
Submitted:
19 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
Non-alcoholic fatty liver disease (NAFLD) is a liver disorder that has become a global health concern due to its increasing prevalence. Currently, there is a need for reliable biomarkers to aid in the diagnosis and prognosis of NAFLD. Extracellular vesicles (EVs) are promising candidates in biomarker discovery, as they carry proteins that reflect the pathophysiological state of the liver. In this review, we developed a list of EV proteins that could be used as diagnostic biomarkers for NAFLD. We employed a multi-step strategy that involved reviewing and comparing various sources of information. Firstly, we reviewed papers that have studied EVs proteins as biomarkers in NAFLD, as well as papers that have studied circulating proteins as biomarkers in NAFLD. To further identify potential candidates, we utilized the EV database Vesiclepedia.org to qualify each protein. Finally, we consulted the Human Protein Atlas to search for candidates' localization, focusing on membrane proteins. By integrating these sources of information, we developed a comprehensive list of potential EVs membrane protein biomarkers that could aid in the diagnosis and monitoring of NAFLD. In conclusion, our multi-step strategy for identifying EV-based protein biomarkers for NAFLD provides a comprehensive approach that can also be applied for other diseases. The protein candidates identified through this approach could have significant implications for the development of non-invasive diagnostic tests for NAFLD and improve the management and treatment of this prevalent liver disorder.
Keywords:
Non-alcoholic fatty liver
; Extracellular vesicles
; Biomarkers
; Surface proteins
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is the most common liver disorder globally, with a prevalence of up to 25% in the general population [1]. It is characterized by hepatic steatosis in the absence of significant alcohol intake. NAFLD ranges from non-alcoholic fatty liver (NAFL) to non-alcoholic steatohepatitis (NASH) and liver fibrosis or cirrhosis, with the latter having the highest risk of developing liver-related morbidity and mortality [1]. Therefore, due to its prevalence and potential severity, there is a need for biomarkers that can accurately diagnose, predict disease progression, and monitor response to therapy, particularly for patients with NAFL who are at risk of developing NASH. Liver biopsy has traditionally been the gold standard for the diagnosis and staging of NAFLD, but its invasiveness, cost, and potential complications have led to a search for non-invasive biomarkers [2]. Based on liver biopsies analysis, the grading and staging of NAFLD are crucial for the diagnosis, management, and prognosis of the disease. The grading system classifies the severity of the histological features of NAFLD, including steatosis, ballooning, and inflammation. The staging system assesses the extent of liver fibrosis, which is the most important prognostic factor in NAFLD [3] (Figure 1).
A variety of biomarkers have been proposed. However, none has been universally accepted as a reliable diagnostic tool for NAFLD [4]. Biomarkers for NAFLD can be broadly categorized into three types: (I) imaging biomarkers, (II) circulating biomarkers, and (III) genetic biomarkers. Imaging biomarkers, such as liver fat percentage by Magnetic Resonance Imaging (MRI) using proton density fat fraction (PDFF), liver stiffness measurement and magnetic resonance elastography, have shown promise in the diagnosis and staging of NAFLD [5]. However, these methods are costly and not widely available, limiting their widespread use.
Circulating biomarkers have been extensively studied in NAFLD, with many potential candidates identified. Nevertheless, these biomarkers have limitations in relation with their sensitivity/specificity, predictive value, and standardization. Although widely used and standardized worldwide, the circulating biomarkers Alanine aminotransferase (ALT), Aspartate aminotransferase (AST), and Gamma-glutamyl transferase (GGT) have limitations in their ability to specifically diagnose NAFLD [6]. These biomarkers can indicate liver dysfunction, but they cannot differentiate between various liver diseases or specifically identify NAFLD.
Fatty acid binding protein 4 (FABP4), cytokeratin 18 (CK-18), and fibroblast growth factor 21 (FGF21) have all emerged as potential circulating biomarkers for NAFLD. While they offer improved specificity compared with traditional liver enzymes, they have limitations that need to be addressed. These biomarkers cannot differentiate between different types of liver damage and can be elevated in other conditions that share risk factors with NAFLD [5]. Furthermore, they are still in the research stage and lack diagnostic approval. Future studies should focus on validating these biomarkers in large cohorts, establishing their diagnostic accuracy, and developing standardized protocols for their utilization in clinical practice.
Novel biomarkers that can overcome these limitations are needed to improve the accuracy and specificity of NAFLD diagnosis and management.
Predictive algorithms, such as the fibrosis-4 (FIB-4) index, fibrotic NASH index (FNI), and Forns score, have emerged as potential tools for the diagnosis of NAFLD [7,8]. These algorithms utilize readily available clinical parameters (such as ALT, Body mass index, AST) to predict the presence or severity of NAFLD, offering advantages in terms of cost, accessibility, and standardization. However, despite their utility, there are certain limitations that need to be addressed, including issues related to specificity and the need for further validation in large cohorts.
Genetic biomarkers have also been investigated in NAFLD, with genome-wide association studies (GWAS) identifying several loci associated with NAFLD susceptibility and progression [4]. The scope of these approaches is not related with the purpose of this review.
Extracellular vesicles (EVs) are a heterogeneous population of vesicles that differ in size, content, and function [9]. They are classified based on their biogenesis and size into three main categories: exosomes (30-150 nm), microvesicles (150-1000 nm), and apoptotic bodies (>1000 nm) (Figure 2). EVs are secreted by various cell types, including liver cells, and can be found in various body fluids, such as blood, urine, and bile. EVs are now recognized as critical players in cell-to-cell communication, as they carry various biomolecules that can be delivered to target cells [9].
EVs' role in NAFLD has been widely investigated [10], and several studies have suggested that EVs can be used as biomarkers for the diagnosis and monitoring of NAFLD [11]. The cargo of EVs secreted by liver cells, including hepatocytes, cholangiocytes, and Kupffer cells, can reflect the liver's pathophysiological state in NAFLD. Circulating EVs in NAFLD patients' blood have been shown to carry specific proteins that are related to lipid metabolism, oxidative stress, inflammation, and fibrosis, which are hallmarks of NAFLD progression [12] (Figure 3).
However, most of the studies dedicated to finding biomolecules with diagnostic value in EVs derived from patients with NAFLD are based on classical technology approaches of EVs isolation and purification by ultracentrifugation followed by omic analysis of biomolecules (eg. proteomics, RNAseq, etc.). These approximations are necessary to discover new possible biomolecule candidates, but they are too complex to be taken to the clinic.
New technologies such as EV-Array [13] or Exoview, allow a quick and easy way to study the surface proteins of the EVs [14]. EV-Array is based on the technology of protein microarray and provides the opportunity to detect and phenotype small EVs from unpurified starting material in a high-throughput manner. Exoview is an innovative approach for the characterization of EVs surface proteins. Exoview uses nanoparticle tracking analysis (NTA) to measure the size and concentration of EVs and combines this with a microarray-based assay to analyse EVs surface proteins. The microarray-based assay uses antibodies that are immobilized on a glass slide to capture specific proteins from the EVs. The captured proteins are then detected using fluorescently labelled secondary antibodies. The resulting protein profile of the EVs can provide valuable information about the underlying biology [14]. Recent advances in flow cytometry have led to the development of high-sensitive flow cytometry (HSFC), which allows for the detection of EVs with higher sensitivity and specificity than traditional flow cytometry methods [15]. HSFC uses smaller sample volumes and specialized instrumentation to minimize background noise and increase signal detection. Another advantage of HSFC is its ability to provide information on multiple surface proteins simultaneously [16]. HSFC can detect and quantify the expression of multiple proteins on individual EVs directly in plasma without preceding purification steps, providing a more comprehensive analysis of EV populations.
In this review, our goal was to develop a list of potential EVs surface protein biomarkers that could aid in the diagnosis and monitoring of NAFLD. We summarize the current state of knowledge regarding EV surface proteins as potential biomarkers for NAFLD. Thus, we conducted a comprehensive review of the literature. (I) First, we reviewed scientific bibliography that have studied EV proteins as biomarkers in NAFLD, as well as papers that have studied circulating proteins as biomarkers in NAFLD. (II) Next, to identify potential candidates, we utilized the EV database Vesiclepedia.org, to qualify each protein. (III) Finally, we consulted the Human Protein Atlas to search for the localization of each protein. We focused on membrane proteins. Finally, we categorized the list according to the biological function and/or technical applications in which these EVs proteins were involved. Thus, we have generated Table 1 with 69 protein candidates with the potential to be used as EVs surface biomarkers in NAFLD.
2. Novel Proposed EV Protein Biomarkers
In this category, we included EV surface proteins that have already been proposed as NAFLD biomarkers by different studies.
Vanin-1 is an amidohydrolase that hydrolyzes specifically one of the carboamide linkages in D-pantetheine thus recycling pantothenic acid (vitamin B5) and releasing cysteamine. In a mice study, Povero et. al [18] showed that Lipid-Induced toxicity stimulates hepatocytes to release angiogenic EVs that require Vanin-1 on surface for uptake by endothelial cells. Their data identify hepatocyte-derived EVs as critical signals that contribute to angiogenesis and liver damage in steatohepatitis. Motomura et. al [17], using mice and human in vitro approaches showed vanin-1 upregulation and lipid accumulation in hepatocytes in response to high-fat diet and free fatty acids (FFA).
TREM2. Triggering receptor expressed on myeloid cells 2 (TREM2) is a membrane protein that forms a receptor signaling complex with the TYRO protein tyrosine kinase binding protein. Soluble TREM2 levels in blood could serve as a circulating marker of NAFLD [19]. Interestingly, TREM2 has been found on EVs derived from human neurons [21]. Additionally, TREM2 regulates EVs secretion in liver macrophages during NAFLD progression [20].
ADAMTSL2. ADAMTS like 2 (ADAMTSL2) is “a disintegrin and metalloproteinase with thrombospondin motifs”. It has been described as a soluble biomarker to indicate NAFLD to NASH progression in patients [22].
