
Submitted:
22 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
During embryo development, the endoplasmic reticulum (ER) acts as an important site for protein biosynthesis, however, in vitro culture (IVC) can negatively affect ER homeostasis. Thereby, the aim of our study was to evaluate the effects of supplementation of tauroursodeoxycholic acid (TUDCA), an ER stress inhibitor, in the IVC of bovine embryos. Two experiments were carried out: Exp. 1: evaluation of blastocyst rate, hatching kinetics and gene expression of hatched embryos after being treated with different concentrations of TUDCA (50, 200 or 1,000 μM) in the IVC; Exp. 2: evaluation of the re-expansion, hatching, and gene expression of hatched embryos previously treated with 200 µM of TUDCA at IVC and submitted to vitrification. There was no increase in the blastocyst and hatched blastocyst rates treated with TUDCA in the IVC. However, embryos submitted to vitrification after treatment with 200 µM of TUDCA, increased hatching rate post-warming, together with a down-regulation in the expression of ER stress-related genes and the accumulation of lipids. In conclusion, this work showed that the addition of TUDCA during in vitro culture can improve the cryotolerance of the bovine blastocyst through the putative modulation of ER and oxidative stress.
Keywords:
Endoplasmic reticulum
; endoplasmic reticulum stress
; embryo
; in vitro culture
; tudca
; vitrification
1. Introduction
Disturbances of the endoplasmatic reticulum (ER) homeostasis cause protein folding or misfolding in the ER lumen, a condition called ER stress, which triggers unfolded protein response (UPR) [1]. A paradox of the UPR pathway is that it leads to a response with simultaneous activation of cell survival and pro-apoptotic pathways. Under those ER stress conditions, activation of the UPR reduces unfolded protein load through several pro-survival mechanisms, including the expansion of the ER membrane, the selective synthesis of key components of the protein folding and the quality control machinery and the attenuation of the influx of proteins into the ER [2]. When ER stress is not mitigated and the homeostasis is not restored, the UPR triggers apoptosis. There are three predominant and unique signaling transduction mechanisms among the UPR signaling pathways: inositol-requiring enzyme 1(IRE1), protein kinase RNA (PKR)-like kinase (PERK) and activating transcription factor 6 (ATF6) [3,4]. The IRE1 endoribonuclease is activated through dimerization and transphosphorylation. This leads to the removal of a 26-nucleotide intron from the premature unmodified form of XBP1 (XBP1-u) gene to produce the unmodified XBP1 (XBP1-s) form [5]. XBP1-s moves to the nucleus and induces UPR-responsive genes. XBP1-s is usually regarded as a reliable marker for the induction of the IRE1 pathway of the UPR, because XBP1 is unmodified exclusively under ER stress conditions [6]. However, if ER stress is excessive or prolonged, the UPR fails, and cellular apoptosis is induced by activation of CCAAT-enhancer-binding-protein homologous protein (CHOP), Jun N-terminal kinase, and cleaved caspase 3 [7,8].
Early embryonic stages are (one of) the most critical periods of the mammalian development [9]. These early stages involve various morphological and biochemical changes related to genomic activity and a complex set of physiological processes, many of which are still unknown [10]. These processes are controlled by several molecular mechanisms and pathways that have a fundamental role in the coordination of homeostatic and metabolic processes [11,12]. Within in vitro systems, disturbances in the embryo's culture environment after fertilization can have detrimental effects on embryonic gene expression [13] which, in turn, can have serious implications for the normality of the blastocyst’s physiology. However, the exact influence of in vitro culture conditions during each of these critical events/steps is still unknown.
Recent studies in several species have shown that ER stress in the embryo impairs embryo developmental competence [14,15] and that stress relief can improve the embryo quality [15,16,17,18]. In the bovine species, the supplementation of the in vitro culture (IVC) medium with tauroursodeoxycholic acid (TUDCA) - a bile acid that acts as a potential chemical chaperone against ER stress in vitro [19] - has been an alternative to relieve ER stress and improve the developmental competence in embryos cloned by somatic cell nuclear transfer [16] and embryos subjected to high O2 tension [20].
Nevertheless, little is known about the effects of ER stress on cryogenic tolerance of embryos generated in vitro [21]. It is studied that the in vitro produced embryo (IVPE) has distinct characteristics when compared to its counterpart produced in vivo. These differences between IVPE and in vivo derived from embryos which involve morphological [22] and molecular aspects that affect the embryo quality and development [23] decreasing cryo-tolerance and pregnancy rates [24]. Therefore, maintaining cell viability after warming is a prerequisite to achieve high outcomes within cryopreservation protocols. To help bridge this gap between IVPE and in vivo derived from embryos to improve post-cryopreservation results, the ER stress could be an alternative target for pharmacological approach.
We hypothesized that supplementation of TUDCA during IVC, decreases the endoplasmic reticulum stress of bovine embryos and improves the cryogenic competence of the embryo related to re-expansion and post-warming hatching rate. Thus, the present study aimed: 1) to evaluate the developmental competency of bovine embryos after being treated with TUDCA in IVC; 2) to investigate the effects of TUDCA treatment in IVC on subsequent developmental competency post-warming of the vitrified blastocysts.
2. Results
2.1. The effects of TUDCA during IVC of bovine embryos
2.1.1. Developmental competence
After 7 days of culture, in the absence and in the presence of TUDCA, there was no difference in the rate of blastocysts among the Control group and the treatments with TUDCA. Yet, group T200 presented a higher rate of blastocysts when compared to group T1000 (P=0.0268; Figure 1). When evaluating the embryo hatching kinetics, there was no difference between treatments on day 7. However, on days 8 and 9, the T1000 group (7.3% and 6.95%, respectively) was statistically different from day 8 Control (26.4%) and all treatments by day 9 (23.8%, 24.2% and 27.9%; respectively to Control group, T50 and T200; Figure 2). The results suggested a toxic effect in the group treated with 1,000 µM of TUDCA altering the embryonic development kinetics and, subsequently, decreasing its ability to hatch.
2.1.2. Gene expression
Transcript abundance from 10 genes was significantly affected in hatched blastocyst after TUDCA treatment, with all genes upregulated in the T1000 group in comparison to the other groups (Figure 3). Of the differentially expressed genes, there were transcripts related to endoplasmic reticulum stress (ATF6, EIF2A, and HSPD1), oxidative stress and response to cellular stress (GFPT2, HMOX1 and TXNRD1), metabolism (ACACA, ELOVL5 and SREBF2), the embryo development and cell proliferation (GSK3A; Figure 3). Corroborating with the analysis of the developmental competence, the T1000 group appeared to be toxic to the embryo, increasing the abundance of transcripts related to ER stress and oxidative stress.
2.2. The effects of TUDCA during IVC on post-warmed vitrified blastocysts
2.2.1. Developmental competence
There was no difference in the re-expansion rate evaluated after 12, 24 and 48 hours of warming up (P>0.1; Table 1). Nevertheless, a larger number of hatched blastocysts were observed from TUDCA treatment in the assessment post-warming (Table 1). After 24- and 48-hours post-warming, there was a higher significantly number of hatched blastocysts in the embryos cultivated with TUDCA compared to control (P=0.09 and 0.0423, respectively; Table 1). The results suggested a beneficial effect of treatment with TUDCA by increasing the hatching rates of embryos subjected to vitrification.
2.2.2. Gene expression
The transcript abundance from 11 genes was affected in hatched blastocyst after vitrification and post-warming. When compared to Control group, in T200 group 5 genes were down-regulated, and 6 genes were up-regulated (Figure 4). Down-regulation was observed for genes related to endoplasmic reticulum stress (HSPA5 and XBP1), oxidative stress and response to cellular stress (GLRX2), and some markers for metabolism (PLIN3 and SREBF2; Figure 4). Up-regulation was observed for genes related to oxidative stress and response to cellular stress (CAT, GPX1, NFE2L2 and PRDX1) and metabolism (G6PD and SLC2A3; Figure 4). The treatment with TUDCA could have relieved the ER stress and modulated the lipid metabolism of the hatched embryo, but direct evaluations of these pathways were not performed. String analysis of those genes statistically different reveals a correlation between genes related to endoplasmic reticulum stress and genes related to oxidative stress and response to cellular stress (Figure 5).
