
Submitted:
25 July 2023
Posted:
26 July 2023
You are already at the latest version
Abstract
Exotic plants are usually problematic for the native species where they coexist. This study evaluated the effect of naturalised alien Cenchrus echinatus L. on native plants in the urban vegetation. A field trial was conducted to assess the effect of this species on cover and diversity of the native vegetation. The allelopathic potential of such species was examined. Plots comprising C. echinatus had lower cover of some native species. Lower floristic diversity was observed at more dense cover of this plant. The soil under Cenchrus attained lower N, P and K contents. This soil had no effect on germination and growth of the native species. It also comprised germinable seeds of some species which were absent from the standing vegetation. Exotic C. echinatus may exert a negative effect on the native vegetation of the urban plant communities. Dense cover of Cenchrus may inhibit germination of native species, leading to reduction in their cover. The reduction in cover and diversity of native species was not attributed to allelopathy. These results suggest that naturalised C. echinatus may be more competitor than the native ones, particularly at higher densities. Furthermore, it may represent a threat for the native plants in the urban vegetation.
Keywords:
naturalised alien species
; allelopathy
; Cenchrus echinatus
; weeds
; floristic diversity
; urban ecology
1. Introduction
In many parts of the world, invasion by alien plants usually represents a threat to the natural ecosystems. The alien species caused significant reduction in the resident species' diversity, disturbance in community structure, inhibition in vegetation growth and pronounced modification in soil bioprocesses [1,2,3,4,5].These alien species have been introduced from a certain region to another by human activities either accidently or intentionally [6]. Due to their destructive effects, scientists had to do highlights on these species and understand how biological invasions proceed and affect biodiversity. Consequently, it would be necessary to search for solutions to save the natural ecosystem [7]. Numerous studies have been conducted to show the effects of alien species. Moreover, they were increasing recently. However, most of these studies addressed the effect of invasive species, and little is known about the effects of naturalised ones. Therefore, it was necessary to fill this gap in knowledge.
Invasion success may be attributed to many factors that facilitate the dominance and wide spread of these species. Allelopathy and competition are key factors contributing towards successful establishment of alien species in the invaded communities [8]. Exotics may have stronger competitive ability for resources than native ones, causing adverse impacts or excluding the natives from their community [9,10]. This high competitive ability may be attributed to some of performance-related traits, such as high growth rate, leaf-area allocation, size and fitness, which promote invasion under different environmental conditions [11,12]. Dense cover of a species certainly facilitates competition of exotics. On the other hand, these species may produce allelopathic compounds that can manifest substantial inhibition for the associated plants directly throughout suppressing germination and growth performance, or indirectly by affecting soil microbes [8]. These compounds may be released into the soil via different methods such as exudation from plant roots, leaching from leaves, or decomposition of plant litter [13,14]. Both mechanisms seem to play, at least, a partial role in the substantial decrease in the abundance of resident species in the invaded communities. Since naturalised species are a subcategory under the alien ones and extend through wide areas [15], they may exhibit adverse effects as the same as the alien species do. In this investigation, we will study the effect of apparent cover and allelopathic potential of Cenchrus echinatus L., a naturalised alien species in Egypt, on the native associates. We also will examine its allelopathic potential using its rhizosphere soil that may contain allelopathic compounds.
Globally, the urban expansion continues to grow with significant increment in population growth and human activities. Urban green areas are considered as crucial ingredients of newly urbanized communities that necessarily attract citizens. Urban ecology is also considered as a field of interest for ecologists to study. Due to urbanization and plantation practices, new soils can be derived from old agricultural fields and small drainage canals and introduced into this region to be more suitable for plantations of gardens and parks. This practice may permit dispersal of plant species to this habitat. Consequently, urban green spaces may become hot points for non-native species, and it would be necessary to be in focus [16].
Non-native species are more closely associated with urban environment than resident ones [17]. This association may be due to influx of their propagules [18], habitat heterogeneity [19] and their ability to adapt with high levels of disturbances [20]. For these reasons, the urban environment accommodates high number of exotics that dominate a wide range of habitats and displace the natives [21]. Consequently, the presence of non-native plants in urban vegetation, in particular, may threaten the native species. Several studies emphasized the harmful effects of alien species on the associated species [22,23]. However, studies testing the effect of naturalised aliens in new urban areas are somewhat little.
Cenchrus echinatus L. is an annual weed species, native to tropical America and it has been introduced as a garden weed in Egypt [6]. It has also been recorded in gardens and parks of newly urbanized cities [24,25,26]. This plant could reproduce by seeds which are spread widely within their spiny fruits. Fruits with burrs can easily attach to animals, vehicles and clothing. They may also be dispersed by water and in contaminated agricultural products. It has the high capacity to spread and establish in newly reclaimed areas [27]. This species was recorded as naturalised alien species in Egypt [6,28]. The allelopathic effect of this species was scarcely explored. In this regard, previous studies investigated that the methanol and aqueous extracts from C. echinatus shoot and root extracts can to some extent inhibit the seed germination and growth criteria of some weed and crops [29,30,31]. Nevertheless, this is insufficient baseline work in the field of ecological allelopathy. In addition, its potential negative effects, as an alien species, in different plant communities are still unknown. Therefore, it was necessary to gather enough information about this. Our field observations indicated the presence of such species in the urban gardens and parks, with a clear reduction in cover of the native vegetation. The main hypothesis of this study was that the dense cover of C. echinatus in field may inhibit germination or, at least, reduce cover of the native species. This hypothesis could be tested in different ways. First, the covering areas and diversity of the native species were measured, in a field-based study, in selected sites comprising such species as well as locations completely free from it. Second, testing the seed bank in the soil in different sites dominated by C. echinatus. Existing of germinable/viable seeds in the location dominated by the alien species and free from the species observed in the seed bank may provide a proof for competition and/or allelopathy. As density of Cenchrus increases, the potential measured floristic diversity decreases. This can be assessed by measuring the diversity indices at different densities of Cenchrus. In addition, measurement of nutrient availability in soils of these sites may provide if this species is a strong competitor or not due to its potential nutrient withdraw. Third, the allelopathic potential of this species can be tested using the soil collected from sites with and without Cenchrus, as the rhizosphere soil of this species may contain allelopathic candidates inhibiting germination and growth of some target native species.
