
Submitted:
26 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
Infections with symbiotic single-celled organisms classified as Ciliophora, are commonly reported in various species of invertebrates. Freshwater mussels also play the role of a host for them, being responsible for their biodiversity. Knowledge about these relationships is insufficient. Freshwater mussels are among the most endangered groups of animals. Ciliate endosymbionts in mussels may be responsible for the diseases and increasingly frequent cases of mass mussel deaths and are particularly dangerous for commercially farmed mussels and for restocking of mussels as part of active conservation measures. On the other hand the Ciliophora parasites and commensals living in freshwater mussels is undescribed. Many of these species are likely to become extinct before they are known to science. Recently, two genera of Ciliophora, Conchophthirus sp. and Trichodina sp., have been described in the mantle cavity of U. crassus - an endangered and protected species in the EU. The basis of extensive research on the impact of endosymbiotic ciliates on U. crassus populations is the knowledge of their species composition, proportion of infected individuals, level of infection intensity and distribution in different types of rivers. Such studies were carried out in the mantle cavity of U. crassus from three rivers in three seasons. Cloning, sequencing and functional analysis of the genetic material of Ciliophora was carried out using NGS (Next-Generation Sequencing) analysis based on the hypervariable V4 and V9 regions of the 18SrRNA gene, which enables the identification of taxonomic groups, including genera and species. The most numerous OTUs are common and cosmopolitan species. But some commensals and potentially parasites were found too. Ciliophora associated with mantle cavity form an interesting, hierarchical biocomplex but their interactions with mussels need further studies.
Keywords:
Ciliophora
; DNA barcoding
; NGS sequencing
; endosymbionts
; parasites
; freshwater mussels Unionidae
1. Introduction
The term biodiversity [1] is always intuitively understood to mean the species richness in a given biotic community. Plethora of papers was published trying to understand factors maintaining biodiversity and the mechanisms, which enable human kind to maintain it. Common slogan “diversity makes stability” proposed by [2], refers to the presumed stabilizing role of diversity in ecological systems, and, despite doubts [3], is still commonly used to explain the important role of biological diversity in stabilizing ecosystems. Although anthropogenic species extinction decoupled concern for the loss of biodiversity with concern for ecosystem stability the concept of diversity of species richness was increasingly understood to refer to genetic diversity, diversity of biotic communities and ecosystems functions [4]. This directly lead to the new concept of “biocomplexity” to characterize complex natural systems that harbour biodiversity. In order to maintain the latter, multiple levels of biological organisation has to be studied, as well as interacting feedbacks and nonlinear or hierarchical relations between components of biotic diversity and their ambient environment. This needs not only the break of traditional approach of measuring species diversity, but needs truly transdisciplinary approach [5].
The biodiversity crisis in Antropocen is overwhelming, however, there are areas where situation seems to be especially critical. This refers to freshwater ecosystems [6]. Large freshwater mussels (Bivalvia, Unionida; hereafter, naiads) inhabiting freshwater in high number and biomass [7] provide important ecosystem functions and services in large scale [8,9,10], however, belong to the least known group of animals and are in need of urgent conservation actions [11].
The important role in regulating complexity of biological processes is played by parasites [12] and as was pointed out by [13], parasite biodiversity may be at risk of extinction, especially if their hosts also face extinction, like it was already proposed for the parasites of naiads [14]. The role of parasites in freshwater mussels might be evidently negative, like castrating trematodes [15], however, in many cases their impact on the host is not clear if any [16]. This especially refers to Protozoans, which were identified as endosymbionts of some naiads living in their mantle cavity, however, only in few cases their role was studied and impact determined [14].
Large freshwater mussels are known to influence local biodiversity by their numerous relations with ecosystem and other species [17]. However, from the point of studying biocomplexity, it is very interesting that on one side, naiads occurrence depend on the fish host, because larvae of naiads must complete obligatory phase of parasitic life on the fish [18]. On the other hand it is an interesting question, what diversity might be found within the ecosystem which naiads create within their shell cavity, where e.g. threatened species of naiad can secure safe development of larvae of protected fish species (e.g. bitterling – [19]). The “shell cavity” system can be quite rich, because naiads can be hosts for evidently negative endosymbionts like castrating trematodes. They can harbor endosymbionts which role is not recognized yet, but potentially with large impacts on hosts, like bacteria (e.g. Wolbachia – [20]) or for microorganisms which may have negative effects on their progeny (like glochidia eating ciliate – [21]). This is a rich community of organisms adapted to within-shell ecosystem of freshwater mussels, which diversity and relation to the host and other organisms is not recognized yet. To fill that gap we would like to present the result of our analysis of Ciliophora diversity inhabiting the organism of threatened species of freshwater mussel, which, potentially, may disappear if the threatened host will die-out. We wanted to answer some basic questions on the species composition, seasonal abundance of Ciliophora and their relation to general habitat.
2. Materials and Methods
2.2. Study Object
For the study of Ciliophora endosymbionts in mussels we chose as an object of the study thick-shelled river mussel U. crassus (Bivalvia: Unionidae) – threatened species, listed in Red List of IUCN [22], protected by directives of EU and national low [23]. It is a middle size freshwater mussel, inhabiting only flowing waters, which numbers decreased rapidly in the 20th century. The species is locally numerous and the individuals were collected from the rivers which has safe populations of very high abundance of the mussel, with relevant permissions (General Directorate for Environmental Protection, Poland No. DZP-WG.6401.128.2022.ASZ.1).
2.2. Study Area and Field Sampling
In order to study Ciliophora endosymbionts occurring in the mantle cavity of the U. crassus, 3 rivers were selected in Central Europe (Poland): the San, the Czarna Włoszczowska and the Czarna Hańcza (Table 1.). In accordance with the permits in each of the sampled rivers, individuals were collected 3 times a year: 10 individuals at the beginning of reproduction (May), 10 during the period of larvae release (July), and 10 after breeding period (September). Previous studies have shown, that the populations of U. crassus inhabiting the rivers differ genetically and in terms of the species composition of the microbiome found in their digestive tracts [24].
Specimens were collected if microscopic inspection (Bresser Researcher Trino 40-1000) of a slide from a sample taken from the interior of the mantle cavity, revealed the presence of ciliates. The mussels were transported in containers with cooled and oxygenated water to the laboratory where they were immediately rinsed in the gill cavity with distilled water. The resulting suspension was concentrated on a filter and fixed in 70% ethanol and stored frozen at -20°C for further analysis.
