
Submitted:
14 July 2023
Posted:
24 July 2023
You are already at the latest version
Abstract
Microalgae are a source of carotenoids, phycocyanin, phenol, amino acids, polyunsaturated fatty acids, sulfated polysaccharides, pigments, and other bioactive molecules with antimicrobial, antioxidant, antiviral, antitumor, anti-inflammatory, and fat-burning properties. The increasing incidence of infectious processes, including tuberculosis, caused by antimicrobial resistant strains of microorganisms necessitates the search for alternative sources of compounds with antimicrobial activity, including microalgae. In this study, the antimicrobial properties of marine and freshwater microalgae extracts were evaluated against Gram-positive (Staphylococcus aureus, Enterococcus faecalis, and Streptococcus pyogenes) and Gram-negative (Klebsiella pneumoniae, Acinetobacter baumannii, and Pseudomonas aeruginosa), Mycobacterium tuberculosis strain H37Rv and Mycobacterium smegmatis. Water, dimethyl sulfoxide, and ethanol were used as extractants. The obtained microalgae extract exhibited cytotoxicity against splenocytes and, to a lesser extent, against peritoneal macrophages of CBA mice. The bacteriostatic effect of the microalgae extracts against bacteria and mycobacteria was demonstrated by MTT test. In addition, activation of bacterial and mycobacterial growth was observed in a number of cases with high levels of microalgae extracts in the medium. In an expression model of tuberculosis in mice, a decrease in mycobacterial load in the lungs and spleen was observed. Thus, microalgae have moderate to weak antimicrobial activity.
Keywords:
diatoms
; cyanobacter
; green microalgae
; Gram-positive and Gram-negative bacteria
; Mycobacteria
; antimicrobial potential
1. Introduction
Microalgae, with their enormous biodiversity and ability to adapt to different habitats, are a powerful source of biologically active compounds such as pigments (chlorophylls, carotenoids and phycobilins), proteins, lipids and polysaccharides. The concentrations of carotenoids and chlorophylls depend on the microalgae species and environmental conditions. The process of photosynthesis of bioactive substances in microalgae can be regulated by the type of illumination of cultures in bioreactors, so that red light stimulates the synthesis of polyphenol and green light stimulates the synthesis of flavanoids [1,2]. The disadvantages of microalgae include their ability to produce toxic substances, which limits the range of microalgae that can be used as a source of organic food, feed, nutraceuticals, and pharmaceuticals. However, only Arthrospira platensis, Chlamydomonas reinhardtii, Auxenochlorella protothecoides, Chlorella vulgaris, Dunaliella salina and Euglena gracilis are considered less dangerous for humans and animals due to the ability of microalgae to synthesize toxins. The antibacterial and antifungal activity of extracts of various microalgae has been demonstrated. For example, the growth of Staphylococcus haemolyticus was inhibited by Arthrospira platensis extract, Bacillus subtilis by Calosedrus rupestris, Furcellaria lumbricalis, Ulva intestinalis and Arthrospira platensis, and Streptococcus mutans was suppressed by Ulva intestinalis [3]. Microalgae lipids, such as α-linolenic acid, sulfoquinovosyldiacylglycerol, monogalactosylmonoacylglycerol, sulfoquinovosylmonoacylglycerol, hexadeca-4,7,10,13-tetraenoic acid, palmitoleic acid and lysophosphatidylcholine exhibit antimicrobial and antifouling properties by microbes [4]. Dichloromethane extract of Scenedesmus obliquus potentiated the antimicrobial activity of penicillin’s, fluoroquinolones against Staphylococcus aureus, Escherichia coli and Salmonella typhimurium in vitro [5]. Fatty acids (oleic, linoleic and linolenic acids) and carotenoids (xanthin, neoxanthin, cryptoxanthin and echinenone) of Chlorella vulgaris [6], Skeletonema tropicum and Chaetoceros Pseudocurvisetus extract [7] and Chaetoceros muelleri extract have antifungal activity [8]. In this study, we set out to investigate the antimicrobial potential of different microalgae taxa against bacteria and mycobacteria.
2. Results
2.1. Biochemical Characteristics of the Microalgae
We determined proteins, lipids and carbohydrates in the dry biomass of different microalgae taxa (Table 1). The minimum protein content was observed in N. shiloi and P. viridis; less than 200 mg/g lipids were determined in R. mixta and P. agardhii; higher content of carbohydrates - in R. mixta, P. agardhii and P. virids. Fx was detected only in N. shiloi. Phycobiliproteins were determined for L. cf. ectocarpi, R. mixta and P. agardhii. Furthermore, the microalgae differ in their PUFA content.
Biochemical compounds in A. plathensis are well studied and include proteins (50-70%), lipids (5%), carbohydrates (10-20%), pigments, amino acid, fatty acid and PUFA [9].
In addition, the chemical nature of the bioactive compounds in microalgae extracts was analyzed by fermentative methods and found that in microalgae extracts present proteins, triglycerides and glucose (Figure 1).
2.2. Cytotoxic Potential of the Microalgae Extracts
The aqueous extracts of the different microalgae taxa were tested by MTT assay on peritoneal macrophages and splenocytes of Balb/c mice at a concentration of 1% v/v. A significant cytotoxic activity (up to 80%) tested microalgae extracts on splenocytes were obtained (Figure 2). Meanwhile, a decrease and an increase in metabolic activity (NADPH-dependent oxidoreductase) were observed in peritoneal macrophages of mice.
2.3. Antibacterial activity of microalgae extracts
In order to identify the antibacterial activity of microalgae, we first tested diatom extracts (A. platensis and N. shiloi) with high Fx content (Table 2). For this purpose, 1% v/v of the test extracts were added to the bacteria in the nutrient medium and growth was evaluated after 14 and 21 days. Fx from A. platensis and N. shiloi decreased the growth of Gram-positive (S. aureus, E. faecalis, S. pyogenes) and Gram-negative (Kl. pneumoniae, A. baumannii, Ps. aeruginosa) bacteria compared to control and ceftazidime on days 14 and 21, especially on day 14. The instability and degradation of Fx may result in a decrease in its antibacterial potential on day 21.
Next, we obtained DMSO (1%) extracts from microalgae and tested their antibacterial potential at 1% v/v, 0.55 v/v, and 0.25% v/v in vitro in comparison with ceftazidime and Fx (Table 3). We did not observe any dose-dependent effect of microalgae extracts on bacterial growth in vitro. While Fx show a dose-dependent inhibition of bacterial growth. Moreover, some microalgae extracts possess to increase bacteria growth compared to control and ceftazidime (p < 0.05).
Blue-green microalgae produced wide spectrum of bioactive molecules, including carotenoids and chlorophylls with antimicrobial activity. We compared microalgae extracts and chlorophyllipt (a mixture of chlorophylls from eucalyptus leaves) on bacterial strains resistant to antimicrobial drugs growth inhibition in vitro (Table 4). We found that Fx effectively inhibited bacterial growth compared to chlorophyllipt and ceftazidime (p < 0.05).
Combined information based on in vitro studies of the effect of microalgae extracts, enriched Fx fraction and comparative analysis with chlorophyllipt reflect the presence of biologically active substances with antibacterial activity in the extracts. Taking into account the fact that microalgae extracts contain a wide range of nutrients (proteins, fats and sugars), during experiments we have shown that this affects the antibacterial potential, contributing not to the death but to the survival of bacteria in the presence of high concentrations of extracts in the culture medium.
2.4. Antimycotic Potential of Microalgae
In order to establish antimycotic activity of microalgae extracts, we used two Mycobacteriaceae - Mycobacterium tuberculosis virulent type strain H37Rv and Mycobacterium smegmatis a non-pathogenic microorganism characterized with rapid growth ability.
First, we compared the antifungal activity of Fx and water extract of A. platensis with rifampicin (Figure 3). Only Fx significantly inhibited the growth of Mycobacterium tuberculosis and Mycobacterium smegmatis (p < 0.05).
Then we compared the antimycotic activity of microalgae DMSO extracts with chlorophyllipt and rifampicin, and that extract from N. shiloi, L. ectocarpi and Fx from N. shiloi inhibit mycobacteria growth more effectively compared with chlorophyllipt and rifampicin (Figure 4; P < 0.05). Some microalgae extracts increased mycobacterial growth and some inhibited it. Such a spread of the effect of microalgae extracts may reflect the resistance of pathogenic strains of mycobacteria to the action of toxic agents, the presence of efflux pumps. Stimulation of the growth of mycobacteria may be a consequence of the presence of nutrients in microalgae extracts.
It was also logical to identify the dose-dependent effect of microalgae extracts and Fx on the survival of mycobacteria in vitro (Table 5).
Microalgae extracts have been shown to either inhibit or stimulate the growth of mycobacteria and there is no clear dose-dependency. In some cases, the minimum or maximum concentration of microalgae extracts stimulated or suppressed the growth of mycobacteria.
2.5. Effect of the Fucoxanthin Administration in Mice with Tuberculosis
We then studied the effect of oral administration of Fx in mice with experimental tuberculosis induced by tail vein injection of Mycobacterium tuberculosis H37Rv. Administration of Fx to reduce the number of CFU of Mycobacterium tuberculosis growth on solid medium from lungs at 46% compared with untreated group and at 8% compared with rifampicin-treated group (Figure 5; p < 0.05). Rifampicin and Fx have no effect on Mycobacterium tuberculosis liver compared with untreated mice (p > 0.05). While Fx administration possess to decrease the number of CFU of Mycobacterium tuberculosis growth on solid medium from spleen at 30% compared with untreated group and at 23% compared with rifampicin-treated group (p < 0.05).
