
Submitted:
19 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
LSM4 is an essential yeast gene encoding a component of different LSM complexes involved in the regulation of mRNA splicing, stability and translation. In previous papers we reported that the expression in S. cerevisiae of the K. lactis Lsm4 gene lacking the C-terminal Q/N-rich domain in a Lsm4 null strain S. cerevisiae (Sclsm4D1) restored cell viability. Nevertheless, in this transformed strain we observed some phenotypes which are typical markers of regulated cell death, Reactive Oxygen Species and oxidated RNA accumulation. In this paper we report that a similar truncation operated in the S. cerevisiae LSM4 gene confers to cells the same phenotypes observed with the K.lactis LSM4D1 gene. Up to now there was no evidence on the direct involvement of LSM4 in autophagy. Here we found that the Sclsm4D1 mutant showed defects in the induction of autophagy and it was very sensitive to nitrogen starvation or treatment with low doses of rapamycin, an inducer of autophagy. Moreover, both during nitrogen starvation and ageing, the Sclsm4D1 mutant accumulated cytoplasmic autophagy-related structures, suggesting a role of Lsm4 in Phagophore Assembly Site (PAS) processing and/or vacuolar autophagosome internalization
Keywords:
Autophagy
; Ageing
; LSM4
; Yeast
1. Introduction
The LSM (like-Sm) protein family is a group of evolutionarily conserved proteins that have been found in a wide range of organisms, from bacteria to yeast and humans [1]. LSM4 is an essential gene and Lsm4 is a component of different LSM complexes, which are involved in the regulation of mRNA splicing, stability and translation [2].
LSM4 has also been found to be upregulated in several types of cancer, such as breast cancer, and in early-stage pancreatic ductal adenocarcinoma, where it is associated with poor prognosis [3,4,5].
We previously reported that Lsm4 is involved in ageing and apoptosis in the yeast Saccharomyces cerevisiae, in that cell expressing a truncated protein of the KlLsm4, from the related yeast Kluyveromyces lactis (Kllsm4Δ1) age prematurely and undergo regulated cell death [6,7]. We demonstrated that the C-terminal Q/N-rich domain of KlLSM4 is needed for efficient RNA degradation [7,8] and for P-bodies localization [9,10], while others reported that this region of ScLSM4 is important only together with the Edc3 protein [11]. These differences could be explained by the use of slightly different constructions and the use of yeast strains that can have different genetic backgrounds.
Although the C-terminal domain of Lsm4 in most eukaryotes consists of an arginine–glycine–glycine repeat (RGG) domain rather than a Q/N-rich region and, despite a great degree of variation in the primary sequence, some functions seem conserved through evolution, while others were not. It has been reported that low-complexity polypeptide regions of proteins, including R/G-rich regions, were involved in protein polymerization and aggregation, suggesting a role for Lsm4 C-terminus in these processes [12].
Actually, as observed in yeast, the C-terminal RGG domain of human Lsm4 plays an important role in Processing bodies (P-bodies, PB) accumulation but, differently from yeast, it is not required for the association to Lsm1-7 complex [13].
Similarly to yeast, this region is important for efficient histone mRNA degradation [14,15], but it has also been shown that the Lsm4 RGG domain is not limiting for the degradation of histone H2A mRNA [13]. These contradictory results should be further investigated.
Both genetic and mRNA capture analysis revealed an involvement of Lsm proteins in the autophagic process [16,17,18] and in last years it became clear that mRNA degradation pathways and autophagy are intimately connected [19].
In mutant cells lacking the RNA helicase Dhh1 or the decapping protein Dcp2, autophagy in the presence of nutrients was increased. In fact, when nutrients are present, TORC1 is able to phosphorylate a serine residue on the Dcp2 that, together with the RCK-Dhh1 complex binds messenger RNAs of the ATG genes promoting the removal of the cap and their degradation by the exoribonuclease Xrn1 [20]. When nutrients are scarce or absent, the catalytic activity of TORC1 is reduced and, consequently, the phosphorylation levels of Dcp2p is reduced, leading to decreased decapping activity and stabilization of ATG genes transcripts. Nevertheless, the DHH1 and DCP2 gene were shown to have no influence on the mRNA degradation levels of ATG genes in the absence of nutrients, suggesting that the decapping machinery promotes the degradation of ATG transcripts only in the presence of nutrients, when the autophagy is not necessary [20].
In a more recent study, it was reported that cells lacking the DHH1 gene (dhh1Δ) rapidly lose viability after prolonged nitrogen starvation, indicating that under these conditions there would be a defect in autophagy induction [21].
Dhh1 would therefore be a bidirectional regulator of autophagy: a) under nutrient-rich conditions, Dhh1 coordinates with the mRNA decapping machinery to degrade ATG mRNAs to maintain autophagy at a basal level, b) upon nitrogen starvation, Dhh1 changes its role to facilitate autophagy by promoting the translation of ATG1 and ATG13 mRNAs [21].
A similar bidirectional role in autophagy seems to be played by the CCR4/NOT complex, which under nutrient-rich conditions directly targets some ATG genes mRNAs promoting their degradation through deadenylation while upon starvation, Ccr4-Not switches its role to promote the expression of a different subset of ATG genes required for autophagy induction [22].
Along with the decapping process followed by degradation in the 5’→3’ direction, mRNAs can also be degraded in the 3’→5’ direction by the exosome complex. Although both 5’→3’ and 3’→5’ RNA degradation mechanisms have been extensively studied, little is known about the relationship between the two pathways. The RNA-binding protein Pat1/Mrt1 has been proposed as a possible link between 5’→3’ and 3’→5’ mRNA degradation. Pat1 interacts with the ring-shaped heptameric complex Lsm1-Lsm7 proteins, to form the Pat1-Lsm complex that binds to the 3’ untranslated region (UTR) of oligoadenylated mRNA [23,24] and acts as a decapping enhancer, in protecting the 3’ end of oligoadenylated mRNA from trimming, and in protecting the 3’ end of mRNA from exosome-dependent 3’-5’ degradation [25,26,27,28].
During nitrogen starvation, the Pat1/Lsm complex binds preferentially to an mRNA subset of the ATG genes, protecting them from degradation by the exosome and promoting their accumulation to ensure a robust autophagic response [28]. Indeed, mutants in the LSM1 or PAT1 genes show evident defects in the induction of the autophagic process induced by nitrogen starvation.
The role of Pat1/Lsm complex in protecting from exosome degradation is specific for some ATGs mRNA, in that bulk and specific degradation of subset of mRNAs, notably those encoding amino acid biosynthesis and ribosomal proteins, occurs by nitrogen starvation or rapamycin-induced autophagy in yeast [29,30].
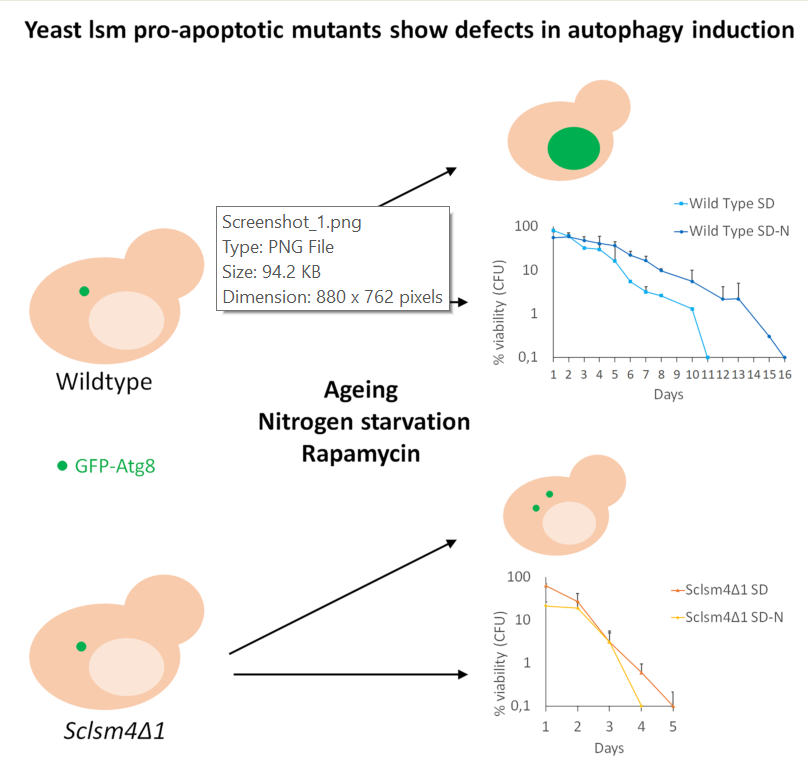
As reported before, the Lsm4 C-terminal Q/N-rich domain is involved in maintaining cell viability during chronological Life Span (CLS). In its absence, we observed all typical markers of regulated cell death, together with Reactive Oxygen Species (ROS) and oxidated RNA accumulation [31]. As these phenotypes were observed expressing the heterologous truncated Kllsm4 gene of K. lactis in the deletion mutant of S. cerevisiae, we decided to construct the corresponding truncated mutant of the LSM4 gene of S. cerevisiae. The expression of truncated S. cerevisiae protein in the absence of LSM4, recapitulated all the phenotypes observed in Kllsm4Δ1 mutant [6] suggesting a role for LSM4 C-terminus in maintaining viability during CLS. We found that the Sclsm4Δ1 mutant showed defects in the induction of autophagy and it was very sensitive to nitrogen starvation or treatment with low doses of rapamycin.
Moreover, both during nitrogen starvation and ageing, the Sclsm4Δ1 mutant accumulated cytoplasmic autophagy-related structures, suggesting a role for Lsm4 in vacuolar Phagophore Assembly Site (PAS), internalization.
2. Results
2.1. Sclsm4Δ1 mutant shows regulated cell death markers and premature ageing
We previously reported that a truncated form of the KlLSM4 gene from the yeast K. lactis (Kllsm4Δ1) was able to restore viability in a S. cerevisiae strain not expressing LSM4. Nevertheless, cells lost viability very soon and showed the markers of regulated cell death [6]. In order to investigate this phenotype in a homologous scenario, we constructed the corresponding Kllsm4Δ1 mutant of the S. cerevisiae LSM4 gene (Sclsm4Δ1), and we expressed it in the S. cerevisae MCY4 strain, which contained the LSM4 gene under the control of Gal1-10 promoter. This strain can grow in galactose, but it cannot grow when glucose is the only carbon source. The expression in such a strain of the Sclsm4Δ1 gene from a centromeric plasmid restored growth on glucose, as also reported when present Kllsm4Δ1. Aimed to check if the phenotypes of the Sclsm4Δ1 mutant were similar to those described for Kllsm4Δ1, we analysed nuclei morphology, intracellular ROS production and maintenance of viability during stationary phase, also defined as Chronological Life Span (CLS). As shown in Figure 1, panel A and B, highly fragmented, enlarged and diffused nuclei, indicative of regulated cell death, were observed in more than 15% of exponentially growing cells and in almost 40% of cells during stationary phase. These percentages are much higher compared to the wild type, which show about 3% and 12% of cells with fragmented nuclei in exponential and stationary phase cells, respectively.
Concerning intracellular ROS accumulation, the percentage of ROS positive cells during exponential phase was about 2% and 12% for the WT and the Sclsm4Δ1 mutant cells. In the latter, the percentage of ROS positive cells during stationary phase reached 60%, about six time more than the WT (Figure1, panels C and D).
One particular phenotype of Kllsm4Δ1 was the early loss of viability during ageing and high sensitivity to oxidative stress. As shown in Figure 1E, also Sclsm4Δ1 cells show a very short CLS, and high sensitivity to hydrogen peroxide treatments (Figure 1, panel E and F).
As a control, to verify that these phenotypes were not due to the expression of Sclsm4Δ1 gene from a centromeric plasmid, we also expressed in the MCY4 strain the full legth gene ScLSM4. As shown in Figure 1E and 1F, the expression of the ScLSM4 gene restored both CLS and hydrogen oxide sensitivity at the same level of the WT strain.
Finally, as also reported for Kllsm4Δ1, Sclsm4Δ1 showed sensitivity to caffeine and acetic acid and lower growth on glycerol medium (Figure 1, panel G). Altogether, these results show that Sclsm4Δ1 mutant recapitulates all the phenotypes showed by Kllsm4Δ1, regard to regulated cell death and premature ageing [32].
2.2. Sclsm4Δ1 mutant is defective in autophagy induction
We previously reported that the over-expression of NEM1, which codes for the catalytic subunit of the yeast nuclear membrane-resident protein phosphatase complex Nem1/Spo7, can suppress most of the mutant phenotypes in the S. cerevisiae lsm4 mutant expressing Kllsm4Δ1 [33]. It has been reported that Nem1 is required for autophagy induction after TORC1 inactivation [34], so we assessed if Lsm4 was involved in the macroautophagy flux through a GFP-Atg8 processing assay [35]. As GFP β-barrel structure is more resistant than Atg8p to vacuolar hydrolysis, the presence of free GFP on western blot indicates that the autophagic process has occurred. Autophagy can be induced under a variety of deprivation conditions, such as depletion of nitrogen and during post-diauxic shift [36]. The autophagic flux has been evaluated upon nitrogen deprivation and during the post-diauxic growth phase, both in wild type and Sclsm4Δ1 mutant cells.
As shown in Figure 2A, during exponential growth autophagy was not observed in the wild type nor in the mutant (Exp, lanes 1 and 4) as only the GFP-Atg8 fusion protein was detected. Protein extracts obtained from cells in the post-diauxic phase (PD, lanes 2 and 5) and nitrogen starvation (SD-N, lanes 3 and 6) showed that free GFP production was reduced in Sclsm4Δ1 cells, suggesting a defect in this mutant in inducing macroautophagy.
Macroautophagy is important for survival during nutrient starvation, and mutants defective in autophagy rapidly lose cell viability after nitrogen starvation [37]. In fact, defective autophagy cells fail to maintain physiological levels of amino acids, and their inability to synthesize new proteins may explain, at least in part, most of the phenotypes associated with autophagy mutants [38]. As from the GFP-Atg8 assay the Sclsm4Δ1 mutant seemed to have important defects in inducing autophagy, we determined the CLS in nitrogen starvation conditions. As shown in Figure 2C, nitrogen starvation increased CLS in a wild type strain. On the contrary, Sclsm4Δ1 cells in SD-N showed a drop in viability already after 1 day and, completely lost viability at day 4, one day before cells maintained in SD.
Another known inducer of autophagy is the antibiotic rapamycin [35]. We found that the Sclsm4Δ1 mutant was highly sensitive even to low doses of rapamycin in that exponential growing cells exposure to 6 nM rapamycin reduced cell viability to 4% within 4 hours, while this is not the case for the wild-type strain, which maintained cell viability equal to untreated cells (Figure 3A).
Similar sensitivity to this drug was found in the lsm1Δ mutant, which is a component of the heptameric ring-shaped complex formed by Lsm1 to Lsm7 [39]. As reported in Figure 3B, the serial dilution assay showed the high sensitivity to 6nM rapamycin of both Sclsm4Δ1 and lsm1Δ cells, compared to their respective wild types CML39-11A and BMA38, suggesting that in both lsm mutant strains autophagy is impaired.
Oxidative stress can induce autophagy both in yeast and in mammalian cells [40] ; at the same time, it has been reported that rapamycin-induced autophagy, confers neuroprotection against aging-induced oxidative stress in old rats [41]. We explored if low doses of rapamycin could protect cells from hydrogen peroxide induced cell death. As shown in Figure 4, the presence of 6nM rapamycin did not protect wild type cells from oxidative stress, as the differences in viability of the treated and untreated samples after exposure to different concentration of H2O2 were not statistically significant. On the other hand, it was not possible to evaluate the protective action of rapamycin in the Sclsm4Δ1 mutant due to its high toxic effect.
During autophagy, bulk cytoplasmic material is sequestered by the phagophore, a double-membrane structure which expands around the cargo forming a sealed, double-membrane vesicle known as the autophagosome (AP). The autophagosome fusion to the vacuole leads to degradation and recycling of the cargo. Autophagic flux can be monitored by the localization of GFP-Atg8, which is delivered to the vacuoles to be degraded. With the aim to have more information on the autophagic step blocked in the Sclsm4Δ1 mutant, we followed the localization of the fusion protein GFP-Atg8 by fluorescence microscopy.
During exponential phase of growth in SD around 1% and 7% of the WT and Sclsm41 mutant cells, respectively, had a single GFP-Atg8 dot denoting the PAS localized near the vacuole membrane (Figure5A, SD exp), being most of the fluorescence uniformly distributed into the cytoplasm. During this growth phase, in the Sclsm4Δ1 mutant there is observed a small percentage of cells showing two or more GFP-Atg8 dots per cell (Figure 5B).
After 4 hours of nitrogen starvation (SD-N) the differences between the wildtype and the mutant increased, with a mean percentage of GFP-Atg8 dots around 3% for the wildtype and 25% for the Sclsm4Δ1 mutant (Figure 5A, SD-N 4h). Moreover, also the number of cells showing ≥2 GFP-Atg8 dots increased in the mutant cells to about 12%, representing half of cell population with GFP-Atg8 dots (Figure 5B).