2. Results
2.1. Floristic Composition
In the vegetation involving Cenchrus echinatus, ten weed species belonging to five families were detected. The half of these species falls within the members of Poaceae, while the remaining species were belonging to other five different families of equal contribution. Besides, seven of these species were annuals (Table 1).
2.2. Effect of Cenchrus echinatus L. on Vegetation
In general, cover of most of the native species was reduced in the Cenchrus-affected plots (Table 1). All annuals were significantly affected (P<0.01). Amongst them, Dactyloctenium aegyptium seemed to be displaced completely from the plots with Cenchrus. Besides, Euphorbia peplus was markedly affected in comparison with the remaining annuals. For perennials, it was clear that cover of the cultivated Cynodon dactylon was significantly declined (P<0.01) in the plots comprising Cenchrus, whilst the remaining perennials were not affected (Table 1).
With respect to the measured diversity indices, there was a remarkable reduction trend for them as the density of Cenchrus increases (Figure 1). The negative effect of this species on plant diversity varied with respect to the measured index and the cover value of Cenchrus. In more detail, the species richness was not affected by the coexistence of Cenchrus. However, the degree of inhibition in the remaining diversity indices (Shannon-Wiener, Evenness and Simpson indices) depended on the cover of the alien species (Figure 1). Simpson index was significantly reduced at the cover of Cenchrus ≥ 70 %, while both evenness and Shannon-Wiener indices were reduced at Cenchrus cover ≥ 80 % (Figure 1). Besides, all of these indices were inversely correlated with the cover of the alien C. echinatus (Table 2).
Regarding the investigation of the soil seed bank in the plots with Cenchrus, four species were found to be emerging (Table 3). These species were absent in the Cenchrus-free plots. The highest numbers of seeds were recorded for Dactyloctenium aegyptium, after which the number of germinating seed of both Amaranthus viridis and Euphorbia paplus were statistically similar. Fewer numbers of seeds were detected for C. echinatus itself (Table 3).
2.3. Effect of C. echinatus on Soil Properties
In terms of soil criteria, the plots with Cenchus attained significantly lower contents of available macronutrient; namely nitrogen, phosphorus and potassium. However, these plots had higher amounts of available zinc (Table 4). The remaining soil properties were statistically similar in all plots.
2.4. Allelopathic Potential of C. echinatus
The results indicated that the soil under Cenchrus had no effect on germination and growth of three target native species (Figure 2).
3. Discussion
The present investigation showed that the patches of the gardens and parks where the alien Cenchrus echinatus was recorded attained lower cover of the native species. This result significantly suggests the negative impact of such exotic species under field conditions. Maybe this is one of the important interesting findings of this study. This observation was similar to that obtained by Hassan and Mohamed [3] who explained the negative effect of naturalised alien Paspalum dilatatum on the native urban vegetation. What we found in this study was the existence of viable/germinable seeds of different native species in the soil under C. echinatus. It is also worth mentioning that those species were completely absent in the above-ground vegetation in the locations where their seeds were found. This result substantially reflects the negative effect of dense-cover of the exotic plant on seed germination of the native ones. Consequently, reduction in cover of the native species may be related to decreased seed germination. On the other hand, the measured floristic diversity in this work was density dependent. In addition, the negative correlation between the cover of C. echinatus and the measured diversity indices was significant. In this regard, it was reported that interference effect of C. echinatus was higher as the density of this weed increases [43]. Germination may be delayed or completely inhibited due to the interspecific neighborhoods [44]. Besides, the negative density-dependent germination was observed by several authors e.g., [45,46].
Allelopathy may have role in weed interference and declining species diversity [47]. Moreover, naturalised alien plants may exert an allelopathic effect that inversely affects cover of the associated species [3,48]. This work showed that the soil under C. echinatus had no effect on germination and growth performance of the target species. This result suggests that allelopathy, as an interference mechanism, is not likely affecting cover and diversity of the native species and, consequently, it is difficult to claim that reduction in these criteria in the native plants in the sites comprising C. echinatus was attributed to allelopathy. This result substantially does not obey our hypothesis about the allelopathic potential of this species.
Our findings showed that some species were more affected by coexistence of the alien species in comparison with others. In particular, the results indicated that Dactyloctenium agyptium was completely displaced from the sites involving C. echinatus. In addition, Euphorbia peplus was highly suffering. This effect could be attributed to rarity of these species. It was shown that rarer species were subjected to more suppression by the community dominants than the introduced grasses [49]. In addition, Zhang and van Kleunen [9] concluded that common naturalised alien species were more competitive than rare native species, and this might lead to loss of rare native species. Furthermore, our field observations indicated the tendency of the alien species to prevail, resulting more inhibition for the native ones. Existence of both native species in the seed bank suggests the inhibitory effect of Cenchrus on germination of these species. Complete displacement of D. aegyptium may be related to the complete inhibition of its seed germination exerted by the alien species under field conditions. Its existence in the seed bank, but its absence in the standing vegetation, suggests the inability of this species, in particular, to cope with conditions of the above-ground vegetation [50], maybe exerted by the dense cover of the alien species.