3. Discussion
In this study, we compared biochemical compounds and the antimicrobial potential of microalgae of different taxa (diatoms, cyanobacteria, green microalgae) and habitats (sea, rivers, lakes) against Gram-positive and Gram-negative bacteria (museum strains sensitive and resistant to antibacterial agents of the clinical isolates of bacteria from multidrug-resistant pulmonary tuberculosis patients, and mycobacteria). In addition, we used microalgae (Leptolynbya cf. ectocarpi, Nanofrustulum shiloi, Platymonas viridis, Planktothrix agardhii, Roholtiella mixta sp. nov.), which, except for Arthrospira platensis, have not been studied for their antimicrobial potential. Thus, secondary metabolites of microalgae Leptolyhbya cf. ectocarpi, such as proteins, carotenoids, phenols and chlorophyll, have the ability to reduce the harmful effect of free radicals, inhibit the activity of matrix metalloproteases (elastase and hyaluronidase) and tyrosinase, increase melatonin production [10]. There is a report on manifestation of antibacterial activity of extract Leptolyngbya sp. extract against MSSA and MRSA only after prior laser treatment of the extract [11,12]. The microalgae Planktothrix agardhii has been shown to have estrogenic activity [13]. The microalgae Roholtiella mixta sp. nov. was only characterized in 2021 and was shown to contain secondary metabolites (β-carotene, zeaxanthin, 3-hydro β-carotene, 3-hydroxyechinenone (4-keto-3-hydroxy-β-carotene), echinenone, caloxanthin, β-cryptoxanthinoleate, and antheraxanthin) [14]. We used unfractionated microalgae extract, except for the enriched fucoxanthin fraction from Arthrospira platensis and Nanofrustulum shiloi obtained by extraction in ethanol.
Microalgae are a valuable source of nutrients, including proteins, fats and sugars. The process of photosynthesis of bioactive substances in microalgae can be regulated by the type of illumination of cultures in bioreactors, so that red light stimulates the synthesis of polyphenol and green light - flavanoids, the speed of aeration, the composition of nutrient media [1,15]. Considering the fact that the incidence of microorganisms resistant to most of the used antimicrobial agents is increasing, it is necessary to search for new sources of biologically active molecules with antimicrobial activity. Microalgae, with their ability to resist and protect against pathogens, are a valuable source of such molecules [16].
Antibacterial compounds produced by cyanobacteria include alkaloids, fatty acids, indoles, macrolides, peptides, phenols, pigments, and terpens [17,18]. The antimicrobial potential of macro- and microalgae depends on the type of algae, extractor and concentration of the extract [19]. Different solvents such as hexane, chloroform, methylene chloride, ethyl acetate, methanol, acetone and water are used as extractants of bioactive molecules from microalgae and showed that hexane extract of Chaetoceros muelleri had the greatest antimicrobial activity including Mycobacterium tuberculosis [8]. The methanol extract of Chlorella sp. UKM8 showed high inhibitory activity ranging from 0.312-6.25 mg/ml against gram positive and gram negative bacteria. The extract contained phenol, hexadecanoic acid, phytol, 9,12-octadecadienoic acid and bicyclo[3.1.1]heptanes, which have antimicrobial potential [20]. The authors analyzed the antimicrobial activity of Scenedesmus subspicatus extracts based on ethanol, methanol, butanol, acetone, dimethyl sulfoxide, and water (solvents of different polarity) [21]. Water and DMSO based extracts have been shown to have better antioxidant and antimicrobial activity. The antibacterial activity of Tetraselmis suecica acid extracts has been observed [16]. The study of antimicrobial potential of microalgae extracts can be conveniently performed using the micro method, as it is easily reproducible and convenient for comparing the study results of different authors [22].
The authors showed the presence of antibacterial activity of extracts of Isochrysis galbana, Scenedesmus sp. NT8c and Chlorella sp. FN1 against gram-positive bacteria in the minimum inhibitory concentration test [23]. The antimicrobial potential of microalgae was shown to be related to the presence of linoleic, oleic, docosahexaenoic and eicosapentaenoic acids in the extracts. A moderate antimicrobial potential of Ettlia pseudoalveolaris extract was shown due to the presence of polyphenol [24]. Nanopeptides (cyclic peptides) possess the main antibacterial activity of the microalgae Oscillatoriales and Nostocales [18]. The authors studied the antimicrobial potential of the extract of Amphidinium carterae against Gram-positive bacteria (Staphylococcus aureus MRSA and MSSA), Gram-negative bacteria (Escherichia coli and Klebsiella pneumoniae), Mycobacterium tuberculosis and Aspergillus fumigatus and showed moderate activity [25]. Amphidinolides from Amphidinium carterae LACW11 show antimicrobial potential against Staphylococcus aureus and Enterococcus faecalis [26]. Lipid complexes from the microalgae Chlorella vulgaris and Arthrospira platensis exhibit antibacterial activity and inhibit the growth of the Gram-positive strain Bacillus subtilis [27,28].
The existence of antifungal potential of microalgae extracts is reported in [6]. It was shown that multidrug-resistant Mycobacterium tuberculosis was sensitive to complexes of Chlorella vulgaris consisting of a combination of unsaturated fatty acids (oleic, linoleic and linolenic acids) and carotenoids (xanthin, neoxanthin, cryptoxanthin and echinenone). Antifungal potential of extracts of diatoms Skeletonema tropicum and Chaetoceros pseudocurvisetus has been noted [7]. The anti-tuberculosis effect of fucoxanthin is based on the inhibition of the enzymes involved in the biosynthesis of the mycobacterial cell wall - UDP-galactopyranose mutase (UGM) and arylamino-N-acetyltransferase [29].
However, not everything is so clear regarding the effectiveness of microalgae protection against bacteria. It is well known that microalgae and bacteria are symbiotic and can mutually influence each other's functional properties [30,31,32,33].
Data present here showed non-fractioned extracts and fucoxanthin from different microalgae taxa were effective against bacteria (Gram-positive and Gram-negative) and mycobacteria (Mycobacterium tuberculosis strain H37Rv and Mycobacterium smegmatis). However, it must be emphasized that some microalgae potentiated bacteria growth.
Limitations of the article include: the use of unrefined fractions of microalgae extracts. These studies are planned in the future, which will identify the most important fractions of microalgae extracts with antimicrobial activity.
4. Materials and Methods
4.1. Microalgae
The objects of the study were microalgae of various systematic groups obtained from the Collective Use Center "Collection of Hydrobionts of the World Ocean" Kovalevsky Research Institute of Biology of the Southern Seas RAS. With the exception of the cyanobacterium Roholtiella mixta (gift of A.A. Goncharov, Vladivostok). Marine cyanobacteria (L. ectocarpi, Leptolyngbya cf. ectocarpi), diatoms (N. shiloi, Nanofrustulum shiloi) and green microalgae (P. viridis, Tetraselmis (Platymonas) viridis Rouchijajnen), freshwater cyanobacteria (P. agardhii, Planktothrix agardhii and; A. platensis, Artrospira platensis) and soil cyanobacteria (R. mixta, Roholtiella mixta sp. nov.) were included in the study. In the first stage of the study, the cultures were adapted to the experimental conditions. The inoculum was grown in 1 L flasks in the accumulation mode. Cultures were grown on mineral nutrient media prepared on sterile seawater and distilled water.
An intensive culture of the diatom alga N. shiloi was obtained on nutrient medium with the following composition (g/L): Na2SiO3 × 9 H20 - 0.150, NaNO3 - 0.3875, NaH2PO4 × 2 H2O - 0.045, Na2EDTA - 0.017, FeSO4 × 7 H2O - 0.030, NaMoO4 × H2O - 1.2∙10-5; CuSO4 × 5 H2O - 2.8∙10-5, ZnSO4 × 7 H2O - 4.4∙10-5, CoCl2 × 6 H2O - 2∙10-5, MnCl2 × 4 H2O - 3.6∙10-5.
For the cultivation of cyanobacteria L. ectocarpi and R. mixta we used the nutrient medium BG11 in our own modification, g/L: NaNO3, 3.75; K2HPO4-3 H2O, 0.065; H3BO3, 2.86×10-3; C₆H₈O₇, 0.006; FeC₆H₅O₇ × 3 H2O, 0.006; Na2EDTA, 0.001; Na2CO3, 0.02; MnCl2 × 4 H2O -1.81×10-5; ZnSO4 × 7 H2O - 0.22×10-5; Na2MoO4 × 2 H2O - 0.4×10-5; CuSO4 × 5 H2O - 0.08×10-5; Co(NO3)2 × 7 H2O - 0.05×10-5.
Green microalgae P. viridis were grown on Trenkenshu nutrient medium with the following composition, g/L: 1.8 - NaNO3; 0.3 - NaH2PO4 × 2 H2O; 0.37 - Na2EDTA; 0.042 - FeC6H5O7 × 3 H2O; 0.008 - MnCl2 × 4H2O; 0.00625 - Co(NO3)2 × 6H2O; 0.00183 - (NH4)6Mo7O24 × 4H2O; 0.00238 - K2Cr2SO4 × 24H2O; 0.00058 - TiO2.