IL13RA1. Interleukin 13 receptor subunit alpha 1 (IL13RA1) is a subunit of the interleukin 13 receptor. Povero et. al [23] showed upregulated levels of IL13RA1 in cirrhotic NASH circulating EVs versus healthy controls. In the same study [23], authors found increased EV levels of IL27RA, ICAM2 and STK16 in cirrotic NASH samples.
3. Metabolism Related Proteins
Metabolism related proteins are critical for the regulation of lipid metabolism, glucose homeostasis, insulin signaling, and inflammatory responses, all of which are disrupted in NAFLD.
CD36 is a multifunctional glycoprotein that acts as receptor/transporter for a broad range of ligands. CD36 increases FFA uptake and, in the liver, it drives hepatosteatosis onset. Clinical studies have reinforced the significance of CD36 by showing increased content in the liver of NAFLD patients [25]. Interestingly, circulating levels of a soluble form of CD36 (sCD36) are abnormally elevated in NAFLD patients [25]. Moreover, we showed that CD36 is expressed on circulating EVs surface and it is related with the delivery of FFA from blood flow to the heart [24]. Bariatric surgery resulted in significantly altered levels of CD36 in circulating EVs of monocyte and endothelial origin [106].
TM4SF5. Transmembrane 4 L six family member 5 (TM4SF5) is a tetraspanin involved in nonalcoholic steatosis and further aggravation of liver disease [28]. TM4SF5 is present on the membranes of different organelles or EVs [26] and cooperates with transporters for fatty acids, amino acids, and monocarbohydrates, thus regulating nutrient uptake into hepatocytes. TM4SF5 can remodel the immune environment by interacting with immune cells during TM4SF5-mediated chronic liver diseases [27]. Interestingly, liver derived EVs with TM4SF5 on their surface target brown adipose tissue for homeostatic glucose clearance [26].
TM6SF2. Transmembrane 6 superfamily member 2 (TM6SF2) encodes for a protein of undetermined function. Genetic studies have reported the association between TM6SF2 variants with hepatic triglyceride content and its impact on NAFLD [29,30].
SLC27A5. Bile acyl–coenzyme A synthetase (Solute carrier family 27 member 5, also known as FATP5) is an isozyme of very long-chain acyl-CoA synthetase. It is considered as a hepatocyte-specific markers in circulating EVs [23]. It has been described a functional association between SLC27A5 and TM4SF4 in fatty acids overload hepatocytes [28]. Expression of SLC27A5 is up-regulated in fat-laden hepatocytes and down-regulated during the progression from NASH to cirrhosis, likely as a result of fat loss occurring during the late stage of the disease. In patients, median levels of SLC27A5-positive circulating EVs were 3-4-fold greater in subjects with NASH compared with healthy controls [23].
SGMS1. Sphingomyelin synthase 1 (SGMS1) catalyzes the reversible transfer of phosphocholine moiety in sphingomyelin biosynthesis. Glucosylceramide (GluCer) accelerates liver steatosis, steatohepatitis, and tumorigenesis [31]. Glucosylceramide stimulates transforming growth factor beta 1 (TGFβ1) activation, which mediates liver fibrosis. Human NASH patients were shown to have higher liver GluCer synthase and higher plasma GluCer levels [31].
GLUT1. Solute carrier family 2 member 1 (SLC2A1) is a facilitative glucose transporter, which is responsible for constitutive or basal glucose uptake. In NASH patient, increased liver GLUT1 levels correlates with higher degree of steatosis [32]. Lu Wan et. al, showed that EVs derived from hepatic stellate cells contain GLUT1 and it is related with metabolic switch of liver nonparenchymal cells [33].
GLUT2. Solute carrier family 2 member 2 (SLC2A2) facilitative hexose transporter that mediates the transport of glucose, fructose and galactose. In NAFLD impaired glucose uptake by the liver is mediated by GLUT2 interference [34].
GLUT5. Solute carrier family 2 member 5 (SLC2A5) functions as a fructose transporter that has only low activity with other monosaccharides. Increased liver levels of GLUT5 were observed in NAFLD induced by high fructose intake in rats [35].
GLUT4. Solute carrier family 2 member 4 (SLC2A4) is an insulin-regulated facilitative glucose transporter, which plays a key role in removal of glucose from circulation. Altered liver levels of GLUT4 has been found in patients with chronic liver disease [37]. We previously demonstrated that EVs derived from cardiomyocytes have GLUT4 on their surface with functional consequences for the cardio-endothelial communication axis [36].
AGTR1. Angiotensin II receptor type 1 (AGTR1) is the receptor for angiotensin II, a vasoconstricting peptide. Hepatic mRNA expression of Agtr1a were downregulated in NAFLD induced rats [40]. Moreover, gene variants of AGTR1 have been related with predisposition to develop NAFLD in patients [38,39]. Bansal et. at, showed AGTR1 presence in circulating EVs from patients under SARS-CoV-2 infection [107].
CAV1. Caveolin 1 (CAV1) is the main component of the caveolae plasma membranes found in most cell types. CAV1 is a crucial regulator of lipid accumulation and metabolism. It is known that hepatocytes CAV1 modulates metabolic gene profiles and function in NAFLD [41]. Increased levels of circulating EVs expressing CAV1 has been reported in melanoma patients [42].
4. Inflammation and Fibrosis
Inflammation and fibrosis are integral processes in the development and progression of NAFLD [108]. Inflammation in the liver is often caused by hepatocyte fat accumulation, which leads to the release of pro-inflammatory cytokines. These cytokines can damage liver cells and trigger an inflammatory response, which in turn can lead to the development of fibrosis [109]. The persistent inflammatory response contributes to hepatocyte injury, insulin resistance, and the recruitment of immune cells, amplifying liver damage. Simultaneously, fibrosis disrupts the liver architecture, impairing liver function, ultimately leading to cirrhosis and hepatocellular carcinoma (HCC) [109].
CXCL10. C-X-C motif chemokine ligand 10 (CXCL10) is a pro-inflammatory cytokine that is involved in a wide variety of processes such as chemotaxis, differentiation, and activation of peripheral immune cells, regulation of cell growth, apoptosis and modulation of angiostatic effects. During hepatocyte lipotoxicity, activated mixed lineage kinase 3 (MLK3) induces the release of CXCL10-bearing vesicles from hepatocytes, which are chemotactic for macrophages [43]. In diabetes, pancreatic beta cells in a pro-inflammatory environment release EVs with CXCL10 on the surface, which induce failure of neighbouring beta cells through activation of the CXCL10/CXCR3 axis [44].
TGFB1. Transforming growth factor beta 1 (TGFB1) is a secreted ligand of the TGF-beta (transforming growth factor-beta) superfamily of proteins. TGFB1 promotes hepatic stellate cells (HSC) activation and extracellular matrix production (ECM), which further contributes to the progression of NAFLD [45]. Interestingly, it has been shown that TGFB1 can be delivered on the surface of mast cell-derived EVs [46].
TGFB2, TGFB3 and their receptors (TGFBR1, TGFBR2, TGFBR3). Transforming growth factor beta 2 and 3 (TGFB1 and TGFB3) are secreted ligands of the TGF-beta (transforming growth factor-beta) superfamily of proteins. Different studies have investigated the implications of these proteins in NAFLD and its progression to NASH in relation to inflammation and fibrosis [48,49], why it is well established that regulation of the TGFB pathway is related to liver pathology. Moreover, Rodrigues-Junior et. al, wrote/published a complete review about TGFB components on EVs surface [47].
ITGB1. Integrin subunit beta 1 (ITGB1). Beta-1 integrins recognize the sequence R-G-D in a wide array of ligands. NASH patient’s livers have increased mRNA expression levels of ITGB1 compared with NAFLD and healthy control [50]. In mice, ITGB1 is released in EVs from hepatocytes under lipotoxic stress and mediates monocyte adhesion to liver sinusoidal endothelial cells to promote hepatic inflammation [51].
TLR2. Toll like receptor 2 (TLR2). Toll-like receptor (TLR) family plays a fundamental role in pathogen recognition and activation of innate immunity. TLR2 and palmitic acid cooperatively activate the inflammasome in Kupffer cells/macrophages in the development of NASH [52].
TLR4. Toll like receptor 4 (TLR4) expression is upregulated in NASH patients, compared with those with NAFLD [53]. Moreover, activation of the TLR4 inflammatory pathway contributes to NAFLD severity and NASH/hepatic fibrosis [54]. Zhang et al. showed evidence indicating TLR4 transference by EVs between dendritic cells [55].
P2RX7. Purinergic receptor P2 × 7 (P2RX7) is a receptor for ATP that acts as a ligand-gated ion channel that is responsible for ATP-dependent lysis of macrophages through the formation of membrane pores permeable to large molecules. P2RX7 regulation is related to the pathophysiological events leading to NAFLD and its inflammatory and fibrotic evolution [56].
P2RY14. Purinergic receptor P2Y14 (P2RY14) is a receptor for UDP-glucose and other UDP-sugar coupled to G-proteins. P2RY14 links hepatocyte death to hepatic stellate cell activation and fibrogenesis in the liver [57].
CXCR4. C-X-C motif chemokine receptor 4 (CXCR4) is a G-protein coupled receptor that transduces a signal by increasing intracellular calcium ion levels and enhancing MAPK1/MAPK3 activation. CXCR4 and its ligand are functionally and mechanistically involved in the progression of liver fibrosis [58,60]. Horizontal transference of CXCR4 by EVs promote hepatocarcinoma cell migration, invasion, and lymphangiogenesis [59].
PDGFRB and PDGFRA. Platelet derived growth factor receptors beta and alpha (PDGFRB and PDGFRA) are tyrosine-protein kinases that act as cell-surface receptors. In NAFLD patients, circulating levels of PDGFRB are progressively enhanced with increasing fibrosis stage and the largest difference was observed in patients with significant fibrosis, compared with no or mild fibrosis [61]. Moreover, increased expression of liver levels of PDGFRA has been associated with NAFLD to NASH progression in patients [31].