One of the most remarkable results was the effect of C. echinatus on soil properties. This study revealed that the sites comprising this species were of lower soil macronutrients, particularly available N, P and K. Such result could reflect the heavy removal of these nutrients made by C. echinatus over the native species. This effect may explain why cover and diversity of native vegetation were declined. Nutrient deficiency may lead to reduction in the overall biodiversity [51]. Furthermore, high plant diversity could be associated with high levels of soil macronutrients [52,53]. On the other hand, this result may also show the better competitive ability of such alien species compared with the native plants. This connotation may be in agreement with that obtained by Tecco et al. [54] who showed a superior competitive ability of alien species over that of natives is often associated with a high ability to acquire and retain resources. Moreover, exotic plants may show better competitive ability than native ones [55].
4. Materials and Methods
4.1. Vegetation Survey
A field study was conducted in the study area described by Hassan [24] during 2018-2019 and repeated in 2021-2022 growing seasons. A total of 80 plots, each of 10 × 1 m, were randomly sampled from the vegetation representing the gardens and parks in the study area. Amongst them, 40 plots were chosen in the sites with C. echinatus (the invaded plots), whereas the remaining plots were selected in the Cenchrus-free sites (control plots). On average, the control plots were selected about 5 m apart from those comprising C. echinatus to be sharing similar environmental conditions. The total number of plots selected was proportional to the area occupied by this alien weed. Plot size was selected on a basis of a similar study performed in this area [25]. In each plot, the names of all species determined were listed. Species identification and nomenclature were carried out using Boulos [32,33,34,35]. The covering area of the species (expressed as m2) was virtually determined, and the relative cover of each species (pi) (expressed as %) was calculated through the equation: relative cover = [cover of a species i / cover of all species] × 100. Besides, the floristic diversity was measured as four diversity indices: species richness (S), i.e. the total number of plant species observed, Shannon-Wiener index (H'), Evenness index (E) and Simpson’s index (D) [36,37] as follows:
- H' = - ∑(pi×ln pi)
- E = [-∑(pi×ln pi)] / ln S
- D = 1 / C and C= ∑pi2
4.2. Soil Seed Bank Sampling
The soil seed bank was sampled in the stands representing dense cover of C. echinatus (cover of Cenchrus ≥ 70 % and the remaining vegetation was covered by the cultivated Cynodon) to avoid existence of seeds of other species from the above-ground vegetation, and to ascertain that the inhibition of germination of the detected seeds could be attributed to presence of the alien species. At such cover %, reduction in plant diversity was highly apparent. Four soil samples each at 5 cm depth were randomly collected from the upper 5 cm of soil around representative individuals from the stands with Cenchrus, while control soils were selected from the stands without Cenchrus. This depth was considered as most seeds have high tendency to be located at this depth [38]. Soil samples were sieved through a 2 mm sieve to remove plant litter. Known volumes of sieved soil samples were spread randomly in a 2 cm layer overlying sterilized soil in 10 cm × 12 cm pots in a greenhouse sharing the prevailing environmental conditions at Beni –Suef University and regularly irrigated. Control pots contained the soils free from Cenchrus for a potential comparison with the seeds detected in Cenchrus-infested soils. The germinable/viable seeds were assessed by monitoring the seedling emergence. Emerging seedlings were counted and left to be juvenile for possible identification by the mentioned Boulos handbooks.
4.3. Soil Analysis
From each plot, three soil samples were collected from 0–20 cm depth and pooled together forming one composite. They were air dried, sieved through 2 mm sieve and stored in plastic bags for analysis. Determination of soil texture, pH, electrical conductivity (EC), and the contents of CaCO3, organic carbon (OC) and availability of some nutrient including N, P, K and Zn was performed.
For determination of soil texture, i.e. the percentage of sand, silt and clay in the inorganic fraction of soil, the hydrometer method was followed [39]. Both pH and EC were measured in soil-water extract using deionized water. Soil pH was measured in the soil extract (1:2.5 w/v) with a digital pH meter (AD 3000), while EC was measured in another soil extract (1:5 w/v) using a conductivity meter (Jenway 3305). The soil content of CaCO3 was estimated using the titration method [40]. Soil organic carbon was measured using Walkley and Black's method [41]. The available soil nutrients including nitrogen, phosphorus, potassium, zinc and copper in the soil samples were determined using Allen's methods [42].
4.4. Allelopathic Potential of C. echinatus
4.4.1. Soil Sampling and Preparation
Soil samples from the rhizosphere of C. echinatus in Cenchrus-dominating plots were collected and exposed to effective shaking inside a plastic bag to get one composite as the treatment soil (the invaded soil). The non-rhizosphere soil samples (control soil) were collected from the sites dominated by Bermuda grass (Cynodon dactylon); the native species cultivated in these gardens and parks for the greening purpose. Both soil types were sieved (2 mm size) to remove the plant litter and placed in plastic pots (10 cm diameter × 15 cm depth each) for near sowing of the target species. Each pot contained about one kg of air-dried soil.
4.4.2. The Target Species
Three native target species namely: Amaranthus viridis (Linn.), Dactyloctenium aegyptium L. Willd. and Eragrostis pilosa (L.) P.Beauv. were used as test species. These species are common weeds in the study area [25]. The preliminary field observation showed that D. aegyptium appeared to have been away/apart from the sites with C. echinatus, whereas the remaining species had lower cover in these sites in comparison with the Cenchrus-free areas. This observation may reflect the impact level of the alien species. Ripe seeds of these species were collected from different locations constituting the gardens and parks of the study area (i.e. the New Beni-Suef city) at the late fruiting stage.
