
Submitted:
24 July 2023
Posted:
25 July 2023
You are already at the latest version
Abstract
N-Acetyl-L-glutamate kinase (NAGK) catalyzes the rate-limiting step in the ornithine/arginine biosynthesis pathway in eukaryotic and bacterial oxygenic phototrophs. NAGK is the most highly conserved target of the PII signal transduction protein in Cyanobacteria and Archaeplastida (red algae and Chlorophyta). However, yet there is still much to be learnt on how NAGK is regulated in vivo. The use of unicellular green Chlamydomonas reinhardtii as a model system has already been instrumental in identifying several key regulation mechanisms that control nitrogen (N) metabolism. With a combination of molecular-genetic and biochemical approaches, we show the existence of the complex CrNAGK control at transcriptional level, which is dependent on N source and N availability. In growing cells, CrNAGK requires CrPII to properly sense the feedback inhibitor arginine. Moreover, we provided primary evidence that CrPII is only partly responsible for regulating CrNAGK activity to adapt to changing nutritional conditions. Collectively, our results suggest that in vivo CrNAGK is tuned at transcriptional and post-translational levels, and CrPII and additional as yet unknown factor(s) are integral parts of this regulation.
Keywords:
N-acetyl-L-glutamate kinase
; arginine biosynthesis
; PII- signal transduction protein
; green algae
1. Introduction
Arginine and the regulation of its metabolism are of great interest in plant biology because this essential amino acid for protein synthesis is also a precursor in the formation of polyamines and nitric oxide that play critical roles in plant development and adaptation to stresses [1,2,3,4]. The rate-limiting step in the ornithine/arginine (Arg) biosynthesis pathway is catalyzed by N-acetyl-L-glutamate kinase (NAGK), which phosphorylates N-acetyl-L-glutamate to N-acetyl-L-glutamyl-phosphate [1]. The enzyme activity is negatively regulated by Arg feedback inhibition in Cyanobacteria and Archaeplastida (red algae and Chlorophyta) [5,6,7,8,9,10]. Through complex formation with protein PII, NAGK gets relieved from inhibition by Arg [5,9,10,11,12], leading to enhanced activity.
PII proteins are among the most highly conserved and widely distributed signal transduction proteins known in all domains of life [11,12,13,14,15]. A phylogenetic analysis of PII homologues in the eukaryotic domain indicated their inheritance from a cyanobacterial endosymbiont, implying their restriction to Archaeplastida [16]. In contrast to Cyanobacteria and red algae, in green algae and land plants, NAGK activity is controlled by the cellular glutamine (Gln) levels via glutamine-dependent PII-NAGK complex formation, which leads to increased enzyme activity [9,10,17]. Gln sensing as the primary product of nitrogen (N) assimilation indicates the specialization of PII from green algae to respond to the cellular N status.
Among the green algae, Chlamydomonas reinhardtii (Chlamydomonas hereinafter) has become a good model system for revealing important facts about the regulation of N metabolism, and has provided important insights for agricultural plants [18,19,20,21]. Progress has also been made in the identification of the genes and proteins of Arg biosynthesis [22,23,24,25]. According to our data, Chlamydomonas shares with higher plants and other green algae the capability of controlling by PII the activity of NAGK in vitro in Gln-dependent manner [10]. Interestingly, in this alga, PII levels are controlled by the nitrogen source [26].
In Cyanobacteria, PII protein is phosphorylated at a seryl residue (S49) located on the large surface exposed T-loop [27]. The different NAGK activity in cyanobacterial cells, results from the different PII phosphorylation states [12]. In green algae, NAGK research has mainly focused on PII-dependent regulation in vitro [9,10,13]. Despite their role in expanding our knowledge on structure and properties of NAGK, the in vitro protein systems have their limitations, at particular their failure to fully recapitulate the native cellular environment. The apparent gap in the information on NAGK regulation in vivo prompted us to investigate this enzyme activity and expression in Chlamydomonas cells grown in various N sources and under N-deprivation.
The present study is the first to address the multiple control of NAGK in vivo, where the signal protein PII is only an integral part of the regulatory network.
2. Results
2.1. CrNAKG expression is dependent on nitrogen source and growth phase
We chose the wild type 6145c for experiments because it is an arginine prototroph and can utilize ammonium, nitrate or nitrite as nitrogen sources [28]. In the first type of analysis, the cell growth was compared on two nitrogen sources, ammonium and nitrate (Figure 1a). Although Chlamydomonas grew well on both media, ammonium supplementation resulted in slightly slower growth and lower final yields.
The As shown in Figure 1b, the relative expression levels of CrNAGK1 were higher in the nitrate-grown cells compared with ammonium-grown cells. Moreover, nitrate led to an approximately 5-fold increase in CrNAGK1 transcript abundance after 4 h (lag phase) and up to approximately 3.3-2.5-fold after 24 h and 48 h (early- and mid-log phases) of acclimation to nitrate. Interestingly, the accumulation of CrNAGK1 was decreased again to the control level after 72 h (late-log phase). In the presence of ammonium, the expression levels of this gene in the lag, early- and mid-log phases were higher in about 2-1.7 times than in the late-log and stationary phases of growth (Figure 1b). Thus, The CrNAGK1 transcription was influenced by the growth phases and N source.
2.2. CrNAKG activity is dependent on nitrogen source
There is evidence that the activity of cyanobacterial NAGK is dependent on nitrogen source [29]. We wanted to find out whether the CrNAGK activity also depends on which external nitrogen source was provided.
In the presence of ammonium, the levels of CrNAGK activity increased at lag phase (4 h) to a peak level of 1.32 U mg-1 protein and then declined slightly as the culture entered the early logarithmic (log) phase of growth, reaching its lowest level during late log phase. (Figure 2a). Interestingly, a significant level of enzyme activity (1.1 U mg-1 protein) was detectable in the stationary phase.
When cells were transferred to a nitrate-containing medium, the CrNAG kinase activity was also increased after 4h but reached a maximum within early and mid-log phase of growth (24h and 48h of incubation) (Figure 2a). During further growth in nitrate-containing medium, the enzyme activity decreased again approximately 3.6 times higher than the control level. It appears likely that the regulation of CrNAGK expression in cells supplemented with ammonium or nitrate is not the only level of enzyme control, especially in the stationary phase of growth.
Chlamydomonas NAGK has been shown to be an arginine-sensitive enzyme [10]. In vitro the activity of arginine-sensitive NAGKs is negatively regulated by arginine feedback inhibition [11,30,31,32]. To assess the overall impact of arginine on the CrNAGK activity in vivo, we quantified the intracellular free arginine content in cells grown on ammonium and nitrate as nitrogen sources. This amino acid content increased after 4h and reached a maximum in the early log-phase of cell growth in both media (Figure 2b). During further growth, the intracellular arginine concentrations decreased. The observed enhanced level of internal arginine in lag, early and mid-lag phases correlated with a high level of CrNAGK activity in these phases (Figure 2a), indicating a relief of this enzyme from inhibition by arginine. In addition, the levels of Gln as an amino acid that controls CrNAGK activity [10] showed the same trend as Arg (Figure 2c). This suggests that PII might be involved in the N-dependent control of the enzyme activiy.
2.3. Nitrite promotes CrNAGK activity
To further explore the relationship between NAGK regulation and N sources, the alga was incubated in 10mM nitrite. The strain 6145c growing in nitrite-supplemented medium did not show significant differences in their growth characteristics compared with cells grown on ammonium (Figure 3a,1a).
In nitrite, cells exhibited the same patterns of CrNAGK1 gene expression and enzyme activity as in cells grown in nitrate (Figure 3b,c). The highest levels of cellular Arg and Gln were obtained in cells grown in the early and mid-log phases (Figure 3d). Therefore, PII might dampen arginine feedback inhibition under these conditions.
2.4. Underexpression of CrPII decreases the activity of CrNAGK but not the CrNAGK1 transcript level
To clarify the role of the CrPII protein in CrNAGK regulation in vivo, we generated CrGLB1-underexpressing transformants as described previously [33]. Two amiRNAGLB1 strains were selected for further research. Western blotting revealed that the CrPII protein levels in the amiRNAGLB1-65 and amiRNAGLB1-88 strains were no higher 5% of those in the parental cells (Figure 4a). To test, if the phenotype was a result of decreased transcript accumulation, RNA was extracted from CC3491, amiRNAGLB1-65 and amiRNAGLB1-88 cells, and analyzed by quantitative real time PCR. The results confirmed a significant knock-down of CrGLB1 mRNA in both transformants (Figure 4b).
The strain CC3491 was unable to grow on nitrate. However, this strain exhibits the growth on ammonium and nitrite as nitrogen sources. Notable, the down-regulation of CrPII did not affect the growth of amiRNAGLB1 cells (Figure S1).
