Submitted:
18 July 2023
Posted:
20 July 2023
You are already at the latest version
Abstract
Tomato brown rugose fruit virus (ToBRFV) is a causal agent of severe emergent diseases in Solanaceae hosts with agronomic relevance, such as tomato and pepper. Herein, we analyzed the seed transmission rate and efficacy of different seed disinfection treatments and performed a comparison of molecular biology techniques seeking rapid and sensitive detection in seed lots. We evaluated several total RNA extraction and RT-PCR protocols using a distinct combination of PCR primers to test for the presence of the ToBRFV Mexican strain in tobacco host. Our results showed that the percentage of seed and seedlings transmission was less than 1% and that a 3% sodium hypochlorite solution was effective as a seed disinfection treatment. Finally, the most sensitive molecular method identified for virus detection consisted of a CTAB-Trizol RNA extraction followed by nested PCR using primers reported by Dovas et al. (2004). Additionally, we tested potential natural hosts in selected Cucurbitaceae and Solanaceae species. Our results showed that the ToBRFV Mexican strain is capable of experimentally infecting eggplant (Solanum melongena), tomatillo (Physalis ixocarpa), and tobacco (Nicotiana rustica).
Keywords:
seed transmission
; tobamovirus
; diagnostic method
; natural hosts
; RT-PCR
1. Introduction
Tomato brown rugose fruit virus (ToBRFV) is a tobamovirus that forms rigid rod-shaped particles. Its genome comprises a positive single-stranded RNA (+ssRNA) genome that contains four open reading frames (ORFs). ORF1 and ORF2 encode the RNA-dependent-RNA polymerase (RdRp), while ORF3 encodes the movement protein (MP) and ORF4 for the capsid protein (CP) [2]. As its viral particles are very stable, it is easily mechanically transmitted. Therefore, cultivation activities are the primary way it spreads in greenhouses [3,4].
As ToBRFV quickly spreads through tomato and pepper-producing zones worldwide, the infected seed has been proposed as an essential source of this virus transmission. Most tobamoviruses are primarily found as contaminants in the coat of seeds. While viral particles are efficiently transmitted when they enter the embryo, those in the seed coat may not be infectious when they separate from the seedling. Although this causes a small transmission percentage, the process can occur as a primary field inoculation and quickly spread through contact [5]. Furthermore, viral particles can remain viable in clay soil and crop wastes for months or even years [6]. The mechanical transmission of ToBRFV from infected seeds to seedlings is very likely responsible for initiating a new infection [7].
Current disinfection methods used in commercial seed production involve chemical treatments with chlorhydric acid (1-9%), calcium hypochlorite (1-5%), sodium hypochlorite (1-3%) [8], tetramethyl thiuram disulfide (TMTD), and trisodium phosphate (10%) [9]. Nevertheless, the continually occurring epidemics show that the treatment of seeds is not having the desired impact, which may be due to the preservation of the virus in the internal tissues of the embryo [5]. In keeping with this, laboratory techniques for tobamoviruses detection in seeds include serological methods such as ELISA, which is a reliable and rapid technique [10,11]. However, it can give false positives and negatives. Molecular methods are also used, such as RT-PCR and qRT-PCR. These methods are more sensitive [12], but as they only detect a genome fragment, performing biological tests of infectivity in indicator plants is suggested as a confirmation [5].
In the case of ToBRFV, the International Seed Federation (ISF) recommends employing qRT-PCR to analyze subsamples of seeds without any disinfectant treatment, followed by bioassays to determine whether local lesions exist in tobacco plants [13]. For instance, the National Center of Phytosanitary Reference (SENASICA) established the following procedure in Mexico. First, seeds should be soaked in distilled water, shaken for 20 minutes, and then germinated to perform the total RNA extraction protocols [14].
Aware of the relevance of emergent diseases associated with potential seed dispersion of ToBRFV Mexican strains to agronomically important Solanaceae hosts, we focused on analyzing the virus seed transmission rate and the efficacy of different seed disinfection treatments. Additionally, we compared molecular biology techniques seeking rapid and sensitive virus detection in seed lots. We evaluated a combination of several total RNA extraction and RT-PCR protocols using a distinct combination of PCR primers reported in the literature. Moreover, we tested potential natural hosts to determine whether or not ToBRFV could infect cucurbits and additional hosts in selected Solanaceae species. Our results showed the specificity of the percentage of Nicotiana rustica seedlings and seeds infected. Herein we are contributing findings that could be useful for better understanding the biology underlying the ToBRFV Mexican strain, its natural hosts, and diagnostic methods with potential application in the routine detection of seeds lots and plants, reducing the risk of dispersion.
2. Materials and Methods
2.1. Plant Materials Used as an Inoculum Source
In September 2020, tomato plants (Solanum lycopersicum L.) showing chlorosis, leaf narrowing, and mosaics (typical symptoms of ToBRFV infection) were collected from commercial greenhouses in Colima, Mexico. The inoculum source tested positive for ToBRFV by RT-PCR. A Nicotiana glutinosa plant was mechanically inoculated using this tissue, and local necrotic lesions were observed. One of these lesions was cut to inoculate a second N. glutinosa plant. This process was repeated twice. A local lesion of the fourth N. glutinosa plant was used to inoculate a tomato plant (var. saladette). Then, 30 days post-inoculation (dpi), the tomato plant was analyzed by RT-PCR for the presence of the virus. The inoculum source was increased by mechanical inoculation on tomato plants to perform the different host bioassays and seed transmission tests.
2.2. RNA Extraction and RT-PCR
Plant material from the inoculum source was divided into two groups for further processing. The first group was macerated in a phosphate buffer (0.1 M potassium phosphate at pH 7.0 + 0.1 M sodium sulfite + 1% of β-mercaptoethanol). This suspension was used to inoculate healthy tomato seedlings mechanically. Meanwhile, the second group was used to extract the total RNA to identify the associated agent by RT-PCR using the specific primers and conditions described by Rodríguez-Mendoza et al. (2018) [15]. The expected fragments were then sequenced.
2.3. Evaluation of Total RNA Extraction Methods
Pepper and tomato seeds were germinated in Petri dishes containing paper towels moistened with sterile distilled water. After 10 to 20 days, when the cotyledonary leaves emerged and expanded, 100 mg of complete seedlings (stem, leaves, and root) of each plant species were separately used to perform total RNA extraction. In addition, three seeds of most of the plant species were planted in 1L Styrofoam cups containing a peat-growing mix. Plants were maintained under greenhouse conditions. After 45 days, 100 mg of foliar tissue (pedicels, leaves, and shoots) was collected to perform total RNA extraction. A total of four RNA extraction methods were evaluated: PlantRNAeasy miniKit®, following the manufacturer's instructions; Trizol®, CTAB2 %, and CTAB2%-Trizol® following the protocol described by Jordon-Thaden et al. (2015) [16] (minor modifications, Appendix A). Plant tissue maceration was performed with liquid nitrogen in a sterile mortar. The amount and quality of the extracted RNA were quantified using a Nanodrop 2000®, and the absorbance measurements 260/280 and 260/230 were recorded.
2.4. Primer Evaluation and Sensitivity Limit
Three replicates of tomato, eggplant (Solanum melongena), and tomatillo (Physalis ixocarpa) seedlings were mechanically inoculated with the tomato inoculum. At 30 dpi, when plants showed chlorosis and mosaic symptoms, total RNA extraction was performed using the previously described CTAB-Trizol protocol. The quality and quantity of the extracted RNA were verified using a Nanodrop 2000®. The final concentration was adjusted to 300 ng/µL. Subsequently, serial dilutions of each total RNA sample were made and adjusted to 10, 1, 0.1, 0.01, 0.001, and 0.0001 ng/µL. Pairs of specific primers, described by Ling et al. (2019) [17] and Rodríguez-Mendoza et al. (2018) [15] and reported by Dovas et al. (2004) [1], were tested. The specifications described in the protocols reported for each primer pair were followed.
2.5. Seed Transmission Test
2.5.1. Acquisition of Seeds
Virus mechanical inoculation was carried out in thirty healthy tobacco plants (N. rustica) at two true leaves stage. Additionally, plants were mock-inoculated as controls. Plants were maintained under greenhouse-controlled conditions with a 12-hours light:12 hours dark cycle at temperatures ranging from 16°C to 24°C until fruiting. Seeds were collected from each plant when the capsules opened and stored at 4°C. ToBRFV infection in tobacco plants was confirmed by total RNA extraction and RT-PCR using the primers reported by Dovas et al. (2004) [1].
2.5.2. Germination Test
Stereoscopic microscopy was used to select vain seeds. Those were divided into three groups of 100 seeds from ToBRFV-infected plants and their respective mock-inoculated controls. Each group was placed into a petri dish containing paper moistened with sterile distilled water. Petri dishes were incubated at 23°C for 72 hours of light, followed by 18° to 24°C for one 12-hour light: 12-hour dark cycle. The sprouted seedlings were counted after 20 days. This procedure was repeated three times.