P. agardhii and A. platensis was grown on standard Zarruk nutrient medium, g/L: 16.8 - NaHCO3; 0.5 - K2HPO4; 2.5 - NaNO3; 1.0 - K2SO4; 1.0 - NaCl; 0.08 - Na2EDTA; 0.01 - FeSO4 × 7 H2O; 0.04 - CaCl2 × 2 H2O; 0.2 - MgSO4 × 7 H2O and 1 ml/L of micronutrient mixture (2.86 - H3BO3; 1.81 - MnCl2 × 4 H2O; 0.222 - ZnSO4 × 7 H2O; 0.079 - CuSO4 × 5 H2O; 0.015 - MoO3; 0.02296 - NH4VO3; 0.04398 - Co(NO3)2 × 6 H2O; 0.096 - K2Cr2(SO4)4 × 24 H2O; 0.04785 - NiSO4 × 7 H2O; 0.01794 -Na2WO4 × 2 H2O and 0.04 - Ti2(SO4)3).
As the density of the culture increased, fresh nutrient medium was added until the volume reached 1 L of each species. After adaptation, the entire volume of each culture was used as inoculum for cultivation in a 10-L flat-parallel photobioreactor. Accumulative cultivation with active cross-linking by air barbotage (1.1 L of air per 1 L of culture per minute) under 24-hour illumination with white fluorescent lamps CЄPIL1LF36W/54-765 was also used to increase the 10-L suspension. The average irradiance of the photobioreactor working area was 150 micromole quanta m-2-s-1 (33 W m-2). Green microalgae and cyanobacteria were cultivated at 28±2°C, diatoms at 20±2°C.
The change of the culture density in the photobioreactor was measured by two methods: measuring the optical density of the suspension in a 0.5 cm cuvette at a wavelength of 750 nm and weighing the crude residue (algal biomass) on analytical balances after centrifugation of the suspension in polypropylene tubes at 1600 g for 15 min [34].
4.2. Chemical Analysis
Lipids were extracted from 10 g of wet microalgae biomass with a clorophorm-ethanol mixture (2:1) until the biomass was completely discolored. the extracts was washed 3-4 times with water to remove non-lipid contaminants. The content of totsl lipids in the cloroform fraction was determined by the gravimetric methods [35]. To determine the fatty acid (FAs) composition in the total lipid extracts and lipid fractions, FAs were hydrolyzed and methylated. The chloroforml fraction was evaporated on a rotary evaporator, and 5 mL of freshly prepared alkali solution in methanol (10 mL 3 N NaOH and 90 mL 90% methanol) was added to the residue. The resulting solution was boiled for 1.5 hours under reflux until complete saponification. The solution wasthen supplemented with several drops of 1% alcoholic solution of phenolphthalein, decontaminated with three 5 mL portions of hexane,neutralized with several drops of 0.1 N HCl, and subjected to reextraction with 5 mL hexane portions in 2-3 batches. The hexane fractions were collected and combined. The hexane solution was evaporated to dryness on a rotary evaporator at a temperature of 30oC. To the residue was added 5 mL 3% solution of hydrogen chloride in methanol (for the FAs esterification). The resulting solution was boiled for 2 hours under reflux, and after cooling, it was subjected to hexane extraction (3 × 5 mL). The hexane layers were combined. Before the determination of fatty acid methyl esters (FAME), the hexane fraction was stored for no longer than one day at a temperature of − 20◦C. The resulting FAs methyl esters (FAMEs) were analyzed using gas chromatography on an instrument Chromatec Crystal 5000.2 equipped with a MS detector and a capillary BPX5 column (60 m × 0.25 mm ×0.25 μm) (SGE Analytical Science, UK). The instrument operated also with a split/splitless injector and a flame-ionization detector, both at 280◦C. Data were analyzed using the NIST 14 library. The Supelco FAME 10 mix 37 (CRM47885, Supelco Inc., USA) certified standard was used to identify FAMEs. The fatty acid contents were estimated in mg per g dry weight.
The content of proteins in microalgae biomass were determined by thr Lowry methods [36]. The presence of carbohydrates in microalgae biomass was done by Dubois methods [37].
The Fx concentration in the microalgae biomass were determined according [38]. The extinction coefficient was taken equal tp 1280 mL/(g*cm) at wave lengtyh 488 nm [39]. The C-phycocyanin and C-phycoerithrin contents were determined using the equations given by Bennet and Bogorad [40].
In DMSO-extracts of microalgae were determined proteins (photometric method with bromocresol green), triglycerides (enzymatic colorometric method GPO-PAP) and glucose (glucose oxidase method GOD-PAP) levels using Vector-Best kit regents (Ruddia).
4.3. Fucoxanthin Extraction
To obtain the crude mass of N. shiloi, the suspension was centrifuged at 1600 g for 15 min. The supernatant was removed. Extraction of fucoxanthin (Fx) was performed from 100 g of crude N. shiloi biomass using a minimum volume of 96% ethanol (200 mL), a 2:1 alcohol:biomass ratio at 38 - 40oC. Under these conditions, an almost complete (90 - 95%) extraction of Fx from the biomass was observed for 2 hours. This resulted in an alcoholic extract with a final concentration of 0.35 mg/mL Fx.
To obtain Fx-enriched oil, the alcohol extract with high Fx concentration was evaporated on a rotary evaporator at 1 kPa and 40°C to a minimum volume (5-7) mL and 200 mL olive oil was added. This mixture was then further evaporated until the final removal of ethanol from the olive oil. Thus, a solution of Fx in oil with a concentration of 0.5 mg/mL was obtained.
4.4. Microalgae extracts
The dry mass of different microalgae taxa was obtained by drying a 3-5 mm layer of biomass on polyethylene placed on a flat surface in a warm air curent (38oC) to a residual humidity of 9-10%. The dry mass of microalgae was stored in a tightly sealed container at - 18oC. Extraction of bioactive compounds from microalgae was carried out from 1g of biomass by a minimum volume of distilled water or 1% DMSO solution (10 mL) at 37oC for 24 hours, then filtering and precipitation by centrifugation at 14000 g, then supernatants were use for in vitro study.
4.5. Cytotoxicity Assay
All experimental procedures were performed in accordance with national and EU guidelines for animal experimentation and approved by the local institutional ethics committee (protocol No. 56).
The MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl trazolium bromide) assay was used to assess the viability of mouse Balb/c peritoneal macrophages and splenocytes. Macrophages were obtained from peritoneal lavage and splenocytes were obtained by spleen homogenization. 105/well of peritoneal macrophages or splenocytes in RPMI 1640 medium supplemented with 10% FCS, 2 mM L-glutamine, 100 U/mL antibiotic/antimycotic in control wells and with 1% v/v of various microalgae extracts in experimental wells were incubated for 24 hours in a humidified state with 5% CO2 at 37oC.
4.6. Antimicrobial Assay
A panel of different bacteria (Kl. pneumoniae, Klebsiella pneumoniae susceptible and Klebsiella pneumoniae resistant (ESBL); S. aureus, Staphylococcus aureus MSSA and MRSA, S. aureus clinical isolate from tuberculosis patient; A. baumannii, Acinetobacter baumannii resistant clinical isolate; E. faecalis, Enterococcus faecalis (VRE) clinical isolate; Ps. aeruginosa, Pseudomonas aeruginosa sensitive and resistant clinical isolate; S. pyogenes, Streptococcus pyogenes resistant clinical isolate) and different mycobacteria (Mycobacterium tuberculosis strain H37Rv and Mycobacterium smegmatis) were used for the MIC (minimum inhibitory concentration) assay. The antimicrobial activity was evaluated after 24 hours, 14 and 21 days of treatment with extracts from different microalgae taxa at the concentration of 1, 0.5 and 0.25% v/v and 35, 17.5 and 8.75 µg/mL Fx. Culture media RPMI 1640 with 10% FCS, ceftazidime (50 µg/mL) and rifampicin (100 µg/mL) were used as control samples. The MIC of microalgae extracts was performed in sterile 96- or 48-well plastic plates. Briefly, an inoculum of microbes (approximately 1.5x107 CFU/mL) in 90 µL and different concentrations of microalgae extracts (1% v/, 0.5% v/v and 0.25% v/v) in 10 µL were added to the wells for 96-well plates, and inoculum of microbes in 0.9 mL and microalgae extracts in 100 µL for 48-well plates, incubated for 24 hours in a humidified state with 5% CO2 at 37oC. Each extract was treated in triplicate. On the next day, day 14, and day 21, 10 µL of MTT was added to all wells and DMSO was added 4 hours later to dissolve the formazan, then the absorbance was detected at 570 nm.
4.7. Mice Model of Tuberculosis
The 15 of 8-12 weeks old Balb/c mice were infected intravenously with 5x106CFU Mycobacterium tuberculosis strain H37Rv. On day 30, the mice were divided into 3 groups of 5 mice each: TB-untreated, Mycobacterium tuberculosis strain H37Rv-injected mice without any treatment (control); Rif, mice treated with rifampicin 0.17 g/mice day in 0.5 mL of 0.9% NaCl solution per os during 5 days, once/day; Fx, 87.5 mg/mL day of fucoxanthin alcohol extract in 0.5 mL were administered per os during 5 days, once/day. On day 8 after treatment initiation, animals were sacrificed by cervical dislocation, and lungs, lien, and spleen were harvested. Samples were homogenized, treated with BD BBLTM MycoPrepTM Reagent (BD, USA), washed with phosphate buffer saline, and plated on Lowenstein-Jensen solid medium. CFU counts were calculated on day 21.
