
Submitted:
17 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
The North African E. pinnata is not known as a traditional medicinal plant but modern research has revealed its extracts richness in anti-oxidants and components of medicinal value. Despite its wide distribution and medicinal value, the species remains taxonomically and phytochemically un-derstudied, especially in Algeria. Here we compare three ecotypes of E. pinnata from the humid, sub-humid and semi-arid areas. The comparison was carried out using classical techniques of multivariate plant morphology, pollen grains size and shape, chromosome numbers, pollen fer-tility, anti-oxidant and antibacterial activities. Significant differences were revealed for all criteria except pollen fertility and antibacterial activity. The semi-arid ecotype showed to be a remote group in relation to the two others except for pollen size where the remote group was the sub-humid ecotype. Pollen size appeared positively correlated with karyotype length. The results (mg GAE/g EXT) of the antioxidant activity tests of the “humid”, “sub-humid” and the “semi-arid” plant groups are as follows, respectively; TPC: 50.79±0.51, 52.04±1.05 and 56.89±0.46; DPPH: 71.18±2.24, 86.39±3.02 and 95.67±2.02; RP: 11.09±1.24, 17.21±0.75 and 25.88±0.26. The inhibition area diameter in the antibacterial activity test varied from 9.25±1.06 mm (Semi-arid plants vs Pseudo-monas aeruginosa) to 12.00±1.41 mm (Sub-humid plants vs Escherichia coli).
Keywords:
Ebenus pinnata
; climate
; morphology
; pollen
; meiosis
; chromosomes
; antioxidants
; antibacterial activity
1. Introduction
The genus Ebenus comprises twenty species, fourteen of which are endemic to Turkey [1,2]. The six other species outside Turkey are E. cretica L. in Crete; E. sibthorpii DC. in the Southeastern Greece and Aegean Sea Islands [3,4]; E. stellata Boiss. in Iran, Oman, Afghanistan, Pakistan and India [4]; E. lagopus Boiss. in Southern Iran [5]; E. armitagei Schweinf. & Taubertin in Libya and Egypt [6–8] and E. pinnata Ait. in Libya, Tunisia, Algeria and Morocco [8,9].
Morphologically, the genus Ebenus distinguishes from other genera of tribe Hedysareae mainly by its corolla shorter than the calyx teeth and its 1-lomented pod enclosed within the calyx tube [4,10,11,12,13]. It constitutes a monophyletic group within the Hedysaroid clade [12] with its ancestral area of origin inferred in the Mediterranean Region [14], and its main center of diversity located in Turkey [15]. The closest species to E. pinnata is E. stellata [15].
Pollen grains in Ebenus species are radially symmetrical, isopolar, tricolpate, prolate or rarely perprolate, intine of 0.5 µm and exine of 1 µm, with reticulate ornamentations and more or less narrow lumina, polar diameter of 24–40 µm, equatorial diameter of 12.4–20 µm, with elliptical outlines on equatorial view, and circular to subcircular outlines on polar view [2,16,17,18]. No specific data were found in literature on E. pinnata pollen grains.
For all Ebenus species so far investigated, the chromosome number is 2n = 2x = 14 [16,19,20,21,22]. The karyotypes are symmetric and include a pair of satellite chromosomes [16,21]. For E. pinnata, two cytotypes have been mentioned: one with 2n = 14 from Morocco and Algeria [19,23,24] and the second with 2n = 18 [25].
Ebenus plants are used in traditional folk medicine to treat various health disorders in Turkey [26,27]. They contain 4.0-13.0% of seed oils including five different fatty acids [28]. Ebenus haussknechtii Bornm. ex Hub.-Mor. is used to prevent skin problems, hypertension and stomach diseases [26]. The analysis of its chemical composition identified various natural compounds (including two flavonoid glycosides and a methylinositol) with very significant antimicrobial activity [26]. As to E. hirsuta Jaub. & Spach., it is used to treat kidney disorders [27,29]. Its aerial parts contain mainly hyperoside, rutin, hesperidin, tannic acid and p-coumaric acid [30]. Its extracts display strong antigenotoxic effects and a significant activity against bacteria and fungi [30]. The authors [30] suggest that E. hirsuta, together with Cytisopsis dorycniifolia, could be beneficial for preparation of high-value pharmaceuticals and functional food ingredients. Similarly, E. laguroides Boiss. and E. macrophylla Jaub. & Spach. have proven to be with significant antioxidant and antibacterial activity due to various chemical compounds, especially, rutin, the dominant component in Ebenus species [31]. Analyses of roots and aerial parts extracts of E. boissieri Barbey have revealed immunomodulatory and antitumor activity inducing apoptosis in breast cancer cells [32] and caspase-mediated apoptotis on cervical cancer cell line Hela [33,34]. E. boissieri can be considered as a source of new anti-apoptotic and therefore anti-carcinogenic agents [32,35]. In addition to its osteoprotective role, E. cretica is very rich in flavonoids and isoflavones such as formononetin [36], maesopsin glucoside (aurone) and other compounds [37]. The results obtained by Kounadi et al. [37] represent valuable information on the beneficial effects of E. cretica, which could be used as a food supplement, a functional food or even as a medicine. E. cretica administration exerts a significant beneficial effect on bone density loss in ovariectomized rats [38]. Mitrocotsa et al. [39] report a long list of important components isolated from E. cretica and E. sibthorpii DC including D-pinitol, quercetin, isorhamnetin glycosides and, especially, rutin-7,4’-di-O-methyl ether and 8,4’-dimethoxy-7-hydroxy-isoflavone, ionyl glycosides icariside B1 and B2. E. stellata is presented as vital for medication and bearing significant antioxidant activity [40]. This plant is also used to treat indigestion in livestock in Pakistan [41]. Phytochemical tests of E. stellata extracts, known for their anticonvulsant activity, have revealed the presence of coumarins, alkaloids, cardiac glycosides, flavonoids, quinone, saponins, steroids, terpenoids and tannins [42,43]. E. stellata extracts have significant antioxidant and antimicrobial (bacteria and fungi) activities [43]. A study by Kiazai et al. [44] has shown that heavy metals concentration in E. stellata plants is below the secure limits according to FAO and WHO, denoting their nontoxicity and suitability to be used as traditional medicines and for synthesis of new drugs. E. armitagei Schweinf. & Taub. appears among plants with moderate in vitro chistosomicidal activity [45]. The study of the chemical composition of E. pinnata plants from Tunisia has detected the presence of several secondary metabolites including ombuoside, kaempferol 3-O-rutinoside, rutin, catechin, picein, β-sitosterol and sitosterol β-glucoside, the first four being with significant antioxidant activity [46,47]. A study by Nouioua and Gaamoune [48] has shown that E. pinnata extract has a relatively high antioxidant activity (IC50 = 12) compared to other taxa from Algeria.
It appears that E. pinnata plants are insufficiently investigated under both taxonomic and phytochemical point of view. In the present study, we compare three Algerian E. pinnata populations from different climate stages (Humid, Sub-humid and Semi-arid) using multivariate whole plant morphology, pollen grains size and shape, pollen fertility, karyology, antioxidant and antibacterial activities. The results are statistically evaluated and discussed.
2. Materials and Methods
2.1. Plant material
The whole plant morphology was studied on plants harvested from populations located in three different climate stages (humid, sub-humid, semi-arid) [49]. The plants were kept fresh using plastic bags and moistened paper until study in the laboratory. More details are given Table 1. Altogether, 126 plants were sampled: 44 specimens from the humid stage, 55 from the sub-humid, and 27 for the semi-arid.
In addition to fresh plants reserved to multivariate morphological analysis, inflorescences were collected from at least five plants at different stages of development: young floral buds for meiosis analysis, flowers just before anthesis for pollen fertility assessment and pollen grains measurements. The material was in situ fixed in 10 ml tubes containing absolute ethanol-glacial acetic acid-chloroform (6:3:1).
The plant material destined to chemical extractions was collected on March 2023 from the same populations as for morphology. 400 to 500 g of fresh leaves and young stems (at the beginning of young inflorescences occurrence) were harvested from each climate stage. The plant material was spread on paper sheets for ten days under sun-free conditions. The sun-free dried plant material was then powdered and stored in sealed glass containers for further use.
2.2. Whole plant morphology analysis
Forty-four quantitative morphological characters (Table 2, character 1-44) were measured on the 126 fresh plants harvested from the three climate stages (see above and Table 1). Measurements were done using a tape measure (stem height), a sliding caliper (stem diameter), a ruler (internodes, leaves, inflorescences…), graph paper (small features such as hairs, flower parts, pods and seeds). A binocular magnifier was used for measurements and observations of minute features. To minimize errors because of character mis-appreciation, the same researcher (T. Zemouri) did all the morphometric scorings. Qualitative traits of color and hairiness are uniform, so they were not included in the analysis.
2.3. Pollen grains size and shape
In situ fixed flowers just before anthesis (see above) were used to recover anthers on a microscope slide containing a drop of lactopropionic orcein prepared according to Dyer [50]. The anthers were dissected under a stereomicroscope to recover pollen grains. After eliminating anther debris, a cover glass was carefully applied over the stain drop. After 10 mn, pollen grains are well stained. The observations were done under an Optica B-353A light microscope. Pollen grains were photographed at 40x magnification. Five flowers from different racemes were used for each population. Altogether, 235 pollen grains (Humid: 116; Sub-humid: 78; Semi-arid: 41) were measured for their polar (P) and equatorial (E) axes; the rate P/E and the product PxE were calculated. Pollen grains shapes and number of apertures were scored.
