Submitted:
19 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
Stand biomass in boreal forests has attained recognition for its crucial role in carbon cycling. In this study, we examined the effect of stand age on above- and belowground biomass allocation in downy birch (Betula pubescens Ehrh.) in three drained peatland sites in Central Finland. We used allometric equations to estimate individual-tree above-ground biomass and its various components, including stem wood, stem bark, living branches, leaves, and dead branches. The above-ground biomass ranged from 10.5 t ha-1 to 276.0 t ha-1 and foliage biomass from 0.009 t ha-1 to 0.6 t ha-1. We found that stand age had a strong correlation with above-ground biomass, but foliage biomass appeared to peak at the middle-aged (50 years) stand compared to the young (10 years) or mature (80 years) stands. We observed coarse root biomass production of 45, 250, and 110 g m-2 in the 0–50 cm peat layer for young, middle-aged, and mature stands, respectively. Most coarse root biomass occurred in the top 20-cm layer (77%). These findings improve our understanding of how biomass allocation patterns – specifically those in foliar biomass– change over time, and the impact on litter inputs into the soil.
Keywords:
downy birch
; biomass allocation
; above-ground biomass
; stand age
; tree diameter
; forest peatland.
1. Introduction
Forests have the potential to positively impact climate change by reducing atmospheric carbon dioxide (CO2) and sequestering C over the short- or mid-term [1]. They act as carbon sinks in terrestrial biomes, comprising about 80% of the global above-ground biomass (AGB) [2,3]. However, due to increasing demands for forest resources, the forest ecosystem is changing rapidly [4,5,6]. Therefore, the tree AGB plays a crucial role in estimating and monitoring forest carbon stocks [7]. Additionally, tree AGB estimations allow us to understand the interaction between forest growth and biomass productivity [8]. Research has shown that tree AGB yield is affected by factors such as stand age, tree species, and site fertility (e.g., the abundance of nitrogen in the soil) [9], with stand age being considered the most important factor [10,11,12,13].
Stand age affects the ability of the stand to sequester carbon by affecting the net primary productivity (NPP). Therefore, the chronosequence method, which represents a type of ‘natural experiment’, is often used to estimate the temporal dynamics of various stand development aspects and to understand the effect of stand age on successional changes [14]. However, while chronosequence studies of birch stands have been limited in Finland [15,16], such studies on birch AGB and belowground biomass (including fine root biomass) have been widely investigated in Southern Estonia [17].
In terms of quantity, foliage (leaves) biomass is not the most important biomass pool in a forest ecosystem. Nevertheless, due to its short lifespan, it has great potential to contribute to carbon dynamics in addition to photosynthesis and respiratory functions. On the other hand, the woody parts of the tree, such as stems, branches, and bark, store carbon for a longer period, as they remain active throughout the tree’s life, thus contributing to forests’ long-term carbon storage [18]. The relationship between stand age and leaf biomass is complicated and the stand structure, such as stand age, stand density, canopy depth, and tree size influence light level and light intensity, and therefore leaf biomass. Starr et al. (2005) [19] reported a weak negative correlation between stand age (35 to over 200 years) and leaf biomass production in Scot’s pine (Pinus sylvestris L.) stands in Finland. However, several studies have shown that foliage biomass increases with the increment of stand age [20,21,22,23].
Forests allocate a significant portion of their net primary productivity (20–65%) belowground for root and mycorrhizal growth and maintenance [24], emphasizing the need for a comprehensive understanding of biomass allocation, particularly belowground, to comprehend and forecast the forests’ response to elevated CO2 [25]. Fine and coarse root fractions are distinguished based on their diameter and exhibit different morphological and functional characteristics, decomposition dynamics, and resource availability responses [26,27]. While fine roots (<2 mm) play a crucial role in resource acquisition and contribute to soil carbon and nitrogen input through rapid turnover, coarse roots (>2 mm) are perennial, support fine root networks, transport water and nutrients, store carbohydrates, and provide physical support for aboveground biomass [24], accounting for up to 80% of belowground biomass [28] and 10–20% of NPP [29,30]. Coarse roots contribute to long-lived wood biomass formation [31], which persists for decades following senescence [32,33,34,35] and is vital for ecosystem carbon storage over the long term [25].
IPCC (2006) [36] classified the compartments of tree AGB as stem, stump, branch, bark, seed, and foliage. Tree-level AGB is generally estimated by either destructive sampling or by allometric models that use tree dimensions as explanatory variables (regression equations) [37]. Destructive sampling is time-consuming, arduous, costly, and highly disruptive to the ecosystem [38]. On the other hand, allometric models are non-destructive and have been considered the most reliable method to estimate tree biomass [39,40,41]. In general, an allometric model estimates traits that are difficult to measure (e.g., AGB) using other traits that are easy to measure (e.g., diameter at breast height (DBH at 1.3 m), tree height) [42,43,44,45]. However, as there have been limited chronosequence studies of birch stands in Finland, especially concerning foliage biomass, we measured leaf biomass in our study area. To measure leaf biomass, which is an important indicator of tree growth, we used a destructive sampling method. This involved collecting samples of the trees and analyzing them in a laboratory to determine the amount of leaf biomass present. By using this method, we were able to obtain more accurate information about the growth patterns of birch stands in our study area.
Birches are an abundant deciduous tree genus in Northern Europe, the Baltic Sea region, and Northwestern Russia [46]. Downy birch (Betula pubescens Ehrh.) and silver birch (Betula pendula Roth.) make up a significant proportion of the total volume of growing stocks in the region, with 16% in Finland, 11–16% in other Nordic countries, and 17–28% in the Baltic countries [47,48]. Downy birch is a primary succession tree on drained peatland sites in Finland and plays a significant role in the pulp and paper industry, as well as contributing to forest biodiversity. Despite being an important tree species in boreal forest ecosystems, the succession and biomass allocation of downy birch is not well understood [49,50].
Our study aimed to investigate the effects of stand age on above- and belowground biomass allocation in downy birch in three drained peatland sites in Central Finland. We used allometric equations [51] and empirical foliage and coarse root measurements to estimate the biomass allocation to stem wood including coarse roots, bark, leaves, and living branches in young, middle-aged, and mature stands. The specific aims of this study were to
- i)
- estimate biomass allocation in the above-ground parts, where we tested the hypotheses that 1) downy birch AGB allocation depends on the stand age, and 2) foliage biomass production would peak at the middle-age stands compared to the young and mature stand, due to the well-established canopy cover, increased nutrient uptake and allocation, and reduced competition.
- ii)
- estimate biomass allocation in the belowground parts, where we tested the hypotheses that 3) coarse root biomass (CRB) would show a consistent pattern with soil depth across all sites, indicating a general trend of root biomass decreasing with increasing soil depth, and 4) coarse root production would be higher in the middle-age stands than the young and mature stand.
2. Materials and Methods
2.1. Study Sites
We selected three naturally established downy birch-dominated stands on forestry drained peatland sites located in Juupajoki, Central Finland (N 61° 50' 36.542", E 24° 17' 37.010"). The mean ages of the trees within the stands were 10, 50, and 80 years, representing a chronology of young, middle-aged, and mature stands, respectively (Table 1). The average annual precipitation and temperature in the area over a 40-year period (1980–2020) were 58.6 ± SD 8.2 mm, and 3.9 ± SD 1.1 ℃, obtained from the nearest weather station data by the Finnish Meteorology Institute (FMI) [52] (Figure 1).
According to the Finnish classification system for drained peatland forests [53], all the sites can be classified as a nutrient-rich Herb-rich or Vaccinium myrtillus type (Rhtkg, MtkgII). The Loukaskorpi site (mature) was originally drained in 1909 (Table 1). The drainage network was further complemented in the 1920s, and ditch network maintenance was in 1964. The Rajasuo site (middle-aged) was drained in 1950 and had pre-commercial thinning conducted in the 1970s. Both the Loukaskorpi and Rajasuo sites also have Norway spruce (Picea abies) as an understory species with varying densities, and the stands have been non-managed for more than 50 years. The Nälkärasinsuo site (young stand) was originally drained in 1936, the tree stand was regenerated by clear cutting in 2006. The naturally established downy birch seedling stand was thinned in 2017, with the remaining stem density of 2767 ha-1 (Table 1). The management and succession history of all sites and their tree stands are well known due to long-term monitoring.
2.2. Tree Measurements and Above-Ground Biomass Estimation
At each study site, we established rectangular sample plots ranging in size from 100–600 m2 depending on the density of the stands in the study area in 2019 (Table 1). We measured the usual tree characteristics such as tree species, tree diameter at breast height (1.3 m, DBH) and height (H) from all the trees by using diameter tape hypsometer (Vertex III and Transponder T3, Haglöf Sweden AB). In total, there were 166 trees in the young stand, 46 trees in the middle-aged stand, and 170 trees in the mature stand.
We estimated the above-ground tree-level biomass using the multivariate biomass models by Repola (2008) [51] for individual tree compartments: stem wood, stem bark, living branches, dead branches, foliage, and total above-ground biomass. Although the data of these models represents mainly trees growing on upland sites, they have been considered suitable for trees of a wide range of stand structures and site properties [54,55,56]. In the biomass models, DBH and H, which are commonly measured in forest inventories, were the independent variables. We summed up the tree-level biomasses to hectare-level (tons, ha-1).
