’s demand for material and energy can be met by eating less feed. As for the CON group, the intestinal oxidative damage was not alleviated and the degree of diarrhea was serious, so they needed to eat more feed to meet the needs of the body in the state of free feeding. Some studies have found that Tau can significantly improve the growth performance of piglets fed a high plant protein diet, but has no effect on piglets fed a high animal protein diet [54]. The reason for this divergence may be that almost all plant feeds do not contain Tau, while animal feeds are rich in Tau [55]. The high concentration of Tau contained in high animal protein diets has met the needs of animals, and additional Tau supplementation has little effect on growth performance at this time. However, the high plant protein diet basically does not contain Tau, and the body has a relatively large demand for Tau. At this time, additional Tau supplementation can significantly improve the growth performance. The study found [56] that after 1% Tau was added to the basic diet containing 7.5% animal protein feed (2.5% whey powder +5% fish meal), the feed to meat ratio of the experimental group on the 10th day after weaning was reduced by 12.5% compared with the control group, and the difference in daily gain was not significant, but there was a trend of improvement. Similarly, it was observed in this experiment that with the increase of Tau supplemental dose, the feed conversion rate showed a linear increase, H-Tau and M-Tau groups were significantly higher than CON group, and the difference in daily gain was not significant, but there was a trend of improvement. These directly indicate that the additional Tau does have a significant improvement effect on intestinal digestion and absorption, and indirectly reflect that the Tau provided in the basic diet supplemented with 15% animal protein feed (5% fish meal +10% whey meal) in this study still cannot meet the needs of the body. And because the price of animal protein feed is much higher than the price of plant protein feed, the proportion of animal protein feed added in the actual production will also be constrained, which leads to the actual production completely rely on the Tau in animal protein feed to meet the needs of the body is unrealistic, we must add additional Tau in the diet to meet the needs of the body.
5. Conclusions
In this study, it was found that adding proper amount of Tau to feed can significantly alleviate intestinal oxidative damage of piglets by enhancing the antioxidant capacity of the body, and repairing the intestinal damage can improve the digestion and absorption ability of piglets. In the concentration range of this trial, the overall improvement was positively correlated with the added dose of Tau, and the improvement effect was most prominent in the H-Tau group (0.4% Tau). It is worth noting that the overall results of this study showed that the repair effect of Tau on intestinal oxidative damage and the improvement effect on feed utilization capacity were linearly correlated with the dosage of Tau. This shows that the dosage of 0.4% Tau added to the feed is not the best dose, and continuing to increase the concentration of Tau in the feed may make the alleviation effect of intestinal oxidative damage better, but the specific effect needs to be confirmed by experiment.
Author Contributions
L.W. and Z.T. Animal Experiments, Sample Collection, Data curation; L.W., Y.Q., Z.H. and C.C. Methodology; Y.Q. and H.D. Data curation; L.W. and Z.T. Writing – Review and Editing; Z.T.: Project administration. WT: Resources, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Natural Science Foundation Project of Chongqing (cstc2021jcyj-msxmX0966), Innovation Research 2035 Pilot Program of Southwest University (SWU-XDPY22005), Fundamental Research Funds for National Key R&D Program of China (2022YFD1300501-3), Sichuan Science and Technology Program (2023YFQ0031, 2023ZHYZ0007).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the License of Experimental Animals (IACAU-20211007-06) of the Animal Experimentation Ethics Committee of Southwest University, Chongqing, China.
Informed Consent Statement
Not applicable.
Data Availability Statement
The 16S rDNA sequencing raw data are available at NCBI under the accession number PRJNA97422 by web link (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA97422). The rest of the raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Acknowledgments
We would like to thank Editage (www.editage.cn) for English language editing.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Ipema, A.F.; Bokkers, E.A.M.; Gerrits, W.J.J.; Kemp, B.; Bolhuis, J.E. Providing live black soldier fly larvae (Hermetia illucens) improves welfare while maintaining performance of piglets post-weaning. Sci Rep. 2021, 11, 7371. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Martins, C.F.; Lopes, P.A.; Palma, M.; Pinto, R.M.A.; Costa, M.; Alfaia, C.M.; Pestana, J.M.; Coelho, D.; Ribeiro, D.M.; Viegas, I.; Almeida, A.M.; Freire, J.P.B.; Prates, J.A.M. Impact of dietary Chlorella vulgaris and feed enzymes on health status, immune response and liver metabolites in weaned piglets. Sci Rep. 2022, 12, 16816. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhu, L.; Zhao, K.; Chen, X.; Xu, J. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J Anim Sci. 2013, 91(3), 1522, Erratum in: J Anim Sci. 2013, 91, 1522. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Péron, V.; Luron, I.; Lallès, J.P.; Sève, B. Weaning Induces Both Transient and Long-Lasting Modifications of Absorptive, Secretory, and Barrier Properties of Piglet Intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- IN, J.; REN, W.K.; WU, X.S. Oxidative stress-mediated signaling pathways: a review. J. Agr. Food Chem. 2013, 11, 132–139. [Google Scholar]
- Huxtable, R.J. Physiological actions of taurine. Physiol Rev. 1992, 72, 101–63. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: an overview. J Nutr. 2006, 136, 1636–1640. [Google Scholar] [CrossRef] [PubMed]
- Kirino, Y.; Goto, Y.; Campos, Y.; Arenas, J.; Suzuki, T. Specific correlation between the wobble modification deficiency in mutant tRNAs and the clinical features of a human mitochondrial disease. Proc Natl Acad Sci U S A. 2005, 102, 7127–32, Epub 2005 May 3. PMID: 15870203; PMCID: PMC1129107. [Google Scholar] [CrossRef]
- Kirino, Y.; Yasukawa, T.; Ohta, S.; Akira, S.; Ishihara, K.; Watanabe, K.; Suzuki, T. Codon-specific translational defect caused by a wobble modification deficiency in mutant tRNA from a human mitochondrial disease. Proc Natl Acad Sci U S A. 2004, 101, 15070–15075, Epub 2004 Oct 11. PMID: 15477592; PMCID: PMC524061. [Google Scholar] [CrossRef]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267, Epub 2019 Mar 30. PMID: 30929309; PMCID: PMC6536957. [Google Scholar] [CrossRef]
- Hansen, S.H.; Grunnet, N. Taurine, glutathione and bioenergetics. Adv Exp Med Biol. 2013, 776, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, Y.; Yan, G.; Bu, Q.; Lv, L.; Yang, Y.; Zhao, J.; Shao, X.; Deng, Y.; Zhu, R.; Zhao, Y.; Cen, X. Protective role of taurine against morphine-induced neurotoxicity in C6 cells via inhibition of oxidative stress. Neurotox Res. 2011, 20, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Hart, G.K.; Dobb, G.J. Effect of a fecal bulking agent on diarrhea during enteral feeding in the critically ill. JPEN J Parenter Enteral Nutr. 1988, 12, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Wu, M.; Tan, B.; Yin, Y.; Li, T.; Xiao, D.; Li, L. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: I. Growth performance, immune function, and antioxidation capacity. J Anim Sci. 2013, 91, 4772–4780, Epub 2013 Aug 21. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Cai, X.; Xu, C.; Luo, Z.; Sheng, Y.; Bao, J.; Chen, X.; Xu, J. Effects of dietary supplementation with N-acetyl cysteine on antioxidant capacities and the expression of inflammatory cytokines in weaned piglets. J Anim Sci. 2016, 15, 634–641. [Google Scholar] [CrossRef]