3. Discussion
Developing embryos may be subjected to several sources of exogenous stress in in vitro culture system [15,31,32,33]. These include oscillating temperature, DNA damage or DNA damaging agents, osmotic stress, and availability of organic osmolytes, oxygen and oxidative stress, hyperglycemia and carbon substrate availability, hyperlipidemia and oxidized lipids, calcium ionophores, cytokines, amino acid deprivation, insulin signaling, and serum components [15,31,32,33]. Cold stress associated with cryopreservation affects embryo development and gene regulation [22,24,34]. These adverse factors negatively impact ER functions and protein synthesis and folding, resulting in the activation of ER stress and the UPR signaling pathways in in vitro produced embryo [14,15,16,17]. Furthermore, there is evidence that IVPEs are more sensitive to cryopreservation than in vivo derived from embryos [48,50] and that this reduced cryotolerance which may be associated with the high lipid content present in the cytoplasm as well as the lipid profile of the cell membrane of these embryos [23,24,48]. Although TUDCA has been demonstrated to exert efficient cytoprotective activity in relieving ER stress [17,18,20,21], recent studies reported on its new potential and molecular modes of action as a weight-reducing agent, modulating lipid metabolism through or independently of ER modulation [51,52]. In the current study, we demonstrated that 200 µM of TUDCA during IVC enhanced the cryotolerance of bovine embryo through the putative modulation of ER and oxidative stress. At 24- and 48-hours post-warming, embryos treated with TUDCA during IVC, had an increase in hatching rate when compared to Control group.
Unlike our results, it was reported in cattle that 10 µM TUDCA was able to improve cryotolerance of embryos after vitrification, increasing hatching rates and decreasing the number of apoptotic cells in the embryo 48-hours post-warming [21]. In this way, the action of TUDCA seems to be highly dependent on the complex combination of variables such as the species, breed, the used concentration, and the in vitro culture conditions.
When we evaluated the 96 markers of transcript abundance in the hatched embryos post-warming, TUDCA treatment induced the decreasing of the mRNA abundance related to ER stress and lipid metabolism pathways. An increase in the mRNA abundance related to antioxidant activity was also observed in embryos in the T200 group. After warming post-vitrification, hatched blastocysts treated with TUDCA showed less mRNA abundance for HSPA5 and XBP1. In several species, the increasing of XBP1-s expression is widely used as a molecular marker of ER stress in vivo and in vitro [53,54]. The increasing mRNA abundance for XBP1-s and HSPA5 was associated with the low competence of embryonic development in several species [18,20,50]. In addition, after warming vitrified embryos treated with TUDCA showed a change in the expression of antioxidants, with high mRNA abundance for CAT, GPX1, NFE2L2 and PRDX1, and less abundance for GLRX2. NFE2L2 is a promising target against oxidative stress, responsible for inducing the expression of several endogenous cytoprotective enzymes [55,56]. In vitro study with the human neuroblastoma SH-SY5Y cell line, observed that TUDCA prevented oxidative stress through the highest expression of NRF2, DJ-1, and antioxidant enzymes heme oxygenase-1 (HO-1) and glutathione peroxidase (GPx) [57], corroborating with our results. The previous study has shown that embryos produced in vivo and cryopreserved undergo greater oxidative stress when compared to embryos that have not been subjected to cryopreservation [34]. In the conditions of this paper, the treatment with TUDCA could have prevented high levels of oxidative stress in vitrified embryos cultured after warming. Collectively, those data could partially explain the higher cell competence to hatch observed with TUDCA treatment, since a mitigated stress (ER and oxidative) improves the cellular activity.
The hatched embryos also modulated markers related to the metabolism. While the mRNA abundance for PLIN3 and SREBF2 decreased, the abundance for G6PD and SLC2A3 were increased with TUDCA treatment.
The reduced post-warming cell viability is also associated with the abnormal amount and/or the type of lipids in the blastomeres that contributes to the occurrence of cryogenic fractures during the freezing process [23,48]. The cell membrane fluidity is related to the lipid profile and the capacity to support cryo-injuries during cryopreservation process [48]. The IVPEs possessing has a different lipid profile from their in vivo derived counterparts [48,50] could have cryotolerance enhanced with changes in the lipid metabolism (e.g., TUDCA treatment). Although the evaluation of the lipid content was not assessed in this work, we cannot rule out that TUDCA treatment may have modulated the lipid content of the hatched embryo (down-regulated mRNA abundance of PLIN3 and SREBF2 in T200 group).
In addition, G6PD (Glucose-6-phosphate dehydrogenase) and SLC2A3 (Solute carrier family 2, facilitated glucose transporter member 3) mRNAs were up-regulated in the embryo treated with TUDCA. It was reported that those genes were essential for pre-implantation embryonic development [58,59,60]. Partially, this finding added data to explain how the TUDCA treatment could improve the post-warming hatching rate of vitrified embryos. Since the mammalian hatching process has involved a coordinated trophectoderm activity and is concomitant with the beginning of hypoblast appearance in bovine species [61], the up-regulation of G6PD and SLC2A3 genes suggest a positive marker after TUDCA treatment.
TUDCA supplementation in IVC was associated with improved embryonic developmental rates in mice [17,30], pigs [18,28], and cows [20,21]. In our experiment, we did not find a significant increase in the rates of blastocyst formation with TUDCA treatments. Similarly, in conditions of low O2 tension (5%), supplementation of 50 µM of TUDCA in IVC of bovine embryos did not modulate embryo competence [20]. However, embryos that were submitted to high tension O2 (20%) in the IVC showed an increased blastocyst rate in cattle and pigs with the supplementation of, respectively, 50 and 200 µM of TUDCA [18,20]. Once again, the potential beneficial effect of TUDCA supplementation seems to be linked with the culture conditions. When stringent condition of the embryo culture is used (e.g., high oxygen tension as source to generate an increase of the reactive oxygen species) the effect of TUDCA to alleviate ER stress was observed [18,20] but, that was not the case of our study. Corroborating with our result, some other recent studies have shown that the increase in reactive oxygen species (ROS), due to O2 tension, is closely related to the increase in ER stress in embryos [20]. In addition, unlike the purpose of our study, TUDCA proved to be beneficial for the development of in vitro produced embryos in conditions where ER stress was chemically or physically actively induced (e.g., using tunicamycin or heat stress, respectively) [18,20,21,35,36,37].
Complementing these results, when we evaluated the hatching kinetics of embryos treated with TUDCA, a significant reduction in the hatched embryos rate was observed with the T1000 group when compared to the other groups (with or without the addition of TUDCA). Contrary to the reported in mice – which had a positive effect on embryonic development and the newborn rate was described with the addition of 1,000 µM of TUDCA in the culture [30] - the higher concentration of TUDCA used in our study proved to be toxic to the bovine embryo and impaired its development. Furthermore, the factor already mentioned (O2 tension linked to the TUDCA effect) seems that the species (mouse or cattle) also plays a role on the upper threshold of the beneficial effect of TUDCA (i.e., when the threshold is exceeded, and the toxic effect is observed).
In the analysis of the mRNA abundance involved in ER stress, oxidative stress, metabolism and embryonic quality, the negative effect on hatching rate of the T1000 group was reinforced.
In case of misfolded proteins in the ER lumen, molecular chaperones (HSPD1 and HSPA5) are activated in order to correct this misfolding and maintain homeostasis in the ER [38; 39]. For instance, in cases of ER stress, HSPA5 dissociates from PERK, ATF6, and IRE1 receptors, activating the UPR pathway [3,4,38]. Activated PERK can recognize and phosphorylate eIF2α, which in turn regulates positively the translation of ATF4, an important inducer of CHOP, GADD34, ATF-3, and genes involved in apoptosis [40]. In the T1000 group, there was an increase on the transcript abundance for HSPD1, ATF6, and EIF2A in hatched embryos. This suggests that the higher concentration of TUDCA of this study paradoxically induced ER stress in the blastocysts. Also, an increase in the gene’s expression involved with oxidative stress (e.g., GFPT2, HMOX1, and TXNRD1) was observed and reinforced the close relationship between ER stress and oxidative stress, where ROS functioned as a mediator of these two events [41,42]. Oxidative stress in embryos could lead to DNA damage [31] and inhibit preimplantation development [43]. For transcripts involved in lipid metabolism (SREBF2, ELOVL5 and ACACA), there was an up-regulation in embryos of the T1000 group. Crosstalk between ER stress and lipid metabolism was well established [44,45,46,47]. Several reports indicated that the pathways that regulate UPR also induce the lipid accumulation in the cell. For instance, the ATF6α pathway plays a role in lipid accumulation interacting with the nuclear form of SREBP-2 [46,47]. In the literature, the lipid accumulation in embryos is associated with lower rates of embryonic survival after cryopreservation and deviations in the relative abundance of transcripts of important genes for embryonic development [23,48]. Additionally, in the corroborating the results, the T1000 group has been shown to increase the transcript abundance for GSK3A (related to embryo development and cell proliferation). GSK3A is a negative regulator in the hormonal control of glucose homeostasis, cell division, proliferation, motility, and survival. In other publications, the highest expression of GSK3A in the embryo is associated with low embryo competence. [49].