Despite the benefits of the equations by Repola (2008) [51], there are also various uncertainties related to the use of uncalibrated DBH and H as explanatory variables, which can result in biased predictions of different biomass compartments when, for example, the distribution of tree species (water and nutrient cycling, competition, etc.) highly dependent on climate deviates from that being in the modeling data [51]. Specifically, the foliage equation was constructed using downy birch with DBHs ranging from 11 to 26 cm [51], hence our tree dimensions fall within the range of data used for the equations of leafless AGB by Repola (2008) [51]. Regarding the other biomass models suitable for boreal conditions, equations presented by Repola (2008) [57] cover a wide range of sites and stand characteristics and are also widely used, particularly in Sweden. However, the models underestimated the litter fall biomass of Scots pine and overestimated it for Norway Spruce in test stands in Southern Finland [58].
2.1. Litterfall
To determine foliage biomass, we measured annual litter fall for three years (2019–2021). Litter falls were collected using seven plastic tube traps per site [59], which were on the ground at a height and diameter of 52 cm and 47.5 cm, respectively. Traps were equipped with textile bags. We drilled six evenly spaced drainage holes (0.5 cm) at the bottom to drain out the water from each bucket and to protect the litter from moisture and rapid decomposition. We further used twelve rectangular traps (0.5 m2, four traps per site) on the ground to collect the herbaceous litter fall. These were constructed of net mesh (1 × 1 cm2). We collected the litter twice per month during the most productive period of the growing season (June–August). All traps remained at the sites during the autumn and winter (September–February), and we collected the litter in the late spring (May) to yield annual estimates. The litter samples were sorted into leaves, needles, branches, and other parts (seeds, bark, buds, etc.) and weighed for fresh mass. The samples were then dried at 105 ℃ for 24 hours, after which we measured the dry mass (ME-T, Mettler Toledo group, Switzerland). We calculated oven-dry foliage (leaves) biomass for the three years (December-January) as t ha-1 yr-1.
2.2. Belowground Biomass
2.2.1. Coarse Root
In October 2020, CRB was measured using the soil coring technique [60,61]. A total of 30 soil cores were obtained across the entire stand, using a cylindrical corer with a diameter of 46 mm. To capture the variability across different depths, the soil cores were divided into five [61] layers (0–10; 10–20; 20–30; 30–40, and 40–50 cm) and subsequently placed in polyethylene bags. The samples were then transported to the laboratory and immediately stored at -18 °C until analysis.
The roots were thoroughly washed to remove any attached soil, and sorted into four categories based on their diameter ranges: d < 2 mm, 2 mm ≤ d < 5 mm, 5 mm ≤ d < 10 mm, and ≥ 10 mm. Additionally, the roots were further divided into tree roots and understory vegetation under a microscope, primarily based on color and ectomycorrhizal tips, as described by Bhuiyan et al. (2017) [62]. The samples were dried at 60 °C and weighed to the nearest 0.001 g. The CRB per square meter was calculated by summing up the average values of coarse root mass for the successive soil layers from the soil cores. However, the fine root fraction (d < 2 mm) was not included in this calculation due to significant losses (65–75%) reported in previous studies [63,64].
2.3. Statistical Analysis
For hypothesis (1), we used Spearman’s rank correlation coefficient to examine the correlation between stand age and above-ground biomass allocation. For hypothesis (2) that the foliage biomass production would peak at the middle-age stands, we used a one-way analysis of variance (ANOVA) to compare the stand age and foliage biomass. We also used General Linear Model (GLM) to test the effect of stand age on measured foliage. We used the LSD Post Hoc test at an alpha level of 0.05 and checked for homogeneity and heteroscedasticity of variances using Levene’s and F tests. For hypothesis (3) that the coarse root biomass (CRB) would show a consistent pattern with soil depth across all sites, indicating a general trend of root biomass decreasing with increasing soil depth, we used repeated measures analysis of variance to evaluate the differences in CRB in the soil depth in all three stands. We considered the stand age to be a between-subjects (grouping) factor, and depth (layer within a core) was the within-subjects (repeated) factor. For hypothesis (4) that the coarse root production would be higher in the middle-aged stands than in the young and mature stands, we used a one-way analysis of variance (ANOVA) and GLM to test the effect of stand age on coarse root biomass production. All analyses were conducted using IBM SPSS Statistics 25 and R software (v3.6.0) [65].
3. Results
3.1. Diameter and Height
We found a wide range of tree DBHs (1–35.5 cm) and heights (1.3– 36.3 m) (Figure 2). The DBH and height of the downy birches increased with the age of the stand. In the mature (80 years old) stand, the mean DBH (12.9 cm ± SE 0.5) was nearly triple that of the young (10 years old) (4.5 cm ± SE 0.2) and 1.5 times higher than in the middle-aged (50 years old) stand (DB-50) (8.9 cm ± SE 0.9) (Table 1). Similarly, the mean tree height was highest in the mature stand (16.7 m ± SE 0.7), four times that of the young seedlings (4.6 m ± SE 0.1) and 1.5 times higher than in the middle-aged stand (11.4 m ± SE 0.8) (Table 1). On the other hand, in all three stands, there was a strong linear relationship between DBH and height (p < 0.001; Figure 2).
3.2. Above-Ground Biomass Allocation
The amount of biomass (AGB) in the different parts of downy birch trees (stem wood, stem bark, living branches, and dead branches) increased as the trees aged (Table 2). Specifically, mature trees had nearly 26 times more AGB (276 ± SE 0.6 t ha-1) than young seedlings (10.5 ± SE 0.2 t ha-1) and 1.5 times more than middle-aged trees (175.4 ± SE 0.3 t ha-1) (Table 2). The total AGB, stem wood, and living branches of downy birch were positively correlated with tree age, while dead branches were negatively correlated with tree age (Table 3). The proportion of stem wood biomass in the total AGB was lowest in young seedlings (65.9%) compared to the middle-aged stand (85.3%) and mature stand (84.9%) (Figure 3). On the other hand, the relative amounts of stem bark and living branch biomass were higher in young seedlings compared to middle and mature stands (Figure 3).
3.1. Foliage Biomass
The biomass of dry foliage (leaves) from litter fall ranged from 0.009 to 0.6 t ha-1, with an average of 0.2 ± SE 0.1 t ha-1 (4 A, Table 4). The effect of stand age on dry foliage biomass was statistically significant, indicating that there is likely a relationship between the age of a stand and the amount of dry foliage biomass it produces (ANOVA, P < 0.05). In the LSD Post Hoc test, the middle-aged stand showed a statistically significant difference, while the young and mature stands did not differ significantly from each other (Appendix: Table A1).
The middle-aged stand appeared to peak in dry foliage biomass more than the other stands during the studied years (Figure 4 A). In 2020, the mean dry foliage biomass yield of the middle-aged stand was highest (mean 0.11 ± SE 0.01 t ha-1), followed by 2021 (mean 0.06 ± SE 0.002 t ha-1) and lowest in 2019 (mean 0.03 ± SE 0.08 t ha-1) (Figure 4 A).
We compared our foliage biomass results with allometric model estimates calculated by equations by Repola (2008) [51] (Figure 4 C). We found that Repola’s equation both underestimated and overestimated foliage biomass across different years and stand ages (Figure 4 A, C). For instance, in the young stand, the equations underestimated the foliage biomass by 4.5 and 1.8 times in 2019 and 2020, respectively, but overestimated it by 2.1 times in 2021 (Figure 4 A, C). Similarly, in the middle-aged stand, Repola’s equation overestimated the foliage biomass by 3.7, 1.1, and 8.3 times in 2019, 2020, and 2021, respectively (Figure 4 A, C). Similar trends were observed in the mature stand (DB-80), where Repola’s model overestimated biomass by 10.1 and 2.2 times in 2019 and 2020, respectively, but underestimated biomass by 1.7 times in 2021 (Figure 4 A, C). The same trend was also observed when comparing fresh foliage and Repola’s equation (Figure 4 B, C). On the other hand, by comparing fresh and dry foliage, we were able to determine the moisture content of the leaves and found that the fresh leaves had lost between 1% and 15% of their water content by weight in different aged stands and years during the drying process (Figure 4 A, B).
3.1. Coarse-Root Biomass
In the middle-aged stand, coarse root biomass production of downy birch stand in the 0–50 cm peat layer totaled 250 g m-2, 45 g m-2 in the young stand, and 110 g m-2 (Table 5) in the matured stand, and supporting our hypothesis that coarse root production was higher in the middle-aged stand (Table 6). All three stands showed significant differences in the CRB allocation by depth (Table 6, Figure 5).
Most of the CRB allocation was in the upper 0–30 cm layer and showed a consistent pattern with depth of soil layer across all sites, supporting our hypothesis of decreasing root biomass with increasing depth (Figure 5). The largest diameter (d ≥ 10 mm) accounted for 68% of the total coarse root biomass. The corresponding shares of the root fractions 5 ≤ d < 10 mm and 2 ≤ d < 5 mm were similar (17% and 15%, respectively). The largest share of the coarse root (55%) was in the upper 10 cm soil layer (Figure 5). Depth and stand age were also statistically significant, indicating differing CRB depth profiles for the stands (Table 6, Figure 5, P < 0.001).
The results of ANOVA showed that the stand age effect on coarse root biomass was significant (P < 0.05); the effect of the depth of the soil layer was also statistically significant (P < 0.001). According to multiple comparison of the mean, all three stands showed significant differences of coarse root biomass (LSD test; P < 0.05; Appendix: Table A2).
4. Discussion
The novelty of the current study is revealing the successional development of downy birch biomass allocation, especially foliage biomass, in peatland forests. We observed that the foliage biomass in the middle-aged stand was significantly higher than in the young and mature stands. This contrasts with Repola (2008) [51], whose model underestimated and overestimated the magnitude of foliage biomass (leaves) for each age class in the present study.
