Submitted:
19 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
Plasmodium vivax causes the second highest number of malaria morbidity and mortality cases in humans. Several biological traits of this parasite species, including the formation of dormant stages (hypnozoites) that persist inside the liver for prolonged periods of time, present an obstacle for intervention measures and create a barrier for the elimination of malaria. Research into the biology of hypnozoites requires efficient systems for parasite transmission, liver stage cultivation and genetic modification. However, P. vivax research is hampered by the lack of an in vitro blood stage culture system, rendering it reliant on in vivo derived, mainly patient material, for transmission and liver stage culture. This has also resulted in a limited capability for genetic modification, creating a bottleneck in investigations into the mechanisms underlying the persistence of the parasite inside the liver. This bottleneck can be overcome through optimal use of the closely related and experimentally more amenable nonhuman primate (NHP) parasite Plasmodium cynomolgi as model system. In this review we discuss the genetic modification tools and liver stage cultivation platforms available for studying P. vivax persistent stages and highlight how their combined use may advance our understanding of hypnozoite biology.
Keywords:
malaria
; hypnozoite
; Plasmodium vivax
; Plasmodium cynomolgi
; transfection
; genetic modification
; liver-stage
1. Introduction
1.1. Hypnozoite Biology
The disease malaria continues to affect a major proportion of the global human population. Data from the World Health Organization (WHO) indicate that in 2021, there were an estimated 247 million malaria cases in 84 malaria endemic countries [1]. More than 600,000 people died from the disease, with most deaths (76%) occurring in children aged under five years. Malaria is caused by protozoan parasites of the genus Plasmodium. Of the five species known to infect humans, by far the deadliest is Plasmodium falciparum. The second most important causative agent of human malaria is Plasmodium vivax. Even though morbidity and mortality numbers are lower than those attributed to P. falciparum, almost half of the world’s population is at risk of getting infected with P. vivax. Most cases have been recorded in the Americas, Southeast Asia, and the Western Pacific regions [2]. Recent reports also indicate widespread transmission of P. vivax in previously presumed resistant Duffy-negative people in Sub-Saharan Africa [3,4,5]. Infection with P. vivax is not trivial, as it can be severe, and it may even lead to death [6,7]. Especially young children and pregnant women are at risk [8].
P. vivax possesses some biological traits that promote a geographically widespread transmission and persistence inside the host. The most striking features are i) early sexual stage development enabling transmission prior to clinical symptoms [9] and ii) the development of dormant stages, so-called hypnozoites, that can remain without symptoms inside the liver for weeks to years before resuming growth, resulting in renewed blood stage infection (relapse) and transmission [10]. P. vivax selectively invades reticulocytes, resulting in low percentages of blood stage infection [11,12]. Hence, it has been suggested that relapse infections originally may have evolved as a mechanism to enable competition with more virulent Plasmodium species [13]. Later in evolution, further adaptation for prolonged persistence in the liver before renewed blood stage infection (‘long latency phenotype’) may have enabled to parasite to survive in temperate areas with shorter mosquito transmission seasons [14].
Hypnozoites are difficult to target with currently available drugs. Only the 8-aminoquinolines primaquine and tafenoquine have been approved as radical cure treatment drugs, but severe side effects, especially in children and pregnant women [15], precludes their widespread use.
Hypnozoites were discovered decades after the other stages of the malaria life cycle [16], and their research is challenging for multiple reasons. Firstly, the location of hypnozoites inside the liver is largely inaccessible for experimentation. Secondly, only a subset of primate malaria species form hypnozoites [17], restricting in vivo studies to NHPs and humans. Finally, by far most of the malaria research funding has been directed to the more deadly P. falciparum and P. vivax has been largely neglected [18]. With the elimination of malaria on the agenda [19], it has been recognized that increased research efforts are needed to tackle the complexities of the life cycle of P. vivax-type parasites [18]. During the past decade, various studies either using the human malaria parasite P. vivax or its sister NHP parasite Plasmodium cynomolgi, have unveiled a first glimpse of hypnozoite biology (reviewed below). This was enabled by technical advances in hypnozoite culture systems, the availability of genome sequences of P. vivax-type parasites and transcriptomics studies that identified genes that may be involved in hypnozoite biology. Follow-up studies dissecting the functional roles of the identified genes will greatly benefit from sophisticated genetic engineering systems.
1.2. Genetic Engineering of Plasmodium
The development of technology to genetically manipulate Plasmodium parasites [20,21] has been a key tool in a wide range of research areas. It has been instrumental in gaining an understanding in biological processes underlying the biology of different stages of the malaria parasite. For example, this technology has identified factors necessary for the regulation of gametocyte development [22,23] and enabled investigations into the functional roles of proteins involved in red blood cell invasion [24]. Furthermore, it has been implemented as tool in drug and vaccine development. For example, it has been used to create genetically attenuated parasites for experimental vaccines [25,26] as a new promising vaccine strategy. Furthermore, the development of bioluminescent Plasmodium reporter lines has enabled miniaturization of drug screening platforms, greatly improving efficiency of screening drugs against different stages of the parasite [27].
Genetic engineering platforms have been developed for a range of Plasmodium species [20,21,27,28,29,30,31]. Stable transfection of malaria parasites traditionally relies on a source of blood stage parasite material for the introduction of DNA, a procedure for introducing the DNA and a method to select for parasites that have taken up the DNA. Depending on the parasite species, this can be done in vitro (P. falciparum) [21], in vivo (P. vivax) [32,33] and the rodent malarias P. berghei, P. yoelii, P. chabaudi [20,28,30] or both (the NHP parasites P. knowlesi [34] and P. cynomolgi [29,35]) (Figure 1). Besides differences in in vitro/in vivo dependence, the level of advancement of developed tools differs between species. At the same time, species-specific biological differences may complicate the extrapolation of findings to other species. For example, between species clear differences exist in gametocyte biology, not only at the level of morphology, maturation rate and longevity, but also in the genes involved in gametocyte development [36].
Investigations into the biology of hypnozoites are particularly hampered by the paucity of model systems. P. vivax genetic engineering has been limited to blood stage parasites. To date, the only model system that has enabled hypnozoite research with genetically engineered parasites is the NHP parasite P. cynomolgi [37,38,39,40,41,42]. In this review, we describe the strengths, weaknesses, and opportunities of the different model systems available for P. vivax research in the context of the application of genetic manipulation technology to hypnozoite research.
2. Liver Stage Models for P. vivax
2.1. P. vivax In Vivo Liver Stage Models
The presence of P. vivax hypnozoites was originally demonstrated in liver biopsies of chimpanzees that had been experimentally infected with P. vivax sporozoites [10]. P. vivax only develops in humans, apes, and has been adapted to grow in some New World monkeys [43,44], and combined with the inaccessible location of hypnozoites and their low quantities inside the liver, this complicates liver stage research. Hence, studies investigating liver stage biology in situ have been limited until the seminal work of Mikholajczak et al [45] demonstrated that complete P. vivax liver stage development could be achieved in a severely immunocompromised FRG KO mouse model transplanted with human hepatocytes. A detailed phenotypic analysis of the liver stage parasites after different time points of infection demonstrated the presence of two distinct parasite populations inside the liver. The small stages were posited to be hypnozoites given their single nucleus, persistence throughout the 21-day time course of the study and differential insensitivity to the schizonticide atovaquone in comparison to growing lier stages. The hypnozoites were shown to slowly increase in size over time, indicating metabolic activity [45]. Following this work, this mouse model has been applied to study the impact of a partially protective P. vivax pre-erythrocytic vaccine on reducing relapses. It was shown that although the vaccine regimen did not completely prevent primary infection, treated mice experienced 62% fewer relapses [46]. Furthermore, another study demonstrated that by first eliminating liver stage schizonts through a schizont-specific drug treatment, recurrence of secondary schizonts could be assessed by microscopy and qRT-PCR, enabling assaying the activity of anti-relapse drugs [47]. Interestingly, the hypnozoite formation frequency in the humanized mouse model was reported to be lower than that observed for P. vivax in vitro platforms [47]. This could be a result of sub-optimal artificial in vitro culture conditions, or differences in parasite clearance rates between in vitro versus in vivo parasites. Since the different platforms use P. vivax sporozoites derived from field isolates, variation in parasite characteristics may also influence the hypnozoite ratios. A side-by-side comparison of the different P. vivax in vitro and in vivo models could possibly give more clues to this.
2.2. P. vivax In Vitro Liver Stage Platforms
P. vivax selectively invades reticulocytes, and as a result the development of a robust long-term in vitro blood stage culture system has proven cumbersome [48,49]. Hence, mosquito transmission needed for liver stage investigations can only be achieved with in vivo derived material from isolates from infected human subjects from endemic areas, or experimentally infected NHPs (with specifically adapted parasite strains) as source for sexual stages. Despite these complications, a wide variety of in vitro liver stage platforms has been developed to study hypnozoite biology and to assay drug activity against these stages, reviewed in [50]. This includes the infection of various types of hepatoma cell lines [51,52], immortalized human hepatocytes (HC-04) [53] or hepatocyte-like cell lines (imHC, [54], and iPSC-derived hepatocyte-like cells (iHCLs, [55]). In addition, primary human hepatocytes (PHH) have been used in different formats: in a coculture system [56,57], as 3D spheroids [58], or in 2D monolayer formats [59,60].