Unlike the T1000 group, the T50 and T200 groups did not significantly affect gene expression in hatched blastocysts when compared to Control, corroborating the results of embryonic development. Although without molecular and cellular evidence of any beneficial effect of TUDCA (T50 and T200 groups), there was no assessment on the pregnancy rate of those embryos (fresh transfer) or the effect of high oxygen culture system.
4. Materials and Methods
4.1. In vitro Production
Bovine ovaries (mainly Bos t. indicus and its crossbreeds) were obtained from a commercial abattoir located at Assis (São Paulo, Brazil) and during the months of August and September. They were transported to the laboratory in sterile saline (0.9% NaCl) at 37°C for 30 minutes at maximum. Cumulus oocyte complexes (COCs) were collected by aspiration of follicles with 3–8 mm in diameter [25]. After sedimentation, COCs were recovered and selected using a stereomicroscope. Only the COCs with a homogeneous cytoplasm and a compact multilayer of cumulus cells were used (grades 1 and 2) [26]. COCs were washed and transferred to 500 µL drops of maturation medium (10 µL/COCs) in 4-well dishes (Nunc, Roskilde, Denmark), which consisted of TCM199 containing Earle salts supplemented with 0.1 IU/mL rhFSH (Gonal-f, Merck Serono, Rockland, MA, USA), 0,22 mg/mL sodium pyruvate, 75 µg/mL amikacin, 4 mg/mL BSA. Drops were incubated at 38.5°C in humidified air with 5% CO2 for 24 h.
Following in vitro maturation (IVM), groups of 25 COCs were transferred to 90 µL drops of Tyrode Albumin Lactate Pyruvate (TALP) supplemented with fatty-acid-free BSA (6 mg/mL), pyruvate (0,22 mg/mL), amikacin (75 µg/mL), heparin (30 µg/mL) and PHE (20 µM penicillamine, 10 µM hypo taurine and 1 µM epinephrine). Oocytes were subjected to in vitro fertilization (IVF) step with frozen-thawed semen from a single sample of a Nellore breed bull (named “Ópio”). Spermatozoa were selected by the Select SPERM (Botupharma Animal Biotechnology, Botucatu, São Paulo, Brazil) method, and the concentration was adjusted to 1 x 106 spermatozoa/mL. Oocytes and spermatozoa were co-incubated under the same conditions as during IVM, and the day of insemination was designated as Day 0. At 18-20 hours post-insemination, presumptive zygotes were denuded from cumulus cells and transferred to 500 µL drops of SOF medium (synthetic oviduct fluid; 10 µL/zygotes) in 4-well dishes, supplemented with pyruvate (0,22 µg/mL), amikacin (75 µg/mL), 2.5% v/v of FCS and BSA (5 mg/mL), and concentrations of TUDCA according to the experimental design described below. All experiments had a Control group (i.e., culture medium without the addition of TUDCA). Subsequently, they were cultivated in physiological oxygen tension (5%) in small, sealed plastic bags with a gas mixture of 5% O2, 5% CO2 and 90% N2 [27], and high humidity in an incubator at 38.5° C. The culture was maintained to all experiments, for 9 days after insemination to reach the hatching stage of the embryos.
4.2. Chemical Treatment
Tauroursodeoxycholic acid sodium salt (TUDCA; Selleckchem, Houston, TX, USA) was dissolved in sterile, distilled water to make a 100 mM stock solution (stored at -80° C). This stock solution was diluted into culture media to make 50 µM, 200 µM, and 1,000 µM solutions of TUDCA at experiment 1 (respectively, groups T50, T200 and T1000) [28,29,30]. The experiment 2 (vitrification) was performed only with the T200 group.
4.3. Target-Transcripts Relative Quantitation: RT-qPCR
4.3.1. RNA isolation and reverse transcription
The total RNA from hatched blastocysts was extracted with the PicoPure RNA Isolation kit (Life Technologies, Foster City, CA, USA) following the manufacturer’s protocol. Extracted RNA was stored at -80°C until further analysis by qPCR. RNA concentration was quantified by a spectrophotometer (Nanodrop, ThermoFischer Scientific, MA, USA).
We used for each sample a pool of 3 hatched blastocysts to reverse transcription. The cDNA synthesis was performed using High-Capacity Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA), following manufacturer’s instructions. DNAse treatment was performed in all samples before reverse transcription as manufactures’ instructions.
4.3.2. Preamplification and quantitative polymerase chain reaction
Gene expression analysis of blastocysts was performed independently, using Applied Biosystems™ TaqMan® Assays, specific for Bos t. taurus. A total of 80 candidate genes was analyzed (Supplementary Material-Table S1). Prior to qPCR thermal cycling, each sample was submitted to sequence-specific preamplification process as follows: 1.25 µL assay mix (TaqMan® Assay was pooled to a final concentration of 0.2X for each of the 96 assays), 2.5 µL TaqMan PreAmp Master Mix (Applied Biosystems, #4391128) and 1.25 µL cDNA (5 ng/µL). The reactions were activated at 95°C for 10 min followed by denaturing at 95ºC for 15s, annealing and amplification at 60ºC for 4 min for 14 cycles. These pre-amplified products were diluted 5-fold prior to RT-qPCR analysis.
Assays and pre-amplified samples were transferred to an integrated fluidic circuits (IFC) plate. For gene expression analysis, the sample solution prepared consisted of 2.25 µL cDNA (pre-amplified products), 2.5 µL of TaqMan Universal PCR Master Mix (2X, Applied Biosystems) and 0.25 µL of 20X GE Sample Loading Reagent (Fluidigm, South San Francisco, CA, USA); and the assay solution: 2.5 µL of 20X TaqMan Gene Expression Assay (Applied Biosystems) and 2.5 µL of 2X Assay Loading Reagent (Fluidigm). The 96.96 Dynamic Array™ Integrated Fluidic Circuits (Fluidigm) chip was used for data collection. After priming, the chip was loaded with 5 µL of each assay solution and 5 µL of each sample solution and was loaded into an automated controller that prepares the nanoliter reactions.
The qPCR thermal cycling was performed in the Biomark HD System (Fluidigm) using the protocol TaqMan GE 96x96 Standard, which consisted of one stage of Thermal Mix (50ºC for 2 min, 70ºC for 20 min and 25ºC for 10 min) followed by a Hot Start stage (50ºC for 2 min and 95ºC for 10 min), followed by 40 cycles of denaturation (95ºC for 15 s), primer annealing and extension (60ºC for 60 s).
4.4. Vitrification of embryos
4.4.1. Embryo freezing
All media used for vitrification and for warming were from Vitrogen Ltd. (Cravinhos, São Paulo, Brazil). The Vitrific® device was purchased from WTA Ltd. (Cravinhos, São Paulo, Brazil).
Quality grades 1 and 2 [26] expanded blastocysts were collected on days 7 and 8 of culture (D7 and D8; n=205). Blastocysts were removed from the culture medium (Control group and T200) and washed three times in holding medium (washing medium at 37ºC). After this, a vitrification solution 1 (V1) was used at room temperature, in which the embryos were submitted to two sequential baths for 8 minutes each. Subsequently, the embryos were transferred to a second solution (vitrification solution 2; V2). In this solution, they remained by just 40 seconds and, then 3 to 5 structures were allocated to the Vitrific® device with the aid of a glass micropipette (approximate inner diameter of 150 μm) to ensure the loading with the minimum possible of the medium. Immediately after that, the structures were deposited on the device, the excess solution was removed, leaving only a thin layer of medium on the structures. Afterwards, the Vitrific® device was plunged directly into the liquid nitrogen (N2) in a vertical position. Finally, the protective cap was placed with the device still submerged in N2. The blastocysts remained vitrified for an average of 12 to 24 hours.