4.8. Statistical Analysis
Data were analyzed using Statistica 10.0 for Windows. In this study, the normality of the distribution was determined by the w-Shapiro-Wilkes criterion, the obtain data were presented in tables as mean ± standard deviation (SD), the data were analyzed by one-way analysis of variance (ANOVA) with a Bonferroni correction (Bonferroni post hoc test) to analyze differences between groups. P-values less than 0.05 were considered statistically significant.
5. Conclusions
In this study, microalgae from marine and freshwater reservoirs belonging to different taxa (diatoms, cyanobacteria and green microalgae) were used. A microalga extracts show toxicity against mouse splenocytes, antimicrobial activity against bacteria and mycobacteria, and the presence of symbiosis between microalgae and bacteria, which can be used to develop new antimicrobial compounds.
Author Contributions
Conceptualisation, A.L. and R.G.; methodology, A.L., R.G., S.Zh. and L.R.; investigation, A.L., R.G., S.Zh. and A.S.; data curation, A.L., A.S., R.G , S .Zh, M.S and O.P; writing-review and editing, A.L. and R.G. All authors gave read and agreed to the published version of the manuscript.
Funding
The work was performed within the framework of the state assignment registration number No. 1023022100045-0, No. 122022800132-1 and No. 121030300149-0.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Novosibirsk Tuberculosis Research Institute MH RF (protocol code 56 and date of approval May 25, 2023).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nieri, P.; Carpi, S.; Esposito, R.; Costantini, M.; Zupo, V. Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry. Nutrients 2023, 15, 464. [Google Scholar] [CrossRef]
- Grubišić, M.; Šantek, B.; Zorić, Z.; Čošić, Z.; Vrana, I.; Gašparović, B.; Čož-Rakovac, R.; Šantek, M.I. Bioprospecting of Microalgae Isolated from the Adriatic Sea: Characterization of Biomass, Pigment, Lipid and Fatty Acid Composition, and Antioxidant and Antimicrobial Activity. Molecules 2022, 27, 1248. [Google Scholar] [CrossRef] [PubMed]
- Tolpeznikaite, E.; Bartkevics, V.; Ruzauskas, M.; Pilkaityte, R.; Viskelis, P.; Urbonaviciene, D.; Zavistanaviciute, P.; Zokaityte, E.; Ruibys, R.; Bartkiene, E. Characterization of Macro- and Microalgae Extracts Bioactive Compounds and Micro- and Macroelements Transition from Algae to Extract. Foods 2021, 10, 2226. [Google Scholar] [CrossRef] [PubMed]
- Cepas, V.; Gutiérrez-Del-Río, I.; López, Y.; Redondo-Blanco, S.; Gabasa, Y.; Iglesias, M.J.; Soengas, R.; Fernández-Lorenzo, A.; López-Ibáñez, S.; Villar, C.J.; et al. Microalgae and Cyanobacteria Strains as Producers of Lipids with Antibacterial and Antibiofilm Activity. Mar. Drugs 2021, 19, 675. [Google Scholar] [CrossRef]
- Zaharieva, M.M.; Zheleva-Dimitrova, D.; Rusinova-Videva, S.; Ilieva, Y.; Brachkova, A.; Balabanova, V.; Gevrenova, R.; Kim, T.C.; Kaleva, M.; Georgieva, A.; et al. Antimicrobial and Antioxidant Potential of Scenedesmus obliquus Microalgae in the Context of Integral Biorefinery Concept. Molecules 2022, 27, 519. [Google Scholar] [CrossRef]
- Kumar, T.; Josephine, A.; Sreelatha, T.; Dusthackeer, V.A.; Mahizhaveni, B.; Dharani, G.; Kirubagaran, R.; Kumar, S.R. Fatty acids-carotenoid complex: An effective anti-TB agent from the chlorella growth factor-extracted spent biomass of Chlorella vulgaris. J. Ethnopharmacol. 2019, 249, 112392. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; de la Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First identification of marine diatoms with anti-tuberculosis activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef]
- Ramos, D.F.; Halicki, P.C.B.; Caprara, C.d.S.C.; Borges, P.; M.D'Oca, C.d.R.; Santos, M.d.F.C.; D'Oca, M.G.M.; Roselet, F.; da Silva, P.E.A.; Abreu, P.C. Chemical Profile and Antimicrobial Activity of the Marine Diatom Chaetoceros muelleri. Chem. Biodivers. 2022, 19, e202100846. [CrossRef]
- Viazau, Y.V.; Manankina, E.E.; Filipchik, E.A.; Goncharik, R.G.; Shalygo, N.V. Effectivness of repeated usage of the modified Zarrouk culture medium for cultivation of Spirulina platensis. Proc. Nat. Acad. Sci. Belarus. Biological series, 2018, 63(4), 426-436. [CrossRef]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria Secondary Metabolites as Biotechnological Ingredients in Natural Anti-Aging Cosmetics: Potential to Overcome Hyperpigmentation, Loss of Skin Density and UV Radiation-Deleterious Effects. Mar. Drugs 2022, 20, 183. [Google Scholar] [CrossRef]
- Vahdati, S.N.; Behboudi, H.; Tavakoli, S.; Aminian, F.; Ranjbar, R. Antimicrobial Potential of the Green Microalgae Isolated from the Persian Gulf. Iran. J. Public Heal. 2022, 51, 1134–1142. [Google Scholar] [CrossRef]
- Al Naim, H.M.; El Semary, N. Laser Treatment Increases the Antimicrobial Efficacy of Cyanobacterial Extracts against Staphylococcusaureus (SA) and Methicillin-resistantStaphylococcus aureus (MRSA). Int. J. Environ. Res. Public Heal. 2022, 19, 13305. [Google Scholar] [CrossRef] [PubMed]
- Štěpánková, T.; Ambrožová, L.; Bláha, L.; Giesy, J.; Hilscherová, K. In vitro modulation of intracellular receptor signaling and cytotoxicity induced by extracts of cyanobacteria, complex water blooms and their fractions. Aquat. Toxicol. 2011, 105, 497–507. [Google Scholar] [CrossRef]
- Abdullin, S.R.; Nikulin, V.Y.; Nikulin, A.Y.; Manyakhin, A.Y.; Bagmet, V.B.; Suprun, A.R.; Gontcharov, A.A. Roholtiella mixta sp. nov. (Nostocales, Cyanobacteria): morphology, molecular phylogeny, and carotenoid content. Phycologia 2021, 60, 73–82. [Google Scholar] [CrossRef]
- Erdoğan, A.; Karataş, A.B.; Demir, D.; Demirel, Z.; Aktürk, M.; Çopur. ; Conk-Dalay, M. Manipulation in Culture Conditions of Nanofrustulum shiloi for Enhanced Fucoxanthin Production and Isolation by Preparative Chromatography. Molecules 2023, 28, 1988. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, F.; Wong, G.; Román, T.; Cárdenas, C.; Alvárez, C.; Schmitt, P.; Albericio, F.; Rojas, V. Identification of Antimicrobial Peptides from the Microalgae Tetraselmis suecica (Kylin) Butcher and Bactericidal Activity Improvement. Mar. Drugs 2019, 17, 453. [Google Scholar] [CrossRef]