4.4.3. Test for Allelopathy
Thirty seeds of the target species were equally disseminated at 0.3 cm depth of soil per pot. The pots were irrigated regularly via misting process when need. The experiment was retained in a protected site under the prevailing environmental conditions in a completely randomized design with four replicates for 30 days. At the end of the test, the numbers of emerging seedlings were determined to calculate the germination percentage. Growth criteria including shoot height, root depth and the total biomass were measured per each individual as indicated by Hassan and Mohamed [3].
4.5. Statistical Analyses
The data were first checked for normality and homogeneity of variances using the Kolmogorov–Smirnoff and Levene's tests, respectively. The results meeting the requirement of normality and homoscedasticity were analysed using independent-samples T test. When field data were not normally distributed, the non-parametric Mann Whitney U test was performed. To evaluate a relationship between the cover of the exotic C. echinatus and the different diversity indices, a Pearson correlation analysis was carried out. All analyses were carried out using SPSS, version 20.0 (IBM Corporation, USA).
5. Conclusions
To great extent, the results of this study followed the main hypothesis proposed. Coexistence of the exotic naturalised Cenchrus echinatus in the urban vegetation led to remarkable decline in cover of most of the associated native plants. Rare species; particularly Dactyloctenium aegyptium and Euphorbia peplus, were more affected in comparison with the remaining native species. The adverse effect of this species on plant diversity varied with respect to the measured index and its cover value. Species richness was not affected at all, whereas the remaining indices were reduced at higher densities of the alien species. Moreover, all of these indices were inversely correlated with cover values of such plant. Interestingly, Presence of this alien species resulted in significant reduction in the available soil macronutrients including nitrogen, phosphorus and potassium. In addition, the soil under this species indicated the existence of germinable /viable seeds of some species which were absent from the standing vegetation. However, the rhizosphere soil under this species had no effect on germination and growth performance of the native target species under greenhouse conditions. These results suggest the negative effect of the alien C. echinatus on the native species in the urban vegetation. Reduction in soil macronutrients may be attributed to heavy removal of these macronutrients made by C. echinatus. Besides, failure of germination of some native species in field may be attributed to the dense cover of Cenchrus. Consequently, the naturalised alien C. echinatus may be more competitor than the native ones. Allelopathic potential of such species seemed to be inactive mechanism affecting cover and diversity of native species. Till now, C. echinatus has not been recorded as an invasive one. However, its negative effects on the native vegetation and soil were addressed. This study substantially indicated that naturalised alien C. echinatus represents a threat for the urban vegetation. Conservation strategies should be devoted to control alien species in the urban ecosystems.
Author Contributions
M.O.H.: Conceived and designed the experiments; Performed the experiments; Analyzed and interpreted the data; Contributed reagents, materials, analysis tools or data, Wrote the paper. H.Y.M.: contributed in writing the manuscript. A.A.: Performed the experiments; Analyzed and interpreted the data; Wrote the manuscript. M.K.O. and B.H.K.: revised the manuscript and provided the funding agency. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors extend their appreciation to the Researchers Supporting Project number (RSP2023R374) King Saud University, Riyadh, Saud Arabia. Hassan also extends his gratitude to his wife for her support and kindness during this work. Hassan would like to thank Nasr H. Gomaa for his guidance and help during this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Castro-Díez, P.; Alonso, Á. Effects of non-native riparian plants in riparian and fluvial ecosystems: a review for the Iberian Peninsula. Limnetica 2017, 36, 525–541. [Google Scholar] [CrossRef]
- Gaertner, M.; Den Breeyen, A.; Hui, C.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: a meta-analysis. Prog. Phys. Geogr. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Hassan, M.O.; Mohamed, H.Y. Allelopathic interference of the exotic naturalized Paspalum dilatatum Poir. threatens diversity of native plants in urban gardens. Flora 2020, 266, 151593. [Google Scholar] [CrossRef]
- Qu, T.; Du, X.; Peng, Y.; Guo, W.; Zhao, C.; Losapio, G. Invasive species allelopathy decreases plant growth and soil microbial activity. PloS one 2021, 16, 1–12. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol. Let. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- El-Beheiry, M.; Hosni, H.; Sharaf El-din, A.; Shaltout, S.K.; Ahmed, D. Updating the checklist of the alien flora in Egypt. Taeckholmia 2020, 40, 41–56. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Invasive species, environmental change and management, and health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef]