Our young seedling stand AGB allocation estimates are of the same magnitude as in Hytönen et al. (2018) [66], where downy birch coppice stand AGB was measured in peat cutaway areas in northern Finland (Age class: A: 10–12 years, B: 15–16 years, C: 22–24 years). The comparability of results can be explained by the similarity of the study sites, which both were peatlands. Similarly, our AGB estimates for the middle-age stand (DB-50) were consistent (175.4 ± SE 0.3 t ha-1) with the study of Uri et al. (2012) [17] for downy birch in Estonia (AGB: 118.6 t ha-1). In their study, the middle-aged stand was 30 years (stands age range: 12–78 years old) compared to the mean age of 50 years in this study. The climate in their study area belongs to the hemiboreal vegetation zone [17] compared to our central boreal forest zone. However, the soil type in the Uri et al. (2012 [17] study was the same as in our study (peatland). Thus, it may imply that the soil type, tree species, and stand age fundamentally control AGB yield despite the dissimilar climate [9].
Our study revealed that the biomass equation by Repola (2008) [51] provided inaccurate estimates of foliage biomasses. This deviation may be due to inter-tree competition within stands, such as stand structure, stand density, self-thinning process, and tree relative growth rate, which can affect the downy birch branch number and biomasses. For example, Mäkelä and Vanninen, (1998) [67] reported that the AGB of Scots pine in Southern Finland varied due to different competition status of the trees (competition for light reduces the density of foliage in the crown). On the other hand, the litter fall in a forest ecosystem provides important insights into the dynamics of nutrient cycling and serves as a primary link between producers and decomposers [68]. However, estimating the amount of branch and foliage biomass at the level of individual trees within a stand can be difficult because trees exhibit a high degree of variability in their size, shape, and foliage density [69]. This variability can make it challenging to accurately estimate the amount of biomass present in branches and leaves, as different trees within the same stand may have vastly different amounts of these materials. Therefore, accurately measuring stand-level branch and foliage biomass requires careful consideration of this variability, as well as the use of appropriate sampling and measurement techniques to ensure accurate and representative estimates [70,71]. Additionally, estimating foliage biomass in matured stands can be difficult because the applicability of biomass equations is often limited to smaller trees, as the variance and potential error increase with stand age [58]. This result could have important implications for forestry management and conservation efforts, as it suggests that the age of a forest stand can play a significant role in determining its productivity and ecological function. Further research may be needed to explore the specific mechanisms underlying this relationship and to identify strategies for optimizing dry foliage biomass production in different stand ages.
The larger tree DBHs and heights in the mature stand compared to the other age classes are reflected in stand-level AGB estimates. Thus, the higher total AGB of the mature stand is perhaps connected with the larger tree DBHs and height compared to the other stands' tree sizes (diameters and heights) [21]. On the other hand, dry foliage was of the same magnitude in young and mature stands, but not in the middle-aged stand. This can be explained by the higher stand density of the middle-aged stand, as stand density affects the production of AGB (Table 1) [17]. The stem numbers of all the stands show that the stand density is high in our non-managed middle-aged and mature stands because of the self-thinning and closed canopy of the trees, thus in pristine downy birch stands, the foliage biomass seems to develop early during succession (Table 1). On the other hand, the chronosequence approach, which examines the development dynamics of tree stands over time, is frequently used in modeling forest succession. However, this approach also highlights the high between-site variability in biotic and abiotic conditions, as well as the influence of stand management and silvicultural practices on stand density, biomass, and yield.
According to Ukonmaanaho et al. (2008) [68], the proportion of senescent leaves’ biomass is closely related to the amount of aboveground litter fall, which can vary between species and years. For example, a smaller litter fall biomass can indicate a longer retention of foliage in the tree canopy. The consistency of our foliage biomass results over successive years suggests that the yields are not coincidental, but accurately reflect the actual leaf mass of the forest. Additionally, the higher annual mean temperature in 2020 (6.2 ℃) compared to 2019 (4.7 ℃) and 2021 (4.1℃) at our study sites may have contributed to the large differences in annual foliage yield [52].
In forest ecosystems, roots are important contributors to net primary production and play a crucial role in carbon cycling [72]. Despite their importance, there is limited empirical data on the belowground coarse root biomass of downy birch, with only a few studies available in the literature [73,74]. However, it is important to consider that various factors can influence coarse root biomass [75], and trees growing in peatlands may allocate more biomass to belowground parts than those growing on mineral soil sites [76]. This was also demonstrated in our study, where the proportion of roots (64%) in a young downy birch stand was higher than the corresponding proportion (ranging from 24% to 59%) in a silver birch stand of similar age growing on mineral soil [61].
The observed distribution of coarse roots, with the majority (77%) located in the upper 20 cm soil layer (Figure 5), is in line with the expected soil nutrient concentration pattern [77]. The tendency for higher concentrations of roots in horizons or patches of higher fertility is well documented in the soil profile [78], and lower soil horizons generally contain fewer roots due to increasingly unfavorable soil factors [75]. Nevertheless, the root system has been shown to adapt to varying environmental conditions, such that trees growing in nutrient-rich forests are expected to develop a higher amount of root biomass [79]. Our findings indicate that coarse root biomass is higher in a more nutrient-rich forest (middle-aged stand) than in nutrient-poor forests (young and mature stands). However, it is important to note that the final conclusions should not be drawn from the 1-year data only.
5. Conclusions
Our study of downy birch stands of different ages showed that as the stand age increased, the leafless above-ground biomass (AGB) also increased. This is consistent with the findings of Konôpka et al. (2020) [20], reflecting that stand age is an indirect measure of stand biomass. The stem wood, living branches, and total AGB of downy birch stands were positively correlated with stand age. Foliage biomass is a good indicator of tree growth, as photosynthesis takes place in foliage. Leaf biomass is also related to growth efficiency (GE, growth/leaf area) which can be expressed as woody parts biomass increment per foliage area [80]. Based on our results, we can conclude that downy birch foliage biomass production peaks at the middle-aged stand compared to the young or mature stands. However, estimating the foliage biomass using Repola’s equation [51] underestimated and overestimated the biomass in each age class. The results of this study suggest that the production and allocation of coarse root biomass in drained peatlands are subject to various factors, including stand age, nutrient availability, and soil depth. These results emphasize the necessity for additional research aimed at exploring the contribution of coarse roots to ecosystem functioning and nutrient cycling. Our findings can improve the understanding of downy birch biomass allocation patterns in drained peatland, specifically foliar biomass, and its impact on litter inputs into the soil. Nevertheless, we recommend further studies to understand the relationship between AGB and stand age and site quality. At the same time, the number of study sites (plots) might be increased to evaluate the AGB differences among stand ages. To further understand the dynamics of downy birch on drained peatlands, we suggest the following three directions for future research:
- Investigating the long-term effects of different management practices, such as successive thinning, on the vegetation succession and biomass accumulation of downy birch on drained peatland sites.
- Investigating the role of understory species, such as Norway spruce, and their interactions with downy birch in terms of competition for resources and potential facilitation in the vegetation succession process on drained peatland sites.
- Comparison of coarse root and fine root biomass between downy birch and other tree species: Future research could compare coarse root and fine root biomass between downy birch and other tree species in these ecosystems and investigate how these differences affect ecosystem functions such as nutrient cycling and carbon sequestration.
Author Contributions
Conceptualization, M.R.K (Md Rezaul Karim); Formal analysis, M.R.K (Md Rezaul Karim); investigation, M.R.K (Md Rezaul Karim) and S.S (Sakari Sarkkola); data curation, M.R.K (Md Rezaul Karim); funding acquisition, M.R.K (Md Rezaul Karim); methodology, M.R.K (Md Rezaul Karim) and S.S (Sakari Sarkkola); supervision, S.S (Sakari Sarkkola) and K.R.G (Katja Rinne-Garmston); visualization, M.R.K (Md Rezaul Karim), K.R.G (Katja Rinne-Garmston), and S.S (Sakari Sarkkola); writing—original draft preparation, M.R.K (Md Rezaul Karim); writing—review and editing, M.R.K (Md Rezaul Karim), K.R.G (Katja Rinne-Garmston), and S.S (Sakari Sarkkola); project administration, S.S (Sakari Sarkkola). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by “Marjatta and Eino Kolli foundation”, and “The Niemi foundation”, (decision number: 20180001). Katja Rinne-Garmston acknowledges funding from the European Research Council (755865) and the Academy of Finland (295319).
Data Availability Statement
Data is contained within the article or supplementary material. Dataset and associated codes used in the main results are available upon reasonable request to the corresponding author.
Acknowledgments
We would like to express gratitude to all laboratory staff at Hyytiälä forestry station in central Finland. Also, many thanks go to Dr. Hanna-Riitta kylmäläinen (lecturer, University of Helsinki) and Professor Raija Laiho for comments on the draft and suggestions and Jacqelin De Faveri for the language revisions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Table A1.
Least Significant Difference–LSD Post Hoc test statistics of measured foliage.
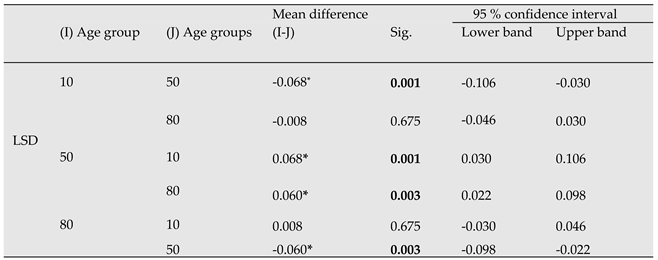 |
Notes: Dependent Variable: Measured foliage (t ha -1); *. The mean difference is significant at the 0.05 level. The P values in bold were statistically significant at an alpha level of 0.05.
Table A2.
Least Significant Difference–Tukey HSD, LSD, and Bonferroni Post Hoc test statistics of coarse root biomass.
