
Submitted:
19 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
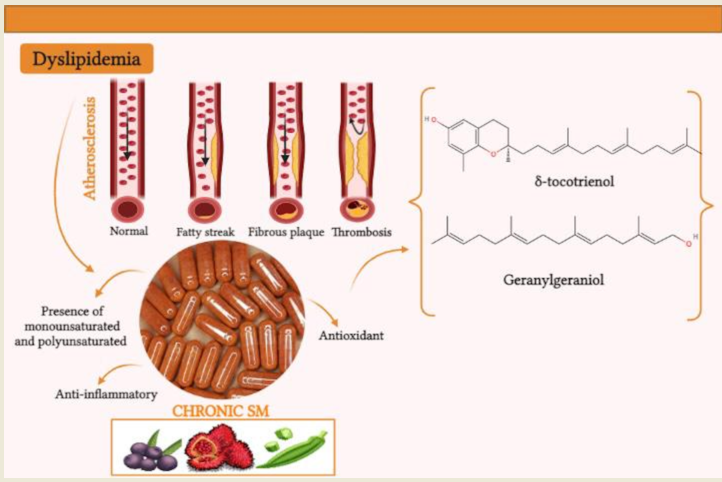
Dyslipidemia presents high levels of serum cholesterol, being characterized as a risk factor for cardiovascular diseases, especially for the development of atherosclerosis. E. oleracea (OFEO), A. esculentus (OFAE), B. orellana (OFBO) and granulated (CHR) oils are rich in bioactive compounds with potential to treat changes in lipid metabolism. This study investigated the effects of treatments with oils from A. esculentus, E. oleracea, B. orellana and granules (Chronic SM®) on saturated fat-induced dyslipidemia Cocos nucifera L. The chromatographic profile showed the majority of unsaturated fatty acids in the tested oils. The quantification of tocotrienols and geranylgeraniol was obtained in OFBO and CHR. Treatments with OFEO, OFAE, OFBO and CHR were able to significantly reduce glycemia with p>0.001, as well as hypertriglyceridemia, total cholesterol and LDL-cholesterol, in addition to increasing HDL-cholesterol. The treatments inhibited the formation of atheromatous plaques in the vascular endothelium of the treated rats. The obtained results suggest that the OFEO, OFAE, OFBO and CHR groups have antidyslipidemic effects and antiatherogenic activity.
Keywords:
Hypercholesterolemia
; Atherosclerosis
; vegetable oils
1. Introduction
Dyslipidemia occurs due to changes in the plasma concentration of low-density lipoproteins (LDL); high density lipoprotein (HDL); and triglycerides (TG) [1]. Dyslipidemia is classified as: isolated hypercholesterolemia, isolated hypertriglyceridemia, mixed hyperlipidemia and low HDL-c [2]. The increase in LDL-c and triglycerides concentrations, and the decrease in HDL-c concentrations is one of the main risk factors for cardiovascular diseases [3].
Many pathological conditions can compromise the cardiovascular system, such as atherosclerosis, a chronic immuno-inflammatory condition, triggered mainly by changes in lipid metabolism and reactive oxygen species (ROS) [4]. Atherosclerosis is characterized by hardening and progressive narrowing of blood vessels, due to the interaction of arterial wall elements, modified lipoproteins, T cells, macrophages and platelets; to the appearance of atherosclerotic plaque, which hinders the supply of blood to the tissues [5].
Multiple epidemiological studies associate diet composition with the main risk factors for dyslipidemias [6]. Currently, there are several drugs capable of controlling hyperlipidemia, but they are expensive and not free of side effects, so the search for therapeutic alternatives of natural origin constitutes promising sources of new functional bioactive substances, for the treatment or as a complement to the treatment. conventional treatment of chronic diseases, such as hyperlipidemias [7,8].
Thus, the population has opted for a diet with functional compounds that have cardioprotective properties, such as unsaturated fatty acids [7]. Unsaturated fatty acids (UFAs) are obtained from the diet through vegetable oils and are classified as monounsaturated (MUFA), with only one double bond, and polyunsaturated (PUFAs), containing two or more double bonds. The ingestion of unsaturated fatty acids can prevent and help the treatment of cardiovascular diseases, acting, therefore, in the reduction of the biochemical parameters, decreasing the plasmatic concentration of triglycerides through the decrease of the synthesis of triglycerides by the liver, as well as in the increase of the activity of the lipoprotein lipase (LPL), accelerating the catabolism of very low density lipoprotein (VLDL) and chylomicrons [9].
Numerous plant species produce oilseeds, such oils have in their composition bioactive properties of great potential in the cosmetics, food and pharmaceutical industry [10]. The vegetable oils from the seeds have essential fatty acids, free of trans fats or cholesterol and fat-soluble vitamins [11], these fatty acids play an activity of great biological importance for human health, acting as regulators of several key genes in the modulation of anti-inflammatory responses. hypercholesterolemic, antioxidant, as well as anti-inflammatory, thus triggering cardioprotective effects [12].
Among these promising oilseed species is Abelmoschus esculentus (L.) Moench. (okra), a shrub, with fruits in a greenish capsule, containing spherical seeds, with high mucilage. The seeds have oil at a concentration of 20 to 40%, which can be compared to the composition of oilseeds such as soybeans, peanuts, sesame [13,14]. The seeds are sources of vitamin E, a powerful natural antioxidant, of great nutritional importance [15], they have phenolic compounds, flavonoids, catechins and oligomeric derivatives as well as being sources of triacylglycerides, phytosterols and phospholipids, they have unsaturated fatty acids, mainly polyunsaturated fatty acids [16,17,18].
Euterpe oleracea Martius., popular açaí tree, is a fruit tree of great socioeconomic relevance in the Amazon, it has smooth globose fruits, violet in color, known as açaí. From the pulp of the fruit, a dark green oil with a distinct aroma reminiscent of açaí is obtained [19]. Açaí oil has 50% of the total dry matter of the pulp, has phenolic compounds, mainly anthocyanins, dietary fiber, in addition to being a good source of minerals such as potassium, magnesium, calcium, phosphorus, sodium, and vitamins E and B1, such as it also has a lipid profile rich in mono- and polyunsaturated fatty acids [20,21].
The species Bixa orellana Linné., known in Brazil as annatto, produces a capsular fruit inside, consisting of reddish orange seeds [22,23]. The oil extracted from the seeds of B. orellana is of great scientific interest due to its high amount of bioactive compounds. It presents in its composition, a lipid fraction of mono and polyunsaturated fatty acids of great biological importance [24]. Seed oil contains compounds such as alkaloids, flavonoids, carotenoids, in addition to having a large amount of tocotrienols, mainly δ-tocotrienol, as well as high concentrations of geranylgeraniol [25,26].
Several studies have attributed promising activities to unsaturated fatty acids, acting in the control and prevention of diseases, such as lipid metabolism disorders, especially in the development of dyslipidemia [27,28]. In this study, our objective was to investigate the possible mechanisms of action of molecular targets and the effects of treatments with oils from A. esculentus, E. oleracea, B. orellana and granules (Chronic SM®) on dyslipidemia induced by saturated fat Cocos nucifera L.in rats.
2. Results and Discussion
2.1. Chemical composition of Euterpe oleracea Martius oils; Abelmoschus esculentus L. Moench.
The ingestion of lipids in the human diet triggers an increase in free fatty acids that can raise cholesterol levels, which contributes to the process of dyslipidemia, being one of the main factors in the occurrence of cardiovascular diseases. The high concentration of saturated fatty acids is among the main lipids that favor hyperlipidemia, raise serum levels of triglycerides (TG), total cholesterol (TC) and low-density lipoprotein cholesterol (LDL-c), being responsible for the development of atherosclerosis and chronic inflammation in blood vessels [1,29].
The oils obtained from plant species present variations in their chemical composition according to the degree of unsaturation of each species, they are rich in essential fatty acids free of trans fats or cholesterol and fat-soluble vitamins. The chemical composition of vegetable oils in relation to the presence of fatty acids can determine the use of these oils for various purposes such as food and/or therapy [20].
Saturated fatty acids (SFA) are obtained in the diet, mainly from fats of animal and vegetable origin, and only have single bonds between the carbons. In turn, unsaturated fatty acids (UFA) can be obtained in the diet through vegetable and fish oils, they are classified as monounsaturated (MUFAs), with only one double bond, and polyunsaturated (PUFAs), containing two or more double bonds and belong to different series, being classified into ω-3, ω-6 and ω-9 [30].
In the analyzes by gas chromatography coupled to mass spectrometry (GC-MS) we can identify six compound retention peaks (Figure 1). According to the OFEO chromatographic data (Table 1), we observed a higher percentage of unsaturated fatty acids (67.83%), with polyunsaturated (5.9%) and monounsaturated (61.27%). Mostly we can find in its composition oleic acid (54.32%), palmitic acid (30.0%), linoleic acid (5.9%), followed by elaidic acid (4.29%), palmitoleic acid (2.62%) and stearic acid (2.29%). %). The data presented are in accordance with the literature, according to [31] Nascimento et al., (2008) 71% of unsaturated fatty acids are present, of which 60.81% are monounsaturated and 10.36% are polyunsaturated. Predominant in its composition oleic acid (56.2%) and palmitic acid (24.1%) [19].
Through chromatographic analysis of the fatty acids present in OFAE, it was possible to identify five compound retention peaks (Figure 2). We can observe a higher percentage of unsaturated fatty acids (61.05%), polyunsaturated (42.12%), monounsaturated (19.38%) (Table 2). This result is in line with other authors, since large amounts of unsaturated lipids (66.32%), especially oleic (20.38%) and linoleic (44.48%) acids, were found in okra grown in northeastern Brazil. Brazil [32], variations of linoleic acid (23.6 to 50.6%) and palmitic acid (10.03 to 36.03%) were also observed [33]. However, we observed in this work the presence mostly of linoleic acid (42.12%) and palmitic acid (33.55%), followed by oleic acid (17.85%), stearic acid (4.96%), gondoic acid (1.53%) in the OFAE, thus presenting form, similarity with those described in the literature.
The calibration curve obtained for total tocotrienols based on delta-tocotrienol presented R2= 0.9904, obtaining the following straight line equation: Y=5000000x-857622 (Figure 3). From the quantification of total tocotrienols it was possible to obtain an average concentration of 617.4 mg/mL, with an average content of 61.7% for OFBO and for CHR it was possible to obtain 46.7 mg/mL with an average content of 48, 7% of total tocotrienols (Table 3).
The calibration curve obtained for geranylgeraniol showed R2= 0.9989, obtaining the following straight line equation: Y=20000000x-400000 (Figure 4). From the quantification of geranylgeraniol present in the samples, it was possible to obtain an average concentration of 872.1 mg/mL for OFBO and an average content of 87.21% of geranylgeraniol, for CHR it was possible to obtain 141.6 mg/mL and an average content 14.16% of geranylgeraniol (Table 4).
2.2. Analysis of clinical and biochemical parameters
Diets rich in saturated fatty acids (SFA) increase insulin resistance and the incidence of cardiovascular disease. While those rich in mono- and polyunsaturated fatty acids play important roles in the adjuvant treatment of heart disease, coronary disease, hypertension, dyslipidemia, type 2 diabetes mellitus and insulin resistance [40,41,42].
MUFAs such as PUFAs act in the modulation of the lipid profile, especially the fatty acids of the ω-3, ω-6 and ω-9 series that are present in the composition of oils, these fatty acids are regulators of the expression of genes involved in lipid metabolism and glucose and adipogenesis, and may act via mediators, such as the peroxisome proliferator activated receptor (PPAR) (α, β and γ), hepatic X receptors (LXRs) (α and β), hepatic nuclear factor receptors (α and β), 4 (HNF4-α) and sterol regulatory element binding proteins (SREBPs) 1 and 2, being important factors in the hepatic metabolism of carbohydrates, fatty acids, triglycerides, cholesterol and bile acids [43,44,45].
Activation of the PPARγ transcription factor is essential in adipogenesis. Certain fatty acids of the ω-3 and ω-6 series can act as PPARγ ligands, and thus control adipogenesis, such as reducing cholesterol and triglyceride levels, in addition to contributing to the reduction of adipose tissue formation [46]. Thus, we can observe in our data, reduction of the body weight of the treatment groups CHR, OFEO, OFAE and OFBO. The reduction in body weight may be associated with the high levels of AGIs present in these oils and in the granules. The CHR (259.57±4.00) and OFBO (243.52±4.25) groups, both with p>0.001, had lower body weight and daily feed intake when compared to the GSC group (Table 5).
The supplementation of UFAs content is also associated with the reduction of hyperplasia and hypertrophy of adipocytes, in the modulation of metabolism through the stimulation of mitochondrial biogenesis and β-oxidation, which can lead to an induction of PPAR-α, consequently the control of differentiation and proliferation of adipose cells [47]. The VEI, SIN, OFEO, OFAE groups also showed significance of p>0.001 when comparing with GSC. The increase in body weight observed in the GSC group is related to the high daily calorie consumption of these animals in a diet composed of SFA, which contributes to the increase in adipose tissue [48].