4.4.2. Warming and culture of cryopreserved embryos
For warming, the Vitrific® device was removed from the N2 with the aid of tweezers. The device’s cap was removed and immediately after, its tip containing the structures was submerged into the warmed solution 1 (D1) at 37° C for 1 minute. Subsequently, the embryos were washed in two drops of warmed solution 2 (D2) and two drops of warmed solution 3 (D3) at room temperature, with an average time of 6 minutes for all steps. After that, the embryos were washed in 5 drops of culture medium. Then, the embryos were transferred to 4-well dishes with 500 µL of culture medium (maximum of 50 structures per well) and they were incubated in 38°C and 5% O2 tension in humidified air (IVC conditions). In the end, embryos were evaluated for re-expansion and hatching rate at 12-, 24- and 48-hours post-warming.
4.5. Experimental Design
4.5.1. Experiment 1: The effects of TUDCA during IVC on developmental competence and gene expression of embryos
To investigate the effect of TUDCA on embryo development and quality, blastocyst and hatched blastocyst rates were analyzed, respectively, on day 7 and days 8-9 (Day 0 being the day of insemination). Hatched blastocysts were collected on days 8-9 for gene expression analysis. After RNA extraction and cDNA reverse transcription, the Biomark HD platform was used to relatively quantify the mRNA abundance of markers of interest. This experiment was replicated five times using 250 presumptive zygotes/treatment.
4.5.2. Experiment 2: The effects of TUDCA during IVC on developmental competence and gene expression of post-warmed vitrified blastocysts
Following the treatment with TUDCA in the culture, expanded blastocysts were collected on days 8-9 and subjected to the technique of vitrification. After 12-24 hours the cryopreserved blastocysts were warmed and cultured at low tension of O2. Embryo re-expansion and hatching rates were analyzed at 12-, 24- and 48-hours post-warming. Hatched embryos were collected 24- and 48-hours post-warming for the analysis of gene expression. Later, RNA extraction and cDNA reverse transcription, the Biomark HD platform was used to relatively quantify the mRNA abundance of markers of interest. This experiment was replicated five times using 88 to 117 vitrified embryos/treatment.
4.6. Statistical analysis
To the embryonic development (experiments 1 and 2), the blastocysts and hatched blastocysts rates were arcsines transformed and subjected to analysis of variance (ANOVA), and the means were compared using the post hoc Tukey test. The normality was assessed with the Shapiro–Wilk test and Bartlett’s test. The results were presented as the mean ± standard error of the mean (SEM). For quantitative PCR data, we calculated the ΔCq values relative to the geometric mean of the best reference genes - i.e., B2M, HPRT1 and PPIA (experiment 1) and ACTB, HPRT1 and PPIA (experiment 2) - among the 96-gene set. Fold-changes were calculated as 2-ΔCq. All analyses were performed using JMP software (SAS Institute, Cary, NC, USA). Statistical significance was determined based on a P-value ≤ 0.1, the significance being considered weak (between 0.051 and 0.1), moderate (between 0.010 and 0.050) or strong (less than 0.01).
5. Conclusions
The present study showed that the addition of TUDCA during in vitro culture can improve the cryotolerance of the bovine blastocyst through the putative modulation of ER and oxidative stress. However, (and) in the culture conditions of this study, there was no observed effect on embryo development with the treatment of 50, 200 and 1,000 µM of TUDCA. Moreover, the highest concentration (1,000 µM), proved to be detrimental to the development and kinetics of the embryo.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, E.M.P. and M.F.G.N; Methodology, E.M.P., C.B.C., F.F.F., P.H.S. and M.F.G.N; Software, E.M.P., C.B.C. and M.F.G.N; Validation, E.M.P., C.B.C., F.F.F., P.H.S., and M.F.G.N; Formal Analysis, E.M.P., C.B.C. and M.F.G.N; Investigation, E.M.P., C.B.C. and M.F.G.N; Resources, M.F.G.N; Data Curation, E.M.P. and M.F.G.N; Writing – Original Draft Preparation, E.M.P. and M.F.G.N; Writing – Review & Editing, E.M.P., C.B.C. and M.F.G.N; Visualization, E.M.P., C.B.C., F.F.F., P.H.S. and M.F.G.N; Supervision, M.F.G.N; Project Administration, E.M.P. and M.F.G.N; Funding Acquisition, M.F.G.N. All authors provided feedback on the manuscript and declare no conflicts of interest. All authors have read and agreed to the published version of the manuscript.
Funding
Supported by Sao Paulo Research Foundation (FAPESP; grant #2012/50533-2).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data of this present study are available from the corresponding authors.
Acknowledgments
The authors would like to thank the Multi-user Laboratory of Phytomedicines, Pharmacology, and Biotechnology (FitoFarmaTec), São Paulo Research Foundation (FAPESP, grant #2012/ 50533-2), and - Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chipurupalli S, Kannan E, Tergaonkar V, D’Andrea R, Robinson N. Hypoxia Induced ER Stress Response as an Adaptive Mechanism in Cancer. Int. J. Mol. Sci. 2019, 20, 749. [Google Scholar] [CrossRef] [PubMed]
- Schroder M, Kaufman RJ. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef] [PubMed]
- Ghemrawi R, Battaglia-Hsu SF, Arnold C. Endoplasmic Reticulum Stress in Metabolic Disorders. Endoplasmic Reticulum Stress in Metabolic Disorders. Cells 2018, 7. [Google Scholar]
- Shen XH, Zhang KZ, Kaufman RJ. The unfolded protein response—A stress signaling pathway of the endoplasmic reticulum. J. Chem. Neuroanat. 2004, 28, 79–92. [Google Scholar] [CrossRef]
- Iwawaki T, Akai R, Kohno K, Miura M. A transgenic mouse model for monitoring endoplasmic reticulum stress. Nat. Med. 2004, 10, 98–102. [Google Scholar] [CrossRef]
- Van Schadewijk A, Van’t Wout EF, Stolk J, Hiemstra PS. A quantitative method for detection of spliced X-box binding protein-1 (XBP1) mRNA as a measure of endoplasmic reticulum (ER) stress. Cell Stress. Chaperones 2011, 17, 275–279. [Google Scholar]
- Szegezdi E, Logue SE, Gorman AM, Samali A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef]
- Van der Kallen CJ, Van Greevenbroek MM, Stehouwer, CD, Schalkwijk, CG. Endoplasmic reticulum stress-induced apoptosis in the development of diabetes: Is there a role for adipose tissue and liver? Apoptosis 2009, 14, 1424–34. [Google Scholar] [CrossRef]
- Zeng F, Schultz RM. RNA transcript profiling during zygotic gene activation in the preimplantation mouse embryo. Dev. Biol. 2005, 283, 40–57. [Google Scholar] [CrossRef]
- Paria BC, Dey SK. Preimplantation embryo development in vitro: Cooperative interactions among embryos and role of growth fac¬tors. Proc. Natl. Acad. Sci. USA 1990, 87, 4756–60. [Google Scholar] [CrossRef]
- O’Neill, C. Evidence for the requirement of autocrine growth fac¬tors for development of mouse preimplantation embryos in vitro. Biol. Reprod. 1997, 56, 229–37. [Google Scholar] [CrossRef] [PubMed]
- Singh M, Chaudhry P, Asselin E. Bridging endometrial receptivity and implantation: Network of hormones, cytokines, and growth factors. J. Endocrinol. 2011, 210, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Rizos D, Ward F, Duffy P, Boland MP, Lonergan P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Michalak M, Gye MC. Endoplasmic reticulum stress in periimplantation embryos. Clin. Exp. Reprod. Med. 2015, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Latham KE Stress signaling in mammalian oocytes and embryos: A basis for intervention and improvement of outcomes. Cell Tissue Res. 2016, 363, 159–67. [CrossRef] [PubMed]