Using micropatterned primary human hepatocytes co-cultured with murine embryonic fibroblasts (MPCC) infected with P. vivax field isolates from Thailand, complete liver stage development was recapitulated in vitro including subsequent reticulocyte infection [56]. Both hypnozoites and liver stage schizonts were observed and characterized at the molecular level. This type of work is challenging, given the presence of liver stage parasites at a low infection rate (which differs between platforms) and a 100-fold smaller genome [61], hence lower amounts of transcripts, compared to the genome of the host cell [62]. These issues were counteracted by enrichment of parasite transcripts using custom made baits against the P. vivax P01 genome. Furthermore, bulk transcriptomics of hypnozoite-enriched samples [56] was achieved by treatment with a PI4K inhibitor known to selectively kill liver stage schizonts [63]. This showed that samples enriched for hypnozoites exhibited reduced transcriptional activity and had lower levels of transcripts encoding functions related to cell division and invasion machinery compared to untreated samples containing both hypnozoites and schizonts, indicative of a quiescent state of hypnozoites [56]. A follow-up of this work using the same platform in combination with Seq-Well single-cell sequencing enabled a more refined transcriptomic analysis of both parasite and host cells [64]. This indicated that hypnozoites can persist in a quiescent state through transcriptional or translational repression and rely on proteolytic activity to remain viable. Furthermore, the results revealed that a subset of sporozoites developing in hepatocytes with reduced metabolism may already be committed for sexual development [64]. Another study investigating P. vivax parasite and host cell transcriptomes at single-cell resolution used primary hepatocytes infected with Cambodian P. vivax parasites combined with a high-throughput droplet-based single cellRNA-seq workflow [65]. The results from this liver stage platform with high infection rates (5.50-6.93% P. vivax-infected hepatocytes at day 5 post sporozoite infection), underscored the notion that hypnozoites are metabolically active, albeit at low levels and found evidence for the existence of hypnozoites in different transcriptomic states. The authors hypothesized that this may reflect heterogeneity in phenotypes ranging from persisting to activating hypnozoites. In the ‘persister’ subgroup of hypnozoites, differential expression of genes involved in post-transcriptional/translational repression was observed, suggesting that these mechanisms may underly quiescence.
2.3. P. cynomolgi Liver Stage Research Platforms
P. cynomolgi is a NHP malaria parasite that is phylogenetically closely related to P. vivax and shares important characteristics of P. vivax including hypnozoite formation [66,67] and early gametocyte development [68]. Notably, the existence of hypnozoites as developmental stage of the life cycle in relapsing parasite species was for the first time demonstrated in biopsies of macaques infected with P. cynomolgi sporozoites [69]. The demonstration that drug activity profiles were highly similar between P. cynomolgi and P. vivax led in the eighties of the past century to the incorporation of large scale drug screening studies with P. cynomolgi sporozoite-induced infections in rhesus monkeys as central step in efforts (which also used patients undergoing P. vivax malaria therapy as well as prison inmate volunteers [70]) to find new drugs effective against hypnozoites [71,72,73,74,75,76,77,78]. In addition to the opportunity to study relapse infections in vivo, an in vitro liver stage culture system was developed for P. cynomolgi [79]. Recently, this in vitro system has been revived and adapted for drug screening drug screening of anti-hypnozoite compounds, setting the stage for the development of similar drug screening systems for P. vivax [80,81]. Furthermore, the availability of these in vitro systems has facilitated access to hypnozoites for biological studies.
Routinely, liver stage culture platforms of P. cynomolgi use monolayers of primary macaque hepatocytes in 96- or 384-well format [42,82]. In these cultures, generally ± 60% of P. cynomolgi M liver stage parasites are present as hypnozoites [81,83]. Cultures infected with P. cynomolgi B strain parasites appeared to contain a higher number of hypnozoites, which may reflect strain dependent variation (although it is not certain whether the M and B lines used were truly separate lines - [84]), or differences in culture conditions and/or hepatocyte cells. Full liver stage development of P. cynomolgi M liver stages was demonstrated in vitro including RBC invasion [41]. By applying a Matrigel cover to 2D monolayer cultures, P. cynomolgi liver stage cultures can be maintained for prolonged periods of time, providing the opportunity to study hypnozoite reactivation in vitro [85]. Hypnozoite reactivation was inferred from the observation that liver stage parasites emerged from cultures that had been drug treated to remove liver stage schizonts [85]. Using microdissection of P. cynomolgi infected liver cells, a first glimpse of the transcriptome of hypnozoites and liver schizonts was reported [86]. The authors suggested a potential role for an AP2-transcription factor which was termed AP2-Q in hypnozoite formation. The function of this transcription factor awaits functional validation.
Besides 2D culture capabilities, a 3D spheroid-cultured hepatocyte system has been developed to better preserve hepatocyte functionality in vitro [58]. Imaging of malaria-infected hepatocytes proved difficult in this system and only possible by time-consuming and data intensive imaging stacking methods. However, following dissociation, P. cynomolgi hypnozoites and liver schizonts obtained from the spheroid cultures could be detected. Furthermore, the system enabled recapitulation of the full life cycle as evidenced by the appearance of infected RBC after overlaying the spheroid-cultured hepatocytes with blood [58].
3. Transfection Technology to Study Hypnozoite Biology
3.1. P. vivax Genetic Engineering Technology
Functional analysis of mechanisms involved in hypnozoite biology depends on the availability of sophisticated tools for genetic engineering. For P. vivax, this type of tools is limited. Only two studies have reported proof of concept of the ability to genetically manipulate P. vivax [32,33]. In the absence of a P. vivax in vitro blood stage culture system, this type of work can only be performed in vivo. Hence, both transfection studies used splenectomized Saimiri boliviensis monkeys as donor to provide blood stage P. vivax as source for transfection.
In the first proof of successful P. vivax transfection, P. vivax Sal I blood stage trophozoites were obtained from a donor monkey [33]. Following percoll purification of trophozoite infected cells, parasites were electroporated with bioluminescence reporter constructs containing the firefly or renilla luciferase gene controlled by heterologous 5’ UTR’s (hrp3 or cam) and 3’ UTR’s (hrp2 or cam) of P. falciparum genes. Following transfection, parasites were maintained in 22h short-term cultures before luciferase measurements. This transient transfection system was successful as evidenced by detection of luciferase signals specifically in extracts from transfected parasites, indicating that these heterologous P. falciparum UTR’s display activity in P. vivax.
Nine years after this first publication, stable transfection of P. vivax was reported in which long-term maintenance of the constructs was achieved using a selectable marker and recipient monkeys to enable selection of drug resistant parasites [32]. Splenectomized Saimiri boliviensis donor monkeys were infected with the pyrimethamine-sensitive P. vivax Chesson line and parasites were electroporated with constructs carrying a pyrimethamine resistant P. vivax dihydrofolate reductase (Pvdhfr) gene and zinc-finger nucleases (ZFNs) to target constructs to the wild type P. vivax dihydrofolate reductase (Pvdhfr) locus. In contrast to the transient transfection study, parasites were not purified, but for each electroporation a leukocyte depleted RBC pellet obtained from a 3 ml blood sample containing P. vivax blood stages at a ± 0.5 % parasitemia was used. The developmental stage of the parasites at the time of harvesting was not mentioned. Following ex vivo electroporation, parasites were inoculated into 4 splenectomized Saimiri boliviensis recipient monkeys. In 1 of 4 recipient monkeys successful editing of the Pvdhfr was shown. An explanation for this suboptimal success rate was that parasitemias in the primates over several weeks are highly variable, depending on susceptibility to infection and immune responses, and can clear spontaneously without treatment [32].
3.2. P. vivax Hypnozoite Research Benefits and Limitations
Obviously, using P. vivax itself as model system is the ideal situation. Robust in vitro [56,60] and in vivo [47] liver stage models are available for studying hypnozoite biology. A major hurdle for P. vivax is the limited arsenal of genetic engineering tools to study functional aspects of hypnozoite biology. This is largely due to the lack of an in vitro blood stage culture system; hence genetic engineering of P. vivax has been fully reliant on the use of in vivo blood stage parasites from infected NHP donors and recipients. Due to the insusceptibility of macaques to P. vivax parasites [87], the two proof-of-concept studies reporting transfection of P. vivax depended on experimentally infected available New World monkeys [32,33]. Hence, a limited availability of these NHPs, ethical reasons, and costs complicate this type of experimental studies. One way to overcome this, may be to attempt using humanized mice as donor and recipients for genetic engineering of P. vivax or to achieve successful in vitro blood stage cultivation of P. vivax.