2.5.3. Estimation of Percentage of Infection in Seeds or Seedlings
The technique described by Albrechtsen (2006) [18] was employed to calculate the percentage (P) of infected seeds or seedlings. In brief, this technique uses the maximum likelihood estimation, in which a population (seeds or seedlings) is divided into subgroups (N) of a known amount (n). P= [1- (Y/N)^1/n]X 100, where Y is the number of negative subgroups. Then, the percentage of infection was calculated in N. rustica seedlings from seeds obtained from ToBRFV-infected plants.
2.5.4. Experimental Method to Determine the Percentage of Infected Seeds
In order to determine the percentage of infected seeds, fifteen subgroups of 250 seeds per Petri dish were placed under the previously described conditions. Then, seedlings were selected using stereoscopic microscopy when sprouted, and the seed coat was separated from the cotyledon. The seedlings were placed in previously sterilized paper envelopes and stored at -20°C. Subsets of 150 were obtained from each Petri dish for a total of 2,250 seedlings. The CTAB-Trizol protocol described by Jordon-Thaden et al. (2015) [16] was employed to obtain total RNA using each subset. The quality and amount of RNA extracted were verified and quantified using a Nanodrop 2000®. Total RNA was adjusted to 300 ng/µL, and 1 µL was employed for reverse transcription, followed by nested PCR using the primers and protocols described by Dovas et al. (2004) [1] (Figure 1). Electrophoresis with a 1.5% agarose gel was performed on the products obtained from the nested PCR. The expected size of the fragments was verified, and the fragments were purified with EXO-SAP according to the manufacturer’s instructions. The purified products were shipped to Macrogen, Korea, for sequencing. The second experiment used stereoscopic microscopy to select 15 subgroups of 150 N. rustica seeds from ToBRFV-positive plants. These were placed in Eppendorf tubes, and total RNA extraction, nested RT-PCR, and purification were performed with each subgroup, as described previously (Figure 1a). Both experiments included two subgroups of 150 seeds or seedlings from healthy N. rustica controls.
2.5.5. Infectivity Tests of Viral Particles in Seeds
Evaluation of infectivity was performed on fifteen subsets comprising 100 seeds treated with 3% sodium hypochlorite for three minutes with respective subsets of untreated seeds. Each group was macerated in liquid nitrogen. The maceration was divided equally into two Eppendorf tubes, one containing 500 µL of phosphate buffer at a pH of 7.0 and the other containing 600 µL of 2% CTAB buffer. Healthy N. rustica plants were mechanically inoculated using carborundum as an abrasive. The CTAB RNA total extraction, followed by nested RT-PCR, was performed on samples in the second tube (Figure 1b). The inoculated N. rustica plants were maintained in a greenhouse at 18°C to 23°C with a 12 hours light:12 hours dark cycle. Plants were visually observed daily to record the appearance of local or systemic symptoms. At 30 dpi, the apical leaves were selected from each plant to extract total RNA and to perform RT-PCR. We included two subgroups of 100 treated seeds, 100 untreated seeds, or 100 seedlings from healthy N. rustica.
2.6. Bioassays of Potential Hosts
To identify potential ToBRFV hosts, mechanical inoculations were performed on five biological replicates from each of the following species; a) Solanaceae family: tomato (Solanum lycopersicum L.), tomatillo (Physalis ixocarpa), tobacco (Nicotiana rustica), pepper (Capsicum annum), eggplant (Solanum melongena L.); b) Cucurbitaceae family; watermelon (Citrullus lanatus), cantaloupe (Cucumis melo L.), squash (Cucurbita pepo), and cucumber (Cucumis sativus L.). Additionally, three plants from each species were mock-inoculated as controls. The plants were maintained under greenhouse conditions and observed daily to record symptoms. After 30 dpi, plants were tested for virus presence using RT-PCR employing the primers described by Dovas et al. (2004) [1] and ELISA, with antibodies against TMV (Agdia®) that have cross-reactions. This experiment was performed thrice: first in October 2019, second in February 2020, and last in April 2020. During the first experiment, the average monthly temperature in the greenhouse ranged from 13°C to 23°C with a 12 hours light:12 hours dark cycle. During the second experiment, the temperature went from 11° to 26° with an 11 hours light:13 hours dark cycle. The temperature during the third experiment ranged from 14° to 29°C with a 13 hours light:11 hours dark cycle.
3. Results
3.1. Evaluation of Total RNA Extraction Methods
When comparing methods, a higher concentration of total RNA was obtained using tomato and pepper seedlings. Notably, the highest amount in nanograms per microliter was obtained with the CTAB-Trizol protocol, while the lowest concentration was obtained with the RNA isolation Kit (Figure 2).
3.2. Primer Evaluation and Sensitivity Limit
Our results show that using total RNA from tomato and tomatillo plants, the primers designed by Ling et al. (2019) [17] detected the virus up to 0.01ng/𝜇L. In contrast, for eggplant RNA samples, the detection limit was 1 ng/𝜇L. In comparison, the primers of Rodríguez-Mendoza et al. (2018) [15] showed a 0.1 ng/𝜇L detection limit in tomato and 0.01 ng/𝜇L in tomatillo. Notwithstanding either of the primers selection, 1 ng/𝜇L was the sensitivity limit for eggplant samples. Remarkably, the sensitivity limit was increased for tomato and tomatillo using the nested RT-PCR technique and the primers described by Dovas et al. (2004) [1]. Using this set of primers, we identified the virus detection limit as 0.0001 ng/𝜇L in tomato, 0.001 ng/𝜇L in tomatillo, and 1 ng/𝜇L in eggplant (Figure 3).
3.3. Seed Transmission
3.3.1. Collection of Seeds and Germination Test
After mechanical inoculation, seed pods from infected tobacco plants were collected for three to four months, collecting an adequate amount of seeds to perform the tests described in this section. As could be expected, the percent seed germination was higher in seeds obtained from control plants (82%) when compared to those infected with ToBRFV (57.7%) (Table 1).
3.3.2. Percentage of Infection in Seeds and Seedlings
ToBRFV was detected in subsample sizes of 150 and 100 seeds, which were found to have a percentage of infection (P) of 0.61% and 0.22%, respectively. Regarding seedlings, positive groups were detected only when analyzing 150 seedlings and not 100, and the percentage of infection (P) was 0.42% (Figure 4a). In addition, the virus was not detected within subgroups of 100 seeds treated with 3% sodium hypochlorite for three minutes.
3.3.3. Infectivity Tests of Viral Particles in Seeds
Regarding the infectivity tests of viral particles in N. rustica seeds, only three subgroups in tobacco leaves showed symptoms (Figure 4b). They were positive by nested RT-PCR (Figure 4b). This could indicate that positive results in seeds are from viable viral particles and can spread an infection. When analyzing subsamples of 100 seedlings and 100 seeds treated with 3% sodium hypochlorite, no positive groups were detected, nor were symptoms observed in inoculated nicotiana leaves.
3.4. Biological Assays to Test Potential Hosts
Symptoms of virus infection were absent for all the Cucurbitaceae tested species (watermelon, cantaloupe, squash, and cucumber). The absence of virus presence was confirmed by negative results obtained by performing both ELISA and nested RT-PCR techniques (Table 2). In the case of the Solanaceae species, tomato and tobacco-infected plants showed symptoms, and virus presence was confirmed as positive. Interestingly, our results showed that only a percentage of the inoculated tomatillo, pepper, and eggplant displayed symptoms (Figure 5); and were confirmed as infected (Table 2).
4. Discussion
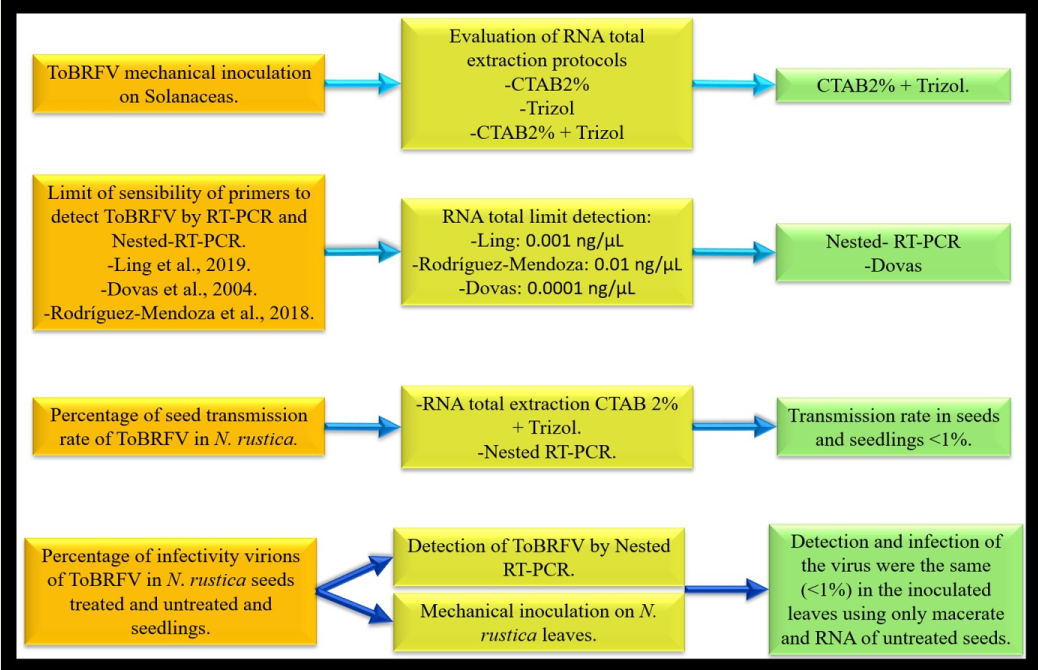
Tobamoviruses can be transmitted by contaminated seeds that are untreated or insufficiently treated with disinfectants to the growing seedling. In this work, we first analyzed virus seed transmission rates and the efficacy of seed disinfection treatments in Nicotiana rustica using a ToBRFV Mexican isolate. Our results showed that the percentage of infection in seeds and seedlings was less than 1% (Figure 6.). Previous experiments demonstrated low seed-to-seedling transmission rates in tomato plants ranging from 0.08% to 1.8% [7,19]. Several disinfection treatments had been tested for ToBRFV removal efficiency from the contaminated seeds. It was found that disinfection treatments such as incubation in sodium hypochlorite solution at 2% for 30 min removed the contaminated ToBRFV virions from the seeds without affecting the germination rate of the treated seeds [7,20,21]. Our experiments identified that sodium hypochlorite solution at 3% was effective as a seed disinfection treatment. Moreover, our findings show that ToBRFV affects the germination of seeds in this host. Although the phenomenon involved is not well known, viruses can affect the physiological performance of the seed [22,23].