2.3. Laboratory analyses
DNA was extracted for all 90 samples using a commercial kit (Sherlock AX Purification Kit, A&A Biotechnology, Poland). The concentration and purity of the isolated DNA was measured using a NanoDrop spectrophotometer. The following were selected for metagenomic sequencing isolates: sample concentration: >= 30 ng/μl and sample purity: OD260/280 = 1.8 - 2.0. For each isolate, amplification of Ciliophora genetic material was carried out. The PCR reaction was carried out using Q5 Hot Start High-Fidelity 2X Master Mix, reaction conditions as according to the manufacturer's recommendations. For amplification of the selected region and library preparation, there were used specific primer sequences forwardfito and reversefito (V4 analysis) and 1391Fciliouc and EukBciliouc (V9 analysis). The selected primers amplified the V4 region (amplicon size 450 bp (base pairs)) and V9 region (about 200 bp) of the 18S rRNA gene. The length of amplified molecular markers and their quality were checked in a 1% agarose gel, with the addition of a DNA intercalating substance. Three samples for each river in each season studied were selected for further analysis. These were samples in which there was the most of DNA. The selected three samples from the same date for a given river were mixed and analysed as one. In this way, two markers (V4 and V9) were examined and DNA data were obtained for Ciliophora for each river in each of the three seasons studied.
Metagenomics of the Ciliophora profile was performed by Next-Generation Sequencing, based on the hypervariable region of V4 (primers provided by Tragin et al. 2017 [25]) and V9 (primers designed by Amaral-Zettler et al. 2009 [26]) of the 18S rRNA gene, which enables identification of taxonomic groups. Metagenomic sequencing was carried out using the Illumina's MiSeq sequencer, using paired-end (PE) technology. Automated preliminary data analysis was performed on the MiSeq instrument using the MiSeq Reporter (MSR) v2.6 software, and the analysis consisted of the two steps: (1) automatic de-multiplexing of samples, (2) generation of fastq files containing raw readings.
Bioinformatic analysis, providing classification of reads to species level (filtering of reads and analysis of sample composition for each taxonomic category of the studied microorganisms based on homology to the databases), was carried out using the QIIME software [27], based on the Pr2 reference sequence database. The analysis consisted of steps: (1) removal of adaptor sequences - cutadapt program, (2) analysis of the quality of the reads and removal of sequences with low quality (quality < 20, minimum length 30) - cutadapt program [28], (3) combination of sequences paired - fastq-join algorithm [29], (4) removal of chimeras of sequences - usearch61 algorithm, (5) clustering on a based on the selected reference sequence base - uclust algorithm, (6) assignment of taxonomy to the selected reference sequence base - uclust algorithm [30].
3. Results and discussion
For all 9 samples, subjected to metagenomic analysis, a sufficient number of sequences was found, allowing accurate insight into the profile of Ciliophora (from 839 to 29239 sequences). The number of isolated OTUs was 98 (at 98% identity). This allowed the assignment of nearly 40 taxa at the genus or species level.
The most numerous reads of OTU of V4 region (Figure 1A) belonged to the Sessilida X sp., followed for Hypotrichia XX sp., Vorticella aequilata, Pseudourostyla cristata, Tokophrya sp., and Urostyla grandis.
The most numerous reads of OTU of V9 region belong also to the Sessilida X sp. followed by Halteria grandinella, Operculariidae X sp., Carchesium polypinum, and Urostyla grandis. (Figure 1B).
Considering the particular taxons which might be of interest from the point of view of mussels conservation, the resulting OTUs confirmed the occurrence of the genera Trichodina sp. and Conchophthirus sp. They were found in U. crassus in earlier microscopic observations which have not showed mussel’s tissue damage caused by ciliate [31]. However the ciliates Conchophthirus sp., Trichodina sp. and Tetrahymena sp. had a range of negative correlations with other endosymbionts species, such as the mites Unionicola intermedia and U. bonze and fish bitterling larvae Rhodeus amarus, occupying the gills and mantle of the unionid mussels Anodonta anatina and Unio pictorum [32]. It was also described that essentially commensal Trichodina sp. may stay abundant in stressed or debilitated fish and may penetrate host tissues causing disease [33]. So such phenomena may be expected in U. crassus. Trichodina sp., on the other hand, is found in large numbers on each date only in the Czarna Włoszczowska river. In the Czarna Hańcza this taxon was found only in spring, and in the San it appears very sparsely or not at all, regardless of the season.
Three OTUs were obtained for Conchophthirus sp., the taxon was detected in samples from all rivers and in all seasons studied, although in low frequency in the San. The taxon was reported earlier in microscopic studies of all native species of unionid hosts in Poland (except for U. crassus), so it seems to be their common endosymbiont, e.g. [34].
The presence of Tetrahymena sp., which species are suspected to kill the mussels larvae glochidia [21], was also confirmed, however, in none of the surveyed rivers it was found in the spring, at the time when glochidia are brooded, and it was not found at all in the San River.
The mean number of the reads in given sample (May, July, September) was increasing during the season (Figure 2A,B) in the most of the trials, although some disturbances of this pattern occurred in the Czarna Włoszczowska river for V9 region (Figure 2B).
The number of OTU (taxons) detected for given river was the highest in the Czarna Włoszczowska, slightly lower in the Czarna Hańcza, and disproportionally lower in the San river. The interesting pattern is visible in the number of OTUs detected in given samples during the season (Figure 3): it is increasing (like reads do) during the season in both lowland rivers, whereas in the San it increases by the order of magnitude: from 2 OTU in May to 26 in September. May and July samples also share very small number of OTU between the rivers, this number increases distinctively as late as in September. That indicates the high influence of season on the development of ciliate communities within mantle cavity.
The species of the highest number of reads belong to cosmopolite species (e.g. Vorticella aequilata, Halteria grandinella, Carchesium polypinum, Pseudourostyla cristata, Urostyla grandis, Figure 1), which seem to be not detrimental to mussels. The difficulty in finding their relation to freshwater mussels stems from the fact, that mussel filtrate water, thus many Ciliophora identified genetically may be not related directly with mussel but simply enter the shell cavity with filtered water. Some of them, like Carchesium polypinum is known as a generalist epibiont and was reported as commensal e.g. on the Ephemera danica Muller 1764, (Ephemeroptera: Ephemeridae) sampled in two small lowland rivers in Poland [35]. In other Polish river Ciliophora belonging Epistylididae (Sessilida) epibiotic ciliates were reported on the signal crayfish Pacifastacus leniusculus [36]. But role one of the ciliate taxon, Tokophrya sp., especially needs to explain, because three species of the genus were found in pathologically changed gills of the Red Swamp Crayfish Procambarus clarkii [37]. Such found ciliate taxons as Sessilida X sp., Hypotrichia XX sp. or Operculariidae X sp. are unknown and needs detailed studies.
