
Submitted:
21 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
This work aimed at producing lactic acid (LA) from sugarcane bagasse after steam explosion at 195 ºC for 7.5 and 15 min. Enzymatic hydrolysis was carried out with Novozymes’ Cellic CTec3 and/or Cellic HTec3, whereas fermentation was performed with Bacillus coagulans DSM2314. Water-washing of pretreated solids before enzymatic hydrolysis improved both hydrolysis and fermentation yields. The presence of xylo-oligosaccharides (XOS) in substrate hydrolysates reduced hydrolysis efficiency, but their effect on fermentation was negligible. The presence of fermentation inhibitors in C5 streams was circumvented by adsorption on activated carbon powder with no detectable sugar losses. High carbohydrates-to-LA conversions (Yp/s) of 0.88 g·g-1 were obtained from enzymatic hydrolysates of water-washed steam-exploded materials that were produced at 195°C, 7.5 min and the use of centrifuged-but-never-washed pretreated solids decreased Yp/s by 16%. However, when the detoxified C5 stream was added at a 10% ratio, Yp/s was raised to 0.93 g·g-1 for an LA productivity of 2.55 g·L-1·h-1. Doubling the pretreatment time caused a decrease in Yp/s to 0.78 g·g-1, but LA productivity was the highest (3.20 g·L-1·h-1). For pretreatment at 195°C for 7.5 min, elimination of water washing seemed feasible, but the use of longer pretreatment times made it mandatory to eliminate fermentation inhibitors.
Keywords:
Sugarcane bagasse
; Steam explosion
; Enzymatic hydrolysis
; Fermentation
; Lactic acid
1. Introduction
Lactic acid is a valuable chemical platform with applications in different industrial sectors such as food, cosmetics, textiles, pharmaceuticals, and chemical synthesis [1]. The global lactic acid market increased from 1,220 kilotons in 2016 to 1,960 kilotons in 2025. This represents the circulation of USD 11.51 billion globally [2,3]. The production of polylactic acid (PLA), a biodegradable polymer, accounts for about 50% of the lactic acid demand [4]. The other half is mainly used as acidulant, preservative, flavoring, emulsifier, and pH regulator in the food industry [5].
Lactic acid has two types of enantiomers (L or D). The pure enantiomers have greater value than the racemic mixture because they are used for special industrial applications. For instance, the L isomer is preferable for food, beverages, and pharma because it is metabolized more rapidly by the human body than the D isomer. L-Lactic acid is used in the synthesis of poly-L-lactic acid (PLLA), while D-lactic acid is used to produce poly-D-lactic acid (PDLA). Both are semi-crystalline bioplastics, while PDLLA, made with the racemic mixture, is amorphous and relatively easy to break down, being ideal for the development of drug delivery systems. However, the L isomer predominates in biomedical applications including bone fixation supports and biodegradable sutures due to its biocompatibility and high mechanical strength [6]. Food packaging, injection molding, and additive manufacturing (3D printing) are other applications in which PLA is widely used [7].
Optically pure lactic acid can be produced by fermentation while chemical synthesis leads to racemic mixtures [8]. Fermentation is also advantageous because it is based on renewable resources and mild process conditions [9]. Several bacteria, fungi, and yeasts can produce lactic acid of high optical purity in high yields [10]. However, lactic acid bacteria (LAB) such as Lactobacillus delbruckii, Lb. rhamnosus, Lb. casei, and Lb. plantarum, and bioengineered bacteria such as Escherichia coli and Corynebacterium spp., are the most widely used for lactic acid fermentation [2,11]. The process is performed by submerged fermentation and the substrate accounts for almost 70% of the production cost. Starch is the predominant raw material for industrial manufacturing, with about 90% of globally traded lactic acid being produced from corn [12]. Thus, the identification of cheaper and widely available substrates is pivotal to reduce process costs [13].
Agro-industrial residues such as sugarcane bagasse (SCB) are lignocellulosic materials with high potential to reduce industrial costs. However, plant cell wall polysaccharides such as cellulose and hemicelluloses must be converted to simple sugars (mostly glucose and xylose) before fermentation. Pretreatment techniques such as hydrothermolysis, steam explosion, acid-catalyzed organosolv, and dilute acid hydrolysis can provide high yields of fermentable sugars in the form of C5 and C6 streams [14]. C5 sugars are obtained in pretreatment acid hydrolysates while C6 sugars derive from enzymatic hydrolysis of pretreated cellulosic materials. However, the use of high pretreatment temperatures (>200 °C) and/or long residence times result in partial dehydration of pentoses and hexoses, causing the release of fermentation inhibitors such as furfural and 5-(hydroxymethyl)furfural (5-HMF) that reduce process yields [15,16]. Such aromatic aldehydes are known to inhibit key enzymes of the microbial carbon metabolism [17]. Other inhibitors may also be released from lignin such as low molar mass phenolic compounds. Also, mild pretreatment severities are already enough to release inhibitory acetic acid (pKa = 4.76) from hemicellulose O-acetyl groups [16]. Weak acids diffuse through the cell membrane and lower the intracellular pH, having implications on cell growth due to their effect on the plasma membrane proton transport activity [18,19].
Steam explosion uses saturated steam at high pressures to produce pretreated cellulosic materials with high accessibility to enzymatic hydrolysis [20]. While enzymatic hydrolysates of water-washed steam-exploded materials are easy to ferment, the C5 fraction normally contains inhibitory concentrations of organic acids (mostly acetic) and furan compounds (mostly furfural) [21]. Oligosaccharides released from hemicelluloses can also act as inhibitors for enzymatic hydrolysis [22] and, in both situations, the release of inhibitory compounds will be largely dependent on pretreatment conditions and feedstock composition. High pretreatment severities will release more fermentation inhibitors by carbohydrate dehydration and lignin hydrolysis, while oligosaccharides will prevail at low severities, particularly when pretreatment is carried out in the absence of an exogenous acid catalyst [23].
Several detoxification techniques have been used to reduce the inhibitory effect of biomass acid hydrolysates. Furans can be removed by physical adsorption [24], liquid-liquid extraction [25], evaporation [26], freeze-drying [27], enzymatic treatments using laccases and other oxidative enzymes [28], or overliming [29], while reducing acetic acid to non-inhibitory concentrations may be more problematic. The most widely used detoxification techniques are adsorption on activated carbon powder and overliming, but their efficiency depends on the type and concentration of fermentation inhibitors that are released in pretreatment liquors [30].
Some strains are more tolerant to inhibitory compounds. In a hydrolysate broth containing 4.01 g·L-1 acetic acid, 0.08 g·L-1 formic acid, 0.05 g·L-1 furfural, and 0.08 g·L-1 5-HMF, B. coagulans IPE22 converted 96% of sugars into LA [31]. Also, B. coagulans JI12 could tolerate up to 20 g·L-1 acetic acid and 4 g·L-1 furfural by metabolizing it to 2-furoic acid [32], while Bacillus sp. P38 was tolerant to 10 g·L-1 furfural and to 6 g·L-1 vanillin or acetic acid [33]. This indicates that Bacillus spp. may be promising organisms to produce L-LA from biomass hydrolysate without the use of a robust detoxification step.
Second-generation lactic acid can be produced by C5 plus C6 fermentation, or by co-fermentation of C5/C6 mixtures. For acid pretreatments such as steam explosion, C6 sugars are mostly arisen from enzymatic hydrolysis of water-washed pretreated materials, while C5 sugars are recovered in pretreatment liquors (C5 streams) that must be detoxified prior to fermentation. B. coagulans has become one of the most popular organisms for this purpose due to its capacity to metabolize C5 sugars via the pentoses phosphate (PP) pathway and to produce optically pure L-LA with high yields [34]. Enzymatic hydrolysis and fermentation can be performed separately or simultaneously. Based on this, different bioprocessing strategies have been designed to produce biobased materials such as separate hydrolysis and fermentation (SHF), separate hydrolysis and co-fermentation (SHCF); simultaneous saccharification and fermentation (SSF), and simultaneous saccharification and co-fermentation (SSCF). SHF and SHCF involve enzymatic hydrolysis of the polysaccharides and subsequent fermentation of the sugars released. By contrast, SSF and SSCF are one-pot methods where enzymatic hydrolysis and microbial fermentation occur simultaneously. Combining these operations results in lower capital costs and higher productivities since sugars are promptly consumed by the fermenting microorganism and enzymes perform better due to lower levels of end-product inhibition [33-40].
Michelson et al. [41] compared the performance and yield of two LA producers, Lb. delbrueckii ssp. lactis DSM 20073 and B. coagulans SIM-7. The latter strain achieved final LA concentrations of 91.5 g·L-1 and 91.6 g·L-1 in batch and fed-batch cultivations for 23 and 21 h, respectively. The LA concentration in 10 h was already 56 g·L-1, whereas comparable results (52 g·L-1) were achieved only in 24 h by DSM 20073. The maximal production rates of SIM-7 and DSM 20073 strains were 9.9 and 5.6 g·L-1·h-1, respectively.