- Han, S.; Zhao, J.; Liu, P.; Wang, K.; Qin, S.; Zhao, Z.; Cui, Y. Two Foreign Antimicrobial Peptides Expressed in the Chloroplast of Porphyridium purpureum Possessed Antibacterial Properties. Mar. Drugs 2022, 20, 484. [Google Scholar] [CrossRef] [PubMed]
- Rojas, V.; Rivas, L.; Cárdenas, C.; Guzmán, F. Cyanobacteria and Eukaryotic Microalgae as Emerging Sources of Antibacterial Peptides. Molecules 2020, 25, 5804. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef]
- Shaima, A.F.; Yasin, N.H.M.; Ibrahim, N.; Takriff, M.S.; Gunasekaran, D.; Ismaeel, M.Y. Unveiling antimicrobial activity of microalgae Chlorella sorokiniana (UKM2), Chlorella sp. (UKM8) and Scenedesmus sp. (UKM9). Saudi J. Biol. Sci. 2022, 29, 1043–1052. [Google Scholar] [CrossRef]
- de Macedo, D.M.; de Barbosa, C.Y.; Pedrosa Brandão, R.M.; das Graças, M.; Gálvez, A.O.; Souza Bezerra, R. Evaluation of antioxidant and antibacterial capacity of green microalgae Scenedesmus subspicatus. Food Sci. Technol. Int. 2019, 25, 318–326. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Bioprospecting for bioactive compounds in microalgae: Antimicrobial compounds. Biotechnol. Adv. 2022, 59, 107977. [Google Scholar] [CrossRef]
- Alsenani, F.; Tupally, K.R.; Chua, E.T.; Eltanahy, E.; Alsufyani, H.; Parekh, H.S.; Schenk, P.M. Evaluation of microalgae and cyanobacteria as potential sources of antimicrobial compounds. Saudi Pharm. J. 2020, 28, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Vornoli, A.; Grande, T.; Lubrano, V.; Vizzarri, F.; Gorelli, C.; Raffaelli, A.; Della Croce, C.M.; Baca, S.Z.; Sandoval, C.; Longo, V.; et al. In Vitro Characterization of Antioxidant, Antibacterial and Antimutagenic Activities of the Green Microalga Ettlia pseudoalveolaris. Antioxidants 2023, 12, 1308. [Google Scholar] [CrossRef] [PubMed]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a New Cytotoxic and Antifungal Amphidinol from the Dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.E.; Murphy, E.; Parkes, R.; Fleming, G.T.A.; Campanile, F.; Thomas, O.P.; Touzet, N. Antibacterial Activity and Amphidinol Profiling of the Marine Dinoflagellate Amphidinium carterae (Subclade III). Int. J. Mol. Sci. 2021, 22, 12196. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, S.; Prosekov, A.; Ivanova, S.; Maslennikov, P.; Andreeva, A.; Budenkova, E.; Kashirskikh, E.; Tcibulnikova, A.; Zemliakova, E.; Samusev, I.; et al. Identification of Metabolites with Antibacterial Activities by Analyzing the FTIR Spectra of Microalgae. Life 2022, 12, 1395. [Google Scholar] [CrossRef]
- Dolganyuk, V.; Andreeva, A.; Sukhikh, S.; Kashirskikh, E.; Prosekov, A.; Ivanova, S.; Michaud, P.; Babich, O. Study of the Physicochemical and Biological Properties of the Lipid Complex of Marine Microalgae Isolated from the Coastal Areas of the Eastern Water Area of the Baltic Sea. Molecules 2022, 27, 5871. [Google Scholar] [CrossRef]
- Šudomová, M.; Shariati, M.A.; Echeverría, J.; Berindan-Neagoe, I.; Nabavi, S.M.; Hassan, S.T.S. A Microbiological, Toxicological, and Biochemical Study of the Effects of Fucoxanthin, a Marine Carotenoid, on Mycobacterium tuberculosis and the Enzymes Implicated in Its Cell Wall: A Link Between Mycobacterial Infection and Autoimmune Diseases. Mar. Drugs 2019, 17, 641. [Google Scholar] [CrossRef]
- Liu, B.; Eltanahy, E.E.; Liu, H.; Chua, E.T.; Thomas-Hall, S.R.; Wass, T.J.; Pan, K.; Schenk, P.M. Growth-promoting bacteria double eicosapentaenoic acid yield in microalgae. Bioresour. Technol. 2020, 316, 123916. [Google Scholar] [CrossRef]
- Dao, G.; Wang, S.; Wang, X.; Chen, Z.; Wu, Y.; Wu, G.; Lu, Y.; Liu, S.; Hu, H. Enhanced Scenedesmus sp. growth in response to gibberellin secretion by symbiotic bacteria. Sci. Total. Environ. 2020, 740, 140099. [Google Scholar] [CrossRef] [PubMed]
- Iida, H.; Aburai, N.; Fujii, K. Microalga–bacteria Community with High Level Carbon Dioxide Acclimation and Nitrogen-fixing Ability. Protist 2023, 174. [Google Scholar] [CrossRef]
- Astafyeva, Y.; Gurschke, M.; Streit, W.R.; Krohn, I. Interplay between the microalgae Micrasterias radians and its symbiont Dyadobacter sp. HH091. Front. Microbiol. 2022, 13, 1006609. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution à l'étude d'une cyanophycée. Influence de divers facteurs physiques et chimiques sur la croissance et la photosynthèse de Spirulina maxima (Setch et Gardner) Geitler: Ph.D thesis. - Paris, 1966. - 114 p. - (A la faculté des sciences de l'université de Paris).
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. j. biochem. Physiol., 1959, 37(8), 911-917.
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Ryabushko, V.I.; Zheleznova, S.N.; Nekhoroshev, M.V. Effect of Nitrogen on Fucoxanthin Accumulation in the Diatom Cylindrotheca closterium (Ehrenb.) Reimann et Lewin. Int. J. Algae 2017, 19, 79–84. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ozaki, Y.; Taminato, M.; Das, S.K.; Mizuno, M.; Yoshimura, K.; Maoka, T.; Kanazawa, K. The distribution and accumulation of fucoxanthin and its metabolites after oral administration in mice. Br. J. Nutr. 2009, 102, 242–248. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
Figure 1.
Content of main biochemical compounds in DMSO-extracts of the different microalgae taxa.
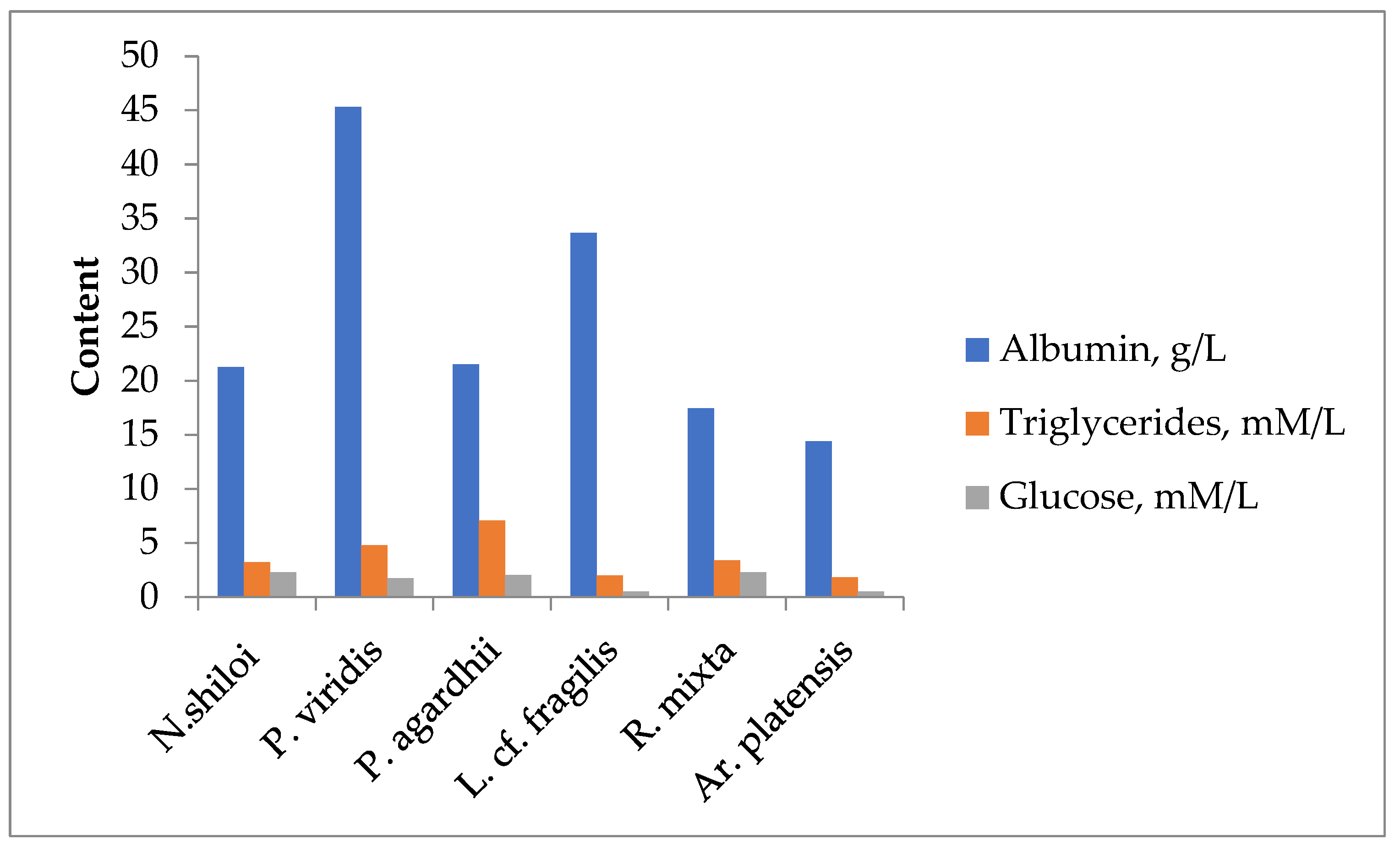
Figure 2.
Cells viability by MTT assay of the different microalgae water-extracts on Balb/c mice peritoneal macrophages and splenocytes after 24 hours exposure. Statistical notations: * = p < 0.05.
Figure 2.
Cells viability by MTT assay of the different microalgae water-extracts on Balb/c mice peritoneal macrophages and splenocytes after 24 hours exposure. Statistical notations: * = p < 0.05.

Figure 3.
Antimycotic potential of microalgae water extracts. Statistical notations: * = p < 0.05 compared with basal mycobacteria growth; # = p < 0.05 with Rifampicin. Rifampicin was added in wells in dose 100 µg/mL, A. platensis in dose 1% v/v and Fx in dose 35 µg/mL.
Figure 3.
Antimycotic potential of microalgae water extracts. Statistical notations: * = p < 0.05 compared with basal mycobacteria growth; # = p < 0.05 with Rifampicin. Rifampicin was added in wells in dose 100 µg/mL, A. platensis in dose 1% v/v and Fx in dose 35 µg/mL.
