
Submitted:
25 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
The discovery of microRNAs and their role in disease today represents a substantial breakthrough that has inspired and propagating research on microRNAs as targets for diagnosis and therapy. Cardiovascular disease is an area where the limitations of early diagnosis and conventional pharmacotherapy are evident and deserve attention. Thus microRNA-based drugs have substantial potential for development. Research and its application can progress considerably as witnessed in preclinical and clinical trials. The use of miRNAs remains experimental at the time of writing but has a promising role in the diagnosis and prognosis of a variety of acute coronary syndrome presentations. Its use, alone or in combination with currently available biomarkers, could be adopted soon, especially if diagnostic ambiguity exists. In this review, we explore the current state of knowledge about microRNAs as potential targets for diagnosis and therapy in the cardiovascular system. We report recent improvements in the acknowledgment and characterization of microRNAs focusing on clinical translation. The new dares and outlooks toward clinical application are discussed.
Keywords:
MicroRNA
; miRNA biomarker
; microRNAs functions
; acute coronary syndrome
; coronary artery disease
Keywords: MicroRNA; miRNA biomarker; microRNAs functions; acute coronary syndrome; coronary artery disease
1. Introduction
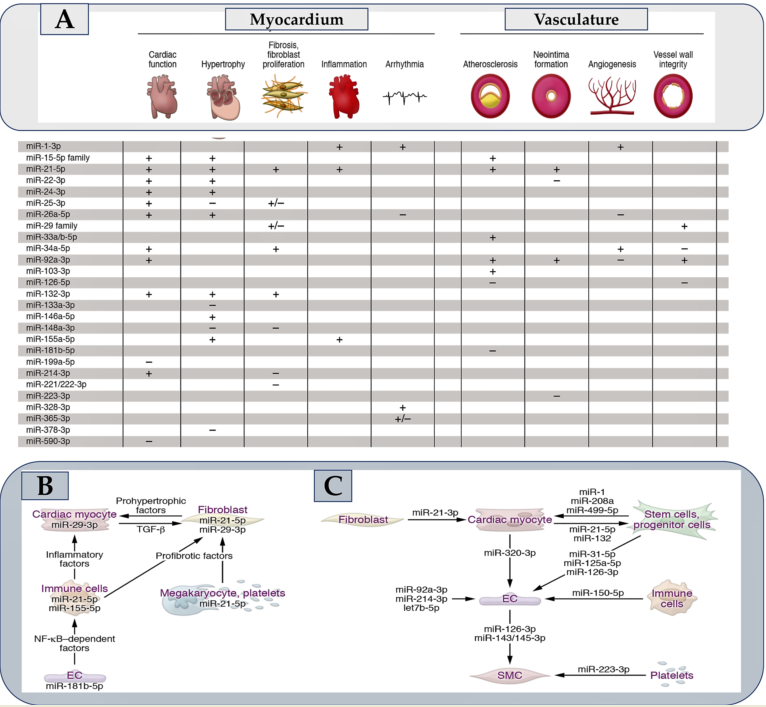
The discovery of a small non-coding RNA in Caenorhabditis elegans with a regulatory function in gene expression at the post-transcriptional level dates back three decades [1,2]. In higher eukaryotes, the subsequent advances in molecular biology applied to the non-coding RNA field have led to the identification of numerous microRNAs, also called miRNAs. Hence, the discovery that they regulate most mammalian mRNAs was a gamble that paid off [3]. However, the questions regarding the number of microRNAs that are expressed in humans remain an open window to this day. Starting in 1973 and following the annotations reported from time to time in the mirBase 22.1 [4] of human microRNAs, it has been observed that many do not resist treatment due to strict criteria such as expression, sequence constraints, or evidence of productive processing of the precursors. This concern has resulted in a significant numerical fluctuation for functional microRNAs in humans that seems to oscillate from 556 (mir-GeneDB 2.0) [5] to 758 [6]. Another non-negligible aspect concerns the fraction of functionally relevant microRNAs that depends on a sufficiently high expression in the tissue. Thus, the fraction of functional microRNAs can be further reduced so that metric disparities in percentages can affect function. It is optimistically possible to hypothesize, albeit on entirely provisional certainties, a critical role in the cardiovascular system of up to 150 microRNAs. Analysis of 30-35 microRNAs have been validated. In vivo experimental models have also been comprehensively performed highlighting the clinical role of many of these candidates and their diagnostic and therapeutic potential. Figure 1 Furthermore, equally promising predictions concern many other candidate microRNAs, implicated in the diagnosis and therapy of cardiovascular diseases.
Although polymorphisms in microRNA biogenesis factors, microRNA genes, or microRNA response elements (MREs) have emerged from the Genome Wide Association Study (GWAS) [49], the pathophysiological consequences have been resolved in a select few. In contrast to this limited knowledge, a wealth of microRNAs that are deregulated or modified in disease can be highlighted. The central question is about the deregulation of a microRNA. Does it cause disease or merely indicate it? MicroRNAs that are actively implicated in disease pathophysiological processes are often associated with elevated baseline steady-state expression, pronounced deregulation in disease (Figure 1), and the preferential presence of both in cells/tissue. For example, subjects with a transverse aortic constriction (TAC) model of ventricular pressure overload have miR-21-5p as the most abundant microRNA in cardiac macrophages. This miRNA is also upregulated 7-fold in the myocardium [50]. Another example concerns miR-29b-3p which is highly expressed in cardiac myocytes and upregulated approximately 3-fold in the presence of TAC [11]. Distinct pathophysiological effects in the myocardium or vasculature following manipulation (Figure 2A) are induced by many of the 30-35 microRNAs that have substantially demonstrated critical cardiovascular roles. Figure 1A fraction of these has a pathophysiological action dependent on the involvement of signaling pathways leading to the secretion of protein factors (Figure 2B), while for others, the action is specifically exosome-mediated as they are themselves secreted in extracellular vesicles (Figure 2C). The impetus due to the growth in the knowledge of microRNAs has gone hand in hand with the increase in the therapeutic development of these molecules in the myocardium and the vascular system [51] Figure 1 and Figure 2.
The aim of this review is to highlight the action of some of these candidate miRNAs in relation to their cardiovascular role and their potential clinical development in the diagnosis and therapy of cardiovascular diseases. Although word count constraints prevent us from a detailed discussion of all microRNAs in Table 1, we address an overview of the roles of microRNAs in the cardiovascular system by focusing on the role of microRNAs as circulating biomarkers in acute coronary syndromes.
2. MicroRNAs Pathophysiologic Surroundings: Insight on Biogenesis, Stability, and Strand Bias of microRNAs
MiRNAs are composed of non-coding RNA sequences that assemble ~22-24 nucleotides. The leading functions exercised concern the fine-tuning of gene expression, the control of tissue growth, and in the government of homeostasis [59,60]. As far as the regulation of gene expression is concerned, the intervention of miRNAs is not only directed towards the cell of origin but operate as main players in the governance of intercellular communication. During the first step, miRNAs are generated starting from pre-miRNA whose synthesis is advocated by the enzyme RNA polymerase II operating on DNA substrate. Subsequently, various enzymes participate in the modifications of the primordial structure of the miRNA and its delivery to the cytoplasm. Here, mature miRNA constitutes the RNA-induced silencing complex (RISC) with Dicer, Argonaute 2, and the transactivation response RNA-binding protein [61,62]. Lastly, miRNAs tie the 3’ untranslated regions of target messenger RNAs (mRNAs) across their 8-nucleotide seed sequence. Semi-complementarity of mature miRNA with target mRNA promotes translational repression, whilst full complementarity of mature miRNA with target mRNA assists mRNA degradation [63,64,65]. Although the pairing of accessories beyond the seed sequence can promote target recognition, only a fraction of microRNAs appears to be dependent on these interactions [66,67,68]. MicroRNA target sites in mRNA are found as microRNA response elements (MREs) primarily within the 3′-UTR and less frequently found in the 5′-UTR or coding regions [66,69]. Unlike the function of lncRNAs or circRNAs, for which different mechanisms of action have been highlighted, microRNAs have two clearly defined activities. the dominant activity is to induce degradation or as a secondary activity promote the translational silencing of target mRNAs [66].
The microRNA folder is supplemented by non-genetic variants, termed isomiRs, which result from different microRNA processing, nucleotide addition, or editing [70,71]. Many cardiovascular isomiRs are recognized, refs. [72,73] with levels fluctuating in disease [72]. For miR-487b-3p and miR-411-5p R isomes, distinct variants and model targetomes were detected [74,75]. The genesis of a microRNA corresponds to its enzymatic degradation at the end of the life cycle. Most microRNAs have been shown to have a significantly longer half-life than mRNAs, as well as considerable variability, depending on the microRNA strand and sequence, cell type, and transaction factors (Figure 1A) [76,77]. Among the latter, targets of microRNAs have also been identified. Although the mechanistic features of target-directed microRNA degradation (TDMD) have been settled [63,78,79] and the meaning of TDMD is well demonstrated in vivo [80], it is difficult to identify mRNAs that engage in TDMD.
miRNAs are decisive for embryonic growth, regular physiology, and function of the cardiovascular apparatus [81,82,83]. Cellular expression standards of miRNAs are modified in assorted cardiovascular disorders which include MI, cardiac hypertrophy, and failure. In addition, hypertension, obesity, hyperlipidemia, and diabetes promote higher levels of miRNAs [83,84,85,86]. A variety of experimental models of atherosclerosis were assessed focusing on the miRNAs’ function. A considerable role of different miRNAs has been advocated in analyzing in vitro and in vivo processes of atherogenesis. Specifically, this includes the elicited phenomena of inflammation, endothelial cell activation, angiogenesis, vascular smooth muscle cell proliferation, migration, and neointima formation have been related to increased miRNA expression [86,87].
The mechanism of active and selective secretion of miRNAs in living cells has been widely described as how this specific function contributes to cell-tissue communication [88,89,90]. The conveyance of secreted miRNAs is ensured by extracellular vectors, which involve specific molecular assets such as Argonaute 2, nucleophosmin, and lipoproteins. Again, miRNAs can alternatively be upregulated into extracellular macrovesicles and subsequently reversed into recipient cells where they alter gene expression [88,91,92]. From these observations, it follows that, contrary to mRNA turnover, miRNAs are molecules that persist in a stable frame for the time of the permanence in the circulation, with optimal advocated protection from endogenous ribonuclease activity [88,89,92]. These findings strengthened the notion that circulating miRNAs could have the potentially functional role of biomarkers capable of detecting the different entities of ACS. Rather, many reports are available that have assessed the diagnostic perspective of miRNAs in these clinical settings. Figure 3.
3. Searching Roles of microRNAs in the Cardiovascular System
Several studies have demonstrated a strong upregulation of miR-21-5p in the weakened human heart [12]. The close correlation of miR-21-5p with the fibrosis process has suggested an upregulation of this molecule also in renal and pulmonary diseases in which fibrotic degeneration is shared as a common denominator. In animal models, miR-21 inhibitors have been observed to work to prevent cardiac fibrosis [12] or neointima formation [13]. While a global miR-21-5p deficiency remained undetected [33,81], the effects of the inhibitors were reiterated by a genetic knockout of miR-21 in non-myocyte cells, [14] thereby pointing out a crucial role in them. Cardiac macrophages and fibroblasts record substantially increased miR-21-5p levels [14,50]. Macrophage-specific miR-21-5p deficient mice were resistant to transverse aortic constriction induced by structural and functional phenotypes, associated with reduced inflammatory phenomena. [50] Steadily, pigs receiving antimiR-21 after ischemia/reperfusion demonstrated improved cardiac function and reduced triggering inflammation [9]. The set of results reported on the role of miR-21-5p in the myocardium gives it a strong profibrotic and proinflammatory function. In accordance, LNA-antimiR-21 is presently undergoing specific evaluations reported in a phase II study for the treatment of fibrotic renal disease.
The miR-29 is considered a family of four nearly identical variants. It has been suggested that it regulates collagen and other matrix proteins, thus making it a designated target for antifibrotic therapies. von Roji and colleagues in a landmark study on miR-29 reported a repressive function exerted on collagen associated with an improvement in cardiac function was offered by miR-29 mimics [15]. This idea has since been reiterated in other organs, resulting in a finale progress of a miR-29 mimic (MRG-201) that is effective in the treatment of idiopathic pulmonary fibrosis. This principle, is, however, aimed at a reduction of collagen expression, antimiR-29b favored the stabilization of the vascular wall subjected to structural modifications that generated abdominal aortic aneurysms in mouse models [16,17]. In contrast to these findings, Sassi and colleagues observed that miR-29 inhibition, rather than elevation, prevents cardiac fibrosis [11]. This unexpected and surprising result surprised experts in the field, but its underlying mechanism was elucidated by the authors. Each of the miR-29 variants is predominantly expressed in cardiac myocytes with high levels in cardiac fibroblasts that are recorded only with prolonged cultivation. The main profibrotic pathophysiological mechanism within the fibroblasts deployed by miR-29 results in the induction of the Wnt pathway for cell hypertrophy and paracrine signaling, Hence, miR-29 inhibition appears appropriate in the myocardium, whereas its elevation is useful for suppressing fibrotic pathways in fibroblasts in patients with skin diseases.
Two independent studies [21,24] explored the elevated expression of miR-92a-3p in endothelial cells and deregulation in mouse models with developed myocardial and vascular tissue lesions. A functional LNA antimiR against miR-92a was shown to promote angiogenesis and tissue repair in these models [24], which was subsequently corroborated in a study based on a porcine ischemia/reperfusion model [21]. These data have been translated, with a first step towards clinical application, following the results reported in a pharmacological study on antimiR-92a, called MRG-110. The enrolled cohort consisted of healthy individuals who received a single drug by intravenous injection [97]. Importantly, given the efficacy of antimiR-92a after intradermal inoculation, which was also found in animal models of skin injury, a second phase I clinical study using this route of administration was designed (ClinicalTrials.gov NCT03603431).
Upregulation of miR-155-5p in immune cells has been reported in patients experiencing cardiac inflammation or in respective animal models [23,26]. Bone marrow transplantation experiments in mice suggested that the proinflammatory activity of miR-155 is linked to macrophages [91], in which NF-κB expression is enhanced. Instead, an increase in miR-146a-3p expression is opposed to this effect [27]. Although miR-155 inhibition improves cardiac inflammation in mice [23,26], the notion of macrophage-specific miR-155 deficiency favoring an arrest of the arteriogenesis process after vascular injury [28] needs to be further analyzed to support these initial findings. Cobomarsen, an antimiR against miR-155, progressed to a phase I study in cutaneous T-cell lymphoma (ClinicalTrials.gov NCT02580552) [98], but a phase II study was halted due to speculative interest in cardiovascular research. Since the clinical data collected would be valuable, they can be used to guide the development of antimiR-155 for cardiovascular therapy.
MiR-132-3p has passed the preclinical phase and moved into the clinical one progressing in efficacy, safety, and benefits very quickly. The most surprising data concerned the genetic deficiency of the miR-132/-212 cluster or of an antagomir against miR-132-3p that prevented TAC-induced pathological cardiac remodeling [25]. Based on these observations, an evaluation of miR-132-3p inhibition was performed in mouse models of heart failure and blood pressure overload was induced [22,25]. In another study using a porcine model of heart failure, authors demonstrated a long persistence within cardiac tissue (t1⁄2 of 3 weeks) of miR-132 with an advantageous safety profile. The authors also validated the derepression of miR-132 targets [22]. Again, in pig models of induced myocardial infarction, the use of antimiR-132 improved cardiac function after myocardial injury [99]. In another pig model, the same effects were achieved to prevent chronic pressure overload [100]. A first study performed in humans, with dose-escalation (phase Ib) in patients with heart failure revealed good tolerability and the first evidence of a therapeutic benefit [101].
4. MicroRNAs as Circulating Biomarkers in Acute Coronary Syndromes
A report by the World Health Organization validated by the Center for Disease Control and Prevention (CDS) has indicated coronary atherosclerotic disease (CAD) and acute coronary syndrome (ACS) alongside its major cause of complications, as the main culprits for morbidity and mortality worldwide [102,103,104]. The diagnostic classification of ACS goes beyond the generic term so that myocardial infarction (MI) is manifested by electrocardiographic signs of elevation or depression of the ST segment (STEMI/NSTEMI) or as unstable angina pectoris (UA). ACS includes a heterogeneous patient population, of which only 50% are admitted to the hospital with chest pain having a cardiac origin [105,106,107,108,109]. This significant heterogeneity, which dominates the cohorts of patients with acute chest pain, has raised concerns to identify highly sensitive and specific diagnostic tools capable of directing toward a precise diagnosis, avoiding the potential onset of complications, and carrying out a precise risk stratification. The clinical presentation, the electrocardiogram, and specific cardiac biomarkers are the cornerstone tools on which the diagnosis of acute forms of ACS is based. The most reliable test for myocardial infarction (MI) and myocardial necrosis is based on serial measurements of serum troponin levels [110]. Although the procedure represents the current standard for diagnosis, it nevertheless retains limitations related to the delay with which the troponin increase occurs. This gap of 1-2 hours, which allows the exclusion of myocardial infarction through the possibility of accessing an early diagnosis, is today the most important clinical challenge to face. Therefore, the need to practice serial measurements during the downtime for the diagnosis of MI, during the prolonged observation of patients in emergency medical and/or cardiology facilities, is the primary factor driving the search for new rule-in and rule-out strategies needed for early detection of MI [111,112]. The benefits associated with using miRNAs as biomarkers were established in a landmark setting by Mitchell and colleagues nearly 14 years ago [113]. The improvement in diagnosis using miRNAs is almost certainly because they are short, non-coding, post-transcriptional regulators of gene expression with remarkable stability in circulation [113]. The diagnostic efficacy of these molecules has been extensively studied and their utility as biomarkers has been proposed for the diagnosis of various diseases including aortic dissection, in which human cytomegalovirus-encoded microRNAs expression profile was found in the plasma of patients [84,85,113,114,115,116,117]. Figure 4.