2.4. Meiosis analysis
In situ fixed young floral buds were used (see above). In a drop of lactopropionic orcein [50] on a slide, after 5-6 minutes in water bath at 60°C, floral buds were dissected to recover the young anthers. Under a cover glass, the anthers were squashed with the thumb to eject the pollen mother cells. The observations were made under an Optica B-353A light microscope to search for under-division mother cells. The best metaphases I and II were photographed at 100x magnification.
2.5. Pollen fertility assessment
In situ fixed flowers (see above), just before anthesis, were used. In a small drop of distilled water on a slide, the anthers of a flower were recovered by dissecting and pressing them with needles to obtain the maximum of pollen grains. After drying the drop of water by slight heating on a hotplate, a small drop of cotton blue [51] was added on the dried spot. Pollen grains stain after 10 mn at room temperature. The observations were made under a coverslip at 40x magnification on an Optica B-353A light microscope. 10 to 30 flowers from different plants were used. 700 to 900 pollen grains per flower were screened. Well stained grains with uniform outlines were considered fertile, whereas the light stained ones, with irregular outlines and relatively small sizes were counted sterile. Pollen fertility rate (TF) is expressed as:
2.6. Plant extract preparation
The dried and finely powdered plant material from each of the three climate stages (5 g for each assay) were extracted with 100 mL of absolute ethanol for 6 hours, or continued until the extract gives no coloration, using a Soxhlet apparatus. At the end of the extraction, the liquid extract was filtered and evaporated in a vacuum at 40°C to complete dryness, using a rotavapor apparatus. The extraction yield was calculated using the following equation:
2.7. Determination of antioxidant activity
2.7.1. Total phenolic content
The total phenolic content of plant extracts was measured by the Folin-Ciocalteu rGAEent assay, using the method described by Singleton et al. [52] with a few modifications. A diluted solution of each extract (200 µL) was mixed with 750 µL of Folin–Ciocalteu rGAEent (previously diluted with water 1:10 v/v). This mixture was maintained at ambient temperature for 5 minutes after which 400 µL of sodium carbonate solution (75 g/L in water) was added. The mixture was left to stand for 1 h at room temperature. The absorption was measured at 765 nm against water blank. The total phenolic contents of the extracts were calculated using the calibration curve of gallic acid standard. Results were given in mg gallic acid equivalent (GAE)/g of dry extract.
2.7.2. DPPH free radical scavenging assay
Free radical scavenging activity was determined using the 2, 2-diphenyl-1-picrylhydrazyl free radical (DPPH) method [53] with some modifications. 25 µL from each extract were added to 975 µL of 100 µM methanolic solution of DPPH. The mixture was shaken and left in the dark at room temperature. After 30 minutes, the absorbance was recorded at 517nm, and compared to the absorbance of blank sample containing 25 µL of methanol and the same amount of DPPH solution. A standard calibration curve was obtained using different gallic acid concentrations. Antioxidant activity was expressed as mg gallic acid equivalent (GAE)/g of dry extract.
2.7.3. Reducing power assay
The method was based on [54] procedures with modifications. 0.125 mL of each extract sample was mixed with 2.5 mL phosphate buffer (0.2 M, pH 6.6) and 2.5 mL of potassium ferricyanide [K3Fe (CN)6] (1%). The mix was incubated in water bath at 50°C for 10 minutes followed by addition of 2.5 mL of trichloroacetic acid (10%) and then centrifuged at 1500 rpm for 10 minutes. Finally, 2.5 mL of the upper layer solution was mixed with distilled water (2.5 mL) and FeCl3 (0.5 mL, 0.1%). The absorbance was recorded at 700 nm. Reducing power assay was expressed as mg gallic acid equivalent (GAE)/g of dry extract.
2.8. Screening of the antibacterial activity
2.8.1. Bacterial strains
The antibacterial activity test of E. pinnata extracts included six foodborne pathogen bacteria provided by Pasteur Institute (Algiers, Algeria) and identified with the ATCC number (American Type Culture Collection). The Gram-negative bacteria: Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Vibrio cholerae ATCC 14035, Salmonella typhi ATCC 14028 and the Gram-positive bacteria: Staphylococcus aureus ATCC 25923 and Methicillin-Resistant S. aureus ATCC 43300 (MRSA) were studied. All strains were grown in nutrient agar (NA) and incubated at 37°C for 18–24 h until the stationary growth phase was reached.
2.8.2. Antibacterial test
The antibacterial activity was conducted using agar well-diffusion method in accordance with the National Committee for Clinical Laboratory Standards [55]. Inoculum containing 106 CFU/mL of each bacterial culture to be tested were evenly spread on the surface of Mueller Hinton agar plates using sterile swabs. Subsequently, wells of 6 mm diameter were punched into the agar medium and filled with 40 μL (5 mg/mL) of plant extract dissolved in dimethyl sulfoxide (DMSO) and allowed to diffuse at room temperature for 2 h. The plates were then incubated in the upright position at 37°C for 24 h. Well containing the same volume of DMSO served as negative control. After incubation, the diameters (mm) of the growth inhibition zones were measured. All tests were repeated in triplicate.
2.9. Statistical evaluation
Whole plant morphology and pollen size data were treated using principal components analysis (PCA) and/or cluster analysis. Before performing cluster analysis, variable values were standardized by centering (Xi = xi – mean). Mean values were compared using unequal N HSD test or t test for paired (dependent) samples. Homogeneity of variances was checked using Levene’s test. Statistical analyses and homogeneity tests were performed using Statistica 8.0 [56].
3. Results
3.1. Whole plant morphology
All the studied populations showed a hemicryptophyte habit (aerial parts dying after fructification and sprouting up from the base the next season) except in rare cases where some plant stems tend to be evergreen, especially in humid spots. All the plants appear the same for their qualitative traits: Greenish-brownish hairy stems, with long upright white hairs; glaucous leaves, thickly hairy leaflets on both sides, appressed hairs; purple corolla parts.
The analysis of the quantitative characters (Table 2, characters 1-44; see SM1 for the raw data matrix) by PCA yielded the scatterplot in Figure 1A (Plane 1-2) where the three studied groups are perfectly separated. On planes 1-3, 1-4 and 1-5 of the PCA (Appendix A), the semi-arid population confirmed to be a separate group from the other two merged groups (humid and sub-humid). The cumulated of the explained variances of the five axes is 68.67%. The characters explaining the separation of the groups on Figure 1A are given in Table 3 and Appendix A.
Table 3 details the morphological comparison of the three groups. There are significant differences for all morphological characters except DI (diameter of inflorescence), WFB (Width of the flower bract), LC2 (Length of the corolla), and LHCT (Length of hairs at the base of the calyx teeth). The morphological comparison of the three groups is better summarized by the cluster analysis in Figure 1B where the “semi-arid” population shows up as a remote group in relation to the closer but different “humid” and “sub-humid” groups.
3.2. Pollen grains size
The results on pollen grains size are summarized in Table 3 (Characters 45-48). There are significant differences between groups for polar axis (P), P/E and PxE but none was revealed for equatorial axis (E). On the PCA scatterplot of Figure 2A (see SM2 for raw data), the three groups appear to be merged in relation to Axis 1, explained by E and P/E. However, in relation to Axis 2 explained by P, it is clear that the “sub-humid” group has most of the highest values of P. Most of the lowest values of P are in the “humid” group, those of the “semi-arid” group are mostly in the middle position between the two other groups. The dendrogram of Figure 2B shows that the “sub-humid” group is far distant from the other two groups. This topology is incongruent with that based on whole plant morphology (Figure 1B) where the “semi-arid” was the remote group. The cluster of the three groups of E. pinnata studied here appears as an outgroup of the other Ebenus species (Figure 2B).
In addition to this statistical evaluation, our observations showed that pollen grains of the studied material are prolate (P = c. 1.5x E), isopolar, radially symmetrical, tricolpate, circular outline on polar view, elliptical outline on equatorial view, surface with reticulate ornamentations (Figure 3G–I) for all the studied material.
3.3. Chromosome numbers, meiotic abnormalities and pollen fertility
Meiosis preparations showed, for all the three gyoups, 7 bivalents in metaphase I and 7 chromosomes in metaphase II (Figure 3A–F) corresponding to the chromosome number of 2n = 2x = 14, with a base number x = 7. On metaphase I, there are often 5 bivalents with circular pairing and two with linear pairing. No multivalents were observed, and chromosome segregation at anaphase-telophase I was regular since metaphase II was always with n = 7.
Meiosis abnormalities were often absent, cytomixis was observed only in a very few cases. Pollen fertility rates were high for all the assessed flowers of the three groups (Table 3 and Appendix C). Pollen fertility rates were of 89.64-98.71% in the “humid” group, 89.01-98.07% in the “sub-humid” and 91.97-97.85% in the “semi-arid” (see SM3 for pollen fertility raw data). No significant differences were detected among groups for pollen fertility (Table 3, line 49).
3.4. Antioxidant activity
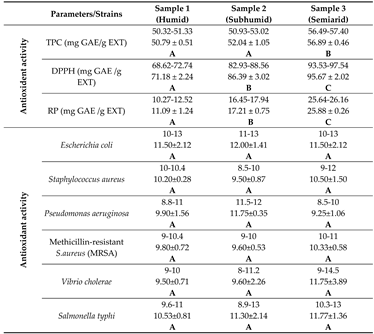
Antioxidant capacities of the studied extracts were evaluated by two in vitro methods: the free DPPH radical scavenging test and the reducing power assay. The results are shown in Table 4.