SPHK1. Sphingosine kinase 1 (SPHK1) catalyzes the phosphorylation of sphingosine to form sphingosine 1-phosphate (SPP), a lipid mediator with both intra- and extracellular functions. SPHK1 mediates hepatic inflammation in HFD mice and initiates proinflammatory signalling in hepatocytes [62]. SPHK1 has been found in EVs derived from the hepatocellular carcinoma cell line SK-Hep1 [63].
ITGA1. Integrin subunit alpha 1 (ITGA1) is a receptor for laminin and collagen. Hepatic insulin resistance associates with increased levels of liver collagen and elevated expression of ITGA1 in hepatocytes isolated from high fat (HF)-fed mice has been shown [64]. Moreover, ITGA1 facilitates hepatic insulin action while promoting lipid accumulation in mice under a HF diet.
ACKR1. Atypical chemokine receptor 1 (ACKR1) controls chemokine levels and localization via high-affinity chemokine binding that is uncoupled from classic ligand-driven signal transduction cascades. A transcriptome analysis of more than 100,000 single human cells revealed distinct endothelial subpopulations that inhabit the liver fibrotic niche. These endothelial cells express ACKR1+, restricted to cirrhotic liver tissue and induce the transmigration of leucocytes [67]. ACKR1 knockdown attenuated leucocyte recruitment by cirrhotic endothelial cells [66]. Importantly, ACKR1 has been found in EVs derived from endothelial cells [65].
GPNMB. Glycoprotein nmb (GPNMB) is a type I transmembrane glycoprotein. GPNMB overexpression ameliorated liver fat accumulation and fibrosis in diet-induced obesity in mice [68]. In patients with non-alcoholic steatohepatitis, serum soluble GPNMB concentrations were higher compared with patients with simple steatosis [68].
SCARB1. Scavenger receptor class B member 1 (SCARB1) is a receptor for different ligands such as phospholipids, cholesterol ester, lipoproteins, phosphatidylserine, and apoptotic cells. In diet-induced obesity in mice, SCARB1 deficiency increased inflammatory dyslipidaemia and adipocytes hypertrophy, and attenuated hepatic steatosis [69]. Angeloni et. al, demonstrate direct evidence of SCARB1 in EVs derived from prostate cancer cells [69].
FGFR1. Fibroblast growth factor receptor 1 (FGFR1) is a tyrosine-protein kinase that acts as cell-surface receptor for fibroblast growth factors. FGFR1 is a central player in the response to liver injury and fibrosis [70].
TNFSF10. TNF superfamily member 10 (TNFSF10) is a cytokine that belongs to the tumor necrosis factor (TNF) ligand family. Liver expression of TNFSF10 is increased in both human and experimental NASH, and fatty murine livers are sensitized to TNFS10F mediated hepatocyte apoptosis [71].
5. NOTCH Pathway
The Notch pathway is a conserved ligand-receptor signaling mechanism involved in the regulation of tissue homeostasis, the maintenance of stem cells in adults and in normal vasculature development and angiogenesis [76]. Notch signaling is activated when specific transmembrane Notch ligands of the Jagged or Delta-like type, found on neighboring cells, engage with the extracellular domain of the receptor by close proximity. Four Notch receptors have been identified in mammalian cells, Notch1-4, which are activated by five canonical ligands, Delta-like 1, 3, and 4 (Dll1, 3, and 4) and Jagged1 and Jagged2 [77,78]. Several studies have shown Notch signaling pathway plays a pivotal role in the regulation of NAFLD progression, from lipid accumulation to fibrosis and cancer [79]. It is well established that all NOTCH components (ligands and receptor) are functionally found in EVs from different sources [74,75].
6. Wnt/β-Catenin Pathway
The Wnt/β-catenin pathway plays a vital role in embryonic development, tissue homeostasis, and cell fate determination. Dysregulation of this pathway has been implicated in numerous human diseases, including cancer and metabolic disorders [110]. There exist 19 secreted Wnt ligands couple to 15 receptors and co-receptors. The canonical Wnt/β-catenin pathway is initiated by the binding of Wnt ligands to the Frizzled (FZD) receptors and co-receptors, such as LRP5/6. This interaction triggers a cascade of intracellular signaling events, leading to the stabilization and accumulation of β-catenin in the cytoplasm as well as posterior nucleus translocation to regulate gene expression [110]. In liver physiology, Wnt/β-catenin pathway plays a critical role to establish and maintain liver zonation [111,112].
Emerging evidence suggests that dysregulation of the Wnt/β-catenin pathway contributes to the development and progression of NAFLD [85]. In the context of NAFLD, aberrant activation of the pathway has been observed in hepatocytes, leading to the accumulation of lipids and the development of hepatic steatosis [85]. Additionally, increased β-catenin signaling has been associated with the activation of hepatic stellate cells and the subsequent progression to fibrosis in NASH. Finally, there are increased evidence showing that different cell types secrete Wnt/β-catenin component in EVs [55,88,113]. Here we highlight the ligands, receptors, and co-receptor which have been directly linked to NAFLD pathology.
Using a mouse model of methionine-choline-deficient diet (MCDD)-induced NASH, Zhu et. al, investigated the Wnt signalling pathways in relation to hepatic glucose oxidation [80]. While liver expression of Wnt1 was unchanged, Wnt3a was significantly reduced in NASH. In contrast, expression of Wnt5a and Wnt11 were increased 3 fold and 20 fold, respectively. Another mice diet-induced obese model of NASH identified increased expression of Wnt2 and RSPO3 (a Wnt pathway co-receptor) in liver pericentral endothelial cells [81]. Xiong et. al, performed single-cell RNA sequencing on non-parenchymal cells isolated from healthy and NASH mouse livers. In that study, secretome gene analysis revealed a highly connected network of intrahepatic signaling and disruption of vascular signaling in NASH. They found altered expression levels of Wnt9b, Wnt2, Wnt4 and Rspo3 [82]. Interestingly, Scavo et. al, found that in circulating EVs FZD7 levels were modulated by lifestyle interventions in patients with NAFLD [83]. In another study, Saponara et. al. showed that in mice, loss of hepatic Wnt/β-catenin activity by Lgr4/5 deletion led to loss of bile acid secretion, cholestatic features, altered lipid homeostasis, and deregulation of lipoprotein pathways, promoting NAFLD [84]. Mutations in LRP6 are one of the major causes of NAFLD induction. Moreover, deprived levels of Wnt1 and higher levels of DKK-1, an inhibitor of the Wnt pathway in plasma, are correlated with a raised risk of hyperlipidemia by antagonizing LRP6 [85]. Finally, different studies have shown β-catenin transference between cells mediated by EVs, which implies an alternative new level of regulation for the pathway, since non ligand-receptor interaction is required for the activation [86,87,88].
7. Plasma/Serum Secreted Proteins in NAFLD
Recent studies have employed proteomic techniques to find plasma and serum secreted proteins that are differentially expressed in patients with NAFLD compared with healthy individuals with the aim to identify new circulating biomarkers. These proteins include markers of liver damage, inflammation, oxidative stress, and metabolic dysfunction.
In a study with NAFLD and NASH patients, Niu et.al. identified deregulated plasma levels of AFM and PIGR [89]. AFM. Afamin (AFM) functions as carrier for hydrophobic molecules in body fluids. It is essential for the solubility and activity of Wnt ligands. Differents groups have identified circulating levels of AFM in patients as predictive biomarker for NAFLD development [89,90]. PIGR. Polymeric immunoglobulin receptor (PIGR) mediates selective transcytosis of polymeric IgA and IgM across mucosal epithelial cells. A corroboration of Niu et. al. results [89] was obtained by Veyel et. al. using a mice NASH model where they found increased circulating levels of PIGR associated with NASH phenotype [93]. PIGR was directly detected in EVs related with cancer biology [91,92].
Using a proteomic approach in plasma and serum samples from NAFLD patients, Corey et. al, identified circulating levels of formimidoyltransferase cyclodeaminase (FTCD) as an indicative biomarker for the NAFLD to NASH progression [22]. FTCD is a folate-dependent enzyme, that displays both transferase and deaminase activity.
8. Inter-Tissue Crosstalk
Inter-tissue crosstalk communication plays a fundamental role in the pathogenesis of NAFLD [38]. The intricate interactions between the liver, adipose tissue, skeletal muscle, and the gut contribute to the development and progression of hepatic steatosis, inflammation, and fibrosis. Excessive adiposity, particularly visceral adipose tissue, is strongly associated with NAFLD development and progression. Adipose tissue secretes various adipokines, EVs, cytokines, and FFAs directly or indirectly influence hepatic lipid metabolism, insulin sensitivity, inflammation, and fibrosis [114]. Skeletal muscle also plays a crucial role in NAFLD. Insulin resistance, a hallmark of NAFLD, is strongly linked to skeletal muscle dysfunction. Impaired glucose and lipid uptake by skeletal muscle led to increased delivery of substrates to the liver, thereby exacerbating hepatic lipid accumulation. Furthermore, skeletal muscle-derived myokines, such as interleukin-6 (IL-6), irisin, and myostatin, have emerged as important regulators of hepatic lipid metabolism and inflammation [115]. In this section, we list proteins related to inter-tissue crosstalk that has been linked to liver pathology.
CD40 is a member of the TNF-receptor superfamily. Its suppression is related to adipose tissue inflammation, while absence of CD40 resulted in aggravated metabolic dysfunction in mice. CD40 expressing CD11c+ dendritic cells contribute to liver inflammation during NASH but are protective against the metabolic syndrome via induction of regulatory T cells [94]. Interestingly, it has been shown that CD40 function is related to adipokines secretion [95].
LRG1. Leucine rich alpha-2-glycoprotein 1 (LRG1) belongs to the leucine-reach repeat proteins family. LRG1 is an adipokine that mediates obesity-induced hepatosteatosis and insulin resistance [96]. While LRG1 is a not a membrane protein, it has been found on circulating EVs surface from cancer patients [97].
