
Submitted:
25 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
The species composition of vultures and their interactions on carcasses of various stages of decomposition is not well understood yet it potentially affects their food acquisition and survival. We collected data from six carcasses between June and December 2021 using camera traps that were set on carcasses undergoing various decomposition states in Sinamatella Camp of Hwange National Park, Zimbabwe. Of interest were the cases in which each vulture species was dominating in interactions which gave it an advantage in terms of food acquisition. Four vulture species were observed (White-backed, White-headed, Lappet-faced and Hooded Vultures). Vulture abundances were greatest on fresh carcasses and least on dry ones. Although dominance behaviors by the White-backed and White-headed Vultures were recorded over all other vulture species, there were no records of the Lappet-faced dominating other vultures. In addition, the Hooded Vultures were mostly dominating non-vulture avian species on advanced decay carcasses. Our results demonstrate how various species may be prone to intense competition that may further place them at disadvantageous positions if food sources decline, more so under climatic shifts and various anthropogenic pressures.
Keywords:
Vultures
; decomposition state
; Camera traps
; dominance
1. Introduction
The importance of vultures has always been recognized ecologically and culturally [1,2,3]. Vultures are keystone species and they are important indicators of terrestrial biodiversity and the environmental health in different landscapes [4,5,6]. The range of African vultures have declined rapidly over the past 30 years. Of the eight species of Africa’s vultures, seven of these species populations have declined by over 80% [7,8] and have been listed as critically endangered by the International Union for the Conservation of Nature) [9]. The extent of vulture declines is complex and varies throughout the range. For example, in West Africa, some species may be extirpated especially in Nigeria, save for a few areas in Ghana and Niger where a substantial number of vultures persist [10]. Declines are also recorded in Kenya, Somalia, South Sudan and Sudan [7,11], although their status is apparently more stable in Uganda, Tanzania and parts of Southern Africa [12]. In Southern Africa, vulture declines have been estimated at 90% (range 75–95%) over the last three decades [7]. Many countries have therefore set species action plans to mitigate pressures facing the vultures. In this regard, studies that focus on the feeding ecology of vultures have also become very important across the African range.
Most African vulture species assemble at dead animals and they play different critical roles on the carcass while feeding. Resource partitioning by vultures may be influenced by many factors, both behavioral and extrinsic [13,14]. Aggressive encounters and interspecific competition have been minimized through various forms of partitioning. The differences in body size and beak morphology have been documented to facilitate the consumption of different body parts on large carcasses [15,16]. Body size also has a strong influence on dominance hierarchies [17] and thus resource partitioning in several guilds of vultures [18]. It is also been noted that the carcass decomposition state has a significant effect on the number and proportions of vultures feeding on it [19].
Spatio-temporal apportioning of resources by vultures has been assessed through studies of differences in habitat use and resource dispersion [20]. For example, the exploitation of carcasses by the Cape Vulture Gyps coprotheres and White-backed Vulture Gyps africanus were dependent on the bird’s abilities to take flight in different vegetation structure types in southern Africa [21]. In other cases outside the African continent, social dominance factors were also found to be critical when the migrant and resident Turkey Vultures Cathartes aura utilized various habitat types and foraging strategies [22]. Resource distribution, whether aggregated or scattered, appears to be important in determining the community composition of scavengers at ungulate carcasses in temperate-zone ecosystems [14]. Competition with predators at carcasses may also influence the scavengers’ community structure at carcasses killed by mammalian predators [23]. There are however fewer studies that explore how the vulture interactions may affect the food acquisition in African vultures.
This study seeks to investigate the vulture composition and interactions on carcasses found in Sinamatella, Hwange National Park, Zimbabwe. We hypothesized that 1) vulture composition while feeding on dead animals is influenced by the carcass species and stage of decomposition and 2) the large bodied vulture species tend to dominate in the inter-species interactions towards food acquisition. This work is important as it provides information on vulture composition in a typical protected area that exists in a mosaic of other land uses such as mining and human settlements.
2. Materials and Methods
The study was carried out in Sinamatella section of Hwange National Park (HNP which is 14 651km2 in size and is the largest National Park in Zimbabwe). HNP is located between latitudes 19°7'26.59"S and longitude 26°35'33"E and has an average altitude of 1000m above sea level. Sinamatella section is managed as an Intensive Protection Zone (IPZ) and it covers an area of 1 328 km2 (as illustrated in Figure 1).