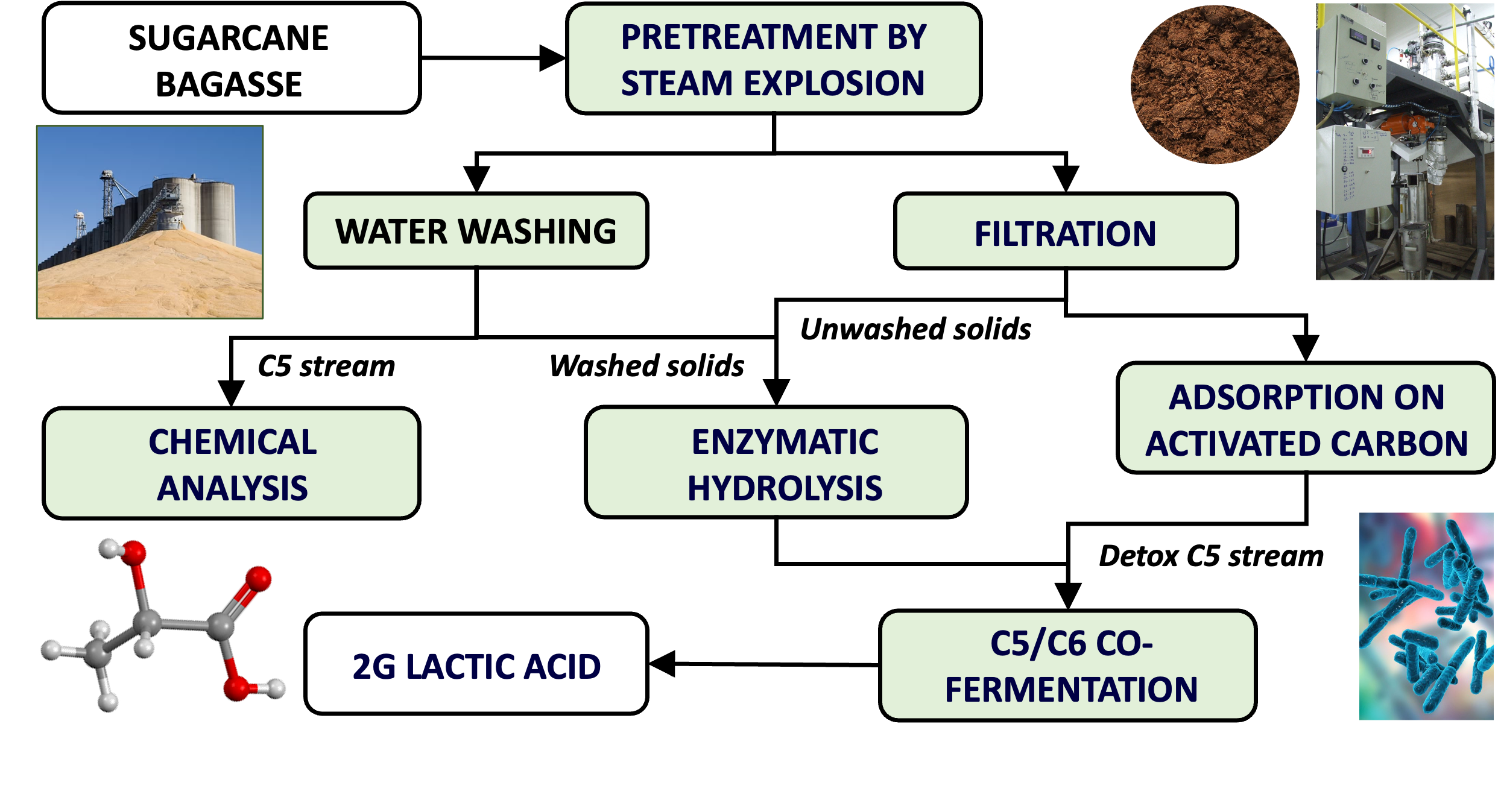
In this present work, different enzymatic hydrolysis and fermentation conditions were used to produce LA from steam exploded SCB. Fermentation inhibitors were removed from C5 streams using physical adsorption on activated carbon powder, while fermentation was carried out with B. coagulans DSM2314 using both SHF and SHCF protocols.
2. Material and Methods
2.1. Sugarcane Bagasse (SCB) Pretreatment and Characterization
Fresh SCB was kindly donated by Raízen (Piracicaba, SP, Brazil). Pretreatment was carried out by steam explosion at 195°C for 7.5 and 15 min using a 10-L stainless-steel high-pressure steam reactor and SCB with a moisture content of 50 wt% [27]. Pretreatment slurries (20-25 wt% total solids) were centrifuged inside a cotton fiber bag to remove water-soluble hemicellulose sugars and low molar mass lignin components (C5 stream). Half of the unwashed centrifuged fiber cake (SEB7.5-UW and SEB15-UW) was reserved for enzymatic hydrolysis, while the other half was water-washed at 5 wt% TS for 1 h at room temperature (~25°C) under constant mechanical stirring, followed by centrifugation to recover the water-washed fiber cake (SEB7.5-WW and SEB15-WW) for its subsequent characterization and enzymatic hydrolysis. Both SEB-UW and SEB-WW substrates, with 35-40 wt% total solids after centrifugation, were stored in vacuum-sealed plastic bags at 4°C prior to chemical characterization and/or enzymatic hydrolysis.
The composition of untreated and pretreated materials was characterized following National Renewable Energy Laboratory (NREL) protocols for total moisture [42], ash [43], total extractives [44], and carbohydrates plus total lignin content (acid-soluble lignin and acid-insoluble lignin) [45]. All reagents, solvents, and chromatographic standards were obtained in analytical grade and used as received. Mass balances and process yields were calculated according to the above-mentioned standard procedures. To this end, theoretical conversion factors were considered to express recovery yields in relation to each raw SCB macromolecular component.
The pretreatment liquor (C5 fraction) was detoxified over activated carbon [46]. Detoxification was performed in 250 mL Erlenmeyer flasks that were loaded with 100 mL of liquid and 10 g of activated carbon powder (Neon®, Brazil) having a surface area of 507.9 m²·g-1 and an average pore size of 1.29 nm. The flasks were covered with Parafilm M to prevent evaporation and placed on a shaker incubator at 25 °C and 120 rpm for 10 min. Then, the suspension was then centrifuged at 2500 rpm for 15 min and aliquots were removed from the supernatant, filtered through a 0.22 μm PVDF filter (Millipore), and analyzed by high performance liquid chromatography (HPLC) to quantify furfural, 5-HMF, acetic acid, formic acid, xylose, glucose, and arabinose, using the chromatographic conditions that are described in Section 2.4. Then, the supernatant was passed through a 0.1-µm ash-less quantitative filter paper (Whatman®) to remove any remaining suspended solids. The detoxification process was repeated 3 times to ensure that acetic acid, furfural, and 5-HMF were brought to non-inhibitory concentrations.
2.2. Enzymatic Hydrolysis
The commercial enzymes used for hydrolysis were provided by Novozymes Latin America (Araucária, SP, Brazil). Cellic CTec3 is a commercial cellulase preparation while Cellic HTec3 contains hemicellulase activity predominantly [20]. Enzyme loading was always based on the wet weight of the commercial enzyme preparation that was added to the reaction system for enzymatic hydrolysis.
Enzymatic hydrolyses of SEB-UW and SEB-WW substrates were performed at 50 °C and 150 rpm for 96 h in acetate buffer (50 mmol·L-1, pH 5.2) using 250 mL Erlenmeyer flasks in a shaker incubator (Ecotron, Infors HT). The best condition was derived from a factorial design that was based on three independent variables in two levels (23): substrate TS (10 and 20 wt%), Cellic CTec3 loading (20 to 60 mg g-1 TS), and Cellic HTec3 addition in a 10% mass ratio (wet basis) in relation to Cellic CTec3 (2 to 6 mg·g-1 TS) [20]. Four quadratic polynomial equations were obtained using the R Studio® 3.4.3 software to describe the mathematical relationship between glucose release (g·L-1) and the selected process variables. The goodness-of-fit of the models was evaluated by determining their adjusted R2. For yield calculations, aliquots were collected at different incubation times and analyzed by HPLC to quantify cellobiose, glucose, and xylose using the chromatographic conditions described in Section 2.4. Hydrolysis yields were determined in percentage by expressing the total glucose release (glucose equivalents) in relation to the total glucose content (quantified as glucans) of the pretreated solids. Xylose was not considered in yield calculations because it was always found in very low quantities.
Enough substrate hydrolysate (C6 stream) for fermentation was obtained by performing the best hydrolysis conditions from the factorial design in a 3.6L Labfors bioreactors (Infors HT, Bottmingen-Basel, Switzerland). Enzymatic hydrolyses of SEB-UW and SEB-WW substrates were performed at 50 °C and 150 rpm for 72 h in acetate buffer (50 mmol·L-1, pH 5.2) using 20 wt% TS and 60 mg g-1 TS of Cellic CTec3, with and without addition of Cellic HTec3 (6 mg·g-1 TS). The total volume of this reaction system was 1000 mL. Aliquots were collected once again at different incubation times and analyzed by HPLC, and hydrolysis yields were determined as described above.
2.3. Microorganism and Fermentation
B. coagulans DSM2314 was acquired as a freeze-dried stock from the Germany Collection of Microorganisms and Cell Cultures (DSMZ, Leibniz Institute, Germany). Cells were grown on Man, Rogosa, and Sharpe (MRS) agar medium and transferred to 50 mL flasks of MRS medium to be cultured for 16 h at 50 °C. The media were pre-sterilized for 15 min at 121 ºC. When the optical density measured at 660 nm reached 2, the pre-culture was added as inoculum to the fermentation, which was carried out at 50 ºC and 150 rpm for 24 h in Multifors 2 bioreactors (Infors HT) that were pre-sterilized empty for 20 min at 121 ºC.
Both SHF and SHCF fermentation experiments were carried out in duplicate in the Multifors 2 bioreactor using a working volume of 300 mL for a total volumetric capacity of 500 mL. SCB hydrolysates (C5, C6 and C5/C6 mixtures) were transferred to the bioreactor vessel and added with 1% yeast extract and 10% (v/v) inoculum (30 mL). Temperature and agitation were set at 50°C and 150 rpm, respectively, and dilute NaOH (5 mol L-1) was used to maintain the fermentation broth at pH 6.0 during the entire reaction course. Fermentation ran for 24 h under anaerobic conditions by continuous N2 purging. Aliquots were taken at different times and analyzed by HPLC for carbohydrates and LA using the conditions described below. LA yields were determined as percentage in relation to the theoretical amount of LA that could have been produced from the fermentable sugars available in substrate hydrolysates.