In the post-diauxic phase there was a little increase in autophagy (Figure 2A) and, as expected, most of the GFP was localized inside the vacuole in the wild type (Figure 5C, PD), with about 3% of cells showing GFP-Atg8 dots. Concerning Sclsm4Δ1 mutant, the percentage of cells showing GFP-Atg8 dots increased up to about 20%, with very few cells showing intravacuolar fluorescence.
After 16h in nitrogen starvation (Figure 5C, SD-N 16h) it was observed a slight increase of cells showing dots in the wild type, while about 35% of Sclsm4Δ1 mutant cells showed cytoplasmic dots and half of them presented 2 or more dots per cell (Figure 5D).
After three days of growth in SD more than 90% of Sclsm4Δ1 mutant cells showed GFP-Atg8 dots, being the number of the wild type cells presenting dots about 13% of the population (Figure 5E, quantification in 5F). This percentages increases a little bit in the wild type incubated for 3 days in SD-N medium, while in the Sclsm4Δ1 mutant cells those presenting GFP-Atg8 dots after 3 days in SD-N medium were the same as after 16h of incubation in SD-N (about 40%, Figure 5F). This could be due to the rapid loss of viability of the Sclsm4Δ1 mutant in SD-N observed already at day 1 (Figure 2C). These data altogether, indicated that Sclsm4Δ1 mutant cells accumulated autophagy-related structures when autophagy was induced by nitrogen starvation or during ageing.
The same experiments were carried out with the lsm1Δ1 mutant with similar results (Supplementary Figure 1), suggesting that the observed autophagy defects are a feature of lsm mutants.
3. Discussion
The Sclsm4Δ1 mutant of S. cerevisiae, which expresses a truncated form of the essential gene LSM4, showed premature ageing, fragmented nuclei and ROS accumulation. The Sclsm4Δ1 mutant showed also high sensitivity to the regulated cell death-inducers acetic acid and caffeine, the same phenotypes as those previously shown for the Kllsm4Δ1 mutant of K. lactis [6,7,42]. It has been reported that mutants hypersensitive to caffeine can also have defects in autophagy [43].
Human LSM4 was indicated among the Differentially Expressed Autophagy-Related-Genes (DE-ARGs) in a study that aimed to find interactions between autophagy and hepatocellular carcinoma (HCC) pathogenesis [44] but, up to now, there is no evidence on the direct involvement of LSM4 in autophagy [44]: our results shown here demonstrate, by mean of GFP-ATG8 fusion protein, that a defect in autophagy induction upon nitrogen starvation was present in the Sclsm4Δ1 mutant.
A confirmation that the Sclsm4Δ1 mutant is defective in autophagy induction is the quick loss of viability in nitrogen starvation and in the presence of low doses of rapamycin.
Rapamycin is known to have a protective effect against oxidative stress, but at 6nM concentration it didn’t exert a protective effect on the wild type cells challenged with hydrogen peroxide, probably because rapamycin concentration employed in this experiment was too low. In the case of the Sclsm4Δ1 mutant it was not possible draw any conclusion due to the elevated toxicity of rapamycin in this mutant. High rapamycin sensitivity was also observed in the deletion mutant of LSM1, encoding a unit of the heptameric ring-shaped complex formed by Lsm1 to Lsm7, which, together with Pat1, is involved in mRNA degradation.
We also investigated the autophagic flux in our mutants by following by fluorescence microscopy the GFP-ATG8 fusion protein, which is a marker for phagophore assembly site (PAS) and autophagosome formation. We showed that GFP-ATG8 dots accumulate in the Sclsm4Δ1 mutant during nitrogen starvation, the diauxic phase and ageing.
It has been reported that under starvation conditions cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction [45]. Actually mitochondrial defects were described for Kllsm4Δ1 mutant in that it accumulated ROS, showed growth arrest on respiratory carbon sources and an aberrant mitochondrial morphology, with a punctuate distribution instead of the normal tubular shape [8,14].
The fact that Sclsm4Δ1 accumulates ROS intracellularly could be associated with autophagy induction defects, as it has been reported that the Atg4 protease activity is inhibited by oxidation in a H2O2 concentration-dependent manner [46]. Nevertheless, autophagosome formation is abolished in atg4Δ cells [47] while the Sclsm4Δ1 mutant, as also lsm1Δ mutant, showed an accumulation of GFP-Atg8 dots, suggesting that the autophagic defects are principally due to defects in a late stage of autophagosome formation preceding the fusion of mature autophagosomes with the vacuole or in the autophagosomes-vacuole fusion process itself.
There are evidences that Pat1-Lsm complex could be involved in these steps as, upon nitrogen starvation, the Pat1-Lsm complex binds and stabilizes a subset of ATG mRNA by preventing their exosome-mediated 3’→5’ degradation. Among these there is Atg1, a serine/threonine kinase homolog to human ULK-Kinase [28,40] that is considered a key regulator of autophagy. Atg1 phosphorylates the Atg4 protease, keeping it inactive preventing the premature release of Atg8 from autophagic membranes, and Ykt6, keeping this SNARE in an inactive state and so regulating also the autophagosome–vacuole fusion [48,49]. Another Atg1 target is Vps34, a class III phosphatidylinositol 3-kinase whose phosphorylation is important for full autophagy activation and cell survival [50]. Vps34, was mislocalized in mutants of the Nem1/Spo7–Pah1 axis, but localized at the right compartments after rapamycin treatment suggesting that Nem1/Spo7complex supports autophagy induction after TORC1 inactivation by nutrient starvation, probably via membrane synthesis [34].
We previously reported that in the Kllsm4Δ1 mutant the ER appear aberrant and the overexpression of NEM1, the catalytic subunit of the Nem1/Spo7–Pah1 axis, could rescue the Kllsm4Δ1 mutant phenotypes, suggesting that the Nem1/Spo7–Pah1 axis could be compromised in the Sclsm4Δ1 mutant [33].
The defects in the Nem1/Spo7–Pah1 axis, together with the possible high degradation of ATG1 mRNA, could concur to the observed autophagy defects in the Kllsm4Δ1 mutant. Nevertheless, to date it is still not possible to determine which is the main pathway affected in the observed Sclsm4Δ1 mutant defects and further investigations will be needed to clarify this important point.
It has been recently reported that phosphorylation of Edc3, a p-bodies component, has an effect on tumours growth and invasion through controlling P-body formation and dynamics [51]. In a genome wide analysis, it has been reported that Atg1 could phosphorylate also Lsm4 [52]. As LSM4 is also involved in some cancers, it will be interesting to use the simple yeast model to find LSM4-targets for the development of antitumoral molecules.
4. Materials and Methods
4.1. Yeast strains, growth conditions and plasmids construction
S. cerevisiae strains used in this work are described in Table 1. Cells were grown at 28°C in YPD (1% yeast extract (BD, #212750), 2% bacto-peptone (BD, #211677), 2% glucose), SD (0,67% yeast nitrogen base without aminoacids (BD, #291940), 2% glucose) supplemented with auxotrophic requirements. For autophagic induction by nitrogen starvation cells were grown in SD-N (0,17% yeast nitrogen base without aminoacids and ammonium sulphate (BD, #233520), 2% glucose). Solid media were obtained by the addition of 2% Bactoagar (BD, #214010).
Plasmid pRS313/Sclsm4Δ1 was obtained by amplifying 868 bp of the ScLSM4 gene, comprising the promoter region and the gene portion encoding the first 84 aminoacids, and then cloning the PCR fragment with BamH1/SacI extremities in the specific site of the vector (primers listed in Table 2). E. coli DH5α cells were used to amplify the vector. MCY4 transformation to give the strain MCY4/Sclsm4Δ1 was performed by ONE-STEP method [53] with ONE-STEP buffer (PEG 3350 40%, LiAc 0,2 M, DTT 0,1 M and ssDNA carrier 0,1 μg/μl (Sigma-Aldrich, D1626)) as transformation mix.
Plasmid pRS313/ScLSM4 was obtained by amplifying 1308 bp of the ScLSM4 gene, comprising the promoter region and the complete coding region, and then cloning the PCR fragment with BamH1/SacI extremities in the specific site of the vector (primers listed in Table 2). E. coli DH5α cells were used to amplify the vector. MCY4 transformation was performed by ONE-STEP method [53] to give the strain MCY4/ScLSM4.
Plasmid pUG36/ATG8 was a courtesy of Tobias Eisenberg and colleagues [54]. Transformation of the selected strains was performed by ONE-STEP method [53].

Table 1.
S. cerevisiae strains used in this work.
| Strain | Genotype | Source |
|---|---|---|
| MCY4 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 | [56] |
| MCY4/Kllsm4Δ1 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/Kllsm4Δ1 | [42] |
| MCY4/Sclsm4Δ1 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/Sclsm4Δ1 | This work |
| CML39-11A | MATa, ade1-101, his3-Δ1, leu2, ura3, trp1-289 | [8] |
| MCY4/ScLSM4 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/ScLSM4 | This work |
| MCY4/Kllsm4Δ1 pUG36/ATG8 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/Kllsm4Δ1, pUG36/ATG8 | This work |
| MCY4/Sclsm4Δ1 pUG36/ATG8 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/Sclsm4Δ1, pUG36/ATG8 | This work |
| CML39-11A pUG36/ATG8 | MATa, ade1-101, his3-Δ1, leu2, ura3, trp1-289 pUG36/ATG8 | This work |
| MCY4/ScLSM4 pUG36/ATG8 | MATα, ade1-101, his3-Δ1, trp1-289, ura3, LEU-GAL1-SDB23 pRS313/ScLSM4, pUG36/ATG8 | This work |
| BMA38 | Matα, ura3-1, leu2-3, -112, ade2-1, can1-100, his3-11, -15, trp1Δ1 | [39] |
| BMA38 lsm1Δ | Matα, ura3-1, leu2-3, -112, ade2-1, can1-100, his3-11, -15, trp1Δ1, lsm1Δ::TRP1 | [39] |
| BY4741 | Mat a, his3Δ1, leu2Δ0, met15Δ0, ura3Δ0 | [57] |
| Lsm1Δ | ||
| Lsm6Δ | ||
| Lsm7Δ | ||
| Atg5Δ | ||
| Dhh1Δ |
Table 2.
Amplification and cloning of the N-terminus truncated ScLSM4 gene.
| Primer Name | Oligonucleotide Sequence |
|---|---|
| BamH1-ScLSM4/Sclsm4Δ1 Fw | 5’-AAAAAAGGATCCGTACGCAGTCACAATGCGG-3’ |
| SacI-ScLSM4 Rv | 5’-GGGGGGAGCTCACCTGTAAACTAAAGGAAAGCTCG-3’ |
| SacI-Sclsm4Δ1 Rv | 5’-GGGGGGAGCTCTTATCTTGCAATTTGATAAACTTGATAAAAGTCC-3’ |
4.2. Viability assays
Stationary cultures of strain MCY4/Sclsm4Δ1 and CML39-11A were tested for microcolony forming ability during chronological lifespan in SD and SD-N media. 3x104 cells were daily plated on a YPD coated slide and analysed with optic microscope after 1-2 days of incubation at 28°C. Cell viability was calculated as the percentage of microcolony forming cells [55].
4.3. Rapamycin treatment
Strains MCY4/Sclsm4Δ1 and CML39-11A were tested for the microcolony forming ability after treatment with 6 nM of rapamycin (Sigma-Aldrich, R8781). Cells were inoculated in SD medium (controls) or SD supplemented with 6 nM of rapamycin (test samples). After 1, 2, 3 and 4 hours of treatment, 3x104 cells were plated on a YPD coated slide and analysed with optic microscope after 1-2 days of incubation at 28°C. Cell viability was calculated as the percentage of microcolony forming cells.
4.4. H2O2 sensitivity test
0,2-0,3 OD600 of culture was incubated for 4h at 28°C with 0,8 mM, 1,2 mM, 3 mM of H2O2 (Sigma-Aldrich, #216763) and then grown on a YPD coated slide for 24h and analysed with optic microscope. Cell viability was calculated as the percentage of microcolony forming cells.
4.5. Caffeine, acetic acid and rapamycin sensitivity test
Serial dilutions of strains MCY4/Sclsm4Δ1 and MCY4/ScLSM4 were spotted on YPD, YPY, YPD+0,25% caffeine, YPD+60 mM acetic acid or YPD+6 nM rapamycin and their viability was detected after 2-3 days of incubation at 28°C. For the testing in other genomic contexts, concentration of the compounds was adapted to maximise the differences between the wildtype and the mutants.
4.6. Fluorescence microscopy
Nuclear morphology was detected with DAPI (Sigma-Aldrich, D8417) staining of exponentially growing cells fixed with 70% (v/v) ethanol. Oxygen reactive species (ROS) were detected by incubating the cells with 5 μg/ml of DHR (Sigma-Aldrich, D1054) for 4h at 28°C and then analysed at fluorescence microscopy (Axioskop 2, Carl Zeiss, Germany). The visualisation of autophagosome formation and translocation was performed using the reporter plasmid pUG36/ATG8 and analysed at the same fluorescence microscopy. The percentage of GFP-ATG8 dots was calculated among the total number of fluorescent-positive cells.
4.7. Protein extraction and Western Blot analysis of autophagy induced cells
Decapping mutant and the wild type strain were grown on SD and SD-N media supplemented with auxotrophic requirements (except for SD-N) at 28°C and then harvested at their logarithmic growth phase (Exp) and post-diauxic phase (after 16h). The amount of cells corresponding to 4 OD600 were washed with H2O, resuspended in 200 μl of NaOH 2 M/β-mercaptoethanol 5% and then chilled on ice for 10′. Protein precipitation was performed with TCA at a final concentration of 8,3%, centrifugation at 13000 rpm for 15′ and pellet suspended in 100 μl of loading buffer (50 mM Tris-HCl pH6,8; 100 mM β-mercaptoethanol; 2% SDS, 0,1% bromophenol blue; 10% glycerol). Samples were then boiled for 5′ and loaded into 12% acrylamide SDS-PAGE gel. A protein marker was loaded in the first lane (Thermo-Fisher, LC5925). Separated proteins were transferred onto nitrocellulose membrane through electroblotting. Ponceau red staining was used as a loading control (0,1% Ponceau S (Sigma-Aldrich, P-3504), 5% acetic acid). Autophagic cargo processing was studied via immunoblotting analysis using anti-GFP antibody (α-mouse-GFP, Santa Cruz Biotechnology, sc-9996) to detect GFP-Atg8, as described [35]. The secondary antibody HRP-associated was sc-2060 Santa Cruz anti-mouse (goat).
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: lsm1Δ mutant shows defects in autophagy-related structures transport to the vacuole.
Author Contributions
Conceptualization, C.M. and C.F.; investigation and data acquisition, B.C., M.S., V.P., U.V.; writing—original draft preparation, C.M.; writing—review and editing, B.C., M.M.B., C.F., C.M.; funding acquisition, C.M., All authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Sapienza University of Rome grant numbers RM11916B4B7F5DDA, RP120172A30C991B and RG12218166E3EE68.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article or Supplementary Material.