We collected data from animal carcasses that were opportunistically found and the site could be reached with reasonable effort (i.e. within 5km from an established road/fire guard). Camera traps (Bushnel 119447 C) were set (one per carcass site over daytime hours only), and programmed to take 2 minute videos after every 5 minutes from the day the carcass was discovered until vultures disappeared from it. The batteries used for these cameras could last for more than 10 days, which was sufficient to cover the whole period until vultures disappeared from the carcass. The camera trap method ensured minimal disturbances to predators/scavengers by humans. For the extraction of data from the camera traps, the Time-lapse software was used where a template of containing the date, time of the day, decomposition state of the carcass, scavenger species, numbers, interactions (both inter and intra-specific) and duration of interaction was designed. The videos from the camera traps were analyzed manually while capturing the information onto the Time Lapse designed data template. In addition to the information captured on Time Lapse, the carcass species, carcass GPS position, the initial carcass decomposition state and vegetation characteristics of the carcass site was also recorded. From the videos, scavenger interactions were recorded as grabbing meat, pecking, chasing away and stalking. The carcass decomposition state was recorded as fresh (defined as the time the animal dies up to the state before signs of bloating show up); active decay (the carcass was at the bloating stage/increasing in size); advanced decay (defined by the shrinking in size of the carcass after its rupture when body fluids flow out and flesh rotting) and dry (when most flesh has been consumed and there is no more water coming from out of the carcass). These classifications were a modification from Valverde, Espín [24]. The captured information from the Time Lapse template was exported as an excel file for analysis.
The vegetation type was categorized into three broad classes i.e. closed woodland (dense vegetation, with trees within 3m of each other), open woodland (sparse vegetation, with trees greater than 3m from each other) and riverine (carcasses on a river bed or within 50m river/stream banks) [25]. The time spent by the scavenger species being engaged in particular interactions was expressed in seconds. A new variable “offender” was derived in which the individual that was dominating/winning a foraging opportunity in an interaction was noted. Another variable “victim” was created in which the individual that was being subdued/losing a feeding opportunity in an interaction was noted. In the cases of intraspecific interactions, the offender and the victim was the same species. A table was used to illustrate the composition of scavengers per carcass under a given decomposition state. Beyond the description of the scavenger community on the carcass, further analysis excluded non-vulture species as our aim was focused on vultures. To separate the observations on a carcass, a new variable “carcass state ID” was developed which was unique for each carcass, at a particular location and during a given decomposition state. The maximum number of vultures recorded per carcass state ID was extracted. The package “vegan” was used to calculate the Shannon diversity index of the vulture community per carcass state ID [26].
The relationships between the response variables (Shannon diversity and vulture abundance) and the dependent variables (vegetation type and decomposition state) were explored using generalized linear models (glms) since the response variables failed to conform to normality assumptions even after transformations attempts. The package ggplot2 were used to illustrate the results.
Subsets of cases under which each vulture species was the offender (i.e. it was benefiting from the interaction in terms food acquisition) were created. In these subsets, the duration of interaction time also failed to meet normality assumptions. Accordingly, glms were performed (one for each vulture species observed) to test if the duration of the offenders` interaction time was affected by the vegetation type, carcass species and the interaction between the victim and the carcass decomposition state. All data sorting and analysis were performed in the R Language and Environment for Statistical Computing [27].
3. Results
3.1. Carcass descriptions and scavenger composition
Six carcasses comprising of the African elephants Loxodonta africana (n=4), kudu Tragelaphus strepsiceros (n=1) and giraffe Giraffa camelopardis (n=1) were monitored. The giraffe carcass was monitored across two decomposition stages (i.e active and advanced decay) and two of the elephant carcasses were monitored across two decomposition stages (i.e one was monitored in the fresh and advanced decay stages while the other was monitored across the advanced decay and dry state) as summarized in Table 1.