5. Insight into the Diagnostic and Prognostic Value of Circulating microRNAs in Coronary Artery Disease. Evaluation of the New Disease Diagnosis Guide for Stable CAD and Acute Coronary Syndrome
Coronary artery disease is the most common manifestation of cardiovascular disease and acute coronary syndrome is related to considerable morbidity and mortality in a wider part of the population worldwide. Several biomarkers for the diagnosis of acute myocardial infarction are known and used. Among these, cardiac troponins are routinely employed as biomarkers in individuals who experience chest pain; however, it lacks sensitivity in the initial hours after the onset of symptoms, which obscures its full diagnostic validity. For this reason, the need to find new biomarkers for the diagnosis of coronary events is still a priority in research aimed at application in the clinical field. Current studies have suggested a substantial role of miRNAs in advocating atherosclerotic plaque generation and their expression is modified during the course of CAD occurrence. While disparate reports have supported the non-superiority of several miRNAs over troponin in the diagnosis of AMI, nevertheless their use may be directed in the early diagnosis and prognosis of patients with CAD. In this scenario, further studies are required. We have summarized recent studies investigating circulating miRNAs as novel biomarkers in CAD, AMI diagnosis, and in-stent restenosis. We also examined the role of miRNAs in CVD risk stratification as well as evaluated their role in the prognosis of patients with acute coronary syndrome [118,119,120,121,122].
5.1. MicroRNAs and Coronary Artery Disease
Discontinuous and relatively discordant results have been reported in different reports involving cohorts of patients with ACS occurring in the two clinical conditions of stable angina (SA) and unstable angina (UA) pectoris. The data that emerged supports a relatively subtle meaning and the absence of a significant diagnostic value capable of explaining the real validity and efficacy of miRNAs in the diagnosis of the various forms of ACS. The diagnostic benefits associated with the investigation of miRNAs as biomarkers in the early diagnosis of unstable angina (UA) were established in a landmark paper from the German Center for Cardiovascular Research 10 years ago [124]. The considerable albeit not conclusive results obtained from this study were because it was conducted in three phases with the enrollment of a large number of patients. In detail, 667 miRNAs were selected in the first screening phase by identifying three arms of patients. Acute coronary syndrome occurred as UA was disclosed in 10 subjects while non-cardiac chest pain (NCCP) in 10 others. The third cohort of patients encompassed 20 healthy subjects that were matched for sex and gender to those patients who exhibited UA. From the initial reference reservoir, screening identified 25 miRNAs interconnected to UA (p > 0.05), and to this, miR-1, miR-208a, and miR-208b were added. All these elements were evaluated in the second phase of replication for the final identification process. The second stage employed an independent patient cohort consisting of 49 UA and 48 NCCP patients. Eight miRNAs including miRNA-19a, miRNA-19b, miRNA-132, miRNA-140-3p, miRNA-142-5p, miRNA-150, miRNA-186, and miRNA-210, were considered critical to enter the final stage, due to their substantial commonality feature to be detected in patients presenting with UA in the first two stages. The third phase involved the validation and supported diagnostic capability of the miRNAs in UA subjects. Confirmation was achieved by examining forty-six UA and sixty-three NCCP patients from the first cohort. The baseline statistical asset was represented by their respective areas under the curve (AUC) which was calculated in patients with UA compared with patients with NCCP. It emerged that MiR-186 showed the best result (AUC=0.78). The second and third best-expressed miRNAs were miR-132 and miR-150, respectively reaching an AUC of 0.91 [124].
More research was conducted within the same period. The aim was to evaluate, in CAD patients compared to healthy controls, both the circulating miRNA profiles and to find the expression of useful miRNAs, thus suggesting correct discrimination between SA and UA pectoris [125]. The authors performed an initial screening of 367 miRNAs by recording key data of three miRNAs with higher expression levels in patients with coronary artery disease (CAD) than in those without the disease. An increase of miRNA-337-5p was reported only in SA, but not in UA. Instead, miRNA-433 and miRNA-485-3p were individually increased in both SA and UA compared to healthy controls. However, a substantial difference was not attributed to any of the three miRNAs regarding the clinical type of ACS hitherto supporting a specificity for SA or UA. The study was also directed at 14 previously investigated miRNAs that had advocated promising evidence. When comparing CAD patients and healthy controls, seven of these miRNAs were found to be dysregulated. The data emerged underlined for miRNA-1, miRNA-122, miRNA-126, miRNA-133a, miRNA-133b, and miRNA-199a substantially increased both in the presence of SA and UA, individually. Instead, miRNA-145 was significantly increased only in patients with UA. The statistical asset for AUC revealed predictive outcomes in the two groups analyzed. In the SA cohort of patients, three miRNAs had a predictive value > 0.85 compared to controls, with respective values of miRNA-1 = 0.918, miRNA-126 = 0.929, and miRNA-485-3p = 0.851. In the UA cohort, the respective AUC values for miRNA-1 = 0.92, miRNA-126 = 0.867 and miRNA-133a = 0.906 were found to be satisfactory (>0.85). The identifiable statistical asset in the AUC area was never considered effective when the two SA and UA arms were compared with the results not achieving diagnostic power. Although the combination of the three best miRNAs with an efficiency of 90.2% and 87.2% respectively could discriminate patients with SA and UA compared to non-pathological controls; however, between the two groups with CAD, the discriminatory capacity never exceeded 66% [125]. The discrepancy in the results appears evident when comparing the former study with that of Fichtlscherer et al., [126] in which the AUC area was assessed by correlating the two clinical conditions of CAD. It was observed that the levels of miR-17, miR-92a, miR-126, miR-145 and miR-155 were decreased in the CAD-SA group compared to healthy controls. Instead, miR-133a levels and miR-208a levels were increased, although the reported values did not suggest significance [126]. In patients with CAD, the diagnostic potential of miRNAs with altered expression, thus constituting a cluster, was evaluated. In the first phase, 13 patients with UA and 13 controls with chest pain who did not demonstrate atherosclerosis during PCI were identified and screened using miRNA profiles. The presence of thirty-four significantly deregulated miRNAs was observed in the UA group. The innovative data of this study concerned the differentiation of miRNAs into clusters vs belonging to a specific family. If some of these miRNAs were derived from gene loci close to each other; they were defined as clusters, such as the miR-106b/25/93 cluster and the miR-17/19b/20a/92a cluster. Instead, miR-21/ family 590-5p was classified as belonging to the same family as it shared 5′ seed sequences with other miRNAs. Out of a total of 34 identified miRNAs, seven (miRNA-106b, miRNA-25, miRNA-92, miRNA-21, miRNA-590-5p, miRNA-126*, and miRNA-451) were selected based on criteria referable to the high levels of circulating abundance, previous findings and cluster definition or same family membership. From a separate cohort 3 groups of subjects 45 patients with UA, 31 patients with AS, and 37 controls were identified [125].
Fichtlscherer et al. [126] and et Ren al. [127] first independently proposed the systematic determination of miRNA dosage, although they advocated substantial differences between acute coronary syndrome clinical settings. The most favorable target to demonstrate the emerging role of miRNAs was to find their up- or down-regulation. miRNA-17, miRNA-20a, miRNA-21, and miRNA-92a were downregulated in patients with CAD due to stable angina [128], while upregulation was recorded in patients with CAD due to unstable angina [127]. Thus, in unstable angina patients, all seven miRNAs were raised as compared to both stable angina and healthy controls even after adjustment for risk factors [59]. In this clinical condition, it is possible to hypothesize that a signature of the miRNA-106b/25 cluster, the miRNA-17/92a cluster, the miRNA-21/590-5p family, miRNA-126*, and miRNA-451 could be used as a biomarker for AU [127].
Recently a report by Su et al. [129] assessed circulating microRNAs (miRNAs) as non-invasive, specific, and sensitive biomarkers for diagnosing coronary artery disease. 203 patients who experienced CAD and 144 age-matched controls (126 high-risk controls and 18 healthy volunteers) were enrolled by using the direct S-Poly(T)Plus method. The advantage of this process provided a sturdy quantification associated with the free RNA extraction procedure, which made plainer strategies, reduced variations, in particular raised accuracy. Direct S-Poly(T)Plus was able to identify novel miRNAs expression profiles of CAD subjects and assess their clinical diagnostic usefulness. Protocol mandated permission to identify twelve differentially expressed miRNAs amongst CAD patients. Likewise, high-risk controls were screened, and the performances of these miRNAs were assessed in validation set-1 with 96 plasma specimens. Ultimately from these 12 miRNAs six, which included miR-15b-5p, miR-29c-3p, miR-199a-3p, miR-320e, miR-361-5p, and miR-378b, were verified in validation set-2 with a sensitivity of 92.8% and a specificity of 89.5%, and the AUC reached 0.971 (95% confidence interval, 0.948-0.993, P < 0.001) in a large cohort for CAD subjects’ diagnosis. These results promote the efficacy of the direct S-Poly(T) Plus method which could be used for the diagnosis of CAD by targeting 12 unique miRNAs. Moreover, plasma fractionation suggested that only small amounts of miRNAs were collected into EVs [129].
5.2. MicroRNAs and Acute Coronary Syndrome
This role was formally recognized in two seminal papers in which assorted separate cardiac and muscle-enriched miRNAs were evaluated and proposed as potential biomarkers of acute MI. [125,130] Subsequently following the emerging evidence, in a larger study dictated to establish the role of miRNAs in the diagnosis of MI, the levels of miRNA-208b and miRNA-499, as well as high sensitivity troponin (hs-Tn), were measured in 510 MI patients and 87 healthy controls [131]. Davaux et al. [131] suggested that both miRNAs were significantly increased in MI patients compared with controls. For miRNA-499 the statistical asset relative to the AUC area was equal to 0.97 when healthy subjects were compared with those who exhibited IM; however, greater diagnostic accuracy was not attributed to the emerged data, compared to the determination of the hs-Tn alone. If in patients in whom a diagnosis of myocardial infarction was erroneously established, and retrospective addition of miRNA-499 was subsequently performed to correct the evaluation, the diagnosis did not substantially improve [131]. A relatively low sensitivity and specificity of miRNAs compared to cTn measurement has the effect of weakening the diagnostic potential of miRNAs. The expression of miRNA-1, miRNA-133a, miRNA-208b, miRNA-499, and cTnT were analyzed in 67 patients with acute MI while 32 constituted the control group of healthy volunteers [132]. In patients with acute MI it was noted that of the four miRNAs analyzed, all were markedly increased within 12 hours of the onset of chest pain. Controls performed on day 14 after acute myocardial injury showed equivalent miRNA levels between groups. Evidence suggested that for all these miRNAs a useful role as biomarkers for AMI could be considered even if lower sensitivity and specificity distinguished the measurement of miRNAs from that of cTnT [132].
A meta-analysis pooled data from these two previously reported studies [132,133] to compare the potential of microRNAs as biomarkers in patients experiencing MI versus values expressed in healthy subjects. In detail, 192 full-text articles were screened from which 19 articles were included in the final analysis. In 15 of these, the 4 miRNAs most frequently reported in the articles were extrapolated and then a detailed evaluation of the studied miRNAs was performed. miRNA-499 was evaluated in 8 studies, miRNA-1 in 7 studies, miRNA-208b in 6 studies, and miRNA-133a in 4 studies, respectively and providing a statistical asset with interesting results. miRNA-208b had a total sensitivity of 0.78 (95% CI: 0.76-0.81), a specificity of 0.88 (95% CI: 0.84-0.91), a diagnostic OR of 48.63 (95% CI: 14.60-162.01) and an AUC value of 0.8965, with P values > 0.01. As for miRNA-1 values, authors disclosed an overall sensitivity of 0.63 (95% CI: 0.59–0.66), a specificity of 0.76 (95% CI: 0.71–0.80), a diagnostic OR of 11.13 (95% CI: 4.09–30.26). The area AUC value was 0.8519 with P values > 0.01. miRNA-133a had a total sensitivity of 0.89 (95%CI: 0.83–0.94; P = 0.0047), specificity of 0.87 (95%CI: 0.79–0.92; P = 0.0262), a diagnostic OR of 54.40 (95%CI: 12.29–240.83, P = 0.0650). The AUC area revealed a value of 0.9434. MiRNA-499 revealed a total sensitivity of 0.88 (95% CI: 0.86-0.90), a specificity of 0.87 (95% CI: 0.84-0.90) with a diagnostic OR of 79.55 (95% CI: 20.20-313.24); Furthermore, the value of the AUC area was 0.9584. The reported values suggested that all miRNAs were consistent as accepted biomarkers of MI. Again, miRNA-133a and miRNA-499 may retain a prominent role as biomarkers for MI, however, the function of miRNA-208b requires further investigation [133].
For a long time, the function of circulating miRNAs has been studied in the cellular components of whole blood and no findings have been provided in the subcomponents. Ward et al. [134] initially used a quantitative real-time polymerase-chain reaction system (qRT-PCR) to investigate the miRNA profiles of whole blood subcomponents in patients with ACS and focused research on plasma, leukocytes, and platelets. Patient cohorts included thirteen AMI patients admitted to the emergency department or undergoing PCI with electrocardiographic signs of ST-segment elevation (STEMI) or non-ST-segment elevation (NSTEMI). Whole blood was recovered from arterial blood samples during PCI. The cell-specific miRNA profiling required by the protocol specifically included the expression of 343 miRNAs that were quantified from whole blood, plasma, peripheral blood mononuclear cells, and platelets. Specifically, there were three different conditions encountered that provided substance to the discussion. Patients who exhibited STEMI revealed higher levels of miRNAs which noted increased expression of miR-25-3p, miR-221-3p, and miR-374b-5p as compared to the NSTEMI cohort. The expression of miRNA 30d-5p non-coding RNA sequences was linked to plasma, platelets, and leukocytes either in STEMI or NSTEMI cohorts. On the contrary miRNAs 221-3p and 483-5p were associated with plasma and platelets only in NSTEMI patients. The authors first advocated that cell-specific miRNAs were substantially different in the cohorts of patients with STEMI and NSTEMI. Likewise, from this study, it emerged that the distribution of miRNA was unique among plasma, platelets, and leukocytes in patients with ischemic heart disease or ACS. Furthermore, given the reported results it was suggested that unique miRNA profiles among the circulating subcomponents characterized patients with myocardial ischemia [134].
Potential bias for broad validation of the diagnostic power of miRNAs in different clinical settings of ACS, including SA, UA, and AMI in patients admitted in ED or undergoing PCI, is represented by a low number of subjects studied. This condition could not considerably assess the interconnection of microRNAs with clinical features and their potential prognostic value. Widera et al. [135] counteracted this concern by assessing the diagnostic and prognostic utility of cardiomyocyte-enriched microRNAs in the circumstances of different clinical settings. Nevertheless, their findings reinvigorated previous speculations about the potential utility of cardiomyocyte-enriched microRNAs as diagnostic or prognostic markers in ACS. In the report, sensitive myonecrosis biomarkers in a larger ACS cohort with 444 subjects were evaluated. Concentration levels of MiR-1, miR-133a, miR-133b, miR-208a, miR-208b, and miR-499 were determined by quantitative reverse transcription PCR in plasma samples obtained after patients’ admission in ED or who received a PCI. Multiple linear regression analysis including clinical variables and hsTnT, miR-1, miR-133a, miR-133b, and miR-208b advocated an independent association with hsTnT levels (all P<0.001). Importantly in patients who exhibited myocardial infarction, higher levels of miR-1, miR-133a, and miR-208b occurred compared to those who had unstable angina. However, all six investigated microRNAs noted a wide range across subjects with unstable angina or myocardial infarction. Levels of non-coding RNA sequences such as miR-133a and miR-208b were crucially related to the risk of death in univariate and age- and gender-adjusted analyses. However, in further statistical discrimination performed after further adjustment for hsTnT, both microRNAs lost their independent association for mortality outcome [135].