Acknowledgments
We thank Dr. Tobias Eisenberg and colleagues for plasmid pUG36/ATG8 and Dr J. D. Beggs for kindly providing the S. cerevisiae strain MCY4. We thank Andrea Marrani for technical help in performing some of the experiments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- C.J. Wilusz, J. Wilusz, Lsm proteins and Hfq: Life at the 3′ end, RNA Biol. 10 (2013) 592–601. [CrossRef]
- W. He, R. Parker, Functions of Lsm proteins in mRNA degradation and splicing, Curr. Opin. Cell Biol. 12 (2000) 346–350. [CrossRef]
- Z. Chen, C. Han, X. Zhou, X. Wang, X. Liao, Y. He, S. Mo, X. Li, G. Zhu, X. Ye, T. Peng, Prognostic value and potential molecular mechanism of the like-Sm gene family in early-stage pancreatic ductal adenocarcinoma, Transl. Cancer Res. 10 (2021) 1744–1760. [CrossRef]
- H.D.K. Ta, WotherJ. Wang, N.N. Phan, N.T. An Ton, G. Anuraga, S.-C. Ku, Y.-F. Wu, C.-Y. Wang, K.-H. Lee, Potential Therapeutic and Prognostic Values of LSM Family Genes in Breast Cancer, Cancers. 13 (2021) 4902. [CrossRef]
- Z.-P. Sun, Z.-G. Tan, C. Peng, Long noncoding RNA LINC01419 promotes hepatocellular carcinoma malignancy by mediating miR-485-5p/LSM4 axis, Kaohsiung J. Med. Sci. 38 (2022) 826–838. [CrossRef]
- C. Mazzoni, P. Mancini, F. Madeo, V. Palermo, C. Falcone, A Kluyveromyces lactis mutant in the essential gene KlLSM4 shows phenotypic markers of apoptosis, FEMS Yeast Res. 4 (2003) 29–35. [CrossRef]
- C. Mazzoni, P. Mancini, L. Verdone, F. Madeo, A. Serafini, E. Herker, C. Falcone, A Truncated Form of KlLsm4p and the Absence of Factors Involved in mRNA Decapping Trigger Apoptosis in Yeast, Mol. Biol. Cell. 14 (2003) 721–729. [CrossRef]
- C. Mazzoni, E. Herker, V. Palermo, H. Jungwirth, T. Eisenberg, F. Madeo, C. Falcone, Yeast caspase 1 links messenger RNA stability to apoptosis in yeast, EMBO Rep. 6 (2005) 1076–1081. [CrossRef]
- C. Mazzoni, I. D’Addario, C. Falcone, The C-terminus of the yeast Lsm4p is required for the association to P-bodies, FEBS Lett. 581 (2007) 4836–4840. [CrossRef]
- M.A.M. Reijns, R.D. Alexander, M.P. Spiller, J.D. Beggs, A role for Q/N-rich aggregation-prone regions in P-body localization, J. Cell Sci. 121 (2008) 2463–2472. [CrossRef]
- C.J. Decker, D. Teixeira, R. Parker, Edc3p and a glutamine/asparagine-rich domain of Lsm4p function in processing body assembly in Saccharomyces cerevisiae, J. Cell Biol. 179 (2007) 437–449. [CrossRef]
- M. Kato, T.W. Han, S. Xie, K. Shi, X. Du, L.C. Wu, H. Mirzaei, E.J. Goldsmith, J. Longgood, J. Pei, N.V. Grishin, D.E. Frantz, J.W. Schneider, S. Chen, L. Li, M.R. Sawaya, D. Eisenberg, R. Tycko, S.L. McKnight, Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels, Cell. 149 (2012) 753–767. [CrossRef]
- M. Arribas-Layton, J. Dennis, E.J. Bennett, C.K. Damgaard, J. Lykke-Andersen, The C-Terminal RGG Domain of Human Lsm4 Promotes Processing Body Formation Stimulated by Arginine Dimethylation, Mol. Cell. Biol. 36 (2016) 2226–2235. [CrossRef]
- V. Palermo, E. Cundari, E. Mangiapelo, C. Falcone, C. Mazzoni, Yeast lsm pro-apoptotic mutants show defects in S-phase entry and progression, Cell Cycle Georget. Tex. 9 (2010) 3991–3996. [CrossRef]
- S. Lyons, A. Ricciardi, A. Guo, C. Kambach, W. Marzluff, The C-terminal extension of Lsm4 interacts directly with the 3′ end of the histone mRNP and is required for efficient histone mRNA degradation, RNA N. Y. N. 20 (2014). [CrossRef]
- M. Costanzo, A. Baryshnikova, J. Bellay, Y. Kim, E.D. Spear, C.S. Sevier, H. Ding, J.L.Y. Koh, K. Toufighi, S. Mostafavi, J. Prinz, R.P. St Onge, B. VanderSluis, T. Makhnevych, F.J. Vizeacoumar, S. Alizadeh, S. Bahr, R.L. Brost, Y. Chen, M. Cokol, R. Deshpande, Z. Li, Z.-Y. Lin, W. Liang, M. Marback, J. Paw, B.-J. San Luis, E. Shuteriqi, A.H.Y. Tong, N. van Dyk, I.M. Wallace, J.A. Whitney, M.T. Weirauch, G. Zhong, H. Zhu, W.A. Houry, M. Brudno, S. Ragibizadeh, B. Papp, C. Pál, F.P. Roth, G. Giaever, C. Nislow, O.G. Troyanskaya, H. Bussey, G.D. Bader, A.-C. Gingras, Q.D. Morris, P.M. Kim, C.A. Kaiser, C.L. Myers, B.J. Andrews, C. Boone, The genetic landscape of a cell, Science. 327 (2010) 425–431. [CrossRef]
- C. Mazzoni, C. Falcone, mRNA stability and control of cell proliferation, Biochem. Soc. Trans. 39 (2011) 1461–1465. [CrossRef]
- S.F. Mitchell, S. Jain, M. She, R. Parker, Global analysis of yeast mRNPs, Nat. Struct. Mol. Biol. 20 (2013) 127–133. [CrossRef]
- E. Delorme-Axford, D.J. Klionsky, On the edge of degradation: Autophagy regulation by RNA decay, Wiley Interdiscip. Rev. RNA. 10 (2019) e1522. [CrossRef]
- G. Hu, T. McQuiston, A. Bernard, Y.-D. Park, J. Qiu, A. Vural, N. Zhang, S.R. Waterman, N.H. Blewett, T.G. Myers, R.J. Maraia, J.H. Kehrl, G. Uzel, D.J. Klionsky, P.R. Williamson, A conserved mechanism of TOR-dependent RCK-mediated mRNA degradation regulates autophagy, Nat. Cell Biol. 17 (2015) 930–942. [CrossRef]
- X. Liu, M. Jin, Z. Yao, A. Bernard, D.J. Klionsky, Bidirectional roles of Dhh1 in regulating autophagy, Autophagy. 15 (2019) 1838–1839. [CrossRef]
- Z. Yin, Z. Zhang, Y. Lei, D.J. Klionsky, Bidirectional roles of the Ccr4-Not complex in regulating autophagy before and after nitrogen starvation, Autophagy. 19 (2023) 415–425. [CrossRef]
- Chowdhury, J. Mukhopadhyay, S. Tharun, The decapping activator Lsm1p-7p–Pat1p complex has the intrinsic ability to distinguish between oligoadenylated and polyadenylated RNAs, RNA. 13 (2007) 998–1016. [CrossRef]
- Chowdhury, S. Kalurupalle, S. Tharun, Pat1 contributes to the RNA binding activity of the Lsm1-7–Pat1 complex, RNA. 20 (2014) 1465–1475. [CrossRef]
- W. He, R. Parker, The yeast cytoplasmic LsmI/Pat1p complex protects mRNA 3′ termini from partial degradation, Genetics. 158 (2001) 1445–1455. [CrossRef]
- S. Tharun, D. Muhlrad, A. Chowdhury, R. Parker, Mutations in the Saccharomyces cerevisiae LSM1 gene that affect mRNA decapping and 3′ end protection, Genetics. 170 (2005) 33–46. [CrossRef]
- Chowdhury, S. Tharun, Activation of decapping involves binding of the mRNA and facilitation of the post-binding steps by the Lsm1-7-Pat1 complex, RNA N. Y. N. 15 (2009) 1837–1848. [CrossRef]
- D. Gatica, G. Hu, N. Zhang, P.R. Williamson, D.J. Klionsky, The Pat1-Lsm complex prevents 3′ to 5′ degradation of a specific subset of ATG mRNAs during nitrogen starvation-induced autophagy, Autophagy. 15 (2019) 750–751. [CrossRef]
- H. Huang, T. Kawamata, T. Horie, H. Tsugawa, Y. Nakayama, Y. Ohsumi, E. Fukusaki, Bulk RNA degradation by nitrogen starvation-induced autophagy in yeast, EMBO J. 34 (2015) 154–168. [CrossRef]
- S. Makino, T. Kawamata, S. Iwasaki, Y. Ohsumi, Selectivity of mRNA degradation by autophagy in yeast, Nat. Commun. 12 (2021) 2316. [CrossRef]
- M. Stirpe, V. Palermo, M. Ferrari, S. Mroczek, J. Kufel, C. Falcone, C. Mazzoni, Increased levels of RNA oxidation enhance the reversion frequency in aging pro-apoptotic yeast mutants, Apoptosis. 22 (2017) 200–206. [CrossRef]
- D. Carmona-Gutierrez, M.A. Bauer, A. Zimmermann, A. Aguilera, N. Austriaco, K. Ayscough, R. Balzan, S. Bar-Nun, A. Barrientos, P. Belenky, M. Blondel, R.J. Braun, M. Breitenbach, W.C. Burhans, S. Büttner, D. Cavalieri, M. Chang, K.F. Cooper, M. Côrte-Real, V. Costa, C. Cullin, I. Dawes, J. Dengjel, M.B. Dickman, T. Eisenberg, B. Fahrenkrog, N. Fasel, K.-U. Fröhlich, A. Gargouri, S. Giannattasio, P. Goffrini, C.W. Gourlay, C.M. Grant, M.T. Greenwood, N. Guaragnella, T. Heger, J. Heinisch, E. Herker, J.M. Herrmann, S. Hofer, A. Jiménez-Ruiz, H. Jungwirth, K. Kainz, D.P. Kontoyiannis, P. Ludovico, S. Manon, E. Martegani, C. Mazzoni, L.A. Megeney, C. Meisinger, J. Nielsen, T. Nyström, H.D. Osiewacz, T.F. Outeiro, H.-O. Park, T. Pendl, D. Petranovic, S. Picot, P. Polčic, T. Powers, M. Ramsdale, M. Rinnerthaler, P. Rockenfeller, C. Ruckenstuhl, R. Schaffrath, M. Segovia, F.F. Severin, A. Sharon, S.J. Sigrist, C. Sommer-Ruck, M.J. Sousa, J.M. Thevelein, K. Thevissen, V. Titorenko, M.B. Toledano, M. Tuite, F.-N. Vögtle, B. Westermann, J. Winderickx, S. Wissing, S. Wölfl, Z.J. Zhang, R.Y. Zhao, B. Zhou, L. Galluzzi, G. Kroemer, F. Madeo, Guidelines and recommendations on yeast cell death nomenclature, Microb. Cell Graz Austria. 5 (2018) 4–31. [CrossRef]
- V. Palermo, M. Stirpe, M. Torella, C. Falcone, C. Mazzoni, NEM1 acts as a suppressor of apoptotic phenotypes in LSM4 yeast mutants, FEMS Yeast Res. 15 (2015). [CrossRef]
- M.A. Rahman, M.G. Mostofa, T. Ushimaru, The Nem1/Spo7-Pah1/lipin axis is required for autophagy induction after TORC1 inactivation, FEBS J. 285 (2018) 1840–1860. [CrossRef]
- D.J. Klionsky, A.K. Abdel-Aziz, S. Abdelfatah, M. Abdellatif, A. Abdoli, S. Abel, H. Abeliovich, M.H. Abildgaard, Y.P. Abudu, A. Acevedo-Arozena, I.E. Adamopoulos, K. Adeli, T.E. Adolph, A. Adornetto, E. Aflaki, G. Agam, A. Agarwal, B.B. Aggarwal, M. Agnello, P. Agostinis, J.N. Agrewala, A. Agrotis, P.V. Aguilar, S.T. Ahmad, Z.M. Ahmed, U. Ahumada-Castro, S. Aits, S. Aizawa, Y. Akkoc, T. Akoumianaki, H.A. Akpinar, A.M. Al-Abd, L. Al-Akra, A. Al-Gharaibeh, M.A. Alaoui-Jamali, S. Alberti, E. Alcocer-Gómez, C. Alessandri, M. Ali, M.A. Alim Al-Bari, S. Aliwaini, J. Alizadeh, E. Almacellas, A. Almasan, A. Alonso, G.D. Alonso, N. Altan-Bonnet, D.C. Altieri, É.M.C. Álvarez, S. Alves, C. Alves da Costa, M.M. Alzaharna, M. Amadio, C. Amantini, C. Amaral, S. Ambrosio, A.O. Amer, V. Ammanathan, Z. An, S.U. Andersen, S.A. Andrabi, M. Andrade-Silva, A.M. Andres, S. Angelini, D. Ann, U.C. Anozie, M.Y. Ansari, P. Antas, A. Antebi, Z. Antón, T. Anwar, L. Apetoh, N. Apostolova, T. Araki, Y. Araki, K. Arasaki, W.L. Araújo, J. Araya, C. Arden, M.-A. Arévalo, S. Arguelles, E. Arias, J. Arikkath, H. Arimoto, A.R. Ariosa, D. Armstrong-James, L. Arnauné-Pelloquin, A. Aroca, D.S. Arroyo, I. Arsov, R. Artero, D.M.L. Asaro, M. Aschner, M. Ashrafizadeh, O. Ashur-Fabian, A.G. Atanasov, A.K. Au, P. Auberger, H.W. Auner, L. Aurelian, R. Autelli, L. Avagliano, Y. Ávalos, S. Aveic, C.A. Aveleira, T. Avin-Wittenberg, Y. Aydin, S. Ayton, S. Ayyadevara, M. Azzopardi, M. Baba, J.M. Backer, S.K. Backues, D.-H. Bae, O.-N. Bae, S.H. Bae, E.H. Baehrecke, A. Baek, S.-H. Baek, S.H. Baek, G. Bagetta, A. Bagniewska-Zadworna, H. Bai, J. Bai, X. Bai, Y. Bai, N. Bairagi, S. Baksi, T. Balbi, C.T. Baldari, W. Balduini, A. Ballabio, M. Ballester, S. Balazadeh, R. Balzan, R. Bandopadhyay, S. Banerjee, S. Banerjee, Á. Bánréti, Y. Bao, M.S. Baptista, A. Baracca, C. Barbati, A. Bargiela, D. Barilà, P.G. Barlow, S.J. Barmada, E. Barreiro, G.E. Barreto, J. Bartek, B. Bartel, A. Bartolome, G.R. Barve, S.H. Basagoudanavar, D.C. Bassham, R.C. Bast, A. Basu, H. Batoko, I. Batten, E.E. Baulieu, B.L. Baumgarner, J. Bayry, R. Beale, I. Beau, F. Beaumatin, L.R.G. Bechara, G.R. Beck, M.F. Beers, J. Begun, C. Behrends, G.M.N. Behrens, R. Bei, E. Bejarano, S. Bel, C. Behl, A. Belaid, N. Belgareh-Touzé, C. Bellarosa, F. Belleudi, M. Belló Pérez, R. Bello-Morales, J.S. de O. Beltran, S. Beltran, D.M. Benbrook, M. Bendorius, B.A. Benitez, I. Benito-Cuesta, J. Bensalem, M.W. Berchtold, S. Berezowska, D. Bergamaschi, M. Bergami, A. Bergmann, L. Berliocchi, C. Berlioz-Torrent, A. Bernard, L. Berthoux, C.G. Besirli, S. Besteiro, V.M. Betin, R. Beyaert, J.S. Bezbradica, K. Bhaskar, I. Bhatia-Kissova, R. Bhattacharya, S. Bhattacharya, S. Bhattacharyya, M.S. Bhuiyan, S.K. Bhutia, L. Bi, X. Bi, T.J. Biden, K. Bijian, V.A. Billes, N. Binart, C. Bincoletto, A.B. Birgisdottir, G. Bjorkoy, G. Blanco, A. Blas-Garcia, J. Blasiak, R. Blomgran, K. Blomgren, J.S. Blum, E. Boada-Romero, M. Boban, K. Boesze-Battaglia, P. Boeuf, B. Boland, P. Bomont, P. Bonaldo, S.R. Bonam, L. Bonfili, J.S. Bonifacino, B.A. Boone, M.D. Bootman, M. Bordi, C. Borner, B.C. Bornhauser, G. Borthakur, J. Bosch, S. Bose, L.M. Botana, J. Botas, C.M. Boulanger, M.E. Boulton, M. Bourdenx, B. Bourgeois, N.M. Bourke, G. Bousquet, P. Boya, P.V. Bozhkov, L.H.M. Bozi, T.O. Bozkurt, D.E. Brackney, C.H. Brandts, R.J. Braun, G.H. Braus, R. Bravo-Sagua, J.M. Bravo-San Pedro, P. Brest, M.