In our results, E. pinnata extract from the semi-arid area (sample 3) had statistically (P <0.05) the highest value of total phenolic content (56.89 ± 0.46 mg GAE/g dry extract), followed by samples 1 and 2 with similar concentrations. The sample 3 also expressed the best scavenging activity (95.67±2.02 mg GAE/g dry extract) and reducing power (25.88±0.26 mg GAE/g dry extract), followed by the sub-humid sample and then the humid one. On cluster analysis (Figure 4), the “semi-arid’ sample behaved as a remote group in relation to the closer “humid” and “sub-humid” ones.
3.5. Antibacterial activity
The antibacterial activity results displayed in Table 4 revealed that all studied extracts were potentially effective in inhibiting microbial growth. Although statistical analysis showed no significant difference between the three extracts, the semi-arid sample seems more active against the four microorganisms tested, namely Staphylococcus aureus, Methicillin-resistant S. aureus, Vibrio cholera and Salmonella typhi with inhibition diameters (ID) of 10.50±1.50, 10.33±0.58, 11.75±3.89 and 11.77±1.36 mm, respectively. On the other hand, the sub-humid sample tends to be more efficient against Escherichia coli and Pseudomonas aeruginosa, with ID of 12.00±1.41 and 11.75±0.35 mm, respectively. The cluster analysis dendrogram based on data Table 4 is shown in Figure 4.
4. Discussion
Our results revealed that, morphologically, the three compared groups were significantly different for most characters, including those of the reproductive system (Table 3, Appendix A). The differences are slight but significant. The multivariate analysis (Figure 1 and Appendix B) showed that the three groups were perfectly discriminated. The “semi-arid” population behaved as a remote group in relation to the other close “humid” and “sub-humid” groups. Is this morphological divergence due to phenotypic plasticity alone? Or is it underpinned by a genetic divergence too? Although the genetic basis is very likely (as suggested by pollen data below), we have no reliable clue, at this point, to clearly addressing the issue. Experimental cultures [57] and molecular genetic analysis [58,59] are required to clarify the question.
Based on pollen data, the three groups discriminated well too (Table 3, APPENDIX A). The “sub-humid” group showed larger pollen size than the two close “humid” and “semi-arid” groups. This situation is well rendered by the cluster analysis on Figure 2B. When compared to other E. species, using literature data [2,15,18], the three groups constituted a separate cluster within which distances between groups are of the same range as those separating other accepted Ebenus species (Figure 2B). This observation supports the hypothesis that the phenotypic differences between the three groups have a genetic basis. Moreover, pollen grain size (as expressed by PxE) of the three groups is highly correlated (R = 0.89, p = 0.30) with total haploid chromosome length (THCL) of the three groups (unpublished data). PxE/THCL (µm2/µm) values are, in increasing order, 229.76/23.82 (humid), 236.64/26.98 (semi-arid) and 253.69/28.33. This is not the case in other Ebenus species since their pollen grain sizes [2,15,18] are not correlated (R = 0.22, p= 0.72) with their haploid total chromosome lengths [16,21] (Appendix C). Pollen size is not always positively correlated to genome size across taxa [60]. A pollen grain is not just plant cells with their nuclei, cytoplasm, membranes and walls. It is above all the exine whose structure and thickness reflect evolutionary history and selective adaptation to various aspects of the environment. The variability of the exine structure and thickness seems to be at the origin of lack of positive correlation of pollen and genome size across taxa.
The chromosome number in our material, as it was repeatedly shown on both metaphase I and metaphase II, is 2n = 2x = 14 (x = 7) as it has already been reported from Morocco [19] and Algeria [23,24]. Preliminary results of an underway karyomorphological study show significant differences in chromosome lengths of material from the three climate stages involved here. On the karyogram reported by Gadoum and Hamma [23], there are two pairs of chromosomes with two pairs of large satellites. The 4 extra chromosomes of the 2n = 18 reported for a Moroccan population by Parra et al. [25] may correspond to those 4 large satellites mistaken for chromosomes.
According to Siddiqui and Alrumman [61], and the references therein, cytomixis is a phenomenon induced by both genetic and environmental factors; it occurs in mutants, hybrids, aneuploids and stressed plants (heat, cold, drought, parasites and pollution), leading to reduced pollen fertility. In our case, very scarce cytomixis and associated abnormalities were encountered while screening meiosis preparations, which is in accordance with the high pollen fertility rates assessed (89.01–98.71%) (Table 3 and Appendix A). From this perspective, it can be drawn that the local E. pinnata populations are in perfect equilibrium with their environment, without any aberrations occurring due to mutation, aneuploidy or hybridization between diverging genotypes.
In the present study, the antioxidant and antimicrobial activities of Ebenus pinnata ethanol extracts revealed an interesting potential that could be used as an alternative medicinal source. Reports concerning E. pinnata phytochemicals and biological activities are very scarce. In the study by Abreu et al. [47], the measurement of antioxidant activity of methanol extract from E. pinnata aerial parts collected from Tunisia using three different tests (DPPH and ABTS radical scavenging activities, and reducing power assay) has revealed an interesting potential. The authors linked this activity specifically to the presence of four phenolic compounds: ombuoside, kaempferol-3-O-rutinoside, rutin, and catechin. Another study from Algeria related to antioxidant parameters of methanol extracts of nine species, Nouioua and Gaamoune [48] have reported that E. pinnata extracts show a relatively high DPPH radical scavenging activity (IC50 of 12.25±2.80 µg/mL) compared to eight other taxa from Eastern Algeria. They have also reported that this plant contains 8.57±0.16 mg GAE/g dry extract of TPC. The latter performances are very low compared with those obtained in the present study (Table 4).
The antioxidant potency of the semiarid extract sample was found to be stronger when compared with the two other samples (humid and sub-humid) for both assays. From these results, it is assumed that its high level of phenolics might have contributed to the observed antioxidant abilities. The present findings are in agreement with other studies reporting a high correlation between total phenolics and antioxidant activity [62,63].
In light of the antimicrobial results, it was observed that all extracts showed comparable power at the analyzed concentration. The main causes of the small differences observed in these results were the various bio-contents of the analyzed samples that were harvested from different bioclimatic stages (humid, sub-humid, and semi-arid). But sometimes, the same species could have different bioactive values since many factors may be responsible of these changes, such as harvesting period, water availability, environmental factors (climate and altitude), and technological factors [64,65]. Indeed, Kabtni et al. [66] have investigated the influence of climate variation on the phenolic composition and antioxidant activity of Medicago minima populations selected from different provenances in Tunisia. They have concluded that the highest phenolic contents are observed in populations from the semi-arid area with a BSK climate and an altitude higher than 550 m, which agrees with the findings of this study. The accumulation of a higher level of phenolic compounds and the expression of the best antioxidant activity for E. pinnata grown under a semiarid climate, characterized by high temperature and low precipitation, can be related to hydric and thermal stresses [67,68].
5. Conclusions
The three plant groups from the humid, sub-humid and semi-arid climate stages are significantly different for all of the whole plant morphology, pollen grains size and anti-oxidant activity but not for antibacterial activity. The semi-arid group revealed to be morphologically and chemically far different from the two closer humid and sub-humid groups. The same chromosome number of 2n = 2x = 14 was repeatedly counted. Pollen grain size seems positively correlated to karyotype length (genome size). Experimental cultures and molecular analysis are required to check the genetic basis hypothesis of the phenotypic heterogeneity of the groups. It would be interesting to compare the chemical composition of the three groups extracts.
Supplementary Materials
The following supporting information is attached to the paper as supplementary files: SM1: Whole plant morphology raw data; SM2: Pollen grains size raw data; SM3: Pollen fertility rates raw data.
Author Contributions
Conceptualization: MS and TZ; Plant material collect: MS and TZ; Morphometrics scoring: TZ; Meiosis and pollen grains analysis: TZ and HB; Antioxidant and antibacterial activities analysis: AC and TZ; Statistic evaluation: MS; Results interpretations, MS, TZ, AC, HB, Original draft preparation: MS and AC (for antioxidant and antibacterial activities); Supervision: MS; Final version preparation MS and TZ. All authors have read and agreed to the published version of the manuscript
Funding
This research received no external funding, The study was carried out in the framework of research activities of the Laboratory of Ecology and Environment (University of Bejaia, Algeria) supported by Direction Générale de la Recherche Scientifique et du Développement Technologique (DGRSDT), Ministry of High Education and Scientific Research (MESRS), Republic of Algeria.
Data Availability Statement
The data used in this research are provided as supplementary files attached to the article.
Acknowledgments
We would like to warmly thank all the technical staffs of the Laboratory of Ecology and Environment and the Laboratory of Microbial Ecology for their full availability during the preparation of this research.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Detailed morphology of groups.
Appendix B
Detailed whole plant morphology PCA results.
Appendix C
Pollen grains size vs karyotype length.
References
- Aytaç Z. (2000). The genus Ebenus L. (Leguminosae/Fabaceae) in Turkey Karaca Arboretum Magazine 5: 145–171.
- Aytaç Z., Yildirim H. (2018). Ebenus zekiyeae (Fabaceae), a new species from Turkey. Annales Botanici Fennici 55(4-6): 25–29. [CrossRef]
- Kaveh A. and Kazampour-Osaloo S. (2012). Cladistic analysis of the genus Ebenus (Fabaceae-Hedysareae) based on morphological data. Taxonomy and Biosystematics 4(11): 61–68. DOI: 20.1001.1.20088906.1391.4.11.7.8.
- Aytaç Z., Suludere Z., Pınar M. (2015). Examination of the leaflets hairs and stoma structures with the electron microscope of the genus Ebenus L. (Leguminosae) in Turkey. Biodicon 8(2): 32–50.
