
Submitted:
24 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
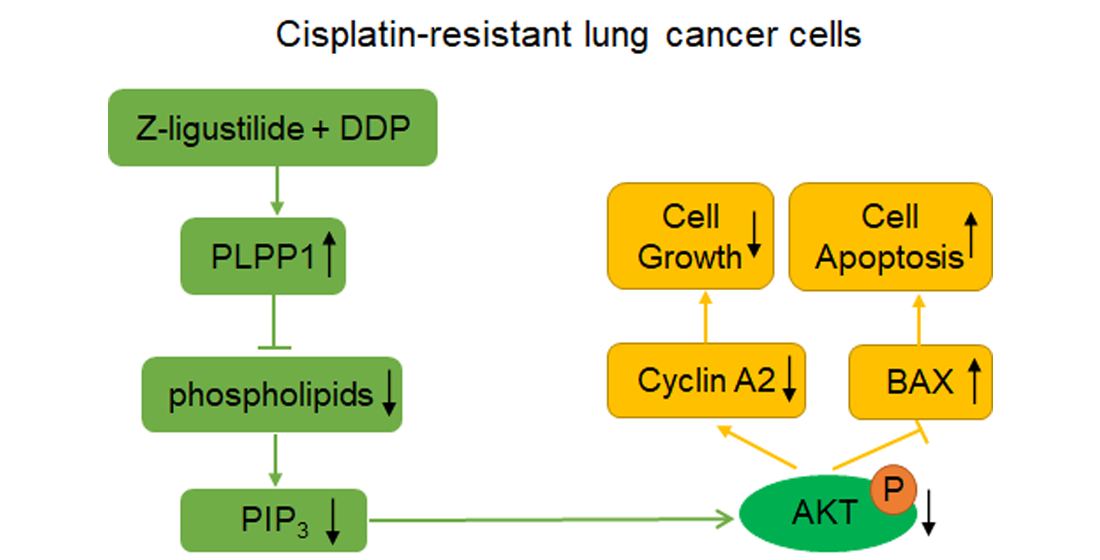
Z-ligustilide is a main active ingredient in the volatile oil of Angelica sinensis, a traditional Chinese medicine. Cisplatin is a commonly used chemotherapy drug for the treatment of lung cancer. Efficacy is often limited by the development of drug resistance after long-term treatment. Here, we investigated the effect of the combination of Z-ligustilide and cisplatin (Z-ligustilide+cisplatin) on resistance of cisplatin-resistant lung cancer cells and its action mechanism. Cell viability was analyzed by cell counting kit-8 (CCK-8) assay. Cell cycle and cell apoptosis were examined by flow cytometry assay. mRNA and protein levels of factors related to cell cycle and apoptosis were analyzed by real-time PCR and western blot. Liquid chromatography-mass spectrometry (LC-MS) analysis and RNA sequencing in A549, A549/DDP and A549/DDP cells treated with the Z-ligustilide+cisplatin were performed. The expression of PLPP1 was analyzed by the Cancer Genome Atlas (TCGA). The correlation between PLPP1 and prognosis was evaluated by immunohistochemistry and Kaplan-Meier (KM) plotter analysis. We found that Z-ligustilide+cisplatin could decrease the cell viability of cisplatin-resistant lung cancer cells. Z-ligustilide+cisplatin induced cell cycle arrest, and promoted cell apoptosis of cisplatin-resistant lung cancer cells. Metabolomics combined with transcriptomics revealed that Z-ligustilide+cisplatin inhibited phospholipid synthesis by upregulating the expression of PLPP1. Furthermore, PLPP1 expression was positively correlated with good prognosis. Knockdown of PLPP1 abolished the effects of Z-ligustilide+cisplatin on cell cycle and apoptosis. Z-ligustilide+cisplatin inhibited the activation of AKT by reducing the levels of PIP3 levels. Z-ligustilide+cisplatin induced cell cycle arrest and promoted cell apoptosis of cisplatin-resistant lung cancer cells by inhibiting PLPP1-mediated phospholipid synthesis.
Keywords:
cisplatin resistance
; Z-Ligustilide
; cell cycle arrest
; cell apoptosis
; phospholipid synthesis
; PLPP1
1. Introduction
Lung cancer is one of the most common malignant tumors in the world, and its incidence and mortality are increasing every year [1]. About 80% lung cancer patients are diagnosed with non-small lung cancer (NSCLC) [2]. Currently, platinum-based chemotherapeutics are the main chemotherapeutic methods for the treatment of lung cancer, among which cisplatin is a common chemotherapy drug [3,4]. During the treatment with cisplatin, patients often develop drug resistance, resulting in poor chemotherapy effect and tumor recurrence [5,6]. Therefore, how to overcome cisplatin resistance, enhance the sensitivity of cancer cells to cisplatin, and develop new therapeutic strategies are urgent clinical problems.
Z-ligustilide belongs to the phthalide class of compounds, and is mostly in the Angelica sinensis and Chuangxiong. Z-Ligustilide has been found to have anti-tumor activity in many tumors. For example, in glioblastoma, Z-ligustilide reduces cell migration by decreasing the expressions of the Rho GTPases [7]. In breast cancer, Z-ligustilide inhibits autophagy and accumulates DNA damages to overcome tamoxifen resistance [8]. In lung cancer, Z-ligustilide inhibits cell proliferation and promotes cell apoptosis by glycolysis [9]. But whether Z-ligustilide can reverse cisplatin resistance in cisplatin-resistant lung cancer has not yet been reported.
Metabolic reprogramming is a hallmark of malignancy [10]. Lipid metabolism reprogramming has gradually been recognized as a key mechanism for promoting cancer cell survival and proliferation [11,12], and is closely related to the development of drug resistance [13]. Gefitinib-resistant and gefitinib-sensitive NSCLC cells exhibited distinct phospholipids compositions, revealing phospholipid remodeling during drug resistance [14].
In this study, through molecular biology experiments, we confirmed that Z-ligustilide combined with cisplatin (Z-ligustilide+cisplatin) reduced the resistance of cisplatin-resistant lung cancer cells. Through lipid metabolomic and transcriptomic analysis, we revealed that PLPP1-mediated phospholipid synthesis participated in the reversal of resistance.
2. Results
2.1. Z-ligustilide alleviated the cisplatin resistance of cisplatin-resistant lung cancer cells
The chemical structures of cisplatin and Z-ligustilide are shown in Figure 1A. The effects of cisplatin and Z-ligustilide on cell viability of lung cancer cells and cisplatin-resistant lung cancer cells were investigated. CCK-8 assay revealed that A549 and H460 cells were more sensitive to cisplatin than A549/DDP and H460/DDP cells (Figure 1B).
Moreover, Z-ligustilide was able to reduce the cell viability of A549 and H460 cells more than that of A549/DDP and H460/DDP cells (Figure 1C). Further, A549/DDP and H460/DDP cells with various concentrations of Z-ligustilide alone or in combination with cisplatin for 24 h were treated, and CCK-8 assay demonstrated that 120 μM Z-ligustilide alone reduced cell viability of cisplatin-resistant lung cancer cells by less than 10%, however, Z-ligustilide combined with 20 μM cisplatin could decrease cell viability by more than 30% (Figure 1D). The results showed that the Z-ligustilide+cisplatin significantly decreased cell viability of cisplatin-resistant lung cancer cells.