Additionally, we compared molecular biology techniques seeking rapid and sensitive virus detection in seed lots (Figure 6). The difference between using subsample sizes of 150 and 100 seeds resulted from an increased viral load when using more seeds, with more positive subgroups being detected. In keeping with this, the virus was not detected within subgroups of 100 seeds after 3% sodium hypochlorite treatment. These results support the proposal that a higher proportion of ToBRFV is found as a contaminant of the seed coat and occasionally of the endosperm [7]. In general, treating seeds with 1-3% sodium hypochlorite (NaOCl) is effective against viruses [8], and it has been corroborated that 2.5% NaOCl for 15 minutes was able to inactivate ToBRFV without affecting seed germination [7]. Nevertheless, the ongoing tobamovirus epidemics demonstrate that treating seeds does not have the desired impact, suggesting that some percentage of the virus is present in the internal tissue of the seeds [5].
Notably, the percentage of a virus transmitted by the seed differs among viral species and variants of the same virus. This percentage can be modified by various factors, including mixed infections, the type of host and the stage of development during which the plant is infected, and the severity and environmental factors that affect both the host and the performance of the virus [24]. In addition, viral accumulation in the host and the transmission type (vectors, contact, seed) can modify the virus's transmission rate. A complex relationship has been observed between viral accumulation in the host and virulence and severity. In keeping with this, more significant viral accumulation cannot be guaranteed to lead to more symptoms [25].
In terms of the efficiency of the vertical transmission of parasites, this is highly dependent on the reproductive potential of the host since the parasite needs to reproduce to infect new individuals [26]. An attenuated virulence could favor the transmission of the virus through the seed. The efficiency of seed transmission has also been associated with greater mobility and multiplication of the virus in the inflorescence [27]. Vertical transmission could be favored by environmental conditions that contribute to reducing the virulence of the virus without controlling the multiplicative level within the plant (tolerance). For example, a higher light intensity favors the accumulation of some viruses in the inflorescence, increasing seed transmission while reducing the host plant's virulence [28].
Nevertheless, of the many factors that influence the percentage of seed transmission, it is known that even if a virus has a ratio as low as 0.001%, this can potentially start an epidemic [24]. In the case of ToBRFV, Davino et al. (2020) [7] found the seed transmission rate was 1.8% and 2.8% in cherry tomatoes.
Regarding the size of the sample and subsamples required to detect ToBRFV, the ISF (2019) [13] recommends analyzing a minimum of 3,000 seeds, which should be divided into subsamples of no more than 250. However, there needs to be more indication as to whether or not this sample size can represent larger seed lots and how they should be sampled. Panno et al. (2020) [4] analyzed 3,000 commercial tomato seeds by lot and divided them into subgroups of 300. This resulted in the detection of ToBRFV in 7% of the lots, and in those cases, all the subgroups were positive. Meanwhile, Fidan et al. (2020) [29] analyzed a lot of 3,000 tomato seeds from symptomatic plants that were artificially infected with ToBRFV, which they divided into 12 subsamples of 250 seeds. They detected the virus in at least one subsample. This work detected the virus in subsamples of 150 and 100 seeds. According to Dombrovsky and Smith (2017) [5], the threshold for detecting Tobamovirus in seeds is one infected out of 249, and 20 subsamples of 100 seeds are required to ensure a 95% likelihood of detecting the virus in minimal infestations of 0.15%.
In the present work, we have observed some advantages and disadvantages that need to be analyzed to determine whether ToBRFV can be detected directly from the seed or the seedling. Seed samples can be immediately investigated, and a more significant number of seeds can be used for each sample processed during extraction, thereby increasing the likelihood of detecting the virus if a few seeds are infected. Nevertheless, non-viable virions, such as contaminants, can be found in the seed, which increases the risk of false positives. In addition, because of the seed tissue's hardness and structural complexity, obtaining a high-quality RNA extraction is more complicated.
While using seedlings makes it possible to increase the replication of the virus and thereby increase the likelihood of detection, it has been found that seedlings remain asymptomatic (with low viral load) and do not express symptoms until handled in intensive production. Meanwhile, analyzing only seedlings without the seed coat eliminates the risk of false positives. The seedling tissue is easier to process, increasing the quality of the extracted RNA. Nonetheless, fewer can be processed since the plant material weighs more in seedlings. There needs to be more information about how many seedlings per subsample need to be processed to detect tobamoviruses.
There are diverse RNA extraction methods whose objective has been focused on obtaining suitable concentrations of the best possible quality since plant tissues may contain high levels of phenolic compounds, polysaccharides, pigments, and RNases [30]. The content of phenols in plants is highly variable and depends on the development stage of plants and the biotic and abiotic environmental factors to which they are exposed. In tomato leaves, the phenol content can decrease with age, but various compounds fluctuate throughout their development [31].
An excellent quality RNA extraction from woody tissues in plants is challenging because they have a high concentration of polysaccharides, polyphenols, and other secondary metabolites. Phenolic compounds bind to proteins and nucleic acids to form large molecular weight complex molecules. Polysaccharides tend to co-precipitate with RNA in the presence of alcohols, remaining as contaminants in the final extraction [32].
The quality and concentration of the RNA obtained during the extractions are critical for accuracy and confidence in plant molecular studies such as RT-PCR [33] and gene expression analysis [34]. However, in mass detection tests, the method's cost will also influence its choice; in this sense, techniques such as CTAB (hexadecyl trimethyl ammonium bromide) and Trizol are cheaper than commercial kits. The CTAB and Trizol methods have also been widely used in various plant species and tissues, with favorable results [33], obtaining good quality and purity and being a fast and economical extraction method for detecting viruses [35]. The CTAB method allows rapid extraction of total RNA from recalcitrant tissue without using toxic chemicals [36]; however, DNA was also extracted by processing the extractions with Trizol afterward, and better quality total RNA extractions can be obtained.
Our results confirmed that the sensitivity limit of each pair of primers was different concerning the plant species. This is probably because the pathogen is expressed and concentrated differently between hosts. In eggplant, the sensitivity limit was the same regardless of the primers used. Regarding tomatillo, the primers of Ling and Rodríguez-Mendoza had the same limit, while for tomato Ling's primers were ten times more sensitive (Figure 3). The Ling primers are designed to amplify the coat protein region, while the Rodríguez-Mendoza primers detected an RdPR region. Magaña-Álvarez et al., (2021) [37] found that primers that amplify the ToBRFV coat protein region have a better performance for virus detection than primers that detect the RdRp region in tomato plants. Nested PCR is known to be more sensitive and reliable than conventional PCR [38]; although it requires more time and reagents to be carried out. As expected, the Dovas primers were 100 times more sensitive with tomato and ten times more sensitive with tomatillo.
The purpose of being able to detect ToBRFV in seeds is to offer producers quality seeds that do not contain more pathogens than the permitted threshold. But the tolerance limits of the virus in seeds depend on certification requirements, and the acceptable level of inoculation is highly debatable. In some cases, the certification of seeds is not affordable or is not available with the infrastructure in developing countries [18]. In the specific case of ToBRFV, given the low percentage of seed transmission that has been reported in tomatoes [7] and considering what we have found in this work (less than 1%), we believe that in addition to certifying seeds through diagnostic tests, it is crucial to verify that all the plants that serve as a source of germplasm are also certified.
Tobamoviruses affect a wide range of species belonging to the Solanaceae and Cucurbitaceae families [5]. In the case of ToBRFV, it has been reported as naturally restricted to tomato and pepper plants. However, it has been possible to experimentally infect multiple plant models, including Nicotiana tabacum, N. benthamiana, N. clevelandii, N. glutinosa, Solanum nigrum, Chenopodium murale, C. amaranticolor, C. quinoa, Petunia hybrid, N. glutinosa, N. occidentalis, N. rustica, and N. sylvestris [39,40]. Although, it has been reported infecting eggplant in Mexico (S. melongena) [40], recent reports indicated not being able to infect this species under controlled conditions, nor were they able to infect potato (S. tuberosum L.) [39].