The mutual relations of the Ciliophora to mussels are usually unknown, which might be a consequence of very large number of ciliates which have been described as symbionts mainly of individuals of metazoan phylla [38]. For that reason, the genetic methods seems to be the only reasonable quantitative approach to study of ecological relations between Ciliophora and their possible hosts. It is important to concentrate on these Ciliophora species, which might have significant negative or positive impact on their hosts. It was demonstrated that some of Ciliophora might be detrimental to molluscs, as a ciliate parasite Tetrahymena rostrata infecting the renal organ of the dusky slug (Arion fuscus) [39], and T. foissneri sp. n. hosted by invasive Lusitanian slug (Arion vulgaris) and T. unionis sp. n. detected in freshwater mussel Unio tumidus [40], as well as, like Tetrahymena glochidiophila n. sp, which attacks larvae of naiads (glochidia; [21]). In our study the some species of Tetrachymena genus were found, however, it might be also possible that species recorded in Poland also can be harmful to glochidia.
It is also worth to mention that many Ciliophora might be highly detrimental to fish [41,42], which rise interesting questions: (1) do mussel host species detrimental to fish, (2) how detrimental they can be to fish species, which are hosts to naiads larvae (e.g. chub Leuciscus cephalus, [43]). The second question is even more important for conservation of the threatened naiads, because their larvae must pass obligatory parasitic period on fish host to complete development [18]. Thus, existence of naiads hosting Ciliophora may depend on the existence of fish hosts of naiads, which Ciliophora may attack [44]. These is a system of parasitism composed on three elements, influencing each other, which forms an interesting example of biocomplexity, considering additional impact of river type.
The increasing pattern of the mean reads per river during the season might simply reflect the population development of Ciliophora after the winter, when temperature of water is very low, likely not advantageous for Ciliophora reproduction, mussels are burrowed in sediment, shell is closed and it is difficult to say if their population grows from some propagules saved inside the mantle cavity or it must develop in each season from the initial colonization from the outside. The similar seasonal population growth of Conchophthirus acuminatus in invasive Dreissenids was observed by Karateyev et al. 2000 [45], although some big seasonal variation was also present. On the opposite, Blazhekovikj-Dimovska & Stojanovski (2020) [46] found rather similar levels of Trichodina sp. infestation intensity between the seasons in two species of carp in temperate climate of Macedonia.
In analysed data, the number of species differ very much between the rivers, depending on their character: lowland rivers are much more rich in both OTUs and reads than the mountainous river San. The Czarna Włoszczowska river, which flows through the agricultural landscape with fish ponds in the river valley, has higher number of OTU than the primaeval and more oligotrophic the Czarna Hańcza. The difference however, is much smaller between each of the lowland rivers in reference to the mountainous river San (Figure 2 and Figure 3). Although it would be difficult to infer about the abundance of OUTs between the rivers, in case of both studied hypervariable regions (V4 and V9) the mean number of reads is strikingly lower for the San river (Figure 2). The differences between rivers in e-DNA of Ciliophora were reported before, e.g. [47], however, differences reported here seems to be outstanding.
Despite that Ciliophora communities are closed within the mantle cavity of the same species, is by no means under the influence of general habitat, which was demonstrated by extreme differences in general intensity of the Ciliophora infection and uniqueness of species composition between lowland rivers and mountainous one. This difference might be related to hydrology of the habitat: lowland rivers are stable and waves of water change are usually mild and prolonged, carrying little sediment, mostly diluting only the floating biota concentration within the channel or not influencing them severely. In mountainous rivers discharge increase cause very short but high flow waves, which have very high energy, large water velocity and transport of sediment of various size, which in conjunction with turbulent flow creates conditions likely leading to destruction of any life forms within the water or on the surface of the channel [48]. Such catastrophes occur frequently in mountains, thus abundance of freshwater biota, especially those small and having no possibility of resistance or escape, must decrease after such an event. If such events happen frequently can keep frequency of Ciliophora occurrence at very low level.
4. Conclusions
Genetic studies of Ciliophora symbionts of naiads should be supplemented with microscope inspection of their tissues in order to find, whether Ciliophora associated with mantle cavity are epibiotas or just random admixture of the water inhaled by the mussel. The diversity of the species largely depends on habitat in terms of site and seasonal change in their prevalence and infestation intensity. At present, it is impossible to determine, how much Ciliophora can be detrimental to mussels and whether mussels can be a storage of ciliate detrimental to their host fish or other orgsanisms, however, they form an interesting, hierarchical biocomplex.
Author Contributions
K.Z.: conceptualization, funding acquisition; M.M., T.Z.: conceptualization (supporting); K.Z., T.Z. writing—original draft preparation, M.M.: molecular analysis methodology; All authors: investigation, writing—review and editing (equal). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Centre, Poland (program MINIATURA 5: the grant no. 2021/05/X/NZ2/01839 to Katarzyna Zając).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
This publication is based upon work from COST Action CA18239, supported by COST (European Cooperation in Science and Technology).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilson, E.O., The current state of biological diversity. Biodiversity, 1988, 521(1), 3-18.
- McArthur, W.H. Observations of the enteric protozoa of Rana pipiens during larval development and metamorphosis. Proceedings of the Iowa Academy of Science (Iowa Acad Sci), 1955, 62: 640-651.