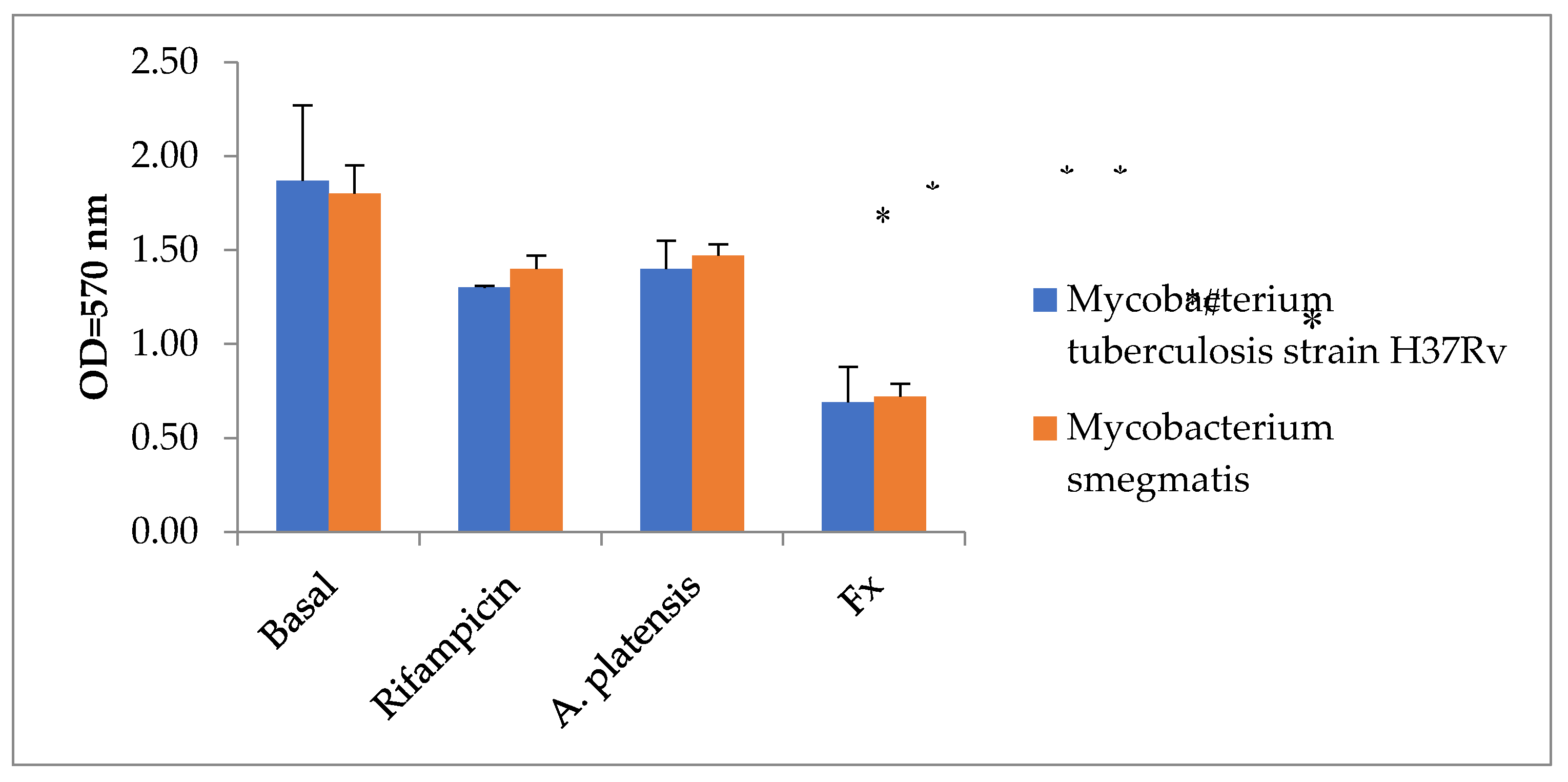
Figure 4.
Comparison on antimycotic activity of the Chlorophyllipt and microalgae DMSO-extracts (M±SD). Statistical notations: * = p < 0.05 compared with basal mycobacteria growth; # = p < 0.05 with Rifampicin. . Rifampicin was added in wells in dose 100 µg/mL, Chlorophyllipt and microalgae extract in dose 1% v/v and Fx in dose 35 µg/mL.
Figure 4.
Comparison on antimycotic activity of the Chlorophyllipt and microalgae DMSO-extracts (M±SD). Statistical notations: * = p < 0.05 compared with basal mycobacteria growth; # = p < 0.05 with Rifampicin. . Rifampicin was added in wells in dose 100 µg/mL, Chlorophyllipt and microalgae extract in dose 1% v/v and Fx in dose 35 µg/mL.
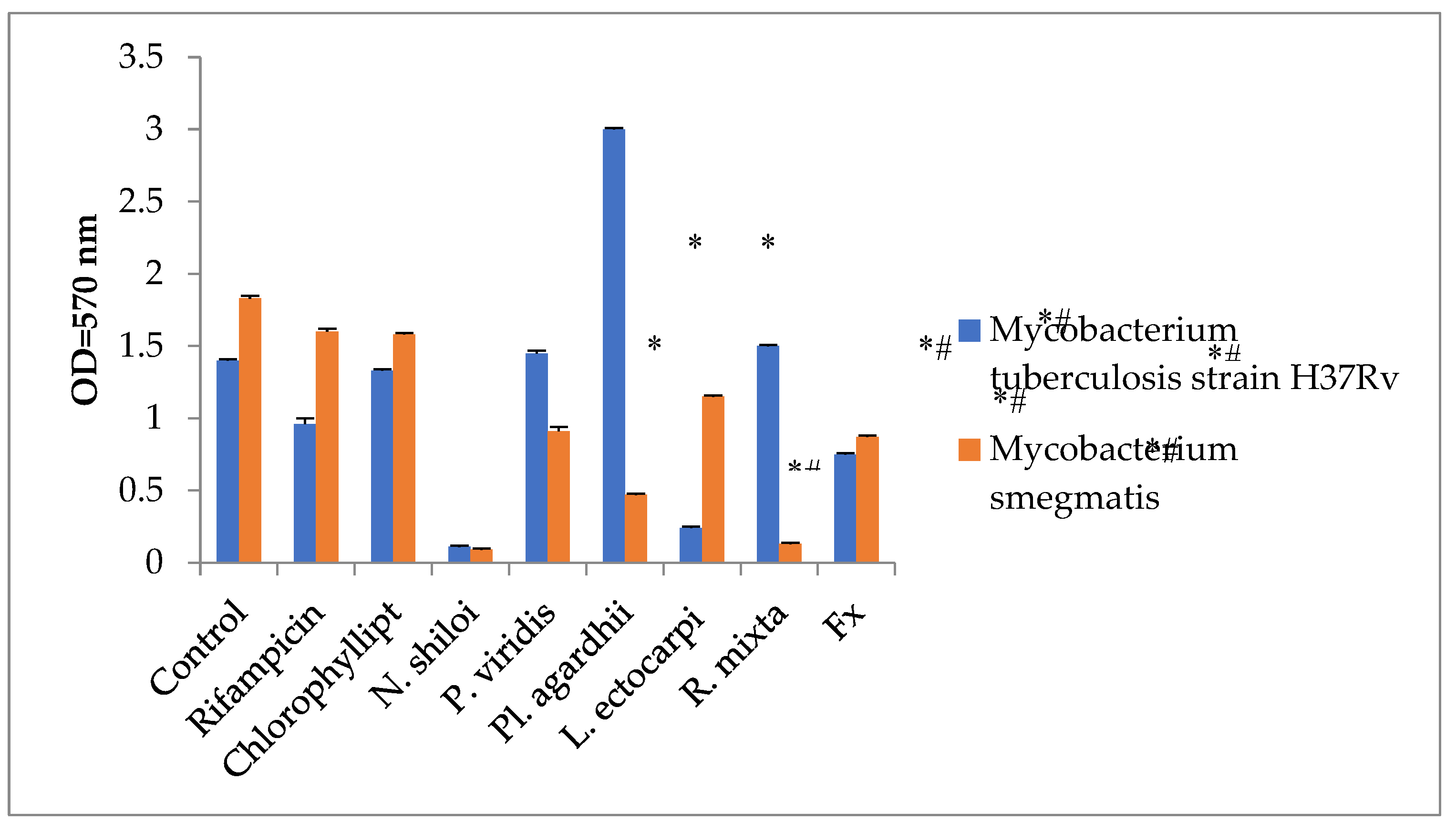
Figure 5.
The number of Mycobacterium tuberculosis CFU growth on Lowenstein-Jensen solid medium from lungs, liver and spleen of the mice with tuberculosis (M±SD). CFU, colony forming units; Rif, Rifampicin-treated group; Fx, Fucoxanthin-treated group. Statistical notations: * = p < 0.05 compared with TB-untreated group; # = p < 0.05 with Rifampicin-treated group.
Figure 5.
The number of Mycobacterium tuberculosis CFU growth on Lowenstein-Jensen solid medium from lungs, liver and spleen of the mice with tuberculosis (M±SD). CFU, colony forming units; Rif, Rifampicin-treated group; Fx, Fucoxanthin-treated group. Statistical notations: * = p < 0.05 compared with TB-untreated group; # = p < 0.05 with Rifampicin-treated group.