We tested potential natural hosts in selected Cucurbitaceae and Solanaceae species. None of the Cucurbitaceae species was infected. Our results show that following recent reports, it was experimentally possible to infect N. rustica [40] with ToBRFV. Moreover, its frequent detection in tomato fields in various countries is consistent with 100% infectivity through the inoculation of this species. Interestingly, although natural ToBRFV infections have not been reported in tomatillo, we identified susceptible, infected plants upon experimental transmission. As tomatillo is a commercial crop in Mexico, this evidence highlights the potential risk posed to Mexican national production.
Regarding eggplant bioassays, only the second and third experiments resulted in systemic infection (20% and 100% of the inoculated plants, respectively). This could be related to the differences in temperature (from 23°C to 29°C) along with the photoperiod conditions. In keeping with this, Panno et al. (2019) [39] could not experimentally infect this vegetable species under controlled conditions with 14 hours of light at 20° to 28°C, while Fidan et al. (2020) [29] detected by RT-PCR to the ToBRFV in artificially inoculated eggplants. Still, all were asymptomatic, using the same temperature but a photoperiod of 16 h of light. Variations in temperature and the photoperiod can determine whether the virus successfully infects these plants and causes symptom appearance. For instance, in the case of pepper bioassays, we identified that the percentage of infectivity increased from 20 to 100% as the temperature increased.
Systemic infection is an outcome of multiple factors such as plant age, temperature, the replication and movement of the virus, and RNA silencing [42,43,44]. Regarding temperature, an ideal range has been found for the virus to become established and systemically replicated, which is specific for each viral species and its host. Generally, some viruses need a minimum temperature for systemic infection. A gradual increase in temperature up to a particular maximum will increase the speed of infection and the number of plants infected [45]. In addition, symptoms can be mitigated at high temperatures since the replication of the genomic RNA decreases, resulting in less viral replication and movement [46].
5. Conclusions
The ToBRFV Mexico strains did not infect cucurbit species in our study. However, they were capable of experimentally infecting tomatillo and eggplant. We found that the best protocol to detect the ToBRFV under these experimental conditions was CTAB-Trizol and the nested RT-PCR using the primers described by Dovas et al. (2004) [1]. The percentage of ToBRFV-infected N. rustica seeds was less than 1%. For the seedlings analysis, the subgroup's size required was found to be at least 150 for the virus to be detected by nested RT-PCR. We recognize that detecting a virus in seeds is very complex when the transmission percentage is low. The success of detecting the virus in seeds, therefore, depends not only on the percentage of virus transmission in a specific host, which is influenced by all the factors mentioned previously but also on the detection technique and the sample type (size of samples and subsamples). In addition, the ISF (2019) [13] protocol specifies that seeds that are analyzed for the detection of ToBRFV should not be treated with any disinfectants. Nevertheless, when seeds are imported or exported, seed companies generally submit them to chemical treatments to prevent phytosanitary problems with germination. For this reason, it is better to analyze recently emerged seedlings.
Author Contributions
Conceptualization, E.J.Z.-M. and K.A.-P.; Methodology, E.J.Z.-M. and C.Y.C.-C.; Formal Analysis, E.J.Z.-M. and D.L.O.-M.; Investigation, E.J.Z.-M. and C.Y.C.-C.; Resources, E.J.Z.-M. and D.L.O.-M.; Writing – Original Draft Preparation, K.A.-P., E.J.Z.-M.; Writing – Review & Editing, HRW, K.A.-P., D.L.O.-M.; Visualization, E.J.Z.-M., K.A.-P.; Supervision, K.A.-P., R.W.H.; Project Administration, R.W.H.; Funding Acquisition, K.A.-P., R.W.H. All authors have read and agreed to the published version of the manuscript.
Funding
K.A.P received financial support from the Fulbright García-Robles Foundation and is a current holder of a fellowship from CONACyT [CVU:227919].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge the Agricultural Parasitology Department of Chapingo University for the facilities to conduct the experiments.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
- (A)
-
Trizol protocol:
- Macerate 100 mg of plant tissue with liquid nitrogen and add it immediately to an Eppendorf tube with 1 mL of cold Trizol. Give a pulse of the vortex and leave them for 5 minutes on ice.
- Add 400 ml of chloroform and mix by inversion 7 times. Leave the tube on ice for 10 minutes.
- Centrifuge by 10 minutes at 13,000 rpm at 4°C.
- Recover carefully the supernatant (approx. 400 µL) and transfer it to a clean Eppendorf tube.
- Add 1.5 V of cold isoamyl alcohol and leave the tube at -20°C for 20 minutes.
- Centrifuge by 15 minutes at 13,000 rpm at 4°C.
- Discard the alcohol taking care of the pellet at the bottom of the tube.
- Wash the pellet with 1 mL of cold ethanol at 90% and centrifuge again for 5 minutes at 13,000 rpm at 4°C.
- Discard all the ethanol, take care of the pellet, and let it dry at room temperature.
- Resuspend and dissolve the pellet in 30 to 50 µL of RNAsa-free distilled water.
- (B)
-
CTAB 2% protocol:
- Macerate 100 mg of plant tissue with liquid nitrogen and add it immediately on an Eppendorf tube with 700 mL of CTAB 2% + PVP (1%) + BME (0.2%). Give a pulse with a vortex.
- Incube at 55°C in a water bath for 10 minutes.
- Add 470 ml of cold chloroform: isoamyl alcohol (24:1) and mix by inversion 7 times.
- Centrifuge by 10 minutes at 13,000 rpm at 4°C.
- Recover carefully the supernatant (approx. 400 µL) and transfer it to a new Eppendorf tube.
- Add 1 V of cold isoamyl alcohol and 1/10 V of sodium acetate (3M) and mix by inversion. Left the tube at -20°C for 20 minutes.
- Centrifuge by 15 minutes at 13,000 rpm at 4°C.
- Discard the liquid taking care of the pellet at the bottom of the tube.
- Wash the pellet with 1 mL of cold ethanol at 90% and centrifuge again for 5 minutes at 13,000 rpm at 4°C.
- Discard all the ethanol, take care of the pellet, and let it dry at room temperature.
- Resuspend and dissolve the pellet in 50 to 100 µL of RNAsa-free distilled water.
- (C)
-
CTAB2% + Trizol
- Follow steps 1 to 5 of section (B) of Appendix 1.
- Add 500 µL of Trizol and mix by inversion 7 times.
- Follow steps 2 to 10 of section (A) of Appendix 1.
References
- Dovas, C.I.; Efthimiou, K., Katis, N.I. Generic detection and differentiation of tobamoviruses by a spot nested RT-PCR-RFLP using dI-containing primers along with homologous dG-containing primers. Journal of Virological Methods 2004, 117, 137–144. [CrossRef]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela N.; Abu-Ras, A.: Ezra, N.; Haberman, A.; Yitzhak L.; Lachman, O.; Dombrovsky, A. A New Israeli Tobamovirus Isolate Infects Tomato Plants Harboring Tm-22 Resistance Genes. PloS One 2017, 12(1), e0170429. PMID: 28107419. [CrossRef]
- Levitzky, N.; Smith, E.; Lachman, O.; Luria, N.; Mizrahi, Y.; Bakelman, H.; Sela, N.; Laskar, O.; Milrot, E.; Dombrovsky, A. The bumblebee Bombus terrestris carries a primary inoculum of Tomato brown rugose fruit virus contributing to disease spread in tomatoes. PLoS ONE 2019, 14(1), e0210871. [CrossRef]
- Panno, S.; Caruso, G.A.; Stefano, B.; Lo Bosco Giosuè, E. R.A.; Salvatore, D. Spread of Tomato Brown Rugose Fruit Virus in Sicily and Evaluation of the Spatiotemporal Dispersion in Experimental Conditions. Agronomy 2020, 10, 834. [CrossRef]
- Dombrovsky, A., Smith, E., 2017. Seed Transmission of Tobamoviruses: Aspects of Global Disease Distribution. In: Advances in seed biology, Jimenez-Lopez, J.C., Ed. IntechOpen. [CrossRef]
- Roberts, A.G., 2014. Plant Viruses: Soil-borne. In: eLS. John Wiley & Sons, Ltd: Chichester. 12 p. [CrossRef]
- Davino, S.; Caruso, A.G.; Bertacca, S.; Barone, S.; Panno, S.. Tomato Brown Rugose Fruit Virus: Seed Transmission Rate and Efficacy of Different Seed Disinfection Treatments. Plants 2020. 9, 1615. [CrossRef]
- Herrera-Vásquez, J.A.; Córdoba-Sellés, M.C.; Cebrián, M.C.; Alfaro-Fernandez, A.; Jordá, C. Seed transmission of Melon necrotic spot virus and efficacy of seed-disinfection treatments. Plant Pathology 2009. 58, 7. [CrossRef]
- Rast, A. Th. B.; Stijger, C.C.M.M. Disinfection of pepper seed infected with different strains of capsicum mosaic virus by trisodium phosphate and dry heat treatment. Plant Pathology 1987, 36 (4), 583-588. [CrossRef]
- Choi, G.S. Occurrence of two tobamovirus diseases in cucurbits and control measures in Korea. Plant Pathology Journal 2001. 17, 243–248. https://www.ppjonline.org/upload/pdf/PPJ021-03-09.pdf.