- Tilman, D., Isbell, F., Cowles, J.M. Biodiversity and ecosystem functioning. Annual review of ecology, evolution, and systematics, 2014, 45, 471-493. https://doi.org/10.1126/science.1064088. [CrossRef]
- Baird Callicott, J., Rozzi, R., Delgado, L., Monticino, M., Acevedo, M. and Harcombe, P. Biocomplexity and conservation of biodiversity hotspots: three case studies from the Americas. Philosophical Transactions of the Royal Society B: Biological Sciences, 2007, 362(1478), 321-333. https://doi.org/10.1098/rstb.2006.1989. [CrossRef]
- Dybas, C.L., From Biodiversity to Biocomplexity: A Multidisciplinary Step toward Understanding Our Environment: Highlights of the 52nd Annual Meeting of the American Institute of Biological Sciences, 24–26 March 2001, Washington, DC. BioScience, 2001, 51(6), 426-430. https://doi.org/10.1641/0006-3568(2001)051[0426:FBTBAM]2.0.CO;2. [CrossRef]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Current Biology, 2019, 29(19), R960-R967. https://doi.org/10.1016/j.cub.2019.08.002. [CrossRef]
- Strayer, D.L., Hunter, D.C., Smith, L.C., Borg, C.K. Distribution, abundance, and roles of freshwater clams (Bivalvia, Unionidae) in the freshwater tidal Hudson River. Freshwater Biology 1994, 3, 239–248. https://doi.org/10.1111/j.1365-2427.1994.tb00858.x. [CrossRef]
- Newton, T.J., Zigler, S.J., Rogala, J.T., Gray, B.R. and Davis, M. Population assessment and potential functional roles of native mussels in the Upper Mississippi River. Aquatic Conservation: Marine and Freshwater Ecosystems, 2011, 21(2), 122-131. https://doi.org/10.1002/aqc.1170. [CrossRef]
- Vaughn, C. C. Ecosystem services provided by freshwater mussels. Hydrobiologia, 2018, 810 (1), 15–27. https://doi.org/10.1007/s10750- 0 17-31. [CrossRef]
- Zieritz, A., Sousa, R., Aldridge, D. C., Douda, K., Esteves, E., Ferreira-Rodríguez, N., Mageroy, J. H., Nizzoli, D., Osterling, M., Reis, J., Riccardi, N., Daill, D., Gumpinger, C., & Vaz, A. S. A global synthesis of ecosystem services provided and disrupted by fresh-water bivalve molluscs. Biological Reviews, 2022, 97, 1967–1998. https://doi.org/10.1111/BRV.12878. [CrossRef]
- Sousa, R., Zając, T., Halabowski, D., Aksenova, O.V., Bespalaya, Y.V., Carvalho, F., Castro, P., Douda, K., da Silva, J.P., Ferreira-Rodríguez, N. and Geist, J. A roadmap for the conservation of freshwater mussels in Europe. Conservation Biology, 2023, 37(2), p.e13994. https://doi.org/10.1111/cobi.13994. [CrossRef]
- Hudson, P.J., Dobson, A.P. and Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends in ecology & evolution, 2006, 21(7), 381-385. https://doi.org/10.1016/j.tree.2006.04.007. [CrossRef]
- Dunn, R. R., Harris, N. C., Colwell, R. K., Koh, L. P., Sodhi, N. S. The sixth mass coextinction: are most endangered species parasites and mutualists? Proceedings of the Royal Society B: Biological Sciences, 2009, 276(1670), 3037-3045. https://doi.org/10.1098/rspb.2009.0413. [CrossRef]
- Brian, J.I., Aldridge, D.C. Mussel parasite richness and risk of extinction. Conservation Biology, 2022, 36(6), p.e13979. https://doi.org/10.1111/cobi.13979. [CrossRef]
- Jokela, J., Uotila, L., Taskinen, J. Effect of the castrating trematode parasite Rhipidocotyle fennica on energy allocation of fresh-water clam Anodonta piscinalis. Functional Ecology 1993, 7, 332-338. https://doi.org/10.2307/2390213. [CrossRef]
- McElwain, A. Are parasites and diseases contributing to the decline of freshwater mussels (Bivalvia, Unionida)?. Freshwater Mollusk Biology and Conservation, 2019, 22(2), 85-89. https://doi.org/10.31931/fmbc.v22i2.2019.85–89. [CrossRef]
- Vaughn, C. C., Nichols, S. J., Spooner, D. E. Community and foodweb ecology of freshwater mussels. Journal of the North American Benthological Society, 2008, 27(2), 409-423. https://doi.org/10.1899/07-058.1. [CrossRef]
- Kat, P.W. Parasitism and the Unionacea (bivalvia). Biological Reviews, 1984, 59(2), 189-207, https://doi.org/1111/j.1469-185X.1984.tb00407.x. [CrossRef]
- Tatoj, K., Ćmiel, A. M., Kwaśna, D., Lipińska, A. M., Zając, K., Zając, T. The endangered thick-shelled river mussel (Unio crassus): a new host species for the European bitterling (Rhodeus amarus). Biodiversity and Conservation, 2017, 26, 1217-1224. https://doi.org/10.1007/s10531-017-1295-y. [CrossRef]
- Mioduchowska, M., Konecka, E., Gołdyn, B., Pinceel, T., Brendonck, L., Lukić, D., Kaczmarek, Ł., Namiotko T., Zając K., Zając T., Jastrzębski J. P., Bartoszek, K. Playing Peekaboo with a Master Manipulator: Metagenetic Detection and Phylogenetic Analysis of Wolbachia Supergroups in Freshwater Invertebrates. International Journal of Molecular Sciences, 2023, 24(11), 9400. https://doi.org/10.3390/ijms24119400. [CrossRef]
- Lynn, D. H., Doerder, F. P., Gillis, P. L., Prosser, R. S. Tetrahymena glochidiophila n. sp., a new species of Tetrahymena (Ciliophora) that causes mortality to glochidia larvae of freshwater mussels (Bivalvia). Diseases of Aquatic Organisms, 2018, 127(2), 125-136. https://doi.org/10.3354/dao03188. [CrossRef]
- Lopes-Lima , M., Kebapçı, U., Van Damme, D. Unio crassus. The IUCN Red List of Threatened Species 2014: e.T22736A42465628. Accessed on 10 July 2023. https://doi.org/10.2305/IUCN.UK.2014-1.RLTS.T22736A42465628.en. [CrossRef]
- Cuttelod, A., Seddon, M. and Neubert, E. European Red List of Non-marine Molluscs. Luxembourg: Publications Office of the European Union, 2011, https://doi.org/10.2779/84538. [CrossRef]
- Mioduchowska, M., Zając, K., Bartoszek, K., Madanecki, P., Kur, J., Zając, T. 16S rRNA-based metagenomic analysis of the gut microbial community associated with the DUI species Unio crassus (Bivalvia: Unionidae). Journal of Zoological Systematics and Evolutionary Research 2020, 58: 615–623. https://doi.org/10.1111/jzs.12377. [CrossRef]
- Tragin M., Zingone A., Vaulot D. Comparison of coastal phytoplankton composition estimated from the V4 and V9 regions of the 18S rRNA gene with a focus on photosynthetic groups and especially Chlorophyta. Environmental Microbiology, 2017, 20, 506–520. https://doi.org/10.1111/1462-2920.13952. [CrossRef]
- Amaral-Zettler, L.A., McCliment, E.A., Ducklow, H.W., Huse, S.M. A method for studying protistan diversity using massively parallel sequencing of V9 hypervariable regions of small-subunit ribosomal RNA genes. PLoS One 2009, 4 (7), e6372. https://doi.org/10.1371/journal.pone.0006372. [CrossRef]
- Caporaso, J.G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F.D., Costello, E.K., Fierer, N., Gonzalez Pena, A., Goodrich, J.K., Gordon, J.I., Huttley, G.A., Kelley, S.T., Knights, D., Koenig, J.E., Ley, R.E., Lozupone, C.A., McDonald, D., Muegge, B.D., Pirrung, M., Reeder, J., Sevinsky, J.R., Turnbaugh, P.J., Walters, W.A., Widmann, J., Yatsunenko, T., Zaneveld, J., Knight, R. QIIME allows analysis of high-throughput community sequencing data. Nature Methods 2010, 7(5), 335- 336. https://doi.org/10.1038/nmeth.f.303. [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. journal, 2011, 17(1), 10-12. https://doi.org/10.14806/ej.17.1.200. [CrossRef]
- Aronesty. ea-utils: “Command-line tools for processing biological sequencing data”, 2011 http://code.google.com/p/ea-utils.