- Boissier E. (1872). Flora Orientalis: Save Enumeratio Plantarum In Oriente a Græcia et Ægypto ad Indfiæ Fines Hucusque Observatarum – vol. 2. APUD EUMDEM, 65, RUE DE LYON.
- Boulos L. (1999). Flora of Egypt. Vol. One (Azollaceae-Oxalidaceae). Cairo, Egypt: Al Hadara Publishing. 340 p.
- Jafri S.M.H. (1980). Fabaceae. In: Flora of Libya, V. 118, S.M.H. Jafri & A. El-Gadi (Eds.), Al Faateh University, FacuIty of Science, Department of Botany, Tripoli.
- Dobignard A., Chatelain C. (2012). Index synonymique de la flore d'Afrique du Nord. Vol.4, Dicotyledoneae : Fabaceae - Nymphaeaceae. Conservatoire et Jardin botaniques de la Ville de Genève, hors-série 11b.
- Quézel P., Santa S. (1962). Nouvelle flore d'Algérie et des régions désertiques méridionales - Tome 1. Ed CNRS. Paris.
- Huber-Morath, A. 1970: Ebenus L. In: Davis PH, Editor. Flora of Turkey and the East Aegean Islands, vol. 3. Edinburgh, UK: Edinburgh University Press, pp. 590–596.
- Çelebi A., Açik L., Aytaç Z. (2009). Biosystematic studies among Ebenus L. species based on morphological, RAPD-PCR and seed protein analyses in Turkey. Pak. J. Bot. 41(5): 2477–2486.
- Amirahmadi A., Kazampour-Osaloo S., Moein F., Kaveh A., Maassoumi A.A. (2014). Molecular systematics of the tribe Hedysareae (Fabaceae) based on nrDNA ITS and plastid trnL-F and matK sequences. Pl. Syst. Evol. 300:729–747. [CrossRef]
- Duan L., Wen J., Yang X., Liu P.L., Arslan E., Ertuğrul K., Chang Z.Y. (2015). Phylogeny of Hedysarum and tribe Hedysareae (Leguminosae: Papilionoideae) inferred from sequence data of ITS, matK, trnL-F and psbA-trnH. Taxon 64:49–64. [CrossRef]
- Kaveh A., Kazampour-Osaloo S. (2015). Estimation of Ebenus species divergence time based on nrDNA ITS and matK cpDNA sequences. 4th National Congress of Plants. 12, 13 May 2015, Tehran, Iran.
- Ghanavati F., Amirabadizadeh H. (2012 Medicinal). Pollen grain morphology in Iranian Hedysareae (Fabaceae). Crop Breeding Journal 2(1): 25–33. [CrossRef]
- Aytaç Z., Ünal F. & Pinar M.N. (2000). Morphological, palynological, and cytotaxonomical study of Ebenus longipes Boiss. & Bal. and E. argentea Siehe ex Bornm. (Leguminosae) from Turkey. Israel Journal of Plant Sciences 48: 321–326. [CrossRef]
- Pinar N.M., Vural C., Zytac Z. (2000). Pollen morphology of Ebenus L. (Leguminosae: subfamily Papilionoideae) in Turkey. Pak. J. Bot. 32(2): 303–310.
- Halbritter H., Auer W., Igersheim A. (2020). Ebenus cretica. In: PalDat - A palynological database. https://www.paldat.org/pub/Ebenus_cretica/305777;jsessionid=B9B8A714C47DE9C0CB28F0595C4D0398; accessed 2021-01-25.
- Molero J., Montserrat-Marti J. M. (1986). Números cromosomáticos de plantas marroquíes. Collectanea Botanica 16: 351–354.
- Davis P.H., Mill R.R., Kit Tan. (1988). Flora of Turkey and the East Aegean Islands, vol. 10 (supplement). Edinburgh, UK: Edinburgh University Press, pp. 381-382.
- Aksoy H., Ünal F., Aytaç Z. (2001). Karyological study on four endemic Ebenus L. taxa (Leguminosae) in Turkey. Caryologia, 54(4): 307–311. [CrossRef]
- Rice et al. 2015. The Chromosome Counts Database (CCDB) – a community resource of plant chromosome numbers. New Phytol. 206(1): 19–26.
- Gadoum N., Hamma A. (2016). Etude cytogénétique de l’espèce Ebenus pinnata Aiton (Fabaceae) du Golfe de Béjaïa et de la Vallée de la Soummam. Mémoire de Master en Sciences Biologiques. Université Abderrahmane Mira de Bejaia, Algérie. 58 p. http://univ-bejaia.dz/dspace/123456789/10254.
- Hadawat A.K., Madani S. (2022). Etude cytogénétique de quelques populations d’Ebenus pinnata Ait. (Fabacées) de la région de Béjaïa. Mémoire de Master en Sciences de l’Environnement. Université Abderrahmane Mira de Bejaia, Algérie. 49 p. http://univ-bejaia.dz/dspace/123456789/21618.
- Parra R., Valdés B., Gordillo I., Venanzi R. (1999). In: Kamari, G., Felber, F. and Garbari, F. (ed.): Meditenanean chromosome number reports – 9 – Fl. Medit. 9: 323–387.
- Uyar Z., Böke N., Türkay E., Koz Ö., Yas A.I., Kırmızıgül S. (2006). Flavonoid glycosides and methylinositol from Ebenus haussknechtii. Nat. Prod. Res. 20(11): 999–1007. [CrossRef]
- Özdemir E., Alpınar K. (2015). An ethnobotanical survey of medicinal plants in western part of central Taurus Mountains: Aladaglar (Nigde–Turkey). Journal of Ethnopharmacology166: 53–65. [CrossRef]
- Azcan N., Saricoban S., Demirci B., Aytaç Z., Baser K.H.C. (2001). Seed oils of fifteen Ebenus taxa growing in Turkey. Chemistry of Natural Compounds 37(3): 253–255. [CrossRef]
- Kültür S., Gürdal B., Sari A., Melikoğlu G., 2021. Traditional herbal remedies used in kidney diseases in Turkey: an overview. Turkish Journal of Botany 45: 269–287. [CrossRef]
- Ceylan R., Katani J., Zengina G., Mati S., Aktumsek A., Boroja T., Stanic S., Vladimir Mihailovic V., Gokalp Ozmen Guler G.O., Boga M., Yılmaz M.A. (2016). Chemical and biological fingerprints of two Fabaceae species (Cytisopsis dorycniifolia and Ebenus hirsuta): Are they novel sources of natural agents for pharmaceutical and food formulations? Industrial Crops and Products 84: 254–262. [CrossRef]
- Bektas E., Kaltalioglu K., Sahin H., Turkmen Z., Kandemir A., (2018). Analysis of phenolic compounds, antioxidant and antimicrobial properties of some endemic medicinal plants. International Journal of Secondary Metabolite 5(2): 75–86. [CrossRef]
- İmir N., Aydemir E., Şimşek E., Göktürk R., Yesilada E., Fişkin K. (2016). Cytotoxic and immunomodulatory effects of Ebenus boissieri Barbey on breast cancer cells. Genetics and Molecular Research 15(1). [CrossRef]
- Simsek E., Imir N., Aydemir E.R., Gokturk R.S., Yesilada E., Fiskin K. (2017). Caspase-mediated apoptotic effects of Ebenus boissieri Barbey extracts on human cervical cancer Cell Line HeLa. Pharmacognosy Magazine 13(50): 254–259. [CrossRef]
- Bhuwan J.C., Vijay J., Archana S.N., Piyush V., Minky M. (2022). Review on documented medicinal plants used for the treatment of cancer; Current Traditional Medicine 8(2): e111021197159. [CrossRef]
- Aydemir E.A., Simsek E., Imir N., Göktürk R.S., Yesilada E., Fiskin K. (2015). Cytotoxic and apoptotic effects of Ebenus boissieri Barbey on human lung cancer cell line A549. Pharmacognosy Magazine, 11(Suppl 1), S37. [CrossRef]
- Ingham J.L., 1978. Flavonoid and isoflavonoid compounds from leaves of sainfoin (Onobrychis viciifolia). Z. Naturforsch. 33: 146–148. [CrossRef]
- Kounadi S., Aligiannis N., Pongratz I., Lelovas P., Ismini D. and Skaltsounis A. (2011). Estrogenic activity of the methanolic extract of Ebenus cretica L. Planta Medica 77 - PM168. [CrossRef]
- Dontas I.., Kounadi S., Aligiannis N., Galanos A., Skaltsounis A. and Lelovas P., (2019). Plant extract administration and mild daily exercise increase bone density of ovariectomized rats. In Abstracts of 14th FELASA congress 2019, PC41, p. 157. Laboratory Animals 53(1): 28–203.
- Mitrocotsa D., Skaltsounis A.-L., S., Harvala C., Tillequin F. (1999). Flavonoid and terpene glycosides from European Ebenus species. Biochemical Systematics and Ecology 27: 305–307. [CrossRef]
- Mandokhail A., Samiullah, Khan K., Tareen A.H., Attiq-Ur-Rehman Kakar A.U.-R, Tariq S., Kakar N., (2020). Determination of antioxidants by four gifferent methods in medicinally jmportant plant Ebenus Stellata of Balochistan. Al-Nahrain Journal of Science.23(4): 13–18. [CrossRef]
- Abidin S.Z.U., Munem A., Khan R., Batiha G.E.-S., Amhad M., Zafar M., Khalil A.A.K., Hetta H.F., Mahmoud M.H., Sami A., Bhatti M.Z. (2021). Ethnoveterinary botanical survey of medicinal plants used in Pashto, Punjabi and Saraiki Communities of Southwest Pakistan. Veterinary Medicine and Science 7(5): 2068–2085. [CrossRef]
- Khodaparast A., Sayyah M., Sardari S. (2012). Anticonvulsant activity of hydroalcoholic extract and aqueous fraction of Ebenus stellata in mice. Iranian Journal of Basic Medical Sciences 15(3):. 811–819.