2.4. Chromatographic Analysis
C5 (from pretreatment) and C6 (from enzymatic hydrolysis) streams and fermentations broths were analyzed at 65 °C using a Shimadzu HPLC, LC-20AD series and a Rezex RHM column (Phenomenex, 300 x 7.8 mm) that was preceded by a Carbo H guard column (300 x 7.8 mm). The column was eluted with 5 mmol L-1 H2SO4 at a flow rate of 0.6 mL min-1. Sample injection (20 µL) was made using a Shimadzu SIL-10AF autosampler. Quantitative analyses were carried out by external calibration using differential refractometry (Shimadzu RID-10A) for carbohydrates and organic acids, while UV spectrophotometry (Shimadzu SPD-M10AVP) at 280 nm was used to quantify furfural and 5-HMF. HPLC calibration curves were based on the analysis of six independent primary standard solutions, and the corresponding linear regression coefficients (R2) were always around 0.99.
2.5. Statistical Analysis
The Tukey's Test (p ≤ 0.05) was applied to evaluate the statistical significance of the experimental data and the experimental design was validated by analysis of variance (ANOVA) using the R Studio® 3.4.3 software [47]. For experiments carried out in two or three replicates, hydrolysis and fermentation yields were expressed as averages with their corresponding standard deviations.
3. Results
3.1. SCB Pretreatment and Characterization
The chemical composition of SCB before and after pretreatment is shown in Table 1. The untreated material had glucans (mostly cellulose), hemicelluloses (mostly xylans), total lignin, total extractives, and ash contents like those already reported elsewhere [20,21,27,31]. Variations in SCB chemical composition are attributed to its source and maturation stage upon harvesting, as well as the edaphoclimatic conditions used for cultivation and the technology used for its industrial processing [48].
Steam explosion at 195°C for 7.5 and 15 min reduced the SCB hemicellulose content by 85.7 and 94.3%, respectively, with a corresponding rise in both glucans and total lignin content (Table 1) [49]. Hemicelluloses were almost entirely depleted of their arabinosyl residues and acetyl groups because arabinose and acetic acid were not detectable by HPLC in sulfuric acid hydrolysates. Pentoses and hexoses were partly detected as furfural and 5-HMF due to dehydration, but in both situations, the reported values were not added to the corresponding polysaccharide quantification because their actual source was not elucidated. Hence, SCB hemicellulose content would be the summation of xylans, arabynosyl residues, pentoses identified as furfural and acetyl groups, totaling 25.5%. Likewise, the total glucan content in Table 2 should be estimated at 38.9%, even though some 5-HMF may have come from hemicelluloses as well. 5-HMF formation was higher in sulfuric acid hydrolysates because furfural was partially involved in side-reactions producing humins, while furans were not formed after acid hydrolysis of steam-treated materials because their hemicellulose content was very low [50,51].
Table 1 reveals the total recovery of SCB components in water-washed steam-exploded materials, in which the mass recovery of pretreated solids was also considered. Glucan recovery was around 90% when steam explosion was carried out at 195°C for 7.5 min but doubling the pretreatment time to 15 min decreased this value by roughly 9%. The hemicellulose content (mostly xylans) in steam-exploded materials was very low and some lignin condensation may have occurred particularly at the highest pretreatment severity, in which lignin recovery was above 100%. Finally, ash recoveries above 100% may have been indicative of partial corrosion and/or abrasion of reactor walls [21,27].
3.2. Chemical Composition of Pretreatment Liquors before and after Detoxification
Pretreatment liquors were characterized by HPLC for their carbohydrate, acetic acid, 5-HMF, and furfural contents before and after detoxification with activated carbon powder (Table 2). Post-hydrolysis of these fractions with dilute sulfuric acid revealed xylose concentrations of 13.2 ± 0.4 and 10.5 ± 0.5 g·L-1 for pretreatments carried out for 7.5 and 15 min, respectively. The presence of acetic acid in pretreatment liquors was due to hemicellulose deacetylation, and its concentration was higher when pretreatment was carried out at more drastic conditions (p < 0.05). Acetic acid coming from O-acetyl groups are partially responsible for the auto-hydrolysis effect, which converts hemicelluloses to mono and oligosaccharides even in the absence of an exogenous acid catalyst. Furfural and 5-HMF derived from pentose and hexose dehydration, respectively, were also detected in pretreatment liquors in concentrations that increased with pretreatment severity. Accumulation of organic acids, furans, and phenolic compounds in pretreatment liquors is not desirable due to their inhibitory effect on hydrolysis and fermentation [52,53].
Controlling microbial inhibition is essential for maximizing biomass conversion. Du et al. [54] identified and quantified 40 potentially toxic compounds after performing eight chemical pretreatment techniques on three different lignocellulosic materials. Fockink et al. [20] demonstrated that the autocatalytic steam-explosion of SCB at 205 ºC induced the accumulation of formic, levulinic and acetic acid in the C5 fraction. In relation to furan compounds, furfural was present in higher concentration, while the accumulation of 5-HMF and 5-methyl-furan (5-MF) was higher at the highest pretreatment temperatures. This is a clear indication that a detoxification stage must be carried out for pretreatment liquors in which the presence of fermentation inhibitors is unavoidable [20.21].
The data presented in Table 2 showed that, by treating SCB pretreatment liquors with activated carbon powder, furfural and 5-HMF concentrations were reduced by 97-98% (p < 0.05), regardless of conditions used for pretreatment. For acetic acid, removals were 55.2 and 57.2% for pretreatments carried out at 195 ºC for 7.5 and 15 min, respectively. By contrast, no sugar losses were observed after adsorption on activated carbon powder at ambient temperature.
In our work, detoxification with activated carbon was highly efficient at room temperature, while other studies required heating to be effective. Lu, Dong, and Yang [55] reported the use of 2% (m·v-1) commercial activated carbon to remove 80% and 87.9% of furfural and 5-HMF present in wood chips acid hydrolysates, respectively. Adsorption was carried out at 90 ºC for 30 min under constant stirring (150 rpm). Miura, Suzuki, and Aoyama [56] detoxified wood acid hydrolysates using adsorption on 10% (m·v-1) activated carbon powder at 30 ºC for 1 h. Around 83% of furfural and other low molar mass phenolic compounds were removed, while carbohydrate and acetic acid concentrations remained practically unaltered. Better detoxification efficiencies may have been due to the better textural properties of the activated carbon used in our studies.
3.3. Enzymatic Hydrolysis
Enzymatic hydrolysis of unwashed (SEB-UW) and water-washed (SEB-WW) substrates was investigated using an experimental design that was based on the following variables: substrate TS, Cellic CTec3 loading, and Cellic HTec3 supplementation. The latter variable was introduced because pretreatment liquors contained oligosaccharides that are known to inhibit total cellulase activity [57,58]. The results obtained after 96 h of hydrolysis were subjected to multiple linear regression analyses to generate mathematical models that could describe trends in relation to the response function, which corresponded to the release of glucose equivalents in the reaction environment (in g·L-1). This was the only response function treated statistically because of its relevance for LA production since high concentrations of fermentable sugars are desirable to achieve high fermentation yields. Table 3 shows the mathematical equations that were generated to fit the experimental data for both SEB-UW and SEB-WW enzymatic hydrolyses, whereas Table 4 presents their corresponding analysis of variance (ANOVA). In general, the quadratic models developed to adjust the enzymatic hydrolysis data were adequate, with adjusted regression coefficients (R2) always above 0.99.
The highest glucan conversion (74.4%) was obtained from SEB15-WW in the presence of Cellic HTec3 for a final glucose concentration of 84.5 g·L-1 after 96 h or hydrolysis (Table 5). This value was 25% higher than what was obtained with the same substrate in the absence of Cellic HTec3 (67.2% glucan conversion for a 78.25 g·L-1 glucose concentration). In fact, Cellic HTec3 improved the hydrolysis performance of Cellic CTec3 in all reaction configurations. Therefore, residual xylans that were retained in steam-exploded materials seemed to have a role in limiting cellulose accessibility. Also, for SEB-UW substrates, additional hemicellulase activity helped converting water-soluble xylo-oligomers to fermentable xylose, justifying the achievement of slightly higher xylose recoveries. However, it is worth noticing that Cellic HTec3 contains some residual cellulase activity (~5 FPU g-1) and this may have been partially responsible for its boosting effect over Cellic CTec3.
SEB-WW was better than SEB-UW for enzymatic hydrolysis, but the difference was minor for substrates produced at the lowest severity (195 ºC, 7.5 min). Comparing the total processing time and the resulting glucose concentration, water washing seems to be dispensable when pretreatment was carried out for 7.5 min, but mandatory when pretreatment was carried out 15 min. This suggests that, by doubling the reaction time, the accumulation of potential hydrolysis inhibitors in the C5 stream became critical and this could not be attributed to XOS because these oligos tend to be almost completely hydrolysed under more drastic pretreatment conditions. On the other hand, in both scenarios, better substrates for hydrolysis were produced when steam explosion was carried out for a longer reaction time or higher pretreatment severity.