3.3. P. cynomolgi Genetic Engineering Technology
3.3.1. P. cynomolgi In Vivo Genetic Engineering Technology
Successful transfection of P. cynomolgi was demonstrated in 1999 in a proof-of-concept study in which the pyrimethamine sensitive P. cynomolgi M line was transfected with episomal constructs that included a pyrimethamine resistant dhfr gene to enable pyrimethamine selection of parasites that have taken up DNA in vivo in recipient monkeys [29]. Blood stage schizont-infected RBC obtained from a donor monkey were ex vivo electroporated with plasmids with a selectable marker gene controlled by UTR’s of P. berghei or P. falciparum (the genome of P. cynomolgi was published years later [88,89]). The material was then pooled and injected into a recipient monkey. After pyrimethamine selection in vivo, a resistant parasite population emerged. Only plasmids containing the P. berghei UTR’s were detected in resistant parasites, indicating that P. berghei dhfr-ts flanking regions may more efficiently control selectable marker gene expression than P. falciparum hrp2/hrp3 regions. A second transfection experiment used this technology to functionally investigate a 95 kDa protein located in the cavaola-vescicle complexes, the CVC protein (pcyphist/cvc-8195), present in both P. vivax and P. cynomolgi [90]. Attempts to target this gene by double crossover recombination using a linearized construct with a selection cassette flanked by homologous regions targeting pcyphist/cvc-8195 failed, indicating that the gene is essential. However, Akinyi et al. did report the first example of integration of a transgene (the RFP reporter gene) into the P. cynomolgi genome, which was shown by the continued expression of RFP in blood stage P. cynomolgi after the original in vivo selection and multiple blood stage passages in rhesus monkeys [90]. PCR and plasmid rescue analyses demonstrated the presence of the reporter gene, but the absence of the plasmid backbone. Given the uneven segregation of episomes, they are rapidly lost in the absence of selection pressure [91]. This complicates liver stage research as it is challenging to maintain drug selection pressure needed to maintain episomal constructs during mosquito transmission. For expression of transgenes during liver stage development, one option is to target constructs for integration into the genome, but this is a less efficient process than episomal transfection. Furthermore, it requires a neutral locus that can be targeted without having detrimental effects in different parts of the life cycle. While such loci have been described for other malaria parasite species [92,93], a neutral P. cynomolgi locus is currently not known. An alternative method for stable, low copy number transgene expression in liver stage parasites is to include a centromere in the transfection construct [94]. This method was successfully applied for the first time in Plasmodium by Iwanaga et al who showed that constructs containing a Plasmodium centromere could be efficiently transfected and maintained throughout the complete life cycle in P. berghei and P. falciparum [95]. By analogy, transfection of P. cynomolgi with a plasmid construct that includes a newly identified P. cynomolgi centromere resulted in maintenance of the construct throughout the parasite life cycle, with a mean of 66% of liver stage parasites expressing the transgenes [40] (Table 1).
3.3.2. Application of P. cynomolgi Vivo Derived Transgenic Parasites to the Study of Hypnozoite Biology
The centromere-containing plasmid as described in the previous paragraph was used for live visualization of liver stage parasites including hypnozoites. Using the GFP-marker expressed from this plasmid, hypnozoite- and schizont-infected hepatocytes could be separated from uninfected hepatocytes by fluorescence-activated cell sorting (FACS) [40] and subjected to transcriptomics. This enabled analysis of host cells of infected versus uninfected cells [96] and a bulk transcriptome analysis of P. cynomolgi liver stages isolated at different timepoints after sporozoite infection [37,39]. It was shown that while hypnozoites progressively decrease transcription, processes involved in quiescence, energy metabolism and maintenance of genome integrity are maintained [37]. These datasets also indicated a massive upregulation of Lisp2 in liver stage schizonts compared to hypnozoites and a follow up study identified this protein as an early marker of liver stage growth [83]. Based on this differential expression pattern of Lisp2, P. cynomolgi dual marker parasites were developed [41,42].
The dual fluorescent P. cynomolgi line that included GFP driven by the constitutive promoter hsp70 and lisp2-driven mCherry provided the unique opportunity to live monitor the development of liver stage parasites including hypnozoites and to follow hypnozoite activation directly and unequivocally in the absence of drug treatment (that would otherwise have been needed to distinguish aborted liver stage schizonts from activated hypnozoites). Through prolonged monitoring of the dual fluorescent P. cynomolgi (up until day 22 post infection of the hepatocyte), almost 40 years after their discovery, it was for the first time formally demonstrated that that hypnozoites can spontaneously activate and resume development after the primary liver stage schizonts have completed development and ruptured [41].
Similarly, a dual bioluminescent parasite line was developed in which the bright Nanoluc luciferase [97] was constitutively expressed under control of the hsp70 promoter and Firefly luciferase was driven by the liver schizont-specific lisp2 promoter [83]. This enabled a robust, ultrafast, and sensitive detection of hypnozoites through enzymatic detection of bioluminescence [42]. This may enable the development of bioluminescence-based high-throughput systems for screening compounds with anti-hypnozoite activity.
To increase our knowledge of mechanisms underlying hypnozoite dormancy, the dual fluorescent P. cynomolgi parasite line described above was used for GFP-fluorescence based single cell FACS isolation of liver stage parasites at early (day 2-6) stages of parasite development [38]. Subsequent single cell RNA-sequencing analysis of the sorted parasites revealed various clusters of parasite populations with different expression patterns. In the putative ‘hypnozoite’ clusters, several genes were identified to be specifically transcribed in hypnozoites. This was validated by RNA-FISH. Strikingly, Chip-seq of sporozoites and blood stages, and ATAC-seq of hypnozoites versus liver schizonts indicated differences in chromatin environment between these parasite forms. Given that the identified hypnozoite-specific transcripts encode putative RNA-binding proteins, it was hypothesized that these epigenetically regulated RNA-binding genes may underly hypnozoite formation and activation processes [38]. This hypothesis needs to be validated by functional studies, which requires sophisticated (conditional) knock-out systems as have been developed for other Plasmodium species [98,99,100], but not yet for P. cynomolgi vivo parasites.
3.3.3. P. cynomolgi In Vitro Genetic Engineering Technology
For P. cynomolgi, an in vitro blood stage culture system has been developed [101,102]. In this system the P. cynomolgi Berok line was adapted to in vitro culture in macaque red blood cells. Recently, this in vitro line was successfully used to edit the putative drug resistance marker MDR1 Y976F using CRISPR-Cas9 [35]. This shows the important proof-of-concept that transfection of the P. cynomolgi vitro line is feasible. However, the authors noted that the methodology still requires optimization, as the editing of Pcymdr1 Y976F was the only successful editing event from a range of other orthologous markers of drug resistance after two years of experimentation.
It is not known whether the in vitro cultured P. cynomolgi line can be directly used for liver stage research. Successful transmission of the in vitro P. cynomolgi Berok line has been shown following an in vivo passage through a monkey [102], but transmission without a monkey passage has not yet been shown (Figure 2). However, the fact that monkey-passaged in vitro P. cynomolgi Berok does give rise to infectious sporozoites demonstrates that the capacity of the vitro line to develop into sexual stages has not been lost.
3.4. P. cynomolgi Hypnozoite Research Benefits and Limitations
Findings obtained with P. cynomolgi as model for P. vivax will always require validation in P. vivax itself. This is especially important in the light of the different relapse characteristics known to exist for P. vivax [14]. P. cynomolgi B/M has short latency characteristics and can thus serve as a model for fast relapsing P. vivax isolates. It is not known whether findings for this frequently relapsing P. cynomolgi can serve as a valid model for the P. vivax long latency lines, and it is unknown whether long-latency P. cynomolgi lines exist. Another limitation of the P. cynomolgi system is that it depends on NHPs for transmission and liver stage research. Genetic engineering of the P. cynomolgi vitro parasite may overcome this [35,102], but transmission without monkey passage has not been reported yet and the blood stage vitro culture still requires macaque red blood cells. Besides these limitations, the P. cynomolgi models provides a unique opportunity: a combined use of robust transfection methods and liver stage cultures enables transgene expression in liver stage cultures, including hypnozoites [37,38,39,40,41,42] (Figure 2). Further development of this technology to include more sophisticated genetic engineering tools (e.g. DiCre [98] and Crispr/Cas9 [103] technologies) as have been described for other Plasmodium species is warranted to enable more detailed investigations into on mechanisms underlying hypnozoite dormancy and activation.
4. Conclusions and outlook
Functional analysis of genes involved in hypnozoite biology is complicated. Current model systems each have their benefits and limitations. Robust liver stage culture platforms enabling the study of P. vivax hypnozoites are in place, but in the absence of a blood stage culture system rely on patient or NHP derived parasites. Transfection of P. vivax is cumbersome and has not been applied to liver stage research. P. cynomolgi is an excellent model for P. vivax. As with any model system biological findings in the model need validation in P. vivax, and it depends on NHPs, but robust liver stage culture systems including hypnozoites are available. Combined with the capacity to genetically manipulate this species, P. cynomolgi is currently the only parasite for which genetic engineering has been applied to study hypnozoite biology.
Through synergistically exploiting the strengths of each model, the complexities associated with P. vivax-type hypnozoite research may be overcome. A variety of tools for the genetic engineering of another closely related NHP parasite P. knowlesi is available [104,105,106,107,108,109,110,111,112]. However, this parasite species does not form hypnozoites. Nonetheless, it can be envisaged for example that P. knowlesi can be used to optimize transfection conditions and to validate genes potentially involved in hypnozoite development (i.e. overexpression of a candidate gene may induce hypnozoite-like features in P. knowlesi liver stage parasites). P. cynomolgi genetic engineering enables direct manipulation and investigation in candidate genes involved in hypnozoite biology. Validation of findings may eventually be performed in P. vivax. Making optimal use of available and improved model systems, may finally provide new insights in the cryptic hypnozoite biology.