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 2010, 26(19), 2460-2461. https://doi.org/10.1093/bioinformatics/btq461 0. [CrossRef]
- Lewisch, E., Arnold, F., Fuehrer, H.P., Harl, J., Thielen, F. and El-Matbouli, M. Parasites and their impact on thick-shelled river mussels Unio crassus from two populations in Luxembourg. Diseases of Aquatic Organisms, 2023, 153, 31-43. https://doi.org/10.3354/dao03718. [CrossRef]
- Brian, J.I., Aldridge, D.C. Abundance data applied to a novel model invertebrate host shed new light on parasite community assembly in nature. Journal of Animal Ecology, 2021, 90(5), 1096-1108. https://doi.org/10.1111/1365-2656.13436. [CrossRef]
- Bruno, D. W., Nowak, B., Elliott, D. G. Guide to the identification of fish protozoan and metazoan parasites in stained tissue sections. Diseases of aquatic organisms, 2006, 70(1-2), 1-36. https://doi.org/10.3354/dao070001. [CrossRef]
- Taskinen, J., Urbańska, M., Ercoli, F., Andrzejewski, W., Ożgo, M., Deng, B., Choo, J. M., Riccardi, N. Parasites in sympatric populations of native and invasive freshwater bivalves. Hydrobiologia, 2021, 848, 3167-3178. https://doi.org/10.1007/s10750-020-04284-0. [CrossRef]
- Grzybkowska, M., Szczerkowska-Majchrzak, E., Dukowska, M., Leszczyńska, J., & Przybylski, M. Ephemera danica (Ephemeroptera: Ephemeridae) as a resource for two commensals: ciliated protozoans (Sessilida) and chironomids (Diptera). Journal of Insect Science, 2016, 16(1), 67. https://doi.org/10.1093/jisesa/iew050. [CrossRef]
- Dobrzycka-Krahel, A., Rolbiecki, L., Karczewski, J., & Skóra, M. E. Variations in host surfaces morphology and biology of ciliate epibionts explaining distribution pattern of epibionts in the invasive signal crayfish Pacifastacus leniusculus (Dana, 1852). Journal of Zoology, 2022, 316 (4), 282-295. https://doi.org/10.1111/jzo.12953. [CrossRef]
- Abd El-Lateif, R. S. Torra, D. E., El Sherry, Y. M. Infestation of External Ciliated Protozoan in Red Swamp Crayfish (Procambarus clarkii). Journal of Advanced Veterinary Research, 2023, 13(2), 197-206.
- Corliss, J.O. Biodiversity, classification, and numbers of species of protists. Nature and human society. The quest for a sustainable world. Proceedings of the 1997 Forum on Biodiversity, Board on Biology, NRC. National Academy Press, Washington, 2000, 130-155.
- Zhang, T., Vďačný, P. Morphological and molecular characterization of the ciliate parasite Tetrahymena rostrata infecting the renal organ of the dusky slug (Arion fuscus). Canadian Journal of Zoology, 2022, 101 (1), 32-45. https://doi.org/10.1139/cjz-2022-0080. [CrossRef]
- Zhang, T., & Vďačný, P. A discovery of two new Tetrahymena species parasitizing slugs and mussels: morphology and multi-gene phylogeny of T. foissneri sp. n. and T. unionis sp. n. Parasitology Research, 2021, 120(7), 2595-2616. https://doi.org/10.1007/s00436-021-07152-5. [CrossRef]
- Khan, R.A. Disease outbreaks and mass mortality in cultured Atlantic cod, Gadus morhua L., associated with Trichodina murmanica (Ciliophora). Journal of Fish Diseases, 2004, 27(3), 181-184. https://doi.org/10.1111/j.1365-2761.2004.00525.x. [CrossRef]
- Madsen, H. C., Buchmann, K., Mellergaard, S. Trichodina sp. (Ciliophora: Peritrichida) in eel Anguilla anguilla in recirculation systems in Denmark: host-parasite relations. Diseases of aquatic organisms, 4, 2000, 2(2), 149-152. https://doi.org/10.3354/dao042149. [CrossRef]
- Abdel-Hafez, G., Lahnsteiner, F., Mansour, N., Licek, E. Pathophysiology of Ichthyophthirius multifiliis infection in rainbow trout (Oncorhynchus mykiss) and chub (Leuciscus cephalus). Journal of comparative pathology, 151(4), 394-399. https://doi.org/10.1016/j.jcpa.2014.08.003. [CrossRef]
- Gaze, W.H., Wootten, R. Ectoparasitic species of the genus Trichodina (Ciliophora: Peritrichida) parasiting British freshwater fish. Folia Parasitologica, 1998, 45(3), 177-190.