-A. Bringer, A. Briones-Herrera, V.C. Broaddus, P. Brodersen, J.L. Brodsky, S.L. Brody, P.G. Bronson, J.M. Bronstein, C.N. Brown, R.E. Brown, P.C. Brum, J.H. Brumell, N. Brunetti-Pierri, D. Bruno, R.J. Bryson-Richardson, C. Bucci, C. Buchrieser, M. Bueno, L.E. Buitrago-Molina, S. Buraschi, S. Buch, J.R. Buchan, E.M. Buckingham, H. Budak, M. Budini, G. Bultynck, F. Burada, J.R. Burgoyne, M.I. Burón, V. Bustos, S. Büttner, E. Butturini, A. Byrd, I. Cabas, S. Cabrera-Benitez, K. Cadwell, J. Cai, L. Cai, Q. Cai, M. Cairó, J.A. Calbet, G.A. Caldwell, K.A. Caldwell, J.A. Call, R. Calvani, A.C. Calvo, M. Calvo-Rubio Barrera, N.O. Camara, J.H. Camonis, N. Camougrand, M. Campanella, E.M. Campbell, F.-X. Campbell-Valois, S. Campello, I. Campesi, J.C. Campos, O. Camuzard, J. Cancino, D. Candido de Almeida, L. Canesi, I. Caniggia, B. Canonico, C. Cantí, B. Cao, M. Caraglia, B. Caramés, E.H. Carchman, E. Cardenal-Muñoz, C. Cardenas, L. Cardenas, S.M. Cardoso, J.S. Carew, G.F. Carle, G. Carleton, S. Carloni, D. Carmona-Gutierrez, L.A. Carneiro, O. Carnevali, J.M. Carosi, S. Carra, A. Carrier, L. Carrier, B. Carroll, A.B. Carter, A.N. Carvalho, M. Casanova, C. Casas, J. Casas, C. Cassioli, E.F. Castillo, K. Castillo, S. Castillo-Lluva, F. Castoldi, M. Castori, A.F. Castro, M. Castro-Caldas, J. Castro-Hernandez, S. Castro-Obregon, S.D. Catz, C. Cavadas, F. Cavaliere, G. Cavallini, M. Cavinato, M.L. Cayuela, P. Cebollada Rica, V. Cecarini, F. Cecconi, M. Cechowska-Pasko, S. Cenci, V. Ceperuelo-Mallafré, J.J. Cerqueira, J.M. Cerutti, D. Cervia, V.B. Cetintas, S. Cetrullo, H.-J. Chae, A.S. Chagin, C.-Y. Chai, G. Chakrabarti, O. Chakrabarti, T. Chakraborty, T. Chakraborty, M. Chami, G. Chamilos, D.W. Chan, E.Y.W. Chan, E.D. Chan, H.Y.E. Chan, H.H. Chan, H. Chan, M.T.V. Chan, Y.S. Chan, P.K. Chandra, C.-P. Chang, C. Chang, H.-C. Chang, K. Chang, J. Chao, T. Chapman, N. Charlet-Berguerand, S. Chatterjee, S.K. Chaube, A. Chaudhary, S. Chauhan, E. Chaum, F. Checler, M.E. Cheetham, C.-S. Chen, G.-C. Chen, J.-F. Chen, L.L. Chen, L. Chen, L. Chen, M. Chen, M.-K. Chen, N. Chen, Q. Chen, R.-H. Chen, S. Chen, W. Chen, W. Chen, X.-M. Chen, X.-W. Chen, X. Chen, Y. Chen, Y.-G. Chen, Y. Chen, Y. Chen, Y.-J. Chen, Y.-Q. Chen, Z.S. Chen, Z. Chen, Z.-H. Chen, Z.J. Chen, Z. Chen, H. Cheng, J. Cheng, S.-Y. Cheng, W. Cheng, X. Cheng, X.-T. Cheng, Y. Cheng, Z. Cheng, Z. Chen, H. Cheong, J.K. Cheong, B.V. Chernyak, S. Cherry, C.F.R. Cheung, C.H.A. Cheung, K.-H. Cheung, E. Chevet, R.J. Chi, A.K.S. Chiang, F. Chiaradonna, R. Chiarelli, M. Chiariello, N. Chica, S. Chiocca, M. Chiong, S.-H. Chiou, A.I. Chiramel, V. Chiurchiù, D.-H. Cho, S.-K. Choe, A.M.K. Choi, M.E. Choi, K.R. Choudhury, N.S. Chow, C.T. Chu, J.P. Chua, J.J.E. Chua, H. Chung, K.P. Chung, S. Chung, S.-H. Chung, Y.-L. Chung, V. Cianfanelli, I.A. Ciechomska, M. Cifuentes, L. Cinque, S. Cirak, M. Cirone, M.J. Clague, R. Clarke, E. Clementi, E.M. Coccia, P. Codogno, E. Cohen, M.M. Cohen, T. Colasanti, F. Colasuonno, R.A. Colbert, A. Colell, M. Čolić, N.S. Coll, M.O. Collins, M.I. Colombo, D.A. Colón-Ramos, L. Combaret, S. Comincini, M.R. Cominetti, A. Consiglio, A. Conte, F. Conti, V.R. Contu, M.R. Cookson, K.M. Coombs, I. Coppens, M.T. Corasaniti, D.P. Corkery, N. Cordes, K. Cortese, M. do C. Costa, S. Costantino, P. Costelli, A. Coto-Montes, P.J. Crack, J.L. Crespo, A. Criollo, V. Crippa, R. Cristofani, T. Csizmadia, A. Cuadrado, B. Cui, J. Cui, Y. Cui, Y. Cui, E. Culetto, A.C. Cumino, A.V. Cybulsky, M.J. Czaja, S.J. Czuczwar, S. D’Adamo, M. D’Amelio, D. D’Arcangelo, A.C. D’Lugos, G. D’Orazi, J.A. da Silva, H.S. Dafsari, R.K. Dagda, Y. Dagdas, M. Daglia, X. Dai, Y. Dai, Y. Dai, J. Dal Col, P. Dalhaimer, L. Dalla Valle, T. Dallenga, G. Dalmasso, M. Damme, I. Dando, N.P. Dantuma, A.L. Darling, H. Das, S. Dasarathy, S.K. Dasari, S. Dash, O. Daumke, A.N. Dauphinee, J.S. Davies, V.A. Dávila, R.J. Davis, T. Davis, S. Dayalan Naidu, F. De Amicis, K. De Bosscher, F. De Felice, L. De Franceschi, C. De Leonibus, M.G. de Mattos Barbosa, G.R.Y. De Meyer, A. De Milito, C. De Nunzio, C. De Palma, M. De Santi, C. De Virgilio, D. De Zio, J. Debnath, B.J. DeBosch, J.-P. Decuypere, M.A. Deehan, G. Deflorian, J. DeGregori, B. Dehay, G. Del Rio, J.R. Delaney, L.M.D. Delbridge, E. Delorme-Axford, M.V. Delpino, F. Demarchi, V. Dembitz, N.D. Demers, H. Deng, Z. Deng, J. Dengjel, P. Dent, D. Denton, M.L. DePamphilis, C.J. Der, V. Deretic, A. Descoteaux, L. Devis, S. Devkota, O. Devuyst, G. Dewson, M. Dharmasivam, R. Dhiman, D. di Bernardo, M. Di Cristina, F. Di Domenico, P. Di Fazio, A. Di Fonzo, G. Di Guardo, G.M. Di Guglielmo, L. Di Leo, C. Di Malta, A. Di Nardo, M. Di Rienzo, F. Di Sano, G. Diallinas, J. Diao, G. Diaz-Araya, I. Díaz-Laviada, J.M. Dickinson, M. Diederich, M. Dieudé, I. Dikic, S. Ding, W.-X. Ding, L. Dini, J. Dinić, M. Dinic, A.T. Dinkova-Kostova, M.S. Dionne, J.H.W. Distler, A. Diwan, I.M.C. Dixon, M. Djavaheri-Mergny, I. Dobrinski, O. Dobrovinskaya, R. Dobrowolski, R.C.J. Dobson, J. Đokić, S. Dokmeci Emre, M. Donadelli, B. Dong, X. Dong, Z. Dong, G.W. Dorn Ii, V. Dotsch, H. Dou, J. Dou, M. Dowaidar, S. Dridi, L. Drucker, A. Du, C. Du, G. Du, H.-N. Du, L.-L. Du, A. du Toit, S.-B. Duan, X. Duan, S.P. Duarte, A. Dubrovska, E.A. Dunlop, N. Dupont, R.V. Durán, B.S. Dwarakanath, S.A. Dyshlovoy, D. Ebrahimi-Fakhari, L. Eckhart, C.L. Edelstein, T. Efferth, E. Eftekharpour, L. Eichinger, N. Eid, T. Eisenberg, N.T. Eissa, S. Eissa, M. Ejarque, A. El Andaloussi, N. El-Hage, S. El-Naggar, A.M. Eleuteri, E.S. El-Shafey, M. Elgendy, A.G. Eliopoulos, M.M. Elizalde, P.M. Elks, H.-P. Elsasser, E.S. Elsherbiny, B.M. Emerling, N.C.T. Emre, C.H. Eng, N. Engedal, A.-M. Engelbrecht, A.S.T. Engelsen, J.M. Enserink, R. Escalante, A. Esclatine, M. Escobar-Henriques, E.-L. Eskelinen, L. Espert, M.-O. Eusebio, G. Fabrias, C. Fabrizi, A. Facchiano, F. Facchiano, B. Fadeel, C. Fader, A.C. Faesen, W.D. Fairlie, A. Falcó, B.H. Falkenburger, D. Fan, J. Fan, Y. Fan, E.F. Fang, Y. Fang, Y. Fang, M. Fanto, T. Farfel-Becker, M. Faure, G. Fazeli, A.O. Fedele, A.M. Feldman, D. Feng, J. Feng, L. Feng, Y. Feng, Y. Feng, W. Feng, T. Fenz Araujo, T.A. Ferguson, Á.F. Fernández, J.C. Fernandez-Checa, S. Fernández-Veledo, A.R. Fernie, A.W. Ferrante, A. Ferraresi, M.F. Ferrari, J.C.B. Ferreira, S. Ferro-Novick, A. Figueras, R. Filadi, N. Filigheddu, E. Filippi-Chiela, G. Filomeni, G.M. Fimia, V. Fineschi, F. Finetti, S. Finkbeiner, E.A. Fisher, P.B. Fisher, F. Flamigni, S.J. Fliesler, T.H. Flo, I. Florance, O. Florey, T. Florio, E. Fodor, C. Follo, E.A. Fon, A. Forlino, F. Fornai, P. Fortini, A. Fracassi, A. Fraldi, B. Franco, R. Franco, F. Franconi, L.B. Frankel, S.L. Friedman, L.F. Fröhlich, G. Frühbeck, J.M. Fuentes, Y. Fujiki, N. Fujita, Y. Fujiwara, M. Fukuda, S. Fulda, L. Furic, N. Furuya, C. Fusco, M.U. Gack, L. Gaffke, S. Galadari, A. Galasso, M.F. Galindo, S. Gallolu Kankanamalage, L. Galluzzi, V. Galy, N. Gammoh, B. Gan, I.G. Ganley, F. Gao, H. Gao, M. Gao, P. Gao, S.-J. Gao, W. Gao, X. Gao, A. Garcera, M.N. Garcia, V.E. Garcia, F. García-Del Portillo, V. Garcia-Escudero, A. Garcia-Garcia, M. Garcia-Macia, D. García-Moreno, C. Garcia-Ruiz, P. García-Sanz, A.D. Garg, R. Gargini, T. Garofalo, R.F. Garry, N.C. Gassen, D. Gatica, L. Ge, W. Ge, R. Geiss-Friedlander, C. Gelfi, P. Genschik, I.E. Gentle, V. Gerbino, C. Gerhardt, K. Germain, M. Germain, D.A. Gewirtz, E. Ghasemipour Afshar, S. Ghavami, A. Ghigo, M. Ghosh, G. Giamas, C. Giampietri, A. Giatromanolaki, G.E. Gibson, S.B. Gibson, V. Ginet, E. Giniger, C. Giorgi, H. Girao, S.E. Girardin, M. Giridharan, S. Giuliano, C. Giulivi, S. Giuriato, J. Giustiniani, A. Gluschko, V. Goder, A. Goginashvili, J. Golab, D.C. Goldstone, A. Golebiewska, L.R. Gomes, R. Gomez, R. Gómez-Sánchez, M.C. Gomez-Puerto, R. Gomez-Sintes, Q. Gong, F.M. Goni, J. González-Gallego, T. Gonzalez-Hernandez, R.A. Gonzalez-Polo, J.A. Gonzalez-Reyes, P. González-Rodríguez, I.S. Goping, M.S. Gorbatyuk, N.V. Gorbunov, K. Görgülü, R.M. Gorojod, S.M. Gorski, S. Goruppi, C. Gotor, R.A. Gottlieb, I. Gozes, D. Gozuacik, M. Graef, M.H. Gräler, V. Granatiero, D. Grasso, J.P. Gray, D.R. Green, A. Greenhough, S.L. Gregory, E.F. Griffin, M.W. Grinstaff, F. Gros, C. Grose, A.S. Gross, F. Gruber, P. Grumati, T. Grune, X. Gu, J.-L. Guan, C.M. Guardia, K. Guda, F. Guerra, C. Guerri, P. Guha, C. Guillén, S. Gujar, A. Gukovskaya, I. Gukovsky, J. Gunst, A. Günther, A.R. Guntur, C. Guo, C. Guo, H. Guo, L.-W. Guo, M. Guo, P. Gupta, S.K. Gupta, S. Gupta, V.B. Gupta, V. Gupta, A.B. Gustafsson, D.D. Gutterman, R. H B, A. Haapasalo, J.E. Haber, A. Hać, S. Hadano, A.J. Hafrén, M. Haidar, B.S. Hall, G. Halldén, A. Hamacher-Brady, A. Hamann, M. Hamasaki, W. Han, M. Hansen, P.I. Hanson, Z. Hao, M. Harada, L. Harhaji-Trajkovic, N. Hariharan, N. Haroon, J. Harris, T. Hasegawa, N. Hasima Nagoor, J.A. Haspel, V. Haucke, W.D. Hawkins, B.A. Hay, C.M. Haynes, S.B. Hayrabedyan, T.S. Hays, C. He, Q. He, R.-R. He, Y.-W. He, Y.-Y. He, Y. Heakal, A.M. Heberle, J.F. Hejtmancik, G.V. Helgason, V. Henkel, M. Herb, A. Hergovich, A. Herman-Antosiewicz, A. Hernández, C. Hernandez, S. Hernandez-Diaz, V. Hernandez-Gea, A. Herpin, J. Herreros, J.H. Hervás, D. Hesselson, C. Hetz, V.T. Heussler, Y. Higuchi, S. Hilfiker, J.A. Hill, W.S. Hlavacek, E.A. Ho, I.H.T. Ho, P.W.-L. Ho, S.-L. Ho, W.Y. Ho, G.A. Hobbs, M. Hochstrasser, P.H.M. Hoet, D. Hofius, P. Hofman, A. Höhn, C.I. Holmberg, J.R. Hombrebueno, C.-W.H. Yi-Ren Hong, L.V. Hooper, T. Hoppe, R. Horos, Y. Hoshida, I.-L. Hsin, H.-Y. Hsu, B. Hu, D. Hu, L.-F. Hu, M.C. Hu, R. Hu, W. Hu, Y.-C. Hu, Z.-W. Hu, F. Hua, J. Hua, Y. Hua, C. Huan, C. Huang, C. Huang, C. Huang, C. Huang, H. Huang, K. Huang, M.L.H. Huang, R. Huang, S. Huang, T. Huang, X. Huang, Y.J. Huang, T.B. Huber, V. Hubert, C.A. Hubner, S.M. Hughes, W.E. Hughes, M. Humbert, G. Hummer, J.H. Hurley, S. Hussain, S. Hussain, P.J. Hussey, M. Hutabarat, H.-Y. Hwang, S. Hwang, A. Ieni, F. Ikeda, Y. Imagawa, Y. Imai, C. Imbriano, M. Imoto, D.M. Inman, K. Inoki, J. Iovanna, R.V. Iozzo, G. Ippolito, J.E. Irazoqui, P. Iribarren, M. Ishaq, M. Ishikawa, N. Ishimwe, C. Isidoro, N. Ismail, S. Issazadeh-Navikas, E. Itakura, D. Ito, D. Ivankovic, S. Ivanova, A.K.V. Iyer, J.M. Izquierdo, M. Izumi, M. Jäättelä, M.S. Jabir, W.T. Jackson, N. Jacobo-Herrera, A.-C. Jacomin, E. Jacquin, P. Jadiya, H. Jaeschke, C. Jagannath, A.J. Jakobi, J. Jakobsson, B. Janji, P. Jansen-Dürr, P.J. Jansson, J. Jantsch, S. Januszewski, A. Jassey, S. Jean, H. Jeltsch-David, P. Jendelova, A. Jenny, T.E. Jensen, N. Jessen, J.L. Jewell, J. Ji, L. Jia, R. Jia, L. Jiang, Q. Jiang, R. Jiang, T. Jiang, X. Jiang, Y. Jiang, M. Jimenez-Sanchez, E.-J. Jin, F. Jin, H. Jin, L. Jin, L. Jin, M. Jin, S. Jin, E.-K. Jo, C. Joffre, T. Johansen, G.V.W. Johnson, S.A. Johnston, E. Jokitalo, M.K. Jolly, L.A.B. Joosten, J. Jordan, B. Joseph, D. Ju, J.-S. Ju, J. Ju, E. Juárez, D. Judith, G. Juhász, Y. Jun, C.H. Jung, S.-C. Jung, Y.K. Jung, H. Jungbluth, J. Jungverdorben, S. Just, K. Kaarniranta, A. Kaasik, T. Kabuta, D. Kaganovich, A. Kahana, R. Kain, S. Kajimura, M. Kalamvoki, M. Kalia, D.S. Kalinowski, N. Kaludercic, I. Kalvari, J. Kaminska, V.O. Kaminskyy, H. Kanamori, K. Kanasaki, C. Kang, R. Kang, S.S. Kang, S. Kaniyappan, T. Kanki, T.