- Raizada, P.; Raghubanshi, A.S.; Singh, J.S. Impact of invasive alien plant species on soil processes: A review. Proc. Nat. Acad. Sci. India, Sect. B. 2008, 78, 288–298. [Google Scholar]
- Zhang, Z.; van Kleunen, M. Common alien plants are more competitive than rare natives but not than common natives. Ecol. Let. 2019, 22, 1378–1386. [Google Scholar] [CrossRef]
- Yuan, L.; Li, J.M.; Yu, F.H.; Oduor, A.M.; van Kleunen, M. Allelopathic and competitive interactions between native and alien plants. Biol. Invasions 2021, 23, 3077–3090. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Let. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: where do we stand?. In Biological invasions.; Nentwig, W.; Springer, Berlin, Heidelberg, 2007, volume 193, pp. 97–125. [CrossRef]
- Callaway, R.M.; Ridenour, W.M.; Laboski, T.; Weir, T.; Vivanco, J.M. Natural selection for resistance to the allelopathic effects of invasive plants. J. Ecol. 2005, 93, 576–583. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. . Non-native species in urban environments: patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef]
- Williams, N.S.; Hahs, A.K.; Vesk, P.A. Urbanisation, plant traits and the composition of urban floras. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 78–86. [Google Scholar] [CrossRef]
- Malavasi, M.; Carboni, M.; Cutini, M.; Carranza, M.L.; Acosta, A.T. Landscape fragmentation, land-use legacy and propagule pressure promote plant invasion on coastal dunes: a patch-based approach. Landsc. Ecol. 2014, 29, 1541–1550. [Google Scholar] [CrossRef]
- Deutschewitz, K.; Lausch, A.; Kühn, I.; Klotz, S. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany. Glob. Ecol. Biogeogr. 2003, 12, 299–311. [Google Scholar] [CrossRef]
- Sukopp, H. Human-caused impact on preserved vegetation. Landsc. urban plan. 2004, 68, 347–355. [Google Scholar] [CrossRef]
- Štajerová, K.; Šmilauer, P.; Brůna, J.; Pyšek, P. Distribution of invasive plants in urban environment is strongly spatially structured. Landsc. Ecol. 2017, 32, 681–692. [Google Scholar] [CrossRef]
- Dalle Fratte, M.; Bolpagni, R.; Brusa, G.; Caccianiga, M.; Pierce, S.; Zanzottera, M.; Cerabolini, B.E. Alien plant species invade by occupying similar functional spaces to native species. Flora 2019, 257, 151419. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.,.....; Lastrucci, L. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 1–13. [CrossRef]
- Hassan, M.O. Leaf litter of Bombax ceiba L. threatens plant cover and floristic diversity in a new urban ecosystem. Flora 2018, 242, 22–30. [Google Scholar] [CrossRef]
- Hassan, M.O.; Hassan, Y.M. Effect of human activities on floristic composition and diversity of desert and urban vegetation in a new urbanized desert ecosystem. Heliyon 2019, 5, e02283. [Google Scholar] [CrossRef]
- Hassan, M.O.; Mohamed, H.Y.; Aboellil, A.H. Allelopathic potential of Ficus retusa L. leaf litter on understory vegetation in urban gardens. Acta Bot. Croat. 2021, 80, 131–139. [Google Scholar] [CrossRef]
- Shehata, H.S. Ecology and allelopathic control of the invasive species Cenchrus echinatus L. in some of newly reclaimed areas in Nile Delta, Egypt. Life Sci J. 2014, 11, 246–260. [Google Scholar]
- Shaltout, K.H.; Hosni, H.A.; El-Kady, H.F.; El-Beheiry, M.A.; Shaltout, S.K. Composition and pattern of alien species in the Egyptian flora. Flora-Morphol. Distrib. Funct. Ecol. Plants 2016, 222, 104–110. [Google Scholar] [CrossRef]
- Nascimento, E.A.; Terrones, M.G.; Morais, S.A.; Chang, R.; Andrade, G.A.; Santos, D.Q.; Pereira, B.H. Allelopathic activity of Cenchrus echinatus L. extracts on weeds and crops. Allelopath. J. 2009, 24, 363–372. [Google Scholar]
- Ma, W.; Miao, S.; Tao, W.; Xu, Z.; Wang, H.; Chen, J.; Huang, L. Study on the Allelopathic Effects of Alien Invasive Species Cenchrus echinatus on Seed Germination and Seedling Growth of Solanaceae Crops. Agric. Sci. Technol. 2014, 15, 885–889. [Google Scholar]
- Favaretto, A.; Scheffer-Basso, S.M.; Perez, N.B. Allelopathy in Poaceae species present in Brazil. A review. Agron. Sustain. Dev. 2018, 38, 1–12. [Google Scholar] [CrossRef]
- Boulos, L. Azollaceae–Oxalidace. In Flora of Egypt.; Al Hadara Publishing: Cairo, Egypt, 1999; Vol. 1. [Google Scholar]
- Boulos, L. Geraniaceae–Boraginaceae. In Flora of Egypt.; Al Hadara Publishing: Cairo, Egypt, 2000; Vol. 2. [Google Scholar]
- Boulos, L. Verbenaceae–Compositae. In Flora of Egypt.; Al Hadara Publishing: Cairo, Egypt, 2002; Vol. 3. [Google Scholar]
- Boulos, L. Monocotyledons: Alismataceae–Orchidaceae. In Flora of Egypt.; Al Hadara Publishing: Cairo, Egypt, 2005; Vol. 4. [Google Scholar]
- Pielou, E.C.; Ecological Diversity. Wiley, London, 1975. [CrossRef]
- Zhang, J.T. Quantitative Methods in Vegetation Ecology. China Science and Technology Press, Beijing, China, 1995.
- Gutterman, Y. Seed Germination in Desert Plants, 1st ed.; Springer Berlin, Heidelberg, Germany, 1993. [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis-advanced Course. Washington department of soil sciences, USA, 1967.
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publishers, Oxford, London, 1989.