- Karatayev, A.Y., Molloy, D.P., Burlakova, L.E. Seasonal dynamics of Conchophthirus acuminatus (Ciliophora, Conchophthiridae) infection in Dreissena polymorpha and D. bugensis (Bivalvia, Dreissenidae). European Journal of Protistology, 2000, 36(4), 397-404, DOI : 10.1016/S0932-4739(00)80045-0. [CrossRef]
- Blazhekovikj-Dimovska, D., Stojanovski, S. Ectoparasitic species of the genus Trichodina (Ciliophora: Peritrichida) parasitizing Macedonian freshwater fish. Acta Biologica, 2020, 27, 11-20. https://doi.org/10.18276/ab.2020.27-02. [CrossRef]
- Kulaš, A., Gulin, V., Kepčija, R.M., Žutinić, P., Perić, M.S., Orlić, S., Kajan, K., Stoeck, T., Lentendu, G., Čanjevac, I., Martinić, I. Ciliates (Alveolata, Ciliophora) as bioindicators of environmental pressure: A karstic river case. Ecological Indicators, 2021, 124, 107430. https://doi.org/10.1016/j.ecolind.2021.107430. [CrossRef]
- Hajdukiewicz, H., Wyżga, B., Amirowicz, A., Oglęcki, P., Radecki-Pawlik, A., Zawiejska, J., Mikuś, P. Ecological state of a mountain river before and after a large flood: Implications for river status assessment. Science of the Total Environment, 2018, 610, 244-257. https://doi.org/10.1016/j.scitotenv.2017.07.162. [CrossRef]
Figure 1.
Mean number of reads at the species level for 9 samples from three rivers (C - Czarna Włoszczowska, H - Czarna Hańcza and S - San) in three seasons (05 - May, 07 - July, 09 - September); A - marker hypervariable V4 region of the 18S rRNA gene, B – marker hypervariable V9 region of the 18S rRNA gene.
Figure 1.
Mean number of reads at the species level for 9 samples from three rivers (C - Czarna Włoszczowska, H - Czarna Hańcza and S - San) in three seasons (05 - May, 07 - July, 09 - September); A - marker hypervariable V4 region of the 18S rRNA gene, B – marker hypervariable V9 region of the 18S rRNA gene.
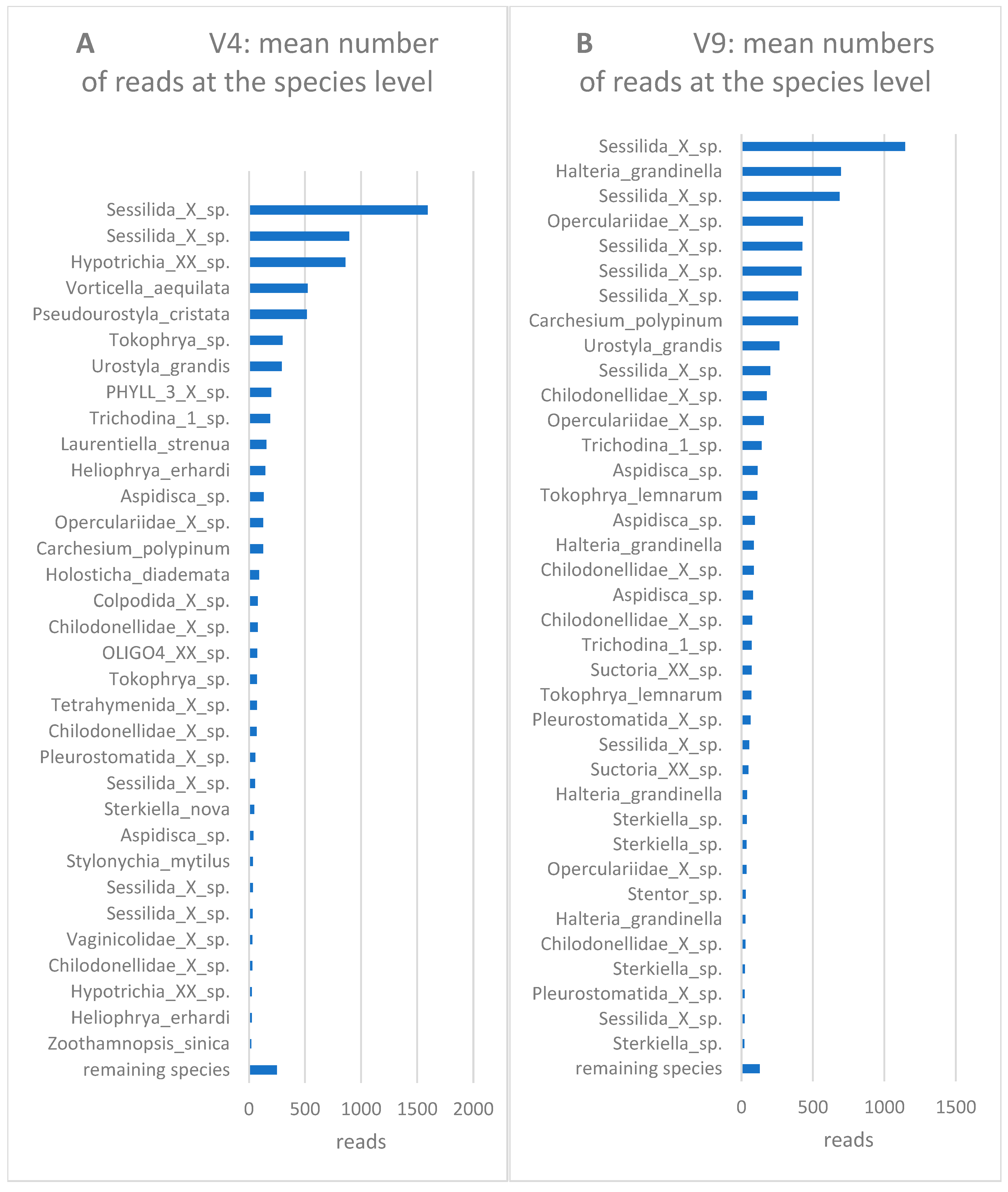
Figure 2.
Mean reads number per OTU (Operational Taxonomic Units) for samples from three rivers (C - Czarna Włoszczowska, H - Czarna Hańcza and S - San) in three seasons (05 - May, 07 - July, 09 September); A - marker hypervariable V4 region of the 18S rRNA gene, B – marker hypervariable V9 region of the 18S rRNA gene.
Figure 2.
Mean reads number per OTU (Operational Taxonomic Units) for samples from three rivers (C - Czarna Włoszczowska, H - Czarna Hańcza and S - San) in three seasons (05 - May, 07 - July, 09 September); A - marker hypervariable V4 region of the 18S rRNA gene, B – marker hypervariable V9 region of the 18S rRNA gene.