Enzymatic hydrolysis of pretreated cellulosic materials was subsequently scaled-up by a factor of 10. Selected hydrolysis condition (20 wt% TS and 60 mg g-1 TS of Cellic CTec3 plus 6 mg g-1 TS of Cellic HTec3) were used in the Infors-HT bioreactor to produce enough substrate hydrolysate for fermentation [20,22,59] Figure 1 shows the hydrolysis profile of both SEB-UW and SEB-WW, with the results given in glucose release in g·L-1. The effects of pretreatment severity, water washing, and Cellic HTec3 supplementation on hydrolysis efficiency were the same as those observed in shake flasks. Cellobiose was always within 1 and 3% of the total glucose release, meaning that the b-glucosidase activity of Cellic CTec3 was high enough to keep it below inhibitory levels. Xylan conversion was higher for SEB7.5-UW and this may have been caused by the xylose concentration of the C5 stream that was retained in the steam-exploded material after centrifugation.
3.4. Fermentation of SEB Pretreatment Liquors
SEB pretreatment liquors and enzymatic hydrolysates were fermented either alone or in combination using SHF and SHCF protocols. Initially, both fractions were fermented separately to evaluate the influence of detoxification on fermentation yields. There was a visual difference in the turbidity of the media (increase in cell biomass) and a significant difference in LA production between undetoxified and detoxified fractions (p < 0.05). Table 6 shows a comparison between the fermentation profiles of SEB7.5 and SEB15 pretreatment liquors.
After fermentation, LA contents in non-detoxified pretreatment liquors were lower than those obtained from detoxified fractions for both pretreatment conditions. LA production from non-detoxified media was 2.3 and 1.7 g·L-1 for SEB7.5 and SEB15, respectively. These values are statistically equal at a 95% confidence level, meaning that pretreatment liquors that do not go through detoxification have similar fermentation performances. By contrast, LA contents from detoxified samples were 13.4 and 6.6 g·L-1, respectively. Lower residence times into the steam reactor led to lower levels of carbohydrate degradation and lower release of fermentation inhibitors in pretreatment hydrolysates. By contrast, the poor fermentation performance of non-detoxified samples was attributed to the presence of inhibitory compounds such as furans, organic acids, and phenolic compounds.
Oliveira et al. [60] used a synthetic media containing xylose and glucose to produce LA, as well as SCB hemicellulose hydrolysates that were obtained by acid hydrolysis at 10% TS using 0.5% (v·v-1) HCl at 140 °C for 15 min. This acid hydrolysate, containing 48 g·L-1 xylose, 7.86 g·L-1 glucose, 0.08 g·L-1 5-HMF, and 0.01 g·L-1 furfural, was supplemented with 20 g·L-1 yeast extract and fermented with B. coagulans 14-300 to produce 56 g·L-1 lactic acid for a theoretical yield of 87%. However, an evaporation step was added to concentrate the acid hydrolysate, and this may have reduced its already low concentration of inhibitory compounds [21,27].
Ahorsu et al. [61] used B. coagulans DSM2314 for the fermentation of nutshell hydrolysates that were obtained by microwave-assisted autohydrolysis. The experiments were carried out at 190 ºC for up to 25 min in an equipment configuration that allowed five reactions to occur simultaneously. An LA productivity of 0.2 g·L-1·h-1 was obtained, which corresponded to a 93% xylose conversion (6 g·L-1 of LA). Furthermore, the presence of 0.45 g·L-1 furfural and 2.42 g·L-1 acetic acid did not affect the B. coagulans DSM2314 fermentation performance. However, the presence of xylo-oligosaccharides was not investigated, although probably present in pretreatment liquors, while fermentation times were up to 48 h.
van der Pol et al. [62] pre-cultured B. coagulans DSM2314 in the presence of fermentation inhibitors using glucose and xylose as carbon source. Inhibitors were found in the following concentrations: 1.6 g·L-1 furfural, 0.2 g·L-1 HMF, and 3.1 g·L-1 acetic acid. Although an increase in LA production was observed in the presence of furfural, fermentation trials were only carried out in synthetic media that simulated the carbohydrate composition of hemicellulose hydrolysates. Furthermore, the observed increase in productivity was not directly associated to furfural consumption or conversion.
A possible explanation for the albeit small formation of LA in the non-detoxified environment (Table 6, Figure 2) would be that, despite the toxicity of furfural being mainly caused by the formation of reactive oxygen species [63,64], several B. coagulans genes are known to encode for enzymes such as superoxide dismutase and catalase that are able to reduce these reactive species to less inhibitory compounds such as 2-furoic acid [65]. Since the DSM2314 strain is catalase-positive, this mechanism may be involved in its tolerance to the presence of furfural [62]. Ye et al. [66] used B. coagulans JI12 to ferment cellulosic and hemicellulosic hydrolysates from oil palm empty fruit bunches that contained glucose and xylose in different proportions (1:10 and 1:1). The strain was able to ferment both hydrolysates in the presence of 4 g·L-1 furfural, which was partially metabolized to 2-furoic acid.
3.5. Fermentation of SEB-UW and SEB-WW Enzymatic Hydrolysates
Table 7 presents the fermentation profile of both SEB-UW and SEB-WW enzymatic hydrolysates. C6 (glucose) fermentation predominated in both systems, but the former involved more C5/C6 co-fermentation because the substrate retained part of the pretreatment liquor (C5 stream) after centrifugation. Carbohydrate-to-LA conversions (Yp/s) of 0.88 and 0.93 g·g-1 were achieved for SEB7.5-WW and SEB7.5-UW, while these values decreased to 0.61 and 0.78 g·g-1 for SEB15-WW and SEB15-UW, respectively. Therefore, UW hydrolysates were not inhibitory to both LA fermentation and microbial growth (see OD600 values in Table 7). Also, B. coagulans DSM2314 consumed both glucose and xylose indistinctively, as demonstrated by the percentages of residual sugar that were detected after fermentation. On the other hand, lower Yp/s values for both SEB15-WW and SEB15-UW hydrolysates were probably due to the higher osmotic stress caused by applying higher initial glucose concentrations in the fermentation media.
For SHCF co-fermentation experiments, the C5/C6 ratio was based on van der Pol et al. [62], which was achieved by combining pretreatment liquors (C5 stream) and SEB-WW enzymatic hydrolysates to achieve 10% C5 and 90% C6. For this, pretreatment liquors were used without any further treatment (e.g., filtration or rotary evaporation) or after physical adsorption on activated carbon power to eliminate most fermentation inhibitors (detoxification). Table 8 presents the fermentation profile for SEB-WW enzymatic hydrolysates to which non-detoxified or detoxified C5 streams were added. Addition of non-detoxified pretreatment liquor to the corresponding SEB7.5-WW enzymatic hydrolysate decreased fermentation efficiency (Yp/s) by ~90%, from 0.88 ± 0.01 to 0.07 ± 0.02 g·g-1. This fact was observed by the low conversion of carbohydrates in the medium and the low cell density at the end of fermentation. Likewise, Yp/s decreased from 0.93 ± 0.01 to 0.07 ± 0.01 g·g-1 after adding the untreated C5 stream to SEB7.5WW hydrolysates. Nevertheless, B. coagulans DSM 2314 was able to produce 4.6 g·L-1 LA in the presence of fermentation inhibitors, demonstrating its ability to adapt to relatively high concentrations of furanic compounds such as furfural. It is important to notice that a possible elimination of washing stage decreased LA production from 64.2 g·L-1 to 48.8 g·L-1 LA for SEB7.5, nearly 24% of reduction. SEB15 also showed decrease of LA, from 56.9 g·L-1 to 52.4 g·L-1, nearly 8%. In fact, these results can be explained by the fact that the washed substrates have had better hydrolysis performance, and that also impacted on the fermentation steps.
Organic acids and phenolic acids are toxic to the bacteria because they can cross the cell membrane, decreasing the intracellular pH and causing damage to cell functions. Upon inhibition, the metabolic energy is spent to maintain homeostasis instead of being used for cell growth. Protein denaturation, metabolism inhibition, and cell death may also occur in the presence of potent inhibitory compounds [67-69].
Cubas-Cano et al. [34] studied the effect of inhibitory compounds on lactic acid fermentation using B. coagulans A162 and DSM2314 strains. Acid hydrolysates from acid-catalysed steam explosion of garden plant waste (180ºC for 10 min plus 60 mg·g-1 H2SO4) were used as carbon source. After pretreatment, the hemicellulose hydrolysate presented the following composition: 5 g L−1 glucose, 15 g L−1 xylose, 3.4 g L−1 arabinose, 0.20 g L−1 furfural, 0.23 g L−1 5-HMF, 1.15 g L−1 acetic acid, and 0.23 g L−1 formic acid. The highest LA productivity of 2.4 g·L-1·h-1 was attained by the A162 strain, which was tolerant to the presence of fermentation inhibitors at these concentrations.
van der Pol et al. [70] applied pSSF for LA production using H2SO4-impregnated steam-exploded SCB (170 ºC for 15 min using 0.72 vol% H2SO4 in relation to SCB dry mass). The substrate was pre-saccharified for 4 to 6 h with an enzyme cocktail containing both xylanase and cellulase activities (15 FPU·g-1 Genencor GC220 in relation to the substrate dry mass), when an inoculum (5%, v·v-1) that was pre-cultured in the presence of furfural was added to the medium. LA production reached 70.4 g·L-1 in 68 h, representing 89.7% carbohydrate conversion and an estimated productivity of 0.98 g·L-1·h-1. LA concentrations above 90 g·L-1 have already been reported in other studies for fermentation times up to 50 h [71].