- Li, J.X.; Liu, S.S.; Gu, Q.S. Transmission efficiency of Cucumber green mottle mosaic virus via seeds, soil, pruning and irrigation water. Journal of Phytopathology 2015, 164(5), 300-309. [CrossRef]
- Kumar, S.; Udaya, S.A.; Nayaka, S.; Lund, O.; Prakash, H. Detection of Tobacco mosaic virus and Tomato mosaic virus in pepper and tomato by multiplex RT–PCR. Letters in Applied Microbiology 2011, 53(3), 359-363. [CrossRef]
- International Seed Federation (ISF), 2019. https://www.worldseed.org/wp-content/uploads/2019/09/Tomato-ToBRFV_2019.09.pdf (accessed on 10 August 2021).
- SENASICA (Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria). 2019. Dirección general de sanidad vegetal- Centro nacional de referencia fitosanitaria, Protocolo de Diagnóstico: Tomato brown rugose fruit virus (ToBRFV). Versión 2.0. http://sinavef.senasica.gob.mx/CNRF/AreaDiagnostico/DocumentosReferencia/Documentos/ProtocolosFichas/Protocolos/VirusFitopatogenos/PD%20ToBRFV%20V.2%20PUB.pdf (accesed on 15 August 2021).
- Rodríguez-Mendoza, J.; García-Ávila, C.J.; López-Buenfil, J.A.; Araujo-Ruiz, K.; Quezada-Salinas, A.; Cambrón-Crisantos, J.M. Identification of Tomato brown rugose fruit virus by RT-PCR from a coding region of replicase (RdRP). Mexican Journal of Phytopathology 2018, 37(2), 1-12. [CrossRef]
- Jordon-Thaden, I.E.; Chanderbali, A.S.; Gitzendanner, M.A.; Soltis D.E. Protocol note. Modified CTAB and Trizol protocols improve RNA extraction from chemically complex embryophyta. Applications in Plant Sciences 2015, 3(5), 1400105. [CrossRef]
- Ling, K.S.; Tian, T.; Gurung, S.; Salati, R.; Gilliard, A. First report of Tomato brown rugose fruit virus infecting greenhouse tomato in the United States. Plant Disease 2019, 103, 1439. [CrossRef]
- Albrechtsen, S.E. Testing methods for seed transmitted viruses: principles and protocols. Publisher: CABI, Wallingford, 2006. 268 p.
- Salem, N.M.; Sulaiman, A.; Samarah, N.; Turina, M.; Vallino, M. Localization and mechanical transmission of Tomato Brown Rugose Fruit Virus in tomato seeds. Plant Disease 2021, 106, 275-281. [CrossRef]
- Samarah, N.; Sulaiman, A.; Salem, N.M.; Turina, M. Disinfection treatments eliminated tomato brown rugose fruit virus in tomato seeds. European Journal of Plant Pathology 2021, 159, 153–162. [CrossRef]
- Chanda, B.; Shamimuzzaman, M.D.; Gilliard, A.; Ling, K.S. Effectiveness of disinfectants against the spread of tobamoviruses: Tomato brown rugose fruit virus and Cucumber green mottle mosaic virus. Virology Journal 2021. 18, 7. [CrossRef]
- Marodin, J.C.; Resende, F.V.; Gabriel, A.; Souza, R.J.De; Resende, J.T.V. De; Camargo, C.K.; Zeist, A.R. Agronomic performance of both virus-infected and virus free garlic with different seed bulbs and clove sizes. Pesquisa Agropecuária Brasileira 2019, 54, e01448. [CrossRef]
- Hemmati, K.; McLean, D. L. Gamete-Seed Transmission of Alfalfa Mosaic Virus and Its Effect on Seed Germination and Yield in Alfalfa Plants. Disease Detection and Losses. AMV in alfalfa seed. Phytopathology 1977, 67, 576-579. https://www.apsnet.org/publications/phytopathology/backissues/Documents/1977Articles/Phyto67n05_576.pdf.
- Mohan, B.G., Baruah, G., Sen, P., Deb, N.P., Kumar, B.B. Host-Parasite Interaction During Development of Major Seed-Transmitted Viral Diseases. In: Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management; Kumar, R.A., Gupta, Eds., Springer, 2020; pp.265-289. [CrossRef]
- Froissart, R.; Doumayrou, J.; Vuillaume, F.; Alizon, S.; Michalakis, Y. The virulence –transmission trade-off in vector-borne plant viruses: a review of (non-)existing studies. Philosophical Transactions of the Royal Society B. 2010, 365, 1907–1918. [CrossRef]
- Lipsitch, M.; Nowak, M.A.; Ebert, D.; May, R.M. The population dynamics of vertically and horizontally transmitted parasites. Proceedings of the Royal Society B: Biological 1995, 260, 321–327. [CrossRef]
- Cobos, A.; Montes, N.; López-Herranz, M.; Gil-Valle, M.; Pagán, I. Within-Host Multiplication and Speed of Colonization as Infection Traits Associated with Plant Virus Vertical Transmission. Journal of virology 2019, 93(23), e01078-19. [CrossRef]
- Montes, N.; Pagán, I. Light Intensity Modulates the Efficiency of Virus Seed Transmission through Modifications of Plant Tolerance. Plants 2019, 8 (304), 15p. [CrossRef]
- Fidan, H.; Pelin, S.; Kubra, Y.; Bengi, T.; Gozde, E.; Ozer, C. Robust molecular detection of the new Tomato brown rugose fruit virus in infected tomato and pepper plant from Turkey. Journal of Integrative Agriculture 2020, 19, 2–11. [CrossRef]
- Wang, H.M.; Yin, W.C.; Wang, C.K.; To, K.Y. Isolation of functional RNA from different tissues of tomato suitable for developmental profiling by microarray analysis. Botanical Studies 2009, 50(2), 115-125. https://ejournal.sinica.edu.tw/bbas/content/2009/2/Bot502-01.pdf.
- Dadáková, K.; Heinrichová, T.; Lochman, J.; Kašparovský, T. Production of Defense Phenolics in Tomato Leaves of Different Age. Molecules 2020, 25(21), 4952. [CrossRef]
- Salzman, R.A.; Fujita, T.; Zhu-Salzman, K.; Hasegawa, P.M.; Bressan, R.A. An improved RNA isolation method for plant tissues containing high levels of phenolic compounds or carbohydrates. Plant Molecular Biology Report 1999, 17, 11-17. [CrossRef]
- Gambino, G.; Perrone, I.; Gribaudo, I. A Rapid and effective method for RNA extraction from different tissues of grapevine and other woody plants. Phytochemical Analysis 2008, 19(6), 520-5. [CrossRef]
- Toni, L.S.; Garcia, A.M.; Jeffrey, D.A.; Jiang, X.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Optimization of phenol-chloroform RNA extraction. MethodsX 2018, 5, 599-608. [CrossRef]
- Mathioudakis, Μ.Μ.; Saponari, M.; Hasiów-Jaroszewska B.; Elbeaino, T.; Koubouris, G. Detection of viruses in olive cultivars in Greece, using a rapid and effective RNA extraction method, for certification of virus-tested propagation material. Phytopathologia Mediterranea 2020, 59(1), 203-211. [CrossRef]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Molecular Biology Reporter 1993, 11, 113–116. [CrossRef]
- Magaña-Álvarez, A.A.; Pérez-Brito, D.; Vargas-Hernández, B.Y.; Ramírez-Pool, J.A.; Núñez-Muñoz, L.A.; Salgado-Ortiz, H.; de la Torre-Almaraz R.; Ruiz- Medrano R.; Xoconostle-Cázares, B. Detection of Tomato brown rugose fruit virus (ToBRFV) in solanaceous plants in Mexico. Journal of Plant Disease Protection 2021, 128, 1627–1635. [CrossRef]
- Shin, Y.; Cho, K.; Cho, H.; Kang, S.; Kim, H.; Kim, Y.; Park, N. Comparison of one-step RT-PCR and a nested PCR for the detection of canine distemper virus in clinical samples. Australian Veterinary Journal 2004, 82(1-2), 83–86. http://dx.doi.org/10.1111/j.1751-0813.2004.tb14651.x.
- Panno, S.; Ruiz-Ruiz, S.; Caruso, A.G.; Alfaro-Fernandez, A.; San Ambrosio, M.I.; Davino, S. Real-time reverse transcription polymerase chain reaction development for rapid detection of Tomato brown rugose fruit virus and comparison with other techniques. PeerJ 2019, 7, e7928. [CrossRef]
- Oladokun, J.; Halabi, M.; Barua, P.; Nath, P.J.P.P. Tomato brown rugose fruit disease: current distribution, knowledge and future prospects. Plant Pathology 2019, 68, 1579-1586. [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). 2019. Reporting Service 2019/192. https://gd.eppo.int/reporting/article-6622 (accessed on 10 January 2022).