- Song BS, Yoon SB, Sim BW, Kim YH; et al. Valproic acid enhances early development of bovine somatic cell nuclear transfer embryos by alleviating endoplasmic reticulum stress. Reprod. Fertil. Dev. 2014, 26, 432–40. [Google Scholar] [CrossRef]
- Zhang JY, Diao YF, Kim HR, Jin DI. Inhibition of endoplasmic reticulum stress improves mouse embryo development. PLoS ONE. 2012, 7, e40433. [Google Scholar]
- Kim JS, Song BS, Lee KS, Kim DH; et al. Tauroursodeoxycholic acid enhances the pre-implantation embryo development by reducing apoptosis in pigs. Reprod. Domest. Anim. 2012, 47, 791–798. [Google Scholar] [CrossRef]
- Lee YY, Hong SH, Lee YJ, Chung SS; et al. Tauroursodeoxycholate (TUDCA), chemical chaperone, enhances function of islets by reducing ER stress. Biochem. Biophys. Res. Commun. 2010, 397, 735–39. [Google Scholar] [CrossRef]
- Yoon SB, Choi SA, Sim BW, Kim JS; et al. Developmental competence of bovine early embryos depends on the coupled response between oxidative and endoplasmic reticulum stress. Biol. Reprod. 2014, 90, 104. [Google Scholar]
- Khatun H, Ihara Y, Takakura K, Egashira K; et al. Role of endoplasmic reticulum stress on developmental competency and cryo-tolerance in bovine embryos. Therio. 2020, 142, 131–37. [Google Scholar] [CrossRef] [PubMed]
- Papis K, Shimizu M, Izaike Y. Factors affecting the survivability of bovine oocytes vitrified in droplets. Therio. 2000, 54, 651–58. [Google Scholar] [CrossRef] [PubMed]
- Abe H, Yamashita S, Satoh T, Hoshi H. Accumulation of cytoplasmic lipid droplets in bovine embryos and cryotolerance of embryos developed in different culture systems using serum-free or serum-containing media. Mol. Reprod. Develop. 2002, 61, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sudano MJ, Paschoal DM, da Silva Rascado T, Crocomo LF; et al. Crucial surviving aspects for vitrified in vitro-produced bovine embryos. Zygote. 2014, 22, 124–31. [Google Scholar] [CrossRef]
- Botigelli RC, Razza EM, Pioltine EM, Fontes PK; et al. Supplementing in vitro embryo production media by NPPC and sildenafil affect the cytoplasmic lipid content and gene expression of bovine cumulus-oocyte complexes and embryos. Reprod. Biol. 2018, 18, 66–75. [Google Scholar] [CrossRef]
- Seneda MM, Esper CR, Garcia JM, de Oliveira JA, Vantini R. Relationship between follicle size and ultrasound-guided transvaginal oocyte recovery. Anim. Reprod. Sci. 2001, 67, 37–43. [Google Scholar] [CrossRef]
- Vajta G, Holm P, Greve T; et al. The Submarine Incubation System, a new tool for in vitro embryo culture. A technique report. Therio. 1997, 48, 1379–85. [Google Scholar] [CrossRef]
- Zhang JY, Diao YF, Oqani RK, Han RX, Jin DI. Effect of endoplasmic reticulum stress on porcine oocyte maturation and parthenogenetic embryonic development in vitro. Biol. Reprod. 2012, 27, 128. [Google Scholar]
- Park HJ, Park JY, Kim JW, Yang SG, Jung JM, Kim MJ, Park JJ, Koo DB. Regulation of the Endoplasmic Reticulum Stress by BIP/GRP78 is involved in Meiotic Maturation of Porcine Oocytes In Vitro. Dev. Reprod. 2017, 21, 407–15. [Google Scholar] [CrossRef]
- Mochizuki M, Miyagi K, Kishigami S. Optimizing treatment of tauroursodeoxycholic acid to improve embryonic development after in vitro maturation of cumulus-free oocytes in mice. PLoS ONE. 2018, 27, 13–e0202962. [Google Scholar]
- Takahashi M, Keicho K, Takahashi H, Ogawa H; et al. Effect of oxidative stress on development and DNA damage in in-vitro cultured bovine embryos by comet assay. Therio. 2000, 54, 137–45. [Google Scholar] [CrossRef] [PubMed]
- Xie Y, Wang F, Puscheck EE, Rappolee DA. Pipetting causes shear stress and elevation of phosphorylated stress-activated protein kinase/jun kinase in preimplantation embryos. Mol. Reprod. Dev. 2007, 74, 1287–94. [Google Scholar] [CrossRef]
- Lin T, Zhang JY, Diao YF, Kang JW; et al. Effects of sorbitol on porcine oocyte maturation and embryo development in vitro. Zygote 2015, 23, 297–306. [Google Scholar] [CrossRef] [PubMed]
- López-Damián EP, Jiménez-Medina JA, Alarcón MA, Lammoglia MA, Hernández A, Galina CS, Fiordelisio T. Cryopreservation induces higher oxidative stress levels in Bos indicus embryos compared with Bos taurus. Therio 2020, 143, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Song BS, Kim JS, Yoon SB, Lee KS; et al. Inactivated Sendai-virus-mediated fusion improves early development of cloned bovine embryos by avoiding endoplasmic-reticulum-stress-associated apoptosis. Reprod. Fertil. Dev. 2011, 23, 826–36. [Google Scholar] [CrossRef] [PubMed]
- Sharma A, Agrawal H, Mullani N, Sandhu A; et al. Supplementation of tauroursodeoxycholic acid during IVC did not enhance in vitro development and quality of buffalo IVF embryos but combated endoplasmic reticulum stress. Therio. 2015, 84, 200–07. [Google Scholar] [CrossRef] [PubMed]
- Lin T, Lee JE, Oqani RK, Kim SY; et al. Tauroursodeoxycholic acid improves pre-implantation development of porcine SCNT embryo by endoplasmic reticulum stress inhibition. Reprod. Biol. 2016, 16, 269–78. [Google Scholar] [CrossRef]
- Wang J, Lee J, Liem D, Ping P. HSPA5 Gene encoding Hsp70 chaperone BiP in the endoplasmic reticulum. Gene. 2017, 618, 14–23. [Google Scholar] [CrossRef]
- Campanella C, Pace A, Caruso Bavisotto C, Marzullo P; et al. Heat shock proteins in Alzheimer’s disease: Role and targeting. Int. J. Mol. Sci. 2018, 19, 2603. [Google Scholar] [CrossRef]
- Harding HP, Zhang YH, Bertolotti A, Zeng HQ, et at. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell. 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Landau G, Kodali VK, Malhotra JD, Kaufman RJ. Detection of oxidative damage in response to protein misfolding in the endoplasmic reticulum. Methods Enzymol. 2013, 526, 231–50. [Google Scholar]
- Van der Vlies D, Makkinje M, Jansens A, Braakman I; et al. Oxidation of er resident proteins upon oxidative stress: Effects of altering cellular redox/antioxidant status and implications for protein maturation. Antioxid. Redox Signal. 2003, 5, 381–87. [Google Scholar] [CrossRef] [PubMed]
- Hausburg MA, Dekrey GK, Salmen JJ, Palic MR, Gardiner CS. Effects of paraquat on development of preimplantation embryos in vivo and in vitro. Reprod. Toxicol. 2005, 20, 239–46. [Google Scholar] [CrossRef] [PubMed]
- Kammoun HL,Chabanon H, Hainault, I; et al. “GRP78 expression inhibits insulin and ER stress-induced SREBP-1c activation and reduces hepatic steatosis in mice,”. The Jour Clin Invest. 2009, 119, 1201–15. [CrossRef] [PubMed]
- Zhang K, Wang S, Malhotra J; et al. “The unfolded protein response transducer IRE1α prevents ER stress-induced hepatic steatosis,”. The EMBO Journal. 2011, 30, 1357–75. [CrossRef]
- Lauressergues E, Bert E, Duriez P, Hum D, Majd Z; et al. Does endoplasmic reticulum stress participate in APD-induced hepatic metabolic dysregulation? Neuropharm. 2012, 62, 784–96. [CrossRef]
- Han J, Kaufman RJ. The role of ER stress in lipid metabolism and lipotoxicity. J. Lipid Res. 2016, 57, 1329–38. [Google Scholar] [CrossRef]
- Sudano MJ, Caixeta ES, Paschoal DM, Martins A; et al. Cryotolerance and global gene-expression patterns of Bos taurus indicus and Bos taurus taurus in vitro-and in vivo-produced blastocysts. Reprod. Fert. Develop. 2014, 26, 1129–41. [Google Scholar] [CrossRef]
- Harris D, Huang B, Rn Oback B. Inhibition of MAP2K and GSK3 signaling promotes bovine blastocyst development and epiblast-associated expression of pluripotency factors 1. Biol Reprod. 2013, 88, 74.