4. Conclusions
Steam explosion of sugarcane bagasse was successfully used to produced second generation lactic acid using SHF and SHCF conversion strategies. The pretreatment liquor (C5 stream), separated from the pretreated solids by centrifugation, was shown to be highly inhibitory to B. coagulans DSM2314, but this problem was circumvented by adsorption of fermentation inhibitors onto activated carbon powder. Furan compounds such as furfural and 5-HMF were almost completely removed, while acetic acid concentrations decreased by nearly 50%. Enzymatic hydrolysates were produced from unwashed and water-washed pretreated solids and fermented to LA under anaerobic conditions. For steam explosion at 195°C for 7.5 min, elimination of this washing stage seemed feasible, but the use of higher pretreatment severities (195°C for 15 min) made it mandatory to avoid fermentation inhibition. The highest LA yields were obtained from enzymatic hydrolysates of water-washed steam-exploded materials where little or no inhibitory effect was found. SHF and SHCF produced 64.2 and 61.4 g∙L-1 LA from materials pretreated for 7.5 min, respectively, while samples pretreated for 15 min gave 56.9 g∙L-1 LA by SHF and 76.7 g∙L-1 by SHCF. Under SHCF conditions, the C5 stream had to be detoxified prior to lactic acid fermentation.
Author Contributions
Conceptualization, W.R.A., T.A.S., and L.P.R.; methodology, W.R.A., T.A.S., and A.Z.F.; validation, W.R.A., T.A.S., and L.P.R.; formal analysis, L.P.R. and A.Z.F.; investigation, W.R.A.; resources, L.P.R.; writing—original draft preparation, W.R.A., T.A.S., and L.P.R.; writing—review and editing, L.P.R; supervision, T.A.S. and L.P.R.; project administration, L.P.R.; funding acquisition, L.P.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Council of Technological and Scientific Development (CNPq, Brazil – Grants 309506/2017-4 and 315930/2021-7), and by the Coordination for the Improvement of Higher Education Personnel (CAPES – Financial Code 001).
Acknowledgments
The authors are grateful to Raízen (Piracicaba, SP, Brazil) and Novozymes Latin America (Araucária, PR, Brazil) for donating batches of sugarcane bagasse and enzymes for hydrolysis, respectively.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Alsaheb, R. A. A.; Aladdin, A.; Othman, N. Z.; Malek, R. A.; Leng, O. M.; Aziz, R.; Enshasy, H. A. El. Lactic Acid Applications in Pharmaceutical and Cosmeceutical Industries. J. Chem. Pharm. Res. 2015, 7, 729–735. [Google Scholar]
- Abedi, E.; Hashemi, S. M. B. Lactic Acid Production – Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Grand View Research. Lactic acid market size, share & trends analysis report by raw material (sugarcane, corn, cassava), by application (PLA, food & beverages, personal care, pharmaceuticals), by region, and segment forecasts, 2022–2030. 2021.
- Singhvi, M.; Joshi, D.; Adsul, M.; Varma, A.; Gokhale, D. D-(-)-Lactic Acid Production from Cellobiose and Cellulose by Lactobacillus lactis Mutant RM2-24. Green Chem. 2010, 12, 1106–1109. [Google Scholar] [CrossRef]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F. J.; Saraiva, J. A.; Sant’Ana, A. S.; Hashemi, S. M. B. Recent Advancements in Lactic Acid Production - a Review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Shah, S. R.; Walker, J. L.; Mikos, A. G. Poly(lactic acid) nanofibrous scaffolds for tissue engineering, Adv. Drug Deliv. Rev. 2016, 107, 206–212. [Google Scholar] [CrossRef]
- Tsuji, H.; Fukui, I. Enhanced Thermal Stability of Poly(Lactide)s in the Melt by Enantiomeric Polymer Blending. Polymer 2003, 44, 2891–2896. [Google Scholar] [CrossRef]
- Komesu, A.; de Oliveira, J. A. R.; Martins, L. H. da S.; Maciel, M. R. W.; Maciel Filho, R. Lactic Acid Production to Purification: A Review. BioResources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Reddy, L. V.; Park, J.; Wee, Y. Homofermentative Production of Optically Pure L -Lactic Acid from Sucrose and Mixed Sugars by Batch Fermentation of Enterococcus faecalis RKY1. Biotechnol. Bioprocess Eng. 2015, 1105, 1099–1105. [Google Scholar] [CrossRef]
- Nwamba, M. C.; Sun, F.; Mukasekuru, M. R.; Song, G.; Harindintwali, J. D.; Boyi, S. A.; Sun, H. Trends and Hassles in the Microbial Production of Lactic Acid from Lignocellulosic Biomass. Environ. Technol. Innov. 2021, 21, 101337. [Google Scholar] [CrossRef]
- Pleissner, D.; Demichelis, F.; Mariano, S.; Fiore, S.; Gutiérrez, I. M. N.; Schneider, R.; Venus, J. Direct production of lactic acid based on simultaneous saccharification and fermentation of mixed restaurant food waste. J. Clean. Prod. 2011, 143, 615–623. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, H.; Cai, D.; Wang, B.; Qin, P.; Wang, Z.; Tan, T. Improvement of L-Lactic Acid Productivity from Sweet Sorghum Juice by Repeated Batch Fermentation Coupled with Membrane Separation. Bioresour. Technol. 2016, 211, 291–297. [Google Scholar] [CrossRef]
- Krishna, B. S.; Sai, S.; Gantala, N.; Tarun, B. Industrial Production of Lactic Acid and Its Applications. Int. J. Biotech Res. 2018, 1, 42–54. [Google Scholar]
- Yoshimura, M.; Byrappa, K. Hydrothermal Processing of Materials: Past, Present and Future. J. Mater. Sci. 2008, 43, 2085–2103. [Google Scholar] [CrossRef]
- Bondesson, P.-M.; Galbe, M.; Zacchi, G. Ethanol and Biogas Production after Steam Pretreatment of Corn Stover with or without the Addition of Sulphuric Acid. Biotechnol. Biofuels 2013, 6, 11. [Google Scholar] [CrossRef]
- Carrasco, C.; Baudel, H. M.; Sendelius, J.; Modig, T.; Roslander, C.; Galbe, M.; Hahn-Hägerdal, B.; Zacchi, G.; Lidén, G. SO2-Catalyzed Steam Pretreatment and Fermentation of Enzymatically Hydrolyzed Sugarcane Bagasse. Enzyme Microb. Technol. 2010, 46, 64–73. [Google Scholar] [CrossRef]
- Wee, Y.; Yun, J.; Kim, D.; Ryu, H. Batch and repeated batch production of L-(þ)-lactic 2254 acid by RKY1 using wood hydrolyzate and corn steep liquor Enterococcus faecalis, J. Ind. Microbiol. Biotechnol. 2006, 33, 431–435. [Google Scholar] [CrossRef]
- Palmqvist, E.; Grage, H.; Meinander, N. Q. Main and Interaction Effects of Acetic Acid, Furfural, and p-Hydroxybenzoic Acid on Growth and Ethanol Productivity of Yeasts. Biotechnol. Bioeng. 1999, 63, 46–55. [Google Scholar] [CrossRef]
- Wang, L.-Q.; Cai, L.-Y.; Ma, Y.-L. Study on Inhibitors from Acid Pretreatment of Corn Stalk on Ethanol Fermentation by Alcohol Yeast. RSC Adv. 2020, 10, 38409–38415. [Google Scholar] [CrossRef]
- Fockink, D. H.; Urio, M. B.; Sa, J. H.; Ramos, L. P. Enzymatic Hydrolysis of Steam-Treated Sugarcane Bagasse: Effect of Enzyme Loading and Substrate Total Solids on Its Fractal Kinetic Modeling and Rheological Properties. Energy Fuels 2017, 31, 6211–6220. [Google Scholar] [CrossRef]
- Neves, P. V; Pitarelo, A. P.; Ramos, L. P. Production of Cellulosic Ethanol from Sugarcane Bagasse by Steam Explosion: Effect of Extractives Content, Acid Catalysis and Different Fermentation Technologies. Bioresour. Technol. 2016, 208, 184–194. [Google Scholar] [CrossRef]
- Xue, S.; Uppugundla, N.; Bowman, M. J.; Cavalier, D.; Da, L.; Sousa, C.; Dale, B. E.; Balan, V. Sugar Loss and Enzyme Inhibition Due to Oligosaccharide Accumulation during High Solids - Loading Enzymatic Hydrolysis. Biotechnol. Biofuels 2015, 8, 195. [Google Scholar] [CrossRef]
- Fockink, D. H.; Sánchez, J. H.; Ramos, L. P. Comprehensive Analysis of Sugarcane Bagasse Steam Explosion Using Autocatalysis and Dilute Acid Hydrolysis (H3PO4 and H2SO4) at Equivalent Combined Severity Factors. Ind. Crop. Prod. 2018, 123, 563–572. [Google Scholar] [CrossRef]
- Horváth, I. S.; Sjöde, A.; Nilvebrant, N. O.; Zagorodni, A.; Jönsson, L. J. Selection of Anion Exchangers for Detoxification of Dilute-Acid Hydrolysates from Spruce. Appl. Biochem. Biotechnol. 2004, 114, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Cantarella, M.; Cantarella, L.; Gallifuoco, A.; Spera, A.; Alfani, F. Comparison of Different Detoxification Methods for Steam-Exploded Poplar Wood as a Substrate for the Bioproduction of Ethanol in SHF and SSF. Process Biochem. 2004, 39, 1533–1542. [Google Scholar] [CrossRef]
- Larsson, S.; Reimann, A.; Nilvebrant, N.-O.; Jönsson, L. J. Comparison of Different Methods for the Detoxification of Lignocellulose Hydrolyzates of Spruce. Appl. Biochem. Biotechnol. 1999, 77, 91–104. [Google Scholar] [CrossRef]
- Pitarelo, A. P.; Fonseca, C. S.; Chiarello, L. M.; Gírio, F. M.; Ramos, L. P. Ethanol Production from Sugarcane Bagasse Using Phosphoric Acid-Catalyzed Steam Explosion. J. Braz. Chem. Soc. 2016, 27, 1889–1898. [Google Scholar] [CrossRef]
- Jurado, M.; Prieto, A.; Martínez-Alcalá, Á.; Martínez, Á. T.; Martínez, M. J. Laccase Detoxification of Steam-Exploded Wheat Straw for Second Generation Bioethanol. Bioresour. Technol. 2009, 100, 6378–6384. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, C.; Lu, M.; Tu, M. Effect of overliming and activated carbon detoxification on inhibitors removal and butanol fermentation of poplar prehydrolysates. Biotechnol. Biofuels 2018, 11, 1–14. [Google Scholar] [CrossRef]