Author Contributions
A.V.-v.d.W. wrote the manuscript. A-M.Z. and C.H.M.K. reviewed and edited the manuscript.
Funding
This research received no external funding.
Acknowledgments
We would like to acknowledge Francisca van Hassel and the Biorender platform for figure design.
Conflicts of Interest
The authors declare no conflict of interest.
References
- (WHO), W.H.O. World malaria report 2022.
- Battle, K.E.; Lucas, T.C.D.; Nguyen, M.; Howes, R.E.; Nandi, A.K.; Twohig, K.A.; Pfeffer, D.A.; Cameron, E.; Rao, P.C.; Casey, D.; et al. Mapping the global endemicity and clinical burden of Plasmodium vivax, 2000-17: a spatial and temporal modelling study. Lancet 2019, 394, 332-343. [CrossRef]
- Baird, J.K. African Plasmodium vivax malaria improbably rare or benign. Trends Parasitol 2022, 38, 683-696. [CrossRef]
- Quaye, I.K.; Aleksenko, L.; Oeuvray, C.; Yewhalaw, D.; Duah, N.; Gyan, B.; Haiyambo, D.H.; Dongho, G.B.D.; Torgby, R.A.; Amoah, L.; et al. The Pan African Vivax and Ovale Network (PAVON): Refocusing on Plasmodium vivax, ovale and asymptomatic malaria in sub-Saharan Africa. Parasitol Int 2021, 84, 102415. [CrossRef]
- Zimmerman, P.A. Plasmodium vivax Infection in Duffy-Negative People in Africa. Am J Trop Med Hyg 2017, 97, 636-638. [CrossRef]
- Baird, J.K. Evidence and implications of mortality associated with acute Plasmodium vivax malaria. Clin Microbiol Rev 2013, 26, 36-57. [CrossRef]
- Price, R.N.; Tjitra, E.; Guerra, C.A.; Yeung, S.; White, N.J.; Anstey, N.M. Vivax malaria: neglected and not benign. Am J Trop Med Hyg 2007, 77, 79-87.
- Phyo, A.P.; Dahal, P.; Mayxay, M.; Ashley, E.A. Clinical impact of vivax malaria: A collection review. PLoS Med 2022, 19, e1003890. [CrossRef]
- Bantuchai, S.; Imad, H.; Nguitragool, W. Plasmodium vivax gametocytes and transmission. Parasitol Int 2022, 87, 102497. [CrossRef]
- Krotoski, W.A.; Collins, W.E.; Bray, R.S.; Garnham, P.C.; Cogswell, F.B.; Gwadz, R.W.; Killick-Kendrick, R.; Wolf, R.; Sinden, R.; Koontz, L.C.; et al. Demonstration of hypnozoites in sporozoite-transmitted Plasmodium vivax infection. Am J Trop Med Hyg 1982, 31, 1291-1293. [CrossRef]
- Hemmer, C.J.; Holst, F.G.; Kern, P.; Chiwakata, C.B.; Dietrich, M.; Reisinger, E.C. Stronger host response per parasitized erythrocyte in Plasmodium vivax or ovale than in Plasmodium falciparum malaria. Trop Med Int Health 2006, 11, 817-823. [CrossRef]
- Moreno-Perez, D.A.; Ruiz, J.A.; Patarroyo, M.A. Reticulocytes: Plasmodium vivax target cells. Biol Cell 2013, 105, 251-260. [CrossRef]
- White, N.J. Why Do Some Primate Malarias Relapse? Trends Parasitol 2016, 32, 918-920. [CrossRef]
- White, N.J. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar J 2011, 10, 297. [CrossRef]
- Baird, J.K.; Battle, K.E.; Howes, R.E. Primaquine ineligibility in anti-relapse therapy of Plasmodium vivax malaria: the problem of G6PD deficiency and cytochrome P-450 2D6 polymorphisms. Malar J 2018, 17, 42. [CrossRef]
- Cox, F.E. History of the discovery of the malaria parasites and their vectors. Parasit Vectors 2010, 3, 5. [CrossRef]
- Cogswell, F.B.; Krotoski, W.A.; Hollingdale, M.R.; Gwadz, R.W. Identification of hypnozoites and tissue schizonts of Plasmodium vivax and P. cynomolgi by the immunoperoxidase method. Am J Trop Med Hyg 1983, 32, 1454-1455. [CrossRef]
- Lover, A.A.; Baird, J.K.; Gosling, R.; Price, R.N. Malaria Elimination: Time to Target All Species. Am J Trop Med Hyg 2018, 99, 17-23. [CrossRef]
- Organization, W.H. Global technical strategy for malaria 2016-2030. . 2015.
- van Dijk, M.R.; Waters, A.P.; Janse, C.J. Stable transfection of malaria parasite blood stages. Science 1995, 268, 1358-1362. [CrossRef]
- Wu, Y.; Sifri, C.D.; Lei, H.H.; Su, X.Z.; Wellems, T.E. Transfection of Plasmodium falciparum within human red blood cells. Proc Natl Acad Sci U S A 1995, 92, 973-977. [CrossRef]
- Kafsack, B.F.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014, 507, 248-252. [CrossRef]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 2014, 507, 253-257. [CrossRef]
- Cowman, A.F.; Baldi, D.L.; Healer, J.; Mills, K.E.; O'Donnell, R.A.; Reed, M.B.; Triglia, T.; Wickham, M.E.; Crabb, B.S. Functional analysis of proteins involved in Plasmodium falciparum merozoite invasion of red blood cells. FEBS Lett 2000, 476, 84-88. [CrossRef]
- Murphy, S.C.; Vaughan, A.M.; Kublin, J.G.; Fishbauger, M.; Seilie, A.M.; Cruz, K.P.; Mankowski, T.; Firat, M.; Magee, S.; Betz, W.; et al. A genetically engineered Plasmodium falciparum parasite vaccine provides protection from controlled human malaria infection. Sci Transl Med 2022, 14, eabn9709. [CrossRef]
- Roestenberg, M.; Walk, J.; van der Boor, S.C.; Langenberg, M.C.C.; Hoogerwerf, M.A.; Janse, J.J.; Manurung, M.; Yap, X.Z.; Garcia, A.F.; Koopman, J.P.R.; et al. A double-blind, placebo-controlled phase 1/2a trial of the genetically attenuated malaria vaccine PfSPZ-GA1. Sci Transl Med 2020, 12. [CrossRef]
- Swann, J.; Corey, V.; Scherer, C.A.; Kato, N.; Comer, E.; Maetani, M.; Antonova-Koch, Y.; Reimer, C.; Gagaring, K.; Ibanez, M.; et al. High-Throughput Luciferase-Based Assay for the Discovery of Therapeutics That Prevent Malaria. ACS Infect Dis 2016, 2, 281-293. [CrossRef]
- Jongco, A.M.; Ting, L.M.; Thathy, V.; Mota, M.M.; Kim, K. Improved transfection and new selectable markers for the rodent malaria parasite Plasmodium yoelii. Mol Biochem Parasitol 2006, 146, 242-250. [CrossRef]
- Kocken, C.H.; van der Wel, A.; Thomas, A.W. Plasmodium cynomolgi: transfection of blood-stage parasites using heterologous DNA constructs. Exp Parasitol 1999, 93, 58-60. [CrossRef]
- Reece, S.E.; Thompson, J. Transformation of the rodent malaria parasite Plasmodium chabaudi and generation of a stable fluorescent line PcGFPCON. Malar J 2008, 7, 183. [CrossRef]
- van der Wel, A.M.; Tomas, A.M.; Kocken, C.H.; Malhotra, P.; Janse, C.J.; Waters, A.P.; Thomas, A.W. Transfection of the primate malaria parasite Plasmodium knowlesi using entirely heterologous constructs. J Exp Med 1997, 185, 1499-1503. [CrossRef]