FNDC5. Fibronectin type III domain containing 5 (FNDC5), also called irisin, is a secreted myokin released from muscle cells during exercise. Several studies have emphasized that obesity is closely related to a disorder of serum irisin. Clinical data have indicated that serum irisin levels are reduced in patients with obesity-related NAFLD [99]. Exercise induces increased amounts of irisin in circulating EVs in mice and humans [98].
AKR1B7. Aldo-keto reductase family 1 member B7 (AKR1B7) catalyzes the NADPH-dependent reduction of a wide variety of carbonyl-containing compounds to their corresponding alcohols. AKR1B7 in EVs derived from metabolically stressed adipocytes induce NASH in mice [100].
FASN. Fatty acid synthetase (FASN) is a multifunctional enzyme that catalyzes the de novo biosynthesis of long-chain saturated fatty acids starting from acetyl-CoA and malonyl-CoA in the presence of NADPH. FASN drives de novo lipogenesis and mediates pro-inflammatory and fibrogenic signalling in NAFLD [101]. FASN was selectively enriched in EVs derived from adipocytes under the hypoxic condition, which may increase lipid accumulation in recipient adipocytes and preadipocytes [102].
FABP4. Fatty acid binding protein (FABP4) is an adipose tissue-secreted adipokine implicated in the regulation of the energetic metabolism and inflammation [104]. High levels of circulating FABP4 have been described in people with obesity, atherogenic dyslipidemia, diabetes, the metabolic syndrome, and NAFLD. Additionally, emerging data from preclinical studies propose FABP4 as a causal actor involved in the disease progression, rather than a mere biomarker for the disease [104]. Preliminary evidence from flow cytometric analyses showed that circulating EVs contain the adipocyte markers FABP4 and adiponectin in human and mouse plasma. This implies that FABP4 was detected on circulating EVs surface [103,105].
9. Perspective
EVs have emerged as potential biomarkers for NAFLD due to their involvement in the development and progression of these conditions. EVs have been implicated in several processes associated with NAFLD, such as lipid metabolism, inflammation, and fibrosis. The unique composition of EV surface proteins makes them attractive candidates in biomarker discovery. One of the key advantages of EVs as biomarkers is their ability to circulate in the bloodstream, providing a minimally invasive way to access disease-related information from the liver. However, the detection and analysis of EVs in blood samples pose significant challenges, as they are heterogeneous and present at low concentrations. Therefore, the identification of specific surface proteins on EVs that are associated with NAFLD could enhance their diagnostic potential. In this regard, circulating EVs have a cell-specific origin which can be reflected on their expression of EV surface proteins. This feature, implies the possibility to study specific circulating EVs subpopulations in order to bring more specific results. For example, we demonstrated that bariatric surgery resulted in significantly altered levels of CD36 in circulating EVs derived from monocyte and endothelial cells [106].
Traditional methods for EV protein analysis, such as Western Blot, mass spectrometry, SomaScan platform, or ELISA, have limited throughput and specificity. However, emerging technologies like EV-Array, Exoview or high-sensitive flow cytometry enable high-throughput analysis and multiplex detection of EVs, allowing simultaneous evaluation of multiple surface proteins. These technologies could boost the translation of EV-based biomarkers into clinical practice. Moreover, the development of novel technologies, such as the use of microfluidics or nanomaterials, could further improve the sensitivity and specificity of EV detection.
The identification and analysis of surface proteins on EVs offer a promising avenue for the development of non-invasive biomarkers for NAFLD (Figure 4). These biomarkers could potentially overcome the limitations of current diagnostic approaches and provide clinicians with a rapid, cost-effective, and reliable tool to assess disease progression and response to therapy. However, further validation studies are needed to confirm the diagnostic and prognostic value of EV surface proteins in larger patient cohorts.
Author Contributions
Conceptualization, N.A.G. and A.H.; writing—review and editing, N.A.G., A.H., M.M., H.G.-K. and C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the ResearchCube program 2022 for “NGDx – Next Generation Diagnostics” from Aalborg University Hospital.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Loomba, R., S.L. Friedman, and G.I. Shulman, Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell, 2021. 184(10): p. 2537-2564. [CrossRef]
- Masoodi, M., et al., Metabolomics and lipidomics in NAFLD: biomarkers and non-invasive diagnostic tests. Nat Rev Gastroenterol Hepatol, 2021. 18(12): p. 835-856. [CrossRef]
- Kechagias, S., et al., Non-invasive diagnosis and staging of non-alcoholic fatty liver disease. Hormones (Athens), 2022. 21(3): p. 349-368. [CrossRef]
- Anstee, Q.M., L. Castera, and R. Loomba, Impact of non-invasive biomarkers on hepatology practice: Past, present and future. J Hepatol, 2022. 76(6): p. 1362-1378. [CrossRef]
- Hernandez Roman, J. and M.S. Siddiqui, The role of noninvasive biomarkers in diagnosis and risk stratification in nonalcoholic fatty liver disease. Endocrinol Diabetes Metab, 2020. 3(4): p. e00127. [CrossRef]
- Byrne, C.D. and G. Targher, NAFLD: a multisystem disease. J Hepatol, 2015. 62(1 Suppl): p. S47-64. [CrossRef]
- Park, H. , et al., Reappraisal of fibrosis-4 index and non-alcoholic fatty liver disease fibrosis score for advanced fibrosis in average-risk population. Front Med (Lausanne), 2022. 9: p. 1024836. [CrossRef]
- Sun, W. , et al., Comparison of FIB-4 index, NAFLD fibrosis score and BARD score for prediction of advanced fibrosis in adult patients with non-alcoholic fatty liver disease: A meta-analysis study. Hepatol Res, 2016. 46(9): p. 862-70. [CrossRef]
- van Niel, G. , et al., Challenges and directions in studying cell-cell communication by extracellular vesicles. Nat Rev Mol Cell Biol, 2022. 23(5): p. 369-382. [CrossRef]
- Wu, D., H. Zhu, and H. Wang, Extracellular Vesicles in Non-alcoholic Fatty Liver Disease and Alcoholic Liver Disease. Front Physiol, 2021. 12: p. 707429. [CrossRef]
- Newman, L.A., K. Muller, and A. Rowland, Circulating cell-specific extracellular vesicles as biomarkers for the diagnosis and monitoring of chronic liver diseases. Cell Mol Life Sci, 2022. 79(5): p. 232. [CrossRef]
- Garcia-Martinez, I. , et al., Insights Into Extracellular Vesicles as Biomarker of NAFLD Pathogenesis. Front Med (Lausanne), 2020. 7: p. 395. [CrossRef]
- Baek, R. and M.M. Jorgensen, Multiplexed Phenotyping of Small Extracellular Vesicles Using Protein Microarray (EV Array). Methods Mol Biol, 2017. 1545: p. 117-127. [CrossRef]
- Breitwieser, K. , et al., Detailed Characterization of Small Extracellular Vesicles from Different Cell Types Based on Tetraspanin Composition by ExoView R100 Platform. Int J Mol Sci, 2022. 23(15). [CrossRef]
- Botha, J., H. R. Pugsley, and A. Handberg, Conventional, High-Resolution and Imaging Flow Cytometry: Benchmarking Performance in Characterisation of Extracellular Vesicles. Biomedicines, 2021. 9(2). [CrossRef]
- Nielsen, M.H. , et al., Acute Exercise Increases Plasma Levels of Muscle-Derived Microvesicles Carrying Fatty Acid Transport Proteins. J Clin Endocrinol Metab, 2019. 104(10): p. 4804-4814. [CrossRef]
- Motomura, W. , et al., Analysis of vanin-1 upregulation and lipid accumulation in hepatocytes in response to a high-fat diet and free fatty acids. J Clin Biochem Nutr, 2012. 51(3): p. 163-9. [CrossRef]
- Povero, D. , et al., Lipid-induced toxicity stimulates hepatocytes to release angiogenic microparticles that require Vanin-1 for uptake by endothelial cells. Sci Signal, 2013. 6(296): p. ra88. [CrossRef]
- Hendrikx, T. , et al., Soluble TREM2 levels reflect the recruitment and expansion of TREM2(+) macrophages that localize to fibrotic areas and limit NASH. J Hepatol, 2022. 77(5): p. 1373-1385. [CrossRef]
- Hou, J. , et al., TREM2 sustains macrophage-hepatocyte metabolic coordination in nonalcoholic fatty liver disease and sepsis. J Clin Invest, 2021. 131(4). [CrossRef]
- Mallach, A. , et al., The influence of the R47H triggering receptor expressed on myeloid cells 2 variant on microglial exosome profiles. Brain Commun, 2021. 3(2): p. fcab009. [CrossRef]
- Corey, K.E. , et al., ADAMTSL2 protein and a soluble biomarker signature identify at-risk non-alcoholic steatohepatitis and fibrosis in adults with NAFLD. J Hepatol, 2022. 76(1): p. 25-33. [CrossRef]
- Povero, D. , et al., Characterization and Proteome of Circulating Extracellular Vesicles as Potential Biomarkers for NASH. Hepatol Commun, 2020. 4(9): p. 1263-1278. [CrossRef]
- Garcia, N.A. , et al., Circulating exosomes deliver free fatty acids from the bloodstream to cardiac cells: Possible role of CD36. PLoS One, 2019. 14(5): p. e0217546. [CrossRef]
- Rada, P. , et al., Understanding lipotoxicity in NAFLD pathogenesis: is CD36 a key driver? Cell Death Dis, 2020. 11(9): p. 802. [CrossRef]
- Jung, J.W. , et al., Liver-originated small extracellular vesicles with TM4SF5 target brown adipose tissue for homeostatic glucose clearance. J Extracell Vesicles, 2022. 11(9): p. e12262. [CrossRef]
- Kim, E. , et al., TM4SF5-dependent crosstalk between hepatocytes and macrophages to reprogram the inflammatory environment. Cell Rep, 2021. 37(7): p. 110018. [CrossRef]