As shown in Figure 5a, downregulation of CrGLB1 led to a decrease of CrNAGK activity both in ammonium-grown and nitrite-grown cells in the lag and log phases. Unexpectedly, there was no obvious difference in the enzyme activity between parent strain and CrGLB1-underexpressing transformants during stationary phase of growth (Figure 5a,b). However, the expression levels of CrNAGK1 gene were similarly to parental strain (Fig. 5b).
At the same time, lower enzyme activity correlated with a lower level of intracellular arginine in GLB1-knockdown cells compared with parental strain in late-log and stationary phases, and log phase and in ammonium- and nitrite-containing medium, respectively (Figure 5c).
2.5. CrNAGK activity is changed in N-starved cells
It was previously shown that both CrGLB1 gene and CrNAGK1 gene are inducible under N deprivation conditions [22,25]. Since nothing is known about the CrNAGK activity in N-deplete cells, we measured it in the strain 6145c. The transfer of the cells from growth medium into N-free medium led to an increase in NAGK activity approximately 1.4-fold after 4 h and up to approximately 2.4-fold after 24 h of acclimation to N-limitation (Figure 6a). However, after 48h, the activity of CrNAGK decreased sharply to the level of 0.3 U mg−1 protein. The observed Arg level in cells after 48 h of N-deprivation was also the lowest (Figure 6a).
To test whether CrPII controls CrNAGK activity in N-starved cells, we compared the enzyme activity in parental strain and CrGLB1-knockdown transformants. When CC3491 cells are submitted to N deprivation for 4h or 24h, this is accompanied by an increase of CrNAGK activity (Figure 6b). The observed enzyme activity of the transformants following elimination of N from the medium for 4h was approximately 1.8 times lower than that of WT. It is interesting that after 24h or 48h of incubation in N-free medium, CrNAGK activity of amiRNAGLB1-65 and amiRNAGLB1-88 cells was very similar to that of parental cells (Figure 6b), suggesting the independence of NAGK regulation from CrPII under these conditions. After long-time N-starvation (24h and 48 h), the observed Arg levels in all strains was about 50-40% of that of N-replete cells (Figure 6c).
Next, we asked the question whether the changes in CrNAGK activity in N-starved cells are accompanied by changes in the transcript levels. The expression levels of the gene of interest were upregulated by N-depletion in both CC3491 and in amiRNA-GLB1 strains (Figure 6d). Importantly, the induction of gene CrNAGK1 was not reduced in CrGLB1-knockdown transformants. These results indicate a role of transcriptional regulation of CrNAGK1 gene in increasing of enzyme activity after 4h and 24 h of N-starvation. In contrast, despite the increase in the amounts of CrNAGK1 transcripts during further incubation in a N-free medium (Figure 6d), the enzyme activity significantly decreased (Figure 6b), hinting at potential additional regulator(s) of NAGK during Chlamydomonas acclimation to N-starvation
3. Discussion
NAGK is the most highly conserved target of PII in photosynthetic organisms [11]. In vitro experiments suggest that a high nitrogen status is sensed by the CrPII protein, and CrPII-CrNAGK complex formation is favored leading to arginine synthesis [10]. However, it remains elusive how these in vitro model accurately mimic cells in vivo. In this work, we report original insights into the NAGK regulation of Chlamydomonas under conditions of N-sufficiency or N-limitation.
Chlamydomonas efficiently uses ammonium, nitrate and nitrite as N sources [18,21]. Wild type cells grown in the media supplemented with ammonium, nitrate or nitrite with acetate (as a carbon source) showed very similar growth with slightly lower final yields in ammonium-containing medium (Figure 1a,3a). The highest levels of CrNAGK activity were seen in cells grown in nitrate and nitrite from early- and mid-log phase. The regulation at transcriptional level may play a role in higher levels of CrNAGK in cells incubated in nitrate or nitrite compared to that in ammonium (Figure 1b,3b). Notably, CrPII is also induced by nitrate and nitrite [26], ensuring the possible coordinating of two interacted proteins. This result is also consistent with the idea that PII-dependent and nitrate/nitrite assimilation pathways are interconnecting [20].
We also demonstrated that a certain increase in total Arg accumulation in all N sources was found during the early-log phase of cell growth, especially in nitrite-containing medium (Figure 2b,3d). In general, an increase in Arg concentrations is expected to limit the accumulation of this amino acid by inhibiting of arginine-sensitive NAGKs [11]. However, there is an additional PII-mediated regulatory mechanism, by which high nitrogen availability activates NAGK and thus promotes Arg synthesis [10]. The most CrNAGK activity in early-log phase supports the theory that the feedback inhibitory effect of Arg on CrNAGK is alleviated by CrPII in growing cells [11]. As mentioned above, PII of Chloroplastida has acquired an additional C-terminal extension that acts as a Gln-binding site and makes NAGK activation by PII Gln-dependent [9,10,17]. At elevated Gln levels, corresponding to N-rich conditions, CrPII appears to activate CrNAGK (Figure 2c, 3d).
To further study the role of PII-dependent control in CrNAGK activity of growing cells, we took advantage of the amiRNA approach [33]. The two CrGLB1-underexpressing strains, that were generated and characterized in this work, exhibited a significantly reduced level of PII protein: ∼95% less than that in parental strains (Figure 4a), which is consistent with the low levels of CrGLB1 transcripts in both transformants (Figure 4b). The amiRNAGLB1-strains had growth curves that were indistinguishable from the representative growth curve of WT (Figure S1).
Compared to WT, the CrNAGK activity was significantly decreased in the amiRNAGLB1 cells from lag and log phases both in ammonium- and in nitrite-containing media (Figure 5a,b). Thus, our results provide primary evidence that the CrPII is a component of CrNAGK regulation in growing cells (Figure 7). Moreover, the CrNAGK1 gene was not impaired in the CrGLB1-underexpressing strains (Figure 5c,d), indicating the role of transcription in the regulation of CrNAGK in addition to PII.
Surprisingly, no detectable difference in CrNAGK activity occurred between parental strains and CrPII-transformants in stationary phase (Figure 5a,b). The possible scenario is N is depleted in stationary phase and increased levels of 2-oxoglutarate [13] may interfere with CrPII in the control of CrNAGK [10,11]. However, we cannot rule out the role of the other signals that might arise in this growth phase.