Oerlemans et al. [136] prospectively studied the role of circulating microRNAs in ACS using a large cohort of patients, so that the performed analysis could statistically reinforce the results emerging from previous studies that were based on a smaller number of patients. Furthermore, previously performed studies were not designed to involve comparisons with stability markers of cardiac damage and were flawed by the presence of appropriate controls. Briefly, 332 patients with suspected ACS admitted to the ED were enrolled for a single-center evaluation on the potential diagnostic value of circulating microRNAs as novel biomarkers which included cardiac miRNAs (miR-1, -208a, and -499), miR -21 and miR-146a. Subjects with STEMI were not considered for the enrollment which included both UA and NSTEMI patients. The levels of all analyzed miRNAs were markedly increased in 106 patients diagnosed with ACS. These values were also confirmed in patients with initially negative high-sensitivity (hs) troponin or symptom onset <3 hours. In particular, three specific miRNAs, MiR-1, miR-499, and miR-21 had a markedly diagnostic value in all patients hospitalized for suspected ACS in combination with increased hs-troponin levels. The AUC value was statistically significant (from AUC = 0.85 to AUC = 0.90; p > 0.001). Two points emerged promoting these three circulating microRNAs as strong predictors of ACS. The former was their independence from clinical co-variables including patient history and cardiovascular risk factors. The second demonstrated, under the highly reliable statistical profile, that the combination of these three miRNAs generated a higher AUC area with a value of 0.94 compared to hs-troponin T (0.89). Findings suggested that patients with NSTEMI had higher levels of miRNA-208a and miRNA-146a than patients with UA. Conversely, increased levels of miRNA-1 and miRNA-499 tended towards a clinical picture of UA and the levels measured in these patients exceeded those recorded in patients with NSTEMI. miRNA-21 was comparable between NSTEMI and UA subjects. Circulating microRNAs detected in patients with non-ST elevation ACS (NSTE-ACS) and with UA provide great potential as new early biomarkers for the management of patients who are suspected of having ACS [136].
Bai et al. [137] have broadened the field of action by recovering a more assorted typology of subjects with ACS. The authors with the use of gene chip technology evaluated miRNA expression in patients with SA, NSTE-ACS, and ST-segment elevation myocardial infarction (STEMI). 5 patients from each group and 5 controls without CAD were enrolled. All subjects had three or more risk factors. Microarray analysis was used to highlight differential miRNA expression which was confirmed by qRT-PCR. Patients in the control group and those included in the SA or NSTE-ACS groups revealed differentially expressed microRNAs that were involved in inflammation, protein phosphorylation, and cell adhesion, compared with miRNAs from STEMI patients. In addition, differentially expressed miRNAs were observed to be related to mitogen-activated protein kinase signaling, calcium ion pathways, and cell adhesion pathways [137]. It is important to dwell on some points which reinforce the novelty of this study. First, significant upregulation of miR-941, miR-363-3p, and miR-182-5p levels were recorded (fold-change: 2.0 or more, P < 0.05) in control, AS or NSTE- ACS, respectively when compared to data for expression levels in patients with STEMI. Second, data from qRT-PCR analysis noted the plasma miR-941 level was elevated in the NSTE-ACS or STEMI cohorts compared with that in patients without CAD (fold-change: 1.65 and 2.28, respectively; P < 0.05). Third, miR-941 expression was cross-matched between groups. This expression was raised considerably in the STEMI group compared to SA (P < 0.01) and NSTE-ACS (P < 0.05) groups. Likewise, miR-941 expression was higher in ACS patients with NSTE-ACS or STEMI, as compared to patients with AS or without ACS or CAD (P < 0,01). Significant differences in miR-182-5p and miR-363-3p expression were not observed. The areas under the receiver operating characteristic curves were 0.896, 0.808, and 0.781 for patients in the control, AS, and ACS-NSTE groups, respectively, compared with those for STEMI patients. In contrast, that for the ACS group compared with the non-ACS group was 0.734. Evidence suggested that miR-941 expression was relatively higher in patients with ACS and STEMI which may be speculative for the role of miR-941 as a potential biomarker of ACS or STEMI [137].
Two studies by Wang et al. [138,139] deserve comprehensive discussion in the context of atherosclerosis-related circulating miRNAs and advocate the role of novel and sensitive predictors for acute myocardial infarction. Although the authors reported altered expressions of circulating miRNA-21-5p, miRNA-361-5p, and circulating miRNA-519e-5p in patients with coronary atherosclerosis through the use of miRNA microarrays. However, there was no solid evidence for a global role of the expression levels of these circulating miRNAs in the early phase of AMI.
In the former study [138] the expressions of circulating miR-21-5p, miR-361-5p, and miR-519e-5p were studied in AMI patients, and at the same time, their clinical applications for AMI diagnosis and monitoring were evaluated. Study cohorts included the first cohort composed of 17 AMI patients and 28 healthy volunteers while the second cohort consisted of 9 AMI patients, 9 ischemic stroke patients, 8 pulmonary embolism patients, and 12 healthy volunteers. Quantitative real-time PCR and ELISA assays to identify plasma miRNA and cardiac troponin I (cTnI) concentrations, respectively, were performed. The evidence proved that in patients with AMI, the plasma levels of miR-21-5p and miR-361-5p were considerably increased while the concentration of circulating miR-519e-5p was decreased. The significant data was represented by the correlated increase of the levels of these circulating miRNAs with the concentrations of plasma cTnI. At an evaluation of the receiver operating characteristics (ROC), it was observed that these three circulating miRNAs retained a remarkable diagnostic precision for AMI highlighting high values of area under the ROC curve (AUC). Therefore, it clearly emerged that the combination of the three miRNAs crucially raised the diagnostic accuracy. Furthermore, cell experiments provided a reasonable explanation about the origin of these plasma miRNAs that may originate from damaged cardiomyocytes promoted by hypoxia. Again, an important finding concerned the raised levels of all three circulating miRNAs in ischemic stroke (IS) and pulmonary embolism (PE) which contrasted the finding of reduced level of plasma miR-519e-5p reported only in patients who exhibited AMI. Crosswise ROC analysis advocated that circulating miR-519e-5p may be a useful biomarker to discriminate AMI from other ischemic diseases. In this report, Wang et al. fixed that circulating miRNAs may be considered novel and powerful biomarkers for AMI and could represent a potential diagnostic tool for AMI [138].
In later work, Wang et al. [139] narrowed down the field of investigation on the relationship between circulating miRNAs and high-risk features in patients with NSTE-ACS using a GRACE (Global Registry of Acute Coronary Events) analysis. RNA was extracted from the whole blood of 199 patients with NSTE-ACS and whole genome miRNA sequencing was performed. To test the validity of the study, 13 high-risk clinical traits were associated with the use of generalized linear models. In this respect, the GRACE risk score is widely validated for mortality in patients with ACS-NSTE. 205 nominally significant (p < 0.05) miRNA risk factor associations were reported. Of note, significant combinations were observed most frequently with chronic heart failure (HF) which stressed 43 miR. Again, the GRACE risk score underscored 30 miR and renal function 32 miR, respectively. Concerns related to the hierarchical cluster analysis highlighted that chronic HF risk score and GRACE were distinguished by a tendency to cluster more closely together, splitting 14 miRNAs with matching fold-change direction. After eliminating a 5% false discovery rate, chronic heart failure had a significant association with lower circulating levels of miR-3135b (p < 0.0006), miR-126-5p (p < 0.0001), miR -142-5p (p = 0.0004) and miR-144-5p (p = 0.0007). Conversely, the raise in GRACE risk score was inversely tailed with miR-3135b levels (p < 0.0001) and positively tailed with miR-28-3p levels (p = 0.0002. Despite the need to validate with more substantial evidence, the results of two miRs emerged in the study, accentuating the evidence that circulating miRs cluster around two potent traits for mortality risk in NSTE-ACS. For MiR-3135b, under expression has been reported in chronic heart failure associated with an increase in GRACE risk score whereas in the miR-28-3p cluster, no known association with cardiovascular disease has been reported [139].
Kaur et al. [140] evaluated dysregulated miRNA biomarkers in CAD after screening 140 original articles that advocated appropriate evidence for data mining. The systematic review pooled data from these reported studies to compare miRNAs identified in patients suffering from ACS matched with stable CAD patients and control populations. The most usually indicated miRNAs in any CAD were miR-1, miR-133a, miR-208a/b, and miR-499 which were also noted in abundance within the heart. These miRNAs performed a decisive function in cardiac physiology. For an in-depth analysis of the studies that compared subjects with symptoms referable to ACS with patients who manifested stable CAD, miR-21, miR-208a/b, miR-133a/b, miR-30 family, miR-19 and miR -20 were reported more routinely as dysregulated in the ACS group. Authors observed that several miRNAs were consistently available across studies alongside their utterances in either the ACS group or stable CAD group, compared to controls. However, some miRNAs have been identified as biomarkers specifically in ACS patients who expressed plasma levels of miR-499, miR-1, miR-133a/b, and miR -208a/b and in stable CAD patients with plasma levels of miR-215, miR-487a and miR-502. Thus, higher plasma levels of miR-21, miR-133, and miR-499 appear to advocate greater potential as biomarkers to distinguish the diagnosis of ACS from stable CAD. Particular attention should be paid to miR-499 which disclosed a connection between the level of its concentration slope and myocardial damage. It is important to underline that although these miRNAs may provide direct guidance toward potential diagnostic biomarkers, the reported results suggest caution in their interpretation. Concerns are related to the conduct of most studies based on candidate-driven predefined assessments of a limited number of miRNAs [140].
Recently, Zhelankin et al. [141] studied circulating microRNAs as non-invasive biomarkers of cardiovascular disease in CAD and ACS. The authors emphasized the concerns about controversy and inconsistency of data reported for some miRNA, which probably was due to pre-analytical and methodological discrepancies that emerged in diverse studies. Quantitative PCR to measure the relative plasma levels of eight circulating miRNAs was used and potentially correlated to the atherosclerosis process. The cohort included 136 adult subjects with clinical onset of CAD or ACS that included NSTEMI and STEMI admitted patients. Controls included outpatient healthy patients or subjects with hypertension without CAD. The plasma levels of miR-21-5p and miR-146a-5p were significantly increased in patients who disclosed ACS while the level of miR-17-5p was reduced in subjects presenting with ACS and stable CAD as compared to control patients which included healthy or hypertensive without CAD, respectively. Within the ACS patient group, no differences were found in the plasma levels of these miRNAs between patients with positive and negative troponins, nor were any differences found between STEMI and NSTEMI. Our results indicate that increased plasma levels of miR-146a-5p and miR-21-5p can be considered general ACS circulating biomarkers and that lowered miR-17-5p can be considered a general biomarker of CAD. Concerning patients included in the ACS group, no differences were revealed in the plasma levels of these miRNAs whilst the group of patients exhibited positive and negative troponin levels, nor were any differences within the subjects’ group who had STEMI and NSTEMI. This evidence suggested that raised plasma levels of miR-146a-5p and miR-21-5p may be classified as general ACS circulating biomarkers and that lowered levels of miR-17-5p may be a general biomarker of CAD [141].
6. Implication of miRNA in Stent Restenosis
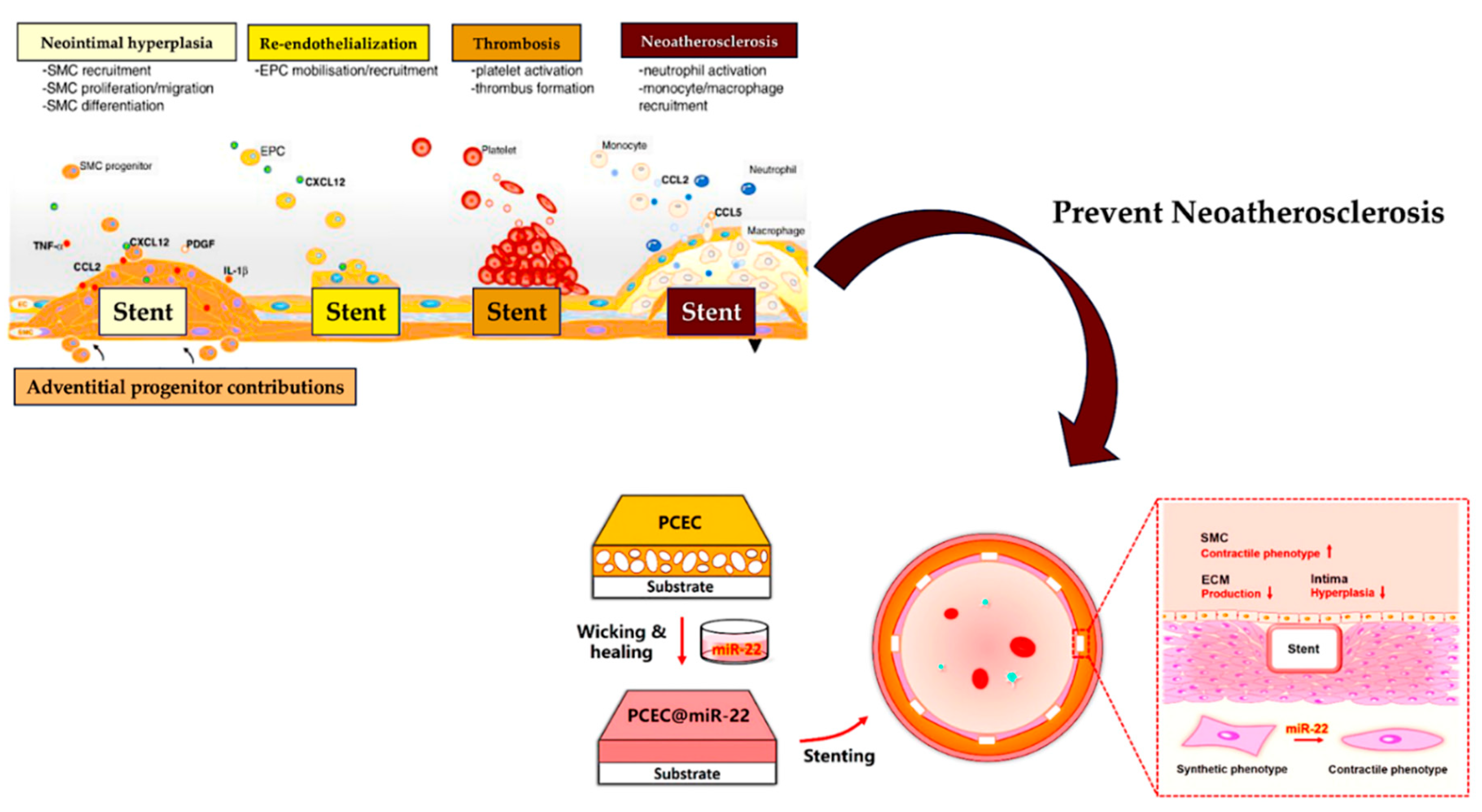
Alterations in miRNA levels have been evaluated in cases of in-stent restenosis which is a complication that occurs after PCI. Both bare metal stents and drug-eluting stents can develop atypical intra-stent neo-proliferation. However, in bare metal stents, the phenomenon of atypical proliferation and smooth muscle cell migration is advocated by the manifestation of vascular lesions that promote out-of-control neointimal formation with subsequent intra-stent restenosis. [142,143,144] Although with the advent of drug-eluting stents, tumors develop less frequently, a significant reduction in the risk of restenosis within the stent is less often desired. However, their major drawback is the increased risk of late thrombosis which is a concern that needs to be addressed. The risk of late thrombosis reported in drug-eluting stents is promoted by the regenerative power of endothelial cells induced by the vascular wall injury and the initiation of the prolonged arterial healing process [145,146,147,148,149].
The modifications of miRNA levels alongside the function of these miRNAs promoted by the implantation of either bare metal or drug-eluting stents have been studied in pig, mouse, and in vitro models [91]. Firstly, in a porcine animal model, miRNA levels were measured in control non-stented and stent-receiving coronary arteries. The results demonstrated the overexpression of several pro-inflammatory miRNAs, including miRNA-21, in stented coronary arteries. Secondly, stent miRNA-21 knockout (KO) mice and stent wild-type mice were compared. In the stented miRNA-21 knockout (KO) arm a reduction in neointima-medial ratio, neointimal thickness, and neointimal area compared to stented wild-type control mice was observed. The subsequent analysis showed an increase in the rate of anti-inflammatory M2 macrophages in the miRNA-21 KO mice arm compared to the wild-type control arm. The significant data emerging from this study advocated a reduction of neointimal proliferation in miRNA-21 stented (KO). This phenomenon was promoted by a substantial shift towards M2 macrophage differentiation associated with reduced smooth muscle cell proliferative and migratory response but without affecting endothelial cell function. Evidence that miRNA-21 promoted inflammation and vessel remodeling after stent implantation was suggested. Therefore, the next step could be the experimentation and subsequent application of drugs that regulate the expression and plasma levels of miRNA-21 that could improve the results of the drug-eluting stents used today for thrombosis [91].