- Park, D. , et al., Tetraspanin TM4SF5 in hepatocytes negatively modulates SLC27A transporters during acute fatty acid supply. Arch Biochem Biophys, 2021. 710: p. 109004. [CrossRef]
- Luo, F., F. Oldoni, and A. Das, TM6SF2: A Novel Genetic Player in Nonalcoholic Fatty Liver and Cardiovascular Disease. Hepatol Commun, 2022. 6(3): p. 448-460. [CrossRef]
- Mann, J.P. , et al., Insights into genetic variants associated with NASH-fibrosis from metabolite profiling. Hum Mol Genet, 2020. 29(20): p. 3451-3463. [CrossRef]
- Li, Z. , et al., Liver sphingomyelin synthase 1 deficiency causes steatosis, steatohepatitis, fibrosis, and tumorigenesis: An effect of glucosylceramide accumulation. iScience, 2021. 24(12): p. 103449. [CrossRef]
- Granja, S.C. , et al., Non-Alcoholic Fatty Liver Disease-Related Hepatocellular Carcinoma: Immunohistochemical Assessment of Markers of Cancer Cell Metabolism. Pathobiology, 2022. 89(3): p. 157-165. [CrossRef]
- Wan, L. , et al., Exosomes from activated hepatic stellate cells contain GLUT1 and PKM2: a role for exosomes in metabolic switch of liver nonparenchymal cells. FASEB J, 2019. 33(7): p. 8530-8542. [CrossRef]
- Shearer, A.M. , et al., PAR2 promotes impaired glucose uptake and insulin resistance in NAFLD through GLUT2 and Akt interference. Hepatology, 2022. 76(6): p. 1778-1793. [CrossRef]
- Roncal-Jimenez, C.A. , et al., Sucrose induces fatty liver and pancreatic inflammation in male breeder rats independent of excess energy intake. Metabolism, 2011. 60(9): p. 1259-70. [CrossRef]
- Garcia, N.A. , et al., Cardiomyocyte exosomes regulate glycolytic flux in endothelium by direct transfer of GLUT transporters and glycolytic enzymes. Cardiovasc Res, 2016. 109(3): p. 397-408. [CrossRef]
- Karim, S. , et al., Dysregulated hepatic expression of glucose transporters in chronic disease: contribution of semicarbazide-sensitive amine oxidase to hepatic glucose uptake. Am J Physiol Gastrointest Liver Physiol, 2014. 307(12): p. G1180-90. [CrossRef]
- de Gracia Hahn, D., A. Duret, and J.P. Mann, An AGTR1 Variant Worsens Nonalcoholic Fatty Liver Disease and the Metabolic Syndrome. Am J Gastroenterol, 2019. 114(4): p. 556-559. [CrossRef]
- Eshraghian, A., S. Iravani, and P. Azimzadeh, The Association between Angiotensin II Type 1 Receptor Gene A1166C Polymorphism and Non-alcoholic Fatty Liver Disease and Its Severity. Middle East J Dig Dis, 2018. 10(2): p. 96-104. [CrossRef]
- van der Graaff, D. , et al., Vasoconstrictor antagonism improves functional and structural vascular alterations and liver damage in rats with early NAFLD. JHEP Rep, 2022. 4(2): p. 100412. [CrossRef]
- Han, M. , et al., Hepatocyte caveolin-1 modulates metabolic gene profiles and functions in non-alcoholic fatty liver disease. Cell Death Dis, 2020. 11(2): p. 104. [CrossRef]
- Logozzi, M. , et al., High levels of exosomes expressing CD63 and caveolin-1 in plasma of melanoma patients. PLoS One, 2009. 4(4): p. e5219. [CrossRef]
- Ibrahim, S.H. , et al., Mixed lineage kinase 3 mediates release of C-X-C motif ligand 10-bearing chemotactic extracellular vesicles from lipotoxic hepatocytes. Hepatology, 2016. 63(3): p. 731-44. [CrossRef]
- Javeed, N. , et al., Pro-inflammatory beta cell small extracellular vesicles induce beta cell failure through activation of the CXCL10/CXCR3 axis in diabetes. Cell Rep, 2021. 36(8): p. 109613. [CrossRef]
- Nair, B. and L.R. Nath, Inevitable role of TGF-beta1 in progression of nonalcoholic fatty liver disease. J Recept Signal Transduct Res, 2020. 40(3): p. 195-200. [CrossRef]
- Shelke, G.V. , et al., Endosomal signalling via exosome surface TGFbeta-1. J Extracell Vesicles, 2019. 8(1): p. 1650458. [CrossRef]
- Rodrigues-Junior, D.M. , et al., Extracellular Vesicles and Transforming Growth Factor beta Signaling in Cancer. Front Cell Dev Biol, 2022. 10: p. 849938. [CrossRef]
- Wallace, S.J. , et al., Understanding the cellular interactome of non-alcoholic fatty liver disease. JHEP Rep, 2022. 4(8): p. 100524. [CrossRef]
- Yang, L. , et al., Transforming growth factor beta signaling in hepatocytes participates in steatohepatitis through regulation of cell death and lipid metabolism in mice. Hepatology, 2014. 59(2): p. 483-95. [CrossRef]
- Albadawy, R. , et al., Clinical Significance of HSPD1/MMP14/ITGB1/miR-6881-5P/Lnc-SPARCL1-1:2 RNA Panel in NAFLD/NASH Diagnosis: Egyptian Pilot Study. Biomedicines, 2021. 9(9). [CrossRef]
- Guo, Q. , et al., Integrin beta(1)-enriched extracellular vesicles mediate monocyte adhesion and promote liver inflammation in murine NASH. J Hepatol, 2019. 71(6): p. 1193-1205. [CrossRef]
- Miura, K. , et al., Toll-like receptor 2 and palmitic acid cooperatively contribute to the development of nonalcoholic steatohepatitis through inflammasome activation in mice. Hepatology, 2013. 57(2): p. 577-89. [CrossRef]
- Sharifnia, T. , et al., Hepatic TLR4 signaling in obese NAFLD. Am J Physiol Gastrointest Liver Physiol, 2015. 309(4): p. G270-8. [CrossRef]
- Sun, B.L. , et al., Involvement of eNAMPT/TLR4 inflammatory signaling in progression of non-alcoholic fatty liver disease, steatohepatitis, and fibrosis. FASEB J, 2023. 37(3): p. e22825. [CrossRef]
- Zhang, Y. , et al., Extracellular Vesicles with Exosome-like Features Transfer TLRs between Dendritic Cells. Immunohorizons, 2019. 3(6): p. 186-193. [CrossRef]
- Rossato, M. , et al., The P2 × 7 Receptor and NLRP3 Axis in Non-Alcoholic Fatty Liver Disease: A Brief Review. Cells, 2020. 9(4). [CrossRef]
- Mederacke, I. , et al., The purinergic P2Y14 receptor links hepatocyte death to hepatic stellate cell activation and fibrogenesis in the liver. Sci Transl Med, 2022. 14(639): p. eabe5795. [CrossRef]
- Boujedidi, H. , et al., CXCR4 dysfunction in non-alcoholic steatohepatitis in mice and patients. Clin Sci (Lond), 2015. 128(4): p. 257-67. [CrossRef]
- Li, M. , et al., Horizontal transfer of exosomal CXCR4 promotes murine hepatocarcinoma cell migration, invasion and lymphangiogenesis. Gene, 2018. 676: p. 101-109. [CrossRef]
- Wang, S. , et al., Emerging Importance of Chemokine Receptor CXCR4 and Its Ligand in Liver Disease. Front Cell Dev Biol, 2021. 9: p. 716842. [CrossRef]
- Lambrecht, J. , et al., A PDGFRbeta-based score predicts significant liver fibrosis in patients with chronic alcohol abuse, NAFLD and viral liver disease. EBioMedicine, 2019. 43: p. 501-512. [CrossRef]
- Geng, T. , et al., SphK1 mediates hepatic inflammation in a mouse model of NASH induced by high saturated fat feeding and initiates proinflammatory signaling in hepatocytes. J Lipid Res, 2015. 56(12): p. 2359-71. [CrossRef]
- Rigogliuso, S. , et al., An active form of sphingosine kinase-1 is released in the extracellular medium as component of membrane vesicles shed by two human tumor cell lines. J Oncol, 2010. 2010: p. 509329. [CrossRef]
- Williams, A.S. , et al., Integrin alpha1-null mice exhibit improved fatty liver when fed a high fat diet despite severe hepatic insulin resistance. J Biol Chem, 2015. 290(10): p. 6546-57. [CrossRef]
- Guo, X. , et al., Endothelial ACKR1 is induced by neutrophil contact and down-regulated by secretion in extracellular vesicles. Front Immunol, 2023. 14: p. 1181016. [CrossRef]
- Nasiri-Ansari, N. , et al., Endothelial Cell Dysfunction and Nonalcoholic Fatty Liver Disease (NAFLD): A Concise Review. Cells, 2022. 11(16). [CrossRef]
- Ramachandran, P. , et al., Resolving the fibrotic niche of human liver cirrhosis at single-cell level. Nature, 2019. 575(7783): p. 512-518. [CrossRef]
- Katayama, A. , et al., Beneficial impact of Gpnmb and its significance as a biomarker in nonalcoholic steatohepatitis. Sci Rep, 2015. 5: p. 16920. [CrossRef]
- Angeloni, N.L. , et al., Pathways for Modulating Exosome Lipids Identified By High-Density Lipoprotein-Like Nanoparticle Binding to Scavenger Receptor Type B-1. Sci Rep, 2016. 6: p. 22915. [CrossRef]
- Ding, B.S. , et al., Divergent angiocrine signals from vascular niche balance liver regeneration and fibrosis. Nature, 2014. 505(7481): p. 97-102. [CrossRef]
- Hirsova, P. , et al., TRAIL Deletion Prevents Liver, but Not Adipose Tissue, Inflammation during Murine Diet-Induced Obesity. Hepatol Commun, 2017. 1(7): p. 648-662. [CrossRef]
- Bacil, G.P. , et al., Unraveling Hepatic Metabolomic Profiles and Morphological Outcomes in a Hybrid Model of NASH in Different Mouse Strains. Antioxidants (Basel), 2023. 12(2). [CrossRef]
- Osada-Oka, M. , et al., Macrophage-derived exosomes induce inflammatory factors in endothelial cells under hypertensive conditions. Hypertens Res, 2017. 40(4): p. 353-360. [CrossRef]
- Gonzalez-King, H. , et al., Hypoxia Inducible Factor-1alpha Potentiates Jagged 1-Mediated Angiogenesis by Mesenchymal Stem Cell-Derived Exosomes. Stem Cells, 2017. 35(7): p. 1747-1759. [CrossRef]
- Gonzalez-King, H. , et al., Non-classical Notch signaling by MDA-MB-231 breast cancer cell-derived small extracellular vesicles promotes malignancy in poorly invasive MCF-7 cells. Cancer Gene Ther, 2022. 29(7): p. 1056-1069. [CrossRef]
- Gridley, T. , Human Genetics. Notch, stroke and dementia. Nature, 1996. 383(6602): p. 673. [CrossRef]
- Olsauskas-Kuprys, R., A. Zlobin, and C. Osipo, Gamma secretase inhibitors of Notch signaling. Onco Targets Ther, 2013. 6: p. 943-55. [CrossRef]
- Sassoli, C. , et al., Mesenchymal stromal cells affect cardiomyocyte growth through juxtacrine Notch-1/Jagged-1 signaling and paracrine mechanisms: clues for cardiac regeneration. J Mol Cell Cardiol, 2011. 51(3): p. 399-408. [CrossRef]
- Xu, H. and L. Wang, The Role of Notch Signaling Pathway in Non-Alcoholic Fatty Liver Disease. Front Mol Biosci, 2021. 8: p. 792667. [CrossRef]
- Zhu, L. , et al., Upregulation of non-canonical Wnt ligands and oxidative glucose metabolism in NASH induced by methionine-choline deficient diet. Trends Cell Mol Biol, 2018. 13: p. 47-56.