2.2. Z-ligustilide+cisplatin induced cell cycle arrest and promoted cell apoptosis of cisplatin-resistant lung cancer cells
The effect of cisplatin and Z-ligustilide on cell proliferation was analyzed using cell cycle assay experiment. The results indicated that the Z-ligustilide+cisplatin induced cell cycle S-phase arrest in cisplatin-resistant lung cancer cells (Figure 2A). Considering that many drugs can induce cell cycle arrest at different stages, eventually leading to cell apoptosis [16,17], we examined the effect of Z-ligustilide and cisplatin on the apoptosis of cisplatin-resistant lung cancer cells. Control (Con), cisplatin, Z-ligustilide, Z-ligustilide+cisplatin were used to treat A549/DDP and H460/DDP cells. Cell apoptosis analysis confirmed that the Z-ligustilide+cisplatin significantly increased the proportion of apoptotic cells compared to the two compounds alone (Figure 2B). Real-time PCR showed a sharp downregulation of cell cycle regulatory proteins Cyclin A and Cyclin E, and a significant upregulation of apoptosis-related protein BAX, downregulation of Bcl-2 in A549/DDP and H460/DDP cells treated with Z-ligustilide+cisplatin (Figure S1). Western blot showed that the expressions of Cyclin A and Cyclin E were downregulated, and the expressions of Cleaved-Caspase 3 and BAX were upregulated in A549/DDP and H460/DDP cells treated with the Z-ligustilide+cisplatin (Figure 2C). Taken together, these data indicated that Z-ligustilide+cisplatin played an important role in inducing cell cycle arrest and cell apoptosis in cisplatin-resistant lung cancer cells.
2.3. Z-ligustilide+cisplatin reduced phospholipid synthesis and increased the expression of PLPP1
To further understand what happened, we wondered whether the resistance of cisplatin-resistant lung cancer cells was associated with lipid metabolism reprogramming, and whether Z-ligustilide+cisplatin could regulate lipid metabolism. Lipid metabolism profile of A549 cells, A549/DDP cells and A549/DDP cells treated with the Z-ligustilide and cisplatin were analyzed by LC-MS. A total of 32 lipids exhibited significant changes among three groups. Heat map showed that phospholipids obviously increased, and TGs decreased in A549/DDP cells compared with A549, however, A549/DDP cells treated with Z-ligustilide+cisplatin reversed the content of phospholipids and TGs (Figure 3A). Pathway enrichment analysis showed that glycerophospholipid metabolism was significantly altered (Figure 3B). Further analysis revealed that A549/DDP cells had higher levels of phosphatidylcholine (PC), phosphatidylethanolamine (PE) and phosphatidylinositol (PI), and lower level of triacylglycerol (TG) than A549 cells. But the Z-ligustilide+cisplatin greatly altered the content of PC, PE, PI and TG in A549/DDP cells (Figure 3C).
To investigate the key factors regulated by Z-ligustilide+cisplatin, we carried out RNA sequencing analysis of A549 cells, A549/DDP cells and A549/DDP cells treated with the Z-ligustilide+cisplatin . Gene set enrichment analysis (GSEA) of RNA sequencing data revealed that genes involved in glycerophospholipid metabolism were highly enriched (Figure 3D). Through RNA sequencing analysis, phospholipid phosphatase 1 (PLPP1) had a significant change in A549 cells, A549/DDP cells and A549/DDP cells treated with Z-ligustilide+cisplatin (Figure 3E). The mRNA level and protein expression of PLPP1 were downregulated in A549/DDP and H460/DDP cells compared with those in A549 and H460 cells, but the Z-ligustilide+cisplatin reversed the expression of PLPP 1 (Figure S2 and Figure 3F).
Next, we explored the expression of PLPP1 in clinical samples. The Cancer Genome Atlas (TCGA) data set analysis showed that PLPP1 had lower expression in 483 lung adenocarcinoma (LUAD) and 486 lung squamous cell carcinoma (LUSC) than in corresponding normal tissues (Figure 4A). The immunohistochemical staining of PLPP1 in 76 lung cancer tissue samples was performed. The typical PLPP1 images are shown in Figure 4B. The results revealed that lung cancer patients with low protein expression of PLPP1 had poor overall survival (Figure 4C). Furthermore, we used the database of Kaplan-Meier (KM) plotter to analyze the association of PLPP1 mRNA expression and prognosis of lung cancer. We found that lung cancer patients with low PLPP1 expression had a shorter overall survival than patients with high PLPP1 expression during the 60-month follow-up (Figure 4D). A more detailed analysis revealed that the overall survival of lung cancer patients with low expression of PLPP1 was shorter than patients with high expression of PLPP1 in the chemotherapy group (Figure 4E). However, overall survival was not significantly different in the non-chemotherapy group (Figure 4F). Together, the results indicated that PLPP1 was low expressed and associated with poor prognosis in lung cancer.
2.4. Z-ligustilide+cisplatin reversed cisplatin resistance through PLPP1-mediated phospholipid synthesis
To investigate whether PLPP1 participated in the effect of Z-ligustilide+cisplatin on resistance of cisplatin-resistant lung cancer cells, we knocked down the expression of PLPP1 in A549/DDP and H460/DDP cells, the mRNA level and protein expression of PLPP1 were examined by real-time PCR and the western blot (Figure S3 and Figure 5A). PLPP1 silence restored Z-ligustilide+cisplatin-induced effects on cell viability, cell cycle and cell apoptosis in A549/DDP and H460/DDP cells (Figure 5B,C). The data demonstrated that PLPP1 was necessary for cell cycle arrest and cell apoptosis induced by the Z-ligustilide+cisplatin.
Phosphatidate phosphatase was reported to be able to regulate phospholipid synthesis [18,19]. LC-MS-based metabolomics showed that PLPP1 silence increased the content of metabolites in phospholipid synthesis in A549/DDP cells. Furthermore, the Z-ligustilide+cisplatin increased the phospholipid synthesis in PLPP1-knockdown A549/DDP cells than in control A549/DDP cells (Figure S4). Relative levels of total PC, PE, PI, PG, PS and TG are shown in Figure 6A. Various components of phospholipids, such as PS, PI, and PC, are involved in the activation of AKT [20,21,22] which plays important roles in cell cycle progression, cell apoptosis and resistance [23]. The western blot experiment confirmed that Z-ligustilide+cisplatin decreased the level of p-AKT, however, PLPP1 silence reversed the expression of p-AKT in A549/DDP and H460/DDP cells (Figure 6B). Subsequently, we examined the PI3K kinase activity and PIP3 levels based on ELISA assay. The results showed that Z-ligustilide+cisplatin had little effect on PI3K kinase activity in cisplatin-resistant lung cells and PLPP1-knockdown cisplatin-resistant lung cells (Figure 6C). But Z-ligustilide+cisplatin decreased the levels of PIP3 in cisplatin-resistant lung cells, knockdown of PLPP1 increased the PIP3 levels in cisplatin-resistant lung cells treated with Z-ligustilide+cisplatin (Figure 6D). The data demonstrated that Z-ligustilide+cisplatin induced the inactivation of AKT by inhibiting PLPP1-mediated phospholipid synthesis.