Regarding the investigated biochemical markers of liver function (AST and ALT) and kidney function (urea and creatinine), it was possible to observe that supplementation with a GSC-induced diet caused a significant increase in the levels of these biochemical markers, both in the liver and in the kidneys (Table 6). However, the treatments positively influenced these results, especially the CHR and OFBO groups demonstrating statistically significant results.
The increase in AST and ALT levels may be due to the administration of saturated fat, associated with increased serum levels of blood cholesterol, as well as the accumulation of fat in the liver, the SFAs generate suppression of the activity of hepatic LDL receptors in messenger RNA (rLDL), blocking intracellular and extracellular cholesterol transport, since as a result of cholesterol reduction, cells via SREBP-2 exhibit increased expression of genes involved in cholesterol synthesis [49,50].
2.3. Analysis of lipid profile and atherogenic index
The lipid profile of the SIN, CHR, OFEO, OFAE and OFBO treatment groups showed significant results, both with ***p>0.001, when compared with the GSC group, which showed high levels of TC, LDL-c and TG (Figure 5). Coconut oil is a rich source of saturated fatty acids and its excess administration triggers the biosynthesis of cholesterol in the liver, causing hypercholesterolemia [51].
A study conducted in Wistar rats submitted to a treatment with coconut oil showed high levels of CT, TG, as well as an increase in LDL-c and a decrease in HDL-c [51]. According to the study [52], it was demonstrated that GSC has a satisfactory effect on the induction of dyslipidemia because it contains 92.64% of saturated fat, which may increase the rate of cholesterol production, as well as triglyceride levels, thus being a well-suited model. described for the induction of hyperlipidemia by GSC in animals.
In the present study, the data demonstrate that the CHR, OFEO, OFAE and OFBO groups act in the prevention of hyperlipidemia induced by GSC. This effect may be associated with the presence of unsaturated, monounsaturated (MUFAs) and polyunsaturated (PUFAs) fatty acids in the composition of the oils tested, as in the granules, it is known that these fatty acids act in the modulation of lipid metabolism at levels of transcription of receptors such as PPARα, present mainly in the liver, essential in the capture and transport of fatty acids and β-oxidation, inducing levels of lipoprotein lipase (LPL), which has the function of hydrolyzing lipoprotein triglycerides into circulating fatty acids [53].
In addition to the AGIs providing great benefits in the lipid profile, the saturated fatty acid stearic (18:0), present in the chemical composition of the species A. esculentus, E. oleracea, B. orellana does not promote hypercholesterolemia, since the dehydrogenation of this fatty acid is faster than chain elongation, causing it to be converted to oleic acid (monounsaturated) in the liver, thus promoting the previous peroxisomal β-oxidation [54].
Another possible response pathway in metabolic homeostasis is through the antioxidant activity indirectly attributed to polyunsaturated fatty acids, which are present in the chemical composition of the studied species. These fatty acids act on vascular endothelial cells, decreasing inflammation and, in turn, the risk of atherosclerosis and cardiovascular disease [55]. They act by preventing lipid peroxidation, eliminating reactive oxygen species (ROS) and reactive nitrogen species (RNS), which are responsible for the oxidation of LDL, a lipoprotein attributed to the formation of foam cells, consequently in the development of atherosclerosis [55,56]. ROS include superoxide anion, hydroxyl radical and hydrogen peroxide. RNS are mainly nitric oxide and peroxynitrite [57,58].
In a study carried out by Dias et al. (2014) [59] the polyphenols present in E. oleracea oil are agents capable of neutralizing reactive oxygen species, thus presenting antioxidant activity. Dong et al. (2014) [60] when obtaining the oil from the seeds of A. esculentus, demonstrated antioxidant activity by the DPPH radical scavenging activity test. The tocotrienols present in the B. orellana species also showed antioxidant properties, improving oxidative stress in metabolic disorders and protecting cellular functions [61], contributing to the decrease of antiproliferative effects, immunoprotection, reducing the risk of cancer and especially cholesterol [62,63]. Tocotrienols are present in CHR and OFBO.
Simvastatin was the drug chosen as the standard drug; it acts as a widely used hypocholesterolemic agent. It is an inactive lactone which is converted into its corresponding β,δ-hydroxy acid in its active form, acting on the pathway of hepatic metabolism by cytochrome P450 after oral administration [64].
Simvastatin known as statin, is a class of more effective drugs for the treatment of lipid alterations, it is a potent inhibitor of the enzyme 3-hydroxy-3-methyl-glutaryl-coenzyme A (HMGCoA) reductase, preventing the formation of mevalonate, which leads to a reduction in the hepatic synthesis of cholesterol, consequently acting in the increase of LDL-c receptors in hepatocytes, thus increasing its uptake from the circulation to restore cholesterol [65,66].
Regarding HDL-c, the results of this study showed a significant increase in the CHR, OFEO, OFAE and OFBO groups when compared with the GSC groups. The CHR group had the highest levels of HDL-c (34.87±3.4), followed by the OFBO group (32.13±4.36), OFAE (31.71±4.15), OFEO (31. 29±3.15), thus indicating that the treatments were able to prevent the increase in LDL-c in this experimental model. These results are possibly related to the lipid composition of the species under study, which present in their composition high levels of monounsaturated fatty acids such as oleic acid and eicosenoic acid (ω-9), and polyunsaturated fatty acids such as α-linoleic acid (ω- 3) and linoleic (ω-6), which can act in the treatment of cardiovascular diseases, as well as in dyslipidemia [30,67,68].
These unsaturated fatty acids are natural ligands of PPARα activation, they can increase the transport of fatty acids, increase the expression of lipoprotein lipase receptors (LPLr), suppress the levels of SREBP-1c, responsible for the regulation of enzymes involved in the synthesis of fatty acids, in addition to increasing the transcription of the main apolipoproteins, such as ApoAI and ApoAII, present in HDL. Unsaturated fatty acids can also inhibit HNF-4α, resulting in reduced expression of genes involved in cholesterol biosynthesis, resulting in addition to increasing the transcription of major apolipoproteins, such as ApoAI and ApoAII, present in HDL [69].
As for the glycemic levels, we can observe significant satisfactory results, the lowest glycemia values were presented in the CHR, OFEO, OFAE and OFBO groups, both with p>0.001 when compared to the GSC (Table 7). The increase in glycemia in the GSC group may be related to the ingestion of SFAs, corroborating the accumulation of body fat, since the excess consumption of carbohydrates and lipids contributes to resistance to insulin action, consequently leading to a chronic increase and gradual increase in glycemia [70].
In addition to its role in lipid homeostasis, PPARα can influence glucose homeostasis, acting by directly regulating gluconeogenesis via stimulation of pyruvate dehydrogenase kinase 4 (PDK4) expression, which favors the use of pyruvate for gluconeogenesis to the detriment of fatty acid synthesis [71]. In animal models of insulin resistance, PPARα agonists increased insulin sensitivity, consequently increasing fatty acid oxidation in the liver, skeletal muscle, and pancreas by endogenous glucose depletion [72].
Glucose homeostasis can also be attributed to the direct action of PPARγ on insulin-stimulated glucose disposal. Activation of PPARγ can increase the expression and translocation to the cell surface of the glucose transporters GLUT1 and GLUT4, thereby increasing glucose uptake into adipocytes and muscle cells, as well as reducing plasma glucose levels [73].
2.4. Formation of atherogenesis
Elevated levels of LDL-c in the blood are recognized as one of the risk factors for cardiovascular disease and are a consequence of the atherosclerosis process [60]. Atherosclerosis is a chronic inflammatory disorder that occurs in response to vascular endothelial aggression caused by an increased concentration of serum LDL in the arteries. After LDL-lipoprotein enters the intima of arteries, they undergo oxidation in the pro-oxidant environment, favored by an increase in reactive oxygen species (ROS) that oxidize LDL-cholesterol into oxidized LDL (oxLDL), thus triggering the process of inflammation and the recruitment of immune cells, especially monocytes, where they differentiate into macrophages and phagocytose the oxLDL, presenting lipids in their interior, and are now called foam cells, thus forming the atherosclerotic plaque [52,74,75].
The consumption of large amounts of saturated fat triggers the atherogenic process, especially in the abdominal aorta region, which is prone to plaque formation. The thoracic and abdominal arteries are the most affected during the process of atherosclerotic plaque formation [52,76].
The data presented in this study demonstrate compliance with the formation of atherosclerotic plaques in the vascular endothelium, as well as the Plasma Atherogenic Index (AI) presented by the GSC group (1.30±0.07) (Figure 6 and Table 7). However, when evaluating aortic atherogenesis, it was possible to demonstrate a great antiatherogenic potential of the CHR, OFEO, OFAE and OFBO treatment groups, due to the absence of plaque formation in the vascular endothelium, corroborating the data obtained from the AI of the treated groups.
This action can be explained by the capacity of the treatment groups (CHR, OFEO, OFAE and OFBO) to present unsaturated fatty acids in their chemical composition, especially of the ω-3 and ω-6 series, as natural ligands of gene targets, which act in biological processes mainly related to lipid metabolism and the inflammatory process, thus playing a key role in several cardiovascular diseases. Contributing effectively to the preservation of endothelial function, reduction of plasma cholesterol, reduction of LDL-c, which is an important lipoprotein in the process of early atherogenesis, and in the stability of the atheroma plaque [77].
The SIN group did not form atherosclerotic plaque. This response can be explained, as simvastatin acts to decrease the levels of oxLDL and macrophages, actions that stabilize the atheromatous plaque, in addition to inhibiting low molecular weight G proteins of the Ras superfamily (Ras and Rho), key proteins involved in cell proliferation, differentiation, apoptosis, migration, contraction and regulation of gene transcription, therefore improving vascular function. Simvastatins reduce the activity of NF-κB in inflammatory and vascular cells, as well as inflammatory biomarkers C-reactive protein (CRP), however they increase the endothelial expression of NO synthase (eNOS), increasing blood flow in the vessels, therefore improving the hemostasis and recovery of endothelium-dependent vasoreactivity [78].
Lecithin, the surfactant used, is described as a complex mixture derived from crude soybean oil, consisting mainly of phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine and phosphatidylinositol, including other lipophilic substances such as glycolipids, triglycerides or fatty acids, in addition to a hydrophilic fraction of phosphoric acid, glycerol, choline and inositol [79,80]. Soy lecithin is used as a dispersant, emulsifier, stabilizer, marketed in different solid pharmaceutical forms, such as capsules, tablets and granules, in the category of dietary supplements [81].
Due to their high lipophilicity, lecithins promote the physical coating of particles, especially those containing fat in their composition, in such a way that the oil, when in an aqueous medium, promotes a reduction in the surface tension between the solid and liquid phases, making these phases mix forming only one, having, therefore, important properties in the drug delivery system [82].
According to the literature, lecithins form fat transport lipoproteins, allowing to reduce blood cholesterol levels. The phospholipids present play an important role during the intestinal absorption of lipids, thus facilitating the formation of micelles, increasing absorption, in addition to having an activating action on the circulation, thus reducing the risk of cardiovascular diseases due to their emulsifying action, which does not allow the deposit of lipids to occur. fat in blood vessels [83,84].
The lecithin group (VEI) used in this study was able to reduce the lipid profile of TC, LDL and TG, and increase HDL showing significant values when compared to the GSC, but when comparing with the treatments (CHR, OFEO, OFAE and OFBO), it was not possible to obtain significant results. In addition to observing increased AI values, as well as the presence of atheroma plaque when analyzed in SEM, presenting, therefore, adverse results in the literature. Thus, we can infer that although lecithin presents responses in lipid levels, it did not demonstrate action in the reduction of atheromatous plaque, however, the treatment groups were more effective in modulating lipid metabolism, as well as antiatherogenic activity obtained from the species under study, this response may be related to the attributed bioavailability, especially to the studied granules (CHR).
AGIs can modulate the inflammatory response by several mechanisms, such as via PPARα which acts by inhibiting the expression of monocyte chemoattractant protein-1 (MCP-1) induced by C-reactive protein (CRP) inhibition of endothelin 1 (ET-1) expression, inhibition of interleukin-6 (IL-6) release induced by interleukin-1 (IL-1) and inhibition of expression of vascular cell adhesion molecule 1 (VCAM-1) induced by liposaccharides (LPS) [85,86,87].
PPARα and γ can also attenuate levels of interferon-gamma (IFN-γ) released by T lymphocytes; it also stimulates expression of the cholesterol efflux regulator protein ABCA1 in foam cells in a liver X receptor (LXR)-dependent manner, promoting ApoA-I-mediated cholesterol efflux; regulates class B type 1 scavenger receptors (SRBI), which plays a role in esterified HDL uptake by the liver and cholesterol efflux from macrophages; as well as induces the secretion of lipoprotein lipase (LPL) and decreases the uptake of glycated LDL by macrophages [87,88,89].