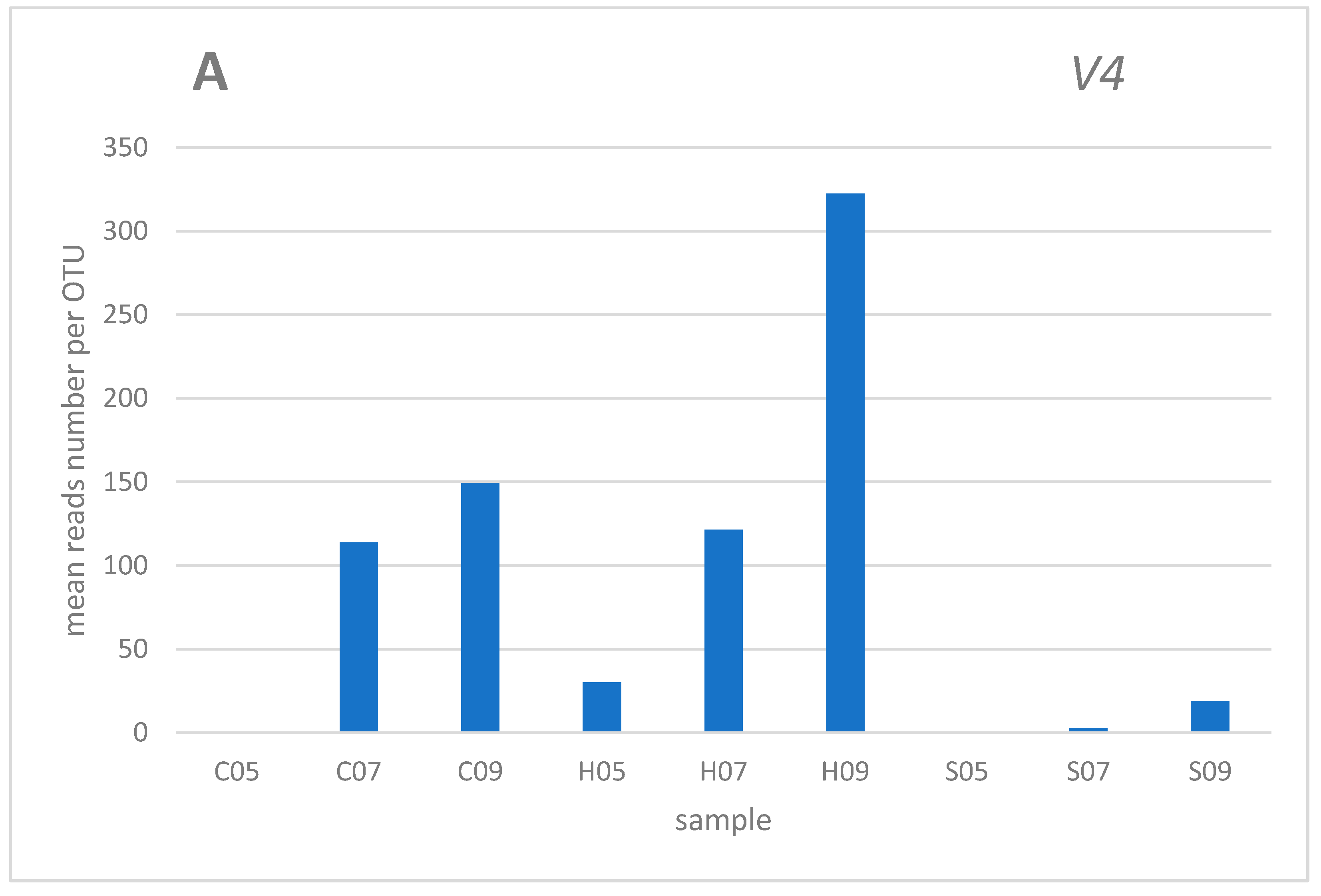
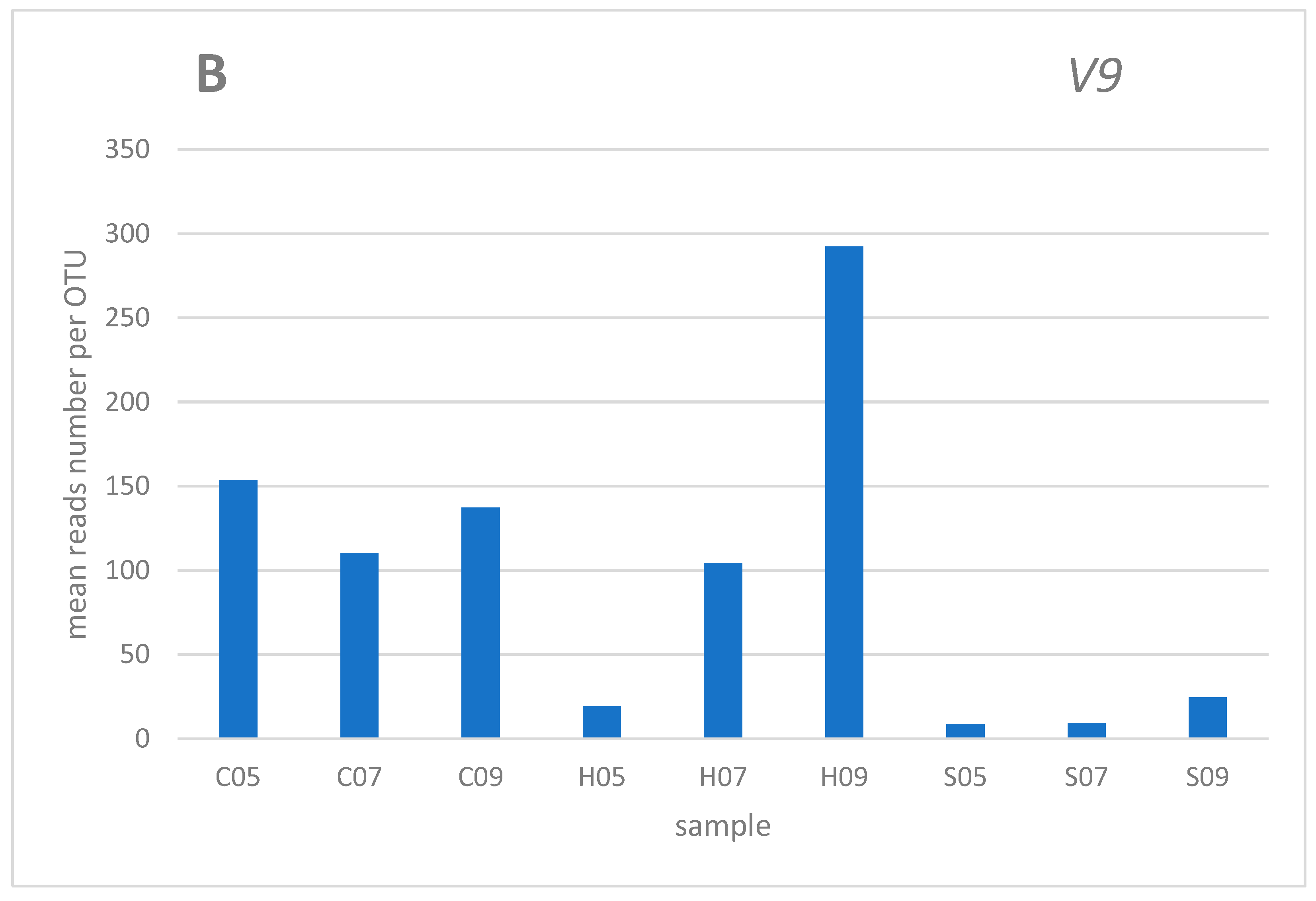
Figure 3.