3. Discussion
Lung cancer is the malignant tumor with the highest morbidity and mortality in the world, and its five-year survival rate is less than 20% [24,25]. The early stage of lung cancer is relatively insidious, and the course of lung cancer develops rapidly. Most of the patients have been in the middle and late stages when they are diagnosed, they cannot undergo surgical resection due to the missed best treatment time [26]. At present, the treatment of NSCLC mostly adopts platinum drug-based combination chemotherapy. The most widely used chemotherapy drug is cisplatin, and cisplatin has the advantages of wide anti-cancer range and strong effect in the treatment of lung cancer. In the process, cisplatin shows a good therapeutic effect, but long-term application will cause tumor cells to develop resistance, resulting in poor therapeutic effect [27]. Therefore, in order to improve clinical treatment effect of patients with lung cancer, it is very important to find new drug against drug resistance.
Z-Ligustilide, also known as 3-butenyl-4,5-dihydro-1 (3H)-isobenzofuranone, is the main active ingredient of Chinese medicine Angelica volatile oil. In recent years, it is found that Z-ligustilide has anti-tumor activity in various tumors. For example, Z-ligustilide can induce cell death and oxidative stress by epigenetically transcriptional regulation of Nrf2 in prostate cancer [28]. In addition, Z-ligustilide can increase the sensitivity of breast cancer to tamoxifen and reduce chemoresistance during breast cancer treatment by inhibiting autophagy and promoting caspase-independent cell death [29]. However, the role of Z-ligustilide in cisplatin-resistant lung cancer has not been reported. In this study, we found that cell viability was obviously reduced, cell proliferation was inhibited and cell apoptosis was increased in cisplatin-resistant lung cancer cells with the treatment of Z-ligustilide+cisplatin.
PLPP1 catalyzed the formation of diacylglycerol (DAG) from phosphatidic acid (PA), and participated in lipid metabolism and signal transduction. It has been reported that loss of the phospholipid phosphatase Pah1 in yeast reduced DAG production, resulting in reducing level and synthesis of TGs and increasing level and synthesis of phospholipids [18,19]. Phospholipids are composed of glycerophospholipids and sphingomyelins. Glycerophospholipids include PC, PE, PS, PG, PI and PA. Changes in PC, PE, and PS can alter the properties of cell membranes and affect signal transduction and biological processes, thereby affecting tumor development and drug resistance [30,31,32]. We discovered that PLPP1 was upregulated in cisplatin-resistant lung cancer cells with the treatment of Z-ligustilide+cisplatin compared with cisplatin-resistant lung cancer cells. Low expression of PLPP1 was strongly associated with poor prognosis. PLPP1 silence alleviated the effects on cell viability, cell cycle, cell apoptosis and phospholipid synthesis mediated by Z-ligustilide+cisplatin. Altered phospholipids are closely related to the activation of the AKT signaling pathway [21]. We observed the inhibition of p-AKT expression and the decrease of PIP3 levels in cisplatin-resistant lung cancer treated with Z-ligustilide+cisplatin, however, the knockdown of PLPP1 reversed the effects. The results demonstrated that the decrease in phospholipids mediated by PLPP1 reduced the levels of PIP3, ultimately inhibiting the activation of AKT.
In summary, our study provided a potential therapeutic strategy for the treatment of cisplatin-resistant lung cancer, and elucidated the specific action mechanism of Z-ligustilide+cisplatin.
4. Materials and Methods
4.1. Cell culture
A549 (human lung adenocarcinoma cell line) was purchased from Shanghai Cell Bank of Chinese Academy of Sciences (Shanghai, China), and A549/DDP (cisplatin-resistant lung adenocarcinoma cell line) was purchased from BeNa Biotechnology Co., Ltd (Hebei, China). H460 (human lung cancer cell line) and H460/DDP (cisplatin-resistant lung cancer cell line) were purchased from YaJi Biological (Shanghai, China). These cells were cultured in DMEM/high glucose medium or RPMI 1640, containing 10% fetal bovine serum, 100 units/mL penicillin and 100 μg/mL streptomycin in an incubator with 5% CO2, 37 °C.
4.2. Reagents and antibodies
Cisplatin and Z-ligustilide were purchased from MedChemExpress Company (Shanghai, China). The following primary antibodies for BAX (Proteintech Technology, Wuhan, China ), PI3K (Proteintech Technology, Wuhan, China), PLPP1 (Proteintech Technology, Wuhan, China) and Cyclin A2 (Proteintech Technology, Wuhan, China), AKT (Cell Signaling Technology, Boston, MA, USA), phosphor-Akt (Ser473) (Cell Signaling Technology, Boston, MA, USA), Cleaved-Caspase 3 (Cell Signaling Technology, Boston, MA, USA) and GAPDH (Cell Signaling Technology, Boston, MA, USA), PI3K p85α (phospho-Tyr607) (Bioworld Technology, Minnesota, USA), Cyclin E1 (HUABIO Technology, Hangzhou, China) and PPAP2A (PLPP1) (Novus Technology, Colorado, USA) were used.
4.3. Cell counting kit-8 assay
Cell viability was detected by cell counting kit-8 (CCK-8) kit (Beyotime, Shanghai, China). Briefly, cells were seeded in 96-well plates containing 3x103 cells and 100 μL medium per well. After the cells adhered, different doses of Z-ligustilide and cisplatin were added to stimulate the cells for 24 h. 10 μL CCK-8 solution was added to each well, and placed in a 37 °C incubator for 1 h in the dark. Then the absorbance was measured at 450 nm.
4.4. Cell apoptosis assay
PE Annexin V Apoptosis Detection Kit (BD Pharmingen, San Diego, CA) was used to assess the percentage of apoptotic cells. Firstly, the cells were treated with cisplatin and Z-ligustilide alone or in combination for 24 h. Then, the cells were washed with PBS, digested with trypsin, and centrifuged. Next, 200 μL 1X Binding Buffer was added to disperse the cells, and 5 μL PE Annexin V and 5 μL 7-AAD were added to mix gently, the cells were incubated at room temperature for 15 min away from light. Finally, the apoptotic cell rates were analyzed by flow cytometry.
4.5. Cell cycle assay
Cisplatin- and Z-ligustilide-stimulated cells were fixed with prechilled 70% ethanol overnight, and then the cells were analyzed by Cycle and Apoptosis Analysis Kit (Beyotime, Shanghai, China) according to the manufacturer’s instruction.
4.6. Liquid chromatography-mass spectrometry analysis
The cells were seeded in 10 cm dishes and stimulated with cisplatin and Z-ligustilide alone or in combination for 24 h. The cells were washed three times with 10 mL PBS, and quenched with liquid nitrogen. Next, cells were collected with 1 mL extractant (80% methanol containing internal standard), and added to chloroform and ultrapure water sequentially, and vortexed for 30 s. Centrifuge at 13000 g for 15 min at 4 °C. 400 μL hydrophobic layer was aspirated and then lyophilized. Quality control (QC) samples were lyophilized in the same volume. Add 30 μL chloroform/methanol (2: 2, v: v) solution, vortex for 30 s to reconstitute, and then add 70 μL acetonitrile/isopropanol/MilliQ water (65: 30: 5) containing 5 mM ammonium acetate, vortex for 30 s to dilute.