- Moraes Barros, R.R.; Straimer, J.; Sa, J.M.; Salzman, R.E.; Melendez-Muniz, V.A.; Mu, J.; Fidock, D.A.; Wellems, T.E. Editing the Plasmodium vivax genome, using zinc-finger nucleases. J Infect Dis 2015, 211, 125-129. [CrossRef]
- Pfahler, J.M.; Galinski, M.R.; Barnwell, J.W.; Lanzer, M. Transient transfection of Plasmodium vivax blood stage parasites. Mol Biochem Parasitol 2006, 149, 99-101. [CrossRef]
- Kocken, C.H.; Ozwara, H.; van der Wel, A.; Beetsma, A.L.; Mwenda, J.M.; Thomas, A.W. Plasmodium knowlesi provides a rapid in vitro and in vivo transfection system that enables double-crossover gene knockout studies. Infect Immun 2002, 70, 655-660. [CrossRef]
- Ward, K.E.; Christensen, P.; Racklyeft, A.; Dhingra, S.K.; Chua, A.C.Y.; Remmert, C.; Suwanarusk, R.; Matheson, J.; Blackman, M.J.; Kaneko, O.; et al. Integrative genetic manipulation of Plasmodium cynomolgi reveals MultiDrug Resistance-1 Y976F associated with increased in vitro susceptibility to mefloquine. J Infect Dis 2022. [CrossRef]
- Ngotho, P.; Soares, A.B.; Hentzschel, F.; Achcar, F.; Bertuccini, L.; Marti, M. Revisiting gametocyte biology in malaria parasites. FEMS Microbiol Rev 2019, 43, 401-414. [CrossRef]
- Bertschi, N.L.; Voorberg-van der Wel, A.; Zeeman, A.M.; Schuierer, S.; Nigsch, F.; Carbone, W.; Knehr, J.; Gupta, D.K.; Hofman, S.O.; van der Werff, N.; et al. Transcriptomic analysis reveals reduced transcriptional activity in the malaria parasite Plasmodium cynomolgi during progression into dormancy. Elife 2018, 7. [CrossRef]
- Toenhake, C.G.; Voorberg-van der Wel, A.; Wu, H.; Kanyal, A.; Nieuwenhuis, I.G.; van der Werff, N.M.; Hofman, S.O.; Zeeman, A.M.; Kocken, C.H.M.; Bartfai, R. Epigenetically regulated RNA-binding proteins signify malaria hypnozoite dormancy. Cell Rep 2023, 42, 112727. [CrossRef]
- Voorberg-van der Wel, A.; Roma, G.; Gupta, D.K.; Schuierer, S.; Nigsch, F.; Carbone, W.; Zeeman, A.M.; Lee, B.H.; Hofman, S.O.; Faber, B.W.; et al. A comparative transcriptomic analysis of replicating and dormant liver stages of the relapsing malaria parasite Plasmodium cynomolgi. Elife 2017, 6. [CrossRef]
- Voorberg-van der Wel, A.; Zeeman, A.M.; van Amsterdam, S.M.; van den Berg, A.; Klooster, E.J.; Iwanaga, S.; Janse, C.J.; van Gemert, G.J.; Sauerwein, R.; Beenhakker, N.; et al. Transgenic fluorescent Plasmodium cynomolgi liver stages enable live imaging and purification of Malaria hypnozoite-forms. PLoS One 2013, 8, e54888. [CrossRef]
- Voorberg-van der Wel, A.M.; Zeeman, A.M.; Nieuwenhuis, I.G.; van der Werff, N.M.; Klooster, E.J.; Klop, O.; Vermaat, L.C.; Kumar Gupta, D.; Dembele, L.; Diagana, T.T.; et al. A dual fluorescent Plasmodium cynomolgi reporter line reveals in vitro malaria hypnozoite reactivation. Commun Biol 2020, 3, 7. [CrossRef]
- Voorberg-van der Wel, A.M.; Zeeman, A.M.; Nieuwenhuis, I.G.; van der Werff, N.M.; Kocken, C.H.M. Dual-Luciferase-Based Fast and Sensitive Detection of Malaria Hypnozoites for the Discovery of Anti-relapse Compounds. Methods Mol Biol 2022, 2524, 397-408. [CrossRef]
- Coatney G.R., C.W.E., Warren M., and Contacos P. G. . The primate malarias [original book published 1971].
- Joyner, C.; Barnwell, J.W.; Galinski, M.R. No more monkeying around: primate malaria model systems are key to understanding Plasmodium vivax liver-stage biology, hypnozoites, and relapses. Front Microbiol 2015, 6, 145. [CrossRef]
- Mikolajczak, S.A.; Vaughan, A.M.; Kangwanrangsan, N.; Roobsoong, W.; Fishbaugher, M.; Yimamnuaychok, N.; Rezakhani, N.; Lakshmanan, V.; Singh, N.; Kaushansky, A.; et al. Plasmodium vivax liver stage development and hypnozoite persistence in human liver-chimeric mice. Cell Host Microbe 2015, 17, 526-535. [CrossRef]
- Schafer, C.; Dambrauskas, N.; Reynolds, L.M.; Trakhimets, O.; Raappana, A.; Flannery, E.L.; Roobsoong, W.; Sattabongkot, J.; Mikolajczak, S.A.; Kappe, S.H.I.; et al. Partial protection against P. vivax infection diminishes hypnozoite burden and blood-stage relapses. Cell Host Microbe 2021, 29, 752-756 e754. [CrossRef]
- Flannery, E.L.; Kangwanrangsan, N.; Chuenchob, V.; Roobsoong, W.; Fishbaugher, M.; Zhou, K.; Billman, Z.P.; Martinson, T.; Olsen, T.M.; Schafer, C.; et al. Plasmodium vivax latent liver infection is characterized by persistent hypnozoites, hypnozoite-derived schizonts, and time-dependent efficacy of primaquine. Mol Ther Methods Clin Dev 2022, 26, 427-440. [CrossRef]
- Bermudez, M.; Moreno-Perez, D.A.; Arevalo-Pinzon, G.; Curtidor, H.; Patarroyo, M.A. Plasmodium vivax in vitro continuous culture: the spoke in the wheel. Malar J 2018, 17, 301. [CrossRef]
- Thomson-Luque, R.; Adams, J.H.; Kocken, C.H.M.; Pasini, E.M. From marginal to essential: the golden thread between nutrient sensing, medium composition and Plasmodium vivax maturation in in vitro culture. Malar J 2019, 18, 344. [CrossRef]
- Valenciano, A.L.; Gomez-Lorenzo, M.G.; Vega-Rodriguez, J.; Adams, J.H.; Roth, A. In vitro models for human malaria: targeting the liver stage. Trends Parasitol 2022, 38, 758-774. [CrossRef]
- Chattopadhyay, R.; Velmurugan, S.; Chakiath, C.; Andrews Donkor, L.; Milhous, W.; Barnwell, J.W.; Collins, W.E.; Hoffman, S.L. Establishment of an in vitro assay for assessing the effects of drugs on the liver stages of Plasmodium vivax malaria. PLoS One 2010, 5, e14275. [CrossRef]
- Hollingdale, M.R.; Collins, W.E.; Campbell, C.C.; Schwartz, A.L. In vitro culture of two populations (dividing and nondividing) of exoerythrocytic parasites of Plasmodium vivax. Am J Trop Med Hyg 1985, 34, 216-222. [CrossRef]
- Sattabongkot, J.; Yimamnuaychoke, N.; Leelaudomlipi, S.; Rasameesoraj, M.; Jenwithisuk, R.; Coleman, R.E.; Udomsangpetch, R.; Cui, L.; Brewer, T.G. Establishment of a human hepatocyte line that supports in vitro development of the exo-erythrocytic stages of the malaria parasites Plasmodium falciparum and P. vivax. Am J Trop Med Hyg 2006, 74, 708-715.