- da Silva Maia, S.; Ribeiro-Rocha, P.R.; Santiago-Castro, T.; Costa da Silva, I.K. E.; Ferreira-Barreto, G.; Torres de Souza, L.; Murga-Orrillo, H.; Abanto-Rodríguez, C. Interference of Cenchrus echinatus and Rottboellia exaltata in growth of cowpea crop. Bioagro 2021, 33, 21–28. [Google Scholar] [CrossRef]
- Tielbörger, K.; Prasse, R. Do seeds sense each other? Testing for density-dependent germination in desert perennial plants. Oikos 2009, 118, 792–800. [Google Scholar] [CrossRef]
- Grundy, A.C.; Mead, A.; Burston, S. Modelling the germination response of weed seeds to burial depth: interactions with seed density, weight and shape. J. Appl. Ecol. 2003, 40, 757–770. [Google Scholar] [CrossRef]
- Hassan, M.O.; Gomaa, N.H.; Fahmy, G.M.; González, L.; Hammouda, O.; Atteya, A.M. Interactions between Sonchus oleraceus L. and some weeds in agroecosystems in Egypt. Ann. Agric. Sci. 2014, 59, 221–228. [Google Scholar] [CrossRef]
- Foy, C.L.; Inderjit. Understanding the role of allelopathy in weed interference and declining plant diversity. Weed Technol. 2001, 15, 873–878. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 1–15. [Google Scholar] [CrossRef]
- Wade, G.L. Grass competition and establishment of native species from forest soil seed banks. Landsc. Urban Plan. 1989, 17, 135–149. [Google Scholar] [CrossRef]
- Anju, M.V.; Warrier, R.R.; Kunhikannan, C. Significance of Soil Seed Bank in Forest Vegetation—A Review. Seeds 2022, 1, 181–197. [Google Scholar] [CrossRef]
- Morgan, J.B.; Connolly, E.L. Plant-Soil Interactions: Nutrient Uptake. Nat. Educ. Knowl. 2013, 4, 2. [Google Scholar]
- Furey, G.N.; Tilman, D. Plant biodiversity and the regeneration of soil fertility. Ecol. 2021, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nat. 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Tecco, P.A.; Díaz, S.; Cabido, M.; Urcelay, C. Functional traits of alien plants across contrasting climatic and land-use regimes: do aliens join the locals or try harder than them? J. Ecol. 2010, 98, 17–27. [Google Scholar] [CrossRef]
- Vilà, M.; Weiner, J. Are invasive plant species better competitors than native plant species? Evidence from pair-wise experiments. Oikos 2004, 105, 229–238. [Google Scholar] [CrossRef]
Figure 1.
Measured diversity indices (Mean ± SE) in relation to the different densities (%) of C. echinatus. Density at 0 value represents Cenchrus-free stands. Values sharing the same letter are statistically similar (at P < 0.05) within the same parameter.
Figure 1.
Measured diversity indices (Mean ± SE) in relation to the different densities (%) of C. echinatus. Density at 0 value represents Cenchrus-free stands. Values sharing the same letter are statistically similar (at P < 0.05) within the same parameter.
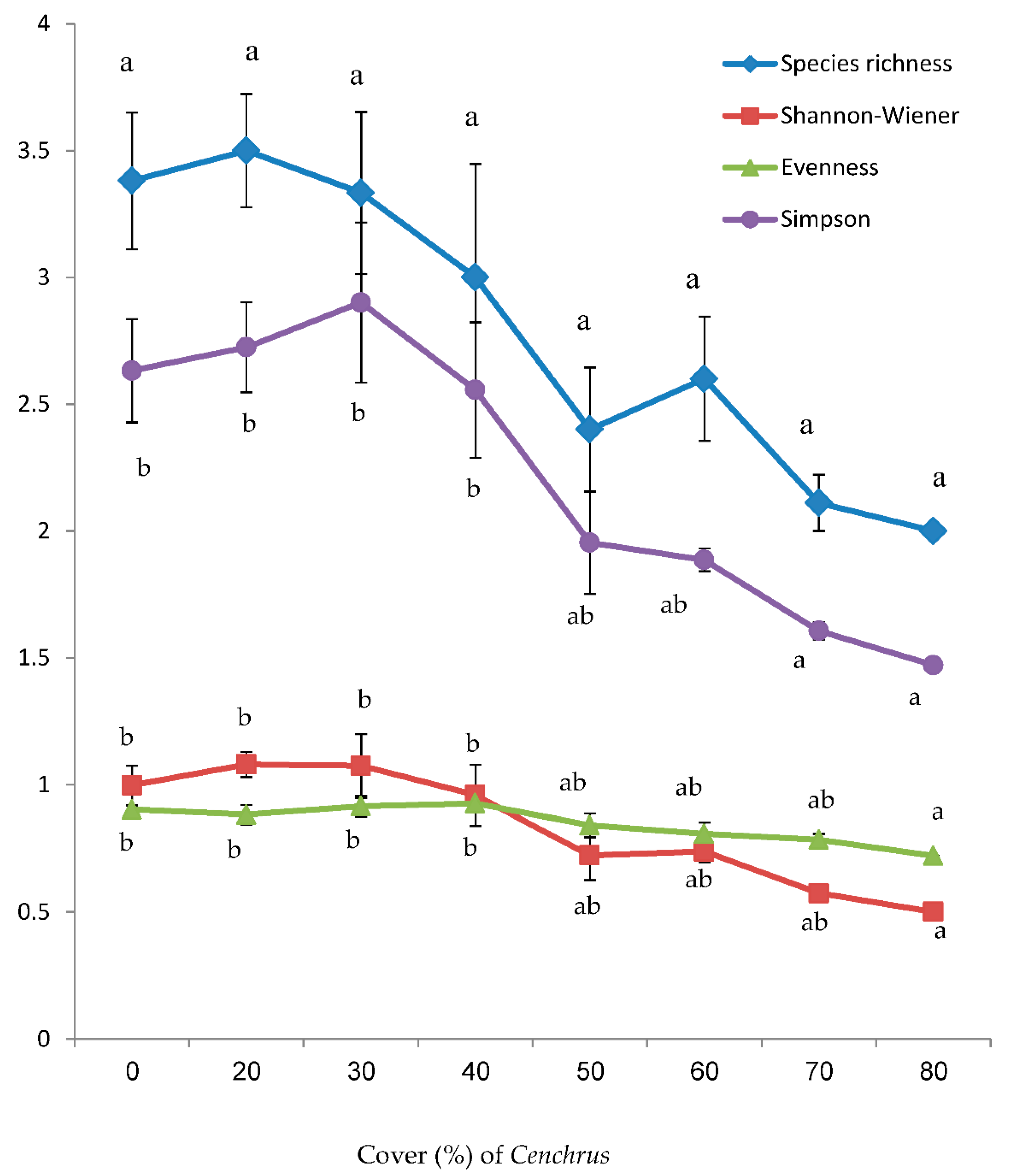
Figure 2.