Table A2.
Least Significant Difference–Tukey HSD, LSD, and Bonferroni Post Hoc test statistics of coarse root biomass.
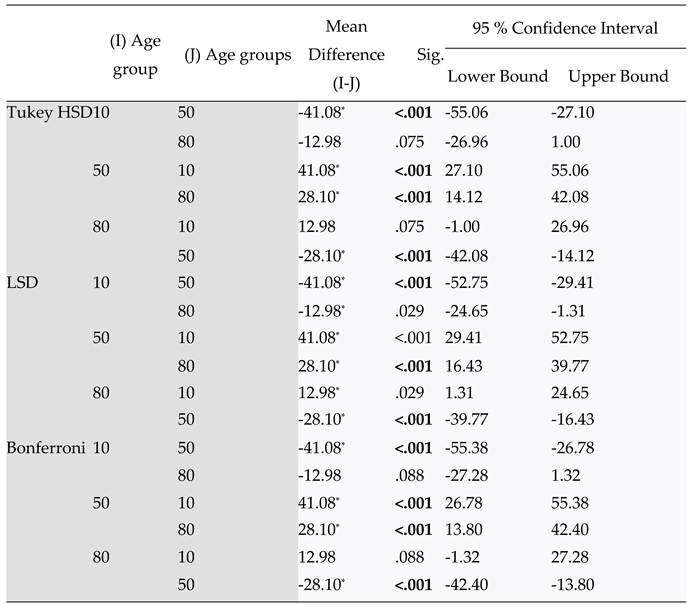 |
Notes: Dependent Variable: Coarse root biomass (g m -2); *. The mean difference is significant at the 0.05 level. The P values in bold were statistically significant at an alpha level of 0.05.
References
- Beedlow, P.A., Tingey, D.T., Phillips, D.L., Hogsett, W.E., Olszyk, D.M., 2004. Rising atmospheric CO 2 and carbon sequestration in forests. Front. Ecol. Environ. 2, 315–322. [CrossRef]
- Saatchi, S.S., Houghton, R.A., Dos Santos Alvalá, R.C., Soares, J.V., Yu, Y., 2007. Distribution of aboveground live biomass in the Amazon basin: AGLB IN THE AMAZON BASIN. Glob. Change Biol. 13, 816–837. [CrossRef]
- Ometto, Jean Pierre, Aguiar, A.P., Assis, T., Soler, L., Valle, P., Tejada, G., Lapola, D.M., Meir, P., 2015. Amazon forest biomass density maps: tackling the uncertainty in carbon emission estimates, in: Ometto, Jean P., Bun, R., Jonas, M., Nahorski, Z. (Eds.), Uncertainties in Greenhouse Gas Inventories. Springer International Publishing, Cham, pp. 95–110. [CrossRef]
- Thuiller, W., Lavergne, S., Roquet, C., Boulangeat, I., Lafourcade, B., Araujo, Miguel.B., 2011. Consequences of climate change on the tree of life in Europe. Nature 470, 531–534. [CrossRef]
- Tian, X., Sohngen, B., Kim, J.B., Ohrel, S., Cole, J., 2016. Global climate change impacts on forests and markets. Environ. Res. Lett. 11, 035011. [CrossRef]
- Perea-Moreno, M.-A., Samerón-Manzano, E., Perea-Moreno, A.-J., 2019. Biomass as Renewable Energy: Worldwide Research Trends. Sustainability 11, 863. [CrossRef]
- Dulamsuren, C., Klinge, M., Bat-Enerel, B., Ariunbaatar, T., Tuya, D., 2019. Effects of forest fragmentation on organic carbon pool densities in the Mongolian forest-steppe. For. Ecol. Manag. 433, 780–788. [CrossRef]
- Chave, J., Andalo, C., Brown, S., Cairns, M.A., Chambers, J.Q., Eamus, D., Fölster, H., Fromard, F., Higuchi, N., Kira, T., Lescure, J.-P., Nelson, B.W., Ogawa, H., Puig, H., Riéra, B., Yamakura, T., 2005. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145, 87–99. [CrossRef]
- Vogel, J.G., Gower, S.T., 1998. Carbon and Nitrogen Dynamics of Boreal Jack Pine Stands With and Without a Green Alder Understory. Ecosystems 1, 386–400. [CrossRef]
- Peichl, M., Arain, M.A., 2007. Allometry and partitioning of above- and belowground tree biomass in an age-sequence of white pine forests. For. Ecol. Manag. 253, 68–80. [CrossRef]
- Donnelly, L., Jagodziński, A.M., Grant, O.M., O’Reilly, C., 2016. Above- and below-ground biomass partitioning and fine root morphology in juvenile Sitka spruce clones in monoclonal and polyclonal mixtures. For. Ecol. Manag. 373, 17–25. [CrossRef]
- Jagodziński, A., Dyderski, M., Gęsikiewicz, K., Horodecki, P., 2018. Tree- and Stand-Level Biomass Estimation in a Larix decidua Mill. Chronosequence. Forests 9, 587. [CrossRef]
- Schepaschenko, D., Moltchanova, E., Shvidenko, A., Blyshchyk, V., Dmitriev, E., Martynenko, O., See, L., Kraxner, F., 2018. Improved Estimates of Biomass Expansion Factors for Russian Forests. Forests 9, 312. [CrossRef]
- Fukami, T., Wardle, D.A., 2005. Long-term ecological dynamics: reciprocal insights from natural and anthropogenic gradients. Proc. R. Soc. B Biol. Sci. 272, 2105–2115. [CrossRef]
- Mälkönen, E., Saaarsalmi, A., 1982. Hieskoivikon biomassatuotos ja ravinteiden menetys kokopuun korjuussa, Folia forestalia. Metsäntutkimuslaitos, Helsinki.
- Wang, J.R., Zhong, A.L., Simard, S.W., Kimmins, J.P., 1996. Aboveground biomass and nutrient accumulation in an age sequence of paper birch (Betula papyrifera) in the Interior Cedar Hemlock zone, British Columbia. For. Ecol. Manag. 83, 27–38. [CrossRef]
- Uri, V., Varik, M., Aosaar, J., Kanal, A., Kukumägi, M., Lõhmus, K., 2012. Biomass production and carbon sequestration in a fertile silver birch (Betula pendula Roth) forest chronosequence. For. Ecol. Manag. 267, 117–126. [CrossRef]
- Thornley, J.H.M., Cannell, M.G.R., 2000. Managing forests for wood yield and carbon storage: a theoretical study. Tree Physiol. 20, 477–484. [CrossRef]
- Starr, M., Saarsalmi, A., Hokkanen, T., Merilä, P., Helmisaari, H.-S., 2005. Models of litterfall production for Scots pine (Pinus sylvestris L.) in Finland using stand, site and climate factors. For. Ecol. Manag. 205, 215–225. [CrossRef]
- Konôpka, B., Pajtík, J., Šebeň, V., Surový, P., Merganičová, K., 2020. Biomass Allocation into Woody Parts and Foliage in Young Common Aspen (Populus tremula L.)—Trees and a Stand-Level Study in the Western Carpathians. Forests 11, 464. [CrossRef]
- Hu, M., Lehtonen, A., Minunno, F., Mäkelä, A., 2020. Age effect on tree structure and biomass allocation in Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies [L.] Karst.). Ann. For. Sci. 77, 90. [CrossRef]
- Yang, B., Xue, W., Yu, S., Zhou, J., Zhang, W., 2019. Effects of Stand Age on Biomass Allocation and Allometry of Quercus Acutissima in the Central Loess Plateau of China. Forests 10, 41. [CrossRef]
- Miksys, V., Varnagirytekabasinskiene, I., Stupak, I., Armolaitis, K., Kukkola, M., Wojcik, J., 2007. Above-ground biomass functions for Scots pine in Lithuania. Biomass Bioenergy 31, 685–692. [CrossRef]
- Landsberg, J.J., Sands, P.J., 2011. Physiological ecology of forest production: principles, processes and models, 1st ed. ed, Terrestrial ecology series. Academic Press/Elsevier, Amsterdam Boston.
- Walker, A.P., De Kauwe, M.G., Medlyn, B.E., Zaehle, S., Iversen, C.M., Asao, S., Guenet, B., Harper, A., Hickler, T., Hungate, B.A., Jain, A.K., Luo, Y., Lu, X., Lu, M., Luus, K., Megonigal, J.P., Oren, R., Ryan, E., Shu, S., Talhelm, A., Wang, Y.-P., Warren, J.M., Werner, C., Xia, J., Yang, B., Zak, D.R., Norby, R.J., 2019. Decadal biomass increment in early secondary succession woody ecosystems is increased by CO2 enrichment. Nat. Commun. 10, 454. [CrossRef]
- Litton, C.M., Raich, J.W., Ryan, M.G., 2007. Carbon allocation in forest ecosystems. Glob. Change Biol. 13, 2089–2109. [CrossRef]
- Poorter, H., Nagel, O., 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Funct. Plant Biol. 27, 1191. [CrossRef]
- Butnor, J.R., Johnsen, K.H., Oren, R., Katul, G.G., 2003. Reduction of forest floor respiration by fertilization on both carbon dioxide-enriched and reference 17-year-old loblolly pine stands: Reduction of forest floor respiration by fertilization. Glob. Change Biol. 9, 849–861. [CrossRef]
- Giardina, C.P., Ryan, M.G., 2002. Total Belowground Carbon Allocation in a Fast-growing Eucalyptus Plantation Estimated Using a Carbon Balance Approach. Ecosystems 5, 487–499. [CrossRef]
- Maier, C.A., Albaugh, T.J., Lee Allen, H., Dougherty, P.M., 2004. Respiratory carbon use and carbon storage in mid-rotation loblolly pine ( Pinus taeda L.) plantations: the effect of site resources on the stand carbon balance: Stand respiration and carbon storage. Glob. Change Biol. 10, 1335–1350. [CrossRef]
- Mobley, M.L., Richter, D. deB., Heine, P.R., 2013. Accumulation and decay of woody detritus in a humid subtropical secondary pine forest. Can. J. For. Res. 43, 109–118. [CrossRef]
- Anderson, P.H., Johnsen, K.H., Butnor, J.R., Gonzalez-Benecke, C.A., Samuelson, L.J., 2018. Predicting longleaf pine coarse root decomposition in the southeastern US. For. Ecol. Manag. 425, 1–8. [CrossRef]
- Clark, D.A., Brown, S., Kicklighter, D.W., Chambers, J.Q., Thomlinson, J.R., Ni, J., 2001. Measuring net primary production in forests: concepts and field methods. Ecol. Appl. 11, 356–370. [CrossRef]
- Johnsen, K.H., Wear, D., Oren, R., Teskey, R.O., Sanchez, F., Will, R., Butnor, J., Markewitz, D., Richter, D., Rials, T., 2001. Meeting global policy commitments: carbon sequestration and southern pine forests. J. For. 99, 14–21.