In turn, the PPARγ in macrophages increases the expression of the scavenger receptor CD36/FAT, responsible for the uptake of oxLDL, being of fundamental importance in the differentiation of macrophages with characteristics of foam cells [90]. Later stages of atherosclerosis are also regulated by PPARα, inhibiting the activation of smooth muscle cell (SMC) proliferation, which is a key event in the development of atherosclerosis and its complications such as tissue factor, an important procoagulant, hence PPARα may block atherothrombosis [91,92].
In addition to the AGIs promoting activity in the inflammatory response, compounds such as tocotrienols are important inflammatory modulators, as they are negative regulators of PPARγ receptors, and can act by suppressing the formation of nitric oxide inducible by an inflammatory signal in macrophages, tumor necrosis factor alpha (TNF-α), as well as inhibition of nuclear factor-κB (NF-κB) activation, thereby stopping tissue inflammation [34,93,94]. In addition to these compounds, geranylgeraniol, present in the chemical composition of CHR and OFBO, can also promote the modulation of PPARγ receptors and can act as an agonist, exert negative regulation via HMG-Coa-reductase and enzymes involved in cholesterol biosynthesis, playing a key role in the development of atherosclerosis [34,35,95].
Inflammation is recognized as a key regulatory process in the face of several risk factors for atherosclerosis. C-reactive protein (CRP) is the inflammatory marker, generated from the liver, it is an acute phase reactant protein, responds immediately after the increase of nonspecific inflammation in the blood vessel, which may reflect in the process of atherosclerotic development, therefore being, recommended as a cardiovascular risk marker [36,37,96]. In view of the results obtained in this study, we can observe increased CRP values in the GSC group (0.84±0.06), which corroborates the results by SEM in the appearance of atheromatous plaque, thus suggesting chronic inflammation in the aortic artery by atherogenic processes. However, we can observe decreased values of the CRP biomarker in the treatment groups CHR (0.41±0.04), OFEO (0.49±0.03), OFAE (0.50±0.03) and OFBO (0 .47±0.04), as well as in the positive control group SIN (0.39±0.04), demonstrating, therefore, a reduction in the inflammatory process and, consequently, antiatherogenic activity, observed in the images obtained from the aortas by SEM.
2.5. Histopathological analyzes
In the histopathological analyses, in this study, the hepatic lobules, portal spaces and well-delineated hepatic veins were easily visualized in the liver, and the hepatocytes formed confluent cords for the central-lobular vein.
The histopathological evaluation showed intense sinusoidal vascular congestion in animals from the GSC and SIN groups (Table 8). This alteration is characterized by the dilation of the hepatic sinusoids [38,39,97].
The occurrence of hepatic steatosis was also observed in the GSC and SIN groups. According to Oda (2008) [98] hepatic steatosis consists of the accumulation of lipids in the cytoplasm of hepatocytes (Figure 7). It is the most frequent hepatic metabolic disorder and results in an imbalance between the synthesis of triacylglycerols from fatty acids and their low secretion by the hepatocyte in the form of lipoproteins. Triacylglycerols accumulate in hepatocytes by one or more of the following pathways: increasing the amount of circulating free fatty acids; reduced β-oxidation, or decreased synthesis or secretion of very low-density lipoproteins, which is primarily responsible for the secretion of triacylglycerols by the liver [99,100].
3. Materials and Methods
3.1. Obtaining oils from Euterpe oleracea Martius.; Abelmoschus esculentus L. Moench.; Bixa orellana Linné and granulated (Chronic SM®).
The oils of Euterpe oleracea Martius. (OFEO), Abelmoschus esculentus L. Moench. (OFAE), Bixa orellana Linné (OFBO) and granules (Chronic SM®) were provided by Ages Bioactive Compounds Co. (São Paulo-SP, Brazil). The extraction method was standardized by the company and is under patent protection confidentiality.
3.2. Obtaining Cocos nucifera L. saturated fat (GSC)
The GSC was obtained from Cocos Empire Company - Mercado Municipal de Belo Horizonte, Minas Gerais, Brazil. The GSC extraction method was performed by mechanically pressing the endocarp, followed by the addition of water 1:1 (v/v) and fat separation by heating at 80 ºC [101].
3.3. Transesterification of oils from Euterpe oleracea Martius.; Abelmoschus esculentus L. Moench. by enzymatic catalysis
In a 25 mL vial, 500 mg of oils, 1.5 mL of ethanol and 50 mg of Candida antarctica lipase (CAL-B ≥ 5,000 U/g) were added. The reaction was maintained at 32°C under magnetic stirring for 24 h. Subsequently, it was transferred to a separatory funnel, the lower phase (glycerol) was separated, while the upper phase (mono-ester) was washed with distilled water (2 x 5 mL). The organic phase was dried with anhydrous sodium sulfate and filtered. Finally, the product was purified by flash silica gel column chromatography with a mixture of ethyl acetate and n-hexane (9:1) as eluent [102].
3.4. Chemical characterization of Euterpe oleracea Martius. oils; Abelmoschus esculentus L. Moench. by gas chromatography
Analyzes by gas chromatography coupled to mass spectrometry (GC-MS) were performed in a Shimadzu/GC 2010 device coupled to a Shimadzu/AOC-5000 auto-injector and a mass detector (Shimadzu MS2010 Plus) with electron impact (70 eV), equipped with a DB-5MS fused silica column (Agilent J&W Advanced 30 m x 0.25 mm x 0.25 µm). (65 kPa). The parameters were: split ratio 1:20, Helium as carrier gas, injection volume of 1.0 μL, injector temperature: 250 ºC, detector temperature at 250 ºC, initial column temperature: 120 ºC, remaining for 2 minutes, heating rate of 5°C/minute up to 270°C. The total analysis time was 36 minutes. The identification of fatty acid esters was identified by comparing the fragmentation spectrum with those contained in the GC-MS library (MS database, NIST 5.0) [103].
3.5. Quantitative analysis of total tocotrienols based on δ-tocotrienol in Bixa orellana Linné oil samples and granules (Chronic SM®).
For the quantitative analysis of the marker of interest, the samples were prepared with 2 solutions of the oil in hexane (Sigma-Aldrich, Lot. MKCF5755, GC grade, Germany) at a concentration of 2 mg/ml and a solution of 1 mg/ml in 10 mL volumetric flasks. Then, 1 mL aliquots of the solution were transferred to each vial and then 50 µL of BSTFA (Sigma-Aldrich, Lot. LC06356V, GC grade, Germany) was added, the derivatizing agent used to increase the volatility of the molecules.
After the derivatization process, the samples were injected into a gas chromatograph coupled to a mass spectrometer (Shimadzu GC/MS – PQ2010SE) equipped with an RTX-5MS fused silica column. For the quantification of total tocotrienols, calibration curves were prepared based on different concentrations that could be approximated by linear regression of the actual concentration of each sample.
The calibration curve was prepared from a 10 mg/mL stock solution of a standard containing 90% delta-tocotrienol and 10% gamma-tocotrienol in hexane. The chosen concentrations were 2.5 mg/mL, 1.0 mg/mL, 0.5 mg/mL, 0.25 mg/mL and 0.1 mg/mL. All injected standards were derivatized with BSTFA to increase the volatility of the molecules and amplify detection by the equipment.
3.6. Quantitative analysis of geranylgeraniol in samples of Bixa orellana Linné oil and granules (Chronic SM®).
For the quantitative analysis of the markers of interest, the samples were prepared in 2 solutions: a 1 mg/mL solution of the granules (Chronic SM®) and a solution containing 0.5 mg/mL of the standardized oil, both in dichloromethane (Sigma -Aldrich, Lot. MKCF5755, GC grade, Germany) in 10 mL volumetric flasks. Then, 1 mL aliquots of each solution were filtered and transferred to each vial.
The samples were injected into a gas chromatograph coupled to a mass spectrometer (Shimadzu GC/MS – PQ2010SE) equipped with an RTX-5MS fused silica column. For the quantification of geranylgeraniol, a calibration curve was prepared based on different concentrations that could be approximated by linear regression of the actual concentration of each sample.
The calibration curve was prepared from a stock solution of 5 mg/mL of a standard of geranylgeraniol in dichloromethane. The chosen concentrations were 4 mg/mL, 2.0 mg/mL, 1.0 mg/mL, 0.5 mg/mL, 0.25 mg/mL and 0.1 mg/mL.
3.7. Animals and Ethical Aspects
This study was approved by the Animal Use Ethics Committee of the Federal University of Amapá CEUA/UNIFAP under Protocol nº 03/2021 on June 2, 2021. The animals used were male Wistar rats (Rattus norvegicus albinus) from the Animal Investigation Multidisciplinary Center (CEMIB) from the University of Campinas-UNICAMP. Rats were kept in polyethylene cages placed in a ventilated, temperature-controlled cabinet (25 ± 2 °C), light/dark cycle (12/12 hours), with free access to standard rodent food and water ad libitum.
3.8. Treatments and Induction of Dyslipidemia
Hyperlipidemia was induced by 2ml of (GCS) daily for 40 days in all treatment groups. The animals were randomly divided into seven groups (n = 8 per group) and treated orally for 40 days: (1) treated with 0.5 mL of distilled water and 2 mL/day (GSC); (2) treated with vehicle-5% lecithin (VEI); (3) treated with simvastatin 20mg/kg (SIN); (4) treated with 200mg/kg of granules (CHR); (5) treated with 200mg/kg (OFEO); (6) treated with 200mg/kg (OFAE); (7) treated with 200mg/kg (OFBO).
3.9. Biochemical Analyzes
The animals' body weight and feed intake were measured every day throughout the treatment to calculate the weekly average. At the end of the experiment for the biochemical analyses, on the 41st day, the animals were fasted for 12 hours and anesthetized with a combination of Ketamine and Xylazine (Cristália- Chemicals and Pharmaceuticals Ltda, Brazil) at a dose of 60 and 100 mg/kg respectively intraperitoneally. Blood samples (1.5 ml) were collected from the ocular plexus and centrifuged for 10 minutes (5000 rpm) for analysis of aspartate aminotransferase (AST), alanine aminotransferase (ALT), total cholesterol (TC) and fractions (LDL and HDL), triglycerides (TG), urea, glucose and creatinine. The atherogenic index (AI) was calculated: Log (Triglycerides/HDL-Cholesterol) [109]. All tests were performed using LabTest kits and automated biochemical analysis equipment model BS 380 (Mindray Biomedicina Electronics Co., Ltd.).
3.10. Analysis by Scanning Electron Microscopy (SEM)
Aortic isolation was performed from the aortic arch to the iliac bifurcation. The thoracic region was divided into 0.5 cm sections for analysis using an equipment (SEM, Hitachi Model-TM3030PLUS) to detect atherogenic processes [56].
3.11. Histopathological Analyzes
For histopathological evaluation, the liver was fixed in a 10% buffered formalin solution for 48 hours and subsequently dehydrated in an alcohol battery, cleared in xylene and embedded in paraffin (in lentils, Inlab brand) according to the methodology described by [110].
Afterwards, the blocks were cut in a rotary microtome (Slee Medical) with disposable razors (Slee) at a thickness of 4 and 5µm. The cuts were arranged on glass slides for optical microscopy, which were left for 25 minutes in an oven at 60ºC for drying and better adhesion of the material that was stained with HE (Harris hematoxylin-LABORCLIN and yellowish eosin-INLAB). The slides were observed in optical microscopes Olympus-micronal BX41 and photographed with an MDCE-5C USB 2.0 camera (digital).
Hepatic alterations were evaluated and quantified on the following histological characteristics: sinusoidal vascular congestion, necrosis and hepatic steatosis.
The intensity of the histopathological characteristics was evaluated according to the methodology described by Rhoden et al., (2000) [111], in which the occurrences of alterations were expressed in crosses (0 to 3+), obtained through the average of three fields random microscopes, being evaluated at a magnification of 200 times, considering the following graduation:
- 0+: no changes
- 1+: slight intensity changes (less than 25% of the analyzed field);
- 2+: moderate intensity changes (25 to 50% of the analyzed field);
- 3+: severe intensity changes (more than 50% of the analyzed field).
The data obtained were expressed as mean ± standard deviation and statistically analyzed using the Kruskal-Wallis test, followed by the Dunn method, always considering a p<0.05.
3.12. Statistical analysis
The results of the dyslipidemia induction experiments were expressed as mean ± standard deviation. Groups were compared using Analysis of Variance (One-Way ANOVA) followed by Tukey's multiple comparison post-test and p<0.05 was considered statistically significant between groups. Statistical programs used were GraphPad Instat and Prism (version 7.0).