UPLC-Q-Exactive HF MS was used for nontargeted lipidomic analysis. The column was an ACQUITY UPLC C8 Column (100 x 2.1 mm, 1.7 μm). The column temperature was 60℃ and the flow rate was 0.3 mL/min. The injection chamber temperature was 10℃. Injection volume was 5 μL. Mobile phase A was 60% acetonitrile/MilliQ water containing 10 mM ammonium acetate, and phase B was 10% acetonitrile/isopropanol containing 10 mM ammonium acetate. Mass spectrometry ion source sheath gas flow rate was 45 arb, and auxiliary gas flow rate was 10 arb. Spray voltage was 3.5 kV (ESI+) and 3.0 kV (ESI-). Capillary temperature was 320℃, and auxiliary gas temperature was 350 °C. Resolution was 12e4. The full scanning modes was used, and scan ranges was 200-1100 m/z (ESI+) and 120-1600 m/z (ESI-).
4.7. RNA sequencing
Total RNA was isolated using a TRIzol total RNA extractibn kit (Tiangen, Beijing, China), which yielded > 2 μg of total RNA per sample. RNA quality was examined by 0.8% agarose gel electrophoresis and spectrophotometry. Illumina HiSeq library construction was performed according to the manufacturer's instructions (Illumina, California, USA). Oligo-dT primers were used to transverse mRNA to obtain cDNA (APExBIO, Houston, USA). The second cDNA strand was synthesized by amplifying the cDNA, and was purified using magnetic beads. After library construction, library fragments were enriched by PCR amplification and selected according to a fragment size of 350-550 bp. The library was quality-assessed using an Agilent 2100 Bioanalyzer (Agilent, California, USA). The library was sequenced using the Illumina NovaSeq 6000 sequencing platform (Paired end150) to generate raw reads.
4.8. Western blot
Cells were harvested and lysed with RIPA (Beyotime, Shanghai, China) containing protein and phosphatase inhibitors (APExBIO, Houston, USA). Total protein extract (20 μg) was loaded in 4-20% SDS-PAGE. The protein was transferred to PVDF membrane, and was blocked with 5% milk. Subsequently, the protein was incubated with primary antibodies and second HRP linked antibodies. Finally, the protein was visualized using ECL Chemiluminescence Chromogenic Solution (Tanon, Shanghai, China).
4.9. Real-time PCR
Total RNA was isolated with Trizol Reagent Kit (Takara, Tokyo, Japan) according to the manufacturer’s instructions. The cDNA was generated by using PrimeScript RT reagent Kit with gDNA Eraser (Takara, Tokyo, Japan). Real-time PCR was carried out by using TB Green Premix Ex Taq II (Takara, Tokyo, Japan) operating on LightCycler 96 Real-time PCR system. The primers sequencings were as follows:
GAPDH (forward): TCCAAAATCAAGTGGGGCGA
GAPDH (reverse): TGATGACCCTTTTGGCTCCC
Cyclin A2 (forward): GATTTCGTCTTCCAGCAGCAG
Cyclin A2 (reverse): ATCTGACAAGCATCGGGACC
Cyclin E1 (forward): AGAGGAAGGCAAACGTGACC
Cyclin E1 (reverse): GAGGCTTGCACGTTGAGTTT
BCL2 (forward): GAACTGGGGGAGGATTGTGG
BCL2 (reverse): CCGTACAGTTCCACAAAGGC
BAX (forward): GACATTGGACTTCCTCCGGG
BAX (reverse): GGGACATCAGTCGCTTCAGT
PLPP1 (forward): CTGGCTGGATTGCCTTTTGC
PLPP1 (reverse): AACAGACAGGGTTTCTCCAAGA
4.10. Immunohistochemistry
Human lung cancer tissue chip was purchased from Outdo Biotech (Shanghai, China). The tissue chip used was HLugA180Su04. Immunohistochemical procedures and evaluation methods were performed [15].
4.11. PIP3 measurements
PIP3 levels were analyzed by Human PIP3 ELISA Kit according to the manufacturer’s instructions (COIBO BIO, Shanghai, China).
4.12. PI3K kinase assay
PI3K kinase activity was examined by Human PI3K ELISA Kit according to the manufacturer’s instructions (COIBO BIO, Shanghai, China).
4.13. Statistical analysis
The experiments are repeated three or more times. Data are presented as mean ± SD. Statistical significance was measured by Student’s t-test (unpaired, two-tailed) with p<0.05 indicating significance. Statistical analysis was performed with Prism software (La Jolla, CA, USA), FlowJo software (TreeStar, Ashland, OR), SIMCA-P software (Umetrics, Umea, Sweden) and Multi Experiment Viewer (http://www.tm4.org).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: The Z-ligustilide+cisplatin induced cell apoptosis. mRNA expression of Cyclin A2, Cyclin E1, BAX and Bcl-2 in A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination. **p < 0.01 and ***p < 0.001. Figure S2: The Z-ligustilide+cisplatin reversed the expression of PLPP1. mRNA level of PLPP1 was determined by real-time PCR in A549 and H460, A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination. ***p < 0.001. Figure S3: The validation of PLPP1 knockdown efficiency. mRNA expression of PLPP1 in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells, respectively. ***p < 0.001. Figure S4: Heat map of the differential lipids in shCtrl and PLPP1 knockdown A549/DDP cells with or without Z-ligustilide+cisplatin treatment. Supplementary File: Uncropped western blots.
Author Contributions
P.Y.G. and G.W.X. designed the experiment framework and wrote the manuscript. P.Y.G. conducted the most experiments. J.H.Z. performed the RNA sequencing analysis. Q.L. and X.L.W. conducted the experiments that were associated with metabolomics. W.S.Q. and T.W. assisted in the process of experiments. X.X.S., X.Y.L., J.C. and H.D.Q. provided the guiding suggestions. All authors have read and agreed to the published version of the manuscript.