- Ludovici, K.H., Zarnoch, S.J., Richter, D.D., 2002. Modeling in-situ pine root decomposition using data from a 60-year chronosequence. Can. J. For. Res. 32, 1675–1684. [CrossRef]
- Eggleston, H., 2006. Intergovernmental Panel on Climate Change, National Greenhouse Gas Inventories Programme, and Chikyu Kankyo Senryaku Kenkyu Kikan. 2006. IPCC guidelines for national greenhouse gas inventories. IPCC Guidel. Natl. Greenh. Gas Invent.
- Laar, A. van, Akça, A., 2007. Tree And Stand Biomass. For. Mensurat. 183–199.
- Cunliffe, A.M., McIntire, C.D., Boschetti, F., Sauer, K.J., Litvak, M., Anderson, K., Brazier, R.E., 2020. Allometric Relationships for Predicting Aboveground Biomass and Sapwood Area of Oneseed Juniper (Juniperus monosperma) Trees. Front. Plant Sci. 11, 94. [CrossRef]
- Hosoda, K., Iehara, T., 2010. Aboveground biomass equations for individual trees of Cryptomeria japonica , Chamaecyparis obtusa and Larix kaempferi in Japan. J. For. Res. 15, 299–306. [CrossRef]
- Chojnacky, D.C., Heath, L.S., Jenkins, J.C., 2014. Updated generalized biomass equations for North American tree species. Forestry 87, 129–151. [CrossRef]
- Krofcheck, D., Litvak, M., Lippitt, C., Neuenschwander, A., 2016. Woody Biomass Estimation in a Southwestern U.S. Juniper Savanna Using LiDAR-Derived Clumped Tree Segmentation and Existing Allometries. Remote Sens. 8, 453. [CrossRef]
- Gower, S.T., Kucharik, C.J., Norman, J.M., 1999. Direct and Indirect Estimation of Leaf Area Index, fAPAR, and Net Primary Production of Terrestrial Ecosystems. Remote Sens. Environ. 70, 29–51. [CrossRef]
- Zianis, D., Suomen Metsätieteellinen Seura, Metsäntutkimuslaitos (Eds.), 2005. Biomass and stem volume equations for tree species in Europe, Silva Fennica monographs. Finnish Society of Forest Science, Finnish Forest Research Institute, Helsinki, Finland.
- Trap, J., Bureau, F., Brethes, A., Jabiol, B., Ponge, J.-F., Chauvat, M., Decaëns, T., Aubert, M., 2011. Does moder development along a pure beech (Fagus sylvatica L.) chronosequence result from changes in litter production or in decomposition rates? Soil Biol. Biochem. 43, 1490–1497. [CrossRef]
- Wang, Y., Pyörälä, J., Liang, X., Lehtomäki, M., Kukko, A., Yu, X., Kaartinen, H., Hyyppä, J., 2019. In situ biomass estimation at tree and plot levels: What did data record and what did algorithms derive from terrestrial and aerial point clouds in boreal forest. Remote Sens. Environ. 232, 111309. [CrossRef]
- Niemistö, P., Viherä-Aarnio, A., Velling, P., Heräjärvi, H., Verkasalo, E., 2008. Koivun kasvatus ja käyttö.
- Korhonen, K.T., Ihalainen, A., Heikkinen, J., Henttonen, H., Pitkänen, J., 2007. Suomen metsävarat metsäkeskuksittain 2004–2006 ja metsävarojen kehitys 1996–2006.
- Hynynen, J., Niemisto, P., Vihera-Aarnio, A., Brunner, A., Hein, S., Velling, P., 2010. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 83, 103–119. [CrossRef]
- Wirth, C., Schumacher, J., Schulze, E.-D., 2004. Generic biomass functions for Norway spruce in Central Europe--a meta-analysis approach toward prediction and uncertainty estimation. Tree Physiol. 24, 121–139. [CrossRef]
- Lehtonen, A., Mäkipää, R., Heikkinen, J., Sievänen, R., Liski, J., 2004. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. For. Ecol. Manag. 188, 211–224. [CrossRef]
- Repola, J., 2008. Biomass equations for birch in Finland. Silva Fenn. 42. [CrossRef]
- Finnish Meteorology Institute (FMI). Available online: https://en.ilmatieteenlaitos.fi/download-observations (accessed on 31 May 2022).
- Vasander, H., Laine, J., 2008. Site type classification on drained peatlands, in: Finland-Fenland: Research and Sustainable Utilisation of Mires and Peat. Finnish Peatland Society Maahenki, pp. 146–151.
- Kankare, V., Holopainen, M., Vastaranta, M., Puttonen, E., Yu, X., Hyyppä, J., Vaaja, M., Hyyppä, H., Alho, P., 2013. Individual tree biomass estimation using terrestrial laser scanning. ISPRS J. Photogramm. Remote Sens. 75, 64–75. [CrossRef]
- Ståhl, G., Heikkinen, J., Petersson, H., Repola, J., Holm, S., 2014. Sample-Based Estimation of Greenhouse Gas Emissions From Forests—A New Approach to Account for Both Sampling and Model Errors. For. Sci. 60, 3–13. [CrossRef]
- Repola, J., 2006. Models for vertical wood density of Scots pine, Norway spruce and birch stems, and their application to determine average wood density. Silva Fenn. 40. [CrossRef]
- Marklund, L.G., 1988. Biomass functions for pine, spruce and birch in Sweden. Rapp.-Sver. Lantbruksuniversitet Institutionen Foer Skogstaxering Swed.
- Lehtonen, A., 2005. Estimating foliage biomass in Scots pine (Pinus sylvestris) and Norway spruce (Picea abies) plots. Tree Physiol. 25, 803–811. [CrossRef]
- Laasasenaho, J., 1982. Taper curve and volume functions for pine, spruce and birch. Metsäntutkimuslaitos.
- Vogt, K.A., Grier, C.C., Meier, C.E., Keyes, M.R., 1983. Organic Matter and Nutrient Dynamics in Forest Floors of Young and Mature Abies amabilis Stands in Western Washington, as Affected by Fine-Root Input. Ecol. Monogr. 53, 139–157. [CrossRef]
- Varik, M., Aosaar, J., Ostonen, I., Lõhmus, K., Uri, V., 2013. Carbon and nitrogen accumulation in belowground tree biomass in a chronosequence of silver birch stands. For. Ecol. Manag. 302, 62–70. [CrossRef]
- Bhuiyan, R., Minkkinen, K., Helmisaari, H.-S., Ojanen, P., Penttilä, T., Laiho, R., 2017. Estimating fine-root production by tree species and understorey functional groups in two contrasting peatland forests. Plant Soil 412, 299–316. [CrossRef]
- Lõhmus, K., Lasn, R., Oja, T., 1991. The influence of climatic and soil physical conditions on growth and morphology of Norway spruce roots, in: Developments in Agricultural and Managed Forest Ecology. Elsevier, pp. 233–239.
- Uri, V., Tullus, H., Lõhmus, K., 2002. Biomass production and nutrient accumulation in short-rotation grey alder (Alnus incana (L.) Moench) plantation on abandoned agricultural land. For. Ecol. Manag. 161, 169–179. [CrossRef]
- Team, R. C. R: A Language and Environment for Statistical Computing. 2013.
- Hytönen, J., Aro, L., Jylhä, P., 2018. Biomass production and carbon sequestration of dense downy birch stands on cutaway peatlands. Scand. J. For. Res. 33, 764–771. [CrossRef]
- Mäkelä, A., Vanninen, P., 1998. Impacts of size and competition on tree form and distribution of aboveground biomass in Scots pine. Can. J. For. Res. 28, 216–227. https://doi.org/10.1139/x97-199. [CrossRef]
- Ukonmaanaho, L., Merilä, P., Nöjd, P., Nieminen, T.M., 2008. Litterfall production and nutrient return to the forest floor in Scots pine and Norway spruce stands in Finland.
- Teobaldelli, M., Somogyi, Z., Migliavacca, M., Usoltsev, V.A., 2009. Generalized functions of biomass expansion factors for conifers and broadleaved by stand age, growing stock and site index. For. Ecol. Manag. 257, 1004–1013. [CrossRef]
- Wojtan, R., Tomusiak, R., Zasada, M., Dudek, A., Michalak, K., Wroblewski, L., Bijak, S., Bronisz, K., 2011. Trees and their components biomass expansion factors for Scots pine (Pinus sylvestris L.) of western Poland. Sylwan 155, 236–243.