- Fajinmi, A. A.; Fajinmi, O. B. Incidence of Okra mosaic virus at different growth stages of okra plants (Abelmoschus esculentus (L.) Moench) under tropical condition. Journal of General and Molecular Virology 2010, 2, 28-3.
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, C.M. Effect of temperature on geminivirus-induced RNA Silencing in Plants. Plant Physiology 2005, 138, 1828–1841. [CrossRef]
- Zhang, X.; Singh, J.; Li, D.; Qua, F. Temperature- dependent survival of Turnip crinkle virus-infected Arabidopsis plants relies on an RNA silencing-based defense that requires DCL2, AGO2, and HEN1. Journal of Virolgy, 2012, 12, 6847–6854. [CrossRef]
- Chung, B.N.; Kyung, S.C.; Jeong, J.A.; Jae, H.J.; Ki, S.D.; Kyo-Sun, P. Effects of Temperature on Systemic Infection and Symptom Expression of Turnip mosaic virus in Chinese cabbage (Brassica campestris). Journal of Plant Pathology 2015, 31(4), 363-370. http://dx.doi.org/10.5423/PPJ.NT.06.2015.0107.
- Szittya, G.; Silhavy, D.; Molnár, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Bánfalvi, Z.; Burgyán, J. Low temperature inhibits RNA silencing-mediated defense by the control of siRNA generation. EMBO Journal 2003, 22, 633–640. [CrossRef]
Figure 1.
(a) Procedure to determine the percentage of infection in seeds and seedlings. M1 and M2 represent the mock-inoculated groups. 1-15 depict the fifteen seeds and seedlings groups; (b) Evaluation of the infectivity of viral particles in seeds. M1 and M2 are the mock-inoculated groups. 1-15 depicts the fifteen seeds and seedlings groups. The seed groups were treated with 3% sodium hypochlorite solution for 3 minutes.
Figure 1.
(a) Procedure to determine the percentage of infection in seeds and seedlings. M1 and M2 represent the mock-inoculated groups. 1-15 depict the fifteen seeds and seedlings groups; (b) Evaluation of the infectivity of viral particles in seeds. M1 and M2 are the mock-inoculated groups. 1-15 depicts the fifteen seeds and seedlings groups. The seed groups were treated with 3% sodium hypochlorite solution for 3 minutes.
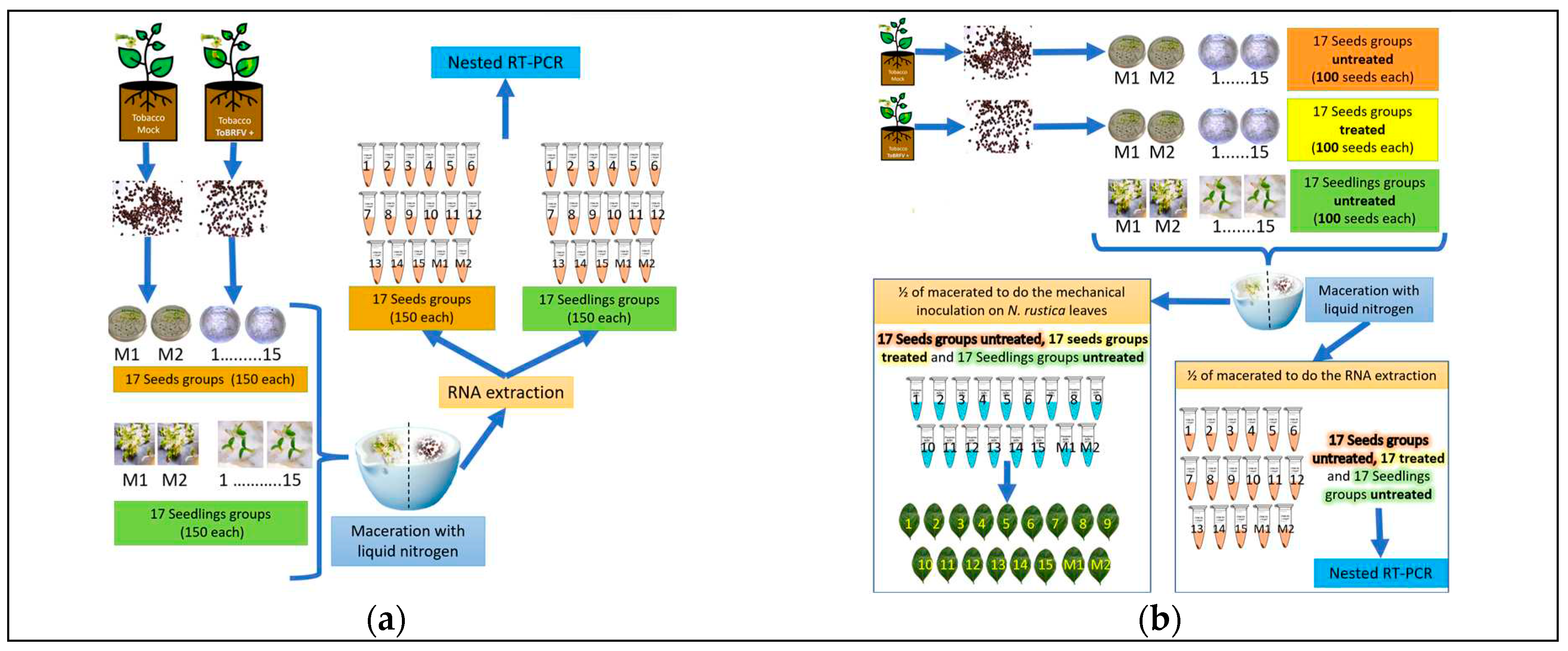
Figure 2.
Evaluation of total RNA extraction methods and its Nanodrop 2000® lectures of each sample.
Figure 2.
Evaluation of total RNA extraction methods and its Nanodrop 2000® lectures of each sample.
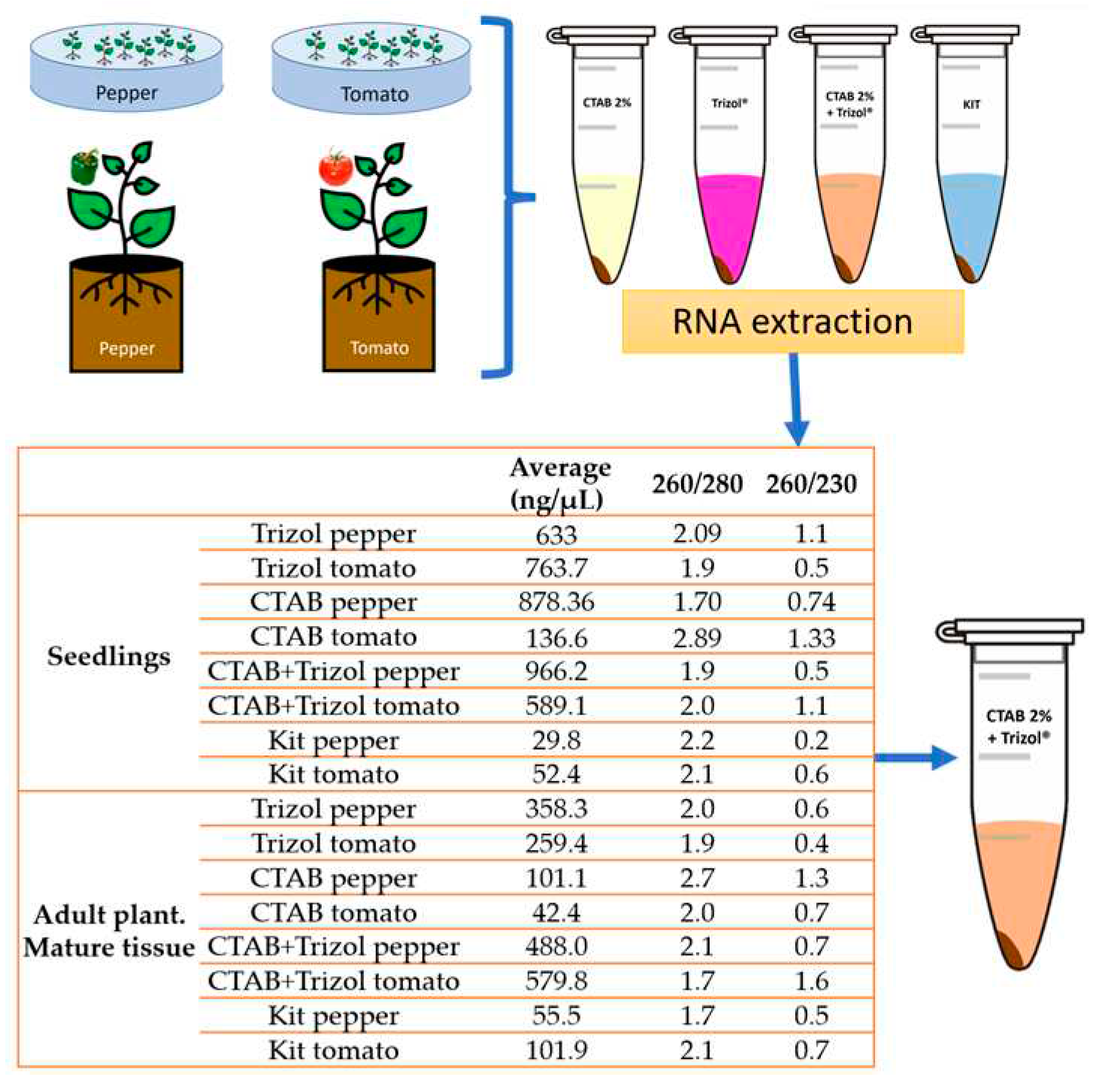
Figure 3.