Funding
The study is supported by the CAS “Light of West China” Program, the National Natural Science Foundation of China (21934006), the Youth Innovation Promotion Association CAS (2021186) and Dalian Science and Technology Talent Innovation Support Program (2022RQ026).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare no conflict of interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J Clin 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Kocher, F.; Hilbe, W.; Seeber, A.; Pircher, A.; Schmid, T.; Greil, R.; Auberger, J.; Nevinny-Stickel, M.; Sterlacci, W.; Tzankov, A.; et al. Longitudinal analysis of 2293 NSCLC patients: a comprehensive study from the TYROL registry. Lung Cancer 2015, 87, 193–200. [Google Scholar] [CrossRef]
- Rosell, R.; Cecere, F.; Santarpia, M.; Reguart, N.; Taron, M. Predicting the outcome of chemotherapy for lung cancer. Curr Opin Pharmacol 2006, 6, 323–331. [Google Scholar] [CrossRef]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat Rev Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef]
- Liu, M.Y.; Li, X.Q.; Gao, T.H.; Cui, Y.; Ma, N.; Zhou, Y.; Zhang, G.J. Elevated HOTAIR expression associated with cisplatin resistance in non-small cell lung cancer patients. J Thorac Dis 2016, 8, 3314–3322. [Google Scholar] [CrossRef]
- Li, W.; Liu, X.; Zhang, G.; Zhang, L. [Role of SOX4 on DDP Resistance in Non-small Cell Lung Cancer Cell of A549]. Zhongguo Fei Ai Za Zhi 2017, 20, 298–302. [Google Scholar] [CrossRef]
- Yin, J.; Wang, C.; Mody, A.; Bao, L.; Hung, S.H.; Svoronos, S.A.; Tseng, Y. The Effect of Z-Ligustilide on the Mobility of Human Glioblastoma T98G Cells. PLoS One 2013, 8, e66598. [Google Scholar] [CrossRef]
- Ma, H.; Li, L.; Dou, G.; Wang, C.; Li, J.; He, H.; Wu, M.; Qi, H. Z-ligustilide restores tamoxifen sensitivity of ERa negative breast cancer cells by reversing MTA1/IFI16/HDACs complex mediated epigenetic repression of ERa. Oncotarget 2017, 8, 29328–29345. [Google Scholar] [CrossRef]
- Jiang, X.; Zhao, W.; Zhu, F.; Wu, H.; Ding, X.; Bai, J.; Zhang, X.; Qian, M. Ligustilide inhibits the proliferation of non-small cell lung cancer via glycolytic metabolism. Toxicol Appl Pharmacol 2021, 410, 115336. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368. [Google Scholar] [CrossRef]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, Q.; Halim, A.; Song, G. Targeting lipid metabolism of cancer cells: A promising therapeutic strategy for cancer. Cancer Lett 2017, 401, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.W.; Wilkins, O.; Bang, S.; Ung, M.; Li, J.; An, J.; Del Genio, C.; Canfield, K.; DiRenzo, J.; Wells, W.; et al. CD36-Mediated Metabolic Rewiring of Breast Cancer Cells Promotes Resistance to HER2-Targeted Therapies. Cell Rep 2019, 29, 3405–3420. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Lee, M.Y.; Choi, D.Y.; Lee, J.W.; You, S.; Lee, K.Y.; Kim, J.; Kim, K.P. Phospholipids of tumor extracellular vesicles stratify gefitinib-resistant nonsmall cell lung cancer cells from gefitinib-sensitive cells. Proteomics 2015, 15, 824–835. [Google Scholar] [CrossRef]
- Feng, X.; Liu, H.; Zhang, Z.; Gu, Y.; Qiu, H.; He, Z. Annexin A2 contributes to cisplatin resistance by activation of JNK-p53 pathway in non-small cell lung cancer cells. J Exp Clin Cancer Res 2017, 36, 123. [Google Scholar] [CrossRef] [PubMed]
- Gamet-Payrastre, L.; Li, P.; Lumeau, S.; Cassar, G.; Dupont, M.A.; Chevolleau, S.; Gasc, N.; Tulliez, J.; Terce, F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res 2000, 60, 1426–1433. [Google Scholar] [PubMed]
- Zhong, S.; Li, Y.G.; Ji, D.F.; Lin, T.B.; Lv, Z.Q. Protocatechualdehyde Induces S-Phase Arrest and Apoptosis by Stimulating the p27(KIP1)-Cyclin A/D1-CDK2 and Mitochondrial Apoptotic Pathways in HT-29 Cells. Molecules 2016, 21, 934. [Google Scholar] [CrossRef]
- Carman, G.M.; Han, G.S. Phosphatidate phosphatase regulates membrane phospholipid synthesis via phosphatidylserine synthase. Adv Biol Regul 2018, 67, 49–58. [Google Scholar] [CrossRef]
- Kwiatek, J.M.; Han, G.S.; Carman, G.M. Phosphatidate-mediated regulation of lipid synthesis at the nuclear/endoplasmic reticulum membrane. Biochim Biophys Acta Mol Cell Biol Lipids 2020, 1865, 158434. [Google Scholar] [CrossRef]
- Kim, H.Y.; Huang, B.X.; Spector, A.A. Phosphatidylserine in the brain: metabolism and function. Prog Lipid Res 2014, 56, 1–18. [Google Scholar] [CrossRef]
- Rueda-Rincon, N.; Bloch, K.; Derua, R.; Vyas, R.; Harms, A.; Hankemeier, T.; Khan, N.A.; Dehairs, J.; Bagadi, M.; Binda, M.M.; et al. p53 attenuates AKT signaling by modulating membrane phospholipid composition. Oncotarget 2015, 6, 21240–21254. [Google Scholar] [CrossRef] [PubMed]
- Pein, H.; Koeberle, S.C.; Voelkel, M.; Schneider, F.; Rossi, A.; Thurmer, M.; Loeser, K.; Sautebin, L.; Morrison, H.; Werz, O.; et al. Vitamin A regulates Akt signaling through the phospholipid fatty acid composition. FASEB J 2017, 31, 4566–4577. [Google Scholar] [CrossRef] [PubMed]
- Revathidevi, S.; Munirajan, A.K. Akt in cancer: Mediator and more. Semin Cancer Biol 2019, 59, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Nasim, F.; Sabath, B.F.; Eapen, G.A. Lung Cancer. Med Clin North Am 2019, 103, 463–473. [Google Scholar] [CrossRef]
- Jones, G.S.; Baldwin, D.R. Recent advances in the management of lung cancer. Clin Med (Lond) 2018, 18, s41–s46. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, V.M.L.; Carvalho, L. Heterogeneity in Lung Cancer. Pathobiology 2018, 85, 96–107. [Google Scholar] [CrossRef]
- Jonna, S.; Subramaniam, D.S. Molecular diagnostics and targeted therapies in non-small cell lung cancer (NSCLC): an update. Discov Med 2019, 27, 167–170. [Google Scholar]
- Su, Z.Y.; Khor, T.O.; Shu, L.; Lee, J.H.; Saw, C.L.; Wu, T.Y.; Huang, Y.; Suh, N.; Yang, C.S.; Conney, A.H.; et al. Epigenetic reactivation of Nrf2 in murine prostate cancer TRAMP C1 cells by natural phytochemicals Z-ligustilide and Radix angelica sinensis via promoter CpG demethylation. Chem Res Toxicol 2013, 26, 477–485. [Google Scholar] [CrossRef]
- Qi, H.; Jiang, Z.; Wang, C.; Yang, Y.; Li, L.; He, H.; Yu, Z. Sensitization of tamoxifen-resistant breast cancer cells by Z-ligustilide through inhibiting autophagy and accumulating DNA damages. Oncotarget 2017, 8, 29300–29317. [Google Scholar] [CrossRef]
- Dobrzynska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol Cell Biochem 2005, 276, 113–119. [Google Scholar] [CrossRef]
- Lee, S.; Jang, W.J.; Choi, B.; Joo, S.H.; Jeong, C.H. Comparative metabolomic analysis of HPAC cells following the acquisition of erlotinib resistance. Oncol Lett 2017, 13, 3437–3444. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Samadzadeh, K.M.; Nitin, N. Rapid assessment of drug resistance of cancer cells to gefitinib and carboplatin using optical imaging. Anal Biochem 2016, 504, 50–58. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Z-ligustilide inhibited cell viability of cisplatin-resistant lung cancer cells. (A) Chemical structures of Cisplatin and Z-ligustilide (Z-LIG). (B) Cell viability of A549 cells and cisplatin-resistant A549 (A549/DDP) cells, H460 cells and cisplatin-resistant H460 (H460/DDP) cells after treatment with cisplatin. (C) Cell viability of A549 cells and A549/DDP, H460 cells and H460/DDP cells after treatment with Z-ligustilide. (D) Cell viability of A549/DDP cells and H460/DDP cells after treatment with Z-ligustilide+cisplatin. ns, no significant, *p < 0.05, **p < 0.01 and ***p < 0.001.