Four vulture species were observed i.e. White-backed Vultures Gyps africanus, Lappet-faced Vultures Torgos tracheliotus, Hooded Vultures Necrosyrtes monachus, White-headed Vultures Trigonoceps occipitalis. In addition to vulture species, the following scavenger species were also observed during the study: Yellow-billed Kite Milvus aegyptius, Marabou Stork Leptoptilos crumenifer, Pied Crow Corvus albus and spotted hyenas Crocuta crocuta. The proportions of all the scavenging communities per carcass during each decomposition stage is shown in Figure 2. The Lappet-faced Vultures and White-backed Vultures were present at all carcasses and at all decomposition states.
The proportions of hyenas and marabou storks were greatest in the advanced stages of the giraffe carcass in riverine, and on the kudu carcass in open woodlands. The diversity of vulture species were not significantly different across vegetation types and decomposition states (p > 0.05 in both cases). However, when considering vulture abundance, there was a significant interaction between vegetation type and decomposition states (Wald Chisq = 7.746; df = 2, p = 0.021). In closed woodlands, vulture abundances were significantly higher on fresh carcasses when compared to the active decay stage while in open woodland, there were more vultures on the advanced decay carcasses when compared to the dry states (Table 2, Figure 3).
3.2. Vulture interactions
Five types of interaction were observed i.e. stalking, pecking, chasing away, running away and grabbing from each other. Dominance by the White-backed Vultures were recorded across all vulture species. The only cases in which the Lappet-faced Vultures were captured dominating other birds were on intraspecific interactions. The frequencies of these interactions are summarized in Table 3.
3.2.1. Cases of the Hooded Vulture dominating interactions
The Hooded Vultures were observed winning in intraspecific interactions and against other non-vulture avian species. During the intraspecific interactions, the time taken to interact were significantly longer in fresh carcasses when compared to the dry carcasses (β= 3.072, SE= 0.053, p<0.001), and the Hooded Vultures also spent significantly longer time dominating over non-vulture avian on advanced decaying carcasses when compared to the active stages (β=3.304, SE= 0.127, p = 0.011) as shown in Figure 4 and Appendix A.
3.2.2. Cases of White-headed Vultures dominating interactions
The White-headed Vultures were observed dominating all vulture species except the White-backed Vultures. Their dominant behavior was recorded for significantly longer periods in the closed woodlands when compared to open woodlands and riverine vegetation (p < 0.0001 in both cases). Similarly, the White-headed Vulture’s dominant behaviors were recorded for significantly longer periods on the kudu carcass when compared to the African elephant ones (β = 0.285, SE = 0.084, p < 0.0001). There was a significant interaction of the victim and decomposition state (Wald Chi Square = 837.44, df = 7, p < 0.0001), with significantly more time being spent in dominant activities over non-vulture avian species on carcasses undergoing active decay, and least on dry ones (Figure 5). Also, when the White-headed Vultures were interacting with the Lappet-faced Vultures, they spent significantly more time on the fresh carcass states when compared to the dry and advanced decay states (p < 0.0001 in both cases) as shown in Appendix A.
3.2.3. Cases of White-backed Vultures dominating interactions
The White-backed Vultures spent significantly more time when they were dominating over White-headed Vultures on actively decaying carcasses when compared to fresh and advanced decaying ones (Wald Chisq = 987.2, df = 9, p < 0.0001), as illustrated in Figure 6. Also, when they interacted with non-vulture species, significantly longer time was taken during the active decay when compared to the advanced decay stages. The dominant interaction time lags were significantly shorter when the White-backed Vultures were feeding on carcasses in open woodlands when compared to the closed woodlands and riverine habitats (Appendix A). The greatest time recorded in intraspecific interactions were on fresh carcasses when compared to the rest.
3.2.4. Cases of Lappet-faced Vultures dominating interactions
The Lappet-faced Vultures were never recorded dominating over other scavenging species, and the best model explaining the time spent in dominant behaviors only retained the vegetation type and the carcass species. Their dominating interactions (amongst themselves) took significantly more time in the closed woodlands when compared to the other vegetation classes (β= 3.664, SE = 0.012, p < 0.0001) as shown in Figure 7a. Also, interactions on dry carcasses were significantly shorter when compared to the fresh and actively decaying carcasses (Figure 7b and Appendix A).