4. Conclusions
Based on the results, we report that the CHR, OFEO, OFAE and OFBO treatment groups can significantly reduce blood glucose, total cholesterol, triglycerides and LDL levels, as well as have been shown to increase the ability to synthesize HDL, in models of induced dyslipidemia. GSC. The antiatherogenic capacity of CHR, OFEO, OFAE and OFBO was demonstrated by the reduction of C-reactive protein, an inflammatory marker, absence of atheromatous plaque formation in the aorta, evaluated by scanning electron microscopy and atherogenic index.
The antihyperlipidemic and antiatherogenic potential of the tested groups can be explained by pharmacological mechanisms already described, based on their main chemical compounds, such as the presence of unsaturated fatty acids, in addition to the presence of tocotrienols and geranylgeraniol, it is known that these substances are reported as natural ligands of important targets in the transcriptional regulation of metabolic pathways, therefore promoting antioxidant, anti-inflammatory activities and modulation of the lipid profile.
5. Patents
The extraction method and the standardization of the oils and granules are under patent protection confidentiality by the company Ages Bioactive Compounds Co. (São Paulo-SP, Brazil).
Author Contributions
PF: Conceptualization, Methodology, Data Curation, Statistical Analysis, Writing and Editing. AL: Conceptualization, Methodology, Data Curation, Draft Writing. FC: Conceptualization, Methodology. AK: Conceptualization, Methodology. HO: Conceptualization, Methodology. GS: Conceptualization, Methodology, Statistical Analysis, Writing – revision and Editing. JCT: Supervision, Data Curation, Writing – Review and Editing.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Use Ethics Committee of the Federal University of Amapá CEUA/UNIFAP under Protocol No. 03/2021 on June 2, 2021.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful for the collaboration of Ages Bioactive Compounds Co. (São Paulo-SP, Brazil).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Souza, N.A.D.; Vieira, S.A.; Fonsêca, P.C.D.A.; Andreoli, C.S.; Priore, S.E., Franceschini, S.D.C.D.C. Familial dyslipidemia and factors associated with changes in the lipid profile in children. Science & Public Health. 2019, 24, 323-332.
- Duraipandiyan, V.; Al-Dhabi, N.A.; Irudayaraj, S.S.; Sunil, C. Hypolipidemic activity of friedelin isolated from Azima tetracantha in hyperlipidemic rats. Revista Brasileira de Farmacognosia. 2016, 26, 89–93. [Google Scholar] [CrossRef]
- Herrington, W.; Lacey, B.; Sherliker, P.; Armitage, J.; Lewington, S. Epidemiology of atherosclerosis and the potential to reduce the global burden of atherothrombotic disease. Circulation research. 2016, 118, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U. Oxidative stress in vascular disease: causes, defense mechanisms and potential therapies. Nature clinical practice Cardiovascular medicine. 2008, 5, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Datta, M.; Majumder, R.; Chattopadhyay, A.; Bandyopadhyay, D. Protective effect of melatonin in atherosclerotic cardiovascular disease: A comprehensive review. Melatonin Research. 2021, 4, 408–430. [Google Scholar] [CrossRef]
- Lottenberg, A.M.P. Importance of the dietary fat on the prevention and control of metabolic disturbances and cardiovascular disease. Arquivos Brasileiros de Endocrinologia & Metabologia 2009, 53, 595–607. [Google Scholar]
- ossain MA; Focken U; Becker K. Evaluation of an unconventional legume seed, Sesbania aculeata, as a dietary protein source for common carp, Cyprinus carpio L. Aquaculture. 2001, 198, 129-140.
- Gonçalves, M.D.C.R.; Diniz, M.D.F.; Borba, J.D.C.; Nunes, X.P.; Barbosa-Filho, J.M. Eggplant (Solanum melongena L.): myth or reality in the fight against the dislipidemy? Revista Brasileira de Farmacognosia. 2006, 16, 252–257. [Google Scholar]
- De Lima Viana, D.E.; Dantas, M.M.; da Silva Menezes, M.E. Omega-3 and omega-6 series fatty acids and their use in the treatment of cardiovascular diseases: a review. Revista Saúde & Ciência Online. 2016, 5, 65–83. [Google Scholar]
- Sarquis, I.R.; Sarquis, R.S.; Marinho, V.H.; Neves, F.B.; Araújo, I.F.; Damasceno, L.F.; Ferreira, I.M. Carapa guianensis Aubl.(Meliaceae) oil associated with silk fibroin, as alternative to traditional surfactants, and active against larvae of the vector Aedes aegypti. Industrial Crops and Products 2020, 157, 112931. [Google Scholar] [CrossRef]
- Prado, J.M.; Assis, A.R.; Maróstica-Júnior, M.R.; Meireles, M.A.A.P.R. Manufacturing cost of supercritical-extracted oils and carotenoids from Amazonian plants. Journal of food process engineering 2010, 33, 348–369. [Google Scholar] [CrossRef]
- Infante, R.E.; Radhakrishnan, A. Continuous transport of a small fraction of plasma membrane cholesterol to endoplasmic reticulum regulates total cellular cholesterol. Elife. 2017, 6, e25466. [Google Scholar] [CrossRef]
- Arapitsas, P. Identification and quantification of polyphenolic compounds from okra seeds and skins. Food Chemistry. 2008, 110, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Paes HMF; Esteves BDS; Sousa EFD. Determination of water requirement of okra in Campos dos Goytacazes, R.J. Revista Ciência Agronômica. 2012, 43, 256-261.
- Al-Kanani, E.A.S.; Al-Hilifi, S.A.H.; Al-Kareem, AH. The nutritional composition and vitamin E of three Iraqi okra (Abelmoschus esculentus L.) seeds oil. In: IOP Conference Series: Earth and Environmental Science. IOP Publishing. 2019, 012058.
- Holser, R.A.; Bost, G.; Vanboven, M. Phytosterol composition of hybrid hibiscus seed oils. Journal Agricultural and Food Chemistry. 2004, 52, 2546–2548. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yu, W.; Li, Y.; Prasad, N.; Tang, Z. Antioxidant activity of extract and its major constituents from okra seed on rat hepatocytes injured by carbon tetrachloride. BioMed research international 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Petropoulos, S.; Fernandes, Â.; Barros, L.; Ciric, A.; Sokovic, M.; Ferreira, I.C. Chemical composition, nutritional value and antioxidant properties of Mediterranean okra genotypes in relation to harvest stage. Food Chemistry. 2018, 242, 466–4743. [Google Scholar] [CrossRef]
- Pacheco-Palencia, L.A.; Talcott, S.T.; Safe, S.; Mertens-Talcott, S. Absorption and biological activity of phytochemical-rich extracts from acai (Euterpe oleracea Mart.) pulp and oil in vitro. Journal of agricultural and food chemistry. 2008, 56, 3593–3600. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.O.; Silva, L.S.; de Brito Magalhães, C.L.; de Figueiredo, B.B.; Costa, D.C.; Silva, M.E.; Pedrosa, M.L. The hypocholesterolemic activity of acai (Euterpe oleracea Mart.) is mediated by the enhanced expression of the ATP-binding cassette, subfamily G transporters 5 and 8 and low-density lipoprotein receptor genes in the rat. Nutrition Research. 2012, 32, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Matta, F.V.; Grace, M.; Lila, M.A.; Ward, N.I.; Felipe-Sotelo, M.; Esposito, D. Phenolic content, anti-inflammatory properties, and dermal wound repair properties of industrially processed and non-processed acai from the Brazilian Amazon. Food & Function 2020, 11, 4903–4914. [Google Scholar]
- Vilar, D.A.; Vilar, M.A.S.; Moura TFAL; Raffin, F.N.; Oliveira MR; Franco CFO. Tradicional uses, chemical constituints, and biological activities of Bixa orellana L.: a review. The Scientific World Journal. 2014, 2014: 857292.
- Aparecido, L.E.D.O.; Rolim, G.D.S.; Moraes, J.R.D.S.C.D.; Rocha, H.G.; Lense, G.H.E.; Souza, P.S. Agroclimatic zoning for urucum crops in the state of Minas Gerais, Brazil. Agrometeorology 2012, 77, 193–200. [Google Scholar] [CrossRef]
- Rão, P.P.; Rao, G.N.; Jyothirmayi, T.; Satyanarayana, A.; Karuna, M.S.L.; Prasad, R.B.N. Characterisation of Seed Lipids from Bixa orellana and Trachyspermum copticum. J Am Oil Chem Soc. 2015, 92, 1483–1490. [Google Scholar]
- Santos, O.V.; Corrêa, N.C.F.; Soares, F.A.S.M.; Gioielli, L.; Costa, C.E.F.; Lannes, S.C.S. Chemical evaluation and thermal behavior of Brazil nut oil obtained by different extraction processes. Food Research International. 2012, 47, 253–258. [Google Scholar] [CrossRef]
- Costa, C.K.; Silva, C.B.; Lordello, A.L.L.; Zanin, S.M.W.; Dias, J.F.G.; Miguel, M.D.; Miguel, O.G. Identification of δ tocotrienol and fatty acids in annatto (Bixa orellana Linné) fixed oil. Brazilian Journal of Medicinal Plants. 2013, 15, 508–512. [Google Scholar]
- Mohamed, S. Functional foods against metabolic syndrome (obesity, diabetes, hypertension and dyslipidemia) and cardiovasular disease. Trends in Food Science & Technology 2014, 35, 114–128. [Google Scholar]
- Guha, L.; Bashir, A.; Bhat, I. Unhealthy Heart (Hypercholesterolemia) and Naturally Available Medications: A Comprehensive Review. Biological Sciences. 2022, 2, 172–186. [Google Scholar] [CrossRef]
- Précoma, D.B.; Oliveira, G.M.M.D.; Simão, A.F.; Dutra, O.P.; Coelho, O.R.; Izar, M.C.D.O.; Mourilhe-Rocha, R. Updated cardiovascular prevention guideline of the Brazilian Society of Cardiology-2019. Arquivos brasileiros de cardiologia. 2019, 113, 787–891. [Google Scholar] [CrossRef] [PubMed]
- Nasciutti, P.R.; Costa, A.P.; Júnior, M.S.; Melo, N. ; Carvalho, R Fatty acids and the cardiovascular system. Biosphere Encyclopedia. 2015, 11. [Google Scholar]
- Nascimento, R.J.S.; Couri, S.; Antoniassi, R.; Freitas, S.P. Fatty acids composition of açaí pulp oil obtained by enzymatic technology and hexane. Revista Brasileira de Fruticultura. 2008, 30, 498–502. [Google Scholar] [CrossRef]
- Soares, G.S.F.; Gomes, V.M.; Albuquerque, A.; Dantas, M.B.; Rosenhain, R.; Souza, A.G.; Persunh, D.C.; et al. Spectroscopic and thermooxidative analysis of organic okra oil and seeds from Abelmoschus esculentus. The Scientific World Journal 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Jarret, R.; Wang, M.L.; Levy, I.J. Seed oil and fatty acid content in okra (Abelmoschus esculentus) and related species. Journal of agricultural and food chemistry. 2011, 59, 4019–4024. [Google Scholar] [CrossRef] [PubMed]
- Batista, M.A.; Santos, A.V.T.L.T.; Nascimento, A.L.; Moreira, L.F.; Souza, I.R.S.; Silva, H.R.; Carvalho, J.C.T. Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules 2022, 27, 1584. [Google Scholar] [CrossRef]
- Soricr, G.P.; Folli, F. A combination of PPAR-γ agonists and HMG CoA reductase inhibitors (statins) as a new therapy for the conservative treatment of AAS (aortic aneurysm syndromes). Medical hypotheses. 2009, 73, 614–618. [Google Scholar]
- Zhnag, W. High-sensitivity C-reactive protein modifies the cardiovascular risk of lipoprotein (a) multi-ethnic study of atherosclerosis. Journal of the American College of Cardiology. 2021, 78, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Toyota, K.; Waku, T.; Hirakawa, Y.; Nagasawa, N.; Kasuga, J.I.; Hashimoto, Y.; Miyachi, H.; Morikawa, K. Adaptability and selectivity of human peroxisome proliferator-activated receptor (PPAR) pan agonists revealed from crystal structures. Acta Crystallogr D Biol Crystallogr. 2009, 65, 786–65. [Google Scholar] [CrossRef] [PubMed]
- Ogbe, R.J.; Agbese, S.P.; Abu, A.H. Protective effect of aqueous extract of Lophira lanceolata leaf against cisplatin-induced hepatorenal injuries and dyslipidemia in Wistar rats. Clinical Phytoscience. 2020, 6. [Google Scholar] [CrossRef]
- Fernández-Martinez, E. et al. Dietary chia seeds (Salvia hispanica) improve acute dyslipidemia and steatohepatitis in rats. Journal of food biochemistry 2019, 43, 12986. [Google Scholar] [CrossRef] [PubMed]
- Magriplis, E. Trans fatty acid intake increases likelihood of dyslipidemia especially among individuals with higher saturated fat consumption. Reviews in Cardiovascular Medicine. 2022, 23, 130. [Google Scholar] [CrossRef]
- eegala, S M; Willett, W.C.; Mozaffarian, D. Consumption and health effects of trans fatty acids: a review. Journal of AOAC International. 2002, 92, 1250-1257.