- Zameer S., Ali S., Gulmeena, Tareen R.B. (2022). Identification of volatile constituents and antimicrobial activity of Ebenus stellata. GU. J. Phytosci. 2(4): 214–222.
- Kiazai I., Samiullah, Khan N., Attiq-Ur-Rehman, Abdul Ghaffar, Abdul Baqi. (2019). Determination of heavy metals concentration in Astragalus anisacanthus and Ebenus stellata of Balochistan, Pakistan. Pure and Applied Biology 8(3): 2028–2035. [CrossRef]
- Yousif F., Hifnawy M.S., Soliman G., Boulos L., Labib T., Mahmoud S., Ramzy F., Yousif M., Hassan I., Mahmoud M., El-Hallouty S.M., El-Gendy M., Gohar L., El-Manawaty M., Fayyad W., El-Menshawi B.S. (2007). Large-scale in vitro. screening of Egyptian native and cultivated plants for schistosomicidal activity, Pharmaceutical Biology 45(6): 501–510. [CrossRef]
- Braham H., Ben Jannet H., Castedo L., Mighri Z. (2004). Isolation, for the first time, of a flavonoid glycoside and the (±)-catechin from the aerial parts of Ebenus pinnata. Journal de la Société Chimique de Tunisie 6: 153–160.
- Abreu P.M., Braham H., Ben Jannet H., Mighri Z., Matthew S. (2007). Antioxidant compounds from Ebenus pinnata. Fitoterapia 78: 32–34. [CrossRef]
- Nouioua W., Gaamoune S. (2018). Antioxidant, antimicrobial and anti-inflammatory activities development of methanol extracts of some species growing in the massif of Boutaleb, Setif, Algeria. International Journal of Pharmacy and Natural Medicines 6(1): 15–20.
- Mebarki A. (2005). Hydrologie des bassins de l’Est algérien : Ressources en eau, aménagement et environnement. Thèse de doctorat d’état (Géographie et Aménagement du Territoire, option : Hydrologie), Université Mentouri de Constantine, Algérie. 360 p.
- Dyer A.F. (1963). The use of lacto-propionic orcein in rapid squash. Methods for chromosome preparations. Stain Technol. 38: 85–90. [CrossRef]
- Mertens T.R., Hamnersmith R.L. (1998). Genetic laboratory investigations. Eleventh ed. Prentice Hall inc., Upper Saddle River, New Jersey.
- Singleton V.L., Orthofer, R., Lamuela-Raventos R.M. (1999). Analysis of total phenols and other oxidation substrates and sntioxidants by means of Folin-Ciocalteu RGAEent. Methods in Enzymology 299: 152–178. [CrossRef]
- Blois M.S. (1958). Antioxidant determinations by the use of a stable free radical. Nature 181(4617): 1199–1200. [CrossRef]
- Oyaizu M. (1986). Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Japanese journal of nutrition and dietetics 44(6): 307–315. [CrossRef]
- NCCLS. (1993). Performance standards for antimicrobial disc suspectibility tests. Approved Standard NCCLS Publication M2-A5, Villanova, PA, USA.
- Statsof, Inc. (2007). Statistica (data analysis software system). version 8.0. www. Statsoft.com.
- Fazlioğlu F. (2018). Phenotypic plasticity of ecotypes across habitats. Akademik Ziraat Dergisi 7(2): 253–258. [CrossRef]
- Hu X., Lan S., Song X.,Yang F., Zhang Z., Peng D., Ren M. (2021). Genetic divergence between two sympatric ecotypes of Phalaenopsis pulcherrima on Hainan Island. Diversity 13: 446. [CrossRef]
- Stronen A.V., Norman A.J., Wal E.V., Paquet P.C. (2021). The relevance of genetic structure in ecotype designation and conservation management. Evolutionary Applications 15(2): 185–202. [CrossRef]
- Knight C.A., Clancy R.B., Götzenberger L., Dann L., Beaulieu J.M. (2010). On the relationship between pollen size and genome size. Journal of Botany 2010, Article ID 612017, 7 pages. [CrossRef]
- Siddiqui S., Alrumman S.A. (2022). Methomyl, imbraclaobrid and clethodim induced cytomixis and syncytes behaviors in PMCs of Pisum sativum L: Causes and outcomes. Saudi J. Biol. Sci. 29(9): 103390. [CrossRef]
- Zhang Y., Wang Z. (2009). Phenolic composition and antioxidant activities of two Phlomis species: A correlation study. CR Biologies 332: 816–826. [CrossRef]
- Kumar S., Sandhir R., Ojha S. (2014). Evaluation of antioxidant activity and total phenol in different varieties of Lantana camara leaves. BMC Research Notes 7, 560. [CrossRef]
- Ahuja I., De Vos R.C., Bones A.M., Hall R.D. (2010). Plant molecular stress responses face climate change. Trends in Plant Science 15(12): 664–674. [CrossRef]
- Rabeta M.S., Nur Faraniza R. (2013). Total phenolic content and ferric reducing antioxidant power of the leaves and fruits of Garcinia atrovirdis and Cynometra cauliflora. International Food Research Journal 20: 1691–1696.
- Kabtni S., Sdouga D., Bettaib Rebey I., Save M., Trifi-Farah N., Fauconnier M.L., Marghali S. (2020). Influence of climate variation on phenolic composition and antioxidant capacity of Medicago minima populations. Scientific reports 10(1): 8293. [CrossRef]
- De Abreu I.N., Mazzafera P. (2005). Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiology and Biochemistry 43(3): 241–248. [CrossRef]
- Al-Huqail A., El-Dakak R.M., Sanad M.N., Badr R.H., Ibrahim M.M., Soliman D. & Khan F. (2020). Effects of climate temperature and water stress on plant growth and accumulation of antioxidant compounds in sweet basil (Ocimum basilicum L.) leafy vegetable. Scientifica (Cairo). 2020 Feb 27; 2020: 3808909. [CrossRef]
Figure 1.
Multivariate analysis of morphological characters of three Ebenus pinnata ecotypes. A: PCA based on 44 quantitative traits measured on 126 fresh plants (see Tabs. 1-3 for more details). B. Cluster analysis of centered mean values of the 44 quantitative traits (UPGMA linkage).
Figure 1.
Multivariate analysis of morphological characters of three Ebenus pinnata ecotypes. A: PCA based on 44 quantitative traits measured on 126 fresh plants (see Tabs. 1-3 for more details). B. Cluster analysis of centered mean values of the 44 quantitative traits (UPGMA linkage).
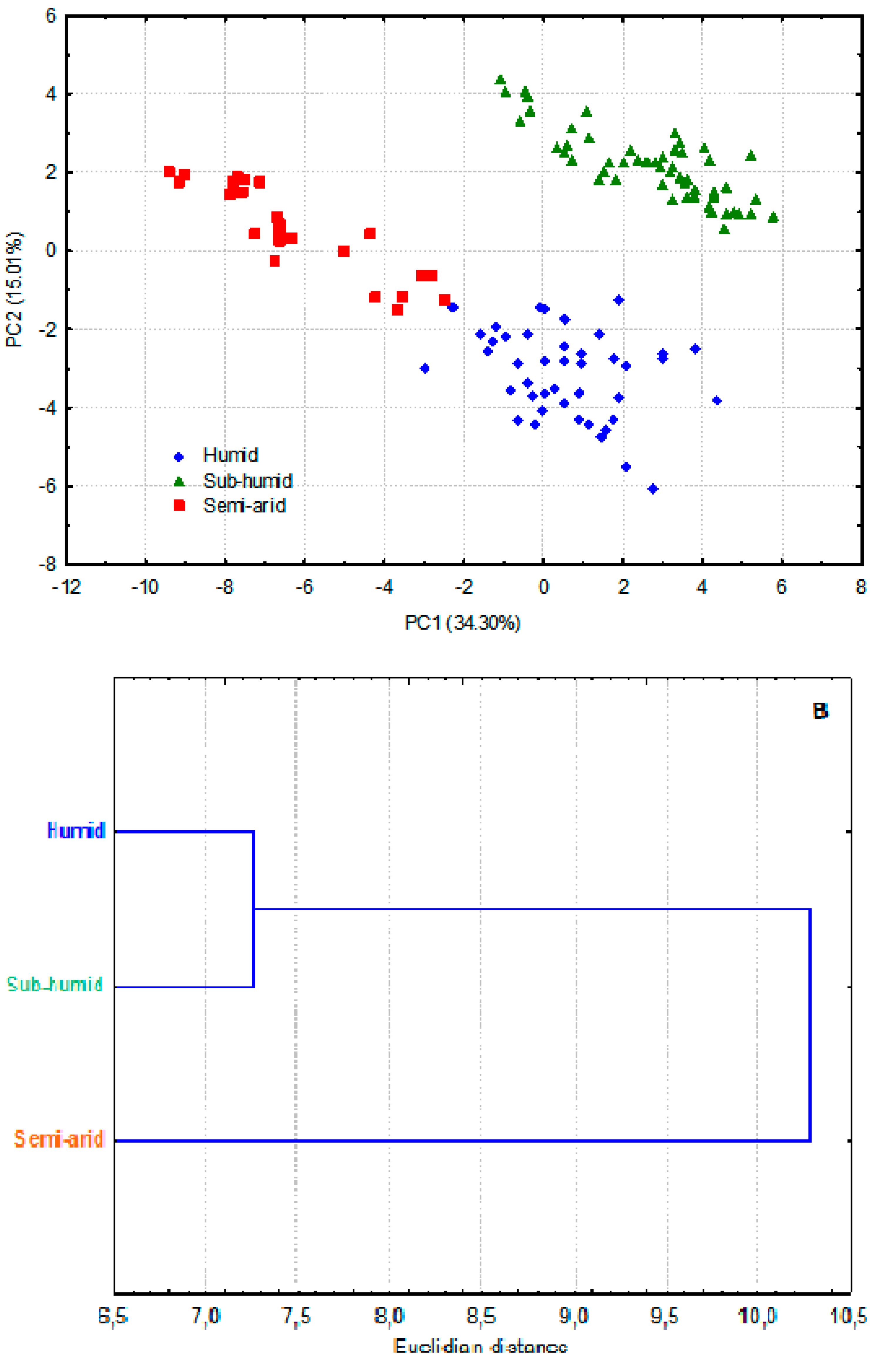
Figure 2.