Germiation (%) and the measured growth criteria (Mean, bars represent standard errors) of the selected target species associated with C. echinatus when planted in Cenchrus-infested and Cenchrus-free soils. .
Figure 2.
Germiation (%) and the measured growth criteria (Mean, bars represent standard errors) of the selected target species associated with C. echinatus when planted in Cenchrus-infested and Cenchrus-free soils. .
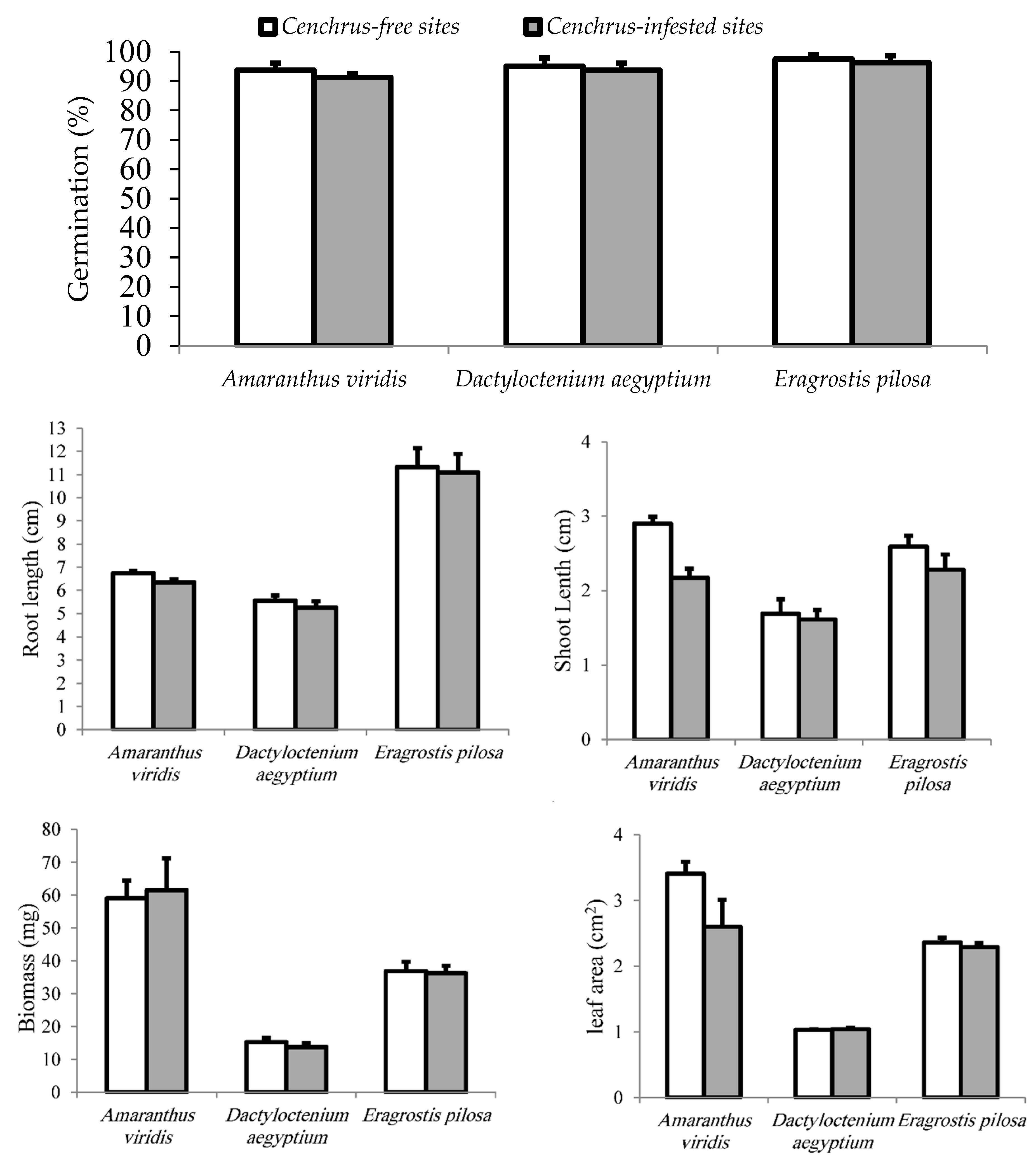

Table 1.
Mean coverage values (m2) (mean ± SE) of the plant species detected in the Cenchrus-free and Cenchrus-infested plots.
Table 1.