- Luo, Z.; Zhu, W.; Guo, Q.; Luo, W.; Zhang, J.; Xu, W.; Xu, J. Weaning Induced Hepatic Oxidative Stress, Apoptosis, and Aminotransferases through MAPK Signaling Pathways in Piglets. Oxid Med Cell Longev. 2016, 2016, 4768541. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Lee, D.S. Taurine Chloramine Prevents Neuronal HT22 Cell Damage Through Nrf2-Related Heme Oxygenase-1. Adv Exp Med Biol. 2017, 975, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.S.; Kim, C. Taurine chloramine administered in vivo increases NRF2-regulated antioxidant enzyme expression in murine peritoneal macrophages. Adv Exp Med Biol. 2013, 775, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Cha, Y.N. Taurine chloramine produced from taurine under inflammation provides anti-inflammatory and cytoprotective effects. Amino Acids. 2014, 46, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Xu, L.; Lu, M.; Chen, Z.; Yuan, Z.; Xu, X.; Guo, X.; Zhang, X.; Sun, H.; You, Q. Structure-Activity and Structure-Property Relationship and Exploratory in Vivo Evaluation of the Nanomolar Keap1-Nrf2 Protein-Protein Interaction Inhibitor. J Med Chem. 2015, 58, 6410–21. [Google Scholar] [CrossRef] [PubMed]
- Tomoda, E.; Nagao, A.; Shirai, Y.; Asano, K.; Suzuki, T.; Battersby, B.J.; Suzuki, T. Restoration of mitochondrial function through activation of hypomodified tRNAs with pathogenic mutations associated with mitochondrial diseases. Nucleic Acids Res. 2023, 17, 139, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Kirino, Y.; Goto, Y.; Campos, Y.; Arenas, J.; Suzuki, T. Specific correlation between the wobble modification deficiency in mutant tRNAs and the clinical features of a human mitochondrial disease. Proc Natl Acad Sci U S A. 2005, 102, 7127–7132, Epub 2005 May 3. PMID: 15870203; PMCID: PMC1129107. [Google Scholar] [CrossRef]
- Ma, Y.; Lv, Q.; Zhao, D.; Wang, J.; Fu, Y.; Li, C.; Wu, G.; Liu, M.; Hu, J.; Lin, S.; Yang, J. Intervention Effect of Taurine on LPS-Induced Intestinal Mechanical Barrier Injury in Piglets. Adv Exp Med Biol. 2022, 1370, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Biehs, B.; Warming, S.; Leong, K.G.; Rangell, L.; Klein, O.D.; De Sauvage, F.J. A reserve stem cell population in small intestine renders Lgr5-positive cells dispensable. Nature. 2011, 478(7368), 255–259, Erratum in: Nature. 2012, 482, 120. PMID: 21927002; PMCID: PMC4251967. [Google Scholar] [CrossRef]
- Chia, L.A.; Kuo, C.J. The intestinal stem cell. Prog Mol Biol Transl Sci. 2010, 96, 157–173, PMID: 21075344; PMCID: PMC4165858. [Google Scholar] [CrossRef]
- Umar, S. Intestinal stem cells. Curr Gastroenterol Rep. 2010, 12, 340–348, PMID: 20683682; PMCID: PMC2965634. [Google Scholar] [CrossRef]
- Lou, W.; Liu, D. Hyperoxia reduces the expression of Bmi1 and Lgr5 in intestinal stem cells of neonatal rats. Basic&Clinical Medicine. 2019, 39, 22–26. [Google Scholar]
- Li, Y.; Ma, S.; Zhang, Y.; Yao, M.; Zhu, X.; Guan, F. (-)-Epicatechin mitigates radiation-induced intestinal injury and promotes intestinal regeneration via suppressing oxidative stress. Free Radic Res. 2019, 53, 851–864, Epub 2019 Jul 12. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, N.; Chartoumpekis, D.V.; Kensler, T.W. Crosstalk between Nrf2 and Notch signaling. Free Radic Biol Med. 2015, 88 (Pt B), 158–167, Epub 2015 May 21. PMID: 26003520; PMCID: PMC4628857. [Google Scholar] [CrossRef]
- Yagishita, Y.; McCallum, M.L.; Kensler, T.W.; Wakabayashi, N. Constitutive Activation of Nrf2 in Mice Expands Enterogenesis in Small Intestine Through Negative Regulation of Math1. Cell Mol Gastroenterol Hepatol. 2021, 11, 503–524, Epub 2020 Sep 5. PMID: 32896624; PMCID: PMC7797379. [Google Scholar] [CrossRef]
- Cao, S.; Wu, H.; Wang, C.; Zhang, Q.; Jiao, L.; Lin, F.; Hu, C. Diquat-induced oxidative stress increases intestinal permeability, impairs mitochondrial function, and triggers mitophagy in piglets. J Anim Sci. 2018, 96, 1795–1805, PMID: 29562342; PMCID: PMC6140957. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Chen, G.; Wang, R.; Peng, J. Oregano essential oil decreased susceptibility to oxidative stress-induced dysfunction of intestinal epithelial barrier in rats. J Funct Foods. 2015, 18, 1191–1199. [Google Scholar]
- Guo, W.; Li, W.; Su, Y.; Liu, S.; Kan, X.; Ran, X.; Cao, Y.; Fu, S.; Liu, J. GPR109A alleviate mastitis and enhances the blood milk barrier by activating AMPK/Nrf2 and autophagy. Int J Biol Sci. 2021, 17, 4271–4284, PMID: 34803497; PMCID: PMC8579459. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, NG.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; Langille, M.G.I.; Douglas, G.M.; Cheng, X.; Rouchka, E.C.; Waigel, S.J.; Dryden, G.W.; Alatassi, H.; Zhang, H.; Haribabu, B.; Vemula, P.K.; Jala, V.R. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat Commun. 2019, 10, 89, PMID: 30626868; PMCID: PMC6327034.51. [Google Scholar] [CrossRef]
- Wen, Z.; Liu, W.; Li, X.; Chen, W.; Liu, Z.; Wen, J.; Liu, Z. A Protective Role of the NRF2-Keap1 Pathway in Maintaining Intestinal Barrier Function. Oxid Med Cell Longev. 2019, 2019, 1759149, PMID: 31346356; PMCID: PMC6617875. [Google Scholar] [CrossRef]
- Chen, H.; Hu, Y.; Fang, Y.; Djukic, Z.; Yamamoto, M.; Shaheen, N.J.; Orlando, R.C.; Chen, X. Nrf2 deficiency impairs the barrier function of mouse oesophageal epithelium. Gut. 2014, 63, 711–719, Epub 2013 May 15. PMID: 23676441; PMCID: PMC3883925. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, J.; Chen, Z.; Huang, K.; Li, Y.; Li, C. Protective effects of sugar cane extract against oxidative damage induced by diquat and mechanism mediated by intestinal microbiota in weaned piglets. Journal of Nanjing Agricultural University. 2020, 3, 514–522. [Google Scholar]
- Baarlen, P.V.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215, Epub 2013 Feb 26. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr Opin Microbiol. 2013, 16, 255–261, Epub 2013 Jul 3. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat Med. 2018, 24, 392–400, PMID: 29634682; PMCID: PMC7043356. [Google Scholar] [CrossRef]
- Dai, J. Effects of Taurine on LPS mediated caecumepithelial tight junction protein, flora andmetabolites in piglets. Shenyang Agricultural University. 2022. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: the food connection. Front Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: human gut microbes associated with obesity. Nature. 2006, 444, 1022–3. [Google Scholar] [CrossRef] [PubMed]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Goossens, H.; Desager, K.N.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: a cross-sectional study. Gut Pathog. 2013, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Yu, Y.; Wan, J.; Mao, Y.; Zhang, H.; Zhang, J.; Tian, X.; Zhao, Q. Progress in research on the relationship between Proteobacteria and the imbalance of mammalian colonic intestinal flora. Chinese Journal of Microecology. 2022, 34, 479–484. [Google Scholar] [CrossRef]
- Råsbäck, T.; Jansson, D.S.; Johansson, K.E.; Fellström, C. A novel enteropathogenic, strongly haemolytic spirochaete isolated from pig and mallard, provisionally designated ‘Brachyspira suanatina’ sp. Environ Microbiol. 2007, 9, 983–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. Effects of Different Fermented Feeds on Growth Performance and Meat Quality and Intestinal Microorganism. Henan Agricultural University. 2019. [CrossRef]
- Bauchart-Thevret, C.; Stoll, B.; Chacko, S.; Burrin, D.G. Sulfur amino acid deficiency upregulates intestinal methionine cycle activity and suppresses epithelial growth in neonatal pigs. Am J Physiol Endocrinol Metab. 2009, 296, 1239–1250. [Google Scholar] [CrossRef]
- Li, L. Diversity and metabolic characteristics of lactic acid bacteria and lactate-degrading bacteria in fermented grains of Luzhou-flavor liquor. Jiangnan University. 2016.