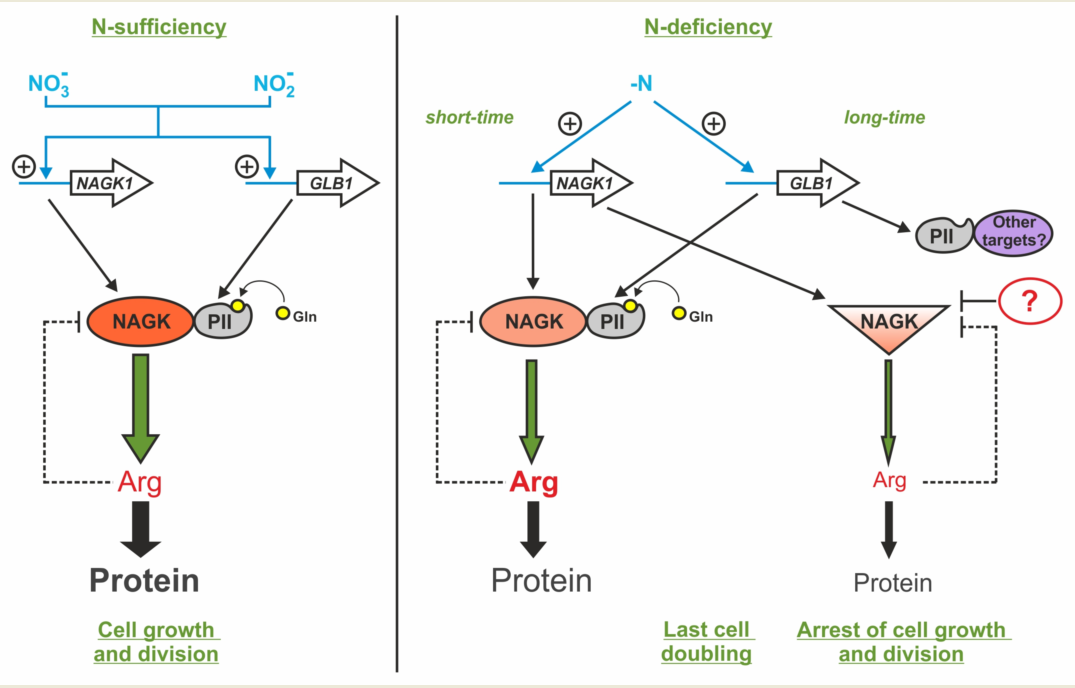
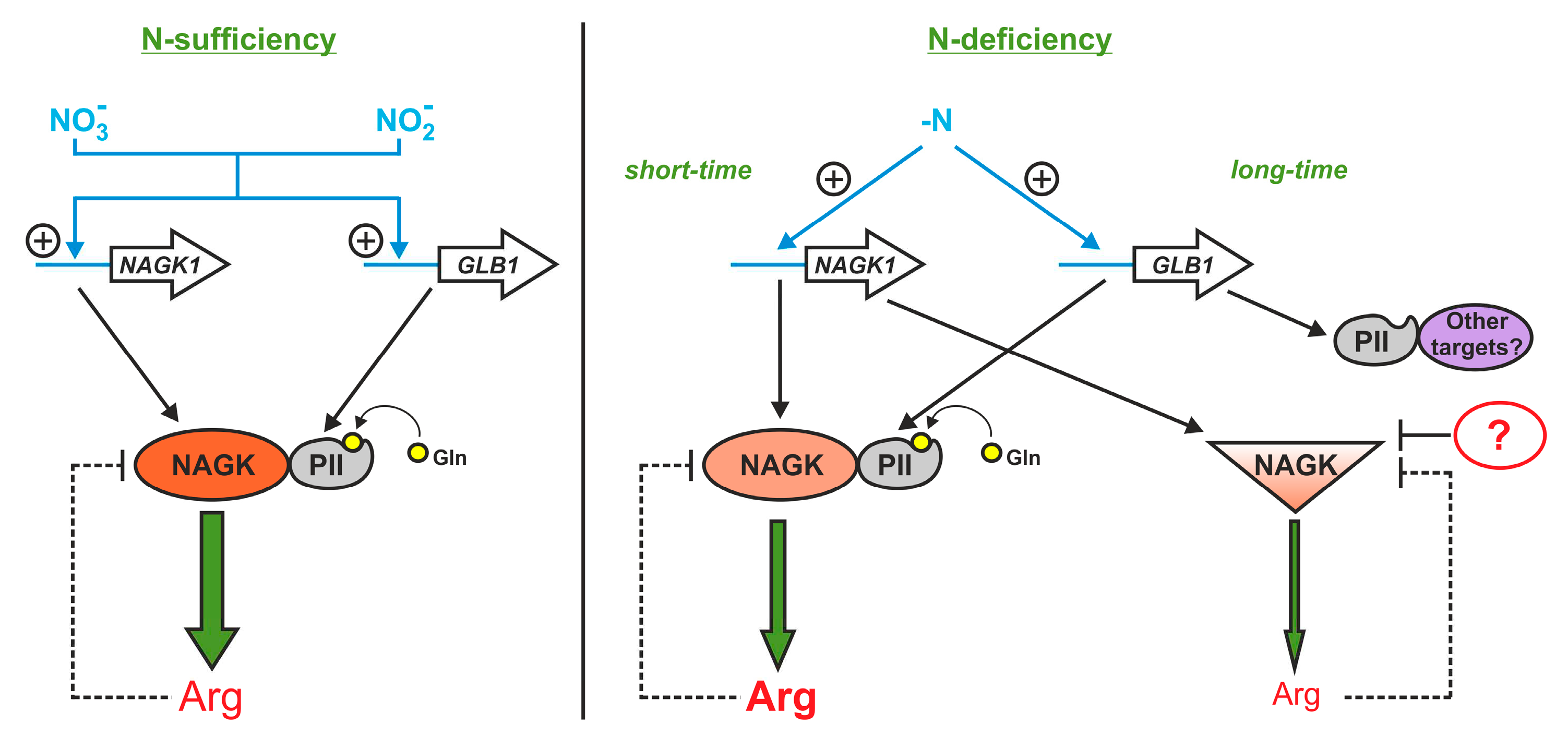
In Chlamydomonas, N supply is critical to the maintenance the growth and division [34]. Under N deprivation, CrNAGK1 and CrGLB1 genes are induced [22,25]. Moreover, in response to -N-shift conditions, CrNAGK activity temporary increased (Figure 6 a,b). A similar transient increase in this enzyme activity following N- deprivation has also been shown in another green alga, Myrmecia incisa [17]. Notably, in the case of amiGLB1 strains, we observed a significant difference in enzyme activity only at 4h of N-deficiency (Figure 6b) Thus, while PII reduction influenced the regulation of CrNAGK in short-time N-starved cells, it appeared not to have a significant effect on the enzyme activity in long-time N-starved cells (Figure 7). Since in Chlamydomonas the cell density approximately doubles within the first 24 h of N starvation [34], the observed PII-dependent CrNAGK regulation can be used to maintain the finite reservoir of intracellular arginine (Figure 6c). This led to an assumption that another mechanism may be responsible for controlling CrNAGK to adapt to long-time N limitation.
Although further research into the molecular mechanisms underlying CrNAGK control is required, this study shows that CrPII is only partly responsible for enzyme activity levels in Chlamydomonas cells.
4. Materials and Methods
4.1. Strains and Growth Conditions
The Chlamydomonas WT strain 6145c was kindly provided by Prof. Emilio Fernández (University of Córdoba, Spain). The strains CC3491 and CC4533 were obtained from Chlamydomonas Resource Center (University of Minnesota, St. Paul, MN, USA).
Cells were grown in tris-acetate-phosphate (TAP) medium [35] with modified trace elements [36] in a chamber (KBWF 240, Binder GmbH, Tuttlinger, Germany) at 22 °C under continuous illumination by white light (fluence rate of 45 μmol/m2 s) with continuous agitation (100 rpm). Depending on the nitrogen source, three variants of the TAP medium were used: with 7.5 mM NH4Cl, 4 mM KNO3 or 10 mM KNO2.
At each harvesting time, the number of cells was recorded by employing a counting chamber. Four hundred cells from each sample were scored for three biological replicates. The number of viable cells was counted microscopically with the use of 0.05% (v/v) Evans blue (Dia-M, Moscow, Russia) as previously described [37]. The number of non-viable (stained) and viable (unstained) cells were determined.
4.2. Generation of GLB1-underexpressing transformants
Screening for GLB1-underexpressing transformants was carried out with the plasmid generated previously [33]. The amiRNA construct (pChlamiRNA3GLB1) or the empty vector [38] was transformed into the cell-wall-deficient CC3491 strain by vortexing with glass beads [39]. The strains were selected on TAP agar containing 10 µg/mL paromomycin (Sigma-Aldrich, Steinheim, Germany) and then screened by Western blotting and RT-qPCR for transformants with reduced abundance of CrPII.
4.3. Quantitative real-time PCR
The total RNA was isolated with Trizol according to the manufacturer’s instructions (Invitrogen, Waltham, MA, USA). DNA contamination was avoided by treatment of the RNA samples with RNase-Free DNase I (Fermentas, Vilnius, Lithuania). Subsequently, concentration and purity of total RNA (260/280 nm ratio) were determined on spectrophotometer (SmartSpec Plus, Bio-Rad, USA). Agarose gel electrophoresis (1.2% agarose, w/v) was performed to visualize the integrity of RNA. Reverse transcription was performed with Revert Aid H Minus First Strand cDNA Synthesis Kit according to the manufacturer’s instructions (Thermo Fisher Scientific, No. K-1631, Rockford, IL, USA). Gene expression analysis was carried out by real-time quantitative RT-PCR (RT-qPCR) on the Light Cycler Instrument (CFX96 Real-Time PCR Detection System, Bio-Rad Laboratories, Singapore) using SYBR Green I following a previously reported protocol [40]. The primer pairs used for RT-qPCR were as follows: 5’- GCAGGCGCTCAACATCAACG-3’ and 5’-CATGCCACCAGCAATGACGC-3’ for CrNAGK1 (Cre01.g015000_4532), 5’-GGCGTCAAGTTCTTCCGCAT-3’ and 5’-GGTTGGAGGGACCGAACTCA-3’ for CrGLB1 (Cre07.g357350_4532) and 5’- CTTCTCGCCCATGACCAC-3’ and 5’-CCCACCAGGTTGTTCTTCAG-3’ for RACK1 (receptor of activated protein kinase C; Cre06.g278222_4532, formerly termed CBLP).
The relative gene expression ratios were normalized with RACK1 (using the ΔCT and ΔΔCT methods [41]. ΔΔCT was used to directly demonstrate the levels of induction and ΔCT to show relative transcript abundances in selected conditions. Controls without template or reverse transcriptase were always included. The accuracy and reproducibility of the real-time assay was determined from low variation in CT values across replicates. Values were obtained from at least three biological replicates; each replicate was analyzed three times.