Evaluation and sensitivity of PCR primers. 1.5% agarose gels electrophoretic analysis of RT-PCR and Nested RT-PCR products (Ling´s primers expected size 842 bp; Rodríguez-Mendoza´s primers expected size 475 bp and Dovas´s primers expected size 400 pb). (-): sterilized water instead RNA. 100pb= 100bp DNA Ladder (Invitrogen®). 1Kb= 1000 bp DNA ladder (Promega®). 10-3 and 10-4 = 0.001 and 0.0001 ng/µL.
Figure 3.
Evaluation and sensitivity of PCR primers. 1.5% agarose gels electrophoretic analysis of RT-PCR and Nested RT-PCR products (Ling´s primers expected size 842 bp; Rodríguez-Mendoza´s primers expected size 475 bp and Dovas´s primers expected size 400 pb). (-): sterilized water instead RNA. 100pb= 100bp DNA Ladder (Invitrogen®). 1Kb= 1000 bp DNA ladder (Promega®). 10-3 and 10-4 = 0.001 and 0.0001 ng/µL.
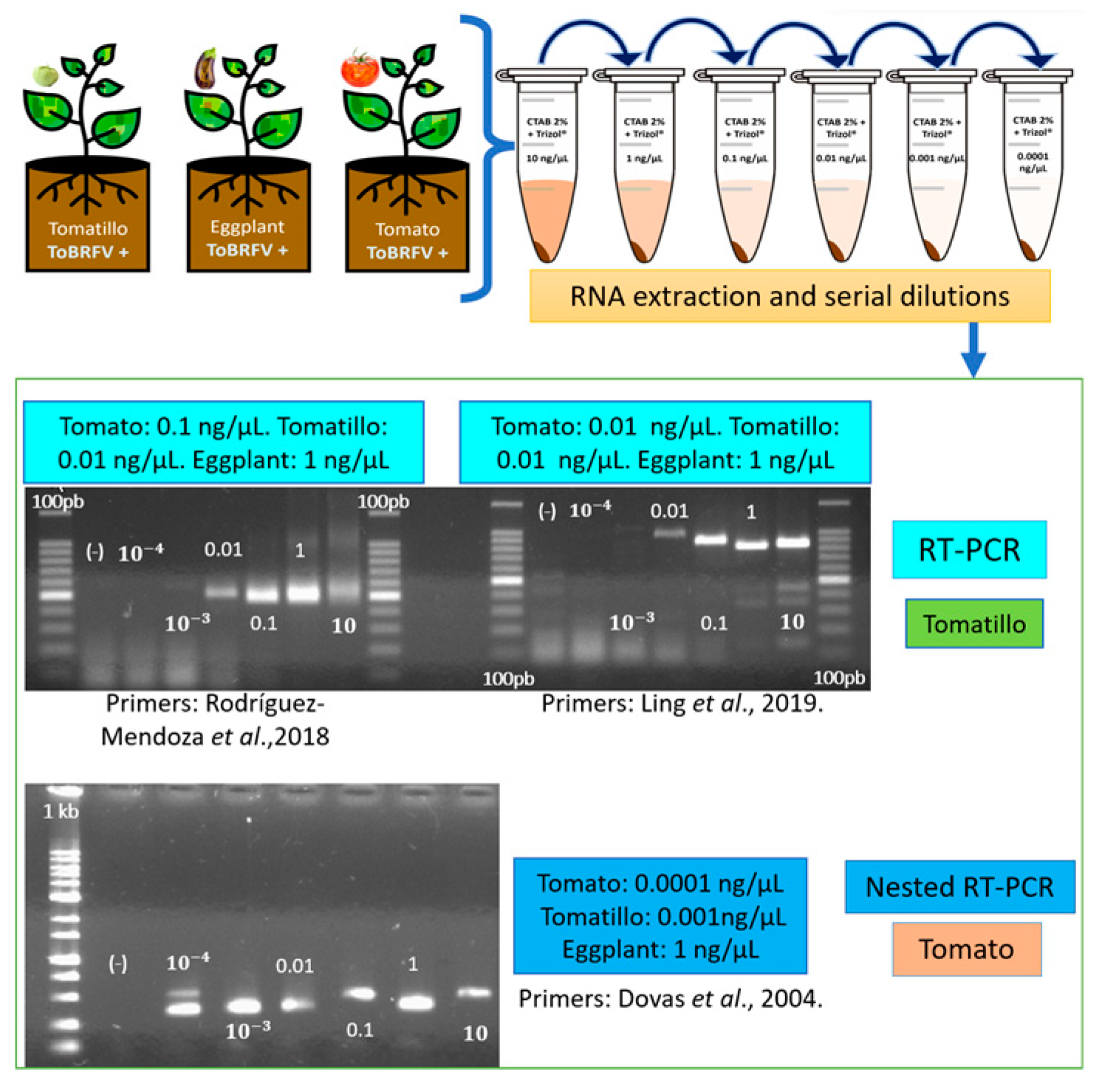
Figure 4.
(a) Nicotiana rustica seeds/seedlings obtained from ToBRFV infected plants to determine the percentage of seed transmission P= [1- (Y/N)^1/n], where N= number of subgroups; n= number of seeds/seedlings in each subgroup, Y= number of negative subgroups by nested RT-PCR. NT= not tested.*Treated with 3% sodium hypochlorite solution for 3 minutes. 1.5% agarose gel electrophoretic analysis of nested RT-PCR products (size expected= 400 pb) obtained from RNAt extracted from subgroups (1-15) of 150 Nicotiana rustica seeds, and 150 seedlings obtained from ToBRFV infected plants; M1 and M2 were obtained from non-infected plants. + = positive control. 100pb= 100bp DNA Ladder (Invitrogen®); (b) N. rustica leaves inoculated with the macerated seed used to get the RNAt of the subgroups of 100 N. rustica seeds obtained from ToBRFV infected plants; the narrow shows the chlorotic lesions. 1.5% agarose gel electrophoretic analysis of nested RT-PCR products (size expected= 400 pb) obtained from the same RNAt extracted from subgroups (1-15) of 100 Nicotiana rustica seeds obtained from ToBRFV infected plants; M1 and M2 were obtained from non-infected plants. + = positive control. 100pb= 100bp DNA Ladder (Invitrogen®).
Figure 4.
(a) Nicotiana rustica seeds/seedlings obtained from ToBRFV infected plants to determine the percentage of seed transmission P= [1- (Y/N)^1/n], where N= number of subgroups; n= number of seeds/seedlings in each subgroup, Y= number of negative subgroups by nested RT-PCR. NT= not tested.*Treated with 3% sodium hypochlorite solution for 3 minutes. 1.5% agarose gel electrophoretic analysis of nested RT-PCR products (size expected= 400 pb) obtained from RNAt extracted from subgroups (1-15) of 150 Nicotiana rustica seeds, and 150 seedlings obtained from ToBRFV infected plants; M1 and M2 were obtained from non-infected plants. + = positive control. 100pb= 100bp DNA Ladder (Invitrogen®); (b) N. rustica leaves inoculated with the macerated seed used to get the RNAt of the subgroups of 100 N. rustica seeds obtained from ToBRFV infected plants; the narrow shows the chlorotic lesions. 1.5% agarose gel electrophoretic analysis of nested RT-PCR products (size expected= 400 pb) obtained from the same RNAt extracted from subgroups (1-15) of 100 Nicotiana rustica seeds obtained from ToBRFV infected plants; M1 and M2 were obtained from non-infected plants. + = positive control. 100pb= 100bp DNA Ladder (Invitrogen®).