- HM, A.E. , et al., Multi-omics characterization of a diet-induced obese model of non-alcoholic steatohepatitis. Sci Rep, 2020. 10(1): p. 1148. [CrossRef]
- Xiong, X. , et al., Landscape of Intercellular Crosstalk in Healthy and NASH Liver Revealed by Single-Cell Secretome Gene Analysis. Mol Cell, 2019. 75(3): p. 644-660 e5. [CrossRef]
- Scavo, M.P. , et al., Exosomal FZD-7 Expression Is Modulated by Different Lifestyle Interventions in Patients with NAFLD. Nutrients, 2022. 14(6). [CrossRef]
- Saponara, E. , et al., Loss of Hepatic Leucine-Rich Repeat-Containing G-Protein Coupled Receptors 4 and 5 Promotes Nonalcoholic Fatty Liver Disease. Am J Pathol, 2023. 193(2): p. 161-181. [CrossRef]
- Shree Harini, K. and D. Ezhilarasan, Wnt/beta-catenin signaling and its modulators in nonalcoholic fatty liver diseases. Hepatobiliary Pancreat Dis Int, 2023. 22(4): p. 333-345. [CrossRef]
- Chairoungdua, A. , et al., Exosome release of beta-catenin: a novel mechanism that antagonizes Wnt signaling. J Cell Biol, 2010. 190(6): p. 1079-91. [CrossRef]
- Dovrat, S. , et al., 14-3-3 and beta-catenin are secreted on extracellular vesicles to activate the oncogenic Wnt pathway. Mol Oncol, 2014. 8(5): p. 894-911. [CrossRef]
- Kalra, H. , et al., Extracellular vesicles containing oncogenic mutant beta-catenin activate Wnt signalling pathway in the recipient cells. J Extracell Vesicles, 2019. 8(1): p. 1690217. [CrossRef]
- Niu, L. , et al., Plasma proteome profiling discovers novel proteins associated with non-alcoholic fatty liver disease. Mol Syst Biol, 2019. 15(3): p. e8793. [CrossRef]
- Pitkanen, N. , et al., Afamin predicts the prevalence and incidence of nonalcoholic fatty liver disease. Clin Chem Lab Med, 2022. 60(2): p. 243-251. [CrossRef]
- Liu, Y., Y. Hu, and L. Deng, The Underlying Roles of Exosome-Associated PIGR in Fatty Acid Metabolism and Immune Signaling in Colorectal Cancer. J Oncol, 2022. 2022: p. 4675683. [CrossRef]
- Tey, S.K. , et al., Patient pIgR-enriched extracellular vesicles drive cancer stemness, tumorigenesis and metastasis in hepatocellular carcinoma. J Hepatol, 2022. 76(4): p. 883-895. [CrossRef]
- Veyel, D. , et al., Biomarker discovery for chronic liver diseases by multi-omics - a preclinical case study. Sci Rep, 2020. 10(1): p. 1314. [CrossRef]
- Aarts, S. , et al., Depletion of CD40 on CD11c(+) cells worsens the metabolic syndrome and ameliorates hepatic inflammation during NASH. Sci Rep, 2019. 9(1): p. 14702. [CrossRef]
- Yuan, M. , et al., CD40L/CD40 Regulates Adipokines and Cytokines by H3K4me3 Modification in Epicardial Adipocytes. J Cardiovasc Pharmacol, 2021. 78(2): p. 228-234. [CrossRef]
- He, S. , et al., LRG1 is an adipokine that mediates obesity-induced hepatosteatosis and insulin resistance. J Clin Invest, 2021. 131(24). [CrossRef]
- Zhong, M.E. , et al., Serum extracellular vesicles contain SPARC and LRG1 as biomarkers of colon cancer and differ by tumour primary location. EBioMedicine, 2019. 50: p. 211-223. [CrossRef]
- Chi, C. , et al., Exerkine fibronectin type-III domain-containing protein 5/irisin-enriched extracellular vesicles delay vascular ageing by increasing SIRT6 stability. Eur Heart J, 2022. 43(43): p. 4579-4595. [CrossRef]
- Wang, Y.D. , et al., New insight of obesity-associated NAFLD: Dysregulated "crosstalk" between multi-organ and the liver? Genes Dis, 2023. 10(3): p. 799-812. [CrossRef]
- Gu, H. , et al., ER stress-induced adipocytes secrete-aldo-keto reductase 1B7-containing exosomes that cause nonalcoholic steatohepatitis in mice. Free Radic Biol Med, 2021. 163: p. 220-233. [CrossRef]
- O'Farrell, M. , et al., FASN inhibition targets multiple drivers of NASH by reducing steatosis, inflammation and fibrosis in preclinical models. Sci Rep, 2022. 12(1): p. 15661. [CrossRef]
- Sano, S. , et al., Lipid synthesis is promoted by hypoxic adipocyte-derived exosomes in 3T3-L1 cells. Biochem Biophys Res Commun, 2014. 445(2): p. 327-33. [CrossRef]
- Gustafson, C.M. , et al., Age- and sex-specific differences in blood-borne microvesicles from apparently healthy humans. Biol Sex Differ, 2015. 6: p. 10. [CrossRef]
- Moreno-Vedia, J. , et al., Unveiling the Role of the Fatty Acid Binding Protein 4 in the Metabolic-Associated Fatty Liver Disease. Biomedicines, 2022. 10(1). [CrossRef]
- Phoonsawat, W. , et al., Adiponectin is partially associated with exosomes in mouse serum. Biochem Biophys Res Commun, 2014. 448(3): p. 261-6. [CrossRef]
- Botha, J. , et al., Bariatric surgery reduces CD36-bearing microvesicles of endothelial and monocyte origin. Nutr Metab (Lond), 2018. 15: p. 76. [CrossRef]
- Bansal, S. , et al., SARS-CoV-2 infection in lung transplant recipients induces circulating exosomes with SARS-CoV-2 spike protein S2. Clin Transl Med, 2021. 11(11): p. e576. [CrossRef]
- Peiseler, M. , et al., Immune mechanisms linking metabolic injury to inflammation and fibrosis in fatty liver disease - novel insights into cellular communication circuits. J Hepatol, 2022. 77(4): p. 1136-1160. [CrossRef]
- Lee, Y.A. and S.L. Friedman, Inflammatory and fibrotic mechanisms in NAFLD-Implications for new treatment strategies. J Intern Med, 2022. 291(1): p. 11-31. [CrossRef]
- Liu, J. , et al., Wnt/beta-catenin signalling: function, biological mechanisms, and therapeutic opportunities. Signal Transduct Target Ther, 2022. 7(1): p. 3. [CrossRef]
- Goel, C., S. P. Monga, and K. Nejak-Bowen, Role and Regulation of Wnt/beta-Catenin in Hepatic Perivenous Zonation and Physiological Homeostasis. Am J Pathol, 2022. 192(1): p. 4-17. [CrossRef]
- Ma, R. , et al., Metabolic and non-metabolic liver zonation is established non-synchronously and requires sinusoidal Wnts. Elife, 2020. 9. [CrossRef]
- Gross, J.C. and L.C. Zelarayan, The Mingle-Mangle of Wnt Signaling and Extracellular Vesicles: Functional Implications for Heart Research. Front Cardiovasc Med, 2018. 5: p. 10. [CrossRef]
- Lee, E., H. Korf, and A. Vidal-Puig, An adipocentric perspective on the development and progression of non-alcoholic fatty liver disease. J Hepatol, 2023. 78(5): p. 1048-1062. [CrossRef]
- Chakravarthy, M.V. , et al., Harnessing Muscle-Liver Crosstalk to Treat Nonalcoholic Steatohepatitis. Front Endocrinol (Lausanne), 2020. 11: p. 592373. [CrossRef]
Figure 1.
Schematic representation of the grading and staging of NAFLD based on liver biopsies analysis. (a) Staging is measure by the fibrosis progression. (b) The grade of severity is classified based on steatosis, ballooning, and inflammation.
Figure 1.
Schematic representation of the grading and staging of NAFLD based on liver biopsies analysis. (a) Staging is measure by the fibrosis progression. (b) The grade of severity is classified based on steatosis, ballooning, and inflammation.