- Aksu DA, Agca C, Aksu S, Bagis H; et al. Gene expression profiles of vitrified in vitro- and in vivo-derived bovine blastocysts. Mol. Reprod. Dev. 2012, 79, 613–625. [Google Scholar] [CrossRef]
- Hua Y, Kandadi MR, Zhu M, Ren J, Sreejayan N. Tauroursodeoxycholic acid attenuates lipid accumulation in endoplasmic reticulum-stressed macrophages. J. Cardiovasc. Pharmacol. 2010, 55, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Cha BH, Kim JS, Ahn JC, Kim HC, Kim BS, Han DK, Park SG, Lee SH. The role of tauroursodeoxycholic acid on adipogenesis of human adipose-derived stem cells by modulation of ER stress. Biomaterials. 2014, 35, 2851–58. [Google Scholar] [CrossRef] [PubMed]
- Lee K, Tirasophon W, Shen X, Michalak M; et al. IRE-1-mediated unconventional mRNA splicing and S2P-mediated ATF6 cleavage merges to regulate XBP1 in signaling the unfolded protein response. Genes. Dev. 2002, 16, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Hosoi T, Ogawa K, Ozawa K. Homocysteine induces X-box-binding protein 1 splicing in the mice brain. Neurochem. Int. 2010, 56, 216–20. [Google Scholar] [CrossRef]
- Wild AC, Moinova HR, Mulcahy RT. Regulation of gamma-glutamyl- cysteine synthetase subunit gene expression by the transcription factor Nrf2. J Biol Chem 1999, 274, 33627–36. [CrossRef]
- Tanaka Y, Aleksunes LM, Yeager RL, Gyamfi MA; et al. NF-E2-related factor 2 inhibits lipid accumulation and oxidative stress in mice fed a high-fat diet. J. Pharmacol. Exp. Ther. 2008, 325, 655–64. [Google Scholar] [CrossRef]
- Moreira S, Fonseca I, Nunes MJ; et al. Nrf2 activation by tauroursodeoxycholic acid in experimental models of Parkinson’s disease. Experimental Neurology. 2017, 295, 77–87. [Google Scholar] [CrossRef]
- Yang HC; et al. Glucose 6-phosphate dehydrogenase deficiency enhances germ cell apoptosis and causes defective embryogenesis in Caenorhabditis elegans. Cell death & disease. 2013, 4, e616. [Google Scholar]
- Wu YH, Lee YH, Shih HY, Chen SH; et al. Glucose-6-phosphate dehydrogenase is indispensable in embryonic development by modulation of epithelial-mesenchymal transition via the NOX/Smad3/miR-200b axis. Cell Death Dis. 2018, 9, 10. [Google Scholar] [CrossRef]
- Purcell SH, Moley KH. Glucose transporters in gametes and preimplantation embryos. Trends Endocrinol. Metab. 2009, 20, 483–489. [Google Scholar] [CrossRef]
- Ortega MS, Kelleher AM, O`Neil E, Benne J; et al. NANOGis required to form the epiblast and maintainpluripotency in the bovine embryo. Mol. Reprod. Develop. 2020, 87, 152–160. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of TUDCA concentrations on the percentage blastocyst. Results were least-squares means ± SEM. Different letters in each bar represent significant difference (P≤0.1). Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
Figure 1.
The effect of TUDCA concentrations on the percentage blastocyst. Results were least-squares means ± SEM. Different letters in each bar represent significant difference (P≤0.1). Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
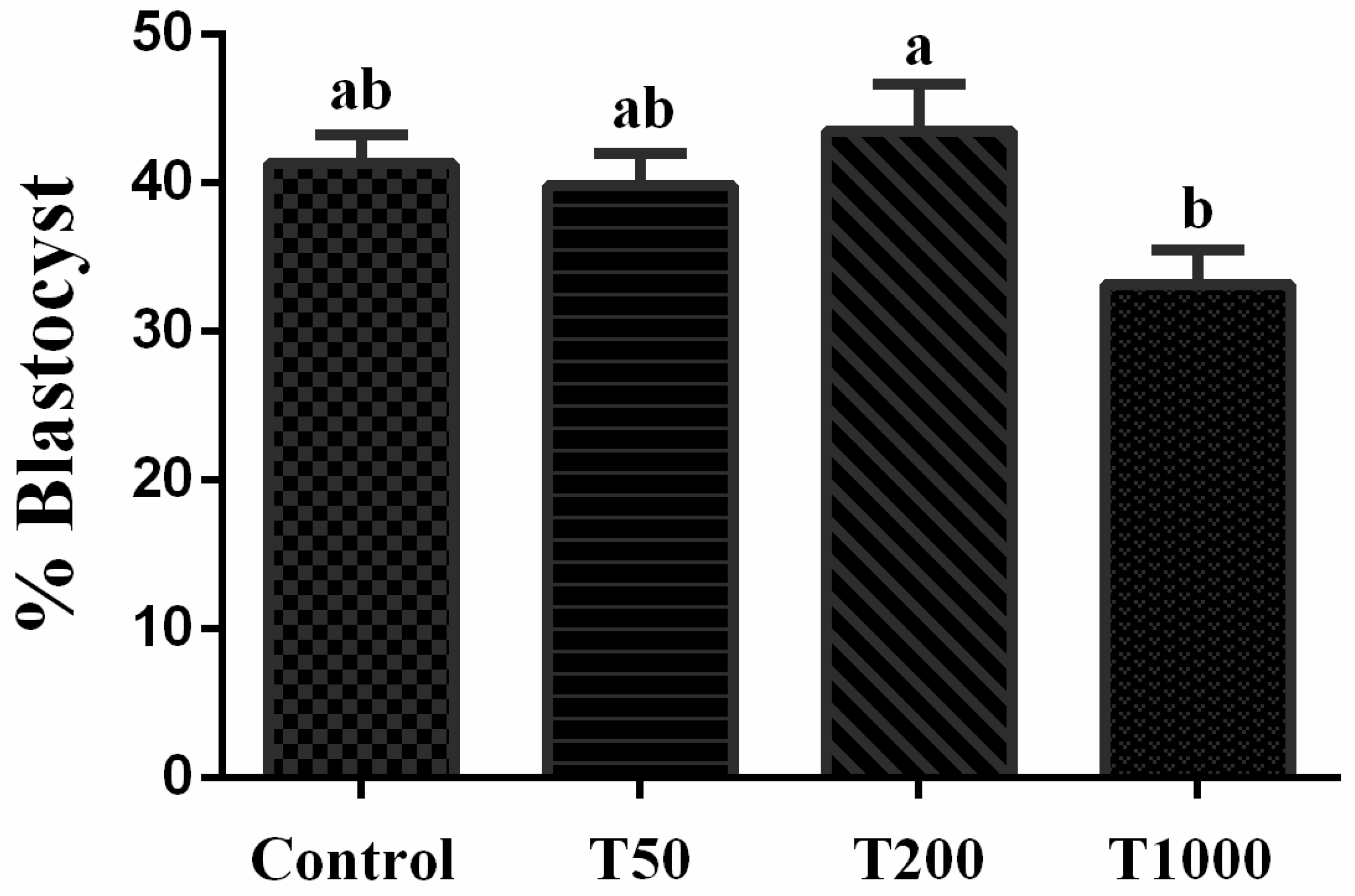
Figure 2.
The effect of TUDCA concentrations on the blastocyst hatching kinetics. Results were least-squares means ± SEM. *P≤0.1. Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
Figure 2.