- Song, Z.; Jiang, N.; Zhang, A. Research progress on biological functions of glutathione. Feed research. 2008, 9, 25–27. [Google Scholar]
- Liu, X. Antioxidative defence mechanism of glutathione on litopenaeus vannamei. Huazhong Agricultural University. 2010.
- Ziegler, D.M. Role of reversible oxidation-reduction of enzyme thiols-disulfides in metabolic regulation. Annu. Rev. Biochem. 1985, 54, 305–329. [Google Scholar] [CrossRef] [PubMed]
- Long, M. A study of effects of N-acetylcysteine on growth and viability of intestinal porcine epithelial cell and the related mechanisms. Wuhan Polytechnic University. 2014.
- Huang, R.; Peng, Z. Effect of diet type and dietary turine supplementation on growth performance of weaning pigs. Cereal Feed Ind. 2008, 9, 44–45. [Google Scholar]
- Spitze, A.R.; Wong, D.L.; Rogers, Q.R.; Fascetti, A.J. Taurine concentrations in animal feed ingredients; cooking influences taurine content. J Anim Physiol Anim Nutr (Berl). 2003, 87, 251–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yu, C.; Lin, M.; Fu, Y.; Zhang, L.; Meng, M.; Xing, S.; Li, J.; Sun, H.; Gao, F.; Zhou, G. Regulation of skeletal muscle protein synthetic and degradative signaling by alanyl-glutamine in piglets challenged with Escherichia coli lipopolysaccharide. Nutrition. 2015, 31, 749–56. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of Tau on mRNA abundance of anti-oxidase gene in liver. Significant difference * p ≤ 0.05,** p ≤ 0.01,*** p ≤ 0.001, **** p ≤ 0.0001.
Figure 1.
Effect of Tau on mRNA abundance of anti-oxidase gene in liver. Significant difference * p ≤ 0.05,** p ≤ 0.01,*** p ≤ 0.001, **** p ≤ 0.0001.
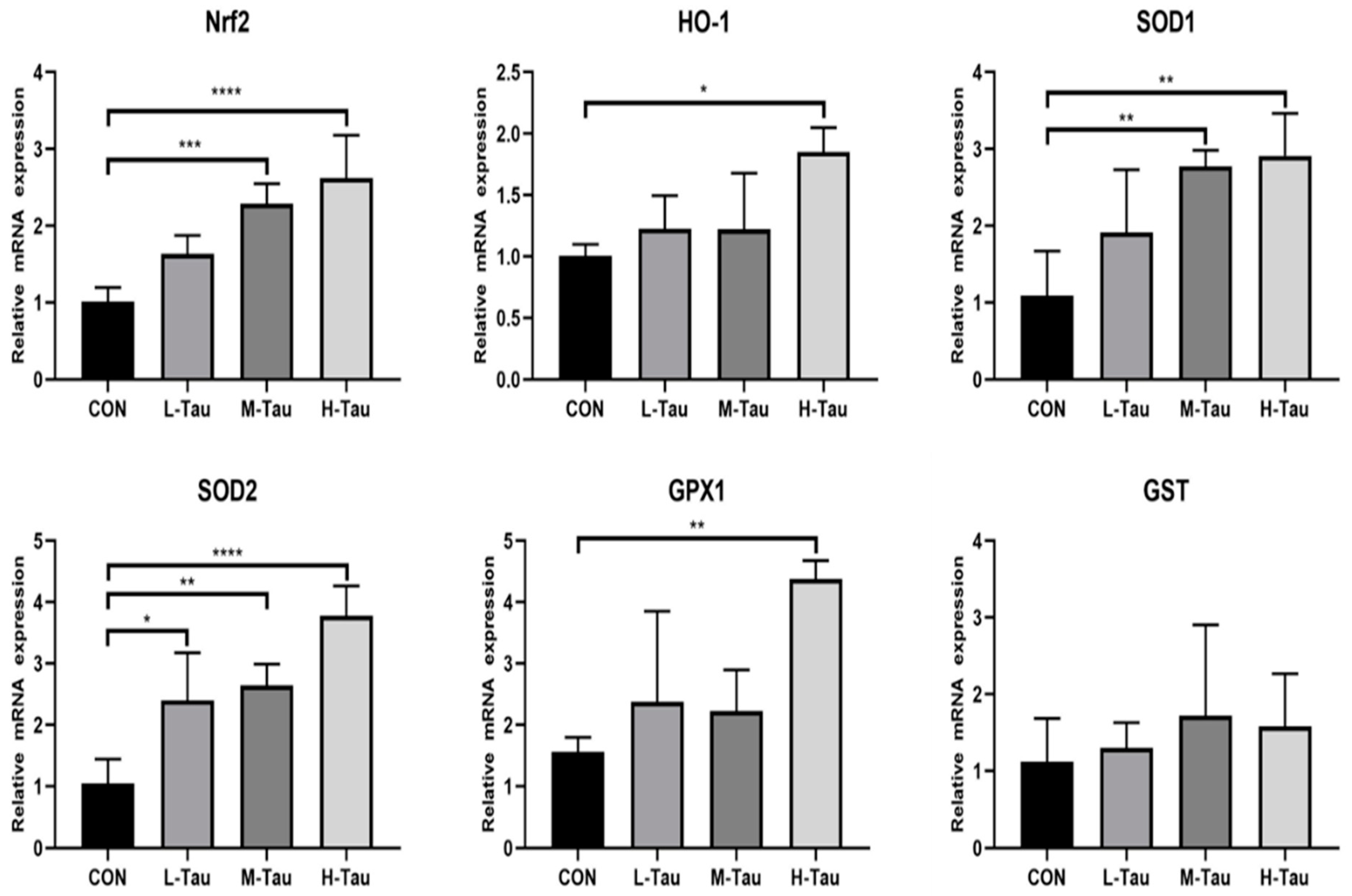
Figure 2.