- Abdel-Rahman, M. A.; Tashiro, Y.; Sonomoto, K. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: overview and limits. J. Biotechnol. 2011, 156, 286–301. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Qi, B.; Luo, J.; Shen, F.; Su, Y.; Khan, R.; Wan, Y. Improving Lactic Acid Productivity from Wheat Straw Hydrolysates by Membrane Integrated Repeated Batch Fermentation under Non-Sterilized Conditions. Bioresour. Technol. 2014, 163, 160–166. [Google Scholar] [CrossRef]
- Ye, L.; Hudari, M. S. Bin; Li, Z.; Wu, J. C. Simultaneous Detoxification, Saccharification and Co-Fermentation of Oil Palm Empty Fruit Bunch Hydrolysate for L-Lactic Acid Production by Bacillus coagulans JI12. Biochem. Eng. J. 2014, 83, 16–21. [Google Scholar] [CrossRef]
- Malacara-Becerra, A.; Melchor-Martínez, E. M.; Sosa-Hernández, J. E.; Riquelme-Jiménez, L. M.; Mansouri, S. S.; Iqbal, H. M. N.; Parra-Saldívar, R. Bioconversion of Corn Crop Residues: Lactic Acid Production through Simultaneous Saccharification and Fermentation. Sustainability 2022, 14, 11799. [Google Scholar] [CrossRef]
- Cubas-Cano, E.; Venus, J.; González-Fernández, C.; Tomás-Pejó, E. Assessment of different Bacillus coagulans strains for L-lactic acid production from defined media and gardening hydrolysates: Effect of lignocellulosic inhibitors, J. Biotechnol. 2020, 323, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Murariu, M.; Dubois, P. PLA composites: From production to properties. Adv. Drug Deliv. Rev. 2016, 107, 17–46. [Google Scholar] [CrossRef]
- Lassalle, V.; Ferreira, M.L. PLA nano- and microparticles for drug delivery: An overview of the methods of preparation. Macromol. Biosci. 2007, 7, 767–783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tsapekos, P.; Alvarado-Morales, M.; Zhu, X.; Zervas, A.; Jacobsen, C.S.; Angelidaki, I. Enhanced fermentative lactic acid production from source-sorted organic household waste: Focusing on low-pH microbial adaptation and bio-augmentation strategy. Sci. Total Environ. 2022, 808, 152129. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, H.; Hasunuma, T.; Ogino, C.; Kondo, A. Bioprocessing of bio-based chemicals produced from lignocellulosic feedstocks. Curr. Opin. Biotechnol. 2016, 42, 30–39. [Google Scholar] [CrossRef]
- Carrillo-Nieves, D.; Rostro Alanís, M. J.; de la Cruz Quiroz, R.; Ruiz, H. A.; Iqbal, H. M. N.; Parra-Saldívar, R. Current status and future trends of bioethanol production from agro-industrial wastes in Mexico. Renew. Sustain. Energy Rev. 2019, 102, 63–74. [Google Scholar] [CrossRef]
- Gauss, W. F.; Suzuki, S.; Takagi, M. Manufacture of Alcohol from Cellulosic Materials Using Plural Ferments. U.S. Patent No. 399 0944, 1976. [Google Scholar]
- Michelson, T.; Kask, K.; Jõgi, E.; Talpsep, E.; Suitso, I.; Nurk, A. L. (+)-Lactic acid producer Bacillus coagulans SIM-7 DSM 14043 and its comparison with Lactobacillus delbrueckii ssp. lactis DSM 20073. Enzyme and microbial technology 2006, 39, 861–867. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Hyman, D.; Payne, C.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Wolfe, J. Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples, Laboratory Analytical Procedure (LAP), Technical Report NREL/TP-510-42621. Natl. Renew. Energy Lab. 2008. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Ash in Biomass, Laboratory Analytical Procedure (LAP), Technical Report NREL/TP-510-42622. Natl. Renew. Energy Lab. 2008. [Google Scholar]
- Sluiter, A.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Extractives in Biomass, Laboratory Analytical Procedure (LAP), Technical Report NREL/TP-510-42619. Natl. Renew. Energy Lab. 2008. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass, Laboratory Analytical Procedure (LAP), Technical Report NREL/TP-510-42618. Natl. Renew. Energy Lab. 2008. [Google Scholar]
- Candido, J. P.; Claro, E. M. T.; de Paula, C. B. C.; Shimizu, F. L.; de Oliveria Leite, D. A. N.; Brienzo, M.; de Angelis, D. D. F. Detoxification of sugarcane bagasse hydrolysate with different adsorbents to improve the fermentative process. World J Microbiol Biotechnol. 2020, 36, 43. [Google Scholar] [CrossRef]
- RStudio, T. RStudio: Integrated Development for R. RStudio, Inc. Available from: http://www.rstudio.com/.2015.
- Andrade, L. P.; Crespim, E.; de Oliveira, N.; de Campos, R. C.; Teodoro, J. C.; Galvão, C. M. A.; Maciel Filho, R. Influence of sugarcane bagasse variability on sugar recovery for cellulosic ethanol production. Bioresour. Technol. 2017, 241, 75–81. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H. Carbohydrate elimination of alkaline-extracted lignin liquor by steam explosion and its methylolation for substitution of phenolic adhesive. Ind. Crops Prod. 2014, 53, 93–101. [Google Scholar] [CrossRef]
- Antal Jr, M. J.; Mok, W. S.; Richards, G. N. Mechanism of formation of 5-(hydroxymethyl)-2-furaldehyde from D-fructose and sucrose. Carbohydr Res. 1990, 199, 91–109. [Google Scholar] [CrossRef]
- Brown, D. W.; Floyd, A. J.; Kinsman, R. G.; Roshanhyphen; Ali, Y. Dehydration reactions of fructose in non-aqueous media. J Chem Technol Biotechnol. 1982, 32, 920–924. [Google Scholar] [CrossRef]
- Hofvendahl, K.; Hahn-Hagerdal, B. Factors affecting the fermentative lactic acid production from renewable resources. Enzyme Microbial Technol. 2000, 26, 87–107. [Google Scholar] [CrossRef]
- Ramos, L. P. The chemistry involved in the pretreatment of lignocellulosic materials. Química Nova 2003, 26, 863–871. [Google Scholar] [CrossRef]
- Du, B.; Sharma, L. N.; Becker, C.; Chen, S. F.; Mowery, R. A.; van Walsum, G. P.; Chambliss, C. K. Effect of varying feedstock–pretreatment chemistry combinations on the formation and accumulation of potentially inhibitory degradation products in biomass hydrolysates. Biotechnol. Bioeng. 2010, 107, 430–440. [Google Scholar] [CrossRef]
- Lu, C.; Dong, J.; Yang, S. T. Butanol Production from wood pulping hydrolysate in an integrated fermentation-gas stripping process. Bioresour. Tech. 2013, 143, 467–475. [Google Scholar] [CrossRef]
- Miura, M.; Suzuki, T.; Aoyama, M. Detoxification of Japanese white birch wood hemicellulose hydrolysate with a carbonaceous sorbent prepared from birch wood hydrolysis residue. Cellulose Chem and Tech. 2016, 50, 265–268. [Google Scholar]
- Sukumaran, R.K.; Singhania, R.R.; Pandey, A. Microbial cellulases production, applications and challenges. J. Sci. Ind. Res. 2005, 64, 832–844. [Google Scholar]
- Mathew, G.M.; Sukumaran, R.K.; Singhania, R.R.; Pandey, A.; 2008. Progress in research on fungal cellulases for lignocellulose degradation. J. Sci. Ind. Res. 2008, 67, 898–907. [Google Scholar]
- Kristensen, J.B.; Felby, C.; Jørgensen, H. Yield-determining factors in high-solids enzymatic hydrolysis of lignocellulose. Biotechnol. Biofuels. 2009, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R. A.; Schneider, R.; Rossel, C.E.V.; Maciel Filho, R.; Venus, J. Polymer grade L-lactic acid production from sugarcane bagasse hemicellulosic hydrolysate using Bacillus coagulans. Bioresour. Technol. Reports 2019, 2019, 26–31. [Google Scholar] [CrossRef]
- Ahorsu, R.; Cintorrino, G.; Medina, F.; Constantí, M. Microwave processes: a viable technology for obtaining xylose from walnut shell to produce lactic acid by Bacillus coagulans. J. Cleaner Production 2019, 231, 1171–1181. [Google Scholar] [CrossRef]
- van der Pol, E.; Springer, J.; Vriesendorp, B.; Weusthuis, R.; Eggink, G. Precultivation of Bacillus coagulans DSM2314 in the presence of furfural decreases inhibitory effects of lignocellulosic by-products during l(+)-lactic acid fermentation. Appl. Microbiol. Biotechnol. 2016, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Feron, V. J.; Til, H. P.; De Vrijer, F.; Woutersen, R. A.; Cassee, F. R.; Van Bladeren, P. J. Aldehydes: occurrence, carcinogenic potential, mechanism of action and risk assessment. Mutation Research 1991, 259, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Allen, S. A.; Clark, W.; McCaffery, J.; M. ; Cai, Z.; Lanctot, A.; Slininger, P.J.; Lewis Liu, Z.; Gorsich, S.W. Furfural induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisae. Biotechnol. Biofuels 2010, 2, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar] [PubMed]
- Ye, L.; Hudari, M. S. B.; Zhou, X.; Zhang, D.; Li, Z.; Wu, J. C. Conversion of acid hydrolysate of oil palm empty fruit bunch to L-lactic acid by newly isolated Bacillus coagulans JI12. App. Microbiol. Biotechnol. 2013, 97, 4831–4838. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Mira, N. P.; Jarboe, L. R. Adaptation and tolerance of bacteria against acetic acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef]
- Othman, M.; Ariff, A.B.; Rios-Solis, L.; Halim, M. Extractive fermentation of lactic acid in lactic acid bacteria cultivation: a review. Front. Microbiol. 2017, 8, 2285. [Google Scholar] [CrossRef]
- van der Pol, E.C.; Eggink, G.; Weusthuis, R.A. Production of L(+)-lactic acid from pretreated sugarcane bagasse using Bacillus coagulans DSM2314 in a simultaneous saccharification and fermentation strategy. Biotechnol. Biofuels 2016, 9, 248. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, J.; Han, X.; Gao, C.; Ma, C.; Tao, F.; Xu, P. Kinetic characteristics of long-term repeated fed-batch (LtRFb) l-lactic acid fermentation by a Bacillus coagulans strain. Eng. Life Sci. 2020, 20, 562–570. [Google Scholar] [CrossRef]
Figure 1.