-D. Kanneganti, A.G. Kanthasamy, A. Kanthasamy, M. Kantorow, O. Kapuy, M.V. Karamouzis, M.R. Karim, P. Karmakar, R.G. Katare, M. Kato, S.H.E. Kaufmann, A. Kauppinen, G.P. Kaushal, S. Kaushik, K. Kawasaki, K. Kazan, P.-Y. Ke, D.J. Keating, U. Keber, J.H. Kehrl, K.E. Keller, C.W. Keller, J.K. Kemper, C.M. Kenific, O. Kepp, S. Kermorgant, A. Kern, R. Ketteler, T.G. Keulers, B. Khalfin, H. Khalil, B. Khambu, S.Y. Khan, V.K.M. Khandelwal, R. Khandia, W. Kho, N.V. Khobrekar, S. Khuansuwan, M. Khundadze, S.A. Killackey, D. Kim, D.R. Kim, D.-H. Kim, D.-E. Kim, E.Y. Kim, E.-K. Kim, H.-R. Kim, H.-S. Kim, null Hyung-Ryong Kim, J.H. Kim, J.K. Kim, J.-H. Kim, J. Kim, J.H. Kim, K.I. Kim, P.K. Kim, S.-J. Kim, S.R. Kimball, A. Kimchi, A.C. Kimmelman, T. Kimura, M.A. King, K.J. Kinghorn, C.G. Kinsey, V. Kirkin, L.A. Kirshenbaum, S.L. Kiselev, S. Kishi, K. Kitamoto, Y. Kitaoka, K. Kitazato, R.N. Kitsis, J.T. Kittler, O. Kjaerulff, P.S. Klein, T. Klopstock, J. Klucken, H. Knævelsrud, R.L. Knorr, B.C.B. Ko, F. Ko, J.-L. Ko, H. Kobayashi, S. Kobayashi, I. Koch, J.C. Koch, U. Koenig, D. Kögel, Y.H. Koh, M. Koike, S.D. Kohlwein, N.M. Kocaturk, M. Komatsu, J. König, T. Kono, B.T. Kopp, T. Korcsmaros, G. Korkmaz, V.I. Korolchuk, M.S. Korsnes, A. Koskela, J. Kota, Y. Kotake, M.L. Kotler, Y. Kou, M.I. Koukourakis, E. Koustas, A.L. Kovacs, T. Kovács, D. Koya, T. Kozako, C. Kraft, D. Krainc, H. Krämer, A.D. Krasnodembskaya, C. Kretz-Remy, G. Kroemer, N.T. Ktistakis, K. Kuchitsu, S. Kuenen, L. Kuerschner, T. Kukar, A. Kumar, A. Kumar, D. Kumar, D. Kumar, S. Kumar, S. Kume, C. Kumsta, C.N. Kundu, M. Kundu, A.B. Kunnumakkara, L. Kurgan, T.G. Kutateladze, O. Kutlu, S. Kwak, H.J. Kwon, T.K. Kwon, Y.T. Kwon, I. Kyrmizi, A. La Spada, P. Labonté, S. Ladoire, I. Laface, F. Lafont, D.C. Lagace, V. Lahiri, Z. Lai, A.S. Laird, A. Lakkaraju, T. Lamark, S.-H. Lan, A. Landajuela, D.J.R. Lane, J.D. Lane, C.H. Lang, C. Lange, Ü. Langel, R. Langer, P. Lapaquette, J. Laporte, N.F. LaRusso, I. Lastres-Becker, W.C.Y. Lau, G.W. Laurie, S. Lavandero, B.Y.K. Law, H.K.-W. Law, R. Layfield, W. Le, H. Le Stunff, A.Y. Leary, J.-J. Lebrun, L.Y.W. Leck, J.-P. Leduc-Gaudet, C. Lee, C.-P. Lee, D.-H. Lee, E.B. Lee, E.F. Lee, G.M. Lee, H.-J. Lee, H.K. Lee, J.M. Lee, J.S. Lee, J.-A. Lee, J.-Y. Lee, J.H. Lee, M. Lee, M.G. Lee, M.J. Lee, M.-S. Lee, S.Y. Lee, S.-J. Lee, S.Y. Lee, S.B. Lee, W.H. Lee, Y.-R. Lee, Y.-H. Lee, Y. Lee, C. Lefebvre, R. Legouis, Y.L. Lei, Y. Lei, S. Leikin, G. Leitinger, L. Lemus, S. Leng, O. Lenoir, G. Lenz, H.J. Lenz, P. Lenzi, Y. León, A.M. Leopoldino, C. Leschczyk, S. Leskelä, E. Letellier, C.-T. Leung, P.S. Leung, J.S. Leventhal, B. Levine, P.A. Lewis, K. Ley, B. Li, D.-Q. Li, J. Li, J. Li, J. Li, K. Li, L. Li, M. Li, M. Li, M. Li, M. Li, M. Li, P.-L. Li, M.-Q. Li, Q. Li, S. Li, T. Li, W. Li, W. Li, X. Li, Y.-P. Li, Y. Li, Z. Li, Z. Li, Z. Li, J. Lian, C. Liang, Q. Liang, W. Liang, Y. Liang, Y. Liang, G. Liao, L. Liao, M. Liao, Y.-F. Liao, M. Librizzi, P.P.Y. Lie, M.A. Lilly, H.J. Lim, T.R.R. Lima, F. Limana, C. Lin, C.-W. Lin, D.-S. Lin, F.-C. Lin, J.D. Lin, K.M. Lin, K.-H. Lin, L.-T. Lin, P.-H. Lin, Q. Lin, S. Lin, S.-J. Lin, W. Lin, X. Lin, Y.-X. Lin, Y.-S. Lin, R. Linden, P. Lindner, S.-C. Ling, P. Lingor, A.K. Linnemann, Y.-C. Liou, M.M. Lipinski, S. Lipovšek, V.A. Lira, N. Lisiak, P.B. Liton, C. Liu, C.-H. Liu, C.-F. Liu, C.H. Liu, F. Liu, H. Liu, H.-S. Liu, H.-F. Liu, H. Liu, J. Liu, J. Liu, J. Liu, L. Liu, L. Liu, M. Liu, Q. Liu, W. Liu, W. Liu, X.-H. Liu, X. Liu, X. Liu, X. Liu, X. Liu, Y. Liu, Y. Liu, Y. Liu, Y. Liu, Y. Liu, J.A. Livingston, G. Lizard, J.M. Lizcano, S. Ljubojevic-Holzer, M.E. LLeonart, D. Llobet-Navàs, A. Llorente, C.H. Lo, D. Lobato-Márquez, Q. Long, Y.C. Long, B. Loos, J.A. Loos, M.G. López, G. López-Doménech, J.A. López-Guerrero, A.T. López-Jiménez, Ó. López-Pérez, I. López-Valero, M.J. Lorenowicz, M. Lorente, P. Lorincz, L. Lossi, S. Lotersztajn, P.E. Lovat, J.F. Lovell, A. Lovy, P. Lőw, G. Lu, H. Lu, J.-H. Lu, J.-J. Lu, M. Lu, S. Lu, A. Luciani, J.M. Lucocq, P. Ludovico, M.A. Luftig, M. Luhr, D. Luis-Ravelo, J.J. Lum, L. Luna-Dulcey, A.H. Lund, V.K. Lund, J.D. Lünemann, P. Lüningschrör, H. Luo, R. Luo, S. Luo, Z. Luo, C. Luparello, B. Lüscher, L. Luu, A. Lyakhovich, K.G. Lyamzaev, A.H. Lystad, L. Lytvynchuk, A.C. Ma, C. Ma, M. Ma, N.-F. Ma, Q.-H. Ma, X. Ma, Y. Ma, Z. Ma, O.A. MacDougald, F. Macian, G.C. MacIntosh, J.P. MacKeigan, K.F. Macleod, S. Maday, F. Madeo, M. Madesh, T. Madl, J. Madrigal-Matute, A. Maeda, Y. Maejima, M. Magarinos, P. Mahavadi, E. Maiani, K. Maiese, P. Maiti, M.C. Maiuri, B. Majello, M.B. Major, E. Makareeva, F. Malik, K. Mallilankaraman, W. Malorni, A. Maloyan, N. Mammadova, G.C.W. Man, F. Manai, J.D. Mancias, E.-M. Mandelkow, M.A. Mandell, A.A. Manfredi, M.H. Manjili, R. Manjithaya, P. Manque, B.B. Manshian, R. Manzano, C. Manzoni, K. Mao, C. Marchese, S. Marchetti, A.M. Marconi, F. Marcucci, S. Mardente, O.A. Mareninova, M. Margeta, M. Mari, S. Marinelli, O. Marinelli, G. Mariño, S. Mariotto, R.S. Marshall, M.R. Marten, S. Martens, A.P.J. Martin, K.R. Martin, S. Martin, S. Martin, A. Martín-Segura, M.A. Martín-Acebes, I. Martin-Burriel, M. Martin-Rincon, P. Martin-Sanz, J.A. Martina, W. Martinet, A. Martinez, A. Martinez, J. Martinez, M. Martinez Velazquez, N. Martinez-Lopez, M. Martinez-Vicente, D.O. Martins, J.O. Martins, W.K. Martins, T. Martins-Marques, E. Marzetti, S. Masaldan, C. Masclaux-Daubresse, D.G. Mashek, V. Massa, L. Massieu, G.R. Masson, L. Masuelli, A.I. Masyuk, T.V. Masyuk, P. Matarrese, A. Matheu, S. Matoba, S. Matsuzaki, P. Mattar, A. Matte, D. Mattoscio, J.L. Mauriz, M. Mauthe, C. Mauvezin, E. Maverakis, P. Maycotte, J. Mayer, G. Mazzoccoli, C. Mazzoni, J.R. Mazzulli, N. McCarty, C. McDonald, M.R. McGill, S.L. McKenna, B. McLaughlin, F. McLoughlin, M.A. McNiven, T.G. McWilliams, F. Mechta-Grigoriou, T.C. Medeiros, D.L. Medina, L.A. Megeney, K. Megyeri, M. Mehrpour, J.L. Mehta, A.J. Meijer, A.H. Meijer, J. Mejlvang, A. Meléndez, A. Melk, G. Memisoglu, A.F. Mendes, D. Meng, F. Meng, T. Meng, R. Menna-Barreto, M.B. Menon, C. Mercer, A.E. Mercier, J.-L. Mergny, A. Merighi, S.D. Merkley, G. Merla, V. Meske, A.C. Mestre, S.P. Metur, C. Meyer, H. Meyer, W. Mi, J. Mialet-Perez, J. Miao, L. Micale, Y. Miki, E. Milan, M. Milczarek, D.L. Miller, S.I. Miller, S. Miller, S.W. Millward, I. Milosevic, E.A. Minina, H. Mirzaei, H.R. Mirzaei, M. Mirzaei, A. Mishra, N. Mishra, P.K. Mishra, M. Misirkic Marjanovic, R. Misasi, A. Misra, G. Misso, C. Mitchell, G. Mitou, T. Miura, S. Miyamoto, M. Miyazaki, M. Miyazaki, T. Miyazaki, K. Miyazawa, N. Mizushima, T.H. Mogensen, B. Mograbi, R. Mohammadinejad, Y. Mohamud, A. Mohanty, S. Mohapatra, T. Möhlmann, A. Mohmmed, A. Moles, K.H. Moley, M. Molinari, V. Mollace, A.B. Møller, B. Mollereau, F. Mollinedo, C. Montagna, M.J. Monteiro, A. Montella, L.R. Montes, B. Montico, V.K. Mony, G. Monzio Compagnoni, M.N. Moore, M.A. Moosavi, A.L. Mora, M. Mora, D. Morales-Alamo, R. Moratalla, P.I. Moreira, E. Morelli, S. Moreno, D. Moreno-Blas, V. Moresi, B. Morga, A.H. Morgan, F. Morin, H. Morishita, O.L. Moritz, M. Moriyama, Y. Moriyasu, M. Morleo, E. Morselli, J.F. Moruno-Manchon, J. Moscat, S. Mostowy, E. Motori, A.F. Moura, N. Moustaid-Moussa, M. Mrakovcic, G. Muciño-Hernández, A. Mukherjee, S. Mukhopadhyay, J.M. Mulcahy Levy, V. Mulero, S. Muller, C. Münch, A. Munjal, P. Munoz-Canoves, T. Muñoz-Galdeano, C. Münz, T. Murakawa, C. Muratori, B.M. Murphy, J.P. Murphy, A. Murthy, T.T. Myöhänen, I.U. Mysorekar, J. Mytych, S.M. Nabavi, M. Nabissi, P. Nagy, J. Nah, A. Nahimana, I. Nakagawa, K. Nakamura, H. Nakatogawa, S.S. Nandi, M. Nanjundan, M. Nanni, G. Napolitano, R. Nardacci, M. Narita, M. Nassif, I. Nathan, M. Natsumeda, R.J. Naude, C. Naumann, O. Naveiras, F. Navid, S.T. Nawrocki, T.Y. Nazarko, F. Nazio, F. Negoita, T. Neill, A.L. Neisch, L.M. Neri, M.G. Netea, P. Neubert, T.P. Neufeld, D. Neumann, A. Neutzner, P.T. Newton, P.A. Ney, I.P. Nezis, C.C.W. Ng, T.B. Ng, H.T.T. Nguyen, L.T. Nguyen, H.-M. Ni, C. Ní Cheallaigh, Z. Ni, M.C. Nicolao, F. Nicoli, M. Nieto-Diaz, P. Nilsson, S. Ning, R. Niranjan, H. Nishimune, M. Niso-Santano, R.A. Nixon, A. Nobili, C. Nobrega, T. Noda, U. Nogueira-Recalde, T.M. Nolan, I. Nombela, I. Novak, B. Novoa, T. Nozawa, N. Nukina, C. Nussbaum-Krammer, J. Nylandsted, T.R. O’Donovan, S.M. O’Leary, E.J. O’Rourke, M.P. O’Sullivan, T.E. O’Sullivan, S. Oddo, I. Oehme, M. Ogawa, E. Ogier-Denis, M.H. Ogmundsdottir, B. Ogretmen, G.T. Oh, S.-H. Oh, Y.J. Oh, T. Ohama, Y. Ohashi, M. Ohmuraya, V. Oikonomou, R. Ojha, K. Okamoto, H. Okazawa, M. Oku, S. Oliván, J.M.A. Oliveira, M. Ollmann, J.A. Olzmann, S. Omari, M.B. Omary, G. Önal, M. Ondrej, S.-B. Ong, S.-G. Ong, A. Onnis, J.A. Orellana, S. Orellana-Muñoz, M.D.M. Ortega-Villaizan, X.R. Ortiz-Gonzalez, E. Ortona, H.D. Osiewacz, A.-H.K. Osman, R. Osta, M.S. Otegui, K. Otsu, C. Ott, L. Ottobrini, J.-H.J. Ou, T.F. Outeiro, I. Oynebraten, M. Ozturk, G. Pagès, S. Pahari, M. Pajares, U.B. Pajvani, R. Pal, S. Paladino, N. Pallet, M. Palmieri, G. Palmisano, C. Palumbo, F. Pampaloni, L. Pan, Q. Pan, W. Pan, X. Pan, G. Panasyuk, R. Pandey, U.B. Pandey, V. Pandya, F. Paneni, S.Y. Pang, E. Panzarini, D.L. Papademetrio, E. Papaleo, D. Papinski, D. Papp, E.C. Park, H.T. Park, J.-M. Park, J.-I. Park, J.T. Park, J. Park, S.C. Park, S.-Y. Park, A.H. Parola, J.B. Parys, A. Pasquier, B. Pasquier, J.F. Passos, N. Pastore, H.H. Patel, D. Patschan, S. Pattingre, G. Pedraza-Alva, J. Pedraza-Chaverri, Z. Pedrozo, G. Pei, J. Pei, H. Peled-Zehavi, J.M. Pellegrini, J. Pelletier, M.A. Peñalva, D. Peng, Y. Peng, F. Penna, M. Pennuto, F. Pentimalli, C.M. Pereira, G.J.S. Pereira, L.C. Pereira, L. Pereira de Almeida, N.D. Perera, Á. Pérez-Lara, A.B. Perez-Oliva, M.E. Pérez-Pérez, P. Periyasamy, A. Perl, C. Perrotta, I. Perrotta, R.G. Pestell, M. Petersen, I. Petrache, G. Petrovski, T. Pfirrmann, A.S. Pfister, J.A. Philips, H. Pi, A. Picca, A.M. Pickrell, S. Picot, G.M. Pierantoni, M. Pierdominici, P. Pierre, V. Pierrefite-Carle, K. Pierzynowska, F. Pietrocola, M. Pietruczuk, C. Pignata, F.X. Pimentel-Muiños, M. Pinar, R.O. Pinheiro, R. Pinkas-Kramarski, P. Pinton, K. Pircs, S. Piya, P. Pizzo, T.S. Plantinga, H.W. Platta, A. Plaza-Zabala, M. Plomann, E.Y. Plotnikov, H. Plun-Favreau, R. Pluta, R. Pocock, S. Pöggeler, C. Pohl, M. Poirot, A. Poletti, M. Ponpuak, H. Popelka, B. Popova, H. Porta, S. Porte Alcon, E. Portilla-Fernandez, M. Post, M.B. Potts, J. Poulton, T. Powers, V. Prahlad, T.K. Prajsnar, D. Praticò, R. Prencipe, M. Priault, T. Proikas-Cezanne, V.J. Promponas, C.G. Proud, R. Puertollano, L. Puglielli, T. Pulinilkunnil, D. Puri, R. Puri, J. Puyal, X. Qi, Y. Qi, W. Qian, L. Qiang, Y. Qiu, J. Quadrilatero, J. Quarleri, N. Raben, H. Rabinowich, D. Ragona, M.J. Ragusa, N. Rahimi, M. Rahmati, V. Raia, N. Raimundo, N.-S. Rajasekaran, S. Ramachandra Rao, A. Rami, I. Ramírez-Pardo, D.B. Ramsden, F. Randow, P.N. Rangarajan, D. Ranieri, H. Rao, L. Rao, R. Rao, S. Rathore, J.A. Ratnayaka, E.A. Ratovitski, P. Ravanan, G. Ravegnini, S.K. Ray, B. Razani, V. Rebecca, F. Reggiori, A. Régnier-Vigouroux, A.S. Reichert, D. Reigada, J.H. Reiling, T. Rein, S. Reipert, R.S. Rekha, H. Ren, J. Ren, W. Ren, T. Renault, G. Renga, K. Reue, K. Rewitz, B. Ribeiro de Andrade Ramos, S.A. Riazuddin, T.M. Ribeiro-Rodrigues, J.