Wang et al. [150] explored the determined function of microRNAs in the interconnected process of myointimal hyperplasia/ in-stent restenosis, using a humanized animal model. Only miR-21 was found upregulated among the candidates proposed in the study. Furthermore, human tissue samples from patients with in-stent restenosis were supported by an increase in miR-21 expression compared to the sample collected from patients with coronary heart disease. Again, miR-21 was systematically restrained via intravenous fluorescein-blocked nucleic acid-anti-miR-21 (anti-21) in the humanized myointimal hyperplasia model used. As anticipated, the suppression of vascular miR-21 was closely related in a dose-dependent manner, in which the reduced obliteration of the lumen followed. Furthermore, anti-21 did not stop the re-endothelialization process. However, systemic administration of anti-miR-21 promoted off-target adverse effects, advocating a decreased expression of miR-21 in the liver, heart, lung, and kidney with concomitant elevation of serum creatinine levels. The usefulness of local suppression of miR-21 was assessed using anti-21-coated stents revealing that the latter effectively reduced the in-stent restenosis as compared with bare metal stents, albeit without significant off-target effects could be noticed. The efficacy of an anti-miR-coated stent for the reduction of in-stent restenosis may provide a promising therapeutic resource [150].
Figure 5.
Schematic illustration of the preparation of the PCEC@miR-22 coated stent. The miR-22 was loaded into PCEC spongy coating based on a wicking action and subsequent self-healing encapsulation.
Figure 5.
Schematic illustration of the preparation of the PCEC@miR-22 coated stent. The miR-22 was loaded into PCEC spongy coating based on a wicking action and subsequent self-healing encapsulation.
7. Limitations
Of the 25 clinical trials on microRNAs with therapeutic potential in cardiovascular disease, 17 of these had phase I or II clinical trials completed [22,97,98,151,152,153,154,155,156,157,158] Although some reported withdrawals, trials that had completed the clinical phase of the study we observed to have different therapeutic developments targeting microRNA for specific indications (e.g., miravirsen, RG-101, cobomarsen, AZD 4076), including the impact in the cardiovascular field (miR-132-3p/inhibitor-CDR132L), proved to be lower than assumed [22,97,98,151,152,153,154,155,156,157,158]. However, it is important to underline that preclinical and clinical data obtained with inhibitory oligonucleotides, including clinical trials that have been discontinued, provide valuable information for the design and performance of microRNA-targeted cardiovascular therapies. Particular attention is paid to miR-17-5p, miR-21-5p, miR-29b-3p, and miR-92a-3p, which have been extensively studied in the laboratory and the clinical field. A special note is the miR-132-3p (CDR132L) inhibitor, which was developed for the treatment of heart failure. [9] The study of CDR132L is well advanced and phase II trials are being planned for which it could become the first drug targeting microRNAs in cardiovascular therapy [9,22,25,101].
8. Conclusions
We have explored the role of miRNAs in the field of cardiovascular disease and discussed many clinical studies targeting microRNAs. In our work, significant results emerged in a published landmark paper that led to the therapeutic application of an antimiR by inoculation. Although the progress made in the last decade suggests that this innovative path for the diagnosis and therapy of cardiovascular diseases should be taken decisively, the results of many microRNAs have yet to be clearly distinguished. It is possible to assume that there may be a wider scope of disease conditions and applications of microRNA therapies than what appears today, through the evaluation of the evidence produced. The body of literature reviewed here is relevant and further scrutiny of microRNAs is required, [159] to validate the function of microRNA candidates in the precise application field of cardiology. We, therefore, expect good results with the combination of microRNA manipulation in cardiovascular disease models, omics technologies, and extensive preclinical testing. These will be key to substantially improving therapeutic development and reducing the risk of dropouts.
Given the progress made in the development of synthetic oligonucleotides, overcoming important hurdles appears to be a solved challenge. Unfortunately, the delivery of these molecules still poses significant points for investigation. This is pertinent to cardiovascular tissues, which does not absorb oligonucleotides efficiently. Another key resolution for the administration of potential drugs can dictate the specific modification of the oligonucleotides to improve both their absorption and their cellular specificity. This part of pharmaceutical research is currently not very advanced and considerable uncertainty exists regarding the optimal concentrations of cellular oligonucleotides. This implies that intense efforts will be needed to obtain the screening of ligands and their chemical connection to oligonucleotides, together with the use of more selective methods that evaluate the concentrations of cellular oligonucleotides.
Finally, concrete wide use of miRNAs remains experimental at the time of writing but has a promising role in the diagnosis, prognosis, and therapy of a variety of cardiovascular disease presentations. Its potential therapeutic role may also be useful for the next generation of stents. Its use, either on its own or coupled with currently available biomarkers may be adopted soon especially if diagnostic ambiguity exists. Further studies are needed to validate its use in the general clinical setting which would need to highlight reproducibility and specificity.
Author Contributions
Conceptualization, F.N.; methodology, F.N. and S.S.A.S.; software, S.S.A.S.; A.A; validation, F.N, S.S.A.S.,V.J; A.A, T.S; formal analysis, F.N., S.S.A.S. V.J.; investigation, F.N., A.A., T.S.; data curation, F.N., S.S.A.S., T.S.; writing—original draft preparation, F.N.; writing—review and editing, F.N., S.S.A.S.,V.J, A.A, T.S.,; visualization, F.N., S.S.A.S., T.S.; supervision, F.N. S.S.A.S., T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochron ic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993, 75, 843–854. [CrossRef]
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 medi- ates temporal pattern formation in C. elegans. Cell. 1993, 75, 855–862.
- Friedman RC, Farh KK, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. https://doi.org/10.1101/gr.082701.108. [CrossRef]
- Kozomara A, Birgaoanu M, Griffiths-Jones S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019;47(d1): D155–D162. https://doi.org/10.1093/nar/gky1141. [CrossRef]
- Fromm B, Domanska D, Høye E, Ovchinnikov V, Kang W, Aparicio-Puerta E, Johansen M, Flatmark K, Mathelier A, Hovig E, Hackenberg M, Friedländer MR, Peterson KJ. MirGeneDB 2.0: the metazo- an microRNA complement. Nucleic Acids Res. 2020, 48, 132–141.
- Kim K, Baek SC, Lee YY, Bastiaanssen C, Kim J, Kim H, Kim VN. A quantitative map of human primary microRNA processing sites. Mol Cell. 2021, 81, 3422–3439. https://doi.org/10.1016/j.molcel.2021.07.002. [CrossRef]
- Matsui M, Prakash TP, Corey DR. Argonaute 2-dependent reg- ulation of gene expression by single-stranded miRNA mimics. Mol Ther. 2016, 24, 946–955.
- Eulalio A, Mano M, Dal Ferro M, Zentilin L, Sinagra G, Zacchigna S, Giacca M. Functional screening identifies miRNAs inducing cardiac regeneration. Nature. 2012, 492, 376–381. [CrossRef]
- Hinkel R, Ramanujam D, Kaczmarek V, Howe A, Klett K, Beck C, Dueck A, Thum T, Laugwitz KL, Maegdefessel L, Weber C, Kupatt C, Engelhardt S.AntimiR-21 prevents myocardial dysfunction in a pig model of ischemia/reperfusion injury. J Am Coll Cardiol. 2020, 75, 1788–1800. [CrossRef]
- Ganesan J, Ramanujam D, Sassi Y, Ahles A, Jentzsch C, Werfel S, Leierseder S, Loyer X, Giacca M, Zentilin L, Thum T, Laggerbauer B, Engelhardt S.MiR-378 controls cardiac hyper- trophy by combined repression of mitogen- activated protein kinase pathway factors. Circula- tion. 2013, 127, 2097–2106.
- Sassi Y, Avramopoulos P, Ramanujam D, Grüter L, Werfel S, Giosele S, Brunner AD, Esfandyari D, Papadopoulou AS, De Strooper B, Hübner N, Kumarswamy R, Thum T, Yin X, Mayr M, Laggerbauer B, Engelhardt S. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activat- ing Wnt signaling. Nat Commun. 2017, 8, 1614.
- Thum T, Gross C, Fiedler J, Fischer T, Kissler S, Bussen M, Galuppo P, Just S, Rottbauer W, Frantz S, Castoldi M, Soutschek J, Koteliansky V, Rosenwald A, Basson MA, Licht JD, Pena JT, Rouhanifard SH, Muckenthaler MU, Tuschl T, Martin GR, Bauersachs J, Engelhardt S. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature. 2008, 456, 980–984. [CrossRef]
- Ji R, Cheng Y, Yue J, Yang J, Liu X, Chen H, Dean DB, Zhang C. MicroRNA expression signature and antisense-mediated depletion reveal an essential role of microRNA in vascular neointimal lesion formation. Circ Res.2007, 100, 1579–1588. https://doi.org/10.1161/circresaha.106.141986. [CrossRef]
- Ramanujam D, Sassi Y, Laggerbauer B, Engelhardt S. Viral vector-based targeting of miR-21 in cardiac nonmyocyte cells reduces pathologic remodeling of the heart. Mol Ther. 2016, 24, 1939–1948. https://doi.org/10.1038/mt.2016.166. [CrossRef]
- van Rooij E, Sutherland LB, Thatcher JE, DiMaio JM, Naseem RH, Marshall WS, Hill JA, Olson EN. Dysregulation of microRNAs after myocardial infarction reveals a role of miR- 29 in cardiac fibrosis. Proc Natl Acad Sci U S A. 2008, 105, 13027–13032.
- Boon RA, Seeger T, Heydt S, Fischer A, Hergenreider E, Horrevoets AJ, Vinciguerra M, Rosenthal N, Sciacca S, Pilato M, van Heijningen P, Essers J, Brandes RP, Zeiher AM, Dimmeler S. MicroRNA-29 in aortic dilation: implications for aneurysm formation. Circ Res. 2011, 109, 1115–1119. https://doi.org/10.1161/circresaha.111.255737. [CrossRef]
- Maegdefessel L, Azuma J, Toh R, Merk DR, Deng A, Chin JT, Raaz U, Schoelmerich AM, Raiesdana A, Leeper NJ, McConnell MV, Dalman RL, Spin JM, Tsao PS. Inhibition of micro- RNA-29b reduces murine abdominal aortic aneurysm development. J Clin Invest. 2012, 122, 497–506. [CrossRef]
- McDonald RA, White KM, Wu J, Cooley BC, Robertson KE, Halliday CA, McClure JD, Francis S, Lu R, Kennedy S, George SJ, Wan S, van Rooij E, Baker AH. miRNA-21 is dysregulated in response to vein grafting in multiple models and genetic ablation in mice attenuates neointima formation. Eur Heart J. 2013, 34, 1636–1643. [CrossRef]
- Eken SM, Christersdottir T, Winski G, Sangsuwan T, Jin H, Chernogubova E, Pirault J, Sun C, Simon N, Winter H, Backlund A, Haghdoost S, Hansson GK, Halle M, Maegdefessel L. miR-29b mediates the chronic inflammatory response in radiotherapy- induced vascular disease. JACC Basic Transl Sci. 2019, 4, 72–82.
- Zeng Z, Xia L, Fan X, Ostriker AC, Yarovinsky T, Su M, Zhang Y, Peng X, Xie Y, Pi L, Gu X, Chung SK, Martin KA, Liu R, Hwa J, Tang WH. Platelet-derived miR-223 promotes a phenotypic switch in arterial injury repair. J Clin Invest. 2019, 129, 1372–1386. [CrossRef]
- Hinkel R, Penzkofer D, Zühlke S, Fischer A, Husada W, Xu QF, Baloch E, van Rooij E, Zeiher AM, Kupatt C, Dimmeler S. Inhibition of microRNA-92a protects against ischemia/reperfusion injury in a large-animal model. Circulation. 2013, 128, 1066–1075.
- Foinquinos A, Batkai S, Genschel C, Viereck J, Rump S, Gyöngyösi M, Traxler D, Riesenhuber M, Spannbauer A, Lukovic D, Weber N, Zlabinger K, Hašimbegović E, Winkler J, Fiedler J, Dangwal S, Fischer M, de la Roche J, Wojciechowski D, Kraft T, Garamvölgyi R, Neitzel S, Chatterjee S, Yin X, Bär C, Mayr M, Xiao K, Thum T. Preclinical development of a miR-132 inhibitor for heart failure treatment. Nat Commun. 2020, 11, 633.
- Corsten MF, Papageorgiou A, Verhesen W, Carai P, Lindow M, Obad S, Summer G, Coort SL, Hazebroek M, van Leeuwen R, Gijbels MJ, Wijnands E, Biessen EA, De Winther MP, Stassen FR, Carmeliet P, Kauppinen S, Schroen B, Heymans S. MicroRNA profiling identifies microRNA-155 as an adverse mediator of cardiac injury and dysfunction during acute viral myocarditis. Circ Res. 2012, 111, 415–425. [CrossRef]
- Bonauer A, Carmona G, Iwasaki M, Mione M, Koyanagi M, Fischer A, Burchfield J, Fox H, Doebele C, Ohtani K, Chavakis E, Potente M, Tjwa M, Urbich C, Zeiher AM, Dimmeler S.MicroRNA-92a controls angio- genesis and functional recovery of ischemic tissues in mice. Science. 2009, 324, 1710–1713.
- Ucar A, Gupta SK, Fiedler J, Erikci E, Kardasinski M, Batkai S, Dangwal S, Kumarswamy R, Bang C, Holzmann A, Remke J, Caprio M, Jentzsch C, Engelhardt S, Geisendorf S, Glas C, Hofmann TG, Nessling M, Richter K, Schiffer M, Carrier L, Napp LC, Bauersachs J, Chowdhury K, Thum T. The miRNA-212/132 family regu- lates both cardiac hypertrophy and cardiomyocyte autophagy. Nat Commun. 2012; 3:1078.
- Heymans S, Corsten MF, Verhesen W, Carai P, van Leeuwen RE, Custers K, Peters T, Hazebroek M, Stöger L, Wijnands E, Janssen BJ, Creemers EE, Pinto YM, Grimm D, Schürmann N, Vigorito E, Thum T, Stassen F, Yin X, Mayr M, de Windt LJ, Lutgens E, Wouters K, de Winther MP, Zacchigna S, Giacca M, van Bilsen M, Papageorgiou AP, Schroen B. Macrophage microRNA-155 promotes cardiac hypertrophy and failure. Circulation. 2013, 128, 1420–1432. [CrossRef]
- Mann M, Mehta A, Zhao JL, Lee K, Marinov GK, Garcia-Flores Y, Lu LF, Rudensky AY, Baltimore D. An NF-κB-microRNA regulatory network tunes macrophage inflammatory responses. Nat Commun. 2017, 8, 851.
- Pankratz F, Bemtgen X, Zeiser R, Leonhardt F, Kreuzaler S, Hilgendorf I, Smolka C, Helbing T, Hoefer I, Esser JS, Kustermann M, Moser M, Bode C, Grundmann S. MicroRNA-155 exerts cell- specific antiangiogenic but proarteriogenic effects during adaptive neovascularization. Circulation. 2015, 131, 1575–1589. [CrossRef]
- Loyer X, Potteaux S, Vion AC, Guérin CL, Boulkroun S, Rautou PE, Ramkhelawon B, Esposito B, Dalloz M, Paul JL, Julia P, Maccario J, Boulanger CM, Mallat Z, Tedgui A. Inhibition of microRNA-92a pre- vents endothelial dysfunction and atherosclerosis in mice. Circ Res. 2014, 114, 434–443.
- 30. Daniel JM, Penzkofer D, Teske R, Dutzmann J, Koch A, Bielenberg W, Bonauer A, Boon RA, Fischer A, Bauersachs J, van Rooij E, Dimmeler S, Sedding DG. Inhibition of miR-92a improves re-endothelialization and prevents neointima formation following vascular injury. Cardiovasc Res. 2014, 103, 564–572. [CrossRef]
- Halkein J, Tabruyn SP, Ricke-Hoch M, Haghikia A, Nguyen NQ, Scherr M, Castermans K, Malvaux L, Lambert V, Thiry M, Sliwa K, Noel A, Martial JA, Hilfiker-Kleiner D, Struman I. MicroRNA-146a is a therapeutic target and biomarker for peripartum cardiomy- opathy. J Clin Invest. 2013, 123, 2143–2154.
- Sun X, Icli B, Wara AK, Belkin N, He S, Kobzik L, Hunninghake GM, Vera MP; MICU Registry; Blackwell TS, Baron RM, Feinberg MW. MicroRNA-181b regulates NF-κB– mediated vascular inflammation. J Clin Invest. 2012, 122, 1973–1990.