4.4. Protein isolation, SDS-PAGE, and Western blotting
Chlamydomonas cells (4 106 cells/ mL in 10 ml) were collected by centrifugation (3,000 g, 5 min) and resuspended in 0.1 M DTT, 0.1 M Na2CO3. Then, 0.66 vol of 5% SDS, 30% sucrose were added. Homogenization of the suspensions was achieved by rapid shaking at room temperature for 20 min. The protein concentration was determined by staining with amido black, using BSA as a standard [42]. After separation of the proteins by SDS-PAGE on a 12% polyacrylamide gel [43], they were transferred to nitrocellulose membranes (Carl Roth, Karlsruhe, Germany) by a semidry blotting (Trans-blot SD, Bio-Rad, Bio-Rad Laboratories, Geylang, Singapore). Blots were blocked in 5% non-fat dry milk in Tris-buffered saline solution with 0.1% Tween 20 prior to an incubation in the presence of primary antibodies. The dilution of the primary antibody was 1:5,000 anti-PII [22] and 1:2,000 anti-HSP70B [44]. As a secondary antibody, the horseradish peroxidase-conjugated anti-rabbit serum (Sigma-Aldrich, No. A054, Steinheim, Germany) was used at a dilution of 1:10,000. The membranes were scanned using Bio-Rad ChemiDocTMMP Imaging System (Bio-Rad, Bio-Rad Laboratories, Singapore).
4.5. NAGK activity assays
To determine NAGK activity, Chlamydmonas strains (2 106 cells/ mL) grown on ammonium, nitrate or nitrite were used. Cells were harvested from lag, log or stationary phase of growth by centrifuging them at 3000 × g for 10 min, and after resuspending in 200 µL of buffer, pH 7.4 (50 mM Tris-Cl, 0.5 mM EDTA, 1 mM DTT and 0.5 mM benzamidine), they were disrupted by glass beads (0.45 mm diameter) using disintegrator (Minilys, Bertin technologies, Montigny-le-Bretonneux, France) The suspension was centrifuged at 20000 × g for 15 min, and the resulting supernatant was used as source of enzyme. All the operations were performed at 0 °C. Protein concentration was assessed by the Pierce™ BCA protein assay kit (No 23227, Thermo Fisher Scientific, Rockford, USA).
The NAGK activity was measured as described [12]. Briefly, freshly prepared protein extracts (0.4 mg) were added to a reaction mixture (400 µL) containing 400 mM NH2OH·HCl, 400 mM Tris (base), 20 mM MgCl2 and 10 mM ATP. The reaction was started by adding 40 mM N-acetyl-L-glutamate. After incubation at 37 °C for 1 h the reaction was stopped by the addition of 400 μL of a solution containing 5% (w/v) FeCl3·6 H2O, 8% (w/v) trichloroacetic acid and 0.3 M HCl. Blank reactions were performed by omitting N-acetyl-glutamate from the assay. The activity was determined spectrophotometrically following the hydroxamate Fe3+ complex formation using a molar extinction coefficient 456/M cm at 450 nm. One unit of NAGK refers to the amount of enzyme required to catalyze the conversion of 1 mmol of N-acetyl-glutamate/min. The specific activity of CrNAGK was expressed as units per mg of protein. Measurements were performed on at least three separate cultures (biological repeats).
4.6. Determination of glutamine content
To determine the intracellular Gln levels, the cells (2 108 cells/mL) were pelleted (4000 g, 5 min) and resuspended in 200 µL of distilled H2O for 20 min at 95 °C. Gln content was measured with Glutamine/Glutamate Determination Kit (No. GLN1, Sigma-Aldrich, Steinheim, Germany) according to the manufacturer’s instructions. Briefly, Gln content was determined spectrophotometrically at 340 nm following enzymatic deamination of L-glutamine and dehydrogenation of L-glutamate with conversion of NAD+ to NADH [45]. The conversion of NAD+ to NADH is proportional to the amount of Glu that is oxidized and, consequently, the amount of Gln converted to Glu in the samples. In addition, endogenous Glu was determined and subtracted from Glu concentration derived by deamination of Gln. The absorbance at 340 nm was measured using a microplate reader CLARIOstar (BMG, Ortenberg, Germany).
4.7. Determination of arginine content
To determine the intracellular Arg levels, the cells (2 106 cells/mL) were pelleted (4000 g, 5 min) and resuspended in 200 µL of distilled H2O for 20 min at 95 °C. Total free Arg was measured as described [46] Briefly, 0.1 mL of 0.2% 8-hydroxyquinoline and 0.1 mL of 2 M NaOH were added to supernatant and reaction mixture was incubated for 10 min on ice. After addition of 0.1 mL of 19% sodium hypochlorite and vortexing for 30 sec the reaction was stopped by 0.1 mL of 40% urea. The absorbance was estimated at 500 nm. Measurements were performed at least in three biological triplicates).
4.8. Statistical analysis
The values for the quantitative experiments described above were obtained from at least three independent experiments with no fewer than three technical replicates. Data represent the mean ± SE. When necessary, statistical analyses were followed by a Student’s t test (p value < 0.01 or < 0.05).
Supplementary Materials
The following supporting information can be downloaded at: the website of this paper posted on Preprints.org., Figure S1: Effects of ammonium and nitrite on cell growth of CC3491 and GLB1-knockdown strains. The growth curves were analyzed in the presence of 7.5 mM NH4Cl or 10 mM KNO2. Values are means ± SE of three independent experiments.
Author Contributions
Data analysis and collection, validation, carrying out experiments, writing—original draft preparation, V.V.; methodology, formal analysis, investigation, T.L.; carrying out experiments, V.S., Conceptualization; writing—original draft preparation, review and editing.; supervision; funding acquisition, E.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by RUSSIAN SCIENCE FOUNDATION, grant number 21-14-00017.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Emilio Fernández (University of Cόrdoba, Spain) for kind providing of the strain 6145c.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study, in the writing of the manuscript and in the decision to publish the results.
References
- Llácer, J.L.; Fita, I.; Rubio, V. Arginine and nitrogen storage. Curr. Opin. Struct. Biol. 2008, 18, 673–681. [Google Scholar] [CrossRef]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Morency, F.; Jones-Overton, C.; Cooke, J. Accumulation pattern and identification of seed storage proteins in zygotic embryos of Pinus strobus and in somatic embryos from different maturation treatments. Physiol. Plant 2004, 121, 682–690. [Google Scholar] [CrossRef]
- Corpas, F.J.; Palma, J.M.; del Río, L.A.; Barroso, J.B. Evidence supporting the existence of L-arginine-dependent nitric oxide synthase activity in plants. New Phytol. 2009, 184, 9–14. [Google Scholar] [CrossRef]
- Beez, S.; Fokina, O.; Herrmann, C.; Forchhammer, K. N-acetyl-L-glutamate kinase (NAGK) from oxygenic phototrophs: P(II) signal transduction across domains of life reveals novel insights in NAGK control. J. Mol. Biol. 2009, 389, 748–758. [Google Scholar] [CrossRef]
- Lapina, T.; Selim, K.A.; Forchhammer, K.; Ermilova, E. The PII signaling protein from red algae represents an evolutionary link between cyanobacterial and Chloroplastida PII proteins. Sci. Rep. 2018, 8, 790. [Google Scholar] [CrossRef]
- Sugiyama, K.; Hayakawa, T.; Kudo, T.; Ito, T.; Yamaya, T. Interaction of N-acetylglutamate kinase with a PII-like protein in rice. Plant Cell Physiol. 2004, 45, 1768–1778. [Google Scholar] [CrossRef]
- Chen, Y.M.; Ferrar, T.S.; Lohmeier-Vogel, E.M.; Morrice, N.; Mizuno, Y.; Berenger, B.; Ng, K.K.; Muench, D.G.; Moorhead, G.B. The PII signal transduction protein of Arabidopsis thaliana forms an arginine-regulated complex with plastid N-acetyl glutamate kinase. J. Biol. Chem. 2006, 281, 5726–5733. [Google Scholar] [CrossRef]
- Minaeva, E.; Forchhammer, K.; Ermilova, E. Glutamine assimilation and feedback regulation of L-acetyl-N-glutamate kinase activity in Chlorella variabilis NC64A results in changes in arginine pools. Protist 2015, 166, 493–505. [Google Scholar] [CrossRef]
- Chellamuthu, V.R.; Ermilova, E.; Lapina, T.; Lüddecke, J.; Minaeva, E.; Herrmann, C.; Hartmann, M.D.; Forchhammer, K. A widespread glutamine-sensing mechanism in the plant kingdom. Cell 2014, 159, 1188–1199. [Google Scholar] [CrossRef]
- Selim, K.; Ermilova, E.; Forchhammer, K. From cyanobacteria to Archaeplastida: new evolutionary insights into PII signalling in the plant kingdom. New Phytol. 2020, 227, 722–731. [Google Scholar] [CrossRef]
- Heinrich, A.; Maheswaran, M.; Ruppert, U.; Forchhammer, K. The Synechococcus elongatus PII signal transduction protein controls arginine synthesis by complex formation with N-acetyl-L-glutamate kinase. Mol. Microbiol. 2004, 52, 1303–1314. [Google Scholar] [CrossRef] [PubMed]