-E. Ricci, R. Ricci, V. Riccio, D.R. Richardson, Y. Rikihisa, M.V. Risbud, R.M. Risueño, K. Ritis, S. Rizza, R. Rizzuto, H.C. Roberts, L.D. Roberts, K.J. Robinson, M.C. Roccheri, S. Rocchi, G.G. Rodney, T. Rodrigues, V.R. Rodrigues Silva, A. Rodriguez, R. Rodriguez-Barrueco, N. Rodriguez-Henche, H. Rodriguez-Rocha, J. Roelofs, R.S. Rogers, V.V. Rogov, A.I. Rojo, K. Rolka, V. Romanello, L. Romani, A. Romano, P.S. Romano, D. Romeo-Guitart, L.C. Romero, M. Romero, J.C. Roney, C. Rongo, S. Roperto, M.T. Rosenfeldt, P. Rosenstiel, A.G. Rosenwald, K.A. Roth, L. Roth, S. Roth, K.M.A. Rouschop, B.D. Roussel, S. Roux, P. Rovere-Querini, A. Roy, A. Rozieres, D. Ruano, D.C. Rubinsztein, M.P. Rubtsova, K. Ruckdeschel, C. Ruckenstuhl, E. Rudolf, R. Rudolf, A. Ruggieri, A.A. Ruparelia, P. Rusmini, R.R. Russell, G.L. Russo, M. Russo, R. Russo, O.O. Ryabaya, K.M. Ryan, K.-Y. Ryu, M. Sabater-Arcis, U. Sachdev, M. Sacher, C. Sachse, A. Sadhu, J. Sadoshima, N. Safren, P. Saftig, A.P. Sagona, G. Sahay, A. Sahebkar, M. Sahin, O. Sahin, S. Sahni, N. Saito, S. Saito, T. Saito, R. Sakai, Y. Sakai, J.-I. Sakamaki, K. Saksela, G. Salazar, A. Salazar-Degracia, G.H. Salekdeh, A.K. Saluja, B. Sampaio-Marques, M.C. Sanchez, J.A. Sanchez-Alcazar, V. Sanchez-Vera, V. Sancho-Shimizu, J.T. Sanderson, M. Sandri, S. Santaguida, L. Santambrogio, M.M. Santana, G. Santoni, A. Sanz, P. Sanz, S. Saran, M. Sardiello, T.J. Sargeant, A. Sarin, C. Sarkar, S. Sarkar, M.-R. Sarrias, S. Sarkar, D.T. Sarmah, J. Sarparanta, A. Sathyanarayan, R. Sathyanarayanan, K.M. Scaglione, F. Scatozza, L. Schaefer, Z.T. Schafer, U.E. Schaible, A.H.V. Schapira, M. Scharl, H.M. Schatzl, C.H. Schein, W. Scheper, D. Scheuring, M.V. Schiaffino, M. Schiappacassi, R. Schindl, U. Schlattner, O. Schmidt, R. Schmitt, S.D. Schmidt, I. Schmitz, E. Schmukler, A. Schneider, B.E. Schneider, R. Schober, A.C. Schoijet, M.B. Schott, M. Schramm, B. Schröder, K. Schuh, C. Schüller, R.J. Schulze, L. Schürmanns, J.C. Schwamborn, M. Schwarten, F. Scialo, S. Sciarretta, M.J. Scott, K.W. Scotto, A.I. Scovassi, A. Scrima, A. Scrivo, D. Sebastian, S. Sebti, S. Sedej, L. Segatori, N. Segev, P.O. Seglen, I. Seiliez, E. Seki, S.B. Selleck, F.W. Sellke, J.T. Selsby, M. Sendtner, S. Senturk, E. Seranova, C. Sergi, R. Serra-Moreno, H. Sesaki, C. Settembre, S.R.G. Setty, G. Sgarbi, O. Sha, J.J. Shacka, J.A. Shah, D. Shang, C. Shao, F. Shao, S. Sharbati, L.M. Sharkey, D. Sharma, G. Sharma, K. Sharma, P. Sharma, S. Sharma, H.-M. Shen, H. Shen, J. Shen, M. Shen, W. Shen, Z. Shen, R. Sheng, Z. Sheng, Z.-H. Sheng, J. Shi, X. Shi, Y.-H. Shi, K. Shiba-Fukushima, J.-J. Shieh, Y. Shimada, S. Shimizu, M. Shimozawa, T. Shintani, C.J. Shoemaker, S. Shojaei, I. Shoji, B.V. Shravage, V. Shridhar, C.-W. Shu, H.-B. Shu, K. Shui, A.K. Shukla, T.E. Shutt, V. Sica, A. Siddiqui, A. Sierra, V. Sierra-Torre, S. Signorelli, P. Sil, B.J. de A. Silva, J.D. Silva, E. Silva-Pavez, S. Silvente-Poirot, R.E. Simmonds, A.K. Simon, H.-U. Simon, M. Simons, A. Singh, L.P. Singh, R. Singh, S.V. Singh, S.K. Singh, S.B. Singh, S. Singh, S.P. Singh, D. Sinha, R.A. Sinha, S. Sinha, A. Sirko, K. Sirohi, E.L. Sivridis, P. Skendros, A. Skirycz, I. Slaninová, S.S. Smaili, A. Smertenko, M.D. Smith, S.J. Soenen, E.J. Sohn, S.P.M. Sok, G. Solaini, T. Soldati, S.A. Soleimanpour, R.M. Soler, A. Solovchenko, J.A. Somarelli, A. Sonawane, F. Song, H.K. Song, J.-X. Song, K. Song, Z. Song, L.R. Soria, M. Sorice, A.A. Soukas, S.-F. Soukup, D. Sousa, N. Sousa, P.A. Spagnuolo, S.A. Spector, M.M. Srinivas Bharath, D. St Clair, V. Stagni, L. Staiano, C.A. Stalnecker, M.V. Stankov, P.B. Stathopulos, K. Stefan, S.M. Stefan, L. Stefanis, J.S. Steffan, A. Steinkasserer, H. Stenmark, J. Sterneckert, C. Stevens, V. Stoka, S. Storch, B. Stork, F. Strappazzon, A.M. Strohecker, D.G. Stupack, H. Su, L.-Y. Su, L. Su, A.M. Suarez-Fontes, C.S. Subauste, S. Subbian, P.V. Subirada, G. Sudhandiran, C.M. Sue, X. Sui, C. Summers, G. Sun, J. Sun, K. Sun, M.-X. Sun, Q. Sun, Y. Sun, Z. Sun, K.K.S. Sunahara, E. Sundberg, K. Susztak, P. Sutovsky, H. Suzuki, G. Sweeney, J.D. Symons, S.C.W. Sze, N.J. Szewczyk, A. Tabęcka-Łonczynska, C. Tabolacci, F. Tacke, H. Taegtmeyer, M. Tafani, M. Tagaya, H. Tai, S.W.G. Tait, Y. Takahashi, S. Takats, P. Talwar, C. Tam, S.Y. Tam, D. Tampellini, A. Tamura, C.T. Tan, E.-K. Tan, Y.-Q. Tan, M. Tanaka, M. Tanaka, D. Tang, J. Tang, T.-S. Tang, I. Tanida, Z. Tao, M. Taouis, L. Tatenhorst, N. Tavernarakis, A. Taylor, G.A. Taylor, J.M. Taylor, E. Tchetina, A.R. Tee, I. Tegeder, D. Teis, N. Teixeira, F. Teixeira-Clerc, K.A. Tekirdag, T. Tencomnao, S. Tenreiro, A.V. Tepikin, P.S. Testillano, G. Tettamanti, P.-L. Tharaux, K. Thedieck, A.A. Thekkinghat, S. Thellung, J.W. Thinwa, V.P. Thirumalaikumar, S.M. Thomas, P.G. Thomes, A. Thorburn, L. Thukral, T. Thum, M. Thumm, L. Tian, A. Tichy, A. Till, V. Timmerman, V.I. Titorenko, S.V. Todi, K. Todorova, J.M. Toivonen, L. Tomaipitinca, D. Tomar, C. Tomas-Zapico, S. Tomić, B.C.-K. Tong, C. Tong, X. Tong, S.A. Tooze, M.L. Torgersen, S. Torii, L. Torres-López, A. Torriglia, C.G. Towers, R. Towns, S. Toyokuni, V. Trajkovic, D. Tramontano, Q.-G. Tran, L.H. Travassos, C.B. Trelford, S. Tremel, I.P. Trougakos, B.P. Tsao, M.P. Tschan, H.-F. Tse, T.F. Tse, H. Tsugawa, A.S. Tsvetkov, D.A. Tumbarello, Y. Tumtas, M.J. Tuñón, S. Turcotte, B. Turk, V. Turk, B.J. Turner, R.I. Tuxworth, J.K. Tyler, E.V. Tyutereva, Y. Uchiyama, A. Ugun-Klusek, H.H. Uhlig, M. Ułamek-Kozioł, I.V. Ulasov, M. Umekawa, C. Ungermann, R. Unno, S. Urbe, E. Uribe-Carretero, S. Üstün, V.N. Uversky, T. Vaccari, M.I. Vaccaro, B.F. Vahsen, H. Vakifahmetoglu-Norberg, R. Valdor, M.J. Valente, A. Valko, R.B. Vallee, A.M. Valverde, G. Van den Berghe, S. van der Veen, L. Van Kaer, J. van Loosdregt, S.J.L. van Wijk, W. Vandenberghe, I. Vanhorebeek, M.A. Vannier-Santos, N. Vannini, M.C. Vanrell, C. Vantaggiato, G. Varano, I. Varela-Nieto, M. Varga, M.H. Vasconcelos, S. Vats, D.G. Vavvas, I. Vega-Naredo, S. Vega-Rubin-de-Celis, G. Velasco, A.P. Velázquez, T. Vellai, E. Vellenga, F. Velotti, M. Verdier, P. Verginis, I. Vergne, P. Verkade, M. Verma, P. Verstreken, T. Vervliet, J. Vervoorts, A.T. Vessoni, V.M. Victor, M. Vidal, C. Vidoni, O.V. Vieira, R.D. Vierstra, S. Viganó, H. Vihinen, V. Vijayan, M. Vila, M. Vilar, J.M. Villalba, A. Villalobo, B. Villarejo-Zori, F. Villarroya, J. Villarroya, O. Vincent, C. Vindis, C. Viret, M.T. Viscomi, D. Visnjic, I. Vitale, D.J. Vocadlo, O.V. Voitsekhovskaja, C. Volonté, M. Volta, M. Vomero, C. Von Haefen, M.A. Vooijs, W. Voos, L. Vucicevic, R. Wade-Martins, S. Waguri, K.A. Waite, S. Wakatsuki, D.W. Walker, M.J. Walker, S.A. Walker, J. Walter, F.G. Wandosell, B. Wang, C.-Y. Wang, C. Wang, C. Wang, C. Wang, C.-Y. Wang, D. Wang, F. Wang, F. Wang, F. Wang, G. Wang, H. Wang, H. Wang, H. Wang, H.-G. Wang, J. Wang, J. Wang, J. Wang, J. Wang, K. Wang, L. Wang, L. Wang, M.H. Wang, M. Wang, N. Wang, P. Wang, P. Wang, P. Wang, P. Wang, Q.J. Wang, Q. Wang, Q.K. Wang, Q.A. Wang, W.-T. Wang, W. Wang, X. Wang, X. Wang, Y. Wang, Y. Wang, Y. Wang, Y.-Y. Wang, Y. Wang, Y. Wang, Y. Wang, Y. Wang, Z. Wang, Z. Wang, Z. Wang, G. Warnes, V. Warnsmann, H. Watada, E. Watanabe, M. Watchon, A. Wawrzyńska, T.E. Weaver, G. Wegrzyn, A.M. Wehman, H. Wei, L. Wei, T. Wei, Y. Wei, O.H. Weiergräber, C.C. Weihl, G. Weindl, R. Weiskirchen, A. Wells, R.H. Wen, X. Wen, A. Werner, B. Weykopf, S.P. Wheatley, J.L. Whitton, A.J. Whitworth, K. Wiktorska, M.E. Wildenberg, T. Wileman, S. Wilkinson, D. Willbold, B. Williams, R.S.B. Williams, R.L. Williams, P.R. Williamson, R.A. Wilson, B. Winner, N.J. Winsor, S.S. Witkin, H. Wodrich, U. Woehlbier, T. Wollert, E. Wong, J.H. Wong, R.W. Wong, V.K.W. Wong, W.W.-L. Wong, A.-G. Wu, C. Wu, J. Wu, J. Wu, K.K. Wu, M. Wu, S.-Y. Wu, S. Wu, S.-Y. Wu, S. Wu, W.K.K. Wu, X. Wu, X. Wu, Y.-W. Wu, Y. Wu, R.J. Xavier, H. Xia, L. Xia, Z. Xia, G. Xiang, J. Xiang, M. Xiang, W. Xiang, B. Xiao, G. Xiao, H. Xiao, H.-T. Xiao, J. Xiao, L. Xiao, S. Xiao, Y. Xiao, B. Xie, C.-M. Xie, M. Xie, Y. Xie, Z. Xie, Z. Xie, M. Xilouri, C. Xu, E. Xu, H. Xu, J. Xu, J. Xu, L. Xu, W.W. Xu, X. Xu, Y. Xue, S.M.S. Yakhine-Diop, M. Yamaguchi, O. Yamaguchi, A. Yamamoto, S. Yamashina, S. Yan, S.-J. Yan, Z. Yan, Y. Yanagi, C. Yang, D.-S. Yang, H. Yang, H.-T. Yang, H. Yang, J.-M. Yang, J. Yang, J. Yang, L. Yang, L. Yang, M. Yang, P.-M. Yang, Q. Yang, S. Yang, S. Yang, S.-F. Yang, W. Yang, W.Y. Yang, X. Yang, X. Yang, Y. Yang, Y. Yang, H. Yao, S. Yao, X. Yao, Y.-G. Yao, Y.-M. Yao, T. Yasui, M. Yazdankhah, P.M. Yen, C. Yi, X.-M. Yin, Y. Yin, Z. Yin, Z. Yin, M. Ying, Z. Ying, C.K. Yip, S.P.T. Yiu, Y.H. Yoo, K. Yoshida, S.R. Yoshii, T. Yoshimori, B. Yousefi, B. Yu, H. Yu, J. Yu, J. Yu, L. Yu, M.-L. Yu, S.-W. Yu, V.C. Yu, W.H. Yu, Z. Yu, Z. Yu, J. Yuan, L.-Q. Yuan, S. Yuan, S.-S.F. Yuan, Y. Yuan, Z. Yuan, J. Yue, Z. Yue, J. Yun, R.L. Yung, D.N. Zacks, G. Zaffagnini, V.O. Zambelli, I. Zanella, Q.S. Zang, S. Zanivan, S. Zappavigna, P. Zaragoza, K.S. Zarbalis, A. Zarebkohan, A. Zarrouk, S.O. Zeitlin, J. Zeng, J.-D. Zeng, E. Žerovnik, L. Zhan, B. Zhang, D.D. Zhang, H. Zhang, H. Zhang, H. Zhang, H. Zhang, H. Zhang, H. Zhang, H. Zhang, H.-L. Zhang, J. Zhang, J. Zhang, J.-P. Zhang, K.Y.B. Zhang, L.W. Zhang, L. Zhang, L. Zhang, L. Zhang, L. Zhang, M. Zhang, P. Zhang, S. Zhang, W. Zhang, X. Zhang, X.-W. Zhang, X. Zhang, X. Zhang, X. Zhang, X. Zhang, X.D. Zhang, Y. Zhang, Y. Zhang, Y. Zhang, Y.-D. Zhang, Y. Zhang, Y.-Y. Zhang, Y. Zhang, Z. Zhang, Z. Zhang, Z. Zhang, Z. Zhang, Z. Zhang, Z. Zhang, H. Zhao, L. Zhao, S. Zhao, T. Zhao, X.-F. Zhao, Y. Zhao, Y. Zhao, Y. Zhao, Y. Zhao, G. Zheng, K. Zheng, L. Zheng, S. Zheng, X.-L. Zheng, Y. Zheng, Z.-G. Zheng, B. Zhivotovsky, Q. Zhong, A. Zhou, B. Zhou, C. Zhou, G. Zhou, H. Zhou, H. Zhou, H. Zhou, J. Zhou, J. Zhou, J. Zhou, J. Zhou, K. Zhou, R. Zhou, X.-J. Zhou, Y. Zhou, Y. Zhou, Y. Zhou, Z.-Y. Zhou, Z. Zhou, B. Zhu, C. Zhu, G.-Q. Zhu, H. Zhu, H. Zhu, H. Zhu, W.-G. Zhu, Y. Zhu, Y. Zhu, H. Zhuang, X. Zhuang, K. Zientara-Rytter, C.M. Zimmermann, E. Ziviani, T. Zoladek, W.-X. Zong, D.B. Zorov, A. Zorzano, W. Zou, Z. Zou, Z. Zou, S. Zuryn, W. Zwerschke, B. Brand-Saberi, X.C. Dong, C.S. Kenchappa, Z. Li, Y. Lin, S. Oshima, Y. Rong, J.C. Sluimer, C.L. Stallings, C.-K. Tong, Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition)1, Autophagy. 17 (2021) 1–382. [CrossRef]
- R. Iwama, Y. Ohsumi, Analysis of autophagy activated during changes in carbon source availability in yeast cells, J. Biol. Chem. 294 (2019) 5590–5603. [CrossRef]
- M. Tsukada, Y. Ohsumi, Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae, FEBS Lett. 333 (1993) 169–174. [CrossRef]
- J. Onodera, Y. Ohsumi, Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation, J. Biol. Chem. 280 (2005) 31582–31586. [CrossRef]
- A.E. Mayes, L. Verdone, P. Legrain, J.D. Beggs, Characterization of Sm-like proteins in yeast and their association with U6 snRNA, EMBO J. 18 (1999) 4321–4331. [CrossRef]
- Y. Lei, Y. Huang, X. Wen, Z. Yin, Z. Zhang, D.J. Klionsky, How Cells Deal with the Fluctuating Environment: Autophagy Regulation under Stress in Yeast and Mammalian Systems, Antioxidants. 11 (2022) 304. [CrossRef]
- A.K. Singh, S. Singh, V.K. Tripathi, A. Bissoyi, G. Garg, S.I. Rizvi, Rapamycin Confers Neuroprotection Against Aging-Induced Oxidative Stress, Mitochondrial Dysfunction, and Neurodegeneration in Old Rats Through Activation of Autophagy, Rejuvenation Res. 22 (2019) 60–70. [CrossRef]
- Mazzoni, C. Falcone, Isolation and study ofKlLSM4 , aKluyveromyces lactis gene homologous to the essential geneLSM4 ofSaccharomyces cerevisiae, Yeast. 18 (2001) 1249–1256. [CrossRef]
- P. Kumar, D. Kundu, A.K. Mondal, V. Nain, R. Puria, Inhibition of TOR signalling in lea1 mutant induces apoptosis in Saccharomyces cerevisiae, Ann. Microbiol. 69 (2019) 341–352. [CrossRef]
- Z. Zhang, Y. Zhang, W. Mo, The Autophagy Related Gene CHAF1B Is a Relevant Prognostic and Diagnostic Biomarker in Hepatocellular Carcinoma, Front. Oncol. 10 (2021). https://www.frontiersin.org/articles/10.3389/fonc.2020.626175 (accessed June 26, 2023).