The effect of TUDCA concentrations on the blastocyst hatching kinetics. Results were least-squares means ± SEM. *P≤0.1. Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
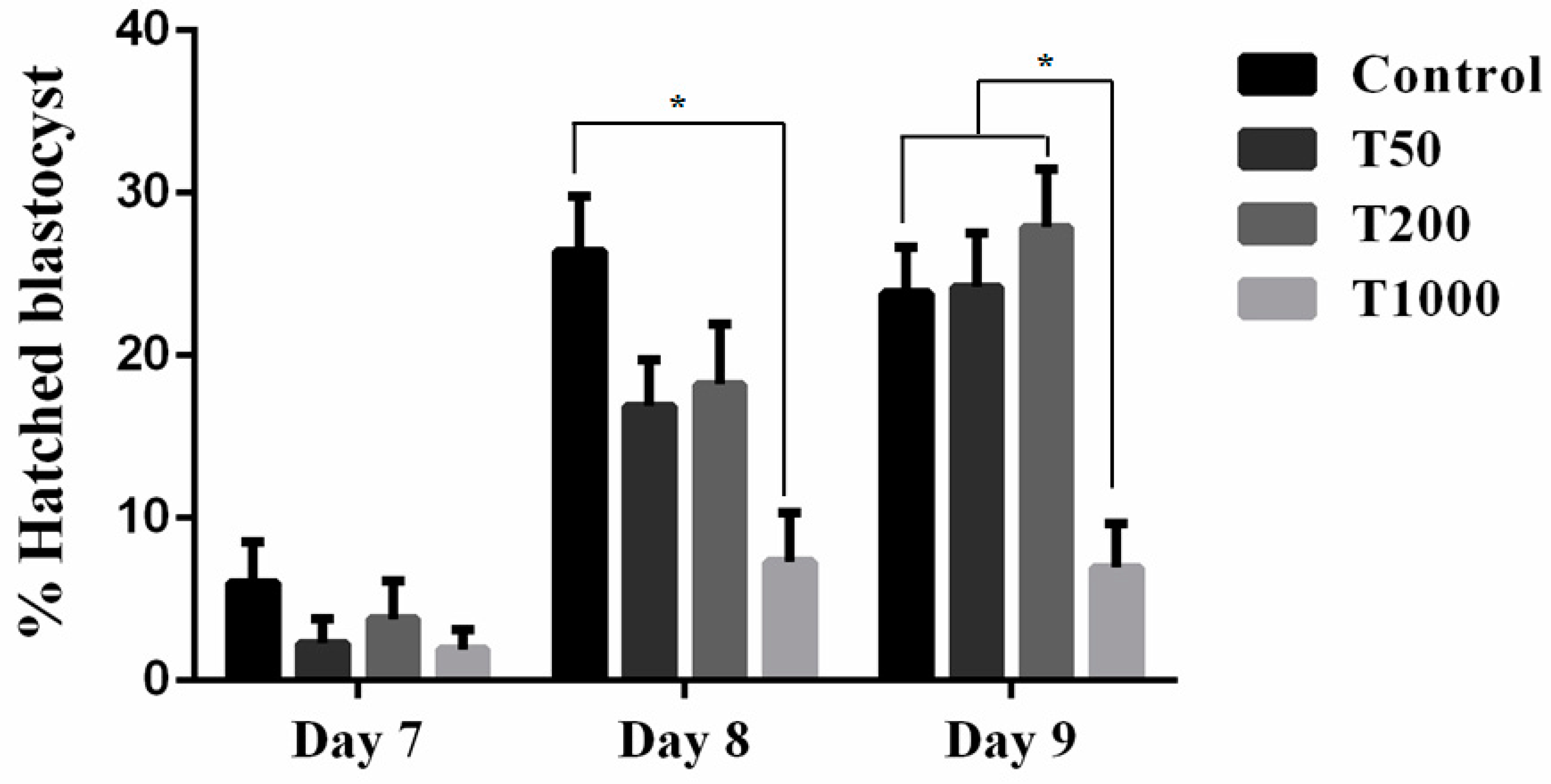
Figure 3.
The effect of TUDCA concentrations on mRNA abundance of differentially expressed genes in the embryo according to the following functional categories: endoplasmic reticulum stress (A), oxidative stress and response to cellular stress (B), related to metabolism (C), and related to the embryo development and cell proliferation (D). Results were least-squares means ± SEM. Different letters in each bar represent a significant difference (P≤0.1). Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
Figure 3.
The effect of TUDCA concentrations on mRNA abundance of differentially expressed genes in the embryo according to the following functional categories: endoplasmic reticulum stress (A), oxidative stress and response to cellular stress (B), related to metabolism (C), and related to the embryo development and cell proliferation (D). Results were least-squares means ± SEM. Different letters in each bar represent a significant difference (P≤0.1). Control = 0 µM of TUDCA; T50 = 50 µM of TUDCA; T200 = 200 µM of TUDCA; T1000 = 1000 µM of TUDCA.
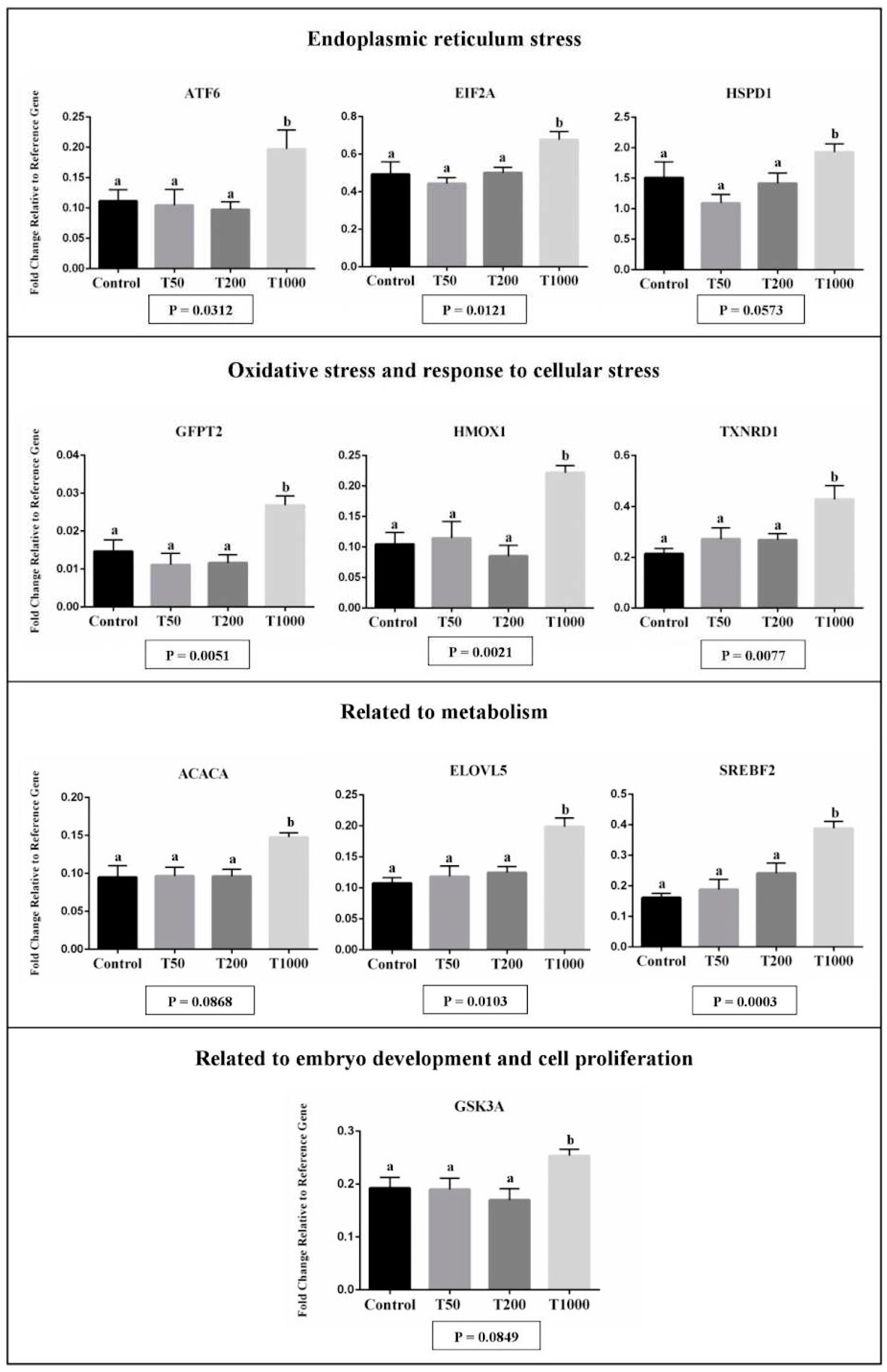
Figure 4.
The effect of TUDCA concentrations on mRNA abundance of differentially expressed genes in the embryo according to the following functional categories: endoplasmic reticulum stress (A), oxidative stress and response to cellular stress (B) and related to metabolism (C). Results were least-squares means ± SEM. The effect of treatment was P≤0.1 for all genes in the figure. Control = 0 µM; T200 = 200 µM...
Figure 4.
The effect of TUDCA concentrations on mRNA abundance of differentially expressed genes in the embryo according to the following functional categories: endoplasmic reticulum stress (A), oxidative stress and response to cellular stress (B) and related to metabolism (C). Results were least-squares means ± SEM. The effect of treatment was P≤0.1 for all genes in the figure. Control = 0 µM; T200 = 200 µM...