- Selim, K.A.; Lapina, T.; Forchhammer, K.; Ermilova, E. Interaction of N-acetyl-L-glutamate kinase with the PII signal transducer in the non-photosynthetic alga Polytomella parva: co-evolution towards a hetero-oligomeric enzyme. FEBS J. 2020, 287, 465–482. [Google Scholar] [CrossRef]
- Leigh, J.A.; Dodsworth, J.A. Nitrogen regulation in bacteria and archaea. Annu. Rev. Microbiol. 2007, 61, 349–377. [Google Scholar] [CrossRef]
- Forchhammer, K.; Selim, K.; Huergo, L. New views on PII signaling: from nitrogen sensing to global metabolic control. Trends Microbiol. 2022, 30, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Chellamuthu, V.R.; Alva, V.; Forchhammer, K. From cyanobacteria to plants: conservation of PII functions during plastid evolution. Planta 2013, 237, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, W.; Sun, L.P.; Zhou, Z.G. Evidence for PII with NAGK interaction that regulates Arg synthesis in the microalga Myrmecia incisa in response to nitrogen starvation. Sci. Rep. 2017, 7, 16291. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Galván, A. Inorganic nitrogen assimilation in Chlamydomonas. J. Exp. Bot. 2007, 58, 2279–2287. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Chamizo-Ampudia, A.; Llamas, A.; Galván, A.; Fernandez, E. Understanding nitrate assimilation and its regulation in microalgae. Front. Plant Sci. 2015, 6, 899. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Llamas, A.; Galván, A.; Fernández, E. Role of nitrate reductase in NO production in photosynthetic eukaryotes. Plants 2019, 8, 56. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Sanz-Luque, E.; Tejada-Jiménez, M.; Llamas, Á; Plouviez, M. ; Guieysse, B.; Fernández, E.; Galván, A. Chlamydomonas reinhardtii, an algal model in the nitrogen cycle. Plants 2020, 9, 903. [Google Scholar] [PubMed]
- Ermilova, E.; Lapina, T.; Zalutskaya, Z.; Minaeva, E.; Fokina, O.; Forchhammer, K. PII signal transduction protein in Chlamydomonas reinhardtii: localization and expression pattern. Protist 2013, 164, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-J.; Wang, H.; Gargouri, M.; Deshpande, R.R.; Skepper, J.N.; Holguin, F.O.; Juergens, M.T.; Shachar-Hill, Y.; Hicks, L.M.; Gang, D.R. The response of Chlamydomonas reinhardtii to nitrogen deprivation: A systems biology analysis. Plant J. 2015, 81, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Wase, N.; Black, P.N.; Stanley, B.A.; DiRusso, C.C. Integrated quantitative analysis of nitrogen stress response in Chlamydomonas reinhardtii using metabolite and protein profiling. J. Proteome Res. 2014, 13, 1373–1396. [Google Scholar] [CrossRef]
- Monteiro, L.d.F.R.; Giraldi, L.A.; Winck, F.V. From Feasting to Fasting: The Arginine Pathway as a Metabolic Switch in Nitrogen-Deprived Chlamydomonas reinhardtii. Cells 2023, 12, 1379. [Google Scholar] [CrossRef]
- Zalutskaya, Z.; Kochemasova, L.; Ermilova, E. Dual positive and negative control of Chlamydomonas PII signal transduction protein expression by nitrate/nitrite and NO via the components of nitric oxide cycle. BMC Plant Biol. 2018, 18, 305. [Google Scholar] [CrossRef]
- Forchhammer, K.; Tandeau de Marsac, N. The PII protein in the cyanobacterium Synechococcus sp. strain PCC 7942 is modified by serine phosphorylation and signals the cellular N-status. J. Bacteriol. 1994, 176, 84–91. [Google Scholar] [CrossRef]
- Bellido-Pedraza, C.M.; Calatrava, V.; Llamas, A.; Fernandez, E.; Sanz-Luque, E.; Galván, A. Nitrous oxide emissions from nitrite are highly dependent on nitrate reductase in the microalga Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2022, 23, 9412. [Google Scholar] [CrossRef]
- Forchhammer, K.; Tandeau de Marsac, N. Functional analysis of the phosphoprotein PII (glnB gene product) in the cyanobacterium Synechococcus sp. strain PCC 7942. J. Bacteriol. 1995, 177, 2033–40. [Google Scholar] [CrossRef]
- Lácer, J.L.; Contreras, A.; Forchhammer, K.; Marco-Marín, C.; Gil-Ortiz, F.; Maldonado, R.; Fita, I.; Rubio, V. The crystal structure of the complex of PII and acetylglutamate kinase reveals how PII controls the storage of nitrogen as arginine. Proc. Natl. Acad. Sci. U.S.A. 2007, 104, 17644–17649. [Google Scholar] [CrossRef]
- Forcada-Nadal, A.; Llácer, J.L.; Contreras, A.; Marco-Marín, C.; Rubio, V. The PII-NAGK-PipX-NtcA regulatory axis of cyanobacteria: a tale of changing partners, allosteric effectors and non-covalent interactions. Front. Mol. Biosci. 2018, 5, 91. [Google Scholar] [CrossRef]
- Forchhammer, K.; Selim, K.A. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 2019, 44, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Zalutskaya, Z.; Kharatyan, N.; Forchhammer, K.; Ermilova, E. Reduction of PII signaling protein enhances lipid body production in Chlamydomonas reinhardtii. Plant Sci. 2015, 240, 1–9. [Google Scholar] [CrossRef]
- Takeuchi, T.; Benning, C. Nitrogen-dependent coordination of cell cycle, quiescence and TAG accumulation in Chlamydomonas. Biotechnol. Biofuels 2019, 12, 292. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.H. Culture and storage methods. In The Chlamydomonas sourcebook. A comprehensive guide to biology and laboratory use; Harris, E., Ed.; Academic Press: San Diego, CA, USA, 1989; pp. 25–63. [Google Scholar]
- Kropat, J.; Hong-Hermesdorf, A.; Casero, D.; Ent, P.; Castruita, M.; Pellegrini, M.; Merchant, S.S.; Malasarn, D. A revised mineral nutrient supplement increases biomass and growth rate in Chlamydomonas reinhardtii. Plant J. 2011, 66, 770–80. [Google Scholar] [CrossRef]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using Evans blue. Plant Cell, Tissue Organ Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Molnar, A.; Bassett, A.; Thuenemann, E.; Schwach, F.; Karkare, S.; Ossowski, S.; Weigel, D. Baulcombe, D. Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas reinhardtii. Plant J. 2009, 58, 165–174. [Google Scholar] [CrossRef]
- Kindle, K.L. High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. U.S.A. 1990, 87, 1228–1232. [Google Scholar] [CrossRef]
- Zalutskaya, Z.; Ostroukhova, M.; Ermilova, E. The Chlamydomonas alternative oxidase 1 is regulated by cadmium stress: new insights into control of expression. Environ. Exp. Bot. 2016, 130, 133–140. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Popov, N.; Schmitt, M.; Schulzeck, S.; Matthies, H. Reliable micromethod for determination of the protein content in tissue homogenates. Acta Biol. Med. Germ. 1975, 34, 1441–1446. [Google Scholar] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Maikova, A.; Zalutskaya, Z.; Lapina, T.; Ermilova, E. The HSP70 chaperone machines of Chlamydomonas are induced by cold stress. J. Plant Physiol. 2016, 204, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Lund, P. L-Glutamine and L-Glutamate: UV-method with glutaminase and glutamate dehydrogenase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; VCH, Verlagsgesellschaft: Weinheim, Germany, 1986; Volume 8, pp. 357–363. [Google Scholar]
- Sakaguchi, S. A new method for the colorimetric determination of arginine. J. Biochem. 1950, 37, 231–236. [Google Scholar] [CrossRef]
Figure 1.
Effects of ammonium and nitrate on cell growth and relative CrNAGK1 gene expression. (a) The growth curves were analyzed in the presence of 7.5 mM NH4Cl or 4 mM KNO3. Values are means ± SE of three independent experiments; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in ammonium- or nitrate-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1.
Figure 1.
Effects of ammonium and nitrate on cell growth and relative CrNAGK1 gene expression. (a) The growth curves were analyzed in the presence of 7.5 mM NH4Cl or 4 mM KNO3. Values are means ± SE of three independent experiments; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in ammonium- or nitrate-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1.

Figure 2.
Effects of ammonium and nitrate on the specific activity of CrNAGK and the total free content of Arg and Gln. (a) Time course of the CrNAGK activity during growth of cells in ammonium- or nitrate-containing medium. Cells were grown as described in Figure 1a; (b) Relationship between nitrogen source and intracellular Arg content; (c) Relationship between nitrogen source and intracellular Gln content. In (b,c): the content of amino acids is expressed in µg per106 cells. Values are means ± SE of three biological replicates.