Effect of Tau on intestinal tissue sections.
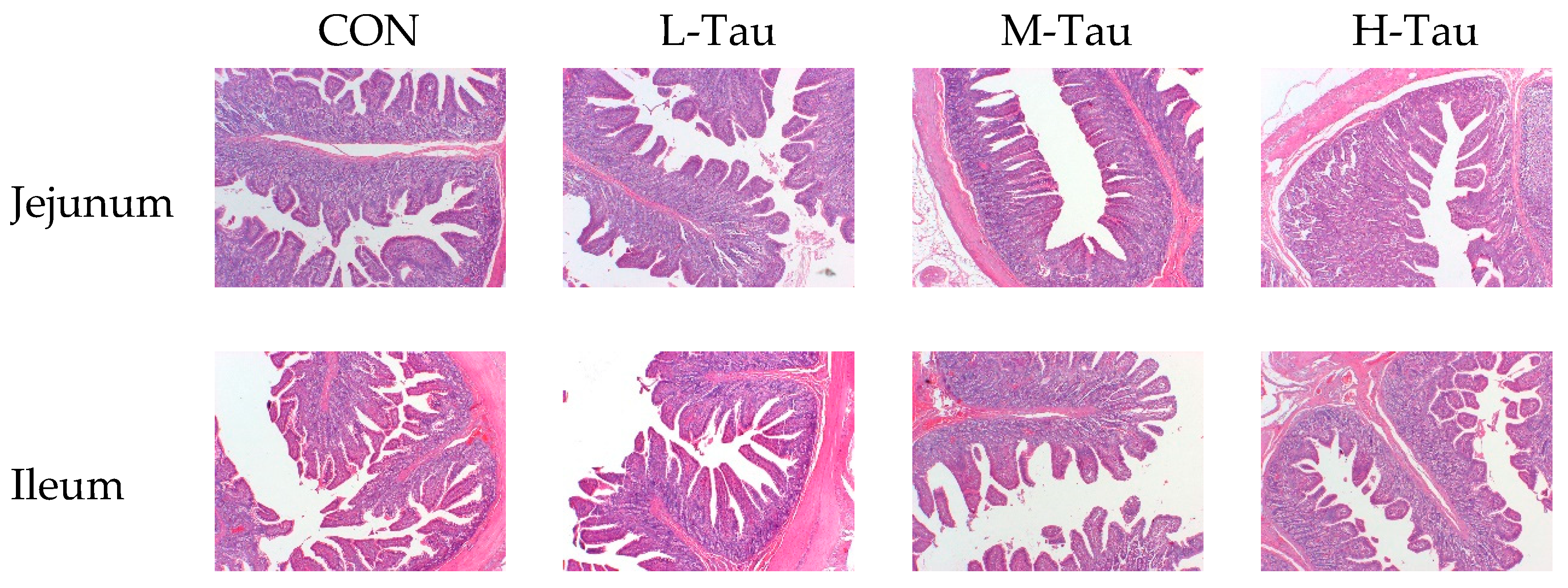
Figure 3.
Effect of Tau on mRNA abundance of tight junction protein gene in liver. Significant difference * p ≤ 0.05,** p ≤ 0.01,*** p ≤ 0.001, **** p ≤ 0.0001.
Figure 3.
Effect of Tau on mRNA abundance of tight junction protein gene in liver. Significant difference * p ≤ 0.05,** p ≤ 0.01,*** p ≤ 0.001, **** p ≤ 0.0001.
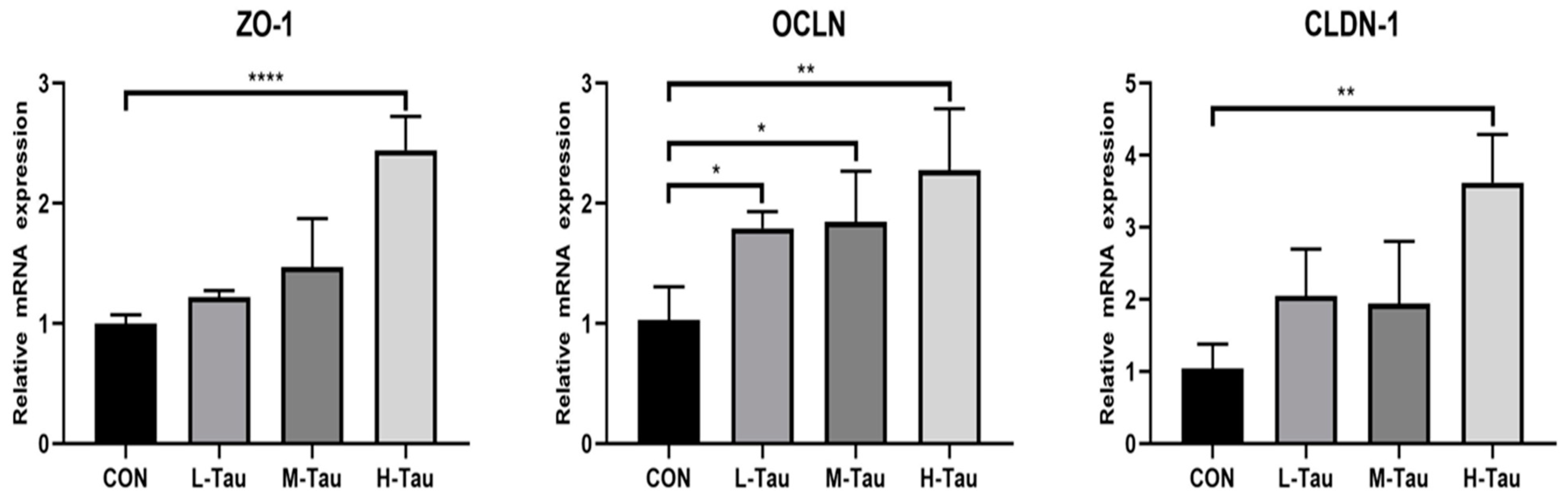
Figure 4.
Venn diagram of microbial ASV in cecal contents of weaned piglets.