- da Costa Martins PA, Salic K, Gladka MM, Armand AS, Leptidis S, el Azzouzi H, Hansen A, Coenen-de Roo CJ, Bierhuizen MF, van der Nagel R, van Kuik J, de Weger R, de Bruin A, Condorelli G, Arbones ML, Eschenhagen T, De Windt LJ. MicroRNA-199b targets the nuclear kinase Dyrk1a in an auto-amplification loop promoting calcineurin/NFAT signalling. Nat Cell Biol. 2010, 12, 1220–1227. https://doi.org/10.1038/ncb2126. [CrossRef]
- Duygu B, Poels EM, Juni R, Bitsch N, Ottaviani L, Olieslagers S, de Windt LJ, da Costa Martins PA. miR-199b-5p is a regulator of left ventricular remodeling following myocardial infarction. Noncoding RNA Res. 2017, 2, 18–26. https://doi.org/10.1016/j.ncrna.2016.12.002. [CrossRef]
- van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, Gerard RD, Richardson JA, Olson EN. A signature pattern of stress- responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proc Natl Acad Sci U S A. 2006, 103, 18255–18260. https://doi.org/10.1073/pnas.0608791103. [CrossRef]
- Aurora AB, Mahmoud AI, Luo X, Johnson BA, van Rooij E, Matsuzaki S, Humphries KM, Hill JA, Bassel-Duby R, Sadek HA, Olson EN. MicroRNA-214 protects the mouse heart from ischemic injury by controlling Ca2+ overload and cell death. J Clin Invest. 2012, 122, 1222–1232. https://doi.org/10.1172/jci59327. [CrossRef]
- Liu X, Xiao J, Zhu H, Wei X, Platt C, Damilano F, Xiao C, Bezzerides V, Boström P, Che L, Zhang C, Spiegelman BM, Rosenzweig A. miR-222 is necessary for exercise- induced cardiac growth and protects against pathological cardiac remodeling. Cell Metab. 2015, 21, 584–595. https://doi.org/10.1016/j.cmet.2015.02.014. [CrossRef]
- Verjans R, Peters T, Beaumont FJ, van Leeuwen R, van Herwaarden T, Verhesen W, Munts C, Bijnen M, Henkens M, Diez J, de Windt LJ, van Nieuwenhoven FA, van Bilsen M, Goumans MJ, Heymans S, González A, Schroen B. MicroRNA-221/222 family counteracts myocardial fibrosis in pressure overload-induced heart failure. Hypertension. 2018, 71, 280–288.
- Carè A, Catalucci D, Felicetti F, Bonci D, Addario A, Gallo P, Bang ML, Segnalini P, Gu Y, Dalton ND, Elia L, Latronico MV, Høydal M, Autore C, Russo MA, Dorn GW 2nd, Ellingsen O, Ruiz-Lozano P, Peterson KL, Croce CM, Peschle C, Condorelli G. MicroRNA-133 controls cardiac hypertrophy. Nat Med. 2007, 13, 613–618.
- Karakikes I, Chaanine AH, Kang S, Mukete BN, Jeong D, Zhang S, Hajjar RJ, Lebeche D. Therapeutic cardiac-targeted delivery of miR-1 reverses pressure overload- induced cardiac hypertrophy and attenuates pathological remodeling. J Am Heart Assoc. 2013, 2, 17–19. https://doi.org/10.1161/jaha.113.000078. [CrossRef]
- Besser J, Malan D, Wystub K, Bachmann A, Wietelmann A, Sasse P, Fleischmann BK, Braun T, Boettger T. MiRNA-1/133a clusters regulate adrenergic control of cardiac repolarization. PLoS One. 2014, 9, e113449.
- Tijsen AJ, van der Made I, van den Hoogenhof MM, Wijnen WJ, van Deel ED, de Groot NE, Alekseev S, Fluiter K, Schroen B, Goumans MJ, van der Velden J, Duncker DJ, Pinto YM, Creemers EE. The microRNA-15 family inhibits the TGFβ-pathway in the heart. Cardiovasc Res. 2014, 104, 61–71. [CrossRef]
- Hullinger TG, Montgomery RL, Seto AG, Dickinson BA, Semus HM, Lynch JM, Dalby CM, Robinson K, Stack C, Latimer PA, Hare JM, Olson EN, van Rooij E. Inhibition of miR-15 pro- tects against cardiac ischemic injury. Circ Res. 2012, 110, 71–81.
- Porrello ER, Mahmoud AI, Simpson E, Johnson BA, Grinsfelder D, Canseco D, Mammen PP, Rothermel BA, Olson EN, Sadek HA. Regulation of neonatal and adult mammalian heart regeneration by the miR-15 family. Proc Natl Acad Sci U S A. 2013, 110, 187–192. doi:10.1073/pnas.1208863110. [CrossRef]
- Fiedler J, Stöhr A, Gupta SK, Hartmann D, Holzmann A, Just A, Hansen A, Hilfiker-Kleiner D, Eschenhagen T, Thum T. Functional microRNA library screening identifies the hypoxamir miR-24 as a potent regulator of smooth muscle cell proliferation and vascularization. Antioxid Redox Signal. 2014, 21, 1167–1176. [CrossRef]
- Meloni M, Marchetti M, Garner K, Littlejohns B, Sala-Newby G, Xenophontos N, Floris I, Suleiman MS, Madeddu P, Caporali A, Emanueli C. Local inhibition of micro- RNA-24 improves reparative angiogenesis and left ventricle remodeling and function in mice with myocardial infarction. Mol Ther. 2013, 21, 1390–1402. [CrossRef]
- Boon RA, Iekushi K, Lechner S, Seeger T, Fischer A, Heydt S, Kaluza D, Tréguer K, Carmona G, Bonauer A, Horrevoets AJ, Didier N, Girmatsion Z, Biliczki P, Ehrlich JR, Katus HA, Müller OJ, Potente M, Zeiher AM, Hermeking H, Dimmeler S. MicroRNA-34a regulates cardiac ageing and function. Nature. 2013, 495, 107–110. [CrossRef]
- Badi I, Mancinelli L, Polizzotto A, Ferri D, Zeni F, Burba I, Milano G, Brambilla F, Saccu C, Bianchi ME, Pompilio G, Capogrossi MC, Raucci A. miR-34a promotes vascular smooth muscle cell calcification by downregulating SIRT1 (sirtuin 1) and AXL (AXl receptor tyro- sine kinase). Arterioscler Thromb Vasc Biol. 2018, 38, 2079–2090.
- Griesemer D, Xue JR, Reilly SK, Ulirsch JC, Kukreja K, Davis JR, Kanai M, Yang DK, Butts JC, Guney MH, Luban J, Montgomery SB, Finucane HK, Novina CD, Tewhey R, Sabeti PC. Genome-wide functional screen of 3′UTR variants uncovers causal vari- ants for human disease and evolution. Cell. 2021, 184, 5247–5260.
- Ramanujam D, Schön AP, Beck C, Vaccarello P, Felician G, Dueck A, Esfandyari D, Meister G, Meitinger T, Schulz C, Engelhardt S. MicroRNA-21-dependent macrophage-to-fibroblast signaling determines the cardiac response to pressure overload. Circu- lation. 2021, 143, 1513–1525. https://doi.org/10.1161/circulationaha.120.050682. [CrossRef]
- Huang CK, Kafert-Kasting S, Thum T. Preclinical and clinical development of noncoding RNA therapeutics for cardio- vascular disease. Circ Res. 2020, 126, 663–678.
- Bang C, Batkai S, Dangwal S, Gupta SK, Foinquinos A, Holzmann A, Just A, Remke J, Zimmer K, Zeug A, Ponimaskin E, Schmiedl A, Yin X, Mayr M, Halder R, Fischer A, Engelhardt S, Wei Y, Schober A, Fiedler J, Thum T. Cardiac fibroblast–derived micro- RNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J Clin Invest. 2014, 124, 2136–2146. https://doi.org/10.1172/jci70577. [CrossRef]
- Wang K, Jiang Z, Webster KA, Chen J, Hu H, Zhou Y, Zhao J, Wang L, Wang Y, Zhong Z, Ni C, Li Q, Xiang C, Zhang L, Wu R, Zhu W, Yu H, Hu X, Wang J. Enhanced cardioprotection by human endometrium mesenchymal stem cells driven by exosomal microRNA-21. Stem Cells Transl Med. 2017, 6, 209–222. https://doi.org/10.5966/sctm.2015-0386. [CrossRef]
- Cheng M, Yang J, Zhao X, Zhang E, Zeng Q, Yu Y, Yang L, Wu B, Yi G, Mao X, Huang K, Dong N, Xie M, Limdi NA, Prabhu SD, Zhang J, Qin G. Circulating myocardial micro- RNAs from infarcted hearts are carried in exosomes and mobilise bone marrow progenitor cells. Nat Commun. 2019, 10, 959. [CrossRef]
- Zheng D, Huo M, Li B, Wang W, Piao H, Wang Y, Zhu Z, Li D, Wang T, Liu K. The role of exosomes and exosomal microRNA in cardiovascular disease. Front Cell Dev Biol. 2021; 8:1–15. https://doi.org/10.3389/fcell.2020.616161.
- Kesidou D, da Costa Martins PA, de Windt LJ, Brittan M, Beqqali A, Baker AH. Extracellular vesicle miRNAs in the promotion of cardiac neovascularisation. Front Physiol. 2020; 11:579892. doi:10.3389/fphys.2020.579892. [CrossRef]
- Ottaviani L, Sansonetti M, da Costa Martins PA. Myocardial cell-to-cell communication via microRNAs. Noncoding RNA Res. 2018, 3, 144–153. [CrossRef]
- Zernecke A, Bidzhekov K, Noels H, Shagdarsuren E, Gan L, Denecke B, Hristov M, Köppel T, Jahantigh MN, Lutgens E, Wang S, Olson EN, Schober A, Weber C. Delivery of microRNA-126 by apoptotic bodies induces CXCL12-dependent vascular protection. Sci Signal. 2009, 2, ra81. doi:10.1126/scisignal.2000610. [CrossRef]
- Small EM, Olson EN. Pervasive roles of microRNAs in cardiovascular biology. Nature. 2011 Jan 20;469(7330):336-42. https://doi.org/10.1038/nature09783. [CrossRef]
- Condorelli G, Dimmeler S. MicroRNAs: components of an integrated system controlling cardiac development, physiology, and disease pathogenesis. Cardiovasc Res. 2008 Sep 1;79(4):551-2. [CrossRef]
- Pratt AJ, MacRae IJ. The RNA-induced silencing complex: a versatile gene-silencing machine. J Biol Chem. 2009 Jul 3;284(27):17897-901. https://doi.org/10.1074/jbc.r900012200. [CrossRef]
- Kobayashi H, Tomari Y. RISC assembly: Coordination between small RNAs and Argonaute proteins. Biochim Biophys Acta. 2016 Jan;1859(1):71-81. https://doi.org/10.1016/j.bbagrm.2015.08.007. [CrossRef]
- Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009 Jan 23;136(2):215-33. https://doi.org/10.1016/j.cell.2009.01.002. [CrossRef]
- Gu S, Jin L, Zhang F, Sarnow P, Kay MA. Biological basis for restriction of microRNA targets to the 3’ untranslated region in mammalian mRNAs. Nat Struct Mol Biol. 2009 Feb;16(2):144-50.
- Liu B, Li J, Cairns MJ. Identifying miRNAs, targets and functions. Brief Bioinform. 2014 Jan;15(1):1-19.
- Bartel DP. Metazoan microRNAs. Cell. 2018, 173, 20–51.
- Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective micro- RNA target sites in mammalian mRNAs. Elife. 2015; 4:1–38. https://doi.org/10.7554/elife.05005. [CrossRef]
- Broughton JP, Lovci MT, Huang JL, Yeo GW, Pasquinelli AE. Pairing beyond the seed sup- ports MicroRNA targeting specificity. Mol Cell. 2016, 64, 320–333.
- Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell. 2007, 27, 91–105. [CrossRef]
- Gebert LFR, MacRae IJ. Regulation of micro- RNA function in animals. Nat Rev Mol Cell Biol. 2019, 20, 21–37. https://doi.org/10.1038/s41580-018-0045-7. [CrossRef]
- Yang A, Bofill-De Ros X, Shao TJ, Jiang M, Li K, Villanueva P, Dai L, Gu S. 3′ Uridylation confers miRNAs with non-canonical target repertoires. Mol Cell. 2019, 75, 511–522.e4. https://doi.org/10.1016/j.molcel.2019.05.014. [CrossRef]
- van der Kwast RVCT, Parma L, van der Bent ML, van Ingen E, Baganha F, Peters HAB, Goossens EAC, Simons KH, Palmen M, de Vries MR, Quax PHA, Nossent AY. Adenosine-to-inosine editing of vasoactive microRNAs alters their targetome and function in ischemia. Mol Ther Nucleic Acids. 2020; 21:932–953. https://doi.org/10.1016/j.omtn.2020.07.020. [CrossRef]
- McGahon MK, Yarham JM, Daly A, Guduric-Fuchs J, Ferguson LJ, Simpson DA, Collins A. Distinctive profile of IsomiR expression and novel microRNAs in rat heart left ventricle. PLoS One. 2013, 8, e65809. https://doi.org/10.1371/journal.pone.0065809. [CrossRef]
- van der Kwast RVCT, van Ingen E, Parma L, Peters HAB, Quax PHA, Nossent AY. Adenosine-to-inosine editing of MicroRNA-487b alters target gene selection after ischemia and promotes neovascu- larization. Circ Res. 2018, 122, 444–456.
- van der Kwast RVCT, Woudenberg T, Quax PHA, Nossent AY. MicroRNA-411 and its 5′-isomiR have distinct targets and functions and are differentially regulated in the vasculature under ischemia. Mol Ther. 2020, 28, 157–170.
- Kingston ER, Bartel DP. Global analyses of the dynamics of mammalian microRNA metabolism. Genome Res. 2019, 29, 1777–1790. https://doi.org/10.1101/gr.251421.119. [CrossRef]
- Marzi MJ, Ghini F, Cerruti B, de Pretis S, Bonetti P, Giacomelli C, Gorski MM, Kress T, Pelizzola M, Muller H, Amati B, Nicassio F. Degradation dynamics of micro- RNAs revealed by a novel pulse-chase approach. Genome Res. 2016, 26, 554–565. https://doi.org/10.1101/gr.198788.115. [CrossRef]
- Han J, LaVigne CA, Jones BT, Zhang H, Gillett F, Mendell JT. A ubiquitin ligase mediates target-directed microRNA decay inde- pendently of tailing and trimming. Science. 2020, 370, eabc9546.
- Shi CY, Kingston ER, Kleaveland B, Lin DH, Stubna MW, Bartel DP. The ZSWIM8 ubiquitin ligase medi- ates target-directed microRNA degradation. Science. 2020, 370, eabc9359.
- Bitetti A, Mallory AC, Golini E, Carrieri C, Carreño Gutiérrez H, Perlas E, Pérez-Rico YA, Tocchini-Valentini GP, Enright AJ, Norton WHJ, Mandillo S, O’Carroll D, Shkumatava A. MicroRNA degradation by a con- served target RNA regulates animal behavior. Nat Struct Mol Biol. 2018, 25, 244–251.
- Kalayinia S, Arjmand F, Maleki M, Malakootian M, Singh CP. MicroRNAs: roles in cardiovascular development and disease. Cardiovasc Pathol. 2021 Jan-Feb; 50:107296. doi:10.1016/j.carpath.2020.107296. [CrossRef]
- Sayed D, Abdullatif M. MicroRNAs in development and disease. Physiol Rev. 2011 Jul;91(3):827-87.