4. Discussion
The carcass detection ability of most vulture species and the landscape of Sinamatella section of HNP allows some discussion of the observed scavenger composition and vulture interactions. Although it is acknowledged that the sample size was relatively low (for example, there was only one observed carcasses for the giraffe and kudu species), the known feeding patterns of vultures allow some analysis given the frequency of capturing of video clips over day time hours in this study (between run rise and sunset).
Vulture diversity and abundance on carcasses and decomposition states
Contrary to the hypothesis of this study that vulture diversity would be different across decomposition states and habitat types, the diversity of vulture species were not different across vegetation types and decomposition states. However, the early stages of carcass decomposition (fresh stage, active and advanced decay stages) had a greater abundance of vultures than the dry stages of decomposition. The study conducted by Mundy, Butchart [6] revealed that the White-backed and the Lappet-faced Vultures normally arrived early at carcasses and have the capacity to open up the carcass skin. Also, these two vulture species have been observed in high numbers at carcasses that still had flesh [4]. This study was probably unable to detect such differences as vultures were already gathered by the time the camera traps were set. Thus a better description would be given if carcasses could be monitored from the time the animal die, and this could be practical when sick/weak animals are followed or where an intact carcass can be experimentally set up. Such experimental set ups would need to be need to be varied across species with soft skins and those with hard ones [28].
Although this study had hypothesized that large bodied carcasses would host more vultures than small ones, this trend was not significant. What is clear however, is that the four vulture species that were recorded in this study seem to be congregating in similar proportions across carcass decomposition states. It therefore suggested that the amount of meat available on a carcass plays an important role in determining the vulture activities on a feeding patch in Sinamatella. To have a better understanding of the effect of carcass body sizes, this study may need to be extended so that small animals are also included. As the savannah system has a large guild of herbivores [29,30], smaller herbivores such as impalas could be experimentally set-up. In such set-ups, other mammalian scavengers may need to be excluded to prolong the longevity of the monitoring period on the carcass [31].
Diversity and abundance of vultures on differing habitat types
As hypothesized, the results from this study showed that there were more vultures in open woodlands than in closed woodlands and riverine habitats. This could be due to the fact that in open woodlands, carcasses were easily located than in the closed woodland and the riverine habitats. Hunter, Durant [32] confirmed that the White-backed Vultures located carrion by sight only and they rarely discovered carcasses concealed in trees or otherwise covered [33]. Indeed this study observed more White-backed Vultures and some marabou storks in carcasses in riverine habitats when compared to the open woodlands. It is however acknowledged that since this study was mostly conducted in the dry season, most trees had shed off their leaves and somehow the effect of vegetation may have been suppressed.
The diversity of vultures were however similar across the habitat types, which was contrary to our hypothesis. We believe that food scarcity in the system [34,35] and the general ability of vultures to discover carcasses [36] probably explain the insignificant results. As already discussed, the status of the trees during the dry season did not result in strongly different scenarios which would be detectable in terms of species diversity – more so for a highly specialized guild like vultures [37].
Vulture dominance and interactions on actively decaying carcasses
The White-backed Vultures were observed dominating the other three vulture species on actively decaying carcasses. The beaks of White-backed Vultures are medium-sized [38] and they are not adapted for tearing tough skin [39] (hence less dominating occasions on fresh carcasses). Thus they are known for eating soft tissues, such as the viscera, using a pulling feeding tactic [6]. The White-backed Vultures are the first to arrive on carcasses in relatively large numbers when compared to other vultures and thus their dominance could be partly due to their numerical advantage.
The Hooded Vultures were observed dominating only in intraspecific interactions and against other non-vulture avian species. This may be accounted to the small body size of this species, which may be linked to their inferiority when competing with the large vultures species like the White-headed and the Lappet-faced Vultures. The average weight of the Hooded Vultures range from 1.5-2.6kg [40] is lighter when compared to the White-backed Vultures 4.2-7.2kg [41], White-headed Vultures (female average weight of4.7kg) [42] and Lappet-faced Vultures (4.4-9.4kg) [43]. These species are unable to access much material on carcasses and instead, they feed primarily on scraps on the ground around carcasses [44]. The Hooded Vultures may be attracted to other vultures landing as a cue to carrion. They are also known for supplementing food from carrion by actively hunting. [4] also noted that Hooded Vulture have a more varied diet, using several food sources other than carrion, but when feeding at a carcass tend to peck on scraps.