Mean coverage values (m2) (mean ± SE) of the plant species detected in the Cenchrus-free and Cenchrus-infested plots.
| Species | Family | Cenchrus-Free Plots | Cenchrus-Infested Plots |
|---|---|---|---|
| Amaranthus viridis L. † | Amaranthaceae | 39.82 ± 4.23 | 15.10** ± 4.41 |
| Apium leptophyllym Pers. † | Apiaceae | 15.60 ± 3.48 | 3.9** ± 0.45 |
| Cynodon dactylon Pers. †† | Poaceae | 142.21 ± 13.25 | 97.38** ± 9.46 |
| Dactyloctenium aegyptium (L.) Willd. † | Poaceae | 3.30 ± 0.64 | 0.0** ± 0.0 |
| Dichanthium annulatum (Forssk.) Stapf †† | Poaceae | 19.28 ± 4.48 | 8.50 ± 3.86 |
| Digitaria sanguinalis (L.) Scop. † | Poaceae | 16.38 ± 3.18 | 11.90 ± 3.27 |
| Eragrostis pilosa (L.) P.Beauv. † | Poaceae | 49.74 ± 8.42 | 14.81** ± 3.50 |
| Oxalis corniculata L. †† | Oxalidaceae | 17.11 ± 6.79 | 9.44 ± 2.33 |
| Euphorbia peplus L. † | Euphorbiaceae | 4.35 ± 0.60 | 0.35** ± 026 |
| Plantago lagopus L. † | Plantaginaceae | 17.00 ± 2.54 | 3.63** ± 2.63 |
| Species | Family | Cenchrus-free plots | Cenchrus-infested plots |
| Amaranthus viridis L. † | Amaranthaceae | 39.82 ± 4.23 | 15.10** ± 4.41 |
| Apium leptophyllym Pers. † | Apiaceae | 15.60 ± 3.48 | 3.9** ± 0.45 |
| Cynodon dactylon Pers. †† | Poaceae | 142.21 ± 13.25 | 97.38** ± 9.46 |
| Dactyloctenium aegyptium (L.) Willd. † | Poaceae | 3.30 ± 0.64 | 0.0** ± 0.0 |
| Dichanthium annulatum (Forssk.) Stapf †† | Poaceae | 19.28 ± 4.48 | 8.50 ± 3.86 |
| Digitaria sanguinalis (L.) Scop. † | Poaceae | 16.38 ± 3.18 | 11.90 ± 3.27 |
| Eragrostis pilosa (L.) P.Beauv. † | Poaceae | 49.74 ± 8.42 | 14.81** ± 3.50 |
| Oxalis corniculata L. †† | Oxalidaceae | 17.11 ± 6.79 | 9.44 ± 2.33 |
| Euphorbia peplus L. † | Euphorbiaceae | 4.35 ± 0.60 | 0.35** ± 026 |
| Plantago lagopus L. † | Plantaginaceae | 17.00 ± 2.54 | 3.63** ± 2.63 |
* P < 0.01 † Annual †† Perennial.
Table 2.
Correlation coefficients (r) between the measured diversity indices and the cover of C. echinatus L.
Table 2.
Correlation coefficients (r) between the measured diversity indices and the cover of C. echinatus L.
| Diversity indices | Cover of Cenchrus |
|---|---|
| Species richness | - 0.54** |
| Shannon index | - 0.70** |
| Evenness | - 0.35* |
| Simpson index | - 0.71** |
* P < 0.05, ** P < 0.01.
Table 3.
Number of germinating seeds of the species per pot detected in the seed bank in the soil dominated by C. echinatus compared with the Cenchrus-free sites.
Table 3.
Number of germinating seeds of the species per pot detected in the seed bank in the soil dominated by C. echinatus compared with the Cenchrus-free sites.
| Species | Life Span | Cenchrus-Free Sites | Cenchrus-Infested Sites |
|---|---|---|---|
| Amaranthus viridis L. | Annual | - | 9.72** ± 1.01 b |
| Cenchrus echinatus L. | Annual | - | 2.73** ± 0.33 a |
| Dactyloctenium aegyptium (L.) Willd. | Annual | - | 14.80** ± 0.90 c |
| Euphorbia peplus L. | Annual | - | 9.64** ± 0.98 b |
- Not detected. ** P < 0.01.
Table 4.
Soil physicochemical properties (mean ± SE) in the Cenchrus-free and Cenchrus-infested plots.
Table 4.
Soil physicochemical properties (mean ± SE) in the Cenchrus-free and Cenchrus-infested plots.
| Soil Properties | Cenchrus-Free Sites | Cenchrus-Infested Sites |
|---|---|---|
| Sand (%) | 48.39 ± 2.15 | 49.00 ±1.60 |
| Silt (%) | 32.62 ±1.58 | 32.02 ±1.50 |
| Clay (%) | 18.99 ± 0.93 | 18.78 ± 0.70 |
| pH | 7.98 ± 0.03 | 7.92 ± 0.03 |
| EC (µS cm-1) | 377.54 ± 15.50 | 377.08 ±10.50 |
| CaCO3 (%) | 2.47 ± 0.09 | 2.56 ± 0.07 |
| OC (%) | 1.97 ± 0.07 | 2.05 ± 0.05 |
| Available nutrients (mg kg-1 soil) |
||
| N | 67.2 ± 1.50 | 51.12** ± 3.90 |
| P | 12.55 ± 0.42 | 10.50** ± 0.45 |
| K | 411.02 ± 5.70 | 361.15** ± 6.20 |
| Zn | 1.60 ± 0.13 | 6.10** ± 0.31 |
| Cu | 2.28 ± 0.13 | 2.45 ± 0.14 |
** P < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