- Pewkliang, Y.; Rungin, S.; Lerdpanyangam, K.; Duangmanee, A.; Kanjanasirirat, P.; Suthivanich, P.; Sa-Ngiamsuntorn, K.; Borwornpinyo, S.; Sattabongkot, J.; Patrapuvich, R.; et al. A novel immortalized hepatocyte-like cell line (imHC) supports in vitro liver stage development of the human malarial parasite Plasmodium vivax. Malar J 2018, 17, 50. [CrossRef]
- Ng, S.; Schwartz, R.E.; March, S.; Galstian, A.; Gural, N.; Shan, J.; Prabhu, M.; Mota, M.M.; Bhatia, S.N. Human iPSC-derived hepatocyte-like cells support Plasmodium liver-stage infection in vitro. Stem Cell Reports 2015, 4, 348-359. [CrossRef]
- Gural, N.; Mancio-Silva, L.; Miller, A.B.; Galstian, A.; Butty, V.L.; Levine, S.S.; Patrapuvich, R.; Desai, S.P.; Mikolajczak, S.A.; Kappe, S.H.I.; et al. In Vitro Culture, Drug Sensitivity, and Transcriptome of Plasmodium Vivax Hypnozoites. Cell Host Microbe 2018, 23, 395-406 e394. [CrossRef]
- March, S.; Ng, S.; Velmurugan, S.; Galstian, A.; Shan, J.; Logan, D.J.; Carpenter, A.E.; Thomas, D.; Sim, B.K.; Mota, M.M.; et al. A microscale human liver platform that supports the hepatic stages of Plasmodium falciparum and vivax. Cell Host Microbe 2013, 14, 104-115. [CrossRef]
- Chua, A.C.Y.; Ananthanarayanan, A.; Ong, J.J.Y.; Wong, J.Y.; Yip, A.; Singh, N.H.; Qu, Y.; Dembele, L.; McMillian, M.; Ubalee, R.; et al. Hepatic spheroids used as an in vitro model to study malaria relapse. Biomaterials 2019, 216, 119221. [CrossRef]
- Mazier, D.; Landau, I.; Druilhe, P.; Miltgen, F.; Guguen-Guillouzo, C.; Baccam, D.; Baxter, J.; Chigot, J.P.; Gentilini, M. Cultivation of the liver forms of Plasmodium vivax in human hepatocytes. Nature 1984, 307, 367-369. [CrossRef]
- Roth, A.; Maher, S.P.; Conway, A.J.; Ubalee, R.; Chaumeau, V.; Andolina, C.; Kaba, S.A.; Vantaux, A.; Bakowski, M.A.; Thomson-Luque, R.; et al. A comprehensive model for assessment of liver stage therapies targeting Plasmodium vivax and Plasmodium falciparum. Nat Commun 2018, 9, 1837. [CrossRef]
- Carlton, J.M.; Adams, J.H.; Silva, J.C.; Bidwell, S.L.; Lorenzi, H.; Caler, E.; Crabtree, J.; Angiuoli, S.V.; Merino, E.F.; Amedeo, P.; et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax. Nature 2008, 455, 757-763. [CrossRef]
- Nurk, S.; Koren, S.; Rhie, A.; Rautiainen, M.; Bzikadze, A.V.; Mikheenko, A.; Vollger, M.R.; Altemose, N.; Uralsky, L.; Gershman, A.; et al. The complete sequence of a human genome. Science 2022, 376, 44-53. [CrossRef]
- Zeeman, A.M.; Lakshminarayana, S.B.; van der Werff, N.; Klooster, E.J.; Voorberg-van der Wel, A.; Kondreddi, R.R.; Bodenreider, C.; Simon, O.; Sauerwein, R.; Yeung, B.K.; et al. PI4 Kinase Is a Prophylactic but Not Radical Curative Target in Plasmodium vivax-Type Malaria Parasites. Antimicrob Agents Chemother 2016, 60, 2858-2863. [CrossRef]
- Mancio-Silva, L.; Gural, N.; Real, E.; Wadsworth, M.H., 2nd; Butty, V.L.; March, S.; Nerurkar, N.; Hughes, T.K.; Roobsoong, W.; Fleming, H.E.; et al. A single-cell liver atlas of Plasmodium vivax infection. Cell Host Microbe 2022, 30, 1048-1060 e1045. [CrossRef]
- Ruberto, A.A.; Maher, S.P.; Vantaux, A.; Joyner, C.J.; Bourke, C.; Balan, B.; Jex, A.; Mueller, I.; Witkowski, B.; Kyle, D.E. Single-cell RNA profiling of Plasmodium vivax-infected hepatocytes reveals parasite- and host- specific transcriptomic signatures and therapeutic targets. Front Cell Infect Microbiol 2022, 12, 986314. [CrossRef]
- Krotoski, W.A.; Bray, R.S.; Garnham, P.C.; Gwadz, R.W.; Killick-Kendrick, R.; Draper, C.C.; Targett, G.A.; Krotoski, D.M.; Guy, M.W.; Koontz, L.C.; et al. Observations on early and late post-sporozoite tissue stages in primate malaria. II. The hypnozoite of Plasmodium cynomolgi bastianellii from 3 to 105 days after infection, and detection of 36- to 40-hour pre-erythrocytic forms. Am J Trop Med Hyg 1982, 31, 211-225.
- Krotoski, W.A.; Garnham, P.C.; Bray, R.S.; Krotoski, D.M.; Killick-Kendrick, R.; Draper, C.C.; Targett, G.A.; Guy, M.W. Observations on early and late post-sporozoite tissue stages in primate malaria. I. Discovery of a new latent form of Plasmodium cynomolgi (the hypnozoite), and failure to detect hepatic forms within the first 24 hours after infection. Am J Trop Med Hyg 1982, 31, 24-35.
- Hawking, F.; Worms, M.J.; Gammage, K. 24- and 48-hour cycles of malaria parasites in the blood; their purpose, production and control. Trans R Soc Trop Med Hyg 1968, 62, 731-765. [CrossRef]
- Krotoski, W.A.; Krotoski, D.M.; Garnham, P.C.; Bray, R.S.; Killick-Kendrick, R.; Draper, C.C.; Targett, G.A.; Guy, M.W. Relapses in primate malaria: discovery of two populations of exoerythrocytic stages. Preliminary note. Br Med J 1980, 280, 153-154. [CrossRef]
- G.R., C. Reminiscences: My Forty-Year Romance with Malaria. Transactions of the Nebraska Academy of Sciences and Affiliated Societies 1985, 222.
- Corcoran, K.D.; Hansukjariya, P.; Sattabongkot, J.; Ngampochjana, M.; Edstein, M.D.; Smith, C.D.; Shanks, G.D.; Milhous, W.K. Causal prophylactic and radical curative activity of WR182393 (a guanylhydrazone) against Plasmodium cynomolgi in Macaca mulatta. Am J Trop Med Hyg 1993, 49, 473-477. [CrossRef]
- Davidson, D.E., Jr.; Ager, A.L.; Brown, J.L.; Chapple, F.E.; Whitmire, R.E.; Rossan, R.N. New tissue schizontocidal antimalarial drugs. Bull World Health Organ 1981, 59, 463-479.
- Deye, G.A.; Gettayacamin, M.; Hansukjariya, P.; Im-erbsin, R.; Sattabongkot, J.; Rothstein, Y.; Macareo, L.; Fracisco, S.; Bennett, K.; Magill, A.J.; et al. Use of a rhesus Plasmodium cynomolgi model to screen for anti-hypnozoite activity of pharmaceutical substances. Am J Trop Med Hyg 2012, 86, 931-935. [CrossRef]
- Dow, G.S.; Gettayacamin, M.; Hansukjariya, P.; Imerbsin, R.; Komcharoen, S.; Sattabongkot, J.; Kyle, D.; Milhous, W.; Cozens, S.; Kenworthy, D.; et al. Radical curative efficacy of tafenoquine combination regimens in Plasmodium cynomolgi-infected Rhesus monkeys (Macaca mulatta). Malar J 2011, 10, 212. [CrossRef]
- Dutta, G.P.; Puri, S.K.; Bhaduri, A.P.; Seth, M. Radical curative activity of a new 8-aminoquinoline derivative (CDRI 80/53) against Plasmodium cynomolgi B in monkeys. Am J Trop Med Hyg 1989, 41, 635-637. [CrossRef]
- Schmidt, L.H. Appraisals of compounds of diverse chemical classes for capacities to cure infections with sporozoites of Plasmodium cynomolgi. Am J Trop Med Hyg 1983, 32, 231-257. [CrossRef]
- Schmidt, L.H.; Fradkin, R.; Genther, C.S.; Rossan, R.N.; Squires, W. Responses of Sporozoite-Induced and Trophozoite-Induced Infections to Standard Antimalarial Drugs. Am J Trop Med Hyg 1982, 31, 646-665.
- Sodeman, T.M.; Contacos, P.G.; Collins, W.E.; Smith, C.S.; Jumper, J.R. Studies on the prophylactic and radical curative activity of RC-12 against Plasmodium cynomolgi in Macaca mulatta. Bull World Health Organ 1972, 47, 425-428.