- Castedo-Dorado, F., Gómez-García, E., Diéguez-Aranda, U., Barrio-Anta, M., Crecente-Campo, F., 2012. Aboveground stand-level biomass estimation: a comparison of two methods for major forest species in northwest Spain. Ann. For. Sci. 69, 735–746.
- Prescott, C., Godbold, D.L., Helmisaari, H.-S., Addo-Danso, S.D., 2016. Introduction to Forests, roots and soil carbon. For. Ecol. Manag. 359, 321.
- Hunziker, M., Sigurðsson, B.D., Halldórsson, G., Schwanghart, W., Kuhn, N., 2014. Biomass allometries and coarse root biomass distribution of mountain birch in southern Iceland.
- Uri, V., Kukumägi, M., Aosaar, J., Varik, M., Becker, H., Morozov, G., Karoles, K., 2017. Ecosystems carbon budgets of differently aged downy birch stands growing on well-drained peatlands. For. Ecol. Manag. 399, 82–93. [CrossRef]
- Cairns, M.A., Brown, S., Helmer, E.H., Baumgardner, G.A., 1997. Root biomass allocation in the world’s upland forests. Oecologia 111, 1–11. [CrossRef]
- Laiho, R., Finér, L., 1996. Changes in root biomass after water-level drawdown on pine mires in southern Finland. Scand. J. For. Res. 11, 251–260. [CrossRef]
- Uri, V., Lõhmus, K., Ostonen, I., Tullus, H., Lastik, R., Vildo, M., 2007. Biomass production, foliar and root characteristics and nutrient accumulation in young silver birch (Betula pendula Roth.) stand growing on abandoned agricultural land. Eur. J. For. Res. 126, 495–506. [CrossRef]
- Sutton, R., Tinus, R.W., 1983. Root and root system terminology. For. Sci. 29, a0001-z0001.
- Vanninen, P., Ylitalo, H., Sievänen, R., Mäkelä, A., 1996. Effects of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.). Trees 10, 231–238. [CrossRef]
- Gersonde, R.F., O’Hara, K.L., 2005. Comparative tree growth efficiency in Sierra Nevada mixed-conifer forests. For. Ecol. Manag. 219, 95–108. [CrossRef]
Figure 1.
Mean air temperature (A) and precipitation (B) of the study area in Juupajoki, Finland (Finnish Meteorology Institute (FMI)) [52].
Figure 1.
Mean air temperature (A) and precipitation (B) of the study area in Juupajoki, Finland (Finnish Meteorology Institute (FMI)) [52].
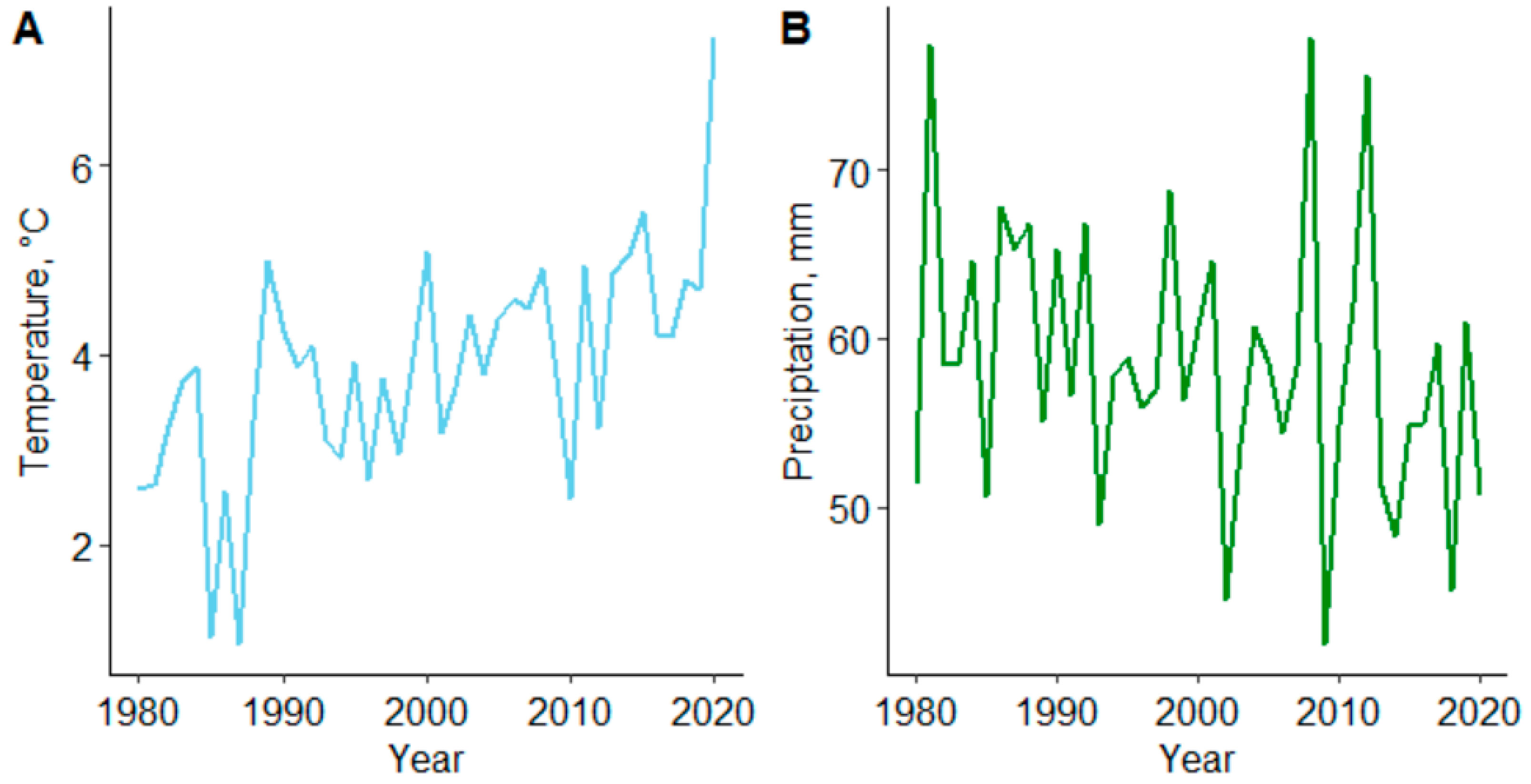
Figure 2.
Relationship between the downy birch tree height and diameter at breast height (DBH) in the studied experimental stands of three mean age classes (10, 50 and 80 years) in Juupajoki, Central Finland in 2019. The shaded areas around each regression line represent 95% confidence intervals.
Figure 2.
Relationship between the downy birch tree height and diameter at breast height (DBH) in the studied experimental stands of three mean age classes (10, 50 and 80 years) in Juupajoki, Central Finland in 2019. The shaded areas around each regression line represent 95% confidence intervals.
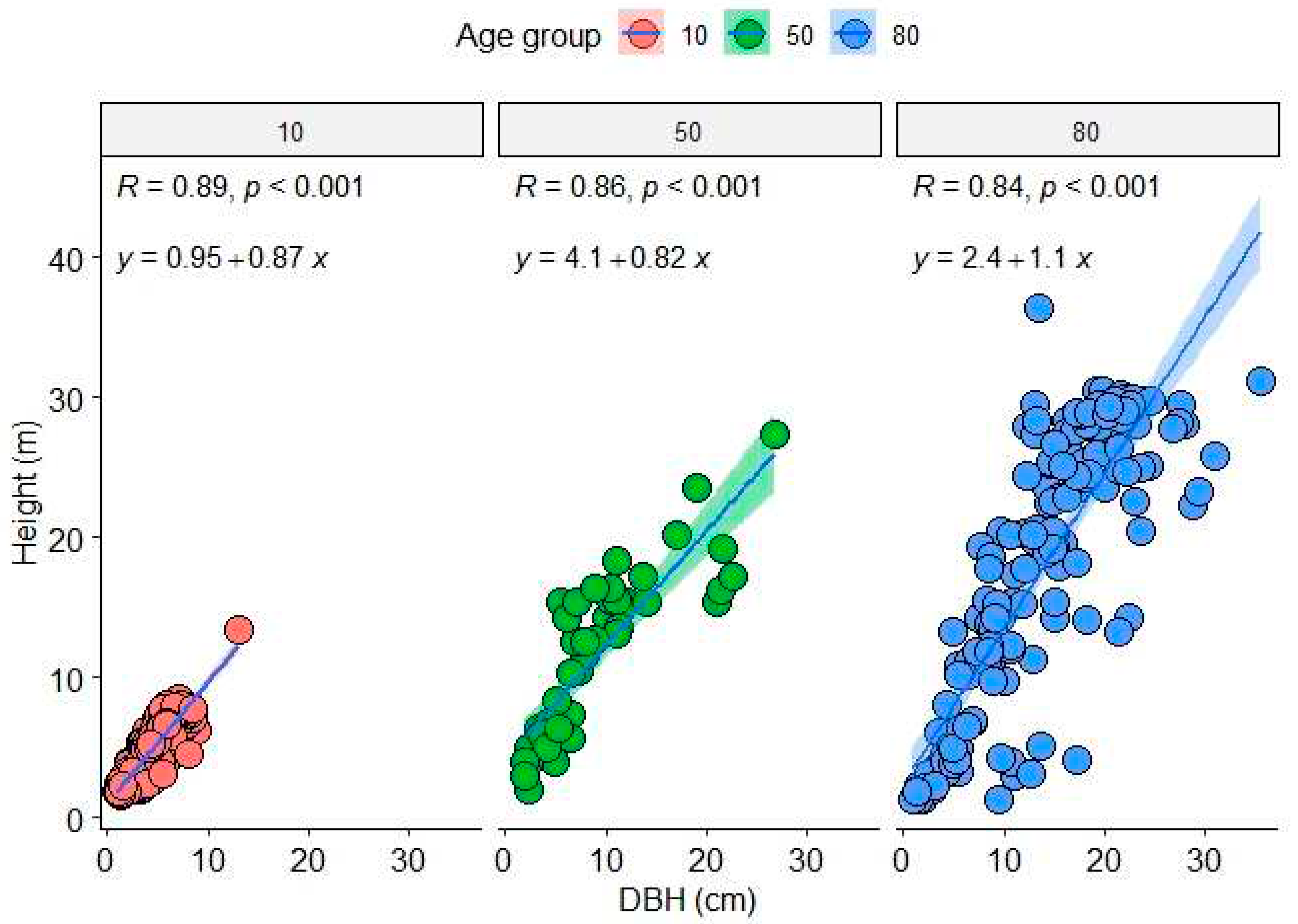
Figure 3.