- Lombardo, Y.B.; Chicco, A.G. Effects of dietary polyunsaturated n-3 fatty acids on dyslipidemia and insulin resistance in rodents and humans. A review. The Journal of nutritional biochemistry. 2006, 17, 1–13. [Google Scholar] [CrossRef]
- Hertz, R.; Magenheim, J.; Berman, I.; Bar-Tana, J. Fatty acyl-CoA thioesters are ligands of hepatic nuclear factor-4α. Nature. 1998, 392, 512–516. [Google Scholar] [CrossRef]
- Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocrine reviews. 1999, 20, 649–688. [Google Scholar]
- Ou, J.; Tu, H.; Shan, B.; Luk, A.; DeBose-Boyd, R.A.; Bashmakov, Y.; Brown, M.S. Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proceedings of the National Academy of Sciences. 2001, 98, 6027–6032. [Google Scholar] [CrossRef]
- Queiroz, J.C.F.D.; Alonso-Vale, M.I.C.; Curi, R.; Lima, F.B. Control of adipogenesis by fatty acids. Brazilian Archives of Endocrinology & Metabology 2009, 53, 582–594. [Google Scholar]
- Hensler, M.; Bardova, K.; Jilkova, Z.M.; Wahli, W.; Meztger, D.; Chambon, P.; Flachs, P. Inhibition of fat cell proliferation by n-3 fatty acids in dietary obese mice. Lipids in health and disease. 2011, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.K.; Ding, J.; Lee, J. S; Garcia, M.; Kanaya, A.M.; Tylavsky, F.A.; Kritchevsky, S.B. Dietary fat and cholesterol and risk of cardiovascular disease in older adults: the Health ABC Study. Nutrition, metabolism and cardiovascular diseases. 2011, 21, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, A.; Kalita, H.; Kalita, M.C.; Devi, R. Withdrawal from high-carbohydrate, high-saturated-fat diet changes saturated fat distribution and improves hepatic low-density-lipoprotein receptor expression to ameliorate metabolic syndrome in rats. Nutrition. 2017, 38, 5–101. [Google Scholar] [CrossRef]
- Shen, Y.; Zhao, Z.; Zhang, L.; Shi, L.; Shahriar, S.; Chan, R.B.; Min, W. Metabolic activity induces membrane phase separation in endoplasmic reticulum. Proceedings of the National Academy of Sciences. 2017, 114, 13394–13399. [Google Scholar] [CrossRef] [PubMed]
- Celeski, I.C.; Fergutz, J.K.; Pereira, E.M.; Zétola, M.; Bazzo, G.C.; Pezzini, B. R. Evaluation of the induction of hyperlipidemia in rats by dexamethasone and coconut oil. Infarma. 2013, 25, 126–131. [Google Scholar] [CrossRef]
- Faria e Souza, B.S.; Carvalho, H.O.; Taglialegna, T.; Barros, A.S.A.; da Cunha, E.L.; Ferreira, I.M.; Carvalho, J.C.T. Effect of Euterpe oleracea Mart.(AÇAÍ) oil on dyslipidemia caused by Cocos nucifera L. saturated fat in wistar rats. Journal of medicinal food 2017, 20, 830–837. [Google Scholar] [CrossRef]
- Duez, H.; Lefebvre, B.; Poulain, P.; Torra, I.P.; Percevault, F.; Luc, G.; Staels, B. Regulation of human ApoA-I by gemfibrozil and fenofibrate through selective peroxisome proliferator-activated receptor α modulation. Arteriosclerosis, thrombosis, and vascular biology 2005, 25, 585–591. [Google Scholar] [CrossRef]
- Lottenberg, A.M.P. Importance of dietary fat in the prevention and control of metabolic disorders and cardiovascular disease. Brazilian Archives of Endocrinology & Metabology 2009, 53, 595–607. [Google Scholar]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacological research. 2008, 57, 57,451–455. [Google Scholar] [CrossRef]
- Thanan, R.; Oikawa, S.; Hiraku, Y.; Ohnishi, S.; Ma, N.; Pinlaor, S.; Murata, M. Oxidative stress and its significant roles in neurodegenerative diseases and cancer. International journal of molecular sciences. 2014, 16, 193–217. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Bitto, A. Oxidative stress: harms and benefits for human health. Oxidative medicine and cellular longevity. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Engström, K.; Saldeen, A.S.; Yang, B.; Mehta, J.L.; Saldeen, T. Effect of fish oils containing different amounts of EPA, D.H.A, and antioxidants on plasma and brain fatty acids and brain nitric oxide synthase activity in rats. Upsala journal of medical sciences 2009, 114, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.M.D.S.; Noratto, G.; Martino, H.S.D.; Arbizu, S.; Peluzio, M.D.C.G.; Talcott, S.; Mertens-Talcott, S.U. Pro-apoptotic activities of polyphenolics from açai (Euterpe oleracea Martius) in human SW-480 colon cancer cells. Nutrition and cancer. 2014, 66, 1394–1405. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Zhang, J.G.; Tian, K.W.; Pan, W.J.; Wei, Z.J. The fatty oil from okra seed: supercritical carbon dioxide extraction, composition and antioxidant activity. Current Topics in Nutraceuticals Research. 2014, 2, 75. [Google Scholar]
- Chin, K.Y.; Pang, K.L.; Soelaiman, I.N. Tocotrienol and its role in chronic diseases. Anti-inflammatory nutraceuticals and chronic diseases 2016, 97–130. [Google Scholar]
- Zhang, R.; Shen, W.; Wei, X.; Zhang, F.; Shen, C.; Wu, B.; Deng, X. Simultaneous determination of tocopherols and tocotrienols in vegetable oils by GC-MS. Analytical Methods. 2016, 8, 7341–7346. [Google Scholar] [CrossRef]
- Sabitha, V.; Ramachandran, S.; Naveen, K. R; Panneerselvam, K. Antidiabetic and antihyperlipidemic potential of Abelmoschus esculentus (L.) Moench. in streptozotocin-induced diabetic rats. Journal of pharmacy and bioallied sciences 2011, 3, 397. [Google Scholar]
- Polonini, H.C.; Santos, F.C.D.; Vaz, U.P.; Brandão, M.A.F.; Raposo, N.R.B.; Ferreira, A.D.O. Development and validation of analytical method for determination of simvastatin in capsules. Química Nova 2011, 34, 516–519. [Google Scholar] [CrossRef]
- Veillard, N.R.; Mach, F. Statins: the new aspirin? Cellular and Molecular Life Sciences CMLS. 2002, 59, 1771–1786. [Google Scholar] [CrossRef]
- Bonfim, M.R.; Oliveira, A.S.B.; Amaral, S.L.D.; Monteiro, H.L. Treatment of dyslipidemias with statins and physical exercise: recent evidence of muscle responses. Brazilian Archives of Cardiology. 2015, 104, 324–331. [Google Scholar]
- Barbosa, D.S.; Nunes, L.B.; Godeny, P.; Lozovoy, M.A.B.; Dichi, I. Effects and mechanisms of action of n-3 polyunsaturated fatty acids in the prevention of cardiovascular diseases. UNIPAR Health Science Archives. 2007, 1, 3. [Google Scholar]
- De Lima Viana, D.E.; Dantas, M.M.; da Silva Menezes, M. E. Omega-3 and omega-6 series fatty acids and their use in the treatment of cardiovascular diseases: a review. Revista Saúde & Ciência Online 2016, 5, 65–83. [Google Scholar]
- Frederiksen, K.S.; Wulff, E.M.; Sauerberg, P.; Mogensen, J.P.; Jeppesen, L.; Fleckner, J. Prediction of PPAR-α ligand-mediated physiological changes using gene expression profiles. Journal of lipid research. 2004, 3, 592–601. [Google Scholar] [CrossRef]
- Jung, C.H.; Choi, K.M. Impact of high-carbohydrate diet on metabolic parameters in patients with type 2 diabetes. Nutrients. 2017, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Peters, J.M.; Harris, R.A. Adaptive increase in pyruvate dehydrogenase kinase 4 during starvation is mediated by peroxisome proliferator-activated receptor α. Biochemical and biophysical research communications. 2001, 287, 91–396. [Google Scholar] [CrossRef]
- Guerre-Millo, M., Gervois, P., Raspé, E., Madsen, L., Poulain, P., Derudas, B., .. & Staels, B. Peroxisome proliferator-activated receptor α activators improve insulin sensitivity and reduce adiposity. Journal of Biological Chemistry 2000, 275, 16638-16642.
- Kramer, D.; Shapiro, R.; Adler, A.; Bush, E.; Rondinone, C.M. Insulin-sensitizing effect of rosiglitazone (BRL-49653) by regulation of glucose transporters in muscle and fat of Zucker rats. Metabolism-Clinical and Experimental 2001, 50, 1294–1300. [Google Scholar] [CrossRef]
- Blasi, C. The autoimmune origin of atherosclerosis. Atherosclerosis. 2008, 201, 17–32. [Google Scholar] [CrossRef]
- Rodrigues, A.P.S.; Souza, B.S.F.; Barros, A.S.A.; de Oliveira Carvalho, H.; Duarte, J.; Boettger, L.E.M.; Carvalho, J.C.T. The effects of Rosmarinus officinalis L. essential oil and its nanoemulsion on dyslipidemic Wistar rats. J Appl Biomed. 2018, 18, 126–135. [Google Scholar] [CrossRef]
- Jackson, C.L.; Bennett, M.R.; Biessen, E.A.; Johnson, J.L.; Krams, R. Assessment of unstable atherosclerosis in mice. Arteriosclerosis, thrombosis, and vascular biology 2007, 27, 714–720. [Google Scholar] [CrossRef]
- Mansour, M. The roles of peroxisome proliferator-activated receptors in the metabolic syndrome. Progress in molecular biology and translational science. 2014, 121, 217–266. [Google Scholar]
- Katznelson, L.; Laws Jr, E.R.; Melmed, S.; Molitch, M.E.; Murad, M.H.; Utz, A.; Wass, J.A. Acromegaly: an endocrine society clinical practice guideline. The Journal of Clinical Endocrinology & Metabolism 2014, 99, 3933–3951. [Google Scholar]
- De Moura, J.M.; Gonçalves, L.A.; Petrus, J.C.C.; Viotto, L.A. Degomagem de óleo vegetal por membrana microporosa. Revista de Engenharia de Alimentos. 2005, 70, 473–478. [Google Scholar]
- Schmitt, H. Phospholipids—making lives healthier. Wellness Foods Europe. 2009, 36–41. [Google Scholar]
- Escobar, B. Tableta masticable de lecitina de soya: de subproducto a producto farmacéutico Chewable pill of soy phosphatidylcholine: from by-product to pharmaceutical product Ing. Martha Zoe Lemus Rodríguez, Tec. Amaury Chong Quesada y Tec. Jorge. MEDISAN 2017, 21, 556–563. [Google Scholar]
- Vissotto, F.Z.; Montenegro, F.M.; Santos, J.M.D.; Oliveira, S.J.R.D. Avaliação da influência dos processos de lecitinação e de aglomeração nas propriedades físicas de achocolatado em pó. Food Science and Technology. 2006, 26, 666–671. [Google Scholar] [CrossRef]
- De Luis, D.A.; Aller, R.; Culebras, J. Influence of soy consumption on bone mass. In: Anales de Medicina Interna (Madrid, Spain: 1984). 2007. p. 361-364.