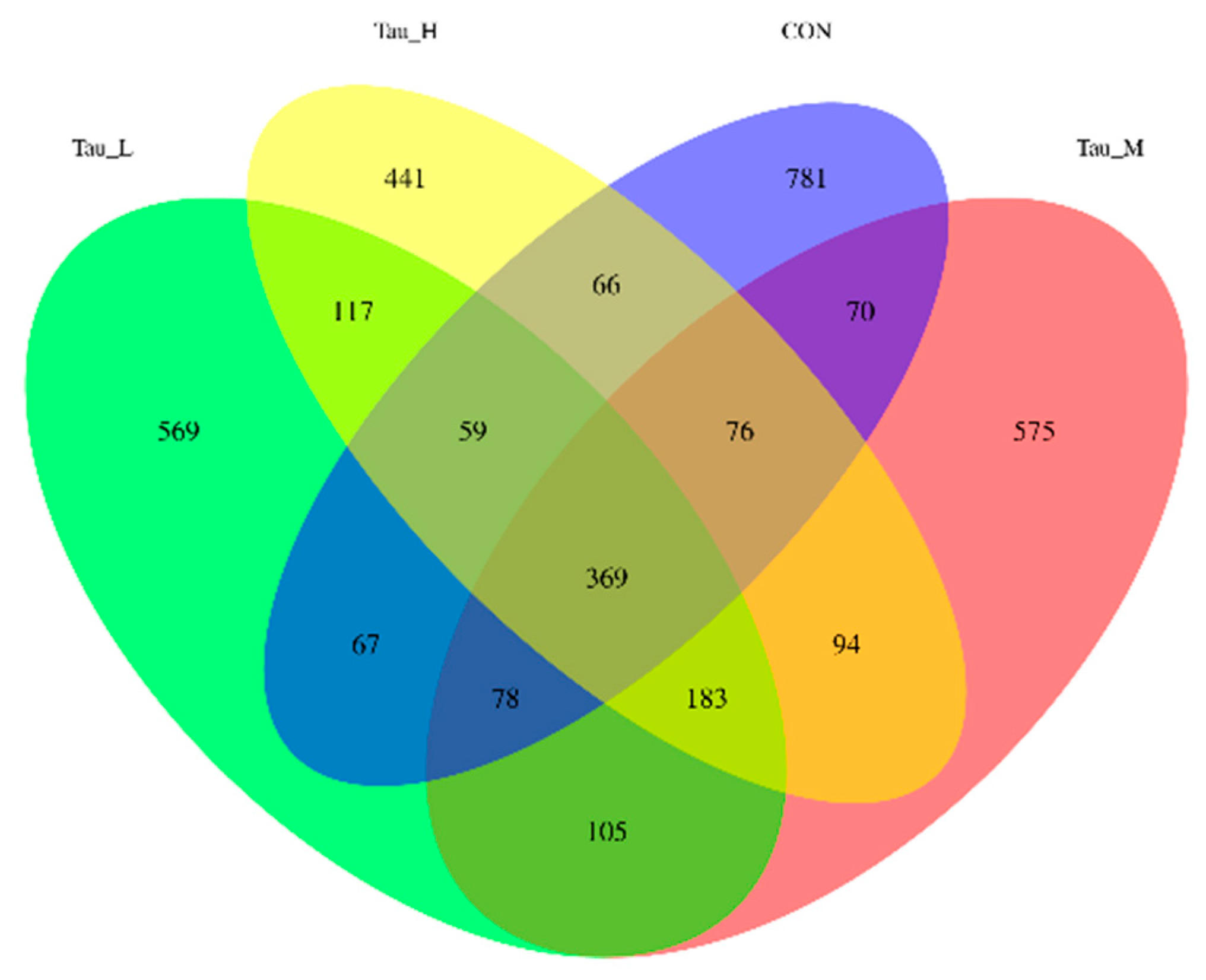
Figure 5.
Microbial clustering map of cecal contents at the phylum level of weaned piglets.
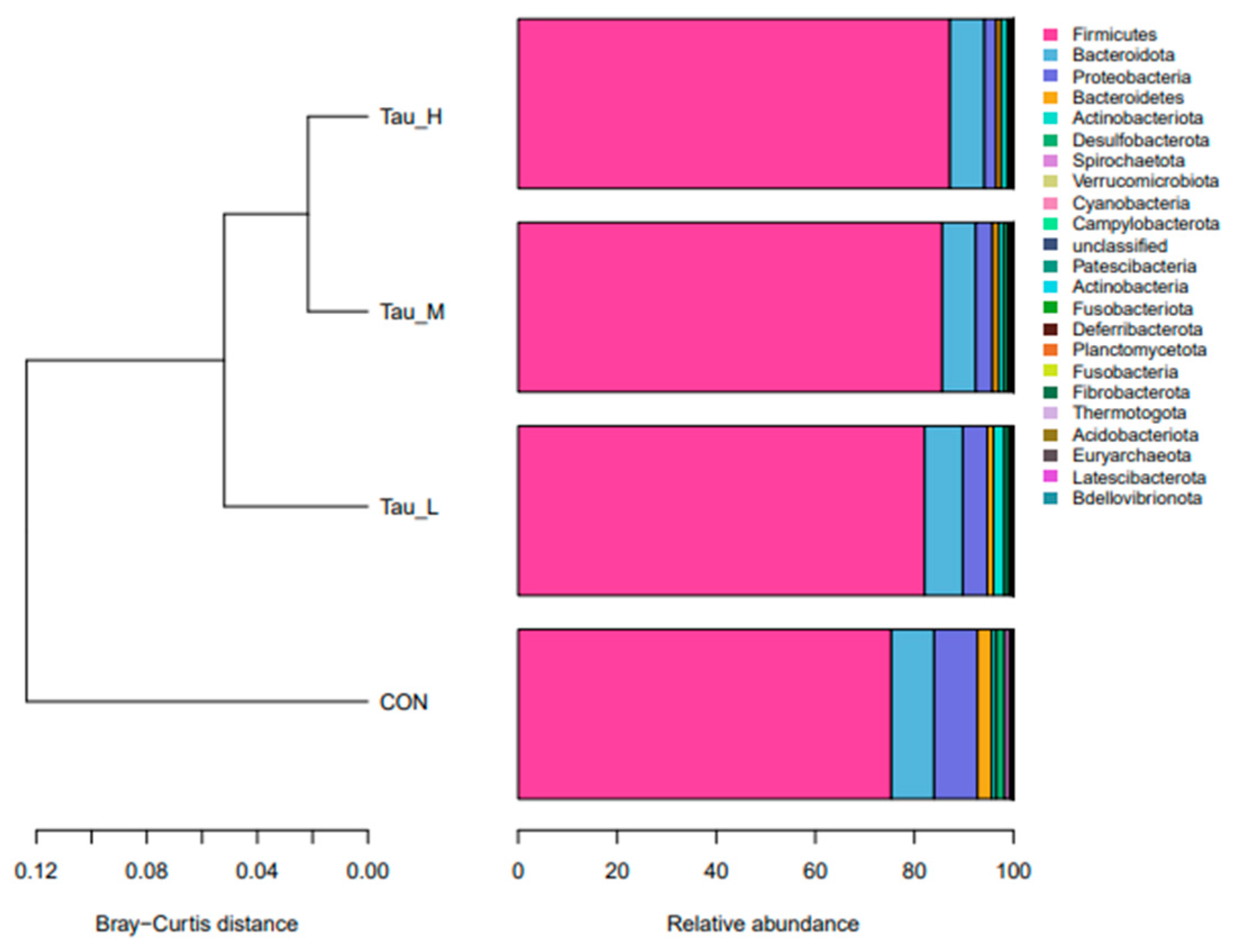
Figure 6.
Plot of microbial PCoA analysis of cecal contents of weaned piglets.