Relative proportions of tree biomass compartments to total aboveground biomasses in each age class – Mature: 80 years, middle-aged: 50 years, and young: 10 years.
Figure 3.
Relative proportions of tree biomass compartments to total aboveground biomasses in each age class – Mature: 80 years, middle-aged: 50 years, and young: 10 years.
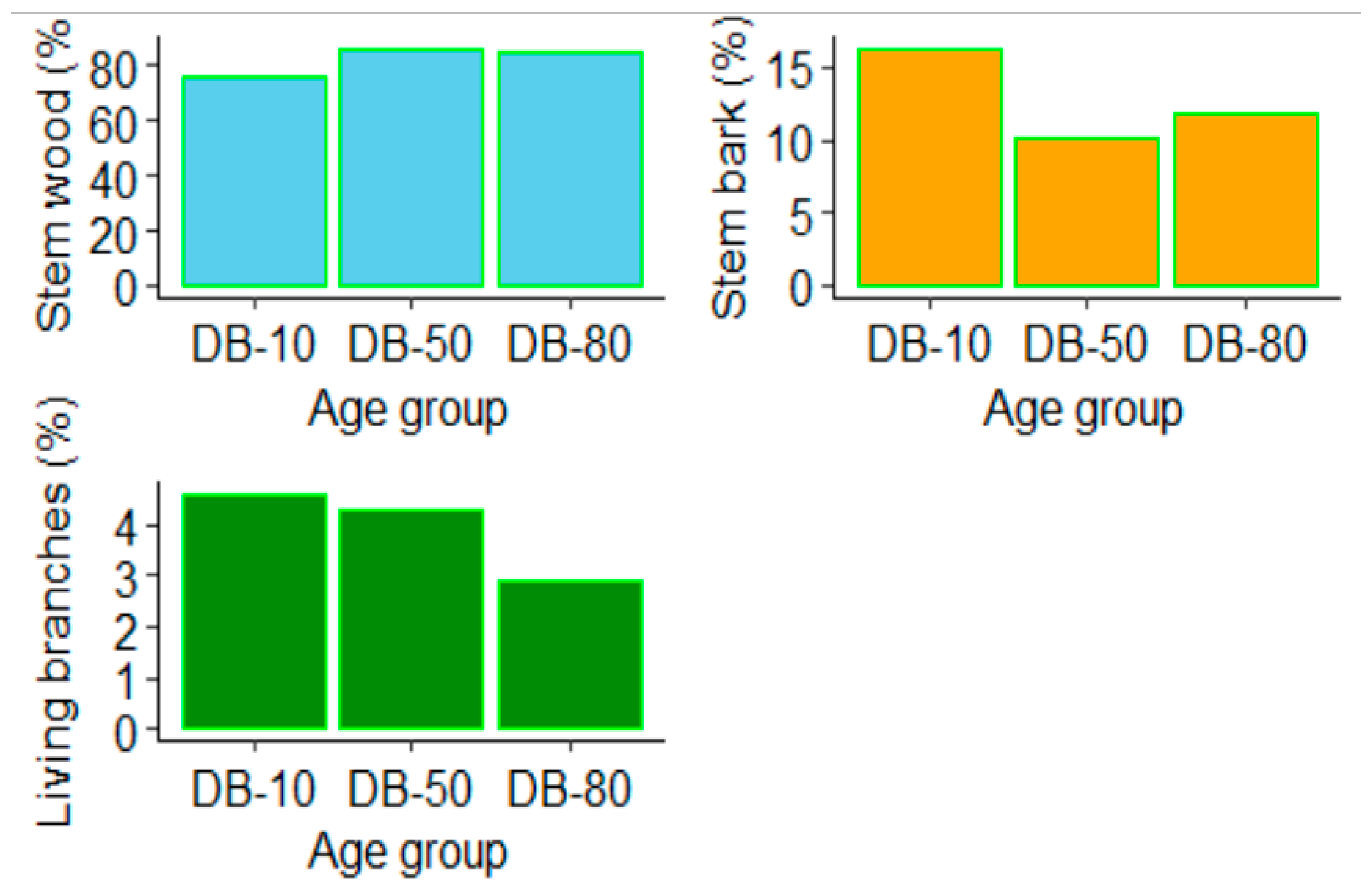
Figure 4.
Mean living foliage biomass (leaves) measured from the captured litter fall in the three different age classes of downy birch in the study area of drained peatlands (A, B); C. Mean foliage biomass estimated by the biomass equation of Repola, (2008) [51] for downy birch. DB-10: Young stand; DB-50: Middle-age stand; DB-80: Mature stand. Bars represent the standard error of means (SE).
Figure 4.
Mean living foliage biomass (leaves) measured from the captured litter fall in the three different age classes of downy birch in the study area of drained peatlands (A, B); C. Mean foliage biomass estimated by the biomass equation of Repola, (2008) [51] for downy birch. DB-10: Young stand; DB-50: Middle-age stand; DB-80: Mature stand. Bars represent the standard error of means (SE).

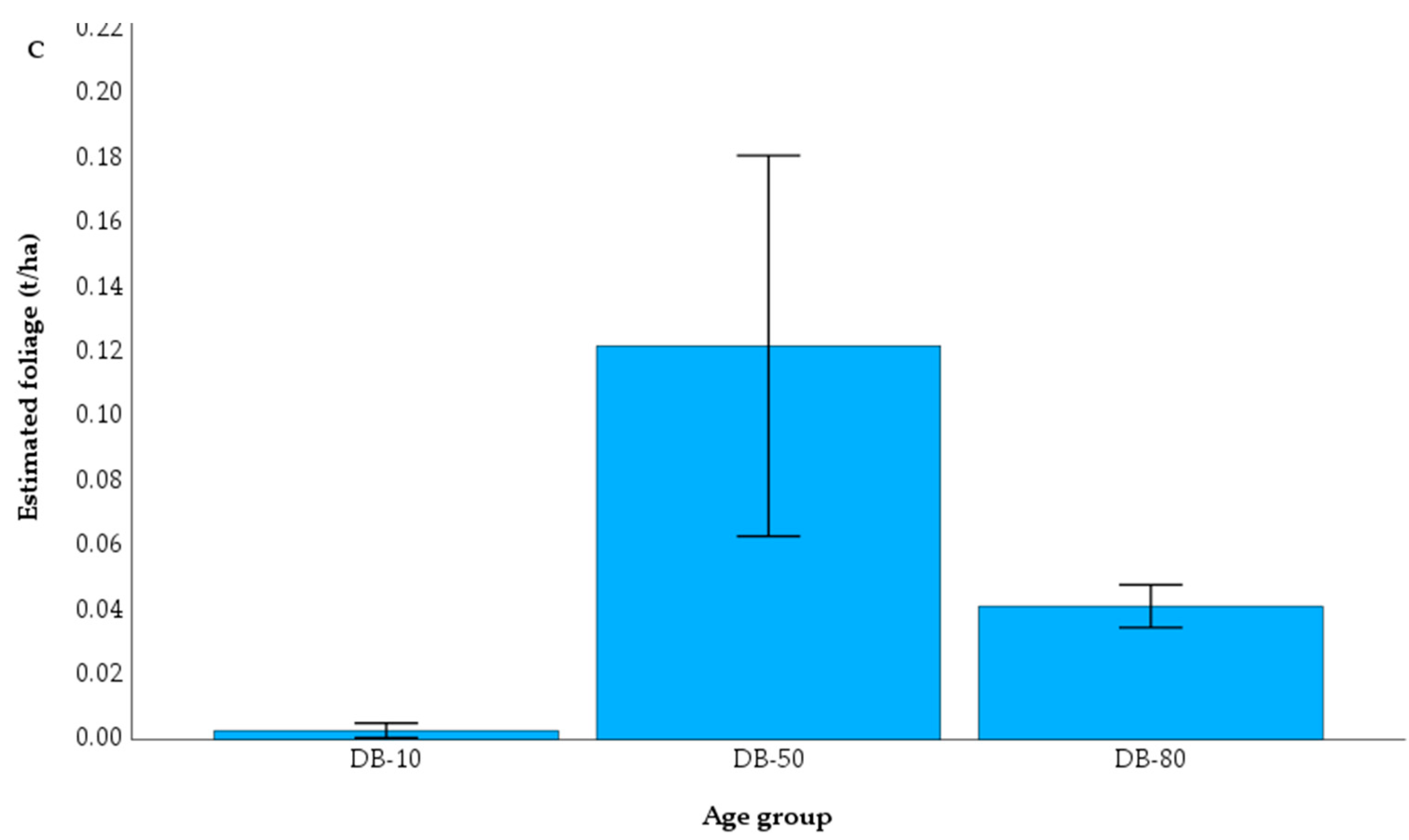
Figure 5.