Figure 1.
Z-ligustilide inhibited cell viability of cisplatin-resistant lung cancer cells. (A) Chemical structures of Cisplatin and Z-ligustilide (Z-LIG). (B) Cell viability of A549 cells and cisplatin-resistant A549 (A549/DDP) cells, H460 cells and cisplatin-resistant H460 (H460/DDP) cells after treatment with cisplatin. (C) Cell viability of A549 cells and A549/DDP, H460 cells and H460/DDP cells after treatment with Z-ligustilide. (D) Cell viability of A549/DDP cells and H460/DDP cells after treatment with Z-ligustilide+cisplatin. ns, no significant, *p < 0.05, **p < 0.01 and ***p < 0.001.
Figure 2.
Z-ligustilide+cisplatin induced S phase arrest and promoted cell apoptosis. (A) Cell cycle analysis of A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination for 24h. (B) PE Annexin V apoptosis analysis of A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination for 24h. (C) Protein expression of Cyclin A2, Cyclin E1, Cleaved-Caspase 3 and BAX in A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination. **p < 0.01 and ***p < 0.001.
Figure 2.
Z-ligustilide+cisplatin induced S phase arrest and promoted cell apoptosis. (A) Cell cycle analysis of A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination for 24h. (B) PE Annexin V apoptosis analysis of A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination for 24h. (C) Protein expression of Cyclin A2, Cyclin E1, Cleaved-Caspase 3 and BAX in A549/DDP and H460/DDP, A549/DDP and H460/DDP treated with cisplatin and Z-ligustilide alone or in combination. **p < 0.01 and ***p < 0.001.
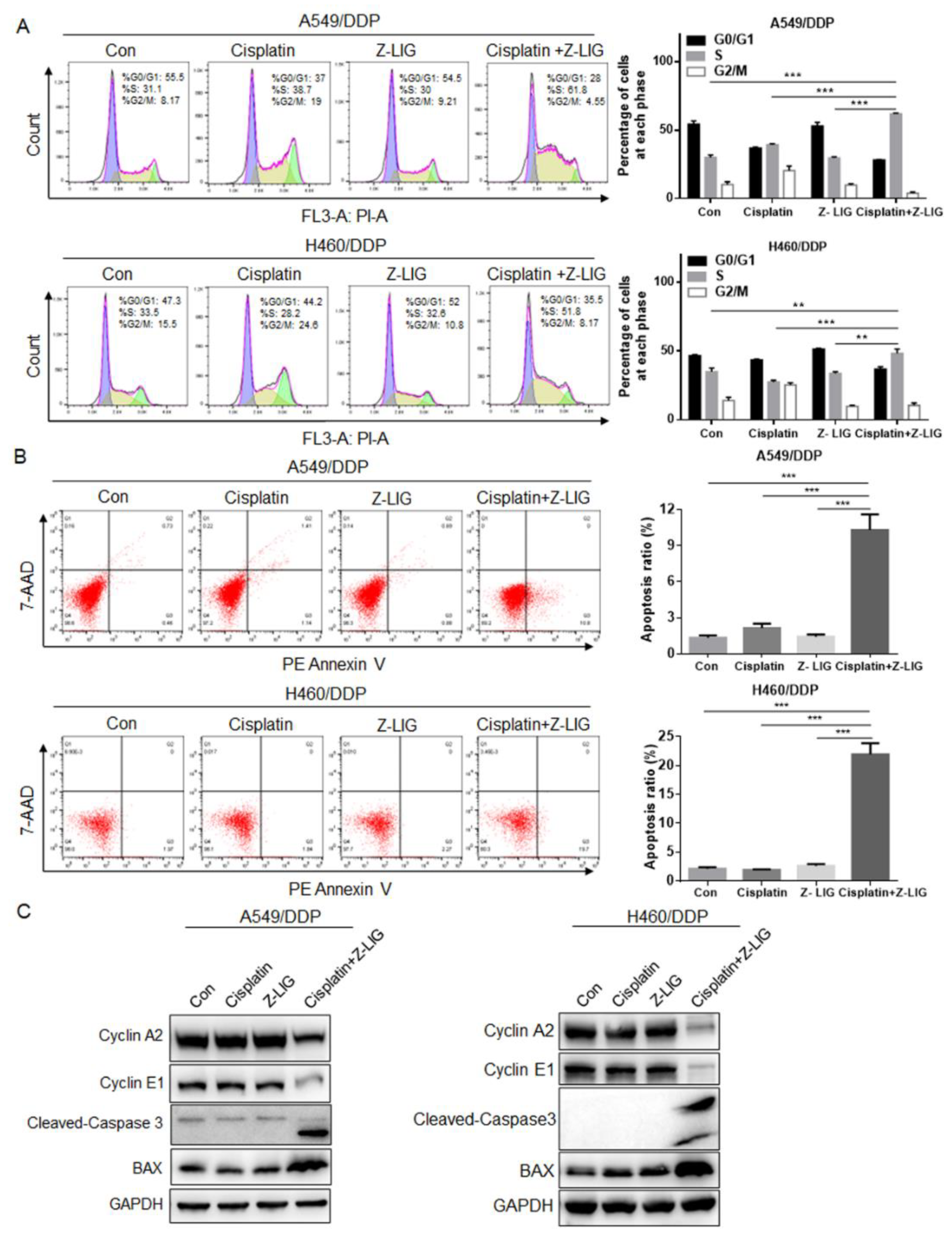
Figure 3.
Z-ligustilide+cisplatin reduced phospholipids contents and upregulated PLPP1 expression. (A) Heat map of the differential lipids in A549, A549/DDP cells and A549/DDP cells Z-ligustilide+cisplatin. (B) Pathway enrichment analysis. (C) Relative enrichment of each lipid class. ***p < 0.001. (D) GSEA analysis of the RNA-seq data. The glycerophospholipid metabolism pathway-targeted genes shows significant overlap. (E) Heat map for RNA-Seq analysis of significantly differentially expressed genes in A549 cells vs A549/DDP cells vs A549/DDP cells treated with Z-ligustilide+cisplatin. (F) Protein level of PLPP1 determined by the western blot.
Figure 3.
Z-ligustilide+cisplatin reduced phospholipids contents and upregulated PLPP1 expression. (A) Heat map of the differential lipids in A549, A549/DDP cells and A549/DDP cells Z-ligustilide+cisplatin. (B) Pathway enrichment analysis. (C) Relative enrichment of each lipid class. ***p < 0.001. (D) GSEA analysis of the RNA-seq data. The glycerophospholipid metabolism pathway-targeted genes shows significant overlap. (E) Heat map for RNA-Seq analysis of significantly differentially expressed genes in A549 cells vs A549/DDP cells vs A549/DDP cells treated with Z-ligustilide+cisplatin. (F) Protein level of PLPP1 determined by the western blot.