Glucose released (in g·L-1) during the enzymatic hydrolysis of steam-exploded substrates for experiments that were carried out in Infors HT bioreactors.
Figure 1.
Glucose released (in g·L-1) during the enzymatic hydrolysis of steam-exploded substrates for experiments that were carried out in Infors HT bioreactors.
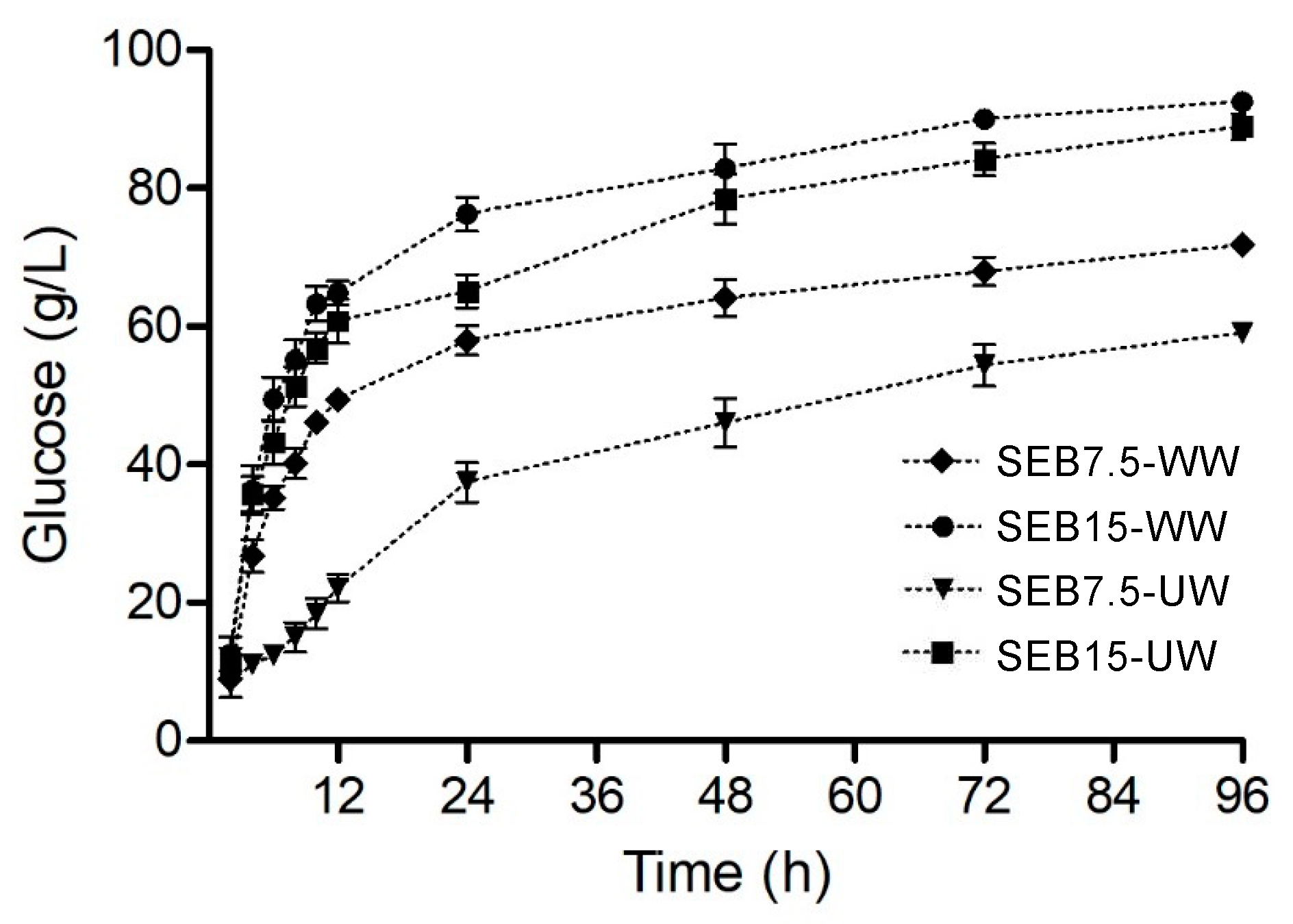

Table 1.
Chemical composition (%) of untreated and water-insoluble steam-exploded SCB, and the corresponding mass recovery (%) of the main SCB components after pretreatment.
Table 1.
Chemical composition (%) of untreated and water-insoluble steam-exploded SCB, and the corresponding mass recovery (%) of the main SCB components after pretreatment.
| Component | Untreated | 195°C, 7.5 min | 195°C, 15 min | |||
| Content | Recovery | Content | Recovery | |||
| Glucans1 | 37.8 ± 0.7 | 54.7 ± 2.4 | 89.7 ± 1.8 | 55.5 ± 0.7 | 82.5 ± 1.1 | |
| Xylans2 | 21.0 ± 0.8 | 3.0 ± 0.4 | 5.9 ± 0.1 | 1.2 ± 0.1 | 3.8 ± 0.1 | |
| Arabinosyl residues2 | 1.3 ± 0.3 | bdl3 | - | - | - | |
| Acetyl groups2 | 2.6 ± 0.1 | bdl3 | - | bdl3 | - | |
| Hexoses identified as 5-HMF4 | 1.1 ± 0.2 | bdl3 | - | bdl3 | - | |
| Pentoses identified as furfural4 | 0.6 ± 0.1 | bdl3 | - | bdl3 | - | |
| Total lignin5 | 22.8 ± 0.7 | 30.5 ± 0.8 | 92.2 ± 1.9 | 31.2 ± 0.2 | 96.6 ± 0.3 | |
| Acid-soluble lignin | 5.0 ± 0.1 | 5.7 ± 0.2 | 70.4 ± 1.5 | 5.9 ± 0.2 | 78.5 ± 0.2 | |
| Acid-insoluble lignin | 19.1 ± 0.6 | 27.8 ± 0.8 | 96.3 ± 0.9 | 28.9 ± 0.1 | 101.4 ± 0.3 | |
| Ash | 4.0 ± 0.1 | 6.1 ± 1.5 | 93.7 ± 1.9 | 6.8 ± 1.1 | 113.8 ± 0.3 | |
| Total | 98.2 | 94.3 | 94.7 | |||
1 Present as β-(1→4)-D-glucans (cellulose). 2 Present as heteroxylan components (hemicelluloses). 3 bdl, below the detection limit of the method. 4 5-HMF and furfural are released in pretreatment liquors by hexose and pentose dehydration, respectively. 5 Summation of acid-soluble and ash-free acid-insoluble lignin.
Table 2.
Chemical composition of pretreatment liquors before and after detoxification.
| Component (g·L-1) | SEB7.51 | SEB151 | |||
| Untreated | Detoxified | Untreated | Detoxified | ||
| Glucose | 2.2 ± 0.1 | 2.2 ± 0.1 | 2.2 ± 0.1 | 2.1 ± 0.1 | |
| Xylose | 11.2 ± 0.2 | 11.1 ± 0.2 | 4.3 ± 0.1 | 4.3 ± 0.2 | |
| Arabinose | 0.6 ± 0.1 | 0.60 ± 0.04 | 0.5 ± 0.1 | 0.4 ± 0.1 | |
| HMF2 | 1.6 ± 0.2 | 0.02 ± 0.02 | 1.6 ± 0.1 | 0.03 ± 0.01 | |
| Furfural2 | 0.8 ± 0.1 | 0.04 ± 0.02 | 1.4 ± 0.3 | 0.03 ± 0.01 | |
| Acetic acid3 | 9.7 ± 0.3 | 4.4 ± 0.5 | 12.1 ± 0.5 | 5.2 ± 0.2 | |
1 SEB7.5 and SEB15, steam explosion of cane bagasse (SEB) at 195 ºC for 7.5 and 15 min; 2 5-HMF and furfural are released in pretreatment liquors by hexose and pentose dehydration; 3 Acetic acid is released in pretreatment liquors by hemicellulose deacetylation.
Table 3.
Mathematical models for glucose release (g/L) from enzymatic hydrolysis experiments that were performed in shake flasks for 96 h.
Table 3.
Mathematical models for glucose release (g/L) from enzymatic hydrolysis experiments that were performed in shake flasks for 96 h.
| Substrate | Equation | R2 |
| SEB7.5-WW | 0.9986 | |
| SEB15-WW | 0.9982 | |
| SEB7.5-UW | 0.9969 | |
| SEB15-UW | 0.9998 |
Table 4.