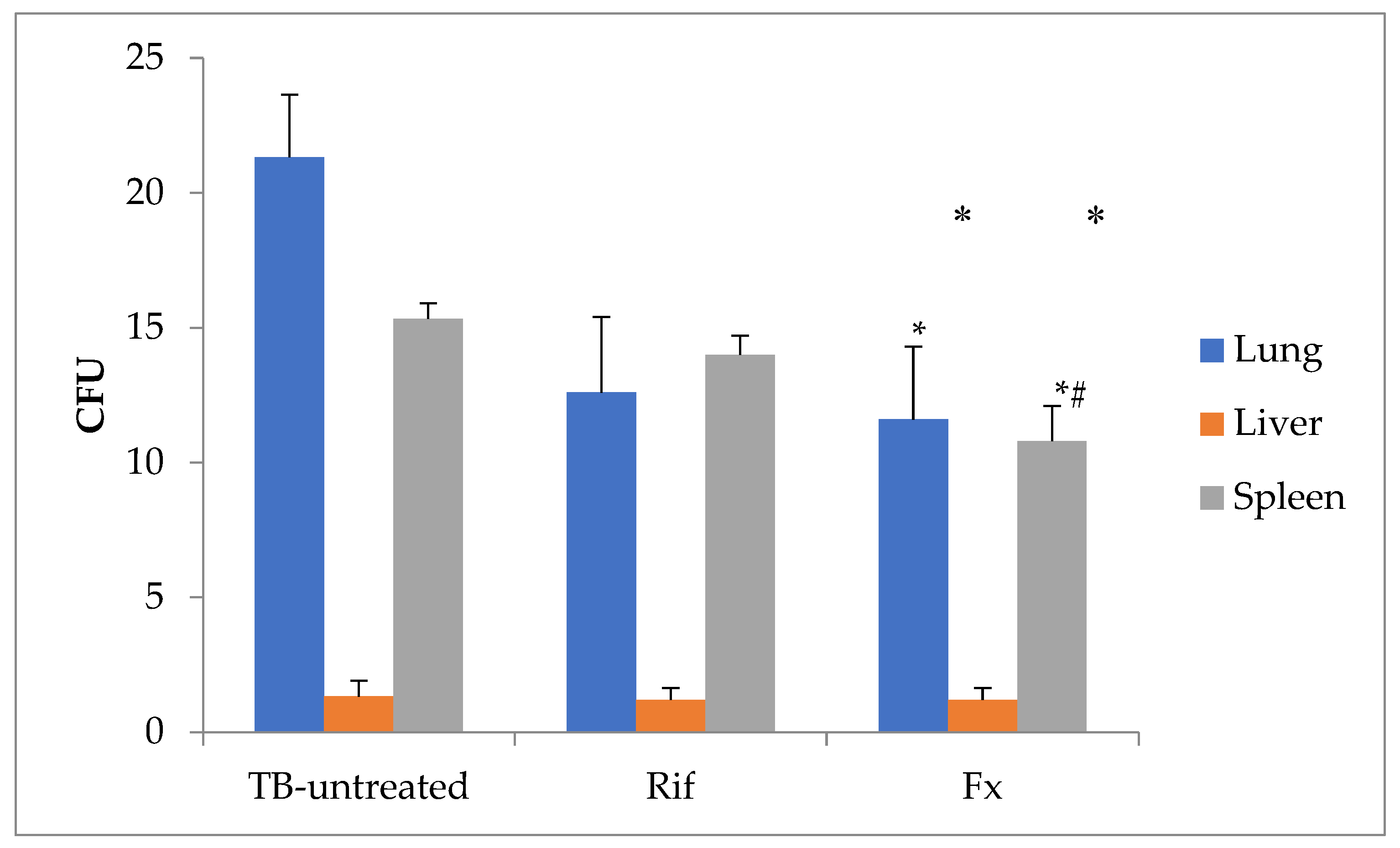

Table 1.
Biochemichal Characteristics of Microalgae (M±SD).
| Content, mg/g of Dry Biomass | L. cf. ectocarpi | N. shiloi | R. mixta sp. Nov. | P. agardhii | P. viridis |
|---|---|---|---|---|---|
| Proteins | 560±5 | 450±5 | 680±10 | 600±10 | 350±10 |
| Lipids | 250±5 | 280±5 | 50±5 | 100±5 | 195±6 |
| Carbohydrates | 50±3 | 70±3 | 110±5 | 180±5 | 340±5 |
| Fx | ND | 15 ±0.5 | ND | ND | ND |
| С-phycoerythrin | 78.8 ± 0.8 | ND | 35 ± 0.,8 | 20 ± 1.2 | ND |
| С-phycocyanin | 22.0 ±0.8 | ND | 140±1 | 88.8 ± 1 | ND |
| PUFA, include: | 50±1.2 | 67±1.25 | 10 ±1.,1 | 11±1.2 | 70±1.25 |
| Eicosapentaenoic acid (C20:5 ω3) | ND | 47.3±0.61 | ND | ND | 4.8±0.7 |
| Arachidonic acid (C20:4ω6) | ND | 8.5 ±0.31 | ND | ND | 0.6±0.6 |
| γ-Linolenic acid (C18:3 n-6) | 18± 0.04 | 0.22± 0.01 | 2.75 ± 0.02 | 1.5 ± 0.01 | 10.7 ± 0.05 |
| Linoleic acid (С18:2 n-6) | 25 ± 0.05 | 1.455±0.04 | 3.16 ± 0.02 | 2.1 ± 0.01 | 2.5 ± 0.02 |
| Oleic acid (C18:1 (n-9) | 0.1± 0.02 | 1.46 ±0.01 | ND | 2 ±0.01 | 0,1± 0,02 |
| α-Linolenic acid (C18:3 n-3) | 10,0 ± 0,05 | ND | ND | ND | 6,5 ± 0,05 |
Note. Fx, fucoxanthin; PUFA, polyunsaturated fatty acid; ND, not determined.
Table 2.
Effect of Fucoxanthin from Diatomeae on bacterial growth in vitro by MTT assay(M±SD).
| Parameters | Kl. pneumoniae (s) | Kl. pneumoniae (r) | S. aureus MSSA | S. aureus MRSA | S. aureus clinical isolate | A. baumannii (r) | E. faecalis (r) | Ps. aeruginosa (s) | Ps. aeruginosa (r) | S. pyogenes(r) |
|---|---|---|---|---|---|---|---|---|---|---|
| On Day 14 | ||||||||||
| Control | 1.18±0.08 | 1.75±0.2 | 1.1±0.09 | 1.47±0.08 | 1.46±0.16 | 0.98±0.17 | 1.08±0.09 | 1.94±0.33 | 1.37±0.33 | 1.88±0.32 |
| Cef | 0.82±0.08* | 1.24±0.04* | 1.03±0.14 | 1.32±0.1* | 1.49±0.13 | 0.7±0.1* | 2.1±0.32 | 1.29±0.03* | 1.3±0.03 | 1.53±0.27 |
| A. platensis | 0.6±0.17* | 0.8±0.05*# | 0.83±0.01*# | 0.96±0.11*# | 0.96±0.04*# | 0.68±0.11* | 1.15±0.13# | 0.49±0.15*# | 0.67±0.03*# | 1.3±0.07* |
| Fx | 0.83±0.1* | 0.66±0.05*# | 0.72±0.08*# | 0.46±0.07*# | 1.16±0.05*# | 0.68±0.14 | 2±0.26 | 0.5±0.06*# | 0.66±0.07*# | 2.07±0.29# |
| On Day 21 | ||||||||||
| Control | 2.52±0.9 | 3.66±0.39 | 4±1 | 4±0.9 | 4±1 | 3.09±1 | 4±1 | 4±0.8 | 4±1 | 4±0.9 |
| Cef | 2.15±0.9 | 3.53±0.54 | 2.07±0.45* | 3.28±0.96 | 1.23±1.03* | 4±0.9 | 4±0.8 | 3.75±0.51 | 3.23±0.97 | 2.39±1.67 |
| A. platensis | 0.54±0.24*# | 2.33±0.1*# | 1.98±0.43* | 2.07±0.51* | 2±0.65* | 0.85±0.29*# | 2.01±0.65*# | 1.99±0.69*# | 1.89±0.79* | 2±0.65* |
| Fx | 1.11±0.03*# | 2.75±0.2*# | 2.26±0.62* | 2.16±0.92 | 2.68±0.34# | 1.06±0.01*# | 2.6±0.34*# | 2.85±0.2*# | 2.89±0.1* | 2.2±0.5* |
Note. Cef, Ceftazidime; Fx, fucoxanthin from N. shiloi; (s), sensitive strain; (r) resistant strain; * = p < 0.05 with control; # = p < 0.05 with Ceftazidime. Ceftazidime was added in wells in dose 50 µg/mL, Fx in dose 35 µg/mL and microalgae extracts in dose 1% v/v.
Table 3.
Dose-dependent effect of microalgae DMSO extracts on bacterial growth in vitro by MTT assay (M±SD).
Table 3.