- S.W. Suzuki, J. Onodera, Y. Ohsumi, Starvation induced cell death in autophagy-defective yeast mutants is caused by mitochondria dysfunction, PloS One. 6 (2011) e17412. [CrossRef]
- M.E. Pérez-Pérez, M. Zaffagnini, C.H. Marchand, J.L. Crespo, S.D. Lemaire, The yeast autophagy protease Atg4 is regulated by thioredoxin, Autophagy. 10 (2014) 1953–1964. [CrossRef]
- E. Hirata, Y. Ohya, K. Suzuki, Atg4 plays an important role in efficient expansion of autophagic isolation membranes by cleaving lipidated Atg8 in Saccharomyces cerevisiae, PLOS ONE. 12 (2017) e0181047. [CrossRef]
- J. Sánchez-Wandelmer, F. Kriegenburg, S. Rohringer, M. Schuschnig, R. Gómez-Sánchez, B. Zens, S. Abreu, R. Hardenberg, D. Hollenstein, J. Gao, C. Ungermann, S. Martens, C. Kraft, F. Reggiori, Atg4 proteolytic activity can be inhibited by Atg1 phosphorylation, Nat. Commun. 8 (2017) 295. [CrossRef]
- S. Barz, F. Kriegenburg, A. Henning, A. Bhattacharya, H. Mancilla, P. Sánchez-Martín, C. Kraft, Atg1 kinase regulates autophagosome-vacuole fusion by controlling SNARE bundling, EMBO Rep. 21 (2020) e51869. [CrossRef]
- 50. Y. Lee, B. Kim, H.-S. Jang, W.-K. Huh, Atg1-dependent phosphorylation of Vps34 is required for dynamic regulation of the phagophore assembly site and autophagy in Saccharomyces cerevisiae, Autophagy. (2023). [CrossRef]
- J.J. Bearss, S.K. Padi, N. Singh, M. Cardo-Vila, J.H. Song, G. Mouneimne, N. Fernandes, Y. Li, M.R. Harter, J.M. Gard, A.E. Cress, W. Peti, A.D. Nelson, J.R. Buchan, A.S. Kraft, K. Okumura, EDC3 phosphorylation regulates growth and invasion through controlling P-body formation and dynamics, EMBO Rep. 22 (2021) e50835. [CrossRef]
- J. Ptacek, G. Devgan, G. Michaud, H. Zhu, X. Zhu, J. Fasolo, H. Guo, G. Jona, A. Breitkreutz, R. Sopko, R.R. McCartney, M.C. Schmidt, N. Rachidi, S.-J. Lee, A.S. Mah, L. Meng, M.J.R. Stark, D.F. Stern, C. De Virgilio, M. Tyers, B. Andrews, M. Gerstein, B. Schweitzer, P.F. Predki, M. Snyder, Global analysis of protein phosphorylation in yeast, Nature. 438 (2005) 679–684. [CrossRef]
- D.-C. Chen, B.-C. Yang, T.-T. Kuo, One-step transformation of yeast in stationary phase, Curr. Genet. 21 (1992) 83–84. [CrossRef]
- T. Eisenberg, H. Knauer, A. Schauer, S. Büttner, C. Ruckenstuhl, D. Carmona-Gutierrez, J. Ring, S. Schroeder, C. Magnes, L. Antonacci, H. Fussi, L. Deszcz, R. Hartl, E. Schraml, A. Criollo, E. Megalou, D. Weiskopf, P. Laun, G. Heeren, M. Breitenbach, B. Grubeck-Loebenstein, E. Herker, B. Fahrenkrog, K.-U. Fröhlich, F. Sinner, N. Tavernarakis, N. Minois, G. Kroemer, F. Madeo, Induction of autophagy by spermidine promotes longevity, Nat. Cell Biol. 11 (2009) 1305–1314. [CrossRef]
- V. Palermo, C. Falcone, C. Mazzoni, Apoptosis and aging in mitochondrial morphology mutants of S. cerevisiae, Folia Microbiol. (Praha). 52 (2007) 479–483. [CrossRef]
- M. Cooper, L.H. Johnston, J.D. Beggs, Identification and characterization of Uss1p (Sdb23p): a novel U6 snRNA-associated protein with significant similarity to core proteins of small nuclear ribonucleoproteins., EMBO J. 14 (1995) 2066–2075.
- C.B. Brachmann, A. Davies, G.J. Cost, E. Caputo, J. Li, P. Hieter, J.D. Boeke, Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications, Yeast Chichester Engl. 14 (1998) 115–132. [CrossRef]
Figure 1.
Sclsm4Δ1 mutant recapitulates the same pleiotropic phenotypes as described for Kllsm4Δ1 mutant. (A) DAPI staining of the CML39-11A (wild type) and Sclsm4Δ1 mutant cells in both exponential and stationary phase. (B) percentage of fragmented nuclei over total cells for the same strains and conditions as in (A) from three independent experiments. (C) Dihydrorhodamine 123 (DHR123) staining of the CML39-11A (wild type) and Sclsm4Δ1 mutant cells in both exponential and stationary phase. (D) Percentage of ROS positive cells over total cells for the same strains and conditions as in (C) from three independent experiments. Data are represented as mean percentage of 700 cells per set ± standard deviation. ***p-value<0.001, ****p-value<0.0001 (E) Chronological life span of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 mutant or the full length LSM4 protein (ScLSM4) cells. Data are represented as the mean of three independent experiments ± standard deviation. (F) Cell viability of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 mutant or the full length LSM4 protein (ScLSM4) cells was measured after exposure to H2O2 at the indicated concentrations for 4 h. Data are represented as the mean of three independent experiments ± standard deviation. *p-value<0.05, **p-value<0.01 (G) 10-fold dilution were spotted on complete solid media containing 2% glycerol (YPY), YPD containing 60 mM acetic acid and 0,25% caffeine and plates were incubated at 28°C for 3 days. YPD was used as growth control.
Figure 1.
Sclsm4Δ1 mutant recapitulates the same pleiotropic phenotypes as described for Kllsm4Δ1 mutant. (A) DAPI staining of the CML39-11A (wild type) and Sclsm4Δ1 mutant cells in both exponential and stationary phase. (B) percentage of fragmented nuclei over total cells for the same strains and conditions as in (A) from three independent experiments. (C) Dihydrorhodamine 123 (DHR123) staining of the CML39-11A (wild type) and Sclsm4Δ1 mutant cells in both exponential and stationary phase. (D) Percentage of ROS positive cells over total cells for the same strains and conditions as in (C) from three independent experiments. Data are represented as mean percentage of 700 cells per set ± standard deviation. ***p-value<0.001, ****p-value<0.0001 (E) Chronological life span of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 mutant or the full length LSM4 protein (ScLSM4) cells. Data are represented as the mean of three independent experiments ± standard deviation. (F) Cell viability of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 mutant or the full length LSM4 protein (ScLSM4) cells was measured after exposure to H2O2 at the indicated concentrations for 4 h. Data are represented as the mean of three independent experiments ± standard deviation. *p-value<0.05, **p-value<0.01 (G) 10-fold dilution were spotted on complete solid media containing 2% glycerol (YPY), YPD containing 60 mM acetic acid and 0,25% caffeine and plates were incubated at 28°C for 3 days. YPD was used as growth control.
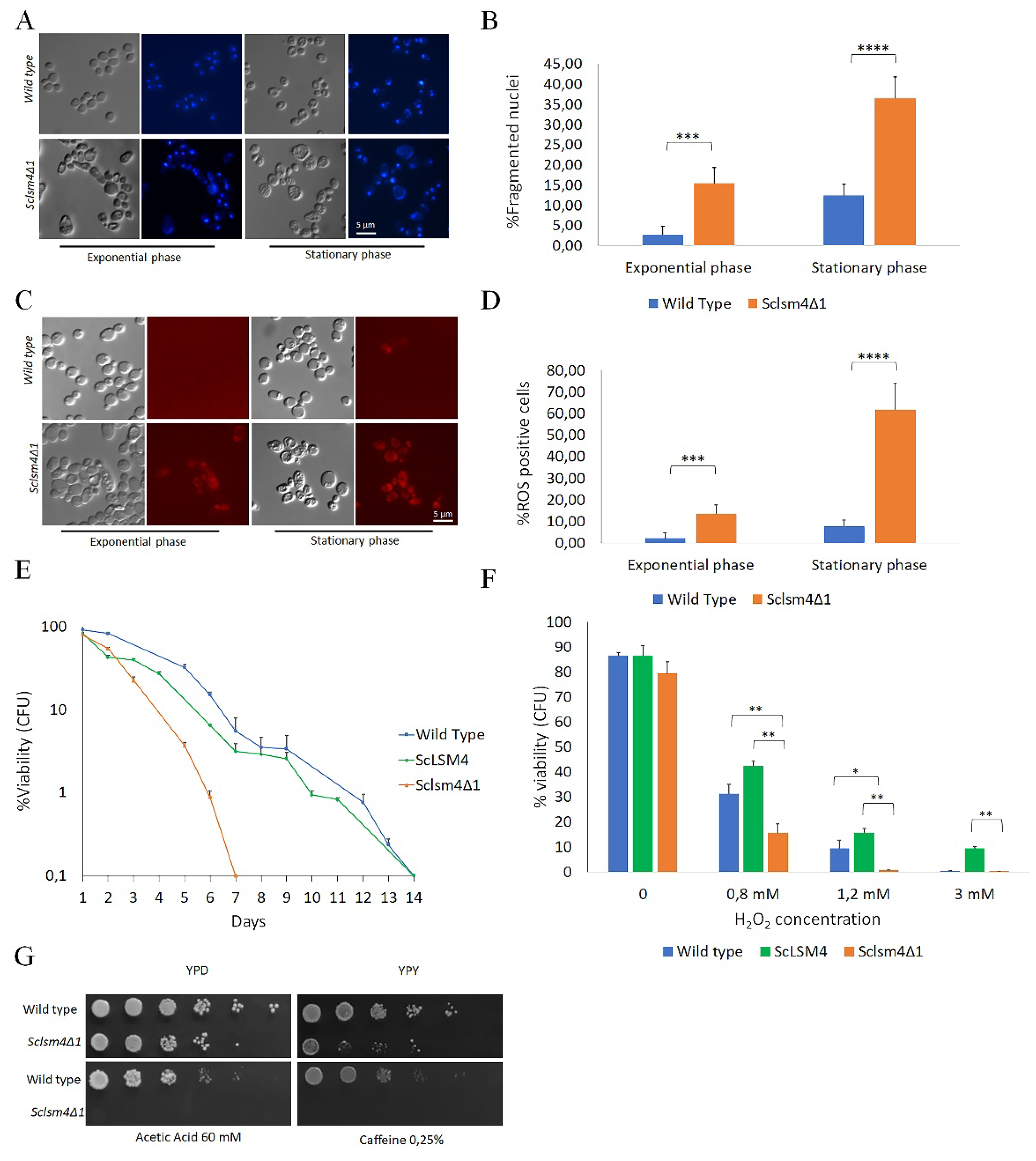
Figure 2.
Sclsm4Δ1 mutant shows defects in autophagy induction. (A) CML39-11A (WT) and mutant Sclsm4Δ1 cells were grown exponentially in SD medium (Exp), then the same amount of cells was centrifuged and resuspended in SD and in SD-N (nitrogen deprivation, -N) medium and further incubated for 16 hours (PD: Post-diauxic phase). Ponceau red staining has been used as a load control. One of three independent experiments is shown. (B) Percentage of autophagy activation was measured as the ratio between free GFP/GFP-ATG8 in three independent experiments. *p-value<0.05. (C) Chronological life span of CML39-11A (WT) and mutant Sclsm4Δ1 cells cultured in standard synthetic medium (SD) or in nitrogen deprived medium (SD-N). Data are represented as the average and standard deviation of three independent experiments and one representative experiment is shown.
Figure 2.
Sclsm4Δ1 mutant shows defects in autophagy induction. (A) CML39-11A (WT) and mutant Sclsm4Δ1 cells were grown exponentially in SD medium (Exp), then the same amount of cells was centrifuged and resuspended in SD and in SD-N (nitrogen deprivation, -N) medium and further incubated for 16 hours (PD: Post-diauxic phase). Ponceau red staining has been used as a load control. One of three independent experiments is shown. (B) Percentage of autophagy activation was measured as the ratio between free GFP/GFP-ATG8 in three independent experiments. *p-value<0.05. (C) Chronological life span of CML39-11A (WT) and mutant Sclsm4Δ1 cells cultured in standard synthetic medium (SD) or in nitrogen deprived medium (SD-N). Data are represented as the average and standard deviation of three independent experiments and one representative experiment is shown.