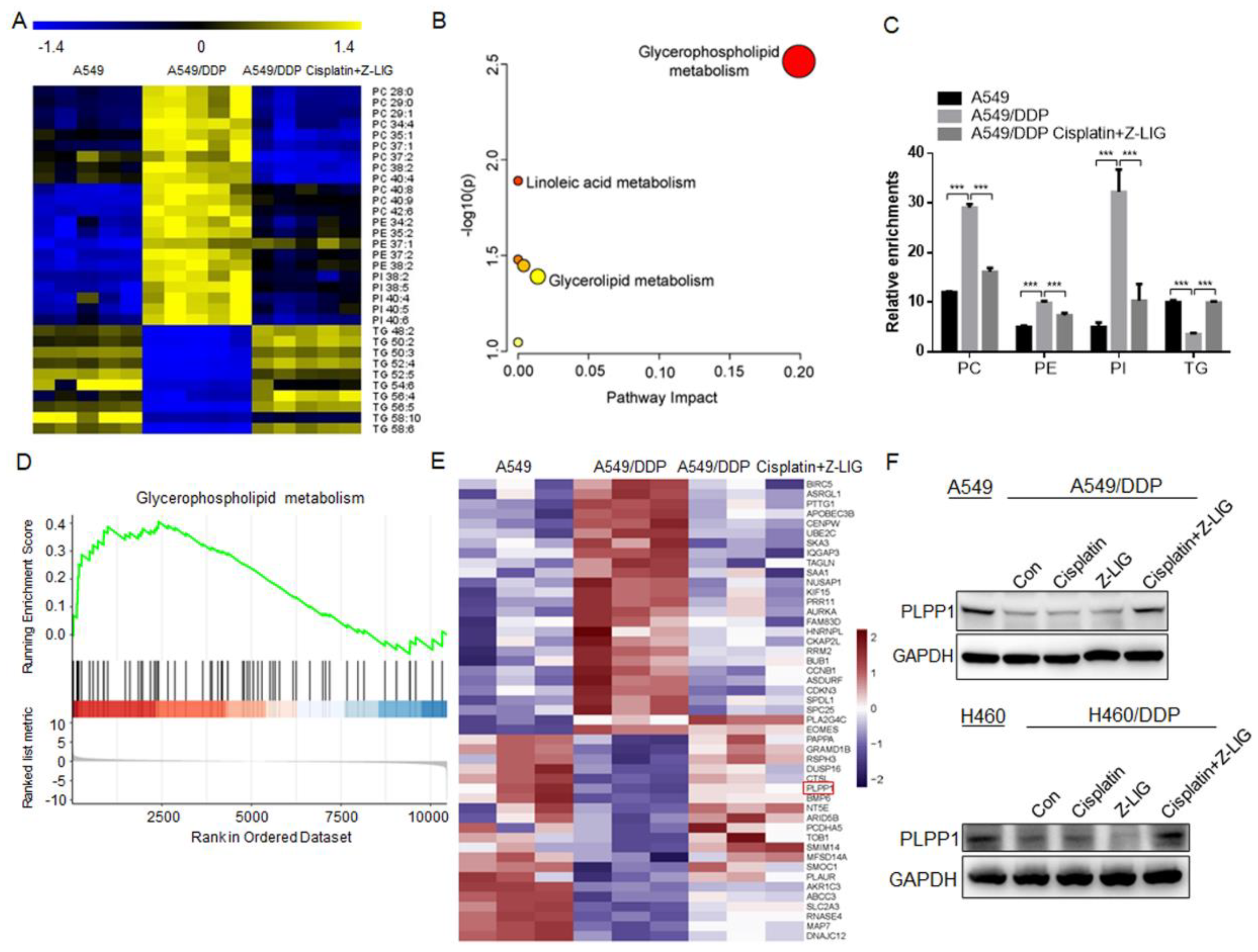
Figure 4.
PLPP1 was low expressed and associated with poor prognosis in lung cancer. (A) PLPP1 expression was detected in LUAD, LUSC, and corresponding normal tissues in the TCGA cohort. LUAD (num (T)=483; num (N)=347). LUSC (num (T)=486; num (N)=338). (B) Representative images of the immunohistochemical staining of PLPP1 in lung cancer samples. Scale bars, 100 μm. (C) The association between the PLPP1 protein expression and overall survival in patients with lung cancer. Low PLPP1 expression (n=41), high PLPP1 expression (n=35). *p < 0.05. (D) The correlation between PLPP1 expression and overall survival in 1451 lung cancer patients. (E) The correlation between PLPP1 expression and overall survival in 116 lung cancer patients administered chemotherapy. (F) The correlation between PLPP1 expression and overall survival in 268 lung cancer patients without chemotherapy.
Figure 4.
PLPP1 was low expressed and associated with poor prognosis in lung cancer. (A) PLPP1 expression was detected in LUAD, LUSC, and corresponding normal tissues in the TCGA cohort. LUAD (num (T)=483; num (N)=347). LUSC (num (T)=486; num (N)=338). (B) Representative images of the immunohistochemical staining of PLPP1 in lung cancer samples. Scale bars, 100 μm. (C) The association between the PLPP1 protein expression and overall survival in patients with lung cancer. Low PLPP1 expression (n=41), high PLPP1 expression (n=35). *p < 0.05. (D) The correlation between PLPP1 expression and overall survival in 1451 lung cancer patients. (E) The correlation between PLPP1 expression and overall survival in 116 lung cancer patients administered chemotherapy. (F) The correlation between PLPP1 expression and overall survival in 268 lung cancer patients without chemotherapy.
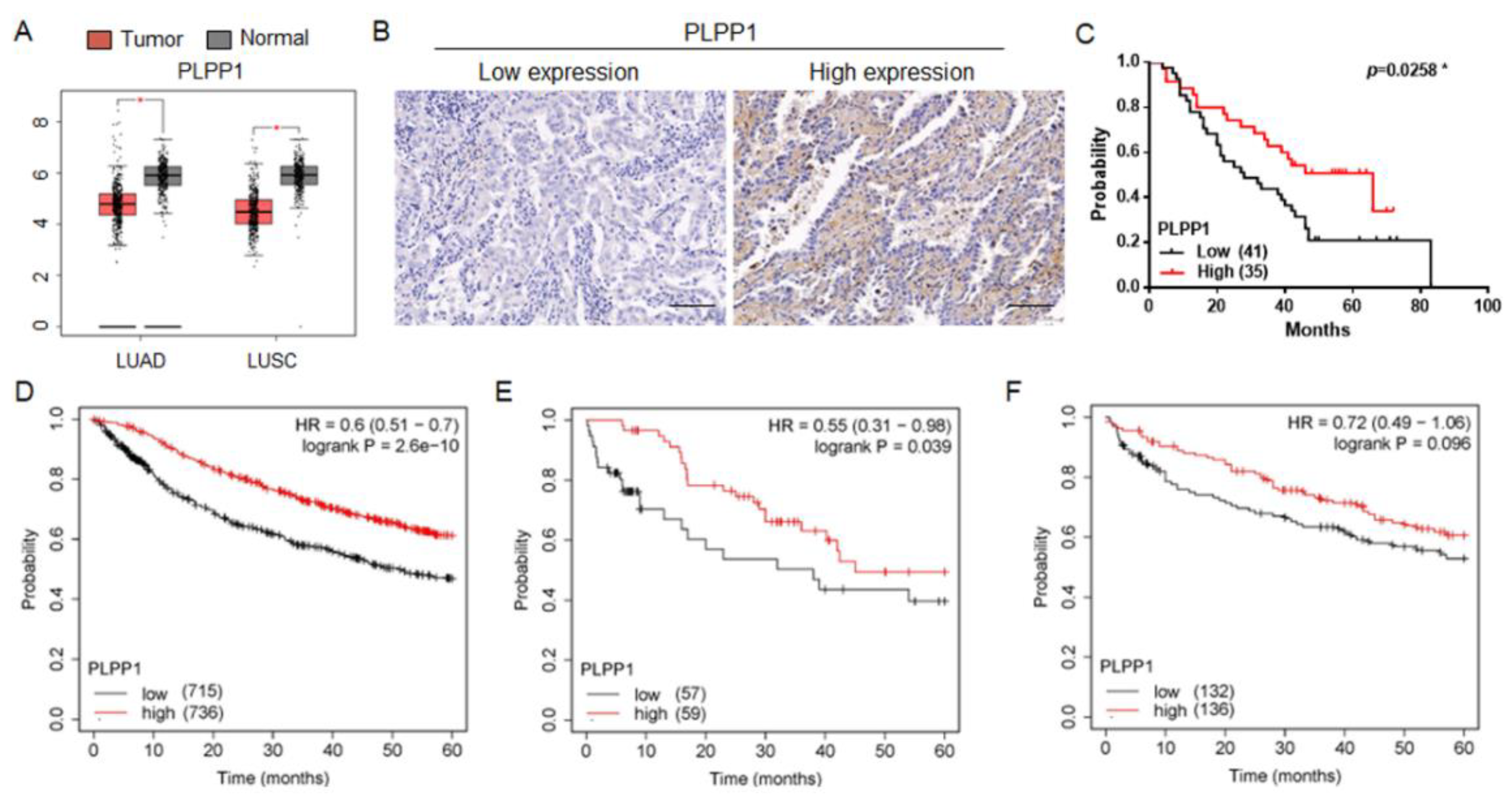
Figure 5.