Mean coarse root production in the drained peatland forests. Error bars are standard error of means. Layer 1: 0-10 cm depth from moss surface; layer 2: 10-20 cm; layer 3: 20-30 cm; layer 4: 30-40 cm; layer 5: 40-50 cm.
Figure 5.
Mean coarse root production in the drained peatland forests. Error bars are standard error of means. Layer 1: 0-10 cm depth from moss surface; layer 2: 10-20 cm; layer 3: 20-30 cm; layer 4: 30-40 cm; layer 5: 40-50 cm.
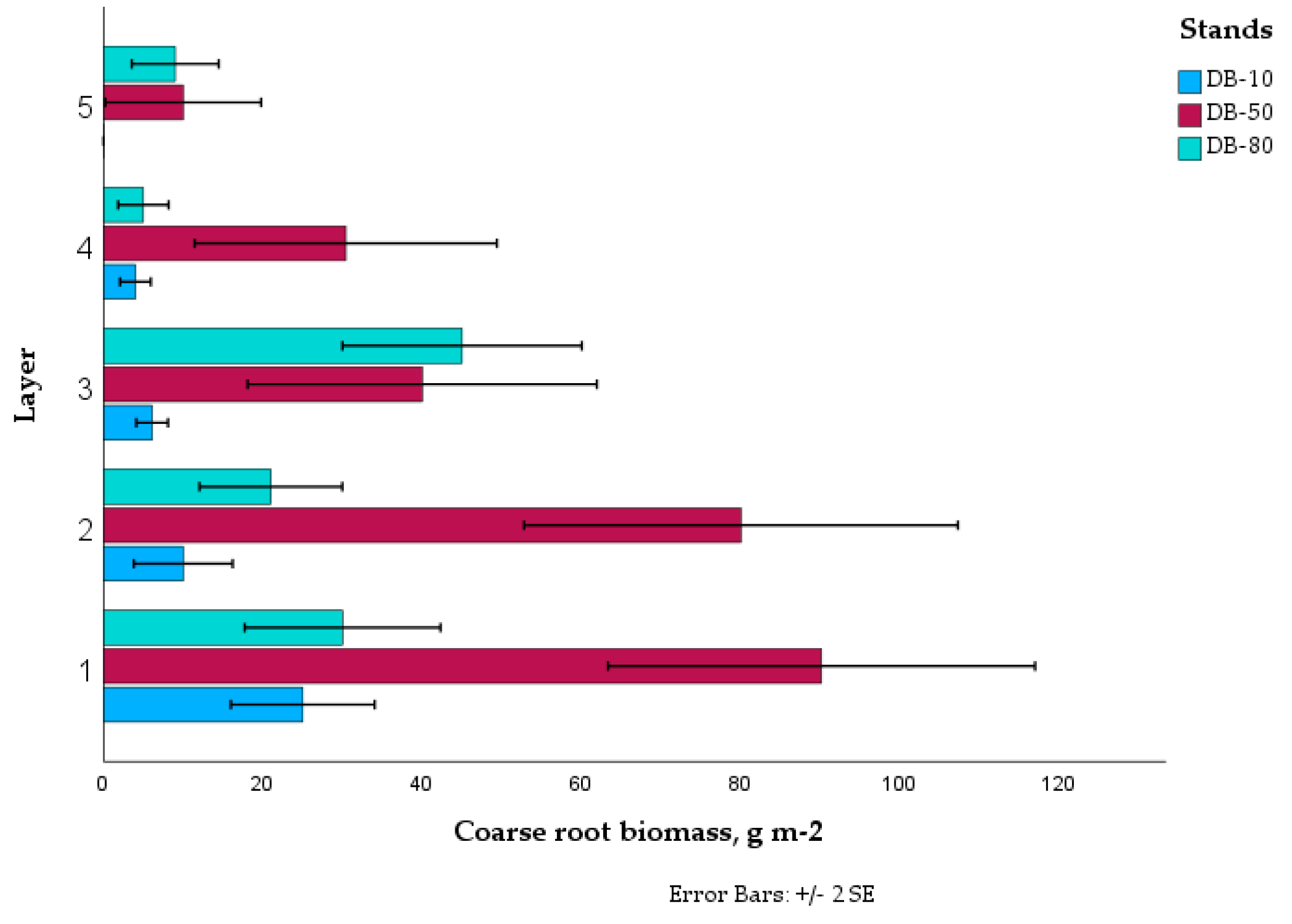
Table 1.
Location, stand characteristics, tree diameter (DBH; diameter at 1.3 m) and tree height of experimental downy birch-dominated (DB) stands on peatlands in Central Finland.
Table 1.
Location, stand characteristics, tree diameter (DBH; diameter at 1.3 m) and tree height of experimental downy birch-dominated (DB) stands on peatlands in Central Finland.
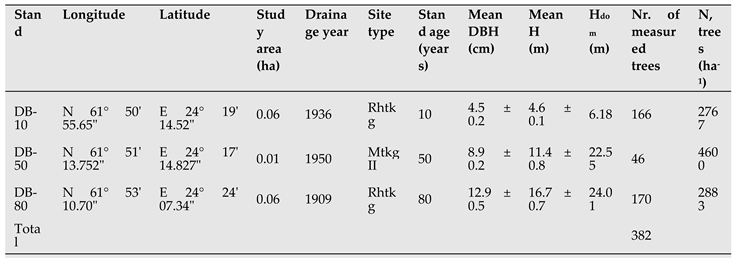 |
Notes: DB-10: Young stand; DB-50: Middle-age stand; DB-80: Mature stand; DBH: diameter at breast height; H: tree height; N: stand density; Hdom: stand dominant height. For DBH and H, the standard error is presented. Rhtkg = herb-rich type; Mtkg = Vaccinium myrtillus-type [53].
Table 2.
Stand-level aboveground biomass by compartment, summed up from tree-level estimates, at the three downy birch-dominated stands representing different age classes.
Table 2.
Stand-level aboveground biomass by compartment, summed up from tree-level estimates, at the three downy birch-dominated stands representing different age classes.
 |
Note: DB-10: Young stand; DB-50: Middle age stand; DB-80: Mature stand; AGB: aboveground biomass.
Table 3.
Correlation matrix of stand age and the different compartments of tree aboveground biomass (t ha-1). SA: stand age, SB: stem biomass, SW: stem wood, LB: living branches, DB: dead branches; AGB: total aboveground biomass.
Table 3.
Correlation matrix of stand age and the different compartments of tree aboveground biomass (t ha-1). SA: stand age, SB: stem biomass, SW: stem wood, LB: living branches, DB: dead branches; AGB: total aboveground biomass.
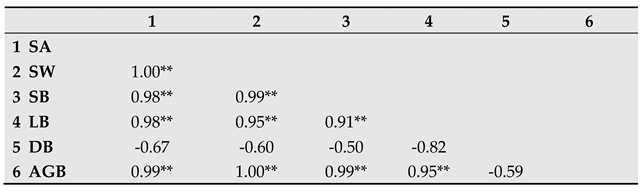 |
**Correlation is significant at the 0.01 level (2-tailed).
Table 4.
Dry foliage biomass (t ha -1) with standard deviation (SD), minima and maxima across the three downy birch-dominated stands and years. N = 6 for all sites.
Table 4.
Dry foliage biomass (t ha -1) with standard deviation (SD), minima and maxima across the three downy birch-dominated stands and years. N = 6 for all sites.
 |
Notes: DB-10: Young stand; DB-50: Middle-age stand; DB-80: Mature stand.
Table 5.
Coarse root production (g m-2) with standard deviation (SD), standard error (SE), minima and maxima by depth in the drained forests. N = 10 for each site.
Table 5.
Coarse root production (g m-2) with standard deviation (SD), standard error (SE), minima and maxima by depth in the drained forests. N = 10 for each site.
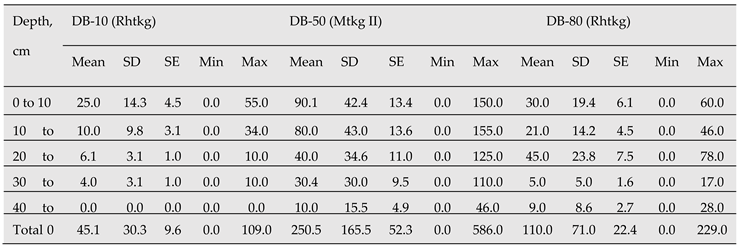 |
Notes: DB-10: Young stand; DB-50: Middle-age stand; DB-80: Mature stand. Rhtkg = herb-rich type; Mtkg = Vaccinium myrtillus-type [53].
Table 6.
Repeated measures analysis of Variance on the effects of stand age (young, middle-aged and mature) on coarse root biomass (g m-2). Data are from cores and were divided into five 10-cm layers per core (within-factor Depth).
Table 6.
Repeated measures analysis of Variance on the effects of stand age (young, middle-aged and mature) on coarse root biomass (g m-2). Data are from cores and were divided into five 10-cm layers per core (within-factor Depth).
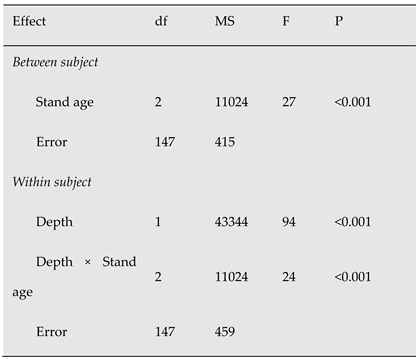 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