The White-headed Vultures were observed dominating other vultures except the White-backed Vultures. Despite the large size of the White-headed Vultures, the numerical advantages of the White-backed Vultures probably play an important role in winning over the White-headed Vultures. Monadjem, Anderson [12] confirmed that White-headed Vultures assemble at carcasses in small numbers (as they are mostly solitary). Kemp, Kirwan [45] also confirmed that the White-headed Vulture is unique among African vultures because it does not rely on large carcasses, but often scouts for smaller carcasses with less competition. Also, Kemp, Kirwan [45] noted that White-headed Vultures do not rely on carrion for survival but rather they are known to hunt and kill their own prey such as stranded fish, mongoose, lizards, snakes, insects, piglets, and birds [46]. Future studies may also assess the contribution of rodents, fish, and lizards to the feeding ecology of vultures.
The Lappet-faced Vulture were never recorded dominating over other vulture species. This is in contracts to other observations, for example, Hockey, Dean [47], noted that they are “dominant over all other species at carcasses”. This may be explained by the fact that the other vulture species actually know the Lappet-faced Vultures are dominant and simply keep out of their way and thus there may be no physical dominance. Hockey, Dean [47] also reported that the Lappet-faced Vultures “may loaf for hours near carcass after feeding”. Thus if the individual Lappet-faced Vultures that were clear on the video footages were actively not feeding then the dominant behavior could have been missed.
5. Conclusions
This study has demonstrated that species such as the Lappet faced and Hooded Vultures are prone to intense competition that may further place them at a disadvantageous positions if food sources decline. With the current climate change projections, this may be the case especially if there is reduction in animal biomass associated with loss of biodiversity. In such scenarios, it would be important for conservation efforts to consider modified vulture restaurants that may offer preferential entrances that will allow all vulture species to access food. There may also be a need to negotiate for a system that the veterinarians certifies some carcasses as “safe for vultures” so that they do not get destroyed especially in communal areas or those surrounding protected areas. This should ideally be coupled with awareness campaigns so that people reduce vulture persecutions and poisoning.
Author Contributions
Conceptualization, T.T. and L.M.; methodology, L.M., S.L and T.M, software, L.M.; validation, L.M. and T.M; formal analysis, T.T. L.M and T.M; investigation, T.T.; resources, S.L.; data curation, L.M.; writing—original draft preparation, L.M.; writing—review and editing, T.T. and T.M; visualization, T.T.; supervision, T.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to the study being priorities by the Zimbabwe Parks and Wildlife Authority, the body which issues the permits to conduct studies in national Parks in Zimbabwe.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to Zimparks for hosting and supporting the first author during her intership period in Sinamatella and Bhejane Trust for providing a vehicle and a driver to the first author.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The combined model results for the four vulture species when they dominated interactions in Sinamatella.
| Model | Variable | Estimate | Std. Error | z value | Pr(>|z|) |