- Cala Calviño, L.; Sánchez Hechavarria, M.E.; García Torres, D.S. Pharmacological aspects of soy lecithin and its possible medical applications. MediSan 2017, 21, 83–95. [Google Scholar]
- Staels, B.; Koenig, W.; Habib, A.; Merval, R.; Lebret, M.; Torra, I. P; Tedgui, A. Activation of human aortic smooth-muscle cells is inhibited by PPARα but not by PPARγ activators. Nature. 1998, 393, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Pasceri, V.; Chang, J.; Willerson, J.T.; Yeh, E.T. Modulation of C-reactive protein–mediated monocyte chemoattractant protein-1 induction in human endothelial cells by anti-atherosclerosis drugs. Circulation. 2001, 103, 2531–2534. [Google Scholar] [CrossRef]
- Rival, Y.; Benéteau, N.; Taillandier, T.; Pezet, M.; Dupont-Passelaigue, E.; Patoiseau, J. F; Delhon, A. PPARα and PPARδ activators inhibit cytokine-induced nuclear translocation of NF-κB and expression of VCAM-1 in EAhy926 endothelial cells. European journal of pharmacology. 2002, 435, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Chinetti, G.; Gbaguidi, F.G.; Griglio, S.; Mallat, Z.; Antonucci, M.; Poulain, P.; Staels, B. CLA-1/SR-BI is expressed in atherosclerotic lesion macrophages and regulated by activators of peroxisome proliferator-activated receptors. Circulation. 2000, 101, 2411–2417. [Google Scholar] [CrossRef]
- Gbaguidi, F.G.; Chinetti, G.; Milosavljevic, D.; Teissier, E.; Chapman, J.; Olivecrona, G.; Staels, B. Peroxisome proliferator-activated receptor (PPAR) agonists decrease lipoprotein lipase secretion and glycated LDL uptake by human macrophages. FEBS letters. 2002, 512, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; HU, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell. 1994, 79, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Marx, N.; Mackman, N.; Schönbeck, U.; Yilmaz, N.; Hombach, V.; Libby, P.; Plutzky, J. PPARα activators inhibit tissue factor expression and activity in human monocytes. Circulation. 2001, 103, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Neve, B.P.; Corseaux, D.; Chinetti, G.; Zawadzki, C.; Fruchart, J.C.; Duriez, P.; Jude, B. PPARα agonists inhibit tissue factor expression in human monocytes and macrophages. Circulation. 2001, 103, 207–212. [Google Scholar] [CrossRef]
- Torabi, S.; Yeganehjoo, H.; Shen, C.L.; Mo, H. Peroxisome proliferator-activated receptor γ down-regulation mediates the inhibitory effect of d-δ-tocotrienol on the differentiation of murine 3T3-F442a preadipocytes. Nutr. Res. 2016, 36, 1345–1352. [Google Scholar] [CrossRef]
- Sun, W.G.; Song, R.P.; Wang, Y.; Ge, S.; Zhang, Y.H.; Wang, H.X.; Ge, S.; Liu, L. X. γ-Tocotrienol-inhibited cell proliferation of human gastric cancer by regulation of nuclear factor-κB activity. Journal of agricultural and food chemistry. 2018, 67, 441–451. [Google Scholar] [CrossRef]
- Matsubara, T.; Takakura, N.; Urata, M.; Muramatsu, Y.; Tsuboi, M.; Yasuda, K.; Kokabu, S. Geranylgeraniol induces PPARγ expression and enhances the biological effects of a PPARγ agonist in adipocyte lineage cells. In vivo. 2018, 32, 1339–1344. [Google Scholar] [CrossRef]
- Aramwit, P.; Supasyndh, O.; Siritienthong, T.; Bang, N. Mulberry leaf reduces oxidation and C-reactive protein level in patients with mild dyslipidemia. BioMed Research International. 2013, 2013. [Google Scholar] [CrossRef]
- preJin, Y-L; Enzan, H.; Kuroda, N.; Hayashi, Y.; Nakayama, H.; Zhang, Y-H; Toi, M.; Miyazaki, E.; Hiroi, M.; Guo, L-M.; Saibara, T. Tissue remodeling following submassive hemorrhagic necrosis in rat livers induced by an intraperitoneal injection of dimethylnitrosamine. Virch. Arch 2003, 442, 39–47. [Google Scholar]
- Oda, E. The metabolic syndrome as a concept of adipose tissue disease. Hypertension Research. 2008, 31, 1283–1291. [Google Scholar] [CrossRef]
- Silva, G.H.; Escanhoela, C.A.F. Nonalcoholic fatty liver disease: pathogenesis and histological findings with emphasis on mitochondrial changes. Journal of Medical Sciences. 2009, 18, 6–5. [Google Scholar]
- Siddiqui, M.S.; Cheang, K.L.; Luketic, V.A.; Boyett, S.; Idowu, M.O.; Patidar Sanyal, A.J. Nonalcoholic steatohepatitis (nash) is associated with a decline in pancreatic beta cell (β-cell) function. Digestive diseases and sciences. 2015, 60, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Handayani, R.; Sulistyo, J.; Rahayu, R.D. Extraction of coconut oil (Cocos nucifera L.) through fermentation system. Biodiversitas Journal of Biological Diversity. 2009, 10.
- Rosset, I.G.; Tavares, M.C.H.; Assaf, E.M.; Porto, A.L.M. Catalytic ethanolysis of soybean oil with immobilized lipase from Candida antarctica and 1H NMR and GC quantification of the ethyl esters (biodiesel) produced. Applied Catalysis A: General 2011, 392, 136–142. [Google Scholar] [CrossRef]
- Rosset, I.G.; Cavalheiro, M.C.H.; Assaf, E.M.; Porto, A.L. Esterificação enzimática de ácido oleico com álcoois alifáticos para a produção de biodiesel por Candida antarctica lipase. Cartas de catálise 2013, 143, 863–872. [Google Scholar]
- Lagunin, A.; Zakharov, A.; Filimonov, D.; Poroikov, V. QSAR modelling of rat acute toxicity on the basis of PASS prediction. Mol. Inform. 2011, 30, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Filimonov, D.A.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.V.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the biological activity spectra of organic compounds using the pass online web resource. Chem. Heterocycl. Compd. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Druzhilovskiy, D.S.; Rudik, A.V.; Filimonov, D.A.; Gloriozova, T.A.; Lagunin, A.A.; Dmitriev, A.V.; Pogodin, P.V.; Dubovskaya, V.I.; Ivanov, S.M.; Tarasova, O.A.; et al. Computational platform Way2Drug: From the prediction of biological activity to drug repurposing. Russ. Chem. Bull. 2017, 66, 1832–1841. [Google Scholar] [CrossRef]
- Rudik, A.V.; Dmitriev, A.V.; Lagunin, A.A.; Filimonov, D.A.; Poroikov, V.V. PASS-based prediction of metabolites detection in biological systems. SAR QSAR Environ. Res. 2019, 30, 751–758. [Google Scholar] [CrossRef]
- Pereira, A.C.M.; Carvalho, H.D.O.; Gonçalves, D.E.S.; Picanço, K.R.T.; Santos, A.V.T.D.L.T.D.; da Silva, R.H.; Braga, F.S.; Bezerra, R.M.; Nunes, A.D.S.; Nazima, M.T.S.T.; e outros Co-tratamento de óleo de urucum purificado ( Bixa orellana L.) e seus grânulos (Chronic®) melhora o perfil lipídico sanguíneo e os efeitos protetores ósseos da testosterona na osteoporose induzida por orquiectomia em ratos Wistar. Moléculas 2021 , 26 , 4720.
- Dobiasova, M. Atherogenic index of plasma [log (triglycerides/HDL-cholesterol)]: theoretical and practical implications. Clinical Chemistry. 2004, 50, 1113–1115. [Google Scholar] [CrossRef]
- Souza, G.C.; Duarte, J.L.; Fernandes, C.P.; Moyado, J.A.V.; Navarrete, A.; Carvalho, J.C.T. Obtainment and Study of the Toxicity of Perillyl Alcohol Nanoemulsion on Zebrafish (Danio rerio). Journal of Nanomedicine Research. 2016, 4, 18–20. [Google Scholar]
- Rhoden, E.L.; Pereira-Lima, L.; Rhoden, C.R.; Lucas, M.L.; Mauri, M.; Zettler, C.G. Analysis of the histopathologic alterations of the livers in Rats pretreated with allopurinol and submitted to hepatic Ischemia: reperfusion. Journal of the Brazilian College of Cirurgiões 2000, 27, 373–377. [Google Scholar]
Figure 1.
Analysis by gas chromatography coupled to mass spectrometry (GC-MS) of Euterpe oleracea oil.
Figure 1.
Analysis by gas chromatography coupled to mass spectrometry (GC-MS) of Euterpe oleracea oil.
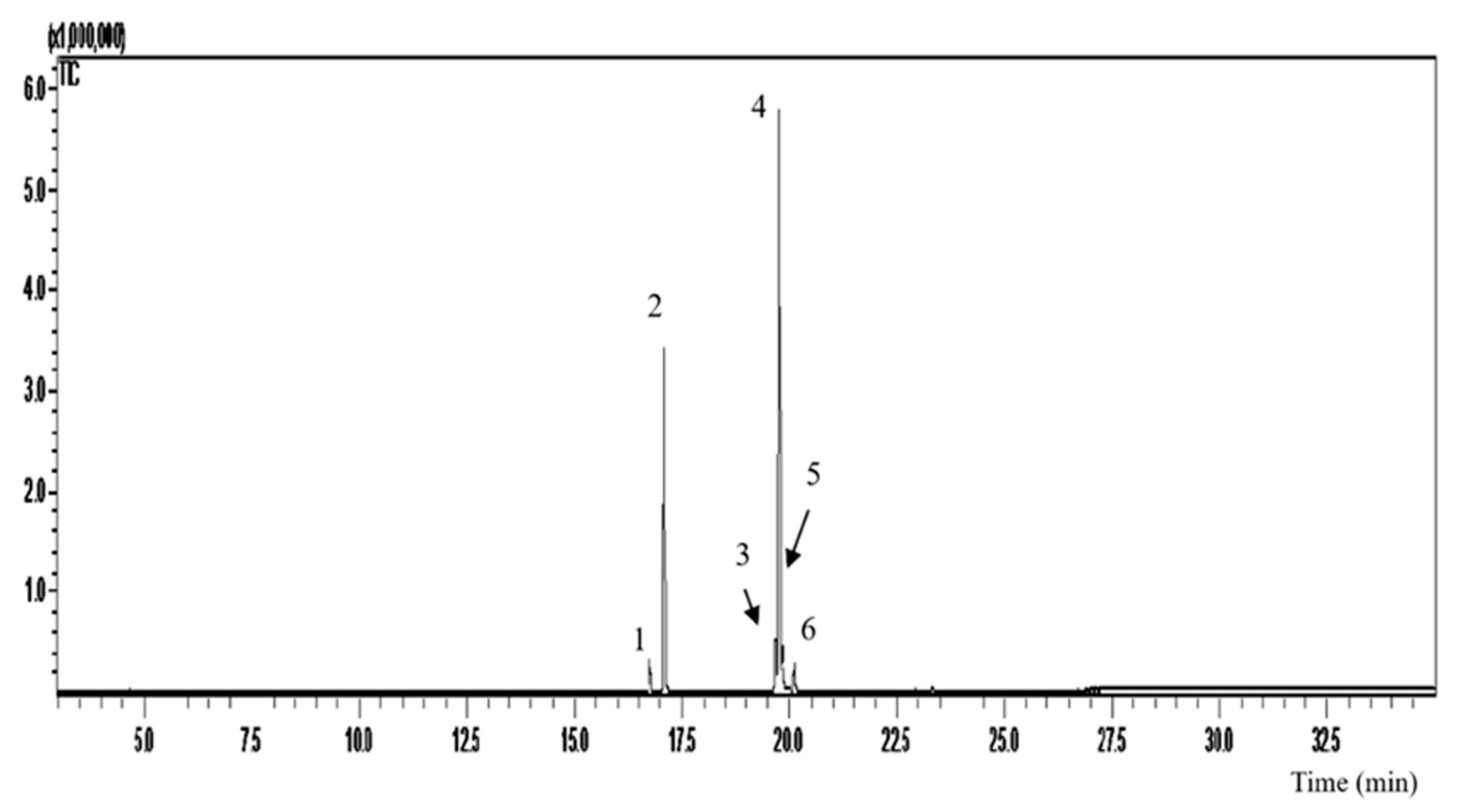
Figure 2.
Analysis by gas chromatography coupled to mass spectrometry (GC-MS) of Abelmoschus esculentus oil.
Figure 2.
Analysis by gas chromatography coupled to mass spectrometry (GC-MS) of Abelmoschus esculentus oil.