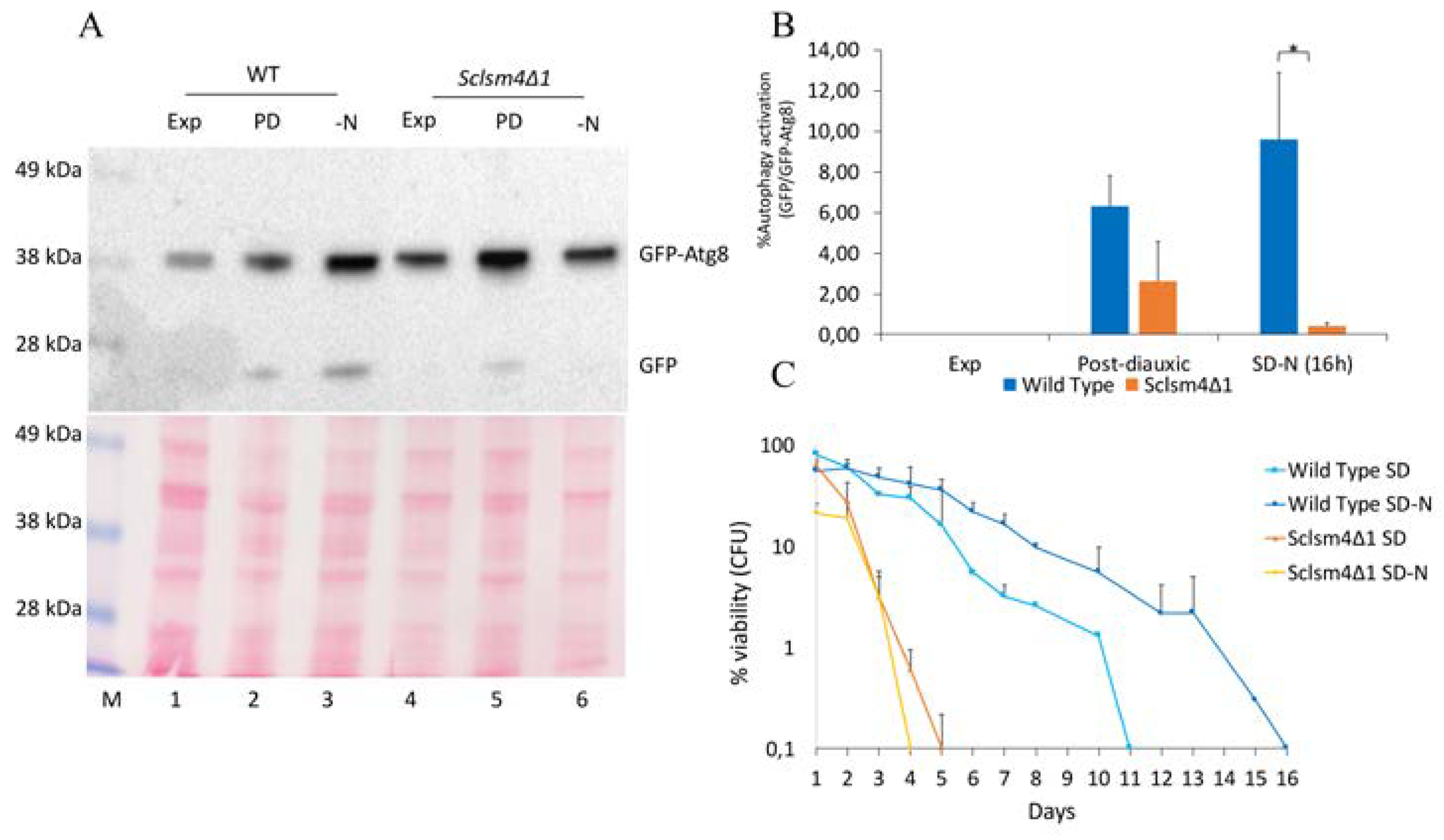
Figure 3.
Sclsm4Δ1 and lsm1 mutants are highly sensitive to rapamycin. (A) Cell viability of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 was measured every hour after treatment with 6 nM of rapamycin in SD medium for 4 hours. Data are represented as the mean of three independent experiments ± standard deviation. **p-value<0.01, ***p-value<0.001 (B) 10-fold dilution of MCY4 expressing Sclsm4Δ1 mutant cells, lsm1Δ mutant and their wild types (CML39-11A and BMA38) were spotted on YPD solid media containing 6 nM rapamycin and incubated at 28°C for 3 days. YPD was used as growth control.
Figure 3.
Sclsm4Δ1 and lsm1 mutants are highly sensitive to rapamycin. (A) Cell viability of the CML39-11A (wild type) and MCY4 expressing Sclsm4Δ1 was measured every hour after treatment with 6 nM of rapamycin in SD medium for 4 hours. Data are represented as the mean of three independent experiments ± standard deviation. **p-value<0.01, ***p-value<0.001 (B) 10-fold dilution of MCY4 expressing Sclsm4Δ1 mutant cells, lsm1Δ mutant and their wild types (CML39-11A and BMA38) were spotted on YPD solid media containing 6 nM rapamycin and incubated at 28°C for 3 days. YPD was used as growth control.
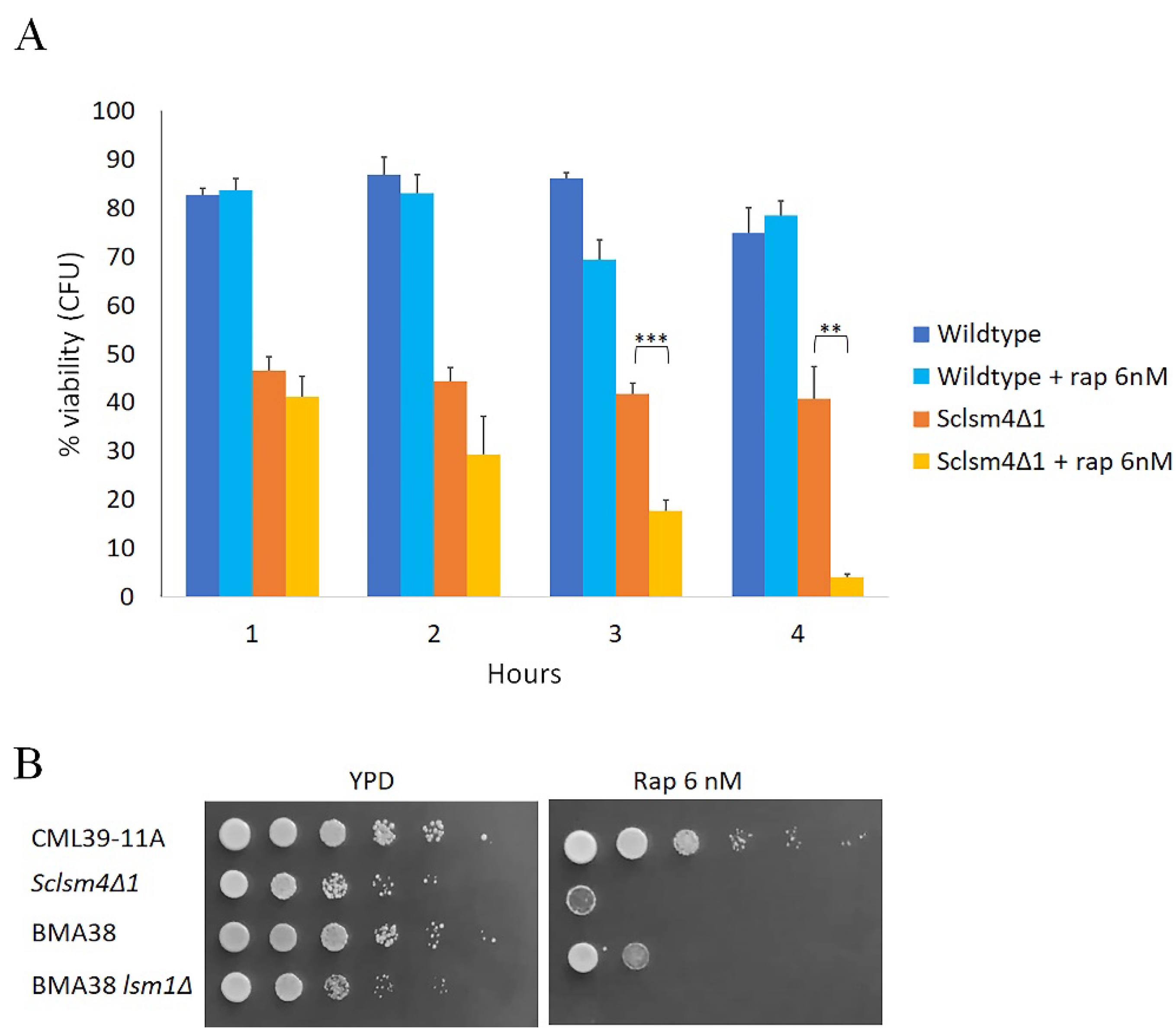
Figure 4.
Treatment with low dose of rapamycin does not protect the cells from oxidative stress. Cell viability of the CML39-11A (wild type) and Sclsm4Δ1 mutant was measured after exposure to H2O2 at the indicated concentrations for 4 h. 6nM rapamycin was added 4h prior exposure to H2O2. Data are represented as the mean of three independent experiments ± standard deviation. ****p-value<0.0001.
Figure 4.
Treatment with low dose of rapamycin does not protect the cells from oxidative stress. Cell viability of the CML39-11A (wild type) and Sclsm4Δ1 mutant was measured after exposure to H2O2 at the indicated concentrations for 4 h. 6nM rapamycin was added 4h prior exposure to H2O2. Data are represented as the mean of three independent experiments ± standard deviation. ****p-value<0.0001.
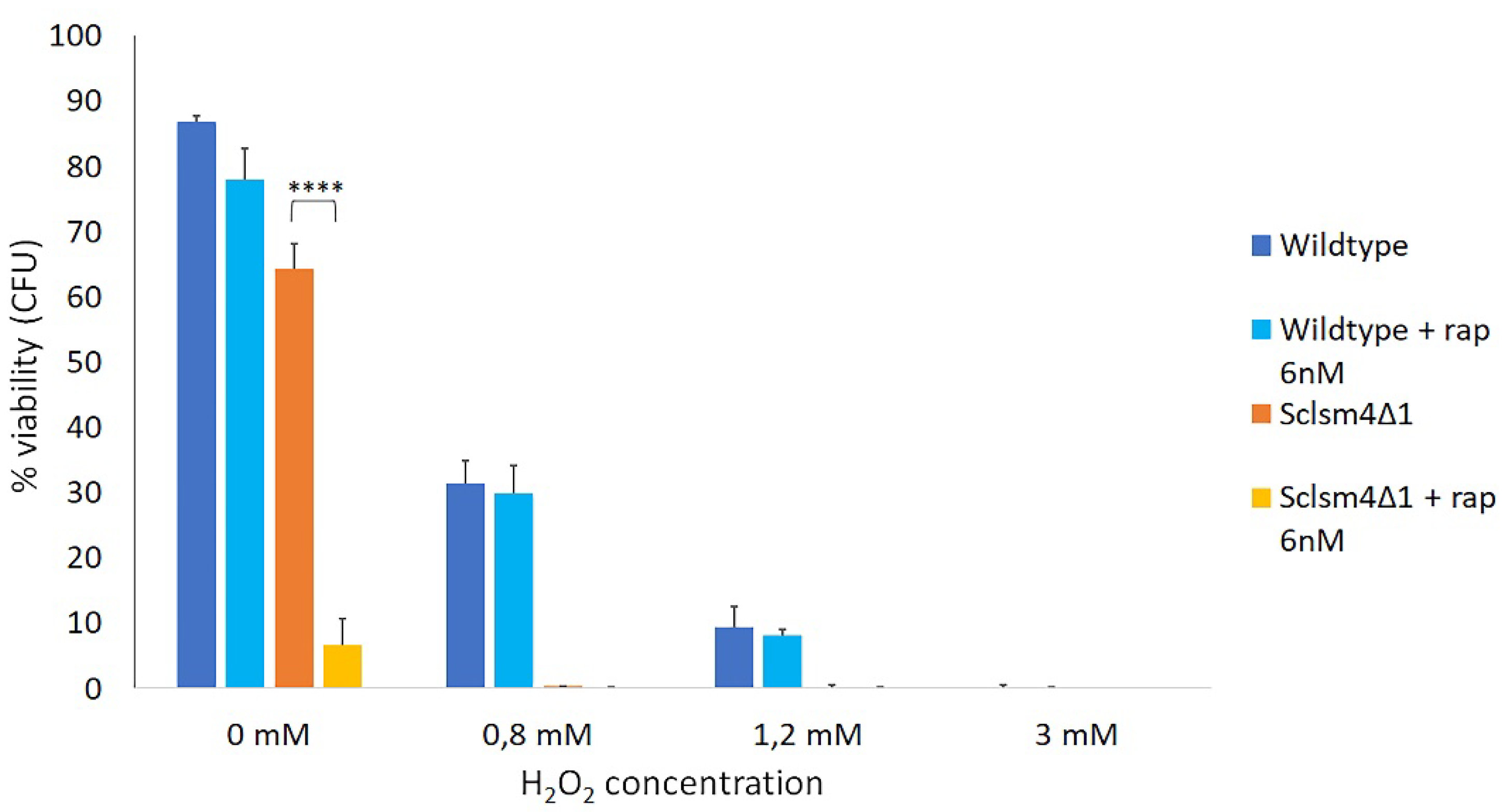
Figure 5.
Sclsm4Δ1 mutant shows defects in autophagy-related structures transport to the vacuole, as indicated by a higher percentage of GFP-ATG8 dots in the cytoplasm during nitrogen starvation and CLS. Wild type CML39-11A and mutant Sclsm4Δ1 cells expressing the fusion protein GFP-ATG8 were observed at the fluorescence microscope during exponential phase in both SD and SD-N medium for 4h (A), during post diauxic phase (PD) and in SD-N for 16h (C) and after 3 days of growth in SD (SD stat) or SD-N (SD-N stat) (E). GFP-Atg8 dots per cell were quantified from three biological replicates (n ≥ 300 cells), and the mean of cells containing one or ≥2 dots was plotted in (B), (D) and (F). Error bars represent standard deviation. *p-value<0.05 **p-value<0.01 ***p-value<0.001 ****p-value<0.0001.
Figure 5.
Sclsm4Δ1 mutant shows defects in autophagy-related structures transport to the vacuole, as indicated by a higher percentage of GFP-ATG8 dots in the cytoplasm during nitrogen starvation and CLS. Wild type CML39-11A and mutant Sclsm4Δ1 cells expressing the fusion protein GFP-ATG8 were observed at the fluorescence microscope during exponential phase in both SD and SD-N medium for 4h (A), during post diauxic phase (PD) and in SD-N for 16h (C) and after 3 days of growth in SD (SD stat) or SD-N (SD-N stat) (E). GFP-Atg8 dots per cell were quantified from three biological replicates (n ≥ 300 cells), and the mean of cells containing one or ≥2 dots was plotted in (B), (D) and (F). Error bars represent standard deviation. *p-value<0.05 **p-value<0.01 ***p-value<0.001 ****p-value<0.0001.
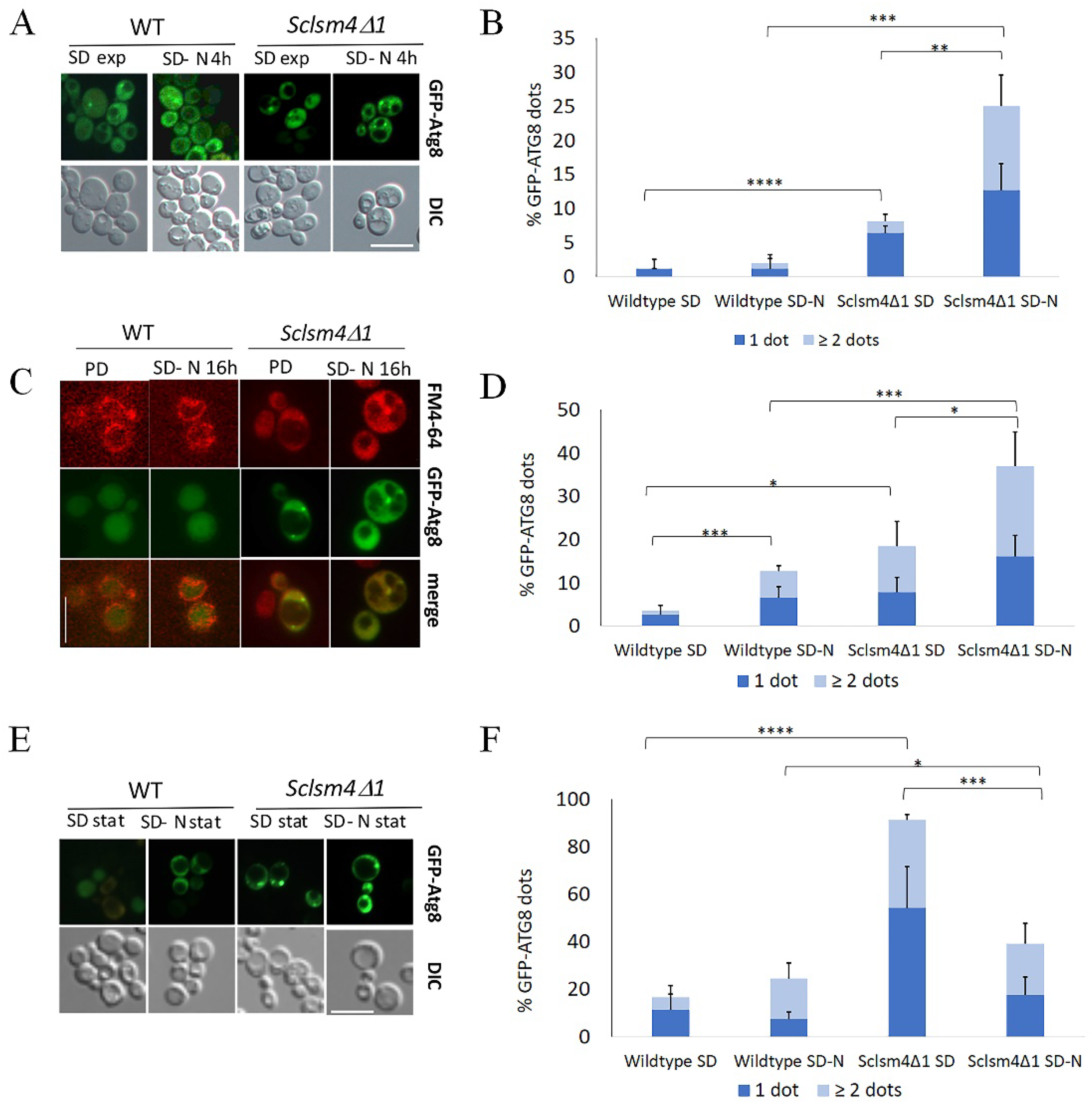
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