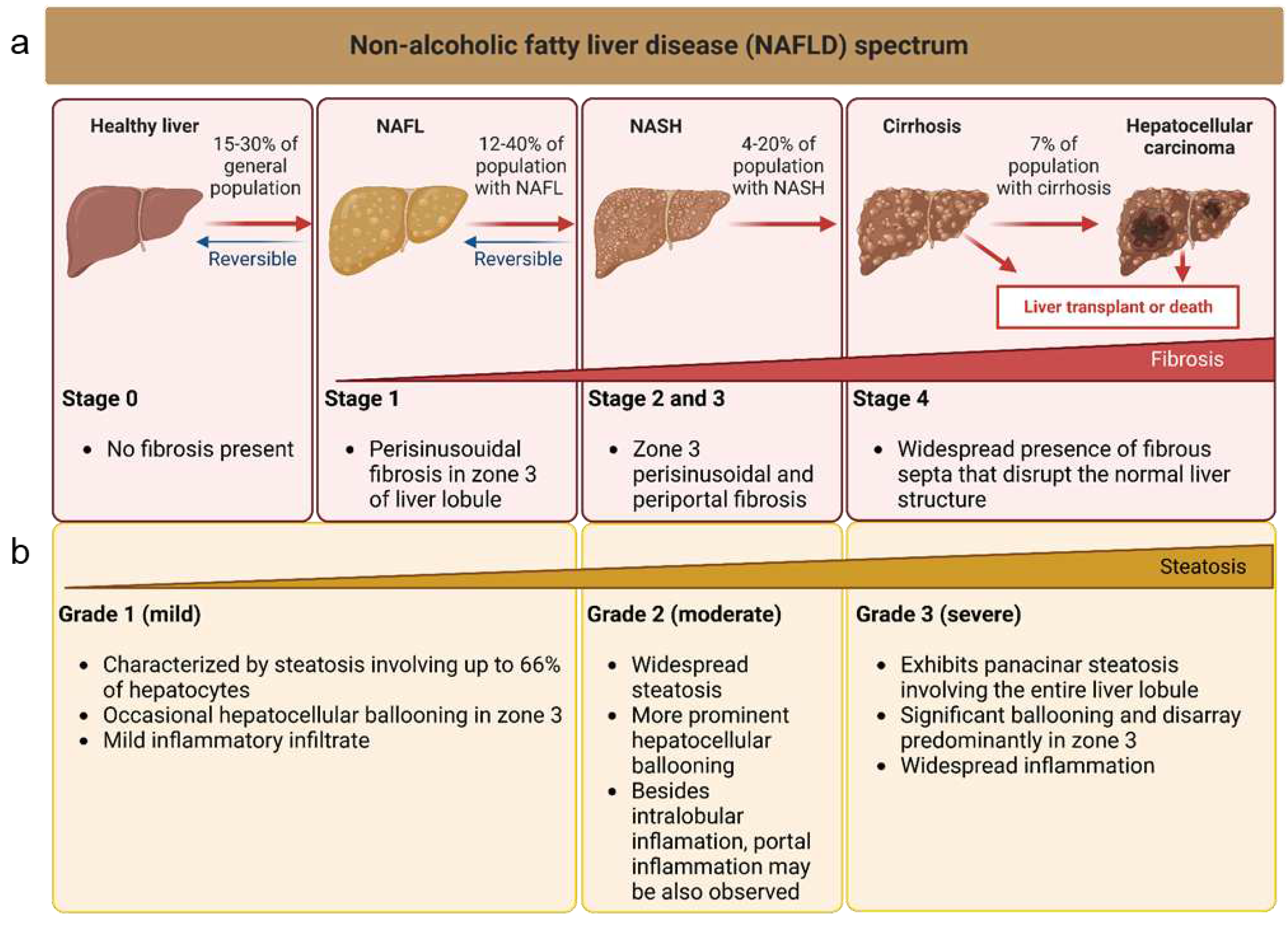
Figure 2.
Schematic representation of the different origins and action mechanisms for EVs. 1. EVs can be produced from the early endosomal pathway (EVs called exosomes around 30-130 nm of diameter) or by evaginations of the plasmatic membrane (EVs called microvesicles around 130-1000 nm of diameter). 2. EVs can be either engulfed by other cells through processes like pynocitosis or phagocytosis. After been engulfed, EVs now inside an endosome, can fuse with the membrane of the endosome and release their content (endosomal scape) degraded by fusion with lysosomes or placed again in the extracellular space (recycling). 3. EVs can also interact directly with receptors to trigger signalling cascades in the receiving cells, or they can directly fuse with the plasmatic membrane of the receiving cells releasing their cytosolic cargo (4.).
Figure 2.
Schematic representation of the different origins and action mechanisms for EVs. 1. EVs can be produced from the early endosomal pathway (EVs called exosomes around 30-130 nm of diameter) or by evaginations of the plasmatic membrane (EVs called microvesicles around 130-1000 nm of diameter). 2. EVs can be either engulfed by other cells through processes like pynocitosis or phagocytosis. After been engulfed, EVs now inside an endosome, can fuse with the membrane of the endosome and release their content (endosomal scape) degraded by fusion with lysosomes or placed again in the extracellular space (recycling). 3. EVs can also interact directly with receptors to trigger signalling cascades in the receiving cells, or they can directly fuse with the plasmatic membrane of the receiving cells releasing their cytosolic cargo (4.).
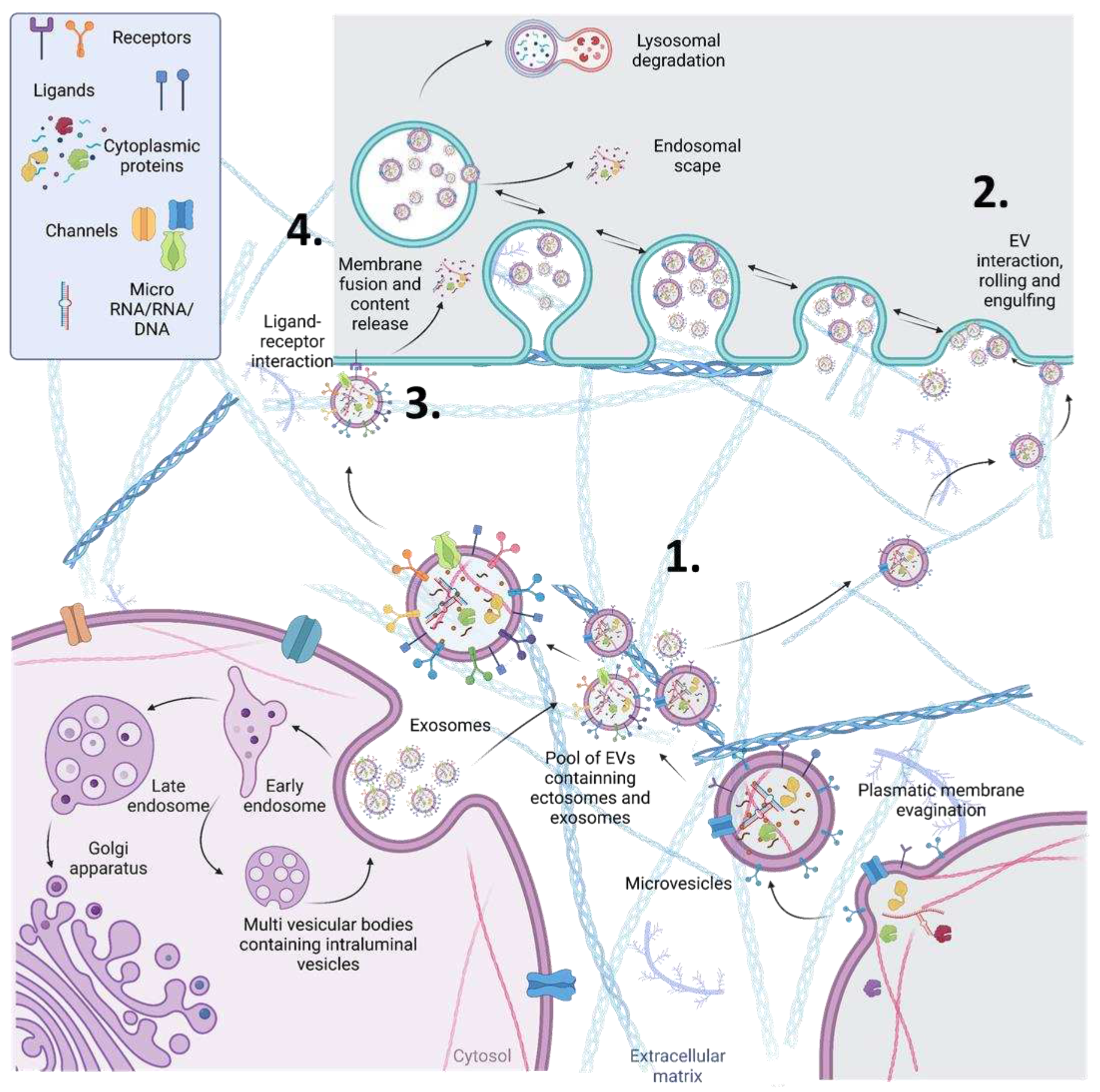
Figure 3.
Liver cells releasing EVs to the circulation. Across different stages of NAFLD pathology, the released EVs have different patterns of surface proteins in the membrane. This differential expression of EV surface proteins could be used as NAFLD biomarkers by measuring from patients’ blood samples.
Figure 3.
Liver cells releasing EVs to the circulation. Across different stages of NAFLD pathology, the released EVs have different patterns of surface proteins in the membrane. This differential expression of EV surface proteins could be used as NAFLD biomarkers by measuring from patients’ blood samples.

Figure 4.
Visualization of the current methods in the diagnosis of NAFLD.