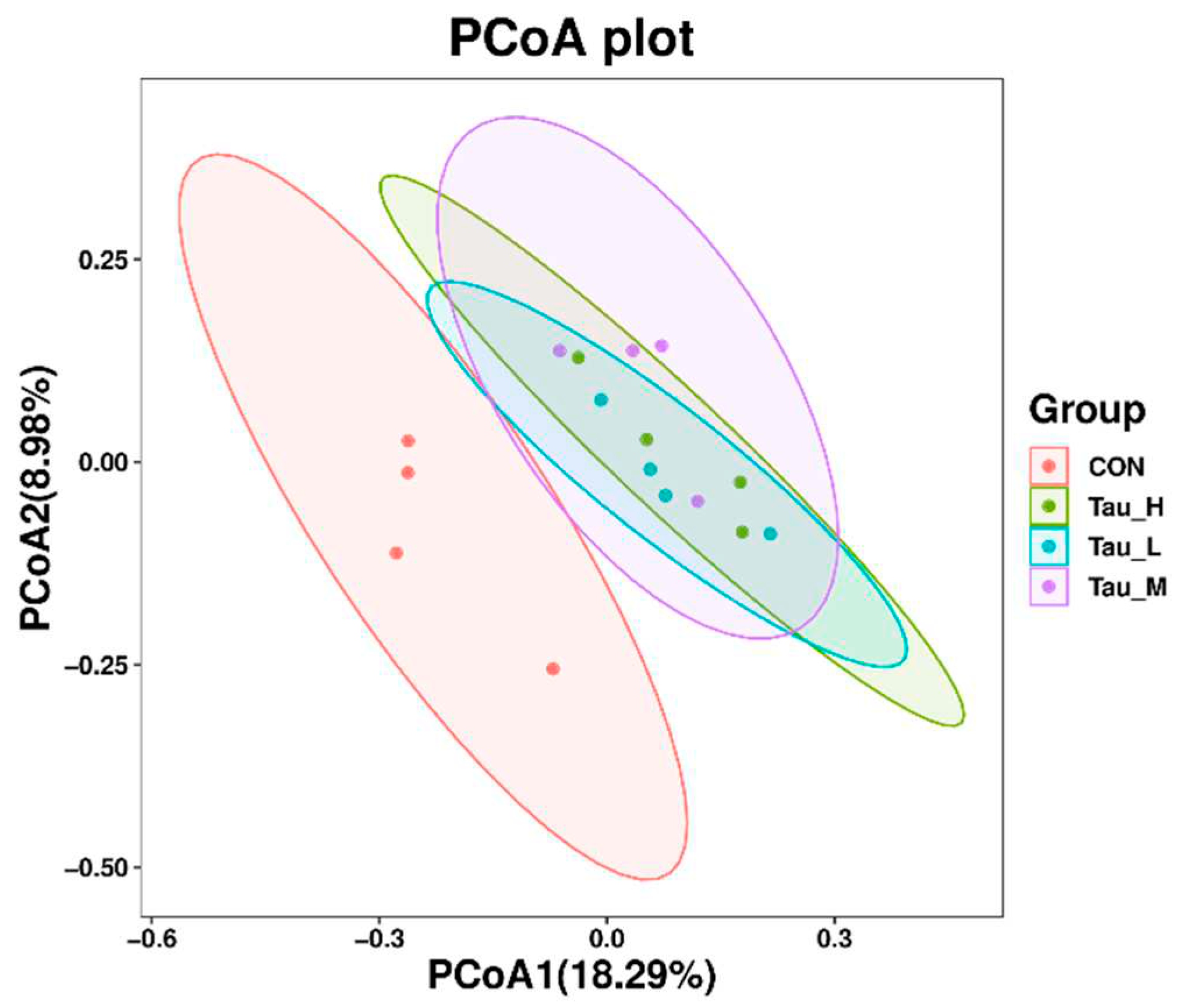

Table 1.
Ingredients and nutrient level of diets.
| Ingredients | Treatments2 | |||
| CON | L-Tau | M-Tau | H-Tau | |
| Corn (%) | 61.49 | 61.49 | 61.49 | 61.49 |
| Soybean meal (%) | 12.00 | 12.00 | 12.00 | 12.00 |
| Puffed soybean meal (%) | 6.08 | 6.08 | 6.08 | 6.08 |
| Fish meal (%) | 5.00 | 5.00 | 5.00 | 5.00 |
| Whey powder (%) | 10.00 | 10.00 | 10.00 | 10.00 |
| Fatty powder (%) | 1.59 | 1.59 | 1.59 | 1.59 |
| Calcium hydrophosphate (%) | 0.93 | 0.93 | 0.93 | 0.93 |
| Mountain flour (%) | 0.64 | 0.64 | 0.64 | 0.64 |
| NaCl (%) | 0.30 | 0.30 | 0.30 | 0.30 |
| L-lysine hydrochloride (%) | 0.57 | 0.57 | 0.57 | 0.57 |
| Methionine (%) | 0.21 | 0.21 | 0.21 | 0.21 |
| Threonine (%) | 0.20 | 0.20 | 0.20 | 0.20 |
| Trace mineral premix (%) | 1.00 | 1.00 | 1.00 | 1.00 |
| Total (%) | 100 | 100 | 100 | 100 |
| Trace mineral premix | ||||
| CuSO4·5H2O (%) | 0.24 | 0.24 | 0.24 | 0.24 |
| KI (%) | 0.01 | 0.01 | 0.01 | 0.01 |
| FeSO4·7H2O (%) | 5.05 | 5.05 | 5.05 | 5.05 |
| MnSO4·H2O (%) | 0.14 | 0.14 | 0.14 | 0.14 |
| Na2SeO3 (%) | 0.02 | 0.02 | 0.02 | 0.02 |
| ZnSO4·7H2O (%) | 4.53 | 4.53 | 4.53 | 4.53 |
| Vitamin complex1 (%) | 3.50 | 3.50 | 3.50 | 3.50 |
| C5H14ClNO (%) | 8.00 | 8.00 | 8.00 | 8.00 |
| Sweetening agent (%) | 6.00 | 6.00 | 6.00 | 6.00 |
| Antioxidant (%) | 2.00 | 2.00 | 2.00 | 2.00 |
| Tau (%) | 0.00 | 20.00 | 30.00 | 40.00 |
| Diluter (%) | 70.51 | 50.51 | 40.51 | 30.51 |
| Total (%) | 100 | 100 | 100 | 100 |
| Nutritional level | ||||
| DE(MJ/kg) | 14.34 | 14.34 | 14.34 | 14.34 |
| CP (%) | 18.42 | 18.42 | 18.42 | 18.42 |
| Ca (%) | 0.78 | 0.78 | 0.78 | 0.78 |
| Total phosphorus (%) | 0.67 | 0.67 | 0.67 | 0.67 |
| Available phosphorus (%) | 0.40 | 0.40 | 0.40 | 0.40 |
| Lysine (%) | 1.42 | 1.42 | 1.42 | 1.42 |
| Methionine (%) | 0.51 | 0.51 | 0.51 | 0.51 |
| Lysine + Cystine (%) | 0.78 | 0.78 | 0.78 | 0.78 |
| Threonine (%) | 0.90 | 0.90 | 0.90 | 0.90 |
| Tryptophan (%) | 0.21 | 0.21 | 0.21 | 0.21 |
1 Provided the following per kilogram of diet: 10500 IU VA; 3000 IU VD; 22.5 IU VE; 3.0 mg VK; 15 mg Pantothenic; 7.5 mg Riboflavin; 1.5 mg Folic Acid; 30.0 mg Niacin; 3.0 mg Thiamine; 4.5 mg VB6; 0.12 mg Biotin; 0.03 mg VB12. 2 Piglets were fed a basal diet with 0.0% (CON), 0.2% taurine (L-Tau), 0.3% taurine (M-Tau), and 0.4% taurine (H-Tau). The follow tables and figures were the same as this table.