| Hooded Vultures | Intercept | 3.63 | 0.04 | 77.895 | <0.0001 |
| Open woodland | -0.695 | 0.034 | -20.304 | < 0.0001 | |
| Riverine | -0.526 | 0.025 | -20.793 | < 0.0001 | |
| Giraffe carcass | - | - | - | - | |
| Kudu carcass | 0.581 | 0.053 | 11.485 | <0.0001 | |
| Lappet-faced Vulture | -0.333 | 0.07 | -4.716 | <0.0001 | |
| Other non-vulture birds | -0.802 | 0.128 | -6.581 | <0.0001 | |
| Advanced decay state | -0.023 | 0.048 | -0.478 | 0.633 | |
| Dry state | -0.775 | 0.113 | -6.853 | <0.0001 | |
| Fresh state | -0.558 | 0.053 | -10.584 | <0.0001 | |
| Lappet-faced Vulture: Advanced decay stage | -0.227 | 0.075 | -3.027 | 0.003 | |
| Other non-vulture birds: Advanced decay stage | 0.326 | 0.127 | 2.556 | 0.011 | |
| Lappet-faced Vulture: Dry stage | - | - | - | - | |
| Other non-vulture birds: Dry stage | - | - | - | - | |
| Lappet-faced Vulture: Fresh stage | 0.624 | 0.077 | 8.06 | <0.0001 | |
| Other non-vulture birds: Fresh stage | - | - | - | - | |
| White-headed | Intercept | 3.219 | 0.071 | 45.522 | <0.0001 |
| Open woodland | -0.36 | 0.024 | -14.864 | <0.0001 | |
| Riverine habitat | -0.563 | 0.038 | -14.752 | <0.0001 | |
| Giraffe carcass | - | - | - | - | |
| Kudu carcass | 0.285 | 0.049 | 0.0584 | <0.0001 | |
| Lappet-faced Vulture | 0.7367 | 0.0744 | 0.099 | <0.0001 | |
| Other non-vulture birds | 0.47 | 0.077 | 6.05 | <0.0001 | |
| White-headed Vulture | -0.27 | 0.079 | -3.427 | 0.001 | |
| Advanced decay stage | -0.366 | 0.076 | -4.82 | <0.0001 | |
| Dry stage | -0.392 | 0.070 | -0.056 | <0.0001 | |
| Fresh stage | -0.269 | 0.076 | -3.549 | 0.001 | |
| Lappet-faced Vulture: Advanced decay stage | -0.441 | 0.081 | -0.054 | <0.0001 | |
| Other non-vulture birds: Advanced decay stage | -0.618 | 0.089 | -0.069 | <0.0001 | |
| White-headed Vulture: Advanced decay stage | 0.722 | 0.084 | 8.573 | <0.0001 | |
| Lappet-faced Vulture: Dry stage | -0.564 | 0.091 | -0.0617 | <0.0001 | |
| Other non-vulture birds: Dry stage | -0.857 | 0.191 | -0.045 | <0.0001 | |
| White-headed Vulture: Dry stage | - | - | - | - | |
| Lappet-faced Vulture: Fresh stage | -0.361 | 0.083 | -0.048 | <0.0001 | |
| Other non-vulture birds: Fresh stage | - | - | - | - | |
| White-headed: Fresh stage | 0.404 | 0.086 | 0.047 | <0.0001 | |
| White-backed | Intercept | 2.974 | 0.057 | 52.547 | <0.0001 |
| Open woodland | -0.283 | 0.020 | -14.358 | <0.0001 | |
| Riverine habitat | 0.043 | 0.017 | 2.504 | 0.012 | |
| Giraffe carcass | - | - | - | - | |
| Kudu carcass | 0.355 | 0.040 | 8.83 | <0.0001 | |
| Lappet-faced Vulture | 0.434 | 0.062 | 6.953 | <0.0001 | |
| Other birds | 0.100 | 0.073 | 1.356 | 0.175 | |
| White-backed Vulture | 0.827 | 0.059 | 13.953 | <0.0001 | |
| White-headed Vulture | 0.991 | 0.060 | 16.444 | <0.0001 | |
| Advanced decay stage | 0.232 | 0.058 | 3.97 | <0.0001 | |
| Dry stage | -1.603 | 0.128 | -12.502 | <0.0001 | |
| Fresh stage | 0.286 | 0.060 | 4.762 | <0.0001 | |
| Lappet-faced Vulture: Advanced decay stage | -0.611 | 0.067 | -9.184 | <0.0001 | |
| Other non-vulture birds: Advanced decay stage | -0.446 | 0.081 | -5.454 | <0.0001 | |
| White-backed Vulture: Advanced decay stage | -0.864 | 0.063 | -13.65 | <0.0001 | |
| White-headed Vulture: Advanced decay stage | -1.518 | 0.066 | -22.878 | <0.0001 | |
| Lappet-faced Vulture: Dry stage | 0.962 | 0.141 | 6.841 | <0.0001 | |
| other on-vulture birds: Dry stage | - | - | - | - | |
| White-backed Vulture: Dry stage | 0.328 | 0.142 | 2.31 | 0.021 | |
| White-headed Vulture: Dry stage | - | - | - | - | |
| Lappet-faced Vulture: Fresh stage | -0.275 | 0.068 | -4.02 | 0.001 | |
| other non-vulture birds: Fresh stage | - | - | - | - | |