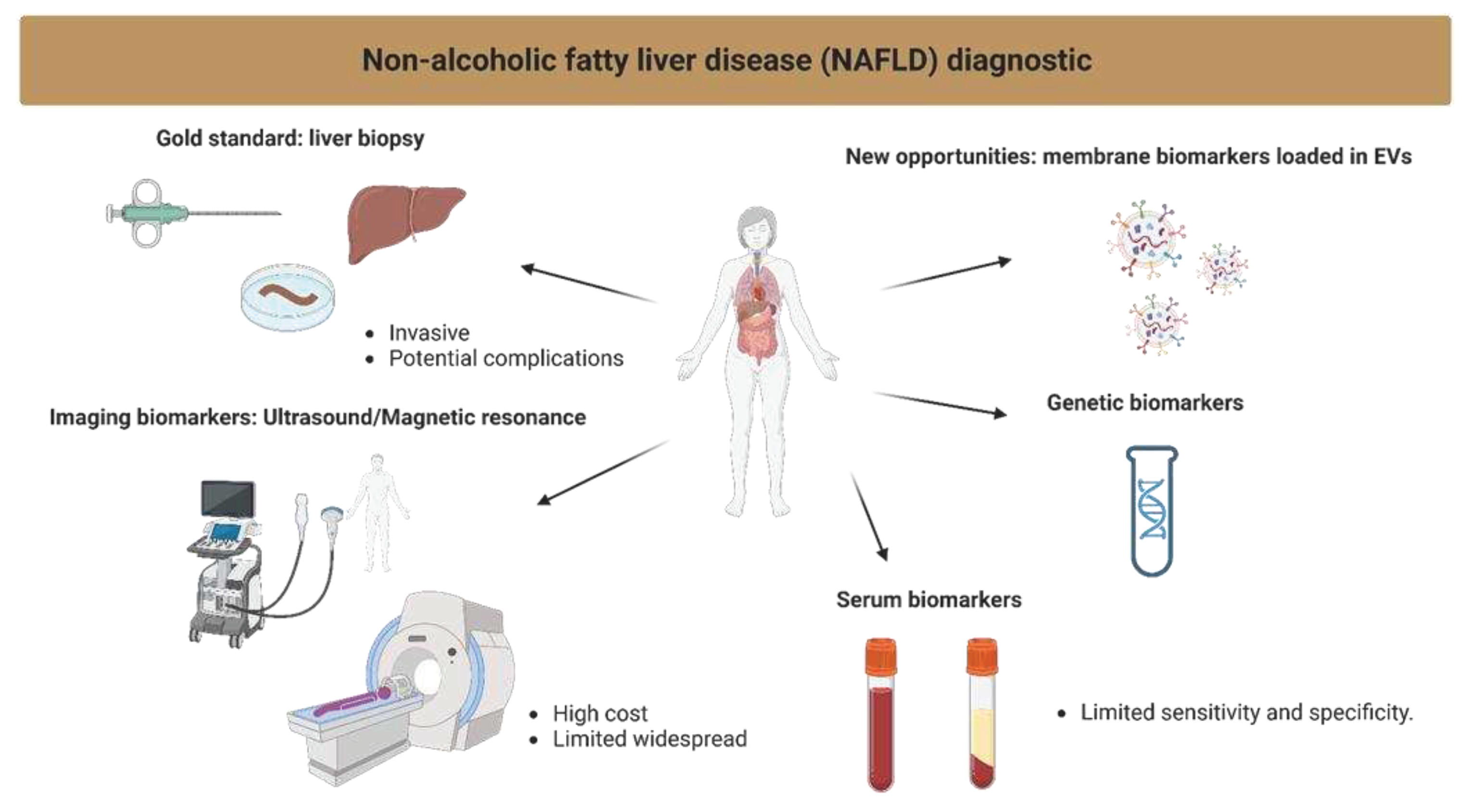

Table 1.
EVs Protein candidates related to NAFLD.
| Mechanism | Name | Description | References |
|---|---|---|---|
| Novel proposed biomarkers | VANIN-1 | Released on EVs surface from lipotoxic hepatocytes | [17,18] |
| TREM2 | Soluble TREM2 levels correlates with NAFLD to NASH progression | [19,20,21] | |
| ADAMTS2 | Soluble ADAMTS2 levels correlates with NAFLD to NASH progression | [22] | |
| IL13RA1 | upregulated levels in circulating EVs in NASH | [23] | |
| IL27RA | upregulated levels in circulating EVs in NASH | [23] | |
| ICAM2 | upregulated levels in circulating EVs in NASH | [23] | |
| STK16 | upregulated levels in circulating EVs in NASH | [23] | |
| Metabolism related proteins | CD36 | circulating levels of a soluble form of CD36 are abnormally elevated in NAFLD patients | [24,25] |
| TM4SF5 | liver derived EVs with TM4SF5 target brown adipose tissue for glucose clearance | [26,27,28] | |
| TM6SF2 | TM6SF2 variants were related to hepatic triglyceride in NAFLD and NASH | [29,30] | |
| SLC27A5 | upregulated levels in circulating EVs in NASH | [23,28] | |
| SGMS1 | NASH patients had higher liver GluCer synthase and higher plasma GluCer levels | [31] | |
| GLUT1 | increased liver GLUT1 levels correlates with higher degree of steatosis in NASH | [32,33] | |
| GLUT2 | Decreased liver levels in NAFLD | [34] | |
| GLUT5 | Increased liver levels in NAFLD induced by high fructose intake in rats | [35,36,37] | |
| GLUT4 | Altered liver levels in patients with chronic liver disease | [36,37] | |
| AGTR1 | gene variants of AGTR1 have been related with predisposition to develop NAFLD | [38,39,40] | |
| CAV1 | hepatocytes CAV1 modulates metabolic gene profiles and function in NAFLD | [41,42] | |
| Inflamation/fibrosis | CXCL10 | hepatocyte lipotoxicity induces the release of CXCL10-bearing vesicles | [43,44] |
| TGFB1 | promotes HSC activation and extracellular matrix production in NAFLD | [45,46] | |
| TGFB2 | drives multiple types of fibrosis during NAFLD to NASH progression | [47,48,49] | |
| TGFBR2 | drives multiple types of fibrosis during NAFLD to NASH progression | [47,48,49] | |
| TGFBR3 | drives multiple types of fibrosis during NAFLD to NASH progression | [47,48,49] | |
| TGFBR1 | drives multiple types of fibrosis during NAFLD to NASH progression | [47,48,49] | |
| ITGB1 | ITGB1 is released in EVs from hepatocytes under lipotoxic stress | [50,51] | |
| TLR2 | activate inflammasome in Kupffer cells/macrophages in NASH development | [52] | |
| TLR4 | contributes to NAFLD severity and NASH/hepatic fibrosis | [53,54,55] | |
| P2 × 7R | related to NAFLD and its inflammatory and fibrotic evolution | [56] | |
| P2Y14R | links hepatocyte death to hepatic stellate cell activation and fibrogenesis | [57] | |
| CXCR4 | functionally and mechanistically involved in the progression of liver fibrosis | [58,59,60] | |
| PDGFRA | increased liver expression levels in NASH patients | [31,61] | |
| PDGFRB | circulating levels of PDGFRB are progressively increased with increasing fibrosis stage | [31,61] | |
| SPHK1 | mediates hepatic inflammation in mice | [62,63] | |
| ITGA1 | facilitates hepatic insulin action while promoting lipid accumulation in mice | [64] | |
| ACKR1 | related to leucocyte recruitment by cirrhotic endothelial cells | [65,66,67] | |
| GPNMB | increased serum levels in NASH | [68] | |
| SCARB1 | SCARB1 deficiency increased inflammatory dyslipidaemia and adipocytes hypertrophy | [69] | |
| FGFR1 | central player in the response to liver injury and fibrosis | [70] | |
| TNFSF10 | increased liver expression levels in NASH | [71] | |
| CD68 | Increased liver infiltration with CD68+ macrophages are related to liver fibrosis | [72,73] | |
| NOTCH | NOTCH1 | directly involved in NAFLD development | [74,75,76,77,78,79] |
| NOTCH2 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| DLL1 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| DLL3 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| DLL4 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| JAG1 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| JAG2 | directly involved in NAFLD development | [74,75,76,77,78,79] | |
| WNT/β-catenin | WNT1 | related to hepatic glucose oxidation in NASH | [80] |
| WNT3a | related to hepatic glucose oxidation in NASH | [80] | |
| WNT5a | related to hepatic glucose oxidation in NASH | [80] | |
| WNT11 | related to hepatic glucose oxidation in NASH | [80] | |
| WNT2 | increased expression levels in liver pericentral endothelial cells in NASH | [81] | |
| RSPO3 | increased expression levels in liver pericentral endothelial cells in NASH | [81] | |
| WNT9b | altered liver expression levels in NASH | [82] | |
| WNT4 | altered liver expression levels in NASH | [82] | |
| FZD7 | modulated levels by lifestyle intervention in NAFLD patients | [83] | |
| LGR4/5 | its activity promotes NAFLD | [84] | |
| LRP6 | Mutations in LRP6 are one of the major causes of NAFLD induction | [85] | |
| DKK1 | related to hyperlipidaemia in NAFLD | [85] | |
| β-catenin | efector of the pathway. It has been found in EVs | [86,87,88] | |
| Plasma/serum secreted proteins in NAFLD | AFM | desregulated plasma levels in NAFLD and NASH patientis | [89,90] |
| PIGR | desregulated plasma levels in NAFLD and NASH patientis | [89,91,92,93] | |
| FTCD | proposed indicative biomarker for NAFLD to NASH progression | [22] | |
| Inter-tissue crosstalk | CD40 | CD40 expressing CD11c+ dendritic cells contribute to liver inflammation in NASH | [94,95] |
| LRG1 | adipokine that mediates obesity-induced hepatosteatosis and insulin resistance | [96,97] | |
| FNDC5 | serum irisin levels are reduced in patients with obesity-related NAFLD | [98,99] | |
| AKR1B7 | AKR1B7 in EVs derived from metabolic stressed adipocytes induce NASH in mice | [100] | |
| FASN | drives de novo lipogenesis, inflammation and fibrogenic signalling in NAFLD | [101,102] | |
| FABP4 | High levels of circulating FABP4 have been described in NAFLD patients | [103,104,105] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