Table 2.
Diarrhea score criteria of piglets [13].
Table 2.
Diarrhea score criteria of piglets [13].
| Degree | Appearance | Moisture | Score |
| Normal | Formed | <70 | 0 |
| Light | Soft | 70-75 | 1 |
| Intermediate | Semi-soild | 75-80 | 2 |
| Heavy | Liquid | >80 | 3 |
Table 3.
Primer sequences of the genes.
| Gene name | ID Number | Primer sequence | Product length (bp) |
| GAPDH | XM_021091114 | F: CGAGATCCCGCCAACATCAAR: CCCCACCCTTCAAGTGAGC | 109 |
| Nrf2 | MH101365.1 | F: TGCAGCTTTTGGCAGAGACAR: AGGAGCAATGAAGACTGGGC | 119 |
| ZO-1 | AJ318101.1 | F: ATTCCTGAGGGAATTGGGCAGR: GCAGGGTTTCACCTTTCTCCT | 94 |
| OCLN | NM_001163647.2 | F: ATATGGTGGAGAGACGCCCAR: AAGGGTAGCCCATACCACCT | 230 |
| SOD2 | NM_214127.2 | F: TTGCAGATTGCCGCTTGTTCR: TTACTTTTTGCAAGCGGCGT | 195 |
| GPX1 | NM_214201.1 | F: GAAGTGTGAGGTGAATGGCGR: TGCGATGTCATTGCGACACA | 145 |
| GST | NM_214300.2 | F: CTCAAGCTTTGGCAAGGGAGR: TGGGCTCTTCGTACACGTTC | 78 |
| HO-1 | NM_001004027.1 | F: TACCGCTCCCGAATGAACACR: GTCACGGGAGTGGAGTCTTG | 209 |
| SOD1 | NM_001190422.1 | F: AAGGCCGTGTGTGTGCTGAAR: AGTGGCCACACCATCTTTGC | 279 |
| CLDN-1 | XM_005670262.3 | F: CTAGTGATGAGGCAGATGAAR: AGATAGGTCCGAAGCAGAT | 250 |
Table 4.
Effect of Tau on antioxidant properties of body.
| Items | Treatments | SEM | P value | |||||
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Blood serum | ||||||||
| SOD (U/mL) | 209.04b | 197.46b | 248.75b | 388.57a | 46.51 | <.001 | 0.007 | 0.809 |
| GPX (U/mL) | 671.39b | 700.93b | 841.29a | 841.86a | 64.46 | 0.003 | 0.686 | 0.118 |
| CAT (U/mL) | 17.40b | 15.72b | 24.16a | 24.14a | 2.57 | 0.001 | 0.546 | 0.009 |
| POD (U/mL) | 6.25b | 6.52b | 8.75ab | 12.99a | 2.82 | 0.007 | 0.204 | 0.996 |
| T-AOC (μmol Trolox/mL) | 0.12c | 0.13bc | 0.15b | 0.19a | 0.02 | <.001 | 0.162 | 0.833 |
| O2•- clearance rate (%) | 17.35b | 16.96b | 17.81ab | 18.64a | 0.73 | 0.013 | 0.119 | 0.457 |
| •OH clearance rate (%) | 3.50b | 3.54b | 3.65b | 3.97a | 0.20 | <.001 | 0.187 | 0.787 |
| GSH (μmol/ mL) | 0.09ab | 0.08b | 0.13ab | 0.19a | 0.06 | 0.033 | 0.310 | 0.717 |
| GSSG (nmol/mL) | 3.67ab | 3.81a | 3.61ab | 2.71b | 0.62 | 0.050 | 0.138 | 0.810 |
| GSH/GSSG | 25.29b | 22.71b | 36.75ab | 80.79a | 30.52 | 0.019 | 0.145 | 0.845 |
| MDA (nmol/L) | 11.2a | 12.5a | 9.66ab | 6.97b | 1.86 | 0.003 | 0.053 | 0.320 |
| NO (μmoL/mL) | 0.34a | 0.34a | 0.25b | 0.24b | 0.04 | 0.001 | 0.935 | 0.100 |
| Liver | ||||||||
| O2•- clearance rate (%) | 17.37b | 16.99b | 18.21a | 18.24a | 0.42 | 0.004 | 0.396 | 0.021 |
| •OH clearance rate (%) | 3.38b | 3.64ab | 3.79ab | 4.00a | 0.23 | 0.005 | 0.867 | 0.763 |
| POD (U/mg Prot) | 4.71ab | 3.93b | 5.62ab | 6.62a | 1.41 | 0.040 | 0.252 | 0.372 |
| GSHS (U/g) | 0.42c | 0.39c | 0.49b | 0.58a | 0.04 | <.001 | 0.013 | 0.099 |
| Jejunal mucosa | ||||||||
| POD (U/mgProt) | 2.91b | 2.94b | 4.79ab | 6.33a | 1.76 | 0.011 | 0.429 | 0.621 |
| O2•- clearance rate (%) | 16.37b | 15.88b | 16.63b | 18.03a | 0.45 | <.001 | 0.002 | 0.572 |
| •OH clearance rate (%) | 3.36b | 3.48b | 3.91a | 4.20a | 0.20 | <.001 | 0.413 | 0.366 |
| GSHS (U/g) | 0.38b | 0.34b | 0.41b | 0.69a | 0.05 | <.001 | <.001 | 0.423 |
a, b, c Values with different letter superscripts within the same row mean significant difference (P<0.05). SOD: superoxide dismutase; GPX: glutathione peroxidase; CAT: catalase; POD: peroxidase; T-AOC: total antioxidant capacity; GSH: glutathione; GSSG: oxidized glutathione; MDA: malondialdehyde; NO: nitric oxide; GSHS: glutathione synthase.
Table 5.
Effect of Tau on intestinal morphology and structure of piglets.
| Items | Treatments | SEM | P value | |||||
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Jejunum | ||||||||
| Weight (g) | 277.01 | 309.64 | 279.62 | 335.56 | 53.33 | 0.245 | 0.670 | 0.237 |
| Weight index (g/kg) | 26.57 | 28.17 | 25.17 | 26.62 | 6.11 | 0.838 | 0.981 | 0.521 |
| Length (cm) | 744.50 | 785.00 | 787.50 | 857.50 | 132.48 | 0.271 | 0.828 | 0.728 |
| Length index (cm /kg) | 71.22 | 70.57 | 71.05 | 68.09 | 14.27 | 0.785 | 0.874 | 0.888 |
| VH (μm) | 235.95c | 286.75bc | 320.24ab | 368.00a | 47.55 | 0.003 | 0.952 | 0.774 |
| CD (μm) | 82.11 | 74.37 | 73.00 | 71.80 | 18.38 | 0.447 | 0.728 | 0.883 |
| VH/CD | 3.46b | 3.98b | 4.65ab | 6.17a | 1.19 | 0.011 | 0.432 | 0.800 |
| Ileum | ||||||||
| Weight (g) | 117.5 | 155.11 | 149.28 | 127.01 | 45.45 | 0.839 | 0.232 | 0.797 |
| Weight index (g/kg) | 11.03 | 13.98 | 12.87 | 10.03 | 3.67 | 0.649 | 0.159 | 0.783 |
| Length(cm) | 238.25 | 366.25 | 373.00 | 287.50 | 104.61 | 0.521 | 0.064 | 0.903 |
| Length index (cm/kg) | 22.79 | 32.77 | 32.49 | 22.68 | 8.34 | 0.974 | 0.035 | 0.970 |
| VH (μm) | 286.69b | 301.40b | 335.97ab | 384.53a | 46.13 | 0.019 | 0.512 | 0.957 |
| CD (μm) | 106.96 | 91.19 | 80.22 | 73.04 | 25.13 | 0.068 | 0.739 | 0.986 |
| VH/CD | 2.92b | 3.56b | 4.61ab | 5.81a | 1.13 | 0.004 | 0.650 | 0.915 |
a, b, c Values with different letter superscripts within the same row mean significant difference (P<0.05). VH: villus height; CD:crypt depth.