Z-ligustilide+cisplatin induced the reversal of resistance mediated by PLPP1. (A) Protein expression of PLPP1 in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells, respectively. (B) Cell viability was measured in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment, ***p < 0.001. (C) Relative protein expressions in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment.
Figure 5.
Z-ligustilide+cisplatin induced the reversal of resistance mediated by PLPP1. (A) Protein expression of PLPP1 in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells, respectively. (B) Cell viability was measured in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment, ***p < 0.001. (C) Relative protein expressions in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment.
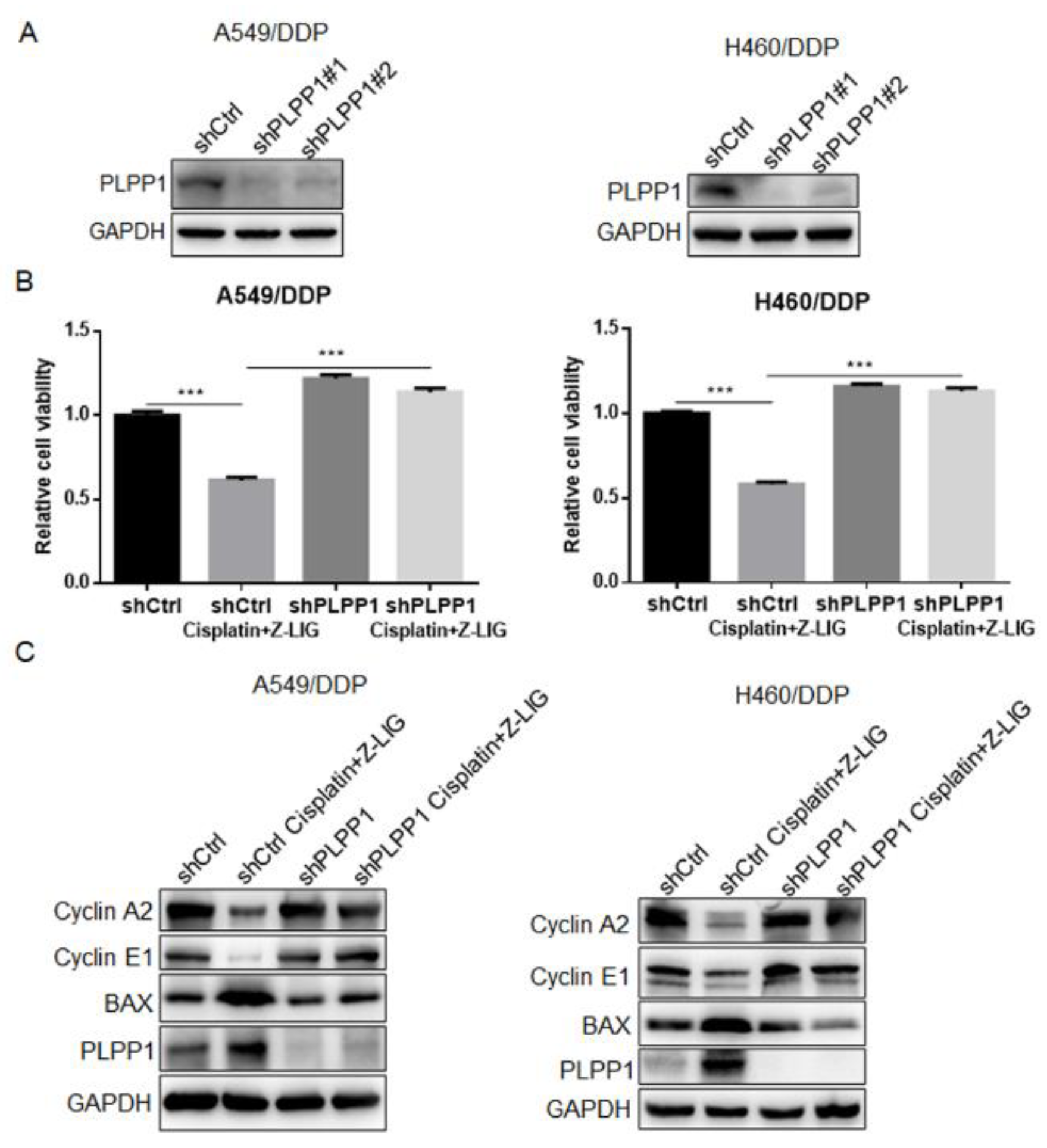
Figure 6.
Z-ligustilide+cisplatin induced the inactivation of AKT by inhibiting PLPP1-mediated phospholipid synthesis. (A) Relative enrichments of each lipid class in shCtrl and PLPP1 knockdown A549/DDP cells with or without Z-ligustilide and cisplatin combination treatment. ns, no significant, ***p < 0.001. (B) Protein expression of p-AKT in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment. (C) and (D) PI3K kinase activity assay and PIP3 levels measurements in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment. ns, no significant, *p < 0.05, **p < 0.01 and ***p < 0.001.
Figure 6.
Z-ligustilide+cisplatin induced the inactivation of AKT by inhibiting PLPP1-mediated phospholipid synthesis. (A) Relative enrichments of each lipid class in shCtrl and PLPP1 knockdown A549/DDP cells with or without Z-ligustilide and cisplatin combination treatment. ns, no significant, ***p < 0.001. (B) Protein expression of p-AKT in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment. (C) and (D) PI3K kinase activity assay and PIP3 levels measurements in shCtrl and PLPP1 knockdown A549/DDP and H460/DDP cells with or without Z-ligustilide and cisplatin combination treatment. ns, no significant, *p < 0.05, **p < 0.01 and ***p < 0.001.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