Figure 2.
Effects of ammonium and nitrate on the specific activity of CrNAGK and the total free content of Arg and Gln. (a) Time course of the CrNAGK activity during growth of cells in ammonium- or nitrate-containing medium. Cells were grown as described in Figure 1a; (b) Relationship between nitrogen source and intracellular Arg content; (c) Relationship between nitrogen source and intracellular Gln content. In (b,c): the content of amino acids is expressed in µg per106 cells. Values are means ± SE of three biological replicates.
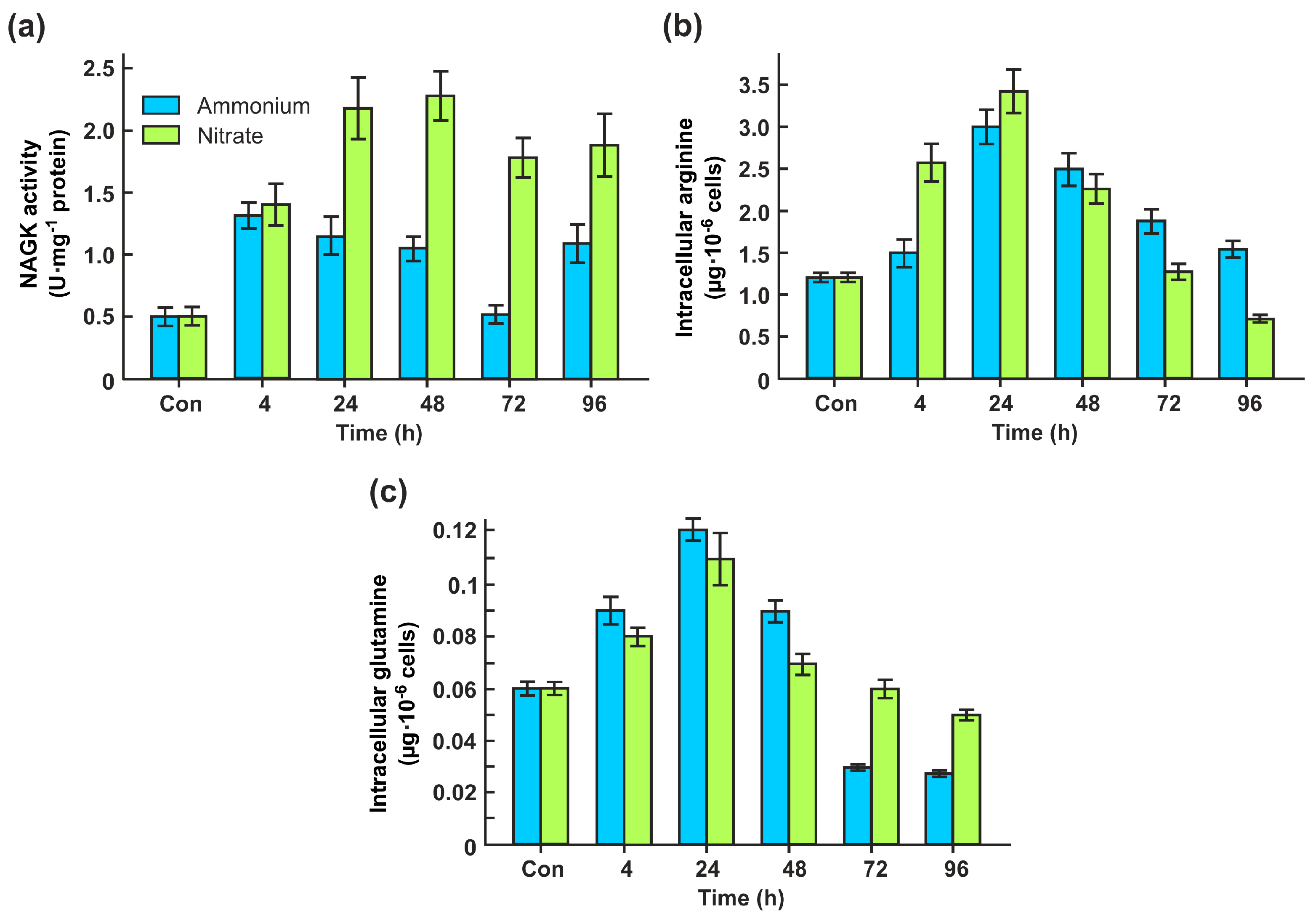
Figure 3.
Effects of nitrite on cell growth, CrNAGK expression and activity, and the total free content of Arg and Gln. (a) The growth curve was analyzed in the presence of 10 mM KNO2. Values are means ± SE of three independent experiments; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in nitrite-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1; (c) Time course of the CrNAGK activity during growth of cells in nitrite-containing medium; (d) Relationship between nitrogen source and intracellular Arg and Gln content. The concentration of amino acids are expressed in µg per106 cells. Values are means ± SE of three biological replicate.
Figure 3.
Effects of nitrite on cell growth, CrNAGK expression and activity, and the total free content of Arg and Gln. (a) The growth curve was analyzed in the presence of 10 mM KNO2. Values are means ± SE of three independent experiments; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in nitrite-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1; (c) Time course of the CrNAGK activity during growth of cells in nitrite-containing medium; (d) Relationship between nitrogen source and intracellular Arg and Gln content. The concentration of amino acids are expressed in µg per106 cells. Values are means ± SE of three biological replicate.
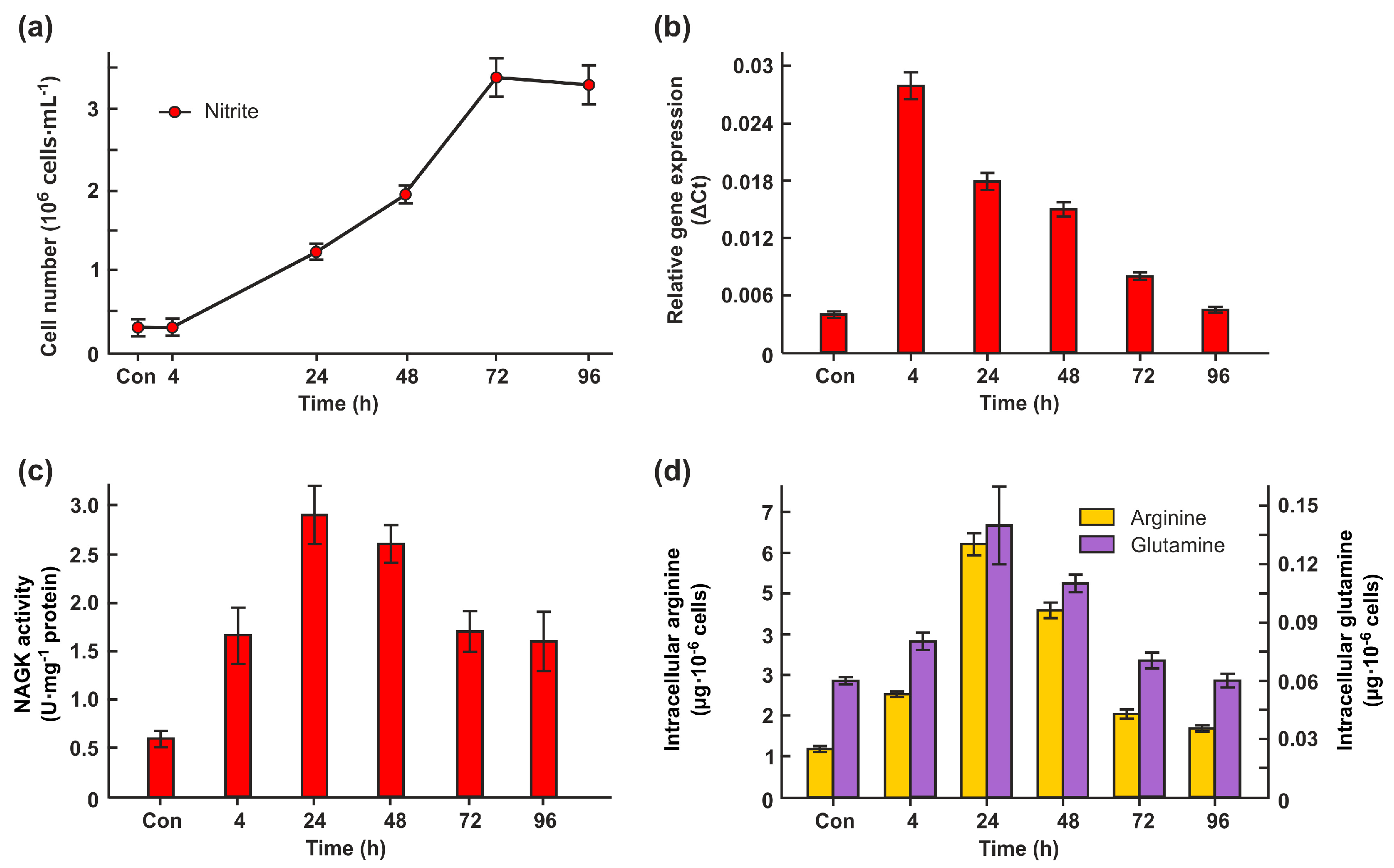
Figure 4.