- Millet, P.; Fisk, T.L.; Collins, W.E.; Broderson, J.R.; Nguyen-Dinh, P. Cultivation of exoerythrocytic stages of Plasmodium cynomolgi, P. knowlesi, P. coatneyi, and P. inui in Macaca mulatta hepatocytes. Am J Trop Med Hyg 1988, 39, 529-534. [CrossRef]
- Dembele, L.; Gego, A.; Zeeman, A.M.; Franetich, J.F.; Silvie, O.; Rametti, A.; Le Grand, R.; Dereuddre-Bosquet, N.; Sauerwein, R.; van Gemert, G.J.; et al. Towards an in vitro model of Plasmodium hypnozoites suitable for drug discovery. PLoS One 2011, 6, e18162. [CrossRef]
- Zeeman, A.M.; van Amsterdam, S.M.; McNamara, C.W.; Voorberg-van der Wel, A.; Klooster, E.J.; van den Berg, A.; Remarque, E.J.; Plouffe, D.M.; van Gemert, G.J.; Luty, A.; et al. KAI407, a potent non-8-aminoquinoline compound that kills Plasmodium cynomolgi early dormant liver stage parasites in vitro. Antimicrob Agents Chemother 2014, 58, 1586-1595. [CrossRef]
- Ashraf, K.; Tajeri, S.; Arnold, C.S.; Amanzougaghene, N.; Franetich, J.F.; Vantaux, A.; Soulard, V.; Bordessoulles, M.; Cazals, G.; Bousema, T.; et al. Artemisinin-independent inhibitory activity of Artemisia sp. infusions against different Plasmodium stages including relapse-causing hypnozoites. Life Sci Alliance 2022, 5. [CrossRef]
- Gupta, D.K.; Dembele, L.; Voorberg-van der Wel, A.; Roma, G.; Yip, A.; Chuenchob, V.; Kangwanrangsan, N.; Ishino, T.; Vaughan, A.M.; Kappe, S.H.; et al. The Plasmodium liver-specific protein 2 (LISP2) is an early marker of liver stage development. Elife 2019, 8. [CrossRef]
- Sutton, P.L.; Luo, Z.; Divis, P.C.S.; Friedrich, V.K.; Conway, D.J.; Singh, B.; Barnwell, J.W.; Carlton, J.M.; Sullivan, S.A. Characterizing the genetic diversity of the monkey malaria parasite Plasmodium cynomolgi. Infect Genet Evol 2016, 40, 243-252. [CrossRef]
- Dembele, L.; Franetich, J.F.; Lorthiois, A.; Gego, A.; Zeeman, A.M.; Kocken, C.H.; Le Grand, R.; Dereuddre-Bosquet, N.; van Gemert, G.J.; Sauerwein, R.; et al. Persistence and activation of malaria hypnozoites in long-term primary hepatocyte cultures. Nat Med 2014, 20, 307-312. [CrossRef]
- Cubi, R.; Vembar, S.S.; Biton, A.; Franetich, J.F.; Bordessoulles, M.; Sossau, D.; Zanghi, G.; Bosson-Vanga, H.; Benard, M.; Moreno, A.; et al. Laser capture microdissection enables transcriptomic analysis of dividing and quiescent liver stages of Plasmodium relapsing species. Cell Microbiol 2017, 19. [CrossRef]
- Tachibana, S.; Kawai, S.; Katakai, Y.; Takahashi, H.; Nakade, T.; Yasutomi, Y.; Horii, T.; Tanabe, K. Contrasting infection susceptibility of the Japanese macaques and cynomolgus macaques to closely related malaria parasites, Plasmodium vivax and Plasmodium cynomolgi. Parasitol Int 2015, 64, 274-281. [CrossRef]
- Pasini, E.M.; Bohme, U.; Rutledge, G.G.; Voorberg-Van der Wel, A.; Sanders, M.; Berriman, M.; Kocken, C.H.; Otto, T.D. An improved Plasmodium cynomolgi genome assembly reveals an unexpected methyltransferase gene expansion. Wellcome Open Res 2017, 2, 42. [CrossRef]
- Tachibana, S.; Sullivan, S.A.; Kawai, S.; Nakamura, S.; Kim, H.R.; Goto, N.; Arisue, N.; Palacpac, N.M.; Honma, H.; Yagi, M.; et al. Plasmodium cynomolgi genome sequences provide insight into Plasmodium vivax and the monkey malaria clade. Nat Genet 2012, 44, 1051-1055. [CrossRef]
- Akinyi, S.; Hanssen, E.; Meyer, E.V.; Jiang, J.; Korir, C.C.; Singh, B.; Lapp, S.; Barnwell, J.W.; Tilley, L.; Galinski, M.R. A 95 kDa protein of Plasmodium vivax and P. cynomolgi visualized by three-dimensional tomography in the caveola-vesicle complexes (Schuffner's dots) of infected erythrocytes is a member of the PHIST family. Mol Microbiol 2012, 84, 816-831. [CrossRef]
- Barale, J.C.; Menard, R. Centromeric plasmids and artificial chromosomes: new kids on the Plasmodium transfection block. Cell Host Microbe 2010, 7, 181-183. [CrossRef]
- Manzoni, G.; Briquet, S.; Risco-Castillo, V.; Gaultier, C.; Topcu, S.; Ivanescu, M.L.; Franetich, J.F.; Hoareau-Coudert, B.; Mazier, D.; Silvie, O. A rapid and robust selection procedure for generating drug-selectable marker-free recombinant malaria parasites. Sci Rep 2014, 4, 4760. [CrossRef]
- van Schaijk, B.C.; van Dijk, M.R.; van de Vegte-Bolmer, M.; van Gemert, G.J.; van Dooren, M.W.; Eksi, S.; Roeffen, W.F.; Janse, C.J.; Waters, A.P.; Sauerwein, R.W. Pfs47, paralog of the male fertility factor Pfs48/45, is a female specific surface protein in Plasmodium falciparum. Mol Biochem Parasitol 2006, 149, 216-222. [CrossRef]
- Iwanaga, S.; Kato, T.; Kaneko, I.; Yuda, M. Centromere plasmid: a new genetic tool for the study of Plasmodium falciparum. PLoS One 2012, 7, e33326. [CrossRef]
- Iwanaga, S.; Khan, S.M.; Kaneko, I.; Christodoulou, Z.; Newbold, C.; Yuda, M.; Janse, C.J.; Waters, A.P. Functional identification of the Plasmodium centromere and generation of a Plasmodium artificial chromosome. Cell Host Microbe 2010, 7, 245-255. [CrossRef]
- Mitchell, G.; Roma, G.; Voorberg-van der Wel, A.; Beibel, M.; Zeeman, A.M.; Schuierer, S.; Torres, L.; Flannery, E.L.; Kocken, C.H.M.; Mikolajczak, S.A.; et al. Transcriptional profiling of hepatocytes infected with the replicative form of the malaria parasite Plasmodium cynomolgi. Malar J 2022, 21, 393. [CrossRef]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem Biol 2012, 7, 1848-1857. [CrossRef]
- Collins, C.R.; Das, S.; Wong, E.H.; Andenmatten, N.; Stallmach, R.; Hackett, F.; Herman, J.P.; Muller, S.; Meissner, M.; Blackman, M.J. Robust inducible Cre recombinase activity in the human malaria parasite Plasmodium falciparum enables efficient gene deletion within a single asexual erythrocytic growth cycle. Mol Microbiol 2013, 88, 687-701. [CrossRef]
- Fernandes, P.; Briquet, S.; Patarot, D.; Loubens, M.; Hoareau-Coudert, B.; Silvie, O. The dimerisable Cre recombinase allows conditional genome editing in the mosquito stages of Plasmodium berghei. PLoS One 2020, 15, e0236616. [CrossRef]
- Knuepfer, E.; Napiorkowska, M.; van Ooij, C.; Holder, A.A. Generating conditional gene knockouts in Plasmodium - a toolkit to produce stable DiCre recombinase-expressing parasite lines using CRISPR/Cas9. Sci Rep 2017, 7, 3881. [CrossRef]
- Christensen, P.; Racklyeft, A.; Ward, K.E.; Matheson, J.; Suwanarusk, R.; Chua, A.C.Y.; Kaneko, O.; Aung, H.L.; Renia, L.; Amanzougaghene, N.; et al. Improving in vitro continuous cultivation of Plasmodium cynomolgi, a model for P. vivax. Parasitol Int 2022, 89, 102589. [CrossRef]
- Chua, A.C.Y.; Ong, J.J.Y.; Malleret, B.; Suwanarusk, R.; Kosaisavee, V.; Zeeman, A.M.; Cooper, C.A.; Tan, K.S.W.; Zhang, R.; Tan, B.H.; et al. Robust continuous in vitro culture of the Plasmodium cynomolgi erythrocytic stages. Nat Commun 2019, 10, 3635. [CrossRef]
- Ghorbal, M.; Gorman, M.; Macpherson, C.R.; Martins, R.M.; Scherf, A.; Lopez-Rubio, J.J. Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR-Cas9 system. Nat Biotechnol 2014, 32, 819-821. [CrossRef]
- Carrasquilla, M.; Adjalley, S.; Sanderson, T.; Marin-Menendez, A.; Coyle, R.; Montandon, R.; Rayner, J.C.; Pance, A.; Lee, M.C.S. Defining multiplicity of vector uptake in transfected Plasmodium parasites. Sci Rep 2020, 10, 10894. [CrossRef]
- Knuepfer, E.; Wright, K.E.; Kumar Prajapati, S.; Rawlinson, T.A.; Mohring, F.; Koch, M.; Lyth, O.R.; Howell, S.A.; Villasis, E.; Snijders, A.P.; et al. Divergent roles for the RH5 complex components, CyRPA and RIPR in human-infective malaria parasites. PLoS Pathog 2019, 15, e1007809. [CrossRef]
- Kocken, C.H.; Zeeman, A.M.; Voorberg-van der Wel, A.; Thomas, A.W. Transgenic Plasmodium knowlesi: relieving a bottleneck in malaria research? Trends Parasitol 2009, 25, 370-374. [CrossRef]
- Mohring, F.; Hart, M.N.; Patel, A.; Baker, D.A.; Moon, R.W. CRISPR-Cas9 Genome Editing of Plasmodium knowlesi. Bio Protoc 2020, 10, e3522. [CrossRef]
- Mohring, F.; Hart, M.N.; Rawlinson, T.A.; Henrici, R.; Charleston, J.A.; Diez Benavente, E.; Patel, A.; Hall, J.; Almond, N.; Campino, S.; et al. Rapid and iterative genome editing in the malaria parasite Plasmodium knowlesi provides new tools for P. vivax research. Elife 2019, 8. [CrossRef]
- Moon, R.W.; Hall, J.; Rangkuti, F.; Ho, Y.S.; Almond, N.; Mitchell, G.H.; Pain, A.; Holder, A.A.; Blackman, M.J. Adaptation of the genetically tractable malaria pathogen Plasmodium knowlesi to continuous culture in human erythrocytes. Proc Natl Acad Sci U S A 2013, 110, 531-536. [CrossRef]
- Moraes Barros, R.R.; Gibson, T.J.; Kite, W.A.; Sa, J.M.; Wellems, T.E. Comparison of two methods for transformation of Plasmodium knowlesi: Direct schizont electroporation and spontaneous plasmid uptake from plasmid-loaded red blood cells. Mol Biochem Parasitol 2017, 218, 16-22. [CrossRef]
- Moraes Barros, R.R.; Thawnashom, K.; Gibson, T.J.; Armistead, J.S.; Caleon, R.L.; Kaneko, M.; Kite, W.A.; Mershon, J.P.; Brockhurst, J.K.; Engels, T.; et al. Activity of Plasmodium vivax promoter elements in Plasmodium knowlesi, and a centromere-containing plasmid that expresses NanoLuc throughout the parasite life cycle. Malar J 2021, 20, 247. [CrossRef]
- Verzier, L.H.; Coyle, R.; Singh, S.; Sanderson, T.; Rayner, J.C. Plasmodium knowlesi as a model system for characterising Plasmodium vivax drug resistance candidate genes. PLoS Negl Trop Dis 2019, 13, e0007470. [CrossRef]
Figure 1.