Analysis of variance (ANOVA) of the mathematical models of Table 3 at a confidence level of 95% (p < 0.05).
Table 4.
Analysis of variance (ANOVA) of the mathematical models of Table 3 at a confidence level of 95% (p < 0.05).
| Substrate | Conditions | Degrees of freedom | RSR* | Adjusted R2 | F-value | p-value |
| SEB-WW | 195°C, 7.5 min | 10 | 0.5219 | 0.9986 | 1702 | 1.459 x 10-14 |
| 195°C, 15 min | 10 | 0.8164 | 0.9969 | 779 | 7.208 x 10-13 | |
| SEB-UW | 195°C, 7.5 min | 10 | 0.9194 | 0.9982 | 1337 | 4.867 x 10-14 |
| 195°C, 15 min | 10 | 0.9317 | 0.9980 | 1207 | 8.156 x 10-14 |
* Residual standard error.
Table 5.
Glucose and xylose release after enzymatic hydrolysis of both SEB-UW and SEB-WW in shake flasks for 96 h using Cellic CTec3 in the absence and presence of 10% Cellic HTec3 (wet basis).
Table 5.
Glucose and xylose release after enzymatic hydrolysis of both SEB-UW and SEB-WW in shake flasks for 96 h using Cellic CTec3 in the absence and presence of 10% Cellic HTec3 (wet basis).
| Parameter | SEB-WW1 | SEB-UW2 | |||
| CTec3 | CTec3/HTec3 | CTec3 | CTec3/HTec3 | ||
| Steam explosion at 195 ºC for 7.5 min | |||||
| Glucose concentration (g·L-1) | 46.17 ± 0.13 | 59.65 ± 0.33 | 45.61 ± 0.30 | 59.50 ± 0.37 | |
| Cellobiose concentration (g·L-1) | 1.50 ± 0.12 | 2.52 ± 0.04 | 1.53 ± 0.12 | 2.25 ± 0.16 | |
| Glucan conversion (%) | 39.26 ± 0.07 | 51.23 ± 0.19 | 38.27 ± 0.17 | 50.15 ± 0.22 | |
| Xylose concentration (g·L-1) | 2.59 ± 0.15 | 3.00 ± 0.09 | 6.24 ± 0.12 | 6.89 ± 0.11 | |
| Xylan conversion | 40.52 ± 0.32 | 46.9 ± 0.19 | 97.5 ± 0.16 | 97.7 ± 0.25 | |
| Steam explosion at 195 ºC for 15 min | |||||
| Glucose concentration (g·L-1) | 78.25 ± 0.28 | 84.52 ± 0.31 | 63.72 ± 0.13 | 75.14 ± 0.11 | |
| Cellobiose concentration (g·L-1) | 3.09 ± 0.12 | 3.40 ± 0.06 | 1.51 ± 0.11 | 2.63 ± 0.12 | |
| Glucan conversion (%) | 67.02 ± 0.16 | 74.44 ± 0.20 | 52.93 ± 0.08 | 63.15 ± 0.10 | |
| Xylose concentration (g·L-1) | 1.78 ± 0.09 | 2.16 ± 0.12 | 2.88 ± 0.07 | 3.30 ± 0.02 | |
| Xylan conversion | 27.8 ± 0.17 | 33.7 ± 0.11 | 45.0 ± 0.31 | 51.6 ± 0.47 | |
1 Water-washed steam-exploded sugarcane bagasse. 2 Unwashed steam-exploded sugarcane bagasse.
Table 6.
Lactic acid fermentation of the pretreatment liquors before and after detoxification with a commercial activated carbon powder.
Table 6.
Lactic acid fermentation of the pretreatment liquors before and after detoxification with a commercial activated carbon powder.
| Parameter | SEB7.5 liquor | SEB15 liquor | |||
| Non-detoxified | Detoxified | Non-detoxified | Detoxified | ||
| Initial Xyl (g·L-1)1 | 11.2 ± 0.2 | 11.1 ± 0.2 | 4.3 ± 0.2 | 4.3 ± 0.1 | |
| Initial Glc (g·L-1)1 | 2.2 ± 0.2 | 2.3 ± 0.1 | 2.2 ± 0.1 | 2.1 ± 0.2 | |
| Initial Ara (g·L-1)1 | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.5 ± 0.2 | 0.4 ± 0.1 | |
| Lactic acid (g·L-1)2 | 2.3 ± 0.6 | 13.4 ± 0.5 | 1.7 ± 0.4 | 6.6 ± 0.6 | |
| Yp/s (g·g-1)3 | 0.17 ± 0.03 | 0.96 ± 0.02 | 0.25 ± 0.10 | 0.97 ± 0.12 | |
| Yx/s (g·g-1)4 | 0.19 ± 0.03 | 0.16 ± 0.04 | 0.09 ± 0.05 | 0.11 ± 0.07 | |
| PLA (g·L-1·h-1)5 | < 0.01 | 0.54 ± 0.02 | < 0.01 | 0.24 ± 0.03 | |
| OD6006 | 1.0 ± 0.3 | 5.9 ± 0.1 | 1.6 ± 0.4 | 3.8 ± 0.6 | |
1 Xyl, Glc, Ara – xylose, glucose, and arabinose. 2 Lactic acid produced by fermentation. 3 Lactic acid yield (gram of product per gram of substrate). 4 Biomass yield (gram of biomass per gram of substrate). 5 Lactic acid productivity (gram of product per liter per hour). 6 Optical densities at 600 nm.
Table 7.
Lactic acid fermentation of enzymatic hydrolysates derived from water-washed (SEB-WW) and unwashed (SEB-UW) pretreatment solids.
Table 7.
Lactic acid fermentation of enzymatic hydrolysates derived from water-washed (SEB-WW) and unwashed (SEB-UW) pretreatment solids.
| Parameter | SEB7.5-WW | SEB7.5-UW | SEB15-WW | SEB15-UW |
| Initial Xyl (g·L-1)1 | 2.1 ± 0.1 | 6.4 ± 0.3 | 0.9 ± 0.7 | 4.9 ± 0.9 |
| Initial Glc (g·L-1)1 | 71.0 ± 0.4 | 59.8 ± 0.3 | 93.0 ± 1.8 | 89.9 ± 1.6 |
| Residual Xyl (g·L-1)1 | 0.2 ± 0.1 | 2.1 ± 0.3 | 0.13 ± 0.07 | 2.4 ± 0.8 |
| Residual Glc (g·L-1)1 | 17.1 ± 1.2 | 14.3 ± 1.6 | 34.6 ± 1.7 | 42.0 ± 1.1 |
| Lactic acid (g·L-1)2 | 64.2 ± 1.3 | 48.8 ± 0.5 | 56.9 ± 1.2 | 52.4 ± 0.8 |
| Yp/s (g·g-1)3 | 0.88 ± 0.01 | 0.74 ± 0.01 | 0.61 ± 0.01 | 0.55 ± 0.02 |
| Yx/s (g·g-1)4 | 0.09 ± 0.02 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.08 ± 0.3 |
| PLA (g·L-1·h-1)5 | 2.68 ± 0.08 | 2.03 ± 0.03 | 2.37 ± 0.07 | 2.19 ± 0.05 |
| OD6006 | 14.6 ± 0.8 | 15.3 ± 0.9 | 14.1 ± 0.3 | 15.9 ± 0.4 |
Table 8.
Lactic acid fermentation of enzymatic hydrolysates derived from water-washed (SEB7.5-WW) pretreatment solids with the addition of non-detoxified and detoxified pretreatment liquors (C5 stream) for a C5/C6 ratio of 1:10.
Table 8.
Lactic acid fermentation of enzymatic hydrolysates derived from water-washed (SEB7.5-WW) pretreatment solids with the addition of non-detoxified and detoxified pretreatment liquors (C5 stream) for a C5/C6 ratio of 1:10.
| Parameter | SEB7.5WW hydrolysates containing: | SEB15WW hydrolysates containing: | |||
| Non-detoxified C5 | Detoxified C5 | Non-detoxified C5 | Detoxified C5 | ||
| Initial Xyl (g·L-1)1 | 6.9 ± 0.7 | 6.4 ± 0.3 | 8.9 ± 0.9 | 5.5 ± 0.6 | |
| Initial Glc (g·L-1)1 | 54.4 ± 0.9 | 59.8 ± 0.3 | 87.2 ± 0.9 | 91.0 ± 1.4 | |
| Residual Xyl (g·L-1)1 | 4.4 ± 0.9 | 1.1 ± 0.5 | 6.2 ± 0.6 | 1.2 ± 0.3 | |
| Residual Glc (g·L-1)1 | 51.9 ± 0.3 | 3.8 ± 0.7 | 85.7 ± 0.4 | 26.4 ± 0.2 | |
| Lactic acid (g·L-1)2 | 4.6 ± 0.3 | 61.4 ± 1.8 | 4.1 ± 0.5 | 76.7 ± 1.4 | |
| Yp/s (g·g-1)3 | 0.07 ± 0.02 | 0.93 ± 0.02 | 0.04 ± 0.02 | 0.78 ± 0.02 | |
| Yx/s (g·g-1)4 | 0.07 ± 0.01 | 0.07 ± 0.02 | 0.07 ± 0.01 | 0.08 ± 0.01 | |
| PLA (g·L-1·h-1)5 | 0.19 ± 0.06 | 2.55 ± 0.11 | 0.17 ± 0.03 | 3.20 ± 0.08 | |
| OD6006 | 3.7 ± 0.2 | 16.2 ± 0.4 | 3.3 ± 0.2 | 16.0 ± 0.7 | |
1-6 See legends in Table 6.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