Venn diagram performing the number of OTUs found in the rivers: marked blue in San, red in Czarna Hańcza, green in Czarna Włoszczowska, black numbers in appropriate place indicate the number of OTUs unique for one river or found in two or three rivers. Sampling in May, July and September; studied two markers V4 and V9.
Figure 3.
Venn diagram performing the number of OTUs found in the rivers: marked blue in San, red in Czarna Hańcza, green in Czarna Włoszczowska, black numbers in appropriate place indicate the number of OTUs unique for one river or found in two or three rivers. Sampling in May, July and September; studied two markers V4 and V9.
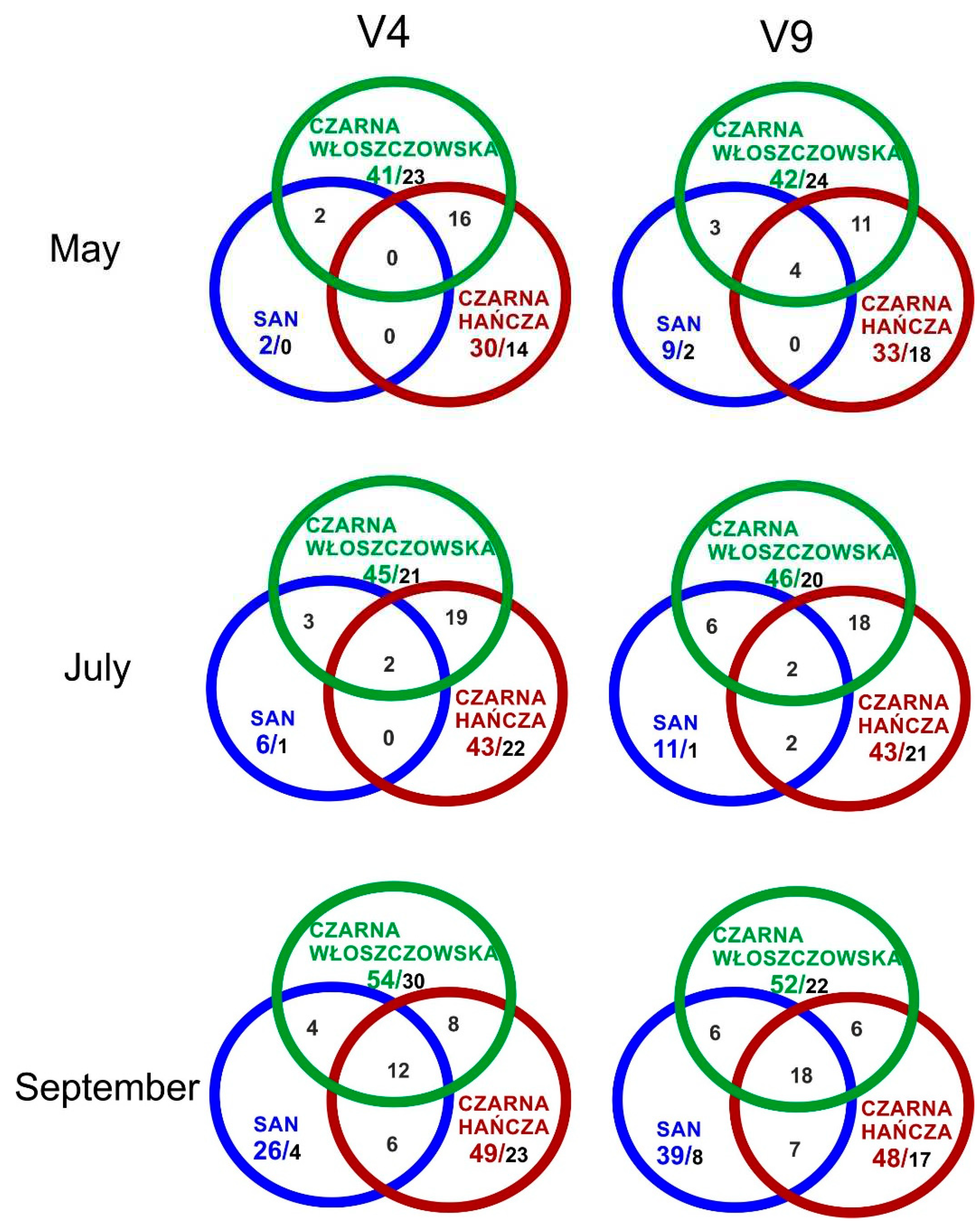

Table 1.
Sampling sites and dates.
| River name | River character | Catchment | GPS of the sampling sites | Dates of sampling |
|---|---|---|---|---|
| San | a dynamic mountainous river in the northern part of the Carpathian Mountains | Vistula | 49.2173 N 22.7174 E |
26.05.2022 |
| 11.07.2022 | ||||
| 13.09.2022 | ||||
| Czarna Włoszczowska | a lowland river flowing through an agricultural landscape | 50.9470 N 19.8464 E |
20.05.2022 | |
| 7.07.2022 22.09.2022 | ||||
| Czarna Hańcza | a pristine lowland river | Neman | 53.9709 N 23.3032 E |
26.05.2022 15.07.2022 14.09.2022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