P. cynomolgi in vivo and in vitro transfection capabilities. Transgenic parasites can be drug selected in vivo or in vitro. To date, successful transmission of P. cynomolgi has only been described following a monkey passage. Transgenic liver stage parasites, including hypnozoites (upper right-hand side, schematic drawing of a hepatocyte with GFP expressing hypnozoite) can be studied in vitro following sporozoite inoculation of cultured primary hepatoyctes.
Figure 1.
P. cynomolgi in vivo and in vitro transfection capabilities. Transgenic parasites can be drug selected in vivo or in vitro. To date, successful transmission of P. cynomolgi has only been described following a monkey passage. Transgenic liver stage parasites, including hypnozoites (upper right-hand side, schematic drawing of a hepatocyte with GFP expressing hypnozoite) can be studied in vitro following sporozoite inoculation of cultured primary hepatoyctes.
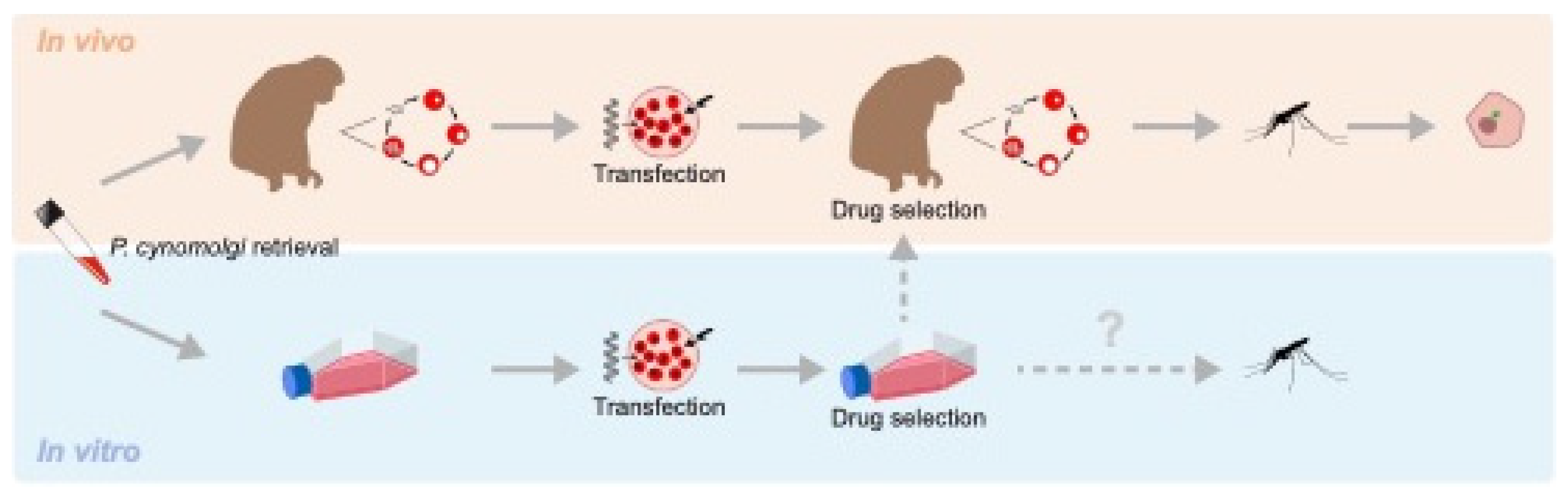
Figure 2.
Access to different stages of the life cycle of P. vivax-type parasites. Stages of the parasite life cycle for which the availability of transgenic parasites has been described are depicted in green. Question marks highlight the current lack of knowledge on the capacity for in vitro cultured parasites to be transmitted without passage through a monkey.
Figure 2.
Access to different stages of the life cycle of P. vivax-type parasites. Stages of the parasite life cycle for which the availability of transgenic parasites has been described are depicted in green. Question marks highlight the current lack of knowledge on the capacity for in vitro cultured parasites to be transmitted without passage through a monkey.
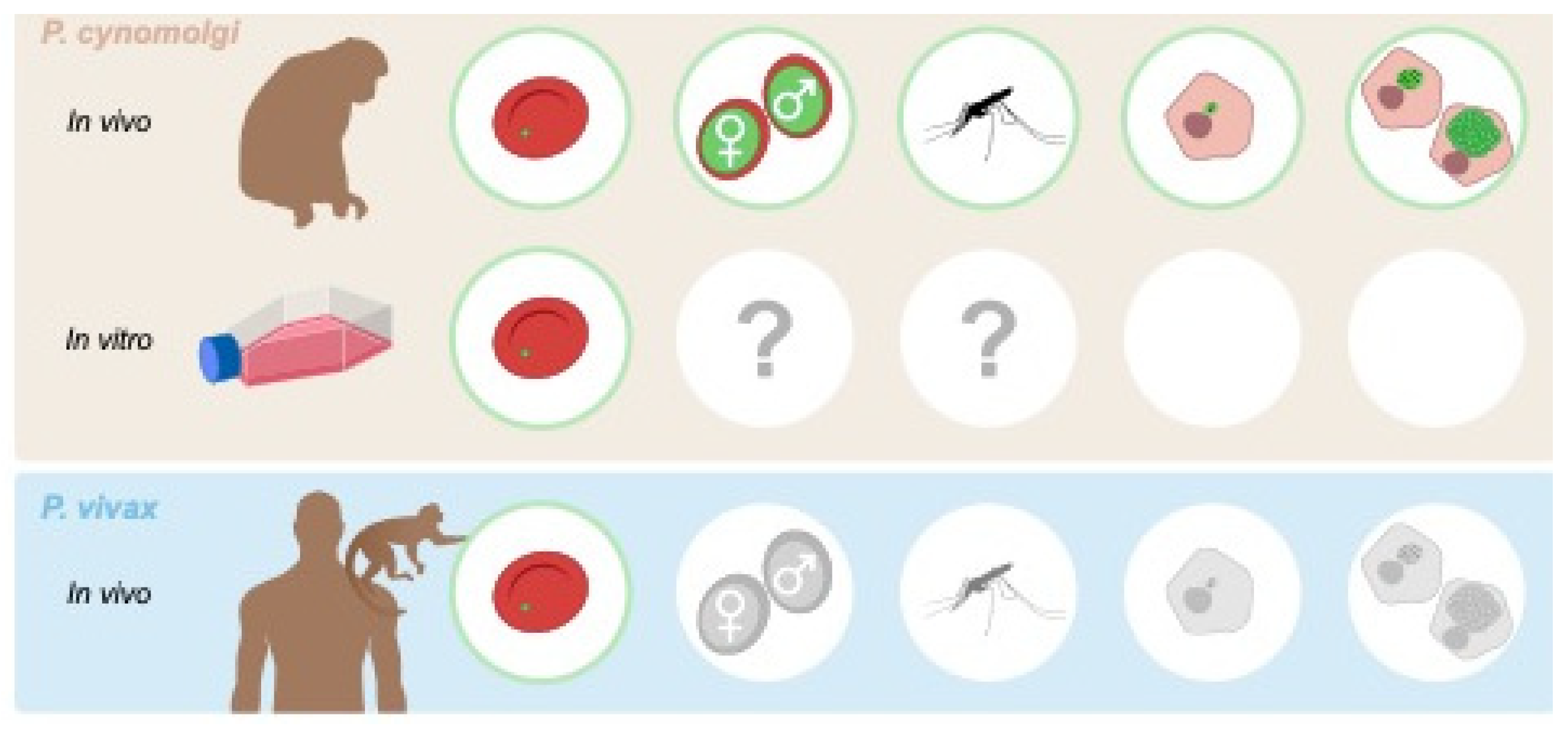

Table 1.
Genetic engineering tools developed for the study of P. vivax-type parasites.
| Material Used for Transfection | Transfection Tools Available | Used to Investigate |
Refs |
|
|---|---|---|---|---|
|
P. vivax vivo |
in vivo derived blood stages | Transient transfection, Zinc-finger mediated recombination | blood stages in vivo |
[32,33] |
|
P. cynomolgi vivo |
in vivo derived blood stages |
Homology directed recombination, episomal, centromere |
blood stages (in vivo); liver stage schizonts and hypnozoites (in vitro) |
[29,37,38,39,40,41,42,90] |
|
P. cynomolgi vitro |
in vitro derived blood stages |
Episomal, Crispr/Cas9 |
blood stages in vitro |
[35] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