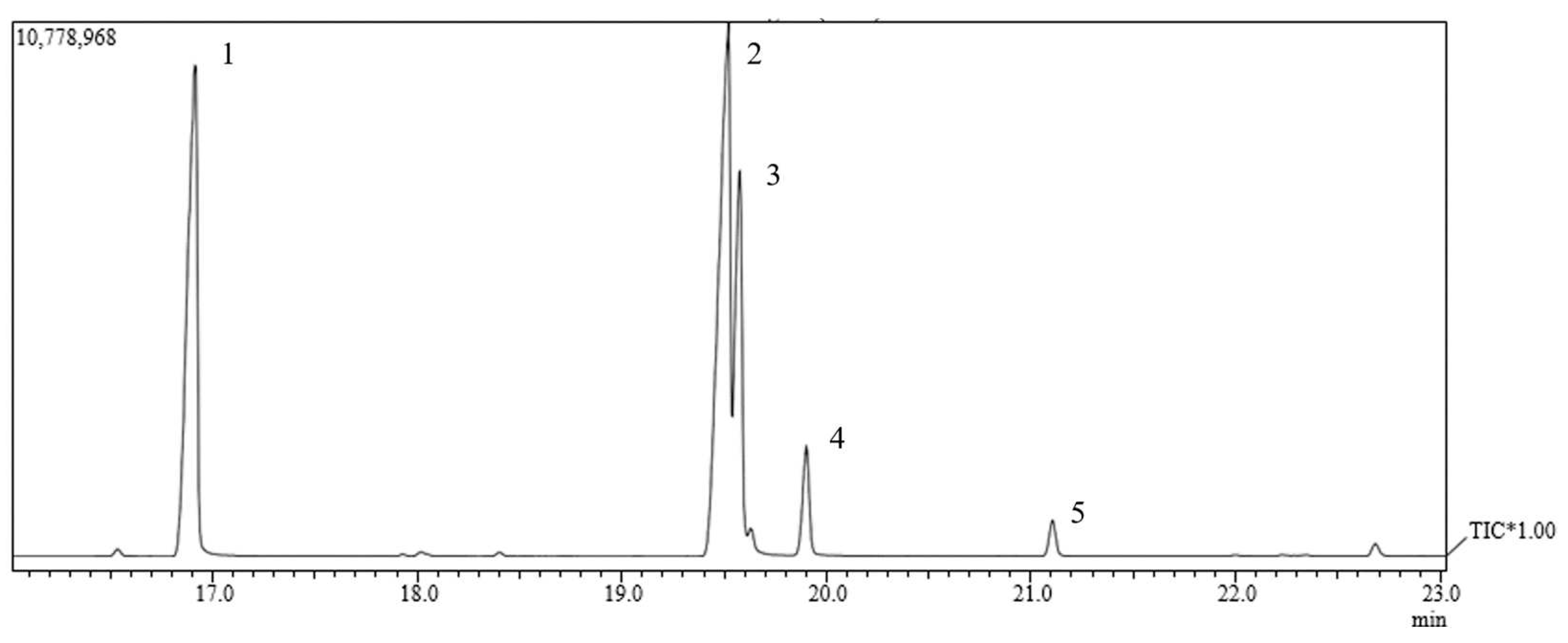
Figure 3.
Calibration curve of total tocotrienols based on delta-tocotrienol.
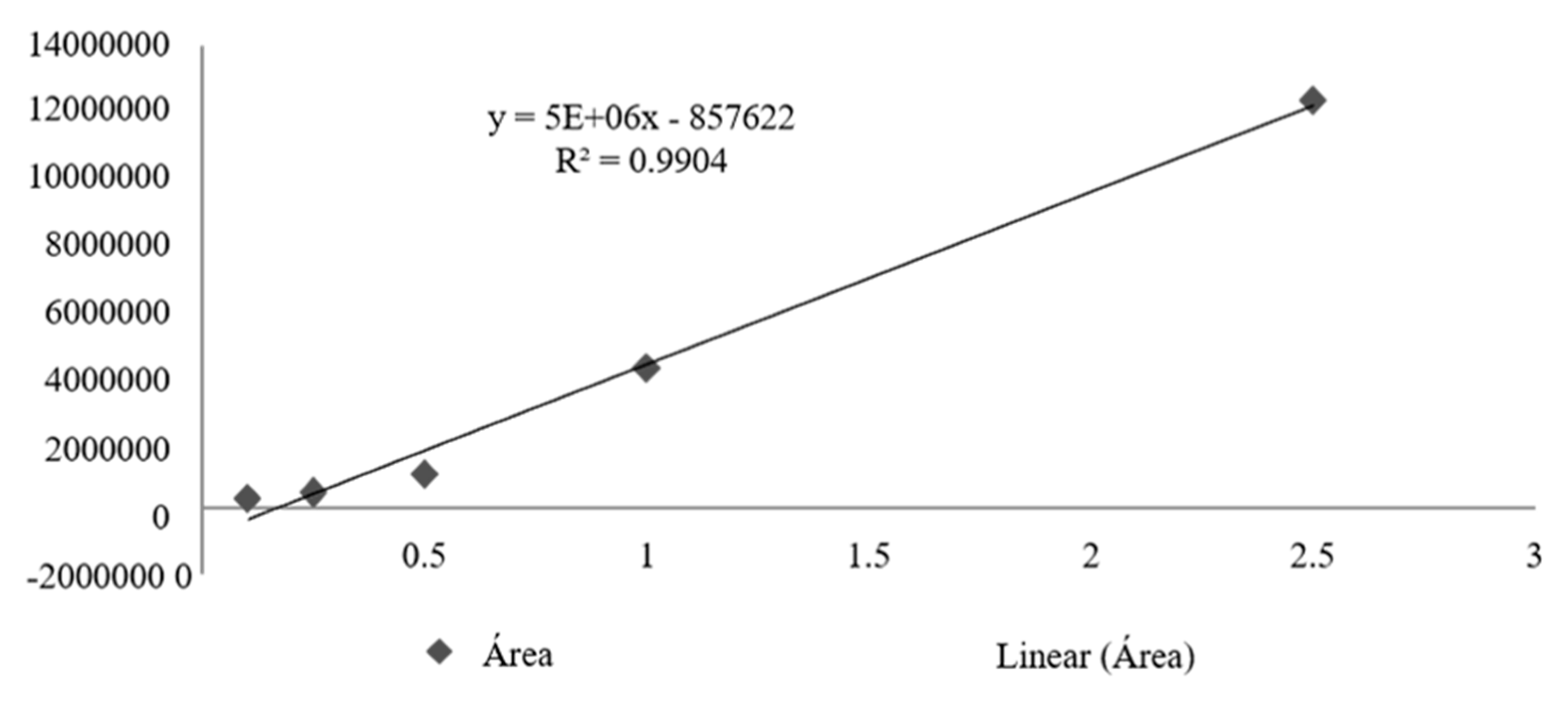
Figure 4.
Calibration curve obtained for geranylgeraniol.
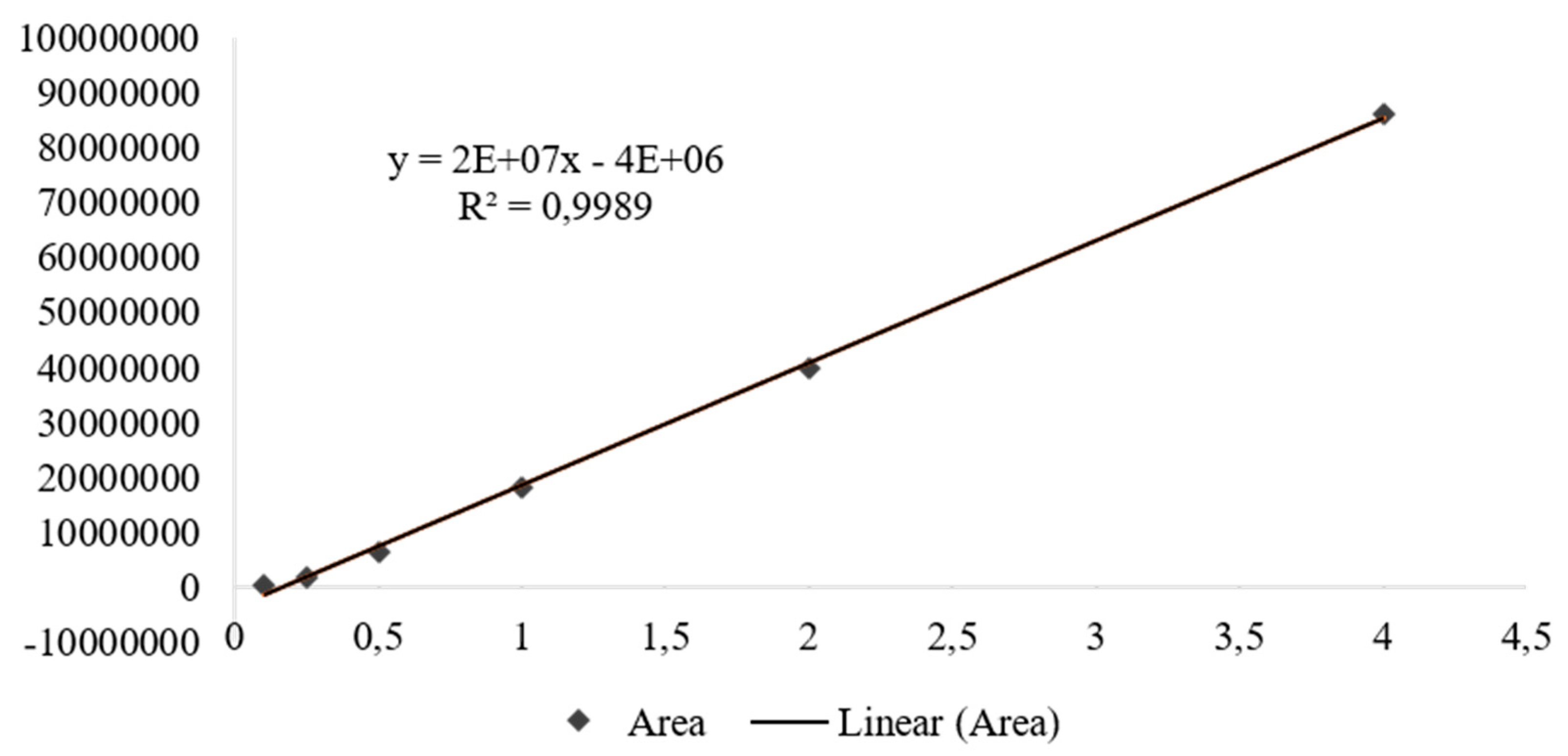
Figure 5.
Effect of CHR, OFEO, OFAE and OFBO treatment on total cholesterol, LDL, HDL and triglycerides in GSC-induced dyslipidemia. Results presented as mean±standard deviation. ***p>0.001 and **p>0.01 represent statistically significant results compared to the GSC group. One-Way ANOVA test followed by the Tukey Multiple Comparisons post-test.
Figure 5.
Effect of CHR, OFEO, OFAE and OFBO treatment on total cholesterol, LDL, HDL and triglycerides in GSC-induced dyslipidemia. Results presented as mean±standard deviation. ***p>0.001 and **p>0.01 represent statistically significant results compared to the GSC group. One-Way ANOVA test followed by the Tukey Multiple Comparisons post-test.
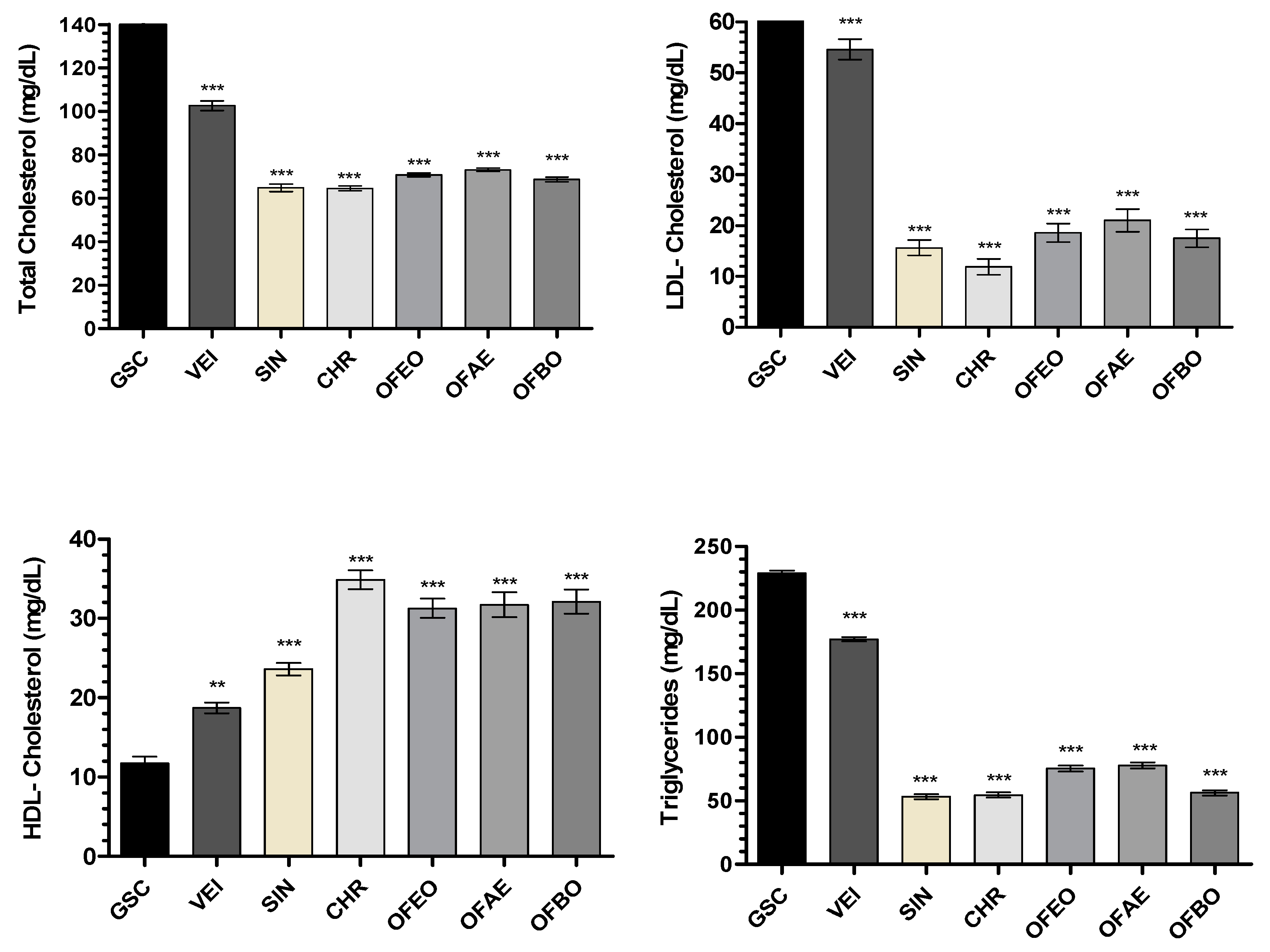
Figure 6.