Characterization of amiRNA-GLB1 strains. (a) CrPII abundance in wild types (CC4533 and CC3491), amiRNAGLB1-65 and amiRNAGLB1-88. Protein levels were analyzed by Western blotting. Each line corresponds to 40 μg of soluble proteins extracted from samples taken from cultures incubated in nitrite-containing medium for 24h. GLB1-66 and GLB1-88 indicate amiRNAGLB1-65 and amiRNAGLB1-88, respectively; (b) RT-qPCR analysis of CrGLB1 transcript levels. Relative expression levels were normalized with the gene expression of RACK1 and calculated using ΔCT. Samples were analyzed from cultures incubated in ammonium- or nitrite-containing medium for 24h.Values are means ± SE of three biological replicates and three technical replicates.
Figure 4.
Characterization of amiRNA-GLB1 strains. (a) CrPII abundance in wild types (CC4533 and CC3491), amiRNAGLB1-65 and amiRNAGLB1-88. Protein levels were analyzed by Western blotting. Each line corresponds to 40 μg of soluble proteins extracted from samples taken from cultures incubated in nitrite-containing medium for 24h. GLB1-66 and GLB1-88 indicate amiRNAGLB1-65 and amiRNAGLB1-88, respectively; (b) RT-qPCR analysis of CrGLB1 transcript levels. Relative expression levels were normalized with the gene expression of RACK1 and calculated using ΔCT. Samples were analyzed from cultures incubated in ammonium- or nitrite-containing medium for 24h.Values are means ± SE of three biological replicates and three technical replicates.
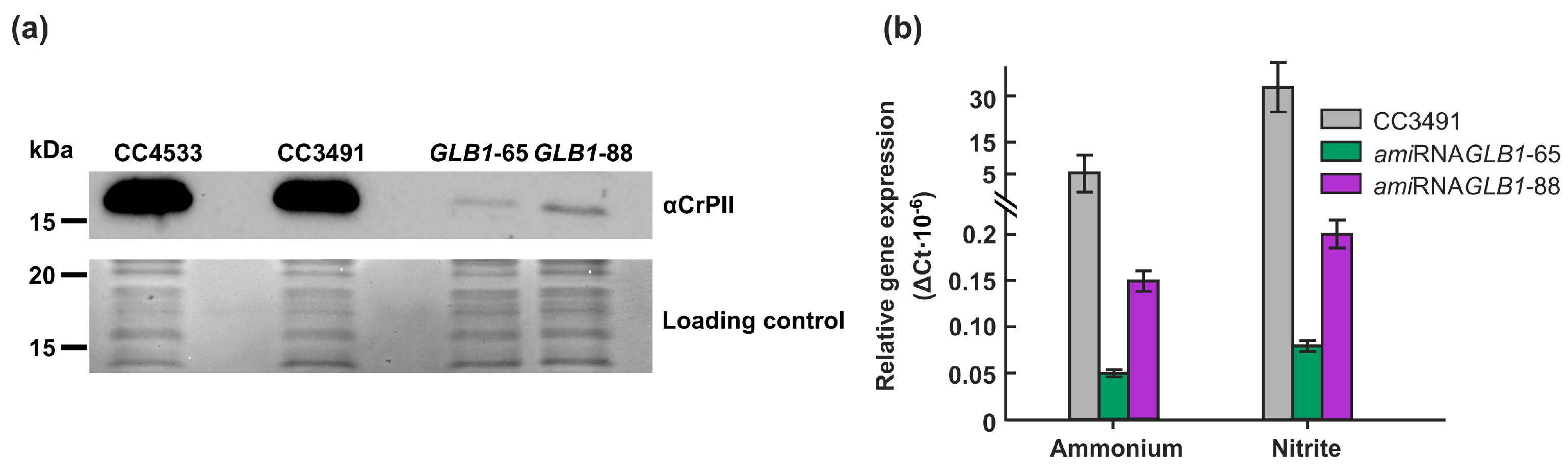
Figure 5.
Effects of ammonium and nitrite on CrNAGK expression and activity, and the total free content of Arg in CC3491 and GLB1-knockdown strains. (a) Time course of the CrNAGK activity during growth of cells in ammonium or nitrite-containing medium; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in ammonium or nitrite-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1; (c) Relationship between nitrogen source and intracellular Arg content. Intracellular Arg concentration at 0 h in each strain is considered as control (set to 100%). Values are means ± SE of three biological replicates. * and **denote significant differences between parental strain and CrGLB1-underexpressing transformants according to the Student’s t test (P value < 0.01 or < 0.05, respectively).
Figure 5.
Effects of ammonium and nitrite on CrNAGK expression and activity, and the total free content of Arg in CC3491 and GLB1-knockdown strains. (a) Time course of the CrNAGK activity during growth of cells in ammonium or nitrite-containing medium; (b) Time course of the CrNAGK1 transcripts accumulation during growth of cells in ammonium or nitrite-containing medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1; (c) Relationship between nitrogen source and intracellular Arg content. Intracellular Arg concentration at 0 h in each strain is considered as control (set to 100%). Values are means ± SE of three biological replicates. * and **denote significant differences between parental strain and CrGLB1-underexpressing transformants according to the Student’s t test (P value < 0.01 or < 0.05, respectively).
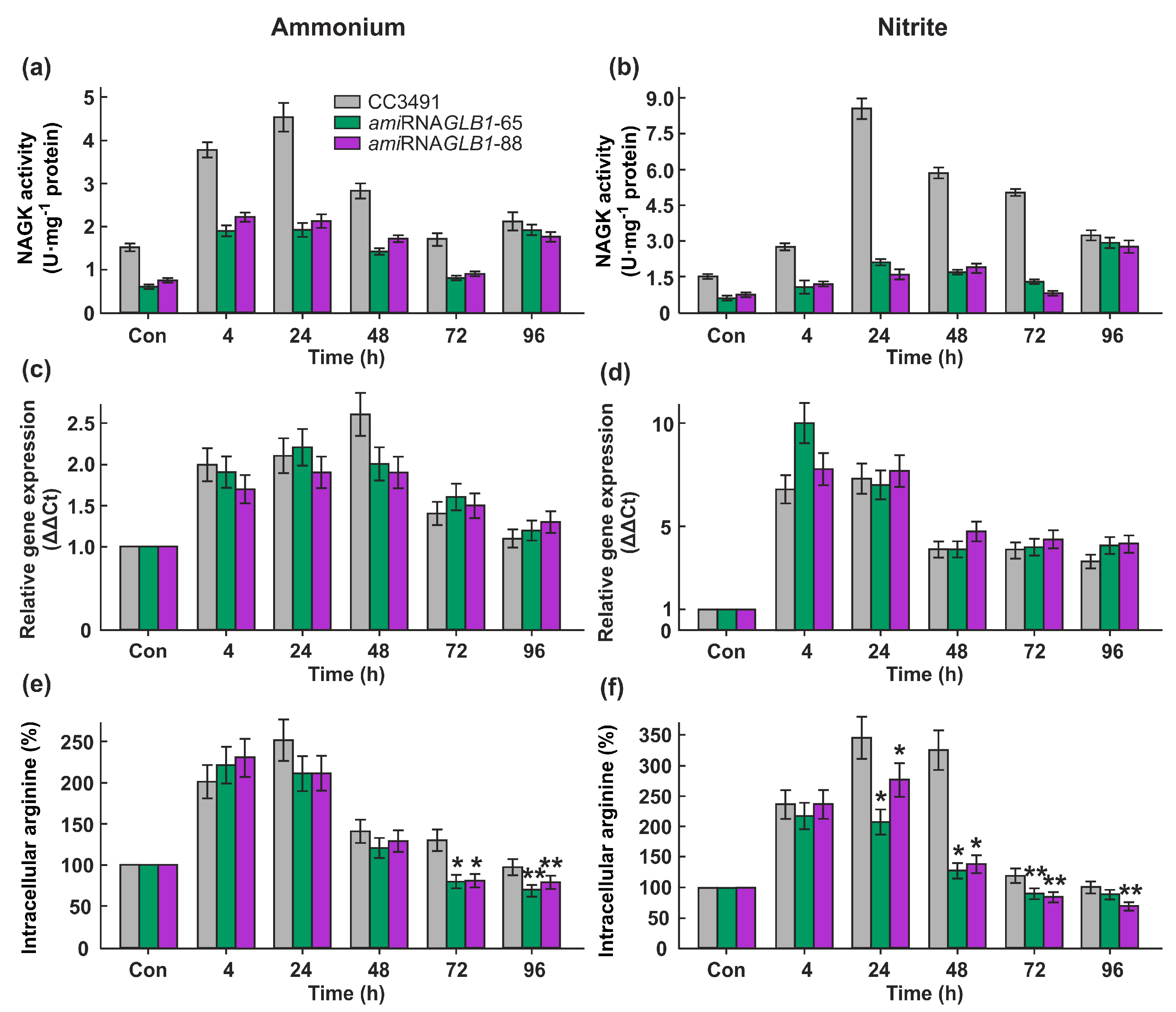
Figure 6.