- Ma Q, Zhang L, Pearce WJ. MicroRNAs in brain development and cerebrovascular pathophysiology.Am J Physiol Cell Physiol. 2019 Jul 1;317(1):C3-C19. https://doi.org/10.1152/ajpcell.00022.2019. [CrossRef]
- Motshwari DD, Matshazi DM, Erasmus RT, Kengne AP, Matsha TE, George C. MicroRNAs Associated with Chronic Kidney Disease in the General Population and High-Risk Subgroups-A Systematic Review. Int J Mol Sci. 2023 Jan 16;24(2):1792. [CrossRef]
- Cheng Y, Du Y, Wang Q, Lv Q, Xue Y, Zhou W, Zhang C, Chen X, Wang D. Human cytomegalovirus-encoded microRNAs expression profile in plasma of patients with aortic dissection. J Cardiothorac Surg. 2023 Jan 18;18(1):39. [CrossRef]
- Quiat D, Olson EN. MicroRNAs in cardiovascular disease: from pathogenesis to prevention and treatment. J Clin Invest. 2013 Jan;123(1):11-8. [CrossRef]
- Wu J, Song J, Wang C, Niu D, Li H, Liu Y, Ma L, Yu R, Chen X, Zen K, Yang Q, Zhang C, Zhang CY, Wang J. Identification of serum microRNAs for cardiovascular risk stratification in dyslipidemia subjects. Int J Cardiol. 2014 Mar 1;172(1):232-4. [CrossRef]
- Vickers KC, Remaley AT. Lipid-based carriers of microRNAs and intercellular communication. Curr Opin Lipidol. 2012 Apr;23(2):91-7. [CrossRef]
- Boon RA, Vickers KC. Intercellular transport of microRNAs. Arterioscler Thromb Vasc Biol. 2013 Feb;33(2):186-92. [CrossRef]
- Caporali A, Miscianinov V, Saif J, Emanueli C. MicroRNA transport in cardiovascular complication of diabetes. Biochim Biophys Acta. 2016 Dec;1861(12 Pt B):2111-2120. https://doi.org/10.1016/j.bbalip.2016.01.010. [CrossRef]
- Mayr M, Zampetaki A, Willeit P, Willeit J, Kiechl S. MicroRNAs within the continuum of postgenomics biomarker discovery. Arterioscler Thromb Vasc Biol. 2013 Feb;33(2):206-14. https://doi.org/10.1161/atvbaha.112.300141. [CrossRef]
- Smith JG, Gerszten RE. Emerging Affinity-Based Proteomic Technologies for Large-Scale Plasma Profiling in Cardiovascular Disease. Circulation. 2017 Apr 25;135(17):1651-1664. https://doi.org/10.1161/circulationaha.116.025446. [CrossRef]
- Santovito D, Egea V, Bidzhekov K, Natarelli L, Mourão A, Blanchet X, Wichapong K, Aslani M, Brunßen C, Horckmans M, Hristov M, Geerlof A, Lutgens E, Daemen MJAP, Hackeng T, Ries C, Chavakis T, Morawietz H, Naumann R, von Hundelshausen P, Steffens S, Duchêne J, Megens RTA, Sattler M, Weber C. Noncanonical inhibition of caspase-3 by a nuclear microRNA confers endo- thelial protection by autophagy in atherosclerosis. Sci Transl Med. 2020, 12, 1–16.
- Li H, Zhan J, Zhao Y, Fan J, Yuan S, Yin Z, Dai B, Chen C, Wang DW. Identification of ncRNA-mediated functions of nucleus-localized miR-320 in cardiomyocytes. Mol Ther Nucleic Acids. 2020; 19:132–143.
- Elbarbary RA, Miyoshi K, Myers JR, Du P, Ashton JM, Tian B, Maquat LE. Tudor-SN-mediated endonucleolytic decay of human cell micro- RNAs promotes G1/S phase transition. Science. 2017, 356, 859–862.
- Yang A, Shao TJ, Bofill-De Ros X, Lian C, Villanueva P, Dai L, Gu S. AGO-bound mature miRNAs are oli- gouridylated by TUTs and subsequently degraded by DIS3L2. Nat Commun. 2020, 11, 1–13.
- Abplanalp WT, Fischer A, John D, Zeiher AM, Gosgnach W, Darville H, Montgomery R, Pestano L, Allée G, Paty I, Fougerousse F, Dimmeler S. Efficiency and target derepres- sion of anti-miR-92a: results of a first in human study. Nucleic Acid Ther. 2020, 30, 335–345.
- Anastasiadou E, Seto AG, Beatty X, Hermreck M, Gilles ME, Stroopinsky D, Pinter-Brown LC, Pestano L, Marchese C, Avigan D, Trivedi P, Escolar DM, Jackson AL, Slack FJ. Cobomarsen, an oligo- nucleotide inhibitor of miR-155, slows DLBCL tumor cell growth in vitro and in vivo. Clin Cancer Res. 2021, 27, 1139–1149.
- Batkai S, Genschel C, Viereck J, Rump S, Bär C, Borchert T, Traxler D, Riesenhuber M, Spannbauer A, Lukovic D, Zlabinger K, Hašimbegović E, Winkler J, Garamvölgyi R, Neitzel S, Gyöngyösi M, Thum T. CDR132L improves systol- ic and diastolic function in a large animal model of chronic heart failure. Eur Heart J. 2021, 42, 192–201.
- Hinkel R, Batkai S, Bähr A, Bozoglu T, Straub S, Borchert T, Viereck J, Howe A, Hornaschewitz N, Oberberger L, Jurisch V, Kozlik-Feldmann R, Freudenthal F, Ziegler T, Weber C, Sperandio M, Engelhardt S, Laugwitz KL, Moretti A, Klymiuk N, Thum T, Kupatt C.AntimiR-132 attenuates myocar- dial hypertrophy in an animal model of percu- taneous aortic constriction. J Am Col Cardiol. 2021, 77, 2923–2935.
- Täubel J, Hauke W, Rump S, Viereck J, Batkai S, Poetzsch J, Rode L, Weigt H, Genschel C, Lorch U, Theek C, Levin AA, Bauersachs J, Solomon SD, Thum T. Novel antisense therapy targeting microRNA-132 in patients with heart failure: results of a first-in-human phase 1b randomized, double-blind, placebo-controlled study. Eur Heart J. 2021, 42, 178–188. https://doi.org/10.1093/eurheartj/ehaa898. [CrossRef]
- Underlying Cause of Death 1999-2020—CDC WONDER. https://wonder.cdc.gov/wonder/help/ucd.html.
- World Health Organization, The Top 10 Causes of Death, 2014.
- Writing Committee Members; Lawton JS, Tamis-Holland JE, Bangalore S, Bates ER, Beckie TM, Bischoff JM, Bittl JA, Cohen MG, Di Maio JM, Don CW, Fremes SE, Gaudino MF, Goldberger ZD, Grant MC, Jaswal JB, Kurlansky PA, Mehran R, Metkus TS Jr, Nnacheta LC, Rao SV, Sellke FW, Sharma G, Yong CM, Zwischenberger BA. 2021 ACC/AHA/SCAI Guideline for Coronary Artery Revascularization: A Report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. J Am Coll Cardiol. 2022 Jan 18;79(2):e21-e129.
- Writing Committee, Michael C. Kontos, James A. de Lemos, Steven B. Deitelzweig, Deborah B. Diercks, M. Odette Gore, Erik P. Hess, Cian P. McCarthy, James K. McCord, Paul I. Musey, Todd C. Villines, and Leesa J. Wright. 2022 ACC Expert Consensus Decision Pathway on the Evaluation and Disposition of Acute Chest Pain in the Emergency Department: A Report of the American College of Cardiology Solution Set Oversight Committee J Am Coll Cardiol. 2022 Nov, 80 (20) 1925–1960.
- Writing Committee Members; Gulati M, Levy PD, Mukherjee D, Amsterdam E, Bhatt DL, Birtcher KK, Blankstein R, Boyd J, Bullock-Palmer RP, Conejo T, Diercks DB, Gentile F, Greenwood JP, Hess EP, Hollenberg SM, Jaber WA, Jneid H, Joglar JA, Morrow DA, O’Connor RE, Ross MA, Shaw LJ.2021 AHA/ACC/ASE/CHEST/SAEM/SCCT/SCMR Guideline for the Evaluation and Diagnosis of Chest Pain: Executive Summary: A Report of the American College of Cardiology/American Heart Association Joint Committee on Clinical Practice Guidelines. J Am Coll Cardiol. 2021 Nov 30;78(22):2218-2261. https://doi.org/10.1161/cir.0000000000001030. [CrossRef]
- Avdikos G, Michas G, Smith SW. From Q/Non-Q Myocardial Infarction to STEMI/NSTEMI: Why It’s Time to Consider Another Simplified Dichotomy; a Narrative Literature Review. Arch Acad Emerg Med. 2022 Oct 1;10(1): e78.
- Miyachi H, Takagi A, Miyauchi K, Yamasaki M, Tanaka H, Yoshikawa M, Saji M, Suzuki M, Yamamoto T, Shimizu W, Nagao K, Takayama M. Current characteristics and management of ST elevation and non-ST elevation myocardial infarction in the Tokyo metropolitan area: from the Tokyo CCU network registered cohort. Heart Vessels. 2016 Nov;31(11):1740-1751. https://doi.org/10.1007/s00380-015-0791-9. [CrossRef]
- Khan AR, Golwala H, Tripathi A, Bin Abdulhak AA, Bavishi C, Riaz H, Mallipedi V, Pandey A, Bhatt DL. Impact of total occlusion of culprit artery in acute non-ST elevation myocardial infarction: a systematic review and meta-analysis. Eur Heart J. 2017 Nov 1;38(41):3082-3089. https://doi.org/10.1093/eurheartj/ehx418. [CrossRef]
- Mitka M. New definition of myocardial infarction puts biomarkers front and center. JAMA. 2012 Oct 17;308(15):1511-2. https://doi.org/10.1001/jama.2012.12794. [CrossRef]
- Möckel M, Giannitsis E, Mueller C, Huber K, Jaffe AS, Mair J, Plebani M, Thygesen K, Lindahl B; Biomarker Study Group of the European Society of Cardiology Acute Cardiovascular Care Association. Editor’s Choice-Rule-in of acute myocardial infarction: Focus on troponin. Eur Heart J Acute Cardiovasc Care. 2017 Apr;6(3):212-217.
- Reichlin T, Cullen L, Parsonage WA, Greenslade J, Twerenbold R, Moehring B, Wildi K, Mueller S, Zellweger C, Mosimann T, Rubini Gimenez M, Rentsch K, Osswald S, Müller C. Two-hour algorithm for triage toward rule-out and rule-in of acute myocardial infarction using high-sensitivity cardiac troponin T. Am J Med. 2015 Apr;128(4):369-79. e4. https://doi.org/10.1016/j.amjmed.2014.10.032. [CrossRef]
- Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O’Briant KC, Allen A, Lin DW, Urban N, Drescher CW, Knudsen BS, Stirewalt DL, Gentleman R, Vessella RL, Nelson PS, Martin DB, Tewari M. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A. 2008 Jul 29;105(30):10513-8. https://doi.org/10.1073/pnas.0804549105. [CrossRef]
- Zampetaki A, Willeit P, Drozdov I, Kiechl S, Mayr M. Profiling of circulating microRNAs: from single biomarkers to re-wired networks. Cardiovasc Res. 2012 Mar 15;93(4):555-62. https://doi.org/10.1093/cvr/cvr266. [CrossRef]
- Felekkis K, Papaneophytou C. Challenges in Using Circulating Micro-RNAs as Biomarkers for Cardiovascular Diseases. Int J Mol Sci. 2020 Jan 15;21(2):561. https://doi.org/10.3390/ijms21020561. [CrossRef]
- Zhao Z, Guo N, Chen W, Wang Z. Leveraging Extracellular Non-coding RNAs to Diagnose and Treat Heart Diseases. J Cardiovasc Transl Res. 2022 Jun;15(3):456-468. https://doi.org/10.1007/s12265-022-10252-x. [CrossRef]
- Shen NN, Wang JL, Fu YP. The microRNA Expression Profiling in Heart Failure: A Systematic Review and Meta-Analysis. Front Cardiovasc Med. 2022 Jun 15; 9:856358. https://doi.org/10.3389/fcvm.2022.856358. [CrossRef]
- Laggerbauer B, Engelhardt S. MicroRNAs as therapeutic targets in cardiovascular disease. J Clin Invest. 2022 Jun 1;132(11): e159179. https://doi.org/10.1172/jci159179. [CrossRef]
- Yan B, Wang H, Tan Y, Fu W. microRNAs in Cardiovascular disease: Small Molecules but Big Roles. Curr. Top Med. Chem. 2019, 19, 1918–1947. [CrossRef]
- Lucas T, Bonauer A, Dimmeler S. RNA Therapeutics in Cardiovascular Disease. Circ Res. 2018 Jul 6;123(2):205-220. https://doi.org/10.1161/circresaha.117.311311. [CrossRef]
- Parizadeh SM, Ferns GA, Ghandehari M, Hassanian SM, Ghayour-Mobarhan M, Parizadeh SMR, Avan A. The diagnostic and prognostic value of circulating microRNAs in coronary artery disease: A novel approach to disease diagnosis of stable CAD and acute coronary syndrome. J Cell Physiol. 2018 Sep ;233(9):6418-6424. https://doi.org/10.1002/jcp.26324. [CrossRef]
- Hang P, Guo J, Sun C, Du Z. MicroRNAs as Candidate Drug Targets for Cardiovascular Diseases. Curr Drug Targets. 2017, 18, 463–472. https://doi.org/10.2174/1389450117666160301101221. [CrossRef]
- Zeller T, Keller T, Ojeda F, Reichlin T, Twerenbold R, Tzikas S, Wild PS, Reiter M, Czyz E, Lackner KJ, Munzel T, Mueller C, Blankenberg S Assessment of microRNAs in patients with unstable angina pectoris. Eur Heart J. 2014 Aug 14;35(31):2106-14.
- D’Alessandra Y, Carena MC, Spazzafumo L, Martinelli F, Bassetti B, Devanna P, Rubino M, Marenzi G, Colombo GI, Achilli F, Maggiolini S, Capogrossi MC, Pompilio G.Diagnostic potential of plasmatic MicroRNA signatures in stable and unstable angina. PLoS One. 2013 Nov 15;8(11): e80345. https://doi.org/10.1371/journal.pone.0080345. [CrossRef]
- Fichtlscherer S, De Rosa S, Fox H, Schwietz T, Fischer A, Liebetrau C, Weber M, Hamm CW, Röxe T, Müller-Ardogan M, Bonauer A, Zeiher AM, Dimmeler S. Circulating microRNAs in patients with coronary artery disease. Circ Res. 2010 Sep 3;107(5):677-84. https://doi.org/10.1161/circresaha.109.215566. [CrossRef]
- Ren J, Zhang J, Xu N, Han G, Geng Q, Song J, Li S, Zhao J, Chen H. Signature of circulating microRNAs as potential biomarkers in vulnerable coronary artery disease. PLoS One. 2013 Dec 5;8(12): e80738. https://doi.org/10.1371/journal.pone.0080738. [CrossRef]
- Lindsell CJ, Anantharaman V, Diercks D, Han JH, Hoekstra JW, Hollander JE, Kirk JD, Lim SH, Peacock WF, Tiffany B, Wilke EK, Gibler WB, Pollack CV Jr; EMCREG-International i*trACS Investigators. The Internet Tracking Registry of Acute Coronary Syndromes (i*trACS): a multicenter registry of patients with suspicion of acute coronary syndromes reported using the standardized reporting guidelines for emergency department chest pain studies. Ann Emerg Med. 2006 Dec;48(6):666-77, 677.e1-9.
- Su M, Niu Y, Dang Q, Qu J, Zhu D, Tang Z, Gou D. Circulating microRNA profiles based on direct S-Poly(T)Plus assay for detection of coronary heart disease. Cell Mol Med. 2020 Jun;24(11):5984-5997. https://doi.org/10.1111/jcmm.15001. [CrossRef]
- Wang GK, Zhu JQ, Zhang JT, Li Q, Li Y, He J, Qin YW, Jing Q. Circulating microRNA: a novel potential biomarker for early diagnosis of acute myocardial infarction in humans, Eur. Heart J. 2010 Mar;31(6):659-66. https://doi.org/10.1093/eurheartj/ehq013. [CrossRef]
- Devaux Y, Vausort M, Goretti E, Nazarov PV, Azuaje F, Gilson G, Corsten MF, Schroen B, Lair ML, Heymans S, Wagner DR. Use of circulating microRNAs to diagnose acute myocardial infarction. Clin Chem. 2012 Mar;58(3):559-67. https://doi.org/10.1373/clinchem.2011.173823. [CrossRef]
- Li YQ, Zhang MF, Wen HY, Hu CL, Liu R, Wei HY, Ai CM, Wang G, Liao XX, Li X. Comparing the diagnostic values of circulating microRNAs and cardiac troponin T in patients with acute myocardial infarction. Clinics (Sao Paulo). 2013 Jan;68(1):75-80. https://doi.org/10.6061/clinics/2013(01)oa12. [CrossRef]
- Cheng C, Wang Q, You W, Chen M, Xia J. MiRNAs as biomarkers of myocardial infarction: a meta-analysis. PLoS One. 2014 Feb 12;9(2): e88566. https://doi.org/10.1371/journal.pone.0088566. [CrossRef]
- Ward JA, Esa N, Pidikiti R, Freedman JE, Keaney JF, Tanriverdi K, Vitseva O, Ambros V, Lee R, McManus DD. Circulating Cell and Plasma microRNA Profiles Differ between Non-ST-Segment and ST-Segment-Elevation Myocardial Infarction. Fam Med Med Sci Res. 2013 Oct 1;2(2):108. https://doi.org/10.4172/2327-4972.1000108. [CrossRef]
- Widera C, Gupta SK, Lorenzen JM, Bang C, Bauersachs J, Bethmann K, Kempf T, Wollert KC, Thum T. Diagnostic and prognostic impact of six circulating microRNAs in acute coronary syndrome. J Mol Cell Cardiol. 2011 Nov;51(5):872-5. https://doi.org/10.1016/j.yjmcc.2011.07.011. [CrossRef]
- Oerlemans MI, Mosterd A, Dekker MS, de Vrey EA, van Mil A, Pasterkamp G, Doevendans PA, Hoes AW, Sluijter JP Early assessment of acute coronary syndromes in the emergency department: the potential diagnostic value of circulating microRNAs. EMBO Mol Med. 2012 Nov;4(11):1176-85.