Table 6.
Effect of Tau on intestinal mucosal barrier of piglets.
| Items | Treatments | SEM | P value | |||||
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Blood serum | ||||||||
| D-LA (umol/L) | 122.51a | 107.10a | 94.74ab | 76.32b | 17.98 | 0.003 | 0.870 | 0.825 |
| Jejunal mucosa | ||||||||
| CLDN-1 (ng/g) | 586.88c | 632.24c | 1017.64b | 1469.29a | 126.29 | <.001 | 0.007 | 0.351 |
| ZO-1(ng/g) | 756.72b | 714.90b | 879.10b | 1291.94a | 133.31 | <.001 | 0.007 | 0.890 |
| OCLN (ng/g) | 39.78c | 32.09c | 66.02b | 136.85a | 9.54 | <.001 | <.001 | 0.841 |
a, b, c Values with different letter superscripts within the same row mean significant difference (P<0.05). D-LA: D-lactose; CLDN-1: closure protein 1; ZO-1: zonula occludens protein 1; OCLN: occludin.
Table 7.
Effect of Tau on cecal microflora diversity of piglets.
| Iteams | Treatments | SEM | P value | |||||
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Alpha diversityin | ||||||||
| observed_otus | 594.00 | 632.25 | 610.00 | 554.50 | 106.52 | 0.566 | 0.396 | 0.911 |
| shannon | 7.19 | 6.73 | 6.30 | 6.35 | 0.77 | 0.111 | 0.520 | 0.788 |
| simpson | 0.97 | 0.95 | 0.92 | 0.93 | 0.04 | 0.202 | 0.507 | 0.528 |
| chao1 | 595.23 | 641.05 | 618.02 | 555.85 | 110.07 | 0.577 | 0.346 | 0.906 |
| pielou_e | 0.78 | 0.72 | 0.68 | 0.70 | 0.07 | 0.084 | 0.319 | 0.775 |
| goods_coverage | 1.00 | 1.00 | 1.00 | 1.00 | 0.00 | 0.809 | 0.032 | 0.872 |
| Phylum levelin | ||||||||
| Firmicutes | 75.33b | 82.06a | 85.60a | 87.17a | 4.17 | 0.001 | 0.240 | 0.898 |
| Proteobacteria | 10.48a | 5.88ab | 4.25b | 2.84b | 3.23 | 0.005 | 0.342 | 0.710 |
| Bacteroidetes | 11.49 | 8.94 | 8.03 | 7.94 | 4.75 | 0.297 | 0.614 | 0.939 |
| Actinobacteriota | 0.94 | 2.23 | 1.06 | 1.26 | 0.82 | 0.914 | 0.216 | 0.060 |
| Spirochaetota | 0.95a | 0.18b | 0.22b | 0.11b | 0.33 | 0.006 | 0.07 | 0.233 |
| Verrucomicrobiota | 0.23 | 0.41 | 0.21 | 0.34 | 0.38 | 0.899 | 0.891 | 0.418 |
| Cyanobacteria | 0.09 | 0.11 | 0.52 | 0.16 | 0.28 | 0.340 | 0.191 | 0.085 |
| Generic level | ||||||||
| Clostridium_sensu_stricto_1 | 5.25b | 24.10a | 37.08a | 34.71a | 11.55 | 0.002 | 0.091 | 0.721 |
| Streptococcus | 13.49 | 5.92 | 2.72 | 5.42 | 6.89 | 0.101 | 0.162 | 0.922 |
| Lachnospiraceae | 6.08 | 5.96 | 4.56 | 2.65 | 4.48 | 0.266 | 0.696 | 0.938 |
| Terrisporobacter | 0.56b | 4.19a | 4.74a | 3.12ab | 1.93 | 0.081 | 0.019 | 0.836 |
| UCG-005 | 5.40a | 1.47b | 2.39b | 2.29b | 1.32 | 0.015 | 0.014 | 0.072 |
| T34_unclassified | 4.93 | 2.38 | 2.25 | 1.08 | 2.42 | 0.053 | 0.579 | 0.533 |
| Agathobacter | 3.33 | 2.53 | 2.44 | 1.62 | 1.69 | 0.190 | 0.990 | 0.707 |
| Turicibacter | 0.42b | 1.93ab | 3.59a | 3.39a | 1.45 | 0.007 | 0.263 | 0.543 |
| Ruminococcus | 3.13 | 1.37 | 1.75 | 1.87 | 1.22 | 0.234 | 0.146 | 0.395 |
| Clostridium_sensu_stricto_2 | 2.15 | 1.57 | 1.81 | 2.51 | 1.61 | 0.716 | 0.440 | 0.918 |
| Streptococcus | 0.53 | 2.52 | 1.74 | 1.78 | 1.45 | 0.380 | 0.203 | 0.292 |
a, b, c Values with different letter superscripts within the same row mean significant difference (P<0.05).
Table 8.
Effect of Tau on growth performance of piglets.
| Items | Treatments | SEM | P value | |||||
| CON | L-Tau | M-Tau | H-Tau | Linear | Quadratic | Cubic | ||
| Initial weight (kg) | 6.75 | 6.74 | 6.73 | 6.60 | 0.76 | 0.662 | 0.812 | 0.916 |
| Final weight (kg) | 12.79 | 13.09 | 13.22 | 13.65 | 1.27 | 0.147 | 0.868 | 0.783 |
| Weight gain (kg) | 6.04 | 6.44 | 6.49 | 7.05 | 1.14 | 0.067 | 0.828 | 0.583 |
| ADG (kg/d) | 201.20 | 214.70 | 216.45 | 235.17 | 38.10 | 0.067 | 0.828 | 0.583 |
| ADFI (kg/d) | 504.91 | 489.61 | 441.18 | 466.17 | 71.20 | 0.118 | 0.373 | 0.278 |
| Feed conversion rate | 0.40b | 0.45ab | 0.49a | 0.51a | 0.09 | 0.007 | 0.678 | 0.916 |
| Diarrhea index | 1.28a | 1.05b | 0.87bc | 0.72c | 0.22 | <.001 | 0.573 | 0.973 |
a, b, c Values with different letter superscripts within the same row mean significant difference (P<0.05). ADG: average daily gain; ADFI: Average daily feed intake.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