Multivariate analysis of pollen size. A. PCA based on three pollen grains traits (polar axis P, equatorial axis E and their rate P/E) measured for the three Ebenus pinnata ecotypes. Axis 1 is explained by E and P/E with respectively R = 0.98 and R = -0.78; Axis 2 is explained by P with R = 0.94. B. Cluster analysis based on the centered mean values of three pollen traits (P, E and P/E).
Figure 2.
Multivariate analysis of pollen size. A. PCA based on three pollen grains traits (polar axis P, equatorial axis E and their rate P/E) measured for the three Ebenus pinnata ecotypes. Axis 1 is explained by E and P/E with respectively R = 0.98 and R = -0.78; Axis 2 is explained by P with R = 0.94. B. Cluster analysis based on the centered mean values of three pollen traits (P, E and P/E).
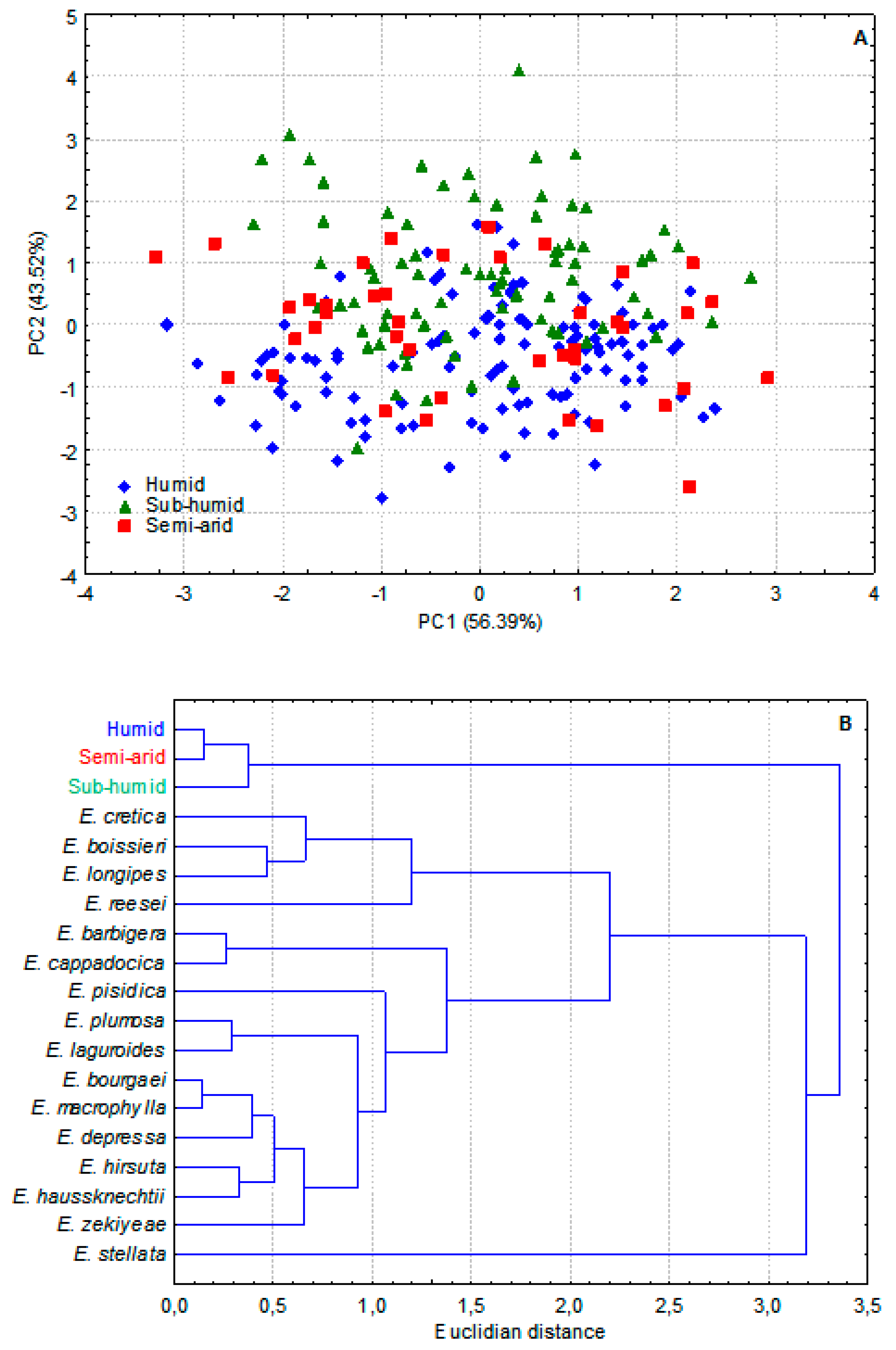
Figure 3.
Haploid phase chromosomes and pollen grains in Ebenus pinnata. A-C: Metaphase I of the “humid”, sub-humid” and “semi-arid” groups respectively (7 bivalents); D-F: Metaphase II in the three groups respectively (7 chromosomes); G: Cotton blue stained fertile pollen grain (on the right) and sterile pollen grain (on the left); H: Polar view of a Lactopropionic orcein stained pollen grain showing three symmetrical colpi and reticulate ornamentations; I: Lactopropionic orcein stained pollen grain with its pollen tube and nucleus (arrow).
Figure 3.
Haploid phase chromosomes and pollen grains in Ebenus pinnata. A-C: Metaphase I of the “humid”, sub-humid” and “semi-arid” groups respectively (7 bivalents); D-F: Metaphase II in the three groups respectively (7 chromosomes); G: Cotton blue stained fertile pollen grain (on the right) and sterile pollen grain (on the left); H: Polar view of a Lactopropionic orcein stained pollen grain showing three symmetrical colpi and reticulate ornamentations; I: Lactopropionic orcein stained pollen grain with its pollen tube and nucleus (arrow).
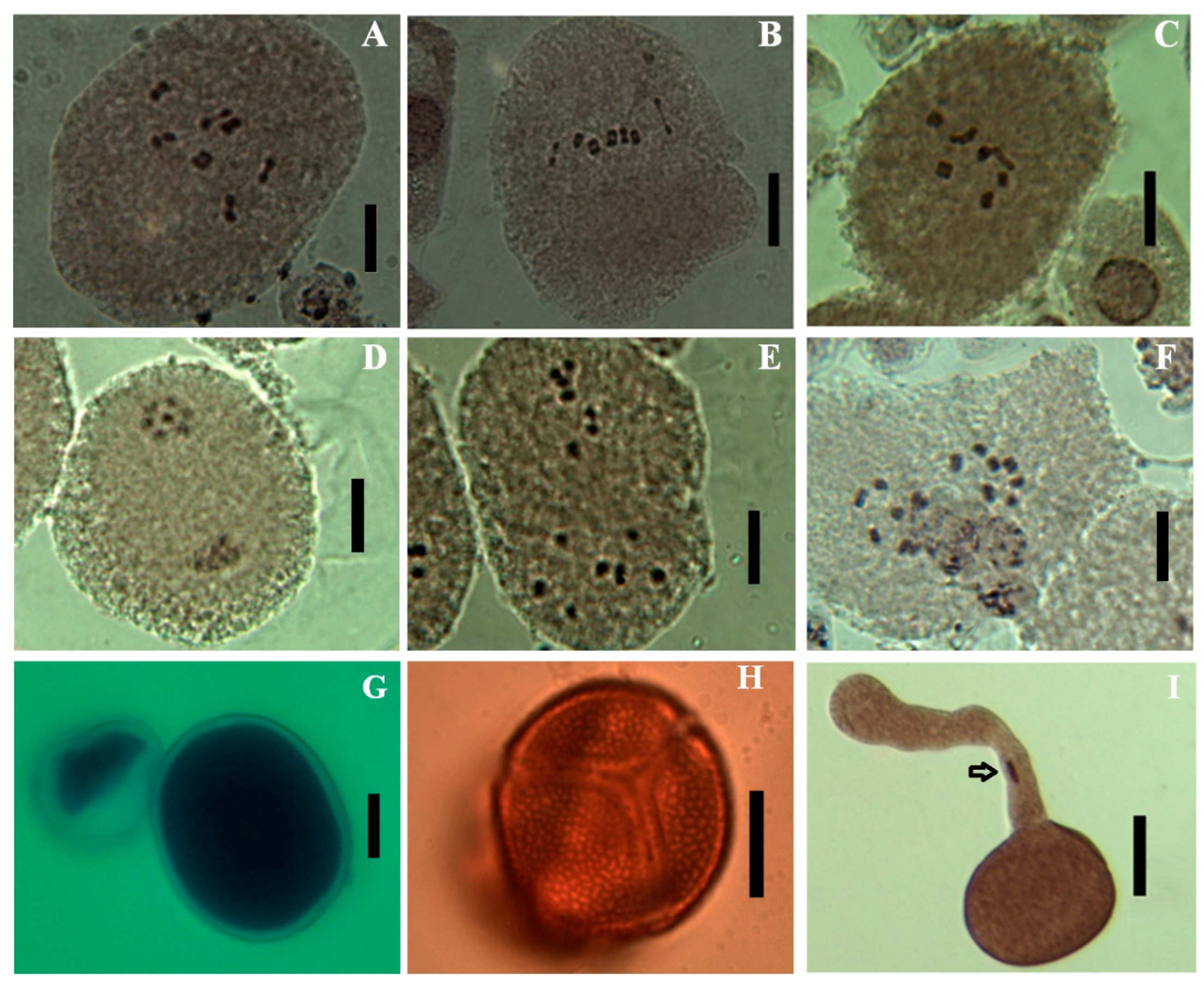
Figure 4.