- Bai R, Yang Q, Xi R, Li L, Shi D, Chen K. miR-941 as a promising biomarker for acute coronary syndrome. BMC Cardiovasc Disord. 2017 Aug 22 ;17(1) :227. [CrossRef]
- Wang F, Long G, Zhao C, Li H, Chaugai S, Wang Y, Chen C, Wang DW. Atherosclerosis-related circulating miRNAs as novel and sensitive predictors for acute myocardial infarction. PLoS One. 2014 Sep 3;9(9):e105734. [CrossRef]
- Wang A, Kwee LC, Grass E, Neely ML, Gregory SG, Fox KAA, Armstrong PW, White HD, Ohman EM, Roe MT, Shah SH, Chan MY. Whole blood sequencing reveals circulating microRNA associations with high-risk traits in non-ST-segment elevation acute coronary syndrome. Atherosclerosis. 2017 Jun; 261:19-25. https://doi.org/10.1016/j.atherosclerosis.2017.03.041. [CrossRef]
- Kaur A, Mackin ST, Schlosser K, Wong FL, Elharram M, Delles C, Stewart DJ, Dayan N, Landry T, Pilote L. Systematic review of microRNA biomarkers in acute coronary syndrome and stable coronary artery disease. Cardiovasc Res. 2020 May 1;116(6):1113-1124. [CrossRef]
- Zhelankin AV, Stonogina DA, Vasiliev SV, Babalyan KA, Sharova EI, Doludin YV, Shchekochikhin DY, Generozov EV, Akselrod AS. Circulating Extracellular miRNA Analysis in Patients with Stable CAD and Acute Coronary Syndromes. Biomolecules. 2021 Jun 29;11(7):962. [CrossRef]
- Chen MS, John JM, Chew DP, Lee DS, Ellis SG, Bhatt DL. Bare metal stent restenosis is not a benign clinical entity. Am Heart J. 2006 Jun;151(6):1260-4. [CrossRef]
- Bainey KR, Norris CM, Graham MM, Ghali WA, Knudtson ML, Welsh RC; APPROACH investigators. Clinical in-stent restenosis with bare metal stents: is it truly a benign phenomenon? Int J Cardiol. 2008 Aug 29;128(3):378-82.
- Park H, Ahn JM, Kang DY, Kim SO, Ko E, Kim TO, Lee PH, Lee SW, Park SW, Park DW, Park SJ. Very Long-term Safety and Effectiveness of Drug-Eluting or Bare-Metal Stents for Left Main Coronary Disease. [CrossRef]
- Joner M, Finn AV, Farb A, Mont EK, Kolodgie FD, Ladich E, Kutys R, Skorija K, Gold HK, Virmani R. Pathology of drug-eluting stents in humans: delayed healing and late thrombotic risk. J Am Coll Cardiol. 2006 Jul 4;48(1):193-202. [CrossRef]
- Nakazawa G, Finn AV, Virmani R. Vascular pathology of drug-eluting stents. Herz. 2007 Jun;32(4):274-80. [CrossRef]
- 8 May; Finn AV, Joner M, Nakazawa G, Kolodgie F, Newell J, John MC, Gold HK, Virmani R. Pathological correlates of late drug-eluting stent thrombosis: strut coverage as a marker of endothelialization. Circulation. 2007 May 8;115(18):2435-41. [CrossRef]
- Montelione N, Catanese V, Nenna A, Jawabra M, Verghi E, Loreni F, Nappi F, Lusini M, Mastroianni C, Jiritano F, Serraino GF, Mastroroberto P, Codispoti FA, Chello M, Spinelli F, Stilo F. The Diagnostic Value of Circulating Biomarkers and Role of Drug-Coated Balloons for In-Stent Restenosis in Patients with Peripheral Arterial Disease. Diagnostics (Basel). 2022 Sep 12;12(9):2207. https://doi.org/10.3390/diagnostics12092207. [CrossRef]
- Nusca A, Viscusi MM, Piccirillo F, De Filippis A, Nenna A, Spadaccio C, Nappi F, Chello C, Mangiacapra F, Grigioni F, Chello M, Ussia GP. In Stent Neo-Atherosclerosis: Pathophysiology, Clinical Implications, Prevention, and Therapeutic Approaches. Life (Basel). 2022 Mar 8;12(3):393. [CrossRef]
- Wang D, Deuse T, Stubbendorff M, Chernogubova E, Erben RG, Eken SM, Jin H, Li Y, Busch A, Heeger CH, Behnisch B, Reichenspurner H, Robbins RC, Spin JM, Tsao PS, Schrepfer S, Maegdefessel L. Local MicroRNA Modulation Using a Novel Anti-miR-21-Eluting Stent Effectively Prevents Experimental In Stent Restenosis. Arterioscler Thromb Vasc Biol. 2015 Sep ;35(9) :1945-53.
- Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A, Zhou Y, Persson R, King BD, Kauppinen S, Levin AA, Hodges MR. Treatment of HCV infec- tion by targeting microRNA. N Engl J Med. 2013, 368, 1685–1694.
- Hong DS, Kang YK, Borad M, Sachdev J, Ejadi S, Lim HY, Brenner AJ, Park K, Lee JL, Kim TY, Shin S, Becerra CR, Falchook G, Stoudemire J, Martin D, Kelnar K, Peltier H, Bonato V, Bader AG, Smith S, Kim S, O’Neill V, Beg MS. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer. 2020, 122, 1630–1637.
- Lee EC, Valencia T, Allerson C, Schairer A, Flaten A, Yheskel M, Kersjes K, Li J, Gatto S, Takhar M, Lockton S, Pavlicek A, Kim M, Chu T, Soriano R, Davis S, Androsavich JR, Sarwary S, Owen T, Kaplan J, Liu K, Jang G, Neben S, Bentley P, Wright T, Patel V. Discovery and preclinical evalua- tion of anti-miR-17 oligonucleotide RGLS4326 for the treatment of polycystic kidney disease. Nat Commun. 2019, 10, 1–14.
- van Zandwijk N, Pavlakis N, Kao SC, Linton A, Boyer MJ, Clarke S, Huynh Y, Chrzanowska A, Fulham MJ, Bailey DL, Cooper WA, Kritharides L, Ridley L, Pattison ST, MacDiarmid J, Brahmbhatt H, Reid G. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: a first-in-man, phase 1, openlabel, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396.
- Deng Y, Campbell F, Han K, Theodore D, Deeg M, Huang M, Hamatake R, Lahiri S, Chen S, Horvath G, Manolakopoulos S, Dalekos GN, Papatheodoridis G, Goulis I, Banyai T, Jilma B, Leivers M. Randomized clinical trials towards a single-visit cure for chronic hepatitis C: Oral GSK2878175 and injectable RG-101 in chronic hepatitis C patients and long-acting injectable GSK2878175 in healthy participants. J Viral Hepat. 2020, 27, 699–708. [CrossRef]
- van der Ree MH, van der Meer AJ, de Bruijne J, Maan R, van Vliet A, Welzel TM, Zeuzem S, Lawitz EJ, Rodriguez-Torres M, Kupcova V, Wiercinska-Drapalo A, Hodges MR, Janssen HL, Reesink HW. Long-term safety and efficacy of microRNA-targeted therapy in chronic hepatitis C patients. Antiviral Res. 2014; 111:53–59. 199.
- van der Ree MH, van der Meer AJ, van Nuenen AC, de Bruijne J, Ottosen S, Janssen HL, Kootstra NA, Reesink HW. Miravirsen dosing in chronic hepatitis C patients results in decreased microRNA-122 levels without affecting other microRNAs in plasma. Aliment Pharmacol Ther. 2016, 43, 102–113. [CrossRef]
- van der Ree MH, de Vree JM, Stelma F, Willemse S, van der Valk M, Rietdijk S, Molenkamp R, Schinkel J, van Nuenen AC, Beuers U, Hadi S, Harbers M, van der Veer E, Liu K, Grundy J, Patick AK, Pavlicek A, Blem J, Huang M, Grint P, Neben S, Gibson NW, Kootstra NA, Reesink HW. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: a phase 1B, double- blind, randomised controlled trial. Lancet. 2017, 389, 709–717. https://doi.org/10.1016/s0140-6736(16)31715-9. [CrossRef]
- Kilikevicius A, Meister G, Corey DR. Reexamining assumptions about miRNA-guided gene silencing. Reexamining assumptions about miRNA-guided gene silencing. Nucleic Acids Res. 2022, 50, 617–634.
Figure 1.
MicroRNAs with critical roles in the cardiovascular system, based on deregulation in disease or disease models and effects of their manipulation in vivo. The different boxes indicate the miRNA type and organism of evaluation (yellow), the disease condition and regulation, (blue), and the key experimental in vivo (green). Abbreviations; AF, atrial fibrillation; AngII, angiotensin II; CAD, coronary artery disease; CnA, human calcineurin subunit A; DCM, dilated cardiomyopathy; HF, heart failure; I/R, cardiac ischemia/reperfusion; CM, cardiac myocyte; EC, endothelial cell; KO, knockout; SMC, smooth muscle cell; MI; myocardial infarction; PPCM, peripartum cardiomyopathy; AAC/TAC, ascending/transverse aortic constriction. Ref. [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48].
Figure 1.
MicroRNAs with critical roles in the cardiovascular system, based on deregulation in disease or disease models and effects of their manipulation in vivo. The different boxes indicate the miRNA type and organism of evaluation (yellow), the disease condition and regulation, (blue), and the key experimental in vivo (green). Abbreviations; AF, atrial fibrillation; AngII, angiotensin II; CAD, coronary artery disease; CnA, human calcineurin subunit A; DCM, dilated cardiomyopathy; HF, heart failure; I/R, cardiac ischemia/reperfusion; CM, cardiac myocyte; EC, endothelial cell; KO, knockout; SMC, smooth muscle cell; MI; myocardial infarction; PPCM, peripartum cardiomyopathy; AAC/TAC, ascending/transverse aortic constriction. Ref. [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48].
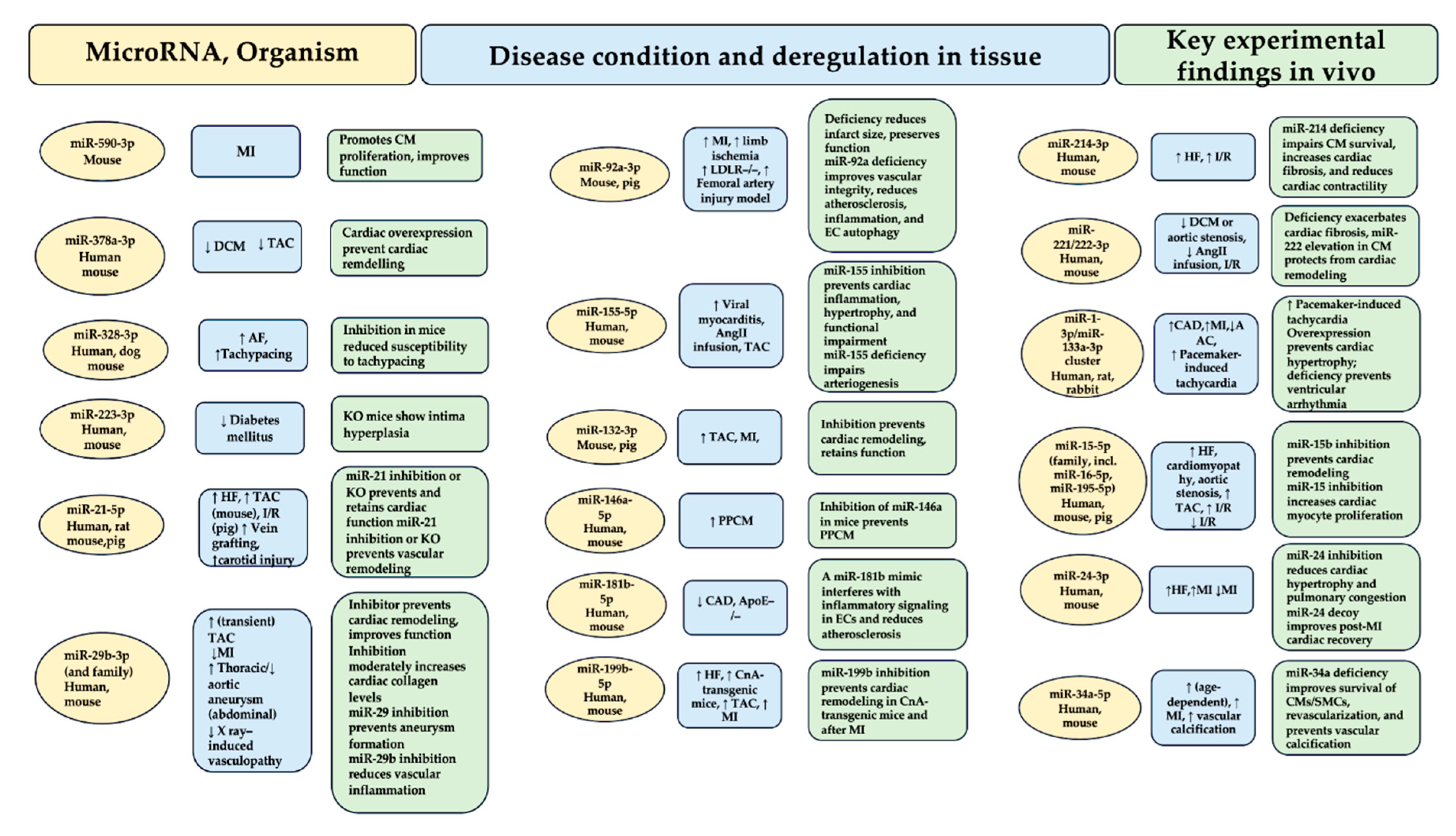
Figure 2.
In section A, the functions of microRNAs in the myocardium and vasculature are epitomized. The process promoted by the suggested microRNA is indicated by the + sign, while the - sign indicates that the pathophysiological process is prevented by the suggested microRNA. The respective columns contain information on the microRNAs that encourage or compromise cardiac function if their elevation or inhibition is recorded. Section B shows examples of microRNAs that control targets involved in cell-cell communication in the cardiovascular system (information gleaned from studies cited in Table 1, Table 2 and Table 3). Section C reports the paracrine roles of microRNA specimens secreted in the cardiovascular system. Atheroprotective effects are advocated by extracellular miR-126-3p (184) and miR-143-3p/miR-145-3p, proangiogenic effects are promoted by exosomal miR-143-3p, miR-222-5p, miR-92a -3p, and miR-214-3p, while miR-320-3p gives the opposite effect. The miR-21 passenger strand (3′) is enriched in exosomes from cardiac fibroblasts, inducing cardiac myocyte hypertrophy, while the miR-21 lead strand, released from endometrial mesenchymal stem cells, is cardioprotective promoting cell survival and angiogenesis. In a retrograde direction, several microRNAs of myocardial origin promote the mobilization of progenitor cells in the bone marrow. miR-223-3p is carried by platelets and regulates the differentiation and proliferation of vascular SMCs. For an overview of these and other cardiovascular microRNAs with proposed paracrine function. Abbreviations; EC, endothelial cell; SMC, smooth muscle cell. Refs. [20,52,53,54,55,56,57,58].
Figure 2.
In section A, the functions of microRNAs in the myocardium and vasculature are epitomized. The process promoted by the suggested microRNA is indicated by the + sign, while the - sign indicates that the pathophysiological process is prevented by the suggested microRNA. The respective columns contain information on the microRNAs that encourage or compromise cardiac function if their elevation or inhibition is recorded. Section B shows examples of microRNAs that control targets involved in cell-cell communication in the cardiovascular system (information gleaned from studies cited in Table 1, Table 2 and Table 3). Section C reports the paracrine roles of microRNA specimens secreted in the cardiovascular system. Atheroprotective effects are advocated by extracellular miR-126-3p (184) and miR-143-3p/miR-145-3p, proangiogenic effects are promoted by exosomal miR-143-3p, miR-222-5p, miR-92a -3p, and miR-214-3p, while miR-320-3p gives the opposite effect. The miR-21 passenger strand (3′) is enriched in exosomes from cardiac fibroblasts, inducing cardiac myocyte hypertrophy, while the miR-21 lead strand, released from endometrial mesenchymal stem cells, is cardioprotective promoting cell survival and angiogenesis. In a retrograde direction, several microRNAs of myocardial origin promote the mobilization of progenitor cells in the bone marrow. miR-223-3p is carried by platelets and regulates the differentiation and proliferation of vascular SMCs. For an overview of these and other cardiovascular microRNAs with proposed paracrine function. Abbreviations; EC, endothelial cell; SMC, smooth muscle cell. Refs. [20,52,53,54,55,56,57,58].