Dose-dependent effect of microalgae DMSO extracts on bacterial growth in vitro by MTT assay (M±SD).
| Parameters | N. shiloi | P. viridis | P. agardhii | L. ectoarpi | R. mixta | Fx |
|---|---|---|---|---|---|---|
| Kl. pneumoniasensitive [Control = 1.38±0.01, Ceftazidime = 0.92±0.01] | ||||||
| 1% v/v | 1.79±0.22 | 1.7±0.04 | 2.18±0.01 | 2.06±0.04 | 2.05±0.04 | 1.4±0.01 |
| 0.5% v/v | 2.11±0.08 | 2.01±0.02 | 2.33±0.02 | 1.91±0.07 | 1,74±0.05 | 1.35±0.01 |
| 0.25% v/v | 1.72±0.01 | 1.75±0.01 | 2.11±0.08 | 1.8±0.07 | 1.29±0.01* | 1.18±0.01* |
| Kl. pneumonia(ESBL) [Control = 1.83±0.02, Ceftazidime = 1.72±0.01] | ||||||
| 1% v/v | 1.3±0.01*# | 1.76±0.01* | 1.7±0.01 | 1.46±0.01 | 1.36±0.02*# | 1.18±0.01*# |
| 0.5% v/v | 1.19±0.01*# | 1.68±0. 01* | 1.68±0.01* | 1.24±0.02*# | 1.23±0.01*# | 0.83±0.01*# |
| 0.25% v/v | 1.22±0.01*# | 1.47±0.01*# | 1.1±0.01*# | 1.04±0.03*# | 0.97±0.01*# | 1.19±0.01*# |
| S. aureusMSSA [Control = 1.92±0.04, Ceftazidime = 0.43±0.01] | ||||||
| 1% v/v | 1.45±0.01* | 1.19±0.01* | 2±0.01 | 1.53±0.01* | 1.47±0.01* | 1.61±0.01* |
| 0.5% v/v | 1.08±0.01* | 1.27±0.01* | 1.47±0.01* | 1.69±0.0* | 1.49±0.01* | 1.53±0.01* |
| 0.25% v/v | 1.34±0.01* | 1.43±0.01* | 1.51±0.01* | 1.38±0.01* | 2.01±0.01 | 1.19±0.01* |
| S. aureusMRSA [Control = 2.11±0.07, Ceftazidime = 2.34±0.01] | ||||||
| 1% v/v | 1.52±0.03*# | 1.41±0.01*# | 2.01±0.02 | 1.92±0.1*# | 2.16±0.03 | 1.35±0.01*# |
| 0.5% v/v | 1.58±0.01*# | 1.08±0.01*# | 1.92±0.01*# | 1.8±0.01*# | 1.83±0.01*# | 1.25±0.01*# |
| 0.25% v/v | 1.59±0.03*# | 1.46±0.01*# | 1.31±0.01*# | 1.58±0.01*# | 1.98±0.01*# | 0.55±0.01*# |
| A. baumanniiresistant [Control = 2.09±0.03, Ceftazidime = 0.92±0.01] | ||||||
| 1% v/v | 2.92±0.01 | 2.39±0.02 | 3.62±0.08 | 2.28±0.01 | 2.23±0.01 | 1.43±0.02* |
| 0.5% v/v | 2.75±0.01 | 2.52±0.12 | 2.68±0.02 | 2.33±0.04 | 2.02±0.18 | 1.18±0.24* |
| 0.25% v/v | 2.15±0.01 | 2.53±0.06 | 2.64±0.05 | 2.18±0.01 | 2.03±0.01 | 0.9±0.1* |
| E. faecalis(VRE) [Control = 2.61±0.03, Ceftazidime = 0.92±0.03] | ||||||
| 1% v/v | 1.67±0.01* | 1.54±0.01* | 2.11±0.01* | 2.16±0.02* | 2.46±0.02* | 1.08±0.01* |
| 0.5% v/v | 1.64±0.01* | 1.82±0.01* | 2±0.01* | 2.18±0.01* | 2.4±0.01* | 1.01±0.01* |
| 0.25% v/v | 1.57±0.01* | 1.56±0. 01* | 1.38±0.01* | 1.69±0.03* | 1.61±0.01* | 0.59±0.01*# |
| Ps. aeruginosasensitive [Control = 1.77±0.01, Ceftazidime =1.4±0.05] | ||||||
| 1% v/v | 2.8±0.04 | 1.65±0.01* | 2.83±0.02 | 1.84±0.01 | 1.55±0.01* | 0.78±0,01*# |
| 0.5% v/v | 1.72±0.01 | 1.45±0.01* | 1.81±0.01 | 1.68±0.01 | 1.55±0.01* | 0.55±0.01*# |
| 0.25% v/v | 1.5±0.01 | 1.41±0.01* | 1.47±0.01* | 1.36±0.01* | 1.59±0.02* | 0.44±0.01# |
| Ps. aeruginosaresistant [Control = 1.73±0.01, Ceftazidime = 1.25±0.01] | ||||||
| 1% v/v | 2.33±0.02 | 1.35±0.01* | 2.28±0.02 | 1.46±0.01* | 0.92±0.01*# | 0.88±0.01*# |
| 0.5% v/v | 1.9±0.01 | 2.13±0.01 | 1,71±0.01 | 1.36±0.03* | 0.75±0.01*# | 0.7±0.01*# |
| 0.25% v/v | 1.64±0.01* | 1.45±0.02* | 1.63±0.01 | 1.19±0.03* | 0.67±0.01*# | 0.43±0.01*# |
| S. pyogenesresistant [Control = 0.98±0.01, Ceftazidime = 0.99±0.01] | ||||||
| 1% v/v | 0.66±0.01* | 0.84±0.01* | 1.05±0.01 | 0.91±0.01* | 0.99±0.03 | 0.47±0.04*# |
| 0.5% v/v | 0.53±0.06* | 0.79±0.03* | 0.95±0.08 | 0.28±0.02*# | 0.68±0.02*# | 0.54±0.04*# |
| 0.25% v/v | 1.55±0.07 | 0.36±0.05* | 1.14±0.06 | 0.99±0.09 | 0.78±0.08*# | 0.5±0.02*# |
Note. Fx, fucoxanthin from N. shiloi; * = p < 0.05 with control; # = p < 0.05 with Ceftazidime. Ceftazidime was added in wells in dose 50 µg/mL, Fx in dose 35-17.5-8.75 µg/mL and microalgae extracts in dose 1% v/v.
Table 4.
Comparison of antimicrobial activity of the Chlorophyllipt and different microalgae DMSO extract (M±SD).
Table 4.
Comparison of antimicrobial activity of the Chlorophyllipt and different microalgae DMSO extract (M±SD).
| Parameters | Control | Ceftazidime | Chlorophyllipt | N.shiloi | P. viridis | P. agardhii | L. ectoarpi | R. mixta | Fx |
|---|---|---|---|---|---|---|---|---|---|
| Kl. pneumoniae | 2.27±0.05 | 0.78±0.01 | 0.98±0.01* | 1.85±0.01* | 1.53±0.01* | 1.78±0.01* | 1.93±0.04* | 1.7±0.01* | 0.77±0.01*& |
| S aureus MRSA | 3.02±0.03 | 2.44±0.01 | 0.72±0.0*# | 0.9±0.01*# | 1.2±0.01*# | 1.93±0.02*# | 0.93±0.01*# | 0.9±0.01*# | 0.6±0.01*#& |
| A. baumannii | 3.49±0.03 | 0.92±0.01 | 1.04±0.01* | 1.6±0.02* | 1.88±0.04* | 1.86±0.0* | 2.27±0.02* | 1.53±0.01* | 0.8±0.01*#* |
| E. faecalis | 2.29±0.02 | 0.92±0.01 | 0.57±0.01*# | 1.41±0.01* | 1.78±0.01* | 1.82±0.26* | 2.06±0.01* | 1.7±0.0* | 2.03±0.02* |
| Ps. aeruginosa | 1.08±0.01 | 0.93±0.01 | 0.67±0.01*# | 0.49±0.01*#& | 1.23±0.02 | 2.18±0.03 | 2.54±0.03 | 0.74±0.001*# | 0.66±0.01*# |
Note. Fx, fucoxanthin from N. shiloi; * = p < 0.05 with control; # = p < 0.05 with Ceftazidime; & = p < 0.05 with Chlorophyllipt. Ceftazidime was added in wells in dose 50 µg/mL, Fx in dose 35 µg/mL and microalgae extracts in dose 1% v/v.
Table 5.
Dose-dependent effect of microalgae extracts on mycobacterium growth in vitro (M±SD).
| Parameters | N. shiloi | P.s viridis | P. agardhii | L. ectoarpi | R. mixta | Fx |
|---|---|---|---|---|---|---|
| Mycobacterium tuberculosis strain H37Rv [Control = 1.38±0.01, Rifampicin 100 µg/mL = 0.96±0.04] | ||||||
| 1% v/v | 1.74±0.01 | 1.72±0.01 | 1.33±0.01* | 1.09±0.01* | 0.62±0.01*# | 1.18±0.01* |
| 0.5% v/v | 0.97±0.01* | 1.41±0.01 | 1.06±0.01* | 0.89±0.01*# | 0.79±0.0*#1 | 0.73±0.01*# |
| 0.25% v/v | 0.67±0.01*# | 1.18±0.01 | 0.77±0.01*# | 0.57±0.01*# | 1.47±0.01 | 2.06±0.01 |
| Mycobacterium smegmatis [Control = 1.82±0.02, Rifampicin = 1.6 ±0.02] | ||||||
| 1% v/v | 1.91±0.01 | 1.08±0.01*# | 2.76±0.01 | 1.12±0.01*# | 0.99±0.01*# | 1.43±0.01*# |
| 0.5% v/v | 0.96±0.01*# | 2.01±0.01 | 0.93±0.01*# | 1.01±0.01*# | 0.76±0.01*# | 1.13±0.01*# |
| 0.25% v/v | 1.46±0.01*# | 1.94±0.01 | 0.84±0.01*# | 0.54±0.01*# | 1.57±0.01*# | 0.99±0.01*# |
Note. Fx, fucoxanthin from N. shiloi; * = p < 0.05 with control; # = p < 0.05 with Rifampicin. Fx was added in wells in dose 35-17.5-8.75 µg/mL.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