Cluster analysis of antioxidant and antibacterial activities of three ecotypes of Ebenus pinnata from different climate stages. A. Antioxidant activity; B. Antibacterial activity. Data source in Table 4. The same topology was obtained when both antioxidant and antibacterial performances were used (not shown).
Figure 4.
Cluster analysis of antioxidant and antibacterial activities of three ecotypes of Ebenus pinnata from different climate stages. A. Antioxidant activity; B. Antibacterial activity. Data source in Table 4. The same topology was obtained when both antioxidant and antibacterial performances were used (not shown).
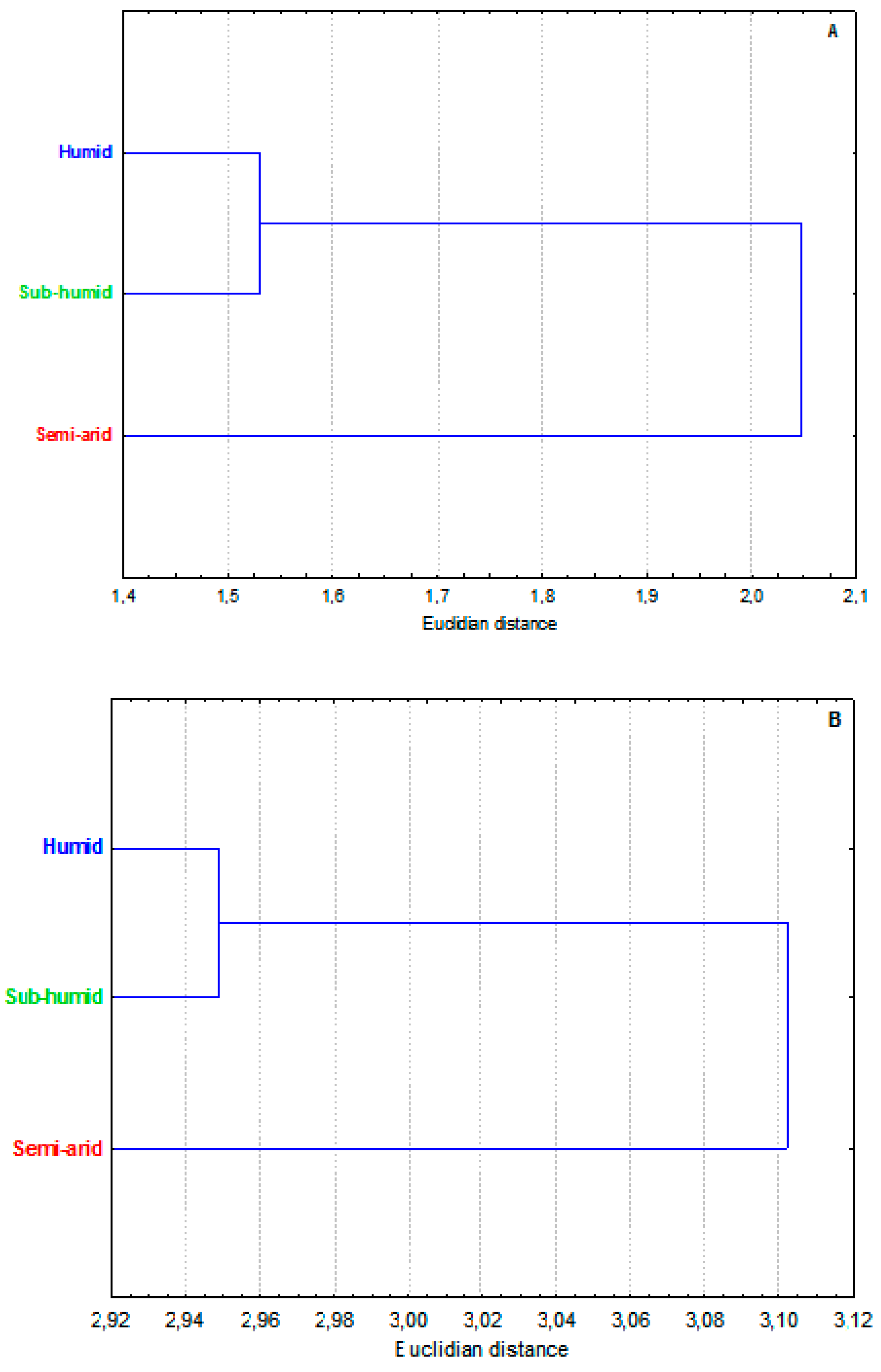

Table 1.
Characteristics, sampling dates, sample sizes and codes of the populations used in multivariate morphological study. H: Humid; SH: Sub-humid SA: Semi-arid.
Table 1.
Characteristics, sampling dates, sample sizes and codes of the populations used in multivariate morphological study. H: Humid; SH: Sub-humid SA: Semi-arid.
| Climate stage | Humid | Sub-humid | Semi-arid |
| Locality name | Kherrata | Semaoun | Boudjelil |
| Population Code | H | SH | SA |
| GPS Localization | 36° 31' 33.30"N 5° 16' 49.73" |
36° 37' 27.16"N 4° 49' 7.20"E |
36° 22' 11"N 4° 26' 48.30"E |
|
Altitude/ Exposure |
612m/East | 195m/West | 270m/East |
| Soil | Red clay | Brown clay | White ground |
| Plant formation | Road bank | Garrigue, Grassland | Grassland, Sparse garrigue |
| Rainfall1 (mm) | 800-1000 | 600-800 | 600-800 |
| Number of plants | 44 | 55 | 27 |
|
Sampling date/ Plant codes |
08/06/2015/ H01-H20 03/06/2015/ H21-H35 15/07/2015/ H236-H44 |
20/05/2015/ SH01-SH15 09/06/2015/ SH16-SH25 09/06/2015/ SH26- SH34 20/05/2015/ SH35-SH47 28/06/2016 SH48-SH55 |
13/05/2015/ SA01-SA13 07/06/2015/ SA14-SA27 |
1 Mebarki [49].
Table 2.
Morphological characters used in the morphological multivariate study. C: Continuous, D: Discrete.
Table 2.
Morphological characters used in the morphological multivariate study. C: Continuous, D: Discrete.
| N° | Coding | Character name | Type | Unit |
|---|---|---|---|---|
| 1 | DBP | Diameter at the base of the plant | C | cm |
| 2 | LLS | Length of the longest stem | C | cm |
| 3 | NS | Number of stems | D | Stems |
| 4 | DLS | Diameter of the longest stem | C | mm |
| 5 | LIN | Length of the third internode | C | cm |
| 6 | NI | Number of inflorescences (racemes) | D | Raceme |
| 7 | LS1 | Length of stipules | C | mm |
| 8 | WS | Width of stipules | C | mm |
| 9 | LL | Length of the leaf (3rd node) | C | cm |
| 10 | WL | Width of the leaf (3rd node) | C | cm |
| 11 | LP | Length of the petiole | C | cm |
| 12 | NPL | Number of pairs of leaflets (3rd node) | D | Pairs |
| 13 | LNLP | Lowest number of leaflets pairs | D | Pairs |
| 14 | HNLP | Highest number of leaflets pairs | D | Pairs |
| 15 | LLB | Length of the leaflet blade | C | cm |
| 16 | WLB | Width of the leaflet blade | C | mm |
| 17 | LIP | Length of the inflorescence peduncle (3rd node) | C | cm |
| 18 | HI | Height of the inflorescence | C | cm |
| 19 | DI | Diameter of the inflorescence | C | cm |
| 20 | LNF | Lowest number of flowers | D | Flower |
| 21 | HNF | Highest number of flowers | D | Flower |
| 22 | LFB | Length of the flower bract | C | mm |
| 23 | WFB | Width of the flower bract | C | mm |
| 24 | LC1 | Length of the flower calyx | C | mm |
| 25 | LC2 | Length of the corolla | C | mm |
| 26 | TLW | Total length of the wing | C | mm |
| 27 | WWB | Width of the wing blade | C | mm |
| 28 | WBW | Width at the base of the wing | C | mm |
| 29 | WMW | Width at the middle of the wing | C | mm |
| 30 | TWK | Total width of the keel | C | mm |
| 31 | LWPK | Length of the widest part of the keel | C | mm |
| 32 | TLK | Total length of the keel | C | mm |
| 33 | TLS | Total length of the standard | C | mm |
| 34 | LSB | Length of the standard blade | C | mm |
| 35 | WSB | Width of the standard blade | C | mm |
| 36 | LSP | Length of the standard ‘petiolule’ | C | mm |
| 37 | LCM | Length of the calyx at maturity | C | mm |
| 38 | LCT | Length of the calyx tube at maturity | C | mm |
| 39 | LHCT | Length of hairs at the base of the calyx teeth | C | mm |
| 40 | LPM | Length of the pod at maturity | C | mm |
| 41 | WPM | Width of the pod at maturity | C | mm |
| 42 | LS2 | Length of the seed | C | mm |
| 43 | WS | Width of the seed | C | mm |
| 44 | LR | Length of the radicle | C | mm |
| Extra variables (Not included in multivariate analyses) | ||||
| 45 | P | Polar diameter of pollen grains | C | µm |
| 46 | E | Equatorial diameter of pollen grains | C | µm |
| 47 | P/E | Rate of P and E diameters of pollen grains | C | |
| 48 | PxE | Product of P *E. | C | µm2 |
| 49 | PF | Pollen fertility rate | C | % |
Table 3.