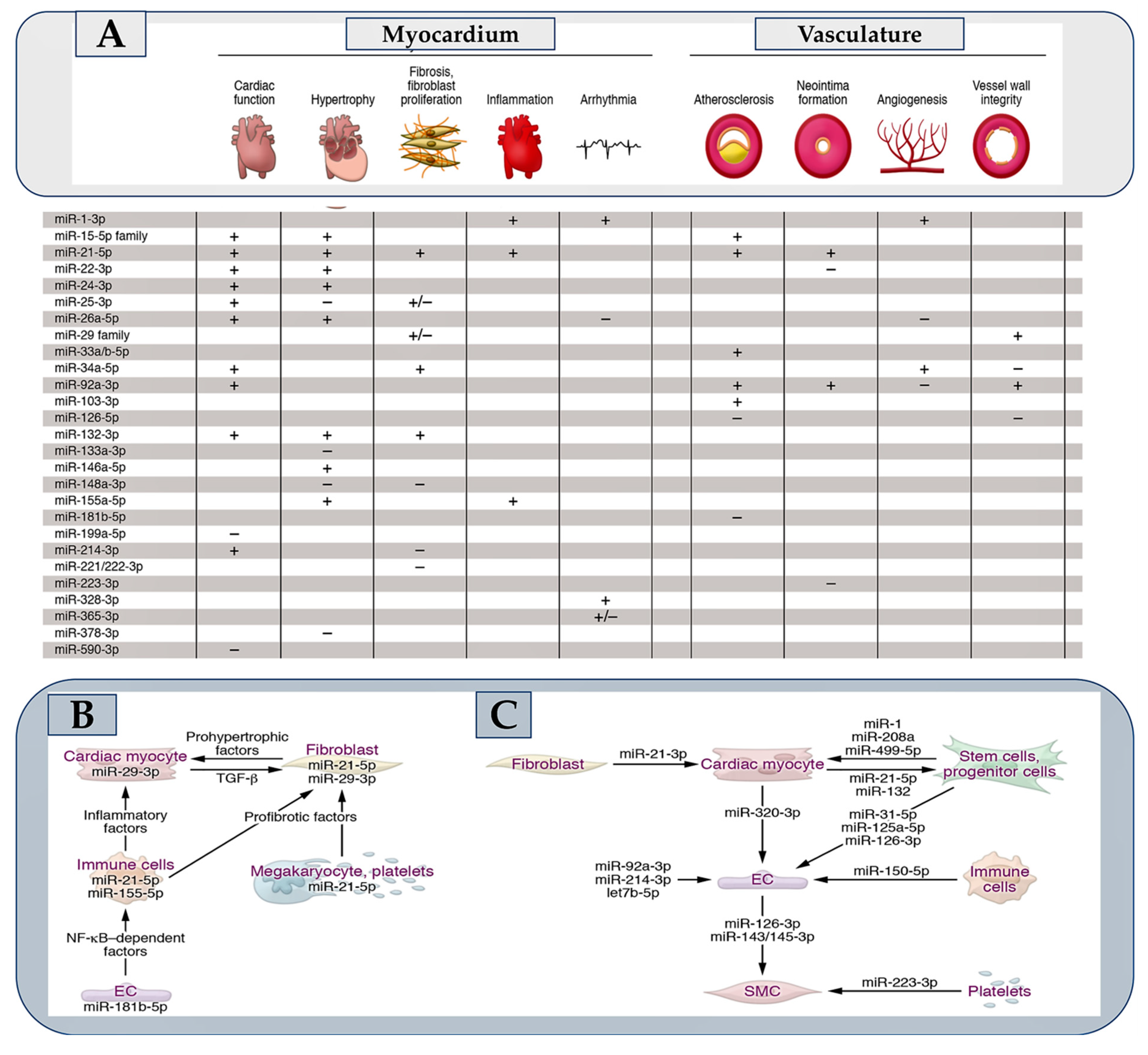
Figure 3.
The biogenesis and function of MicroRNA (miRNA) is reported. Nuclear pre-miRNAs are synthesized and released into the cytoplasm where 3 main steps promote the final synthesis of activated RNAs that progress alongside the production of precursor miRNA (miRNA duplex), RNA-induced silencing complex (RISC complex); and RNA activation (RNAi). A) Canonical elaboration, functional activation, mechanism of action, and degradation pathways of microRNAs are reported. The first step of canonical microRNA biogenesis initiates from greater hairpin RNA molecules (pre-miRNAs), which are produced by RNA Pol II transcription of microRNA genes or clusters, or which occur as part of introns. The second step is based on a microprocessor complex restraining the endonuclease Drosha, the DiGeorge critical region 8 protein (DGCR8), and other factors then cleaves these pre-miRNAs. Finally, the resulting pre-miRNA is exported to the cytoplasm, where the nuclease Dicer works to tailor from 21 to 22 nucleotides in length. Noncanonical mechanisms of microRNA biogenesis are also described, some of which bypass the microprocessor complex or Dicer. Following processing into a duplex of 21-22 nucleotides in length each, one strand called the leader strand, becomes part of the RNA-induced silencing complex (RISC), while the passenger strand (or *-strand) undergoes accelerated degradation. In case both strands are maintained individual functions can be adopted, as demonstrated for cardiovascular miR-21 and miR-126. Another exception is microRNA strands that localize to the nucleus, where they function in unusual ways. The degradation step of microRNAs includes the role of the exonucleases XRN-1, PNPase old-35, and RRP41 or the endonuclease Tudor-SN. The DIS3L2 nuclease degrades a subset of microRNAs after modification by terminal uridyltransferases (TUTases). Mechanisms of target-directed microRNA degradation (TDMD) have been resolved, including the involvement of ubiquitin ligases. (B) Pathways toward the identification and validation of disease-relevant cardiovascular microRNAs are reported. (C) Approaches for identifying microRNA targets are depicted. Refs. [52,70,78,79,93,94,95,96].
Figure 3.
The biogenesis and function of MicroRNA (miRNA) is reported. Nuclear pre-miRNAs are synthesized and released into the cytoplasm where 3 main steps promote the final synthesis of activated RNAs that progress alongside the production of precursor miRNA (miRNA duplex), RNA-induced silencing complex (RISC complex); and RNA activation (RNAi). A) Canonical elaboration, functional activation, mechanism of action, and degradation pathways of microRNAs are reported. The first step of canonical microRNA biogenesis initiates from greater hairpin RNA molecules (pre-miRNAs), which are produced by RNA Pol II transcription of microRNA genes or clusters, or which occur as part of introns. The second step is based on a microprocessor complex restraining the endonuclease Drosha, the DiGeorge critical region 8 protein (DGCR8), and other factors then cleaves these pre-miRNAs. Finally, the resulting pre-miRNA is exported to the cytoplasm, where the nuclease Dicer works to tailor from 21 to 22 nucleotides in length. Noncanonical mechanisms of microRNA biogenesis are also described, some of which bypass the microprocessor complex or Dicer. Following processing into a duplex of 21-22 nucleotides in length each, one strand called the leader strand, becomes part of the RNA-induced silencing complex (RISC), while the passenger strand (or *-strand) undergoes accelerated degradation. In case both strands are maintained individual functions can be adopted, as demonstrated for cardiovascular miR-21 and miR-126. Another exception is microRNA strands that localize to the nucleus, where they function in unusual ways. The degradation step of microRNAs includes the role of the exonucleases XRN-1, PNPase old-35, and RRP41 or the endonuclease Tudor-SN. The DIS3L2 nuclease degrades a subset of microRNAs after modification by terminal uridyltransferases (TUTases). Mechanisms of target-directed microRNA degradation (TDMD) have been resolved, including the involvement of ubiquitin ligases. (B) Pathways toward the identification and validation of disease-relevant cardiovascular microRNAs are reported. (C) Approaches for identifying microRNA targets are depicted. Refs. [52,70,78,79,93,94,95,96].
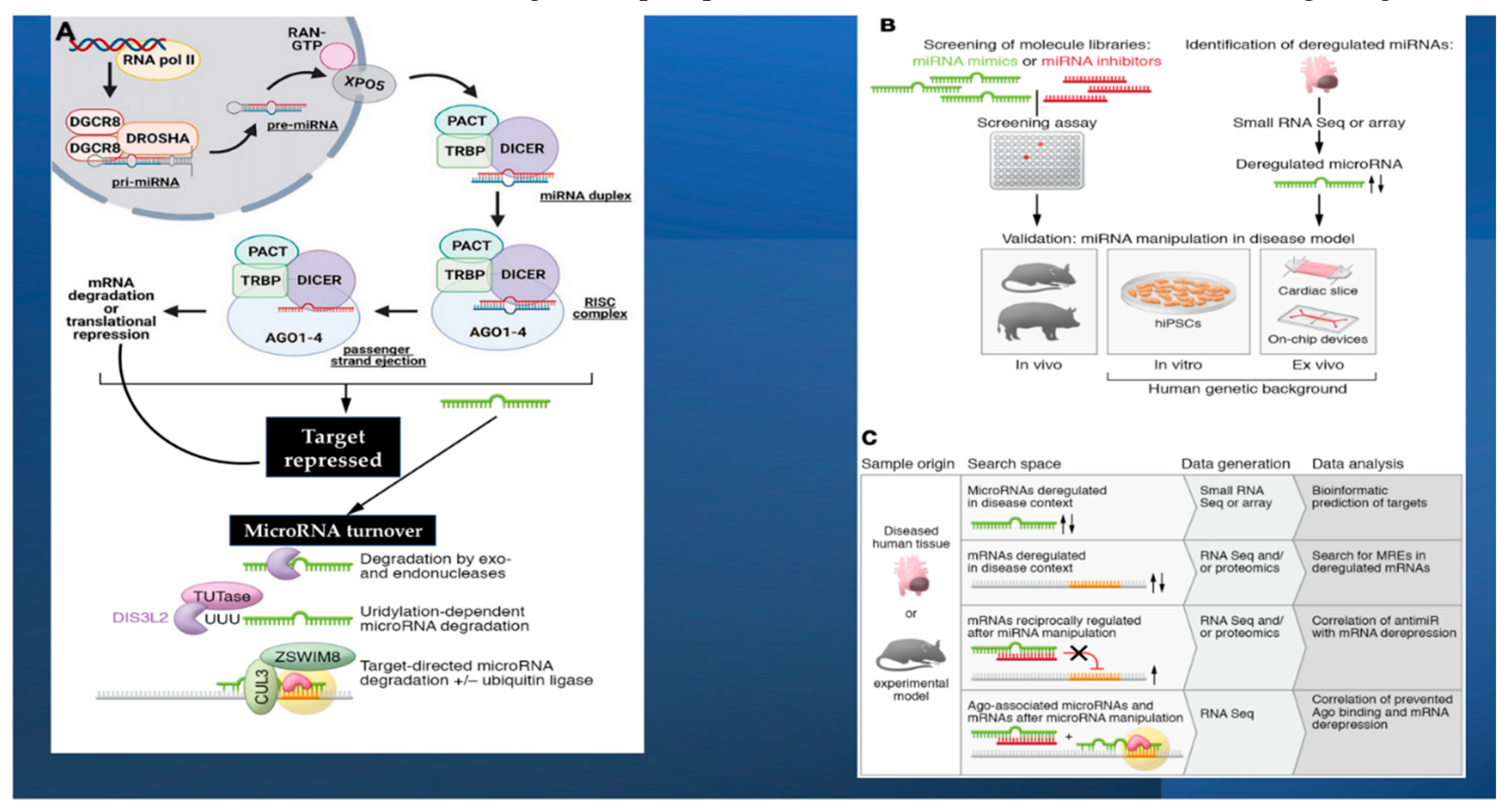
Figure 4.
Above Delivery of miRNAs into the bloodstream is promoted during STEMI. This process is the response to myocardial infarction involving impaired cardiomyocytes that release circulating miRNAs via protein complexes, microvescicles, exosomes, apoptotic bodies and/or mesenchymal stem cells, and thus are upregulated or downregulated. Below. miRNA may serve as a diagnostic, prognostic, and therapeutic pathway in patients with STEMI. Abbreviations: ST, ST Segment; ECG, electrocardiogram; ACS, acute coronary syndrome; STEMI, ST-segment elevation myocardial infarction; RNA, ribonucleic acid; miRNAs, microRNAs; MSCs, mesenchymal stem cells.
Figure 4.
Above Delivery of miRNAs into the bloodstream is promoted during STEMI. This process is the response to myocardial infarction involving impaired cardiomyocytes that release circulating miRNAs via protein complexes, microvescicles, exosomes, apoptotic bodies and/or mesenchymal stem cells, and thus are upregulated or downregulated. Below. miRNA may serve as a diagnostic, prognostic, and therapeutic pathway in patients with STEMI. Abbreviations: ST, ST Segment; ECG, electrocardiogram; ACS, acute coronary syndrome; STEMI, ST-segment elevation myocardial infarction; RNA, ribonucleic acid; miRNAs, microRNAs; MSCs, mesenchymal stem cells.
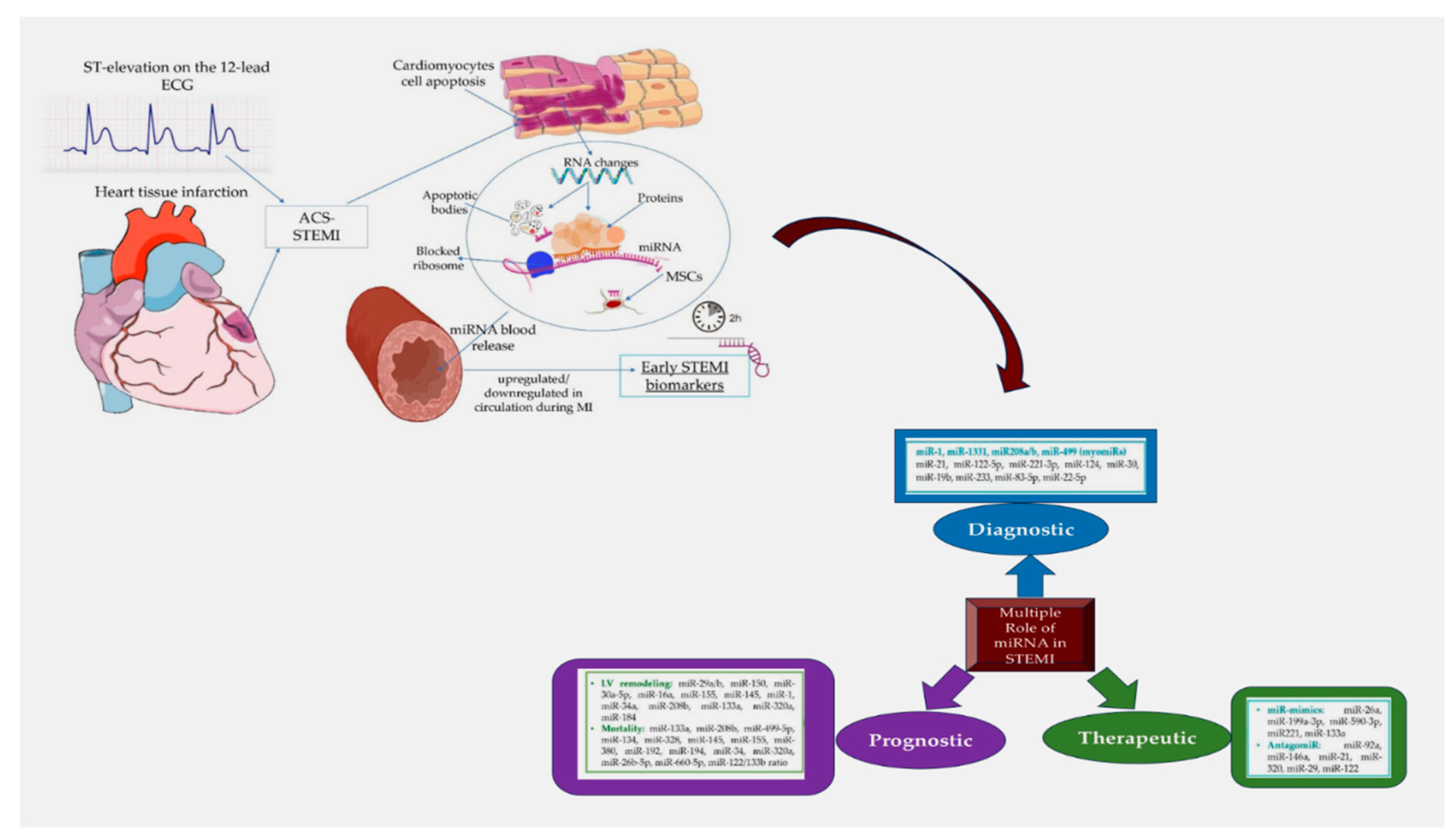
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