SEM images of cross-sections of the thoracic aorta (A) GCS, (B) VEI, (C) SIN, (D) CHR, (E) OFEO, (F) OFAE, (G) OFBO. The white arrows indicate the formation of an atheroma plaque in the vascular endothelium.
Figure 6.
SEM images of cross-sections of the thoracic aorta (A) GCS, (B) VEI, (C) SIN, (D) CHR, (E) OFEO, (F) OFAE, (G) OFBO. The white arrows indicate the formation of an atheroma plaque in the vascular endothelium.
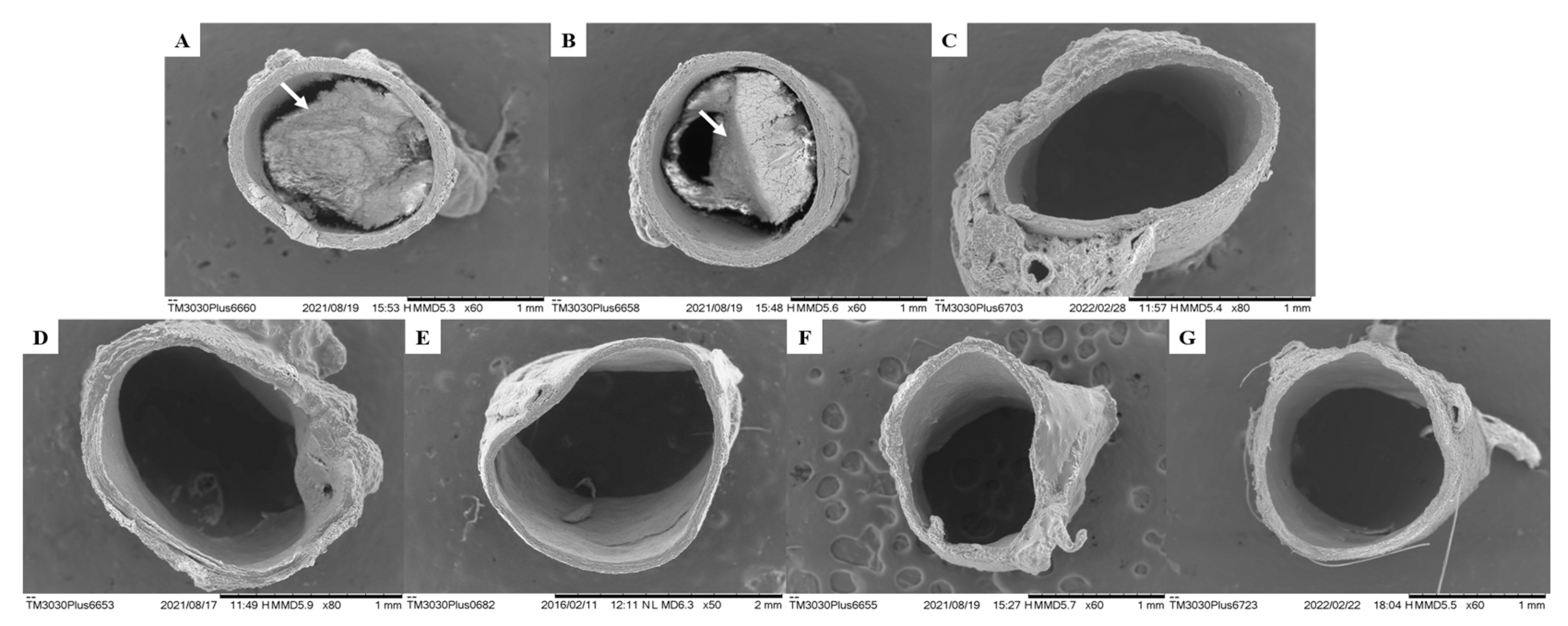
Figure 7.
Histological section of the liver of animals submitted to the coconut oil-induced dyslipidemia test. In A, normal liver, where hepatocytes are observed (H); In B, altered liver, where Hs is observed: hepatic steatosis; In C, altered liver, where Sc is observed: sinusoidal congestion. (H&E).
Figure 7.
Histological section of the liver of animals submitted to the coconut oil-induced dyslipidemia test. In A, normal liver, where hepatocytes are observed (H); In B, altered liver, where Hs is observed: hepatic steatosis; In C, altered liver, where Sc is observed: sinusoidal congestion. (H&E).

Table 1.
Determination of FAEE equivalent to fatty acids (FAEE) in the composition of Euterpe oleracea oil analyzed by GC-MS.
Table 1.
Determination of FAEE equivalent to fatty acids (FAEE) in the composition of Euterpe oleracea oil analyzed by GC-MS.
| aFatty acid (n:i) | Peaks | Retettion time (min) | Concentration (%) |
|---|---|---|---|
| Palmitoleic (16:1) | 1 | 16.75 | 2.62 |
| Palmitic (16:0) | 2 | 18.07 | 30.0 |
| Linoleic (18:3) | 3 | 19.68 | 5.9 |
| (Z)-Oleic (18:1) | 4 | 19.78 | 54.32 |
| Elaidic (18:1) | 5 | 19.84 | 4.29 |
| Stearic (18:0) | 6 | 20.12 | 2.29 |
| ∑Monounsaturated | ** | ** | 61.27 |
| ∑ Polyunsaturated | ** | ** | 5.9 |
| ∑ Unsaturated | ** | ** | 67.83 |
| ∑ Saturated | ** | ** | 32.83 |
1 aFAEE equivalent to the respective fatty acids and (n:i) number of carbon and unsaturation.
Table 2.
Determination of FAEE equivalent to fatty acids (FAEE) in the composition of Abelmoschus esculentus oil analyzed by GC-MS.
Table 2.
Determination of FAEE equivalent to fatty acids (FAEE) in the composition of Abelmoschus esculentus oil analyzed by GC-MS.
| aFatty acid (n:i) | Peaks | Retettion time (min) | Concentration (%) |
|---|---|---|---|
| Palmitic (16:0) | 1 | 19.91 | 33.55 |
| Linoleic (18:2) | 2 | 19.51 | 42.12 |
| Oleic (18:1) | 3 | 19.57 | 17.85 |
| Stearic (18:0) | 4 | 19.90 | 4.96 |
| Eicosenoic (20:1) | 5 | 21.10 | 1.53 |
| ∑Monounsaturated | ** | ** | 19.38 |
| ∑ Polyunsaturated | ** | ** | 42.12 |
| ∑ Unsaturated | ** | ** | 61.5 |
| ∑ Saturated | ** | ** | 40.04 |
2 aFAEE equivalent to the respective fatty acids and (n:i) number of carbon and unsaturation.
Table 3.
Average concentration and average content of total tocotrienols in OFBO and granules (Chronic SM®).
Table 3.
Average concentration and average content of total tocotrienols in OFBO and granules (Chronic SM®).
| Sample | Concentration (mg/mL) | Content (%) |
|---|---|---|
| OFBO | 0,617448233 | 61,74482 |
| CHR | 0,046756 | 46,75485 |
Table 4.
Average concentration and average content of geranylgeraniol of OFBO and granules (Chronic SM®).
Table 4.
Average concentration and average content of geranylgeraniol of OFBO and granules (Chronic SM®).
| Sample | Concentration (mg/mL) | Content (%) |
|---|---|---|
| OFBO | 0,872136 | 87,2136 |
| CHR | 0,1416946 | 14,1694 |
Table 5.
Effect of CHR, OFEO, OFAE and OFBO treatment on clinical parameters of Wistar rats with GSC-induced dyslipidemia.
Table 5.
Effect of CHR, OFEO, OFAE and OFBO treatment on clinical parameters of Wistar rats with GSC-induced dyslipidemia.
| Parameters | Water(ml) | Weight (g) | Food (g) |
| GSC | 269,43±13,26 | 375,02±7,25 | 131,57±6,66 |
| VEI | 252,36±10,89 | 280,73±7,20*** | 84,50±1,55 |
| SIN | 251,60±8,77* | 269,68±4,03*** | 83,40±1,92 |
| CHR | 245,21±7,03** | 259,57±4,00*** | 71,33±2,86*** |
| OFEO | 237,33±8,25*** | 265,41±5,11*** | 83,67±1,97 |
| OFAE | 238,90±8,10*** | 270,39±4,62*** | 96,14±4,22 |
| OFBO | 249,19±5,93* | 243,52±4,25*** | 76,60±1,14*** |
5Results presented as mean±standard deviation. ***p>0.001, **p>0.01 and *p>0.05 represent statistically significant results compared to the GSC group. One-Way ANOVA test followed by the Tukey Multiple Comparisons post-test.
Table 6.
Effect of treatment with CHR, OFEO, OFAE and OFBO on biochemical parameters of Wistar rats with GSC-induced dyslipidemia.
Table 6.
Effect of treatment with CHR, OFEO, OFAE and OFBO on biochemical parameters of Wistar rats with GSC-induced dyslipidemia.
| Parameters | Urea (mg/dL) | Creatinine (mg/dL) | AST (U/dL) | ALT (U/dL) |
| GSC | 38,29±5,96 | 0,90±0,04 | 141,08±7,27 | 59,11±7,37 |
| VEI | 33,29±2,29 | 0,82±0,05* | 136,50±8,64 | 51,97±4,20 |
| SIN | 27,75±5,26** | 0,80±0,04*** | 128,70±8,58* | 46,11±5,65*** |
| CHR | 28,63±5,68* | 0,85±0,03** | 131,60±5,12 | 48,64±3,00** |
| OFEO | 32,00±4,16 | 0,81±0,04** | 132,55±4,25 | 51,44±3,56 |
| OFAE | 30,71±5,19 | 0,83±0,03* | 133,15±6,67 | 53,34±2,59 |
| OFBO | 29,13±3,31* | 0,81±0,05*** | 132,50±5,48 | 50,49±4,26* |
6Results presented as mean±standard deviation. ***p>0.001, **p>0.01 and *p>0.05 represent statistically significant results compared to the GSC group. One-Way ANOVA test followed by the Tukey Multiple Comparisons post-test.
Table 7.
Effect of treatment with CHR, OFEO, OFAE and OFBO on biochemical parameters and Atherogenic Index in the plasma of Wistar rats with dyslipidemia induced by GSC.
Table 7.
Effect of treatment with CHR, OFEO, OFAE and OFBO on biochemical parameters and Atherogenic Index in the plasma of Wistar rats with dyslipidemia induced by GSC.
| Parameters | Glucose (mg/dL) | PCR (mg/dL) | Atherogenic Index of Plasma (AIP) |
|---|---|---|---|
| GSC | 161,29±9,23 | 0,84±0,06 | 1,30±0,07 |
| VEI | 143,00±6,81*** | 0,62±0,04*** | 0,98±0,04*** |
| SIN | 129,25±5,15*** | 0,39±0,04*** | 0,23±0,07*** |
| CHR | 114,88±2,95*** | 0,41±0,04*** | 0,19±0,07*** |
| OFEO | 126,00±3,37*** | 0,49±0,03*** | 0,38±0,03*** |
| OFAE | 122,86±2,04*** | 0,50±0,03*** | 0,39±0,03*** |
| OFBO | 117,13±4,05*** | 0,47±0,04*** | 0,25±0,08*** |
7Results presented as mean±standard deviation. ***p>0.001 represent statistically significant results compared to the GSC group. One-Way ANOVA test followed by the Tukey Multiple Comparisons post-test.
Table 8.
Histopathological study of the liver of animals submitted to the Cocos nucifera L. (GSC) induced dyslipidemia test.
Table 8.
Histopathological study of the liver of animals submitted to the Cocos nucifera L. (GSC) induced dyslipidemia test.
| Group/Change | GSC | VEI | SIN | CHR | OFEO | OFAE | OFBO |
|---|---|---|---|---|---|---|---|
| Sinusoidal congestion | 2a | 0 | 1b | 1 | 0 | 0 | 0 |
| Steatosis | 2c | 1 | 2d | 0 | 1 | 1 | 1 |
| Necrosis | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
8Different letters represent statistically significant difference p<0.05 between groups (Kruskal-Wallis test, followed by Dunn's method).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