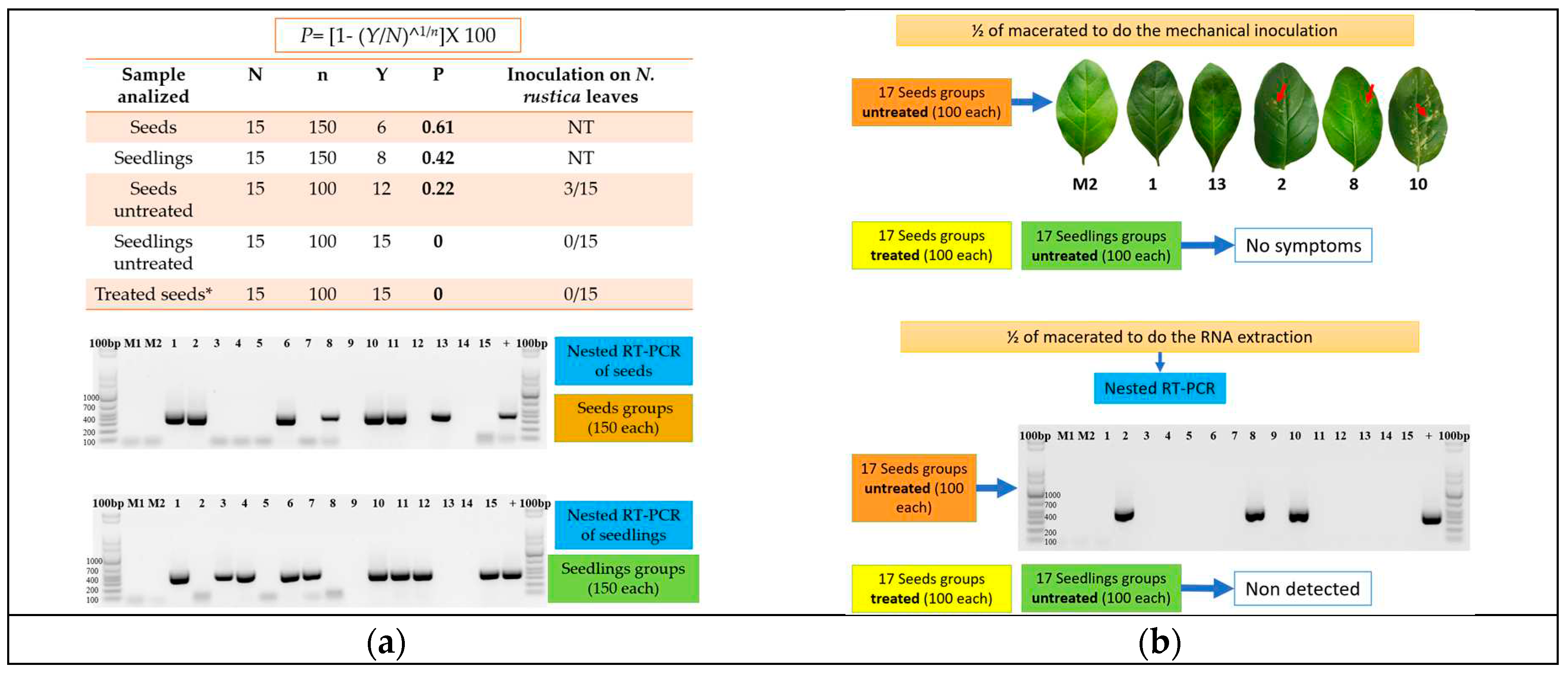
Figure 5.
Symptoms in tomato (1-2A), tobacco (1-2B), tomatillo (1-2C), pepper (1D), and eggplant (1-2 E) infected with ToBRFV after mechanical transmission. Plants showed narrowing (1A, 1C), mosaic (2A), necrosis on stem and leaves (1E, 2E), chlorosis (1B, 2B), chlorotic lesions (2C), and stunting (1D). Mock-inoculated (3A,3B, 3C, 2D and 3E).
Figure 5.
Symptoms in tomato (1-2A), tobacco (1-2B), tomatillo (1-2C), pepper (1D), and eggplant (1-2 E) infected with ToBRFV after mechanical transmission. Plants showed narrowing (1A, 1C), mosaic (2A), necrosis on stem and leaves (1E, 2E), chlorosis (1B, 2B), chlorotic lesions (2C), and stunting (1D). Mock-inoculated (3A,3B, 3C, 2D and 3E).

Figure 6.
General procedures of inoculation and detection of ToBRFV in N. rustica seeds and seedlings and the main results obtained in this investigation.
Figure 6.
General procedures of inoculation and detection of ToBRFV in N. rustica seeds and seedlings and the main results obtained in this investigation.
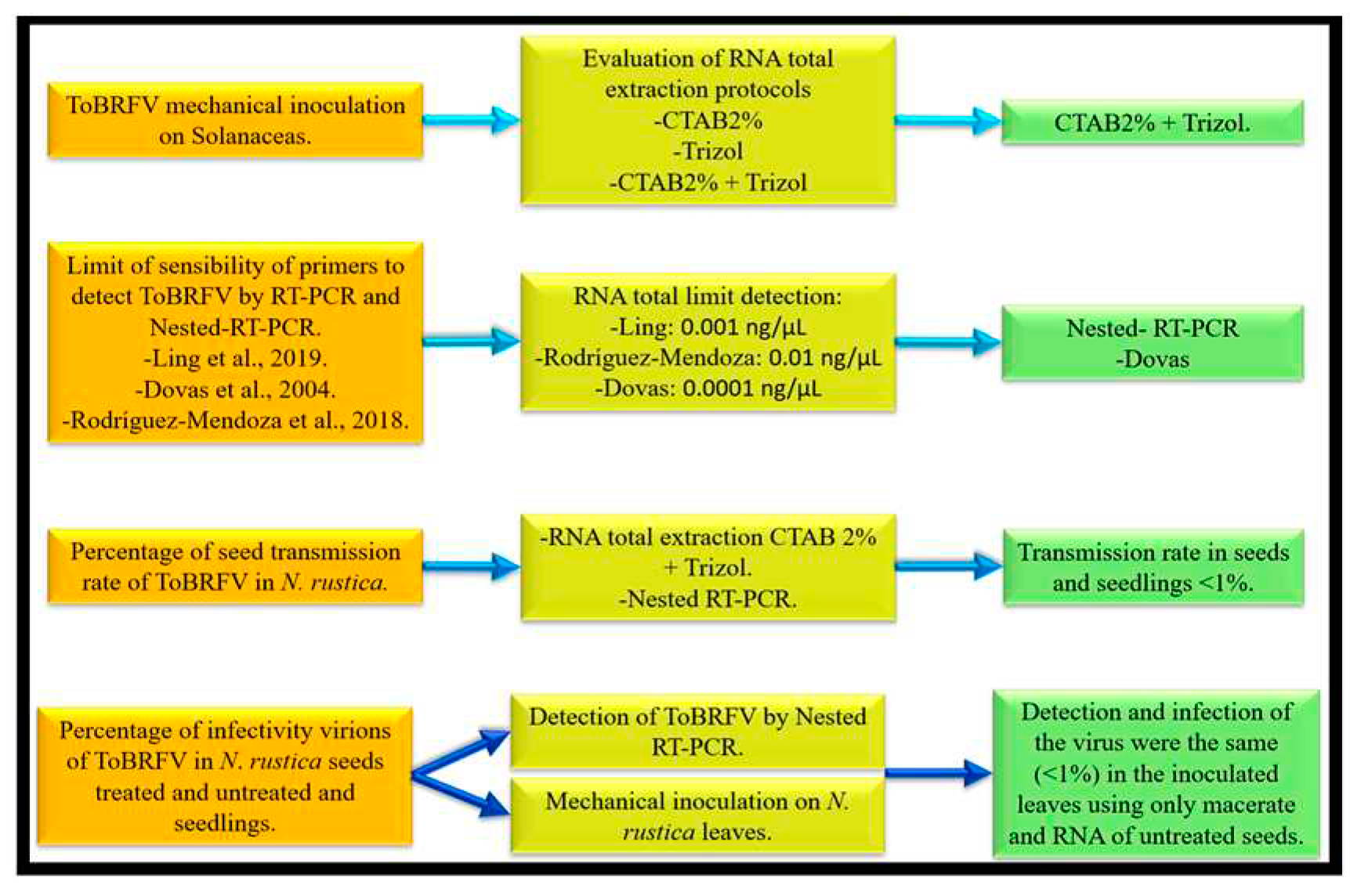

Table 1.
Germination of N. rustica seeds from healthy plants and those infected with ToBRFV.
| Number of Germinated Seeds | ||
|---|---|---|
| Repetition | +ToBRFV | Healthy |
| 1 | 58 | 92 |
| 2 | 56 | 87 |
| 3 | 64 | 88 |
| 4 | 56 | 75 |
| 5 | 74 | 88 |
| 6 | 52 | 82 |
| 7 | 46 | 44 |
| 8 | 63 | 89 |
| 9 | 51 | 93 |
Table 2.
Analysis by nested RT-PCR and ELISA of ToBRFV inoculated plants.
| Inoculated Specie | Nested RT-PCR | ELISA |
|---|---|---|
| Watermelon (Citrullus lanatus) | 0*, 0°, 0" | 0, 0, 0 |
| Cantaloupe (Cucumis melo) | 0, 0, 0 | 0, 0, 0 |
| Squash (Cucurbita pepo) | 0, 0, 0 | 0, 0, 0 |
| Cucumber (Cucumis sativus) | 0, 0, 0 | 0, 0, 0 |
| Pea (Pisum sativum) | 0, 0, 0 | 0, 0, 0 |
| Tomato (Solanum lycopersicum) | 5, 5, 5 | 5, 5, 5 |
| Tomatillo (Physalis ixocarpa) | 3, 4, 3 | 3, 4, 3 |
| Tobacco (Nicotiana rustica) | 5, 5, 5 | 5, 5, 5 |
| Pepper (Capsicum annum) | 2, 4, 5 | 2, 4, 5 |
| Eggplant (Solanum melongena) | 0, 1, 5 | 0, 1, 5 |
Number of positive plants in the first (*), second (°) and third (“) experimental repetition. All the mock-inoculated plants were negative. Plants were analyzed after 25 dpi.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