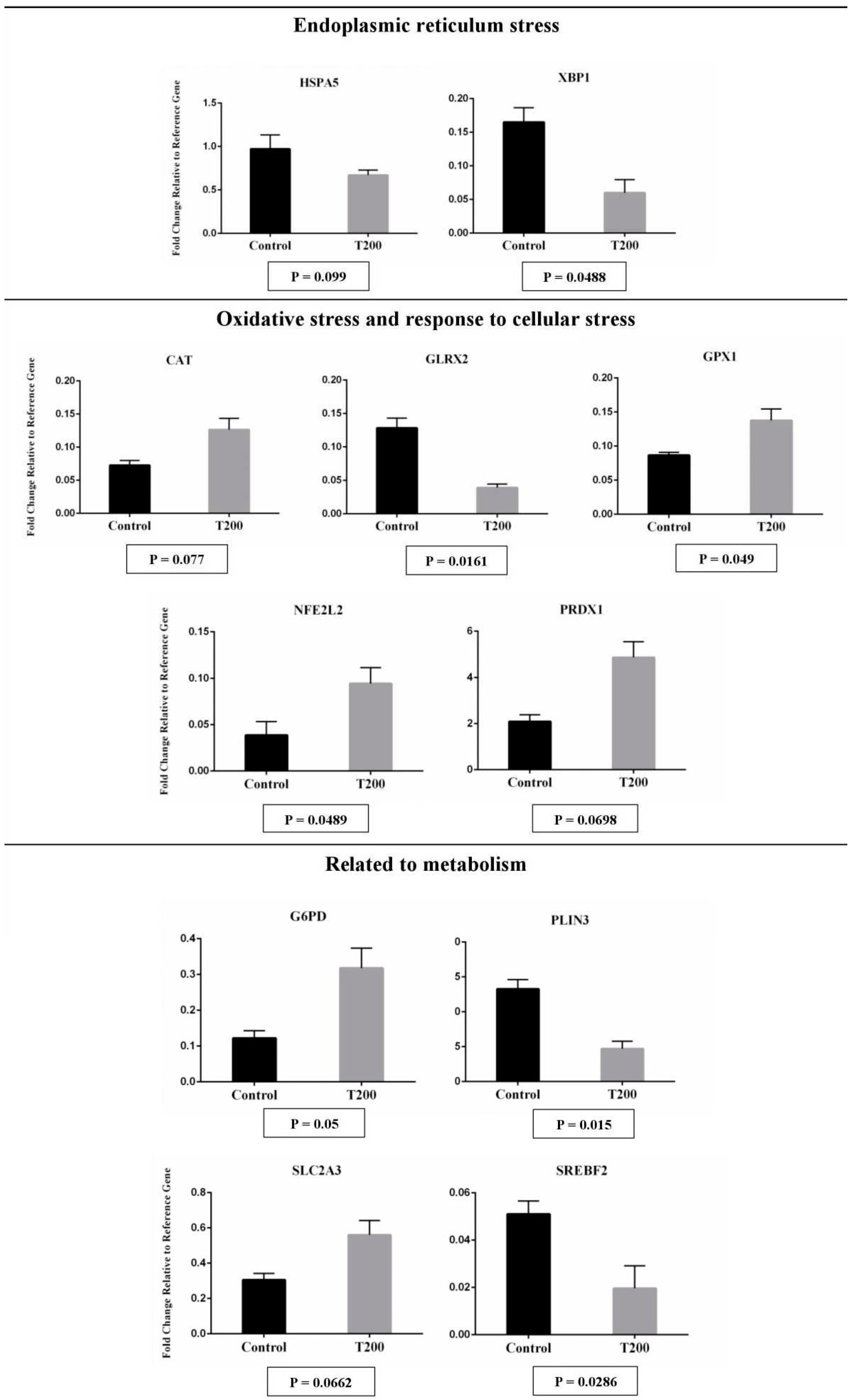
Figure 5.
STRING network of protein-protein interactions between genes with significant difference (P≤0.1) of mRNA abundance among groups. Different line colors represent the relationship between the proteins translated from analysed genes, as described on the legend.
Figure 5.
STRING network of protein-protein interactions between genes with significant difference (P≤0.1) of mRNA abundance among groups. Different line colors represent the relationship between the proteins translated from analysed genes, as described on the legend.
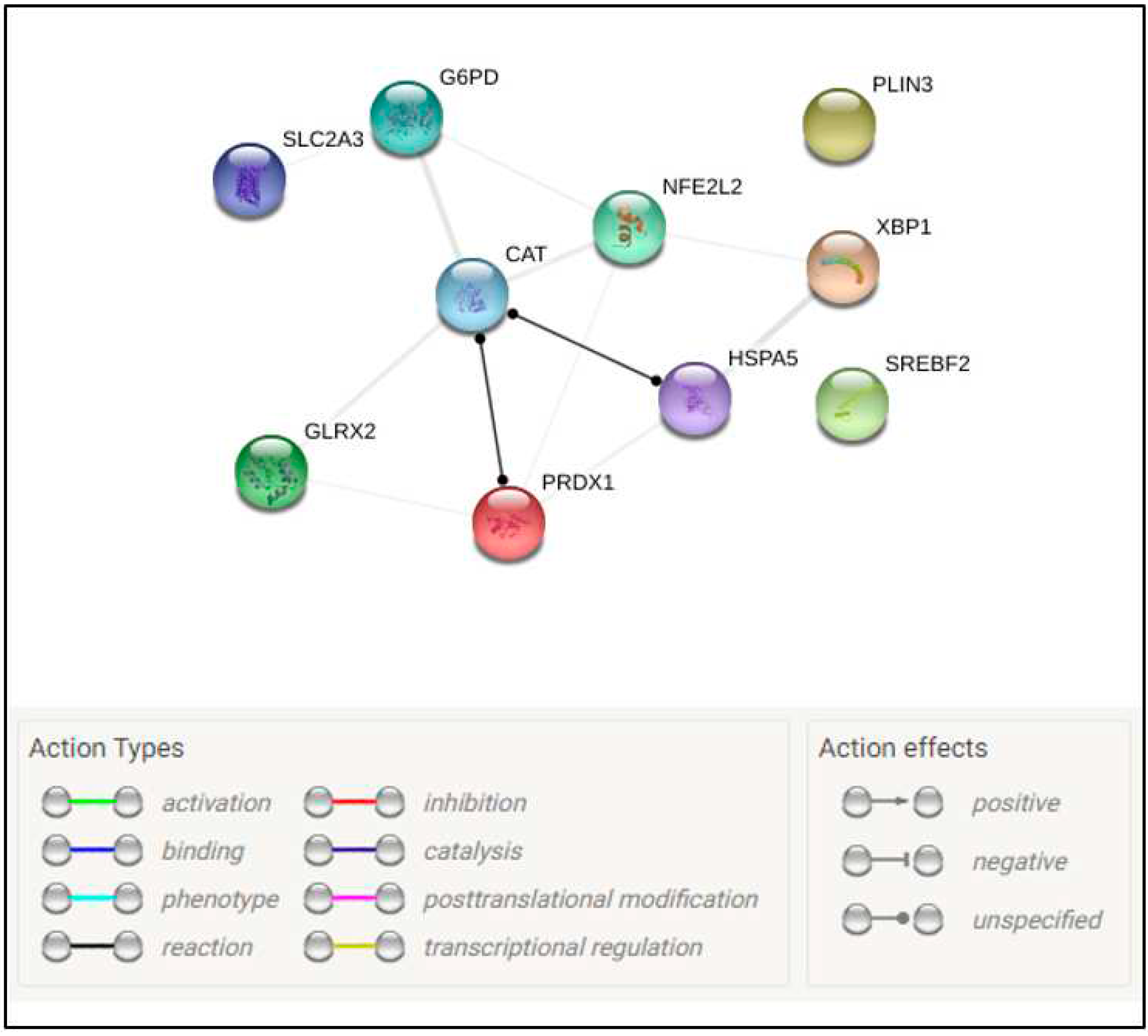

Table 1.
Re-expansion and hatching rate of embryos after vitrification on Days 7 and 8 (only expanded blastocysts) from groups Control or T200 (200 μM of TUDCA). Total of five replicates (mean ± standard error mean).
Table 1.
Re-expansion and hatching rate of embryos after vitrification on Days 7 and 8 (only expanded blastocysts) from groups Control or T200 (200 μM of TUDCA). Total of five replicates (mean ± standard error mean).
 |
Abbreviations: Re-exp.: re-expansion; Hatch.: hatching. Different letters represent a significant difference
(P≤0.1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