Effects of N-deprivation on CrNAGK expression and activity, and the total free content of Arg in wild types and GLB1-knockdown strains. (a) CrNAGK activity and intracellular Arg content in 6145c strain during incubation of in N-free medium; * denotes significant differences between parental strain and CrGLB1-underexpressing transformants according to the Student’s t test (P value < 0.01 or < 0.05, respectively (b) Time course of the CrNAGK activity in parental strain and amiRNAGLB1 strains during incubation of in N-free medium; (c) Intracellular Arg content in parental strain and amiRNAGLB1 strains during incubation of in N-free medium; Intracellular Arg concentration in each strain in N-replete medium is considered as control (set to 100%). Values are means ± SE of three biological replicates; (d) Time course of the CrNAGK1 transcripts accumulation in parental strain and amiRNAGLB1 strains during incubation of in N-free medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1.
Figure 6.
Effects of N-deprivation on CrNAGK expression and activity, and the total free content of Arg in wild types and GLB1-knockdown strains. (a) CrNAGK activity and intracellular Arg content in 6145c strain during incubation of in N-free medium; * denotes significant differences between parental strain and CrGLB1-underexpressing transformants according to the Student’s t test (P value < 0.01 or < 0.05, respectively (b) Time course of the CrNAGK activity in parental strain and amiRNAGLB1 strains during incubation of in N-free medium; (c) Intracellular Arg content in parental strain and amiRNAGLB1 strains during incubation of in N-free medium; Intracellular Arg concentration in each strain in N-replete medium is considered as control (set to 100%). Values are means ± SE of three biological replicates; (d) Time course of the CrNAGK1 transcripts accumulation in parental strain and amiRNAGLB1 strains during incubation of in N-free medium. Values are means ± SE of three biological replicates and three technical replicates and are given as expression level relative to a house-keeping gene RACK1.
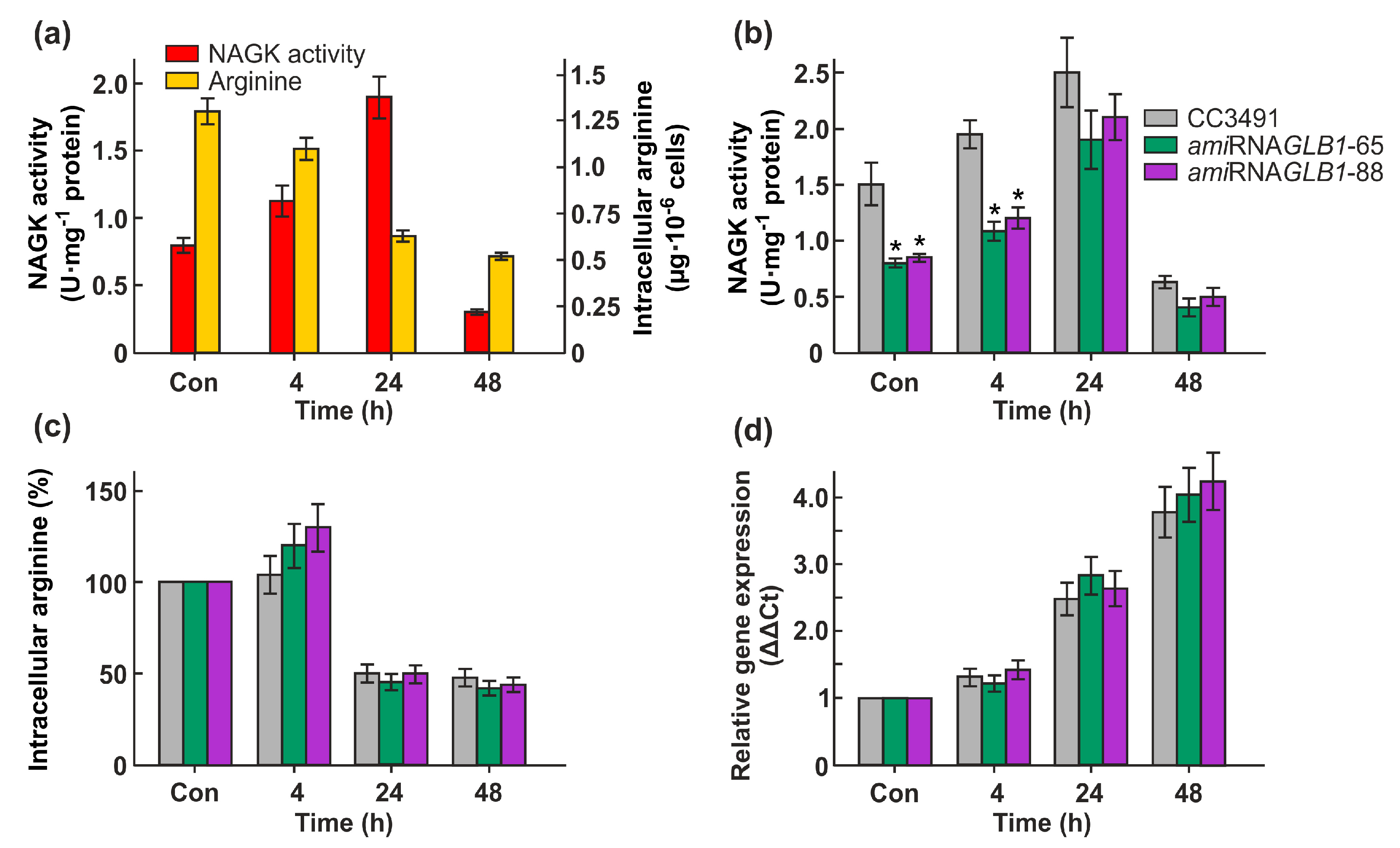
Figure 7.
Anticipated model of CrNAGK regulation under N-sufficiency and N-limitation. When N is available, the L-glutamine concentration increases, resulting in PII-NAGK complex formation. Under these conditions PII alleviates NAGK from Arg feedback inhibition and thereby enhances NAGK activity and Arg production. In addition, nitrate and nitrite upregulate NAGK1 and GLB1 gene expression. When cells became N-deficient for a short period of time, an increase in NAGK and PII levels appears enough to contribute to NAGK-PII complex formation and keep the enzyme active, which in turn results in elevated Arg synthesis. Under N-limitation for a long period of time, NAGK activity is decreasing and becomes the lowest. This could be achieved by releasing signal protein from the complex with NAGK through reduction in Gln content and/or PII sequestration by additional target. The release of PII results in stronger arginine feedback inhibition of NAGK, diminishing energy consumption and flux into arginine. An additional mechanism responsible for negative control of NAGK at posttranslational level is proposed. The positive transcriptional regulation is indicated by (+). The green arrows' width is indicative of the levels of Arg biosynthesis. Blunted lines denote the negative regulation at posttranslational level.
Figure 7.
Anticipated model of CrNAGK regulation under N-sufficiency and N-limitation. When N is available, the L-glutamine concentration increases, resulting in PII-NAGK complex formation. Under these conditions PII alleviates NAGK from Arg feedback inhibition and thereby enhances NAGK activity and Arg production. In addition, nitrate and nitrite upregulate NAGK1 and GLB1 gene expression. When cells became N-deficient for a short period of time, an increase in NAGK and PII levels appears enough to contribute to NAGK-PII complex formation and keep the enzyme active, which in turn results in elevated Arg synthesis. Under N-limitation for a long period of time, NAGK activity is decreasing and becomes the lowest. This could be achieved by releasing signal protein from the complex with NAGK through reduction in Gln content and/or PII sequestration by additional target. The release of PII results in stronger arginine feedback inhibition of NAGK, diminishing energy consumption and flux into arginine. An additional mechanism responsible for negative control of NAGK at posttranslational level is proposed. The positive transcriptional regulation is indicated by (+). The green arrows' width is indicative of the levels of Arg biosynthesis. Blunted lines denote the negative regulation at posttranslational level.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