| White-backed Vulture: Fresh stage | -0.511 | 0.065 | -7.877 | <0.0001 | |
| White-headed Vulture: Fresh stage | -0.762 | 0.067 | -11.448 | <0.0001 | |
| Lappet-faced | Intercept | 3.805 | 0.024 | 158.556 | <0.0001 |
| Open woodland | -0.875 | 0.028 | -31.211 | <0.0001 | |
| Riverine | -0.922 | 0.034 | -27.032 | <0.0001 | |
| Giraffe carcass | - | - | - | - | |
| Kudu carcass | 0.137 | 0.066 | 2.084 | 0.037 | |
| Advanced decay stage | 0.009 | 0.029 | 0.307 | 0.759 | |
| Dry stage | -0.256 | 0.053 | -4.868 | <0.0001 | |
| Fresh stage | -0.225 | 0.031 | -7.155 | <0.0001 |
References
- Hovardas, T. and K. Korfiatis, Towards a critical re-appraisal of ecology education: Scheduling an educational intervention to revisit the ‘balance of nature’metaphor. Science and Education, 2011. 20: p. 1039-1053. [CrossRef]
- McKean, S., M. Mander, N. Diederichs, L. Ntuli, K. Mavundla, V. Williams, and J. Wakelin, The impact of traditional use on vultures in South Africa. Vulture News, 2013. 65: p. 15-36. [CrossRef]
- Thompson, L. and A. Blackmore, A brief review of the legal protection of vultures in South Africa. Ostrich, 2020. 91(1): p. 1-12. [CrossRef]
- Kendall, C., M. Virani, P. Kirui, S. Thomsett, and M. Githiru, Mechanisms of coexistence in vultures: understanding the patterns of vulture abundance at carcasses in Masai Mara National Reserve, Kenya. The Condor, 2012. 114(3): p. 523-531. [CrossRef]
- Carrete, M. and J. Donázar, Application of central-place foraging theory shows the importance of Mediterranean dehesas for the conservation of the cinereous vulture, Aegypius monachus. Biological Conservation, 2005. 126(4): p. 582-590. [CrossRef]
- Mundy, P., D. Butchart, J. Ledger, and S. Piper, The vultures of Africa. Vol. 671. 1992: Academic Press London.
- Ogada, D., P. Shaw, R. Beyers, R. Buij, C. Murn, J. Thiollay, C. Beale, R. Holdo, D. Pomeroy, and N. Baker, Another continental vulture crisis: Africa's vultures collapsing toward extinction. Conservation Letters, 2016. 9(2): p. 89-97. [CrossRef]
- Virani, M., C. Kendall, P. Njoroge, and S. Thomsett, Major declines in the abundance of vultures and other scavenging raptors in and around the Masai Mara ecosystem, Kenya. Biological Conservation, 2011. 144(2): p. 746-752. [CrossRef]
- IUCN, S., IUCN SSC Guiding principles on Creating Proxies of Extinct Species, 2016, IUCN.
- Ogada, D., The power of poison: pesticide poisoning of Africa's wildlife. Annals of the New York Academy of Sciences, 2014. 1322(1): p. 1-20. [CrossRef]
- Ottinger, M., A. Botha, R. Buij, B. Coverdale, M. Gore, R. Harrell, J. Hassell, S. Krüger, C. McClure, and J. Mullinax, A strategy for conserving Old World vulture populations in the framework of One Health. Journal of Raptor Research, 2021. 55(3): p. 374-387. [CrossRef]
- Monadjem, A., M. Anderson, S. Piper, and A. Boshoff. The vultures of southern Africa-quo vadis. in Proceedings of a workshop on vulture research and conservation in southern Africa. Bird of Prey Working Group, Endangered Wildlife Trust, Johannesburg, South Africa. 2004.
- Selva, N. and M. Fortuna, The nested structure of a scavenger community. Proceedings of the Royal Society B: Biological Sciences, 2007. 274(1613): p. 1101-1108. [CrossRef]
- Blazquez, M., J. Sanchez-Zapata, F. Botella, M. Carrete, and S. Eguía, Spatio-temporal segregation of facultative avian scavengers at ungulate carcasses. Acta Oecologica, 2009. 35(5): p. 645-650. [CrossRef]
- Kruuk, H. and M. Turner, Comparative notes on predation by lion, leopard, cheetah and wild dogs in the Serengeti area, East Africa. Mammalia, 1967.