Morphological comparison of Ebenus pinnata plants from different climate stages. Values are expressed as Mean ± SD (See Appendix A for more details). FL1 and FL2 are correlations R with CP1 and CP2 of PCA respectively (Explanatory R values are shown in bold). See Table 2 for trait full names and measure units. Mean values with different capital letters are statistically different (Unequal N HSD Test, α = 0.05). Lowercase letters a and b compare the polar (P) against the equatorial (E) axis of pollen grains (t-Test for dependent samples, α < 0.001). NA: Not attributed.
Table 3.
Morphological comparison of Ebenus pinnata plants from different climate stages. Values are expressed as Mean ± SD (See Appendix A for more details). FL1 and FL2 are correlations R with CP1 and CP2 of PCA respectively (Explanatory R values are shown in bold). See Table 2 for trait full names and measure units. Mean values with different capital letters are statistically different (Unequal N HSD Test, α = 0.05). Lowercase letters a and b compare the polar (P) against the equatorial (E) axis of pollen grains (t-Test for dependent samples, α < 0.001). NA: Not attributed.
| N° | Trait Code |
FL1 | FL2 | Humid | Sub-humid | Semi-arid |
|---|---|---|---|---|---|---|
| 1 | DBP | 0.49 | -0.24 | 0.73±0.19 B | 0.73±0.33 B | 0.55±0.25 A |
| 2 | LLS | 0.80 | 0.10 | 68.22±18.38 B | 89.28±28.93 C | 44.41±19.79 A |
| 3 | NS | 0.23 | -0.60 | 9.55±6.52 B | 4.65±2.47 A | 3.11±2.41 A |
| 4 | DLS | 0.47 | -0.41 | 4.19±1.12 B | 3.75±1.01 AB | 3.16±1.11 A |
| 5 | LIN | 0.67 | 0.16 | 5.27±1.04 B | 6.53±1.57 C | 4.36±1.58 A |
| 6 | NI | 0.52 | -0.39 | 57.57±28.25 B | 47.89±32.23 B | 17.89±23.80 A |
| 7 | LS1 | 0.66 | 0.17 | 10.98±1.67 A | 13.60±2.74 B | 10.10±2.71 A |
| 8 | WS | 0.56 | -0.13 | 4.22±0.75 B | 4.41±0.74 B | 3.64±1.00 A |
| 9 | LL | 0.77 | -0.05 | 10.01±2.06 B | 10.85±2.60 B | 6.91±1.93 A |
| 10 | WL | 0.67 | -0.06 | 5.19±0.80 B | 5.55±0.88 B | 4.64±0.66 A |
| 11 | LP | 0.73 | 0.03 | 4.56±1.21 B | 5.23±1.61 B | 2.96±0.98 A |
| 12 | NPL | 0.43 | -0.07 | 4.73±0.50 B | 4.78±0.50 B | 4.11±0.97 A |
| 13 | LNLP | -0.18 | -0.29 | 2.80±0.55 B | 2.45±0.50 A | 2.81±0.68 B |
| 14 | HNLP | 0.43 | -0.23 | 4.98±0.15 B | 4.91±0.35 B | 4.52±0.89 A |
| 15 | LLB | 0.70 | 0.03 | 2.45±0.33 B | 2.66±0.35 C | 2.15±0.36 A |
| 16 | WLB | 0.22 | -0.63 | 7.77±1.51 B | 6.14±1.21 A | 6.18±1.46 A |
| 17 | LIP | 0.56 | -0.31 | 22.00±3.17 B | 21.29±3.56 B | 18.20±3.94 A |
| 18 | HI | 0.43 | -0.49 | 5.21±1.41 B | 4.40±0.75 A | 3.88±1.61 A |
| 19 | DI | 0.33 | -0.16 | 2.44±0.31 A | 2.40±0.25 A | 2.30±0.31 A |
| 20 | LNF | 0.24 | 0.24 | 11.84±8.05 A | 16.64±7.17 B | 13.56±9.89 AB |
| 21 | HNF | 0.66 | -0.41 | 56.43±11.83 C | 49.44±9.47 B | 33.93±17.42 A |
| 22 | LFB | 0.08 | 0.29 | 8.15±0.62 A | 8.60±0.89 B | 8.46±1.04 AB |
| 23 | WFB | 0.06 | -0.12 | 3.29±0.32 A | 3.25±0.18 A | 3.19±0.43 A |
| 24 | LC1 | -0.05 | 0.25 | 12.72±0.94 A | 13.21±0.90 B | 13.38±1.06 B |
| 25 | LC2 | 0.13 | -0.25 | 8.21±0.44 A | 8.05±0.24 A | 7.98±0.70 A |
| 26 | TLW | 0.40 | 0.87 | 1.79±0.08 A | 3.54±0.13 C | 2.40±0.07 B |
| 27 | WWB | 0.45 | 0.85 | 1.37±0.09 A | 2.45±0.07 C | 1.67±0.05 B |
| 28 | WBW | -0.67 | 0.23 | 1.10±0.10 A | 1.14±0.06 A | 1.38±0.04 B |
| 29 | WMW | -0.02 | 0.89 | 0.43±0.05 A | 0.89±0.07 C | 0.83±0.07 B |
| 30 | TWK | -0.86 | -0.34 | 4.21±0.23 B | 3.62±0.10 A | 4.99±0.09 C |
| 31 | LWPK | -0.86 | -0.25 | 3.47±0.26 B | 3.12±0.08 A | 4.14±0.08 C |
| 32 | TLK | -0.81 | -0.29 | 6.36±0.36 B | 5.94±0.11 A | 6.99±0.08 C |
| 33 | TLS | -0.89 | -0.02 | 6.65±0.45 B | 6.30±0.23 A | 8.04±0.08 C |
| 34 | LSB | -0.86 | -0.29 | 5.13±0.47 B | 4.49±0.12 A | 6.06±0.05 C |
| 35 | WSB | -0.87 | -0.39 | 4.67±0.36 B | 3.36±0.16 A | 6.04±0.10 C |
| 36 | LSP | -0.31 | 0.38 | 1.72±0.47 A | 1.81±0.16 A | 2.02±0.10 B |
| 37 | LCM | -0.42 | 0.67 | 13.19±0.48 A | 14.50±0.89 B | 15.26±0.18 C |
| 38 | LCT | 0.04 | 0.81 | 2.80±0.14 A | 3.29±0.12 C | 3.19±0.12 B |
| 39 | LHCT | -0.15 | 0.22 | 3.27±0.05 A | 3.28±0.07 A | 3.29±0.03 A |
| 40 | LPM | -0.85 | 0.14 | 5.09±0.09 A | 5.08±0.11 A | 5.99±0.10 B |
| 41 | WPM | -0.71 | 0.40 | 3.04±0.10 A | 3.15±0.13 B | 3.46±0.07 C |
| 42 | LS2 | -0.87 | -0.06 | 2.35±0.07 B | 2.19±0.10 A | 3.00±0.08 C |
| 43 | WS | -0.85 | 0.06 | 2.05±0.05 A | 2.02±0.05 A | 2.34±0.05 B |
| 44 | LR | -0.77 | -0.34 | 2.14±0.05 B | 2.02±0.07 A | 2.25±0.05 C |
| 45 | P | NA | NA | 17.95±1.10 Ab | 19.47±1.27 Bb | 18.43±1.08 Ab |
| 46 | E | NA | NA | 12.80±1.18 Aa | 13.03±0.99 Aa | 12.84±1.37 Aa |
| 47 | P/E | NA | NA | 1.41±0.10 A | 1.50±0.11 B | 1.45±0.15 AB |
| 48 | PxE | NA | NA | 230.56±31.60 A | 254.31±30.81 B | 237.16±32.74 A |
| 49 | PF | NA | NA | 94.04±2.64 A | 95.01±2.02 A | 94.97±2.11 A |
Table 4.
Comparison of antioxidant and antibacterial activities of three samples of Ebenus pinnata ethanolic extracts from different climate stages. For each parameter, values are expressed as Min-Max (upper line) and Mean ± SD (lower line). Different capital letters indicate highly significant differences between mean values (LSD test, α = 0.01 for antioxidant parameters, and α = 0.05 for antibacterial activity, with C>B>A). TPC: Total polyphenol content; DPPH: radical scavenging activity; RP: Reducing power assay.
Table 4.
Comparison of antioxidant and antibacterial activities of three samples of Ebenus pinnata ethanolic extracts from different climate stages. For each parameter, values are expressed as Min-Max (upper line) and Mean ± SD (lower line). Different capital letters indicate highly significant differences between mean values (LSD test, α = 0.01 for antioxidant parameters, and α = 0.05 for antibacterial activity, with C>B>A). TPC: Total polyphenol content; DPPH: radical scavenging activity; RP: Reducing power assay.
 |
1 Expressed as the diameter (mm) of the inhibition area.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