- Kendall, C., The early bird gets the carcass: temporal segregation and its effects on foraging success in avian scavengers. The Auk: Ornithological Advances, 2014. 131(1): p. 12-19. [CrossRef]
- Brown, J. and B. Maurer, Body size, ecological dominance and Cope's rule. Nature, 1986. 324(6094): p. 248-250. [CrossRef]
- Gomez, L., D. Houston, P. Cotton, and A. Tye, The role of Greater Yellow-headed Vultures Cathartes melambrotus as scavengers in neotropical forest. Ibis, 1994. 136(2): p. 193-196. [CrossRef]
- Hunter, J., S. Durant, and T. Caro, Patterns of scavenger arrival at cheetah kills in Serengeti National Park Tanzania. African Journal of Ecology, 2007. 45(3): p. 275-281. [CrossRef]
- Harel, R., O. Duriez, O. Spiegel, J. Fluhr, N. Horvitz, W. Getz, W. Bouten, F. Sarrazin, O. Hatzofe, and R. Nathan, Decision-making by a soaring bird: time, energy and risk considerations at different spatio-temporal scales. Philosophical Transactions of the Royal Society B: Biological Sciences, 2016. 371(1704): p. 20150397. [CrossRef]
- Bamford, A., A. Monadjem, M. Anderson, A. Anthony, W. Borello, M. Bridgeford, P. Bridgeford, P. Hancock, B. Howells, and J. Wakelin, Trade-offs between specificity and regional generality in habitat association models: a case study of two species of African vulture. Journal of Applied Ecology, 2009. 46(4): p. 852-860. [CrossRef]
- Platt, S. and T. Rainwater, Noteworthy observations of foraging turkey vultures. The Wilson Journal of Ornithology, 2009: p. 839-841. [CrossRef]
- Pereira, L., N. Owen-Smith, and M. Moleón, Facultative predation and scavenging by mammalian carnivores: Seasonal, regional and intra-guild comparisons. Mammal review, 2014. 44(1): p. 44-55. [CrossRef]
- Valverde, I., S. Espín, P. María-Mojica, and A. García-Fernández, Protocol to classify the stages of carcass decomposition and estimate the time of death in small-size raptors. European Journal of Wildlife Research, 2020. 66(6): p. 93. [CrossRef]
- Rogers, C., A woody vegetation survey of Hwange National Park, Zimbabwe. Department of National Parks and Wildlife Management report, 1993.
- Oksanen, J., F.G. Blanchet, R. Kindt, P. Legendre, P.R. Minchin, R. O’hara, G.L. Simpson, P. Solymos, M. Stevens, and H. Wagner, vegan: Community Ecology Package. R package version 2.0-10. 2022.
- R Development Core Team, R: A language and environment for statistical computing. Vienna, Austria; 2014, 2022.
- Miller, C., M. Pittman, X. Wang, X. Zheng, and J. Bright, Diet of Mesozoic toothed birds (Longipterygidae) inferred from quantitative analysis of extant avian diet proxies. BMC biology, 2022. 20(1): p. 1-37. [CrossRef]
- du Toit, J., Large herbivores and savanna heterogeneity. The Kruger experience: Ecology and management of savanna heterogeneity, 2003. 4(1): p. 292-309.
- Fritz, H., P. Duncan, I.J. Gordon, and A. Illius, Megaherbivores influence trophic guilds structure in African ungulate communities. Oecologia, 2002. 131: p. 620-625. [CrossRef]
- Hill, J., T. DeVault, J. Beasley, O. Rhodes Jr, and J. Belant, Effects of vulture exclusion on carrion consumption by facultative scavengers. Ecology and Evolution, 2018. 8(5): p. 2518-2526. [CrossRef]
- Hunter, J., D. Durant, and T. Caro, Patterns of scavenger arrival at cheetah kills in Serengeti National Park Tanzania. African Journal of Ecology, 2006: p. 1-7. [CrossRef]
- Houston, D., The role of griffon vultures Gyps africanus and Gyps ruppellii as scavengers. Journal of Zoology, 1974. 172(1): p. 35-46. [CrossRef]
- Nyathi, S., J. Olowoyo, and A. Oludare, Perception of scavengers and occupational health hazards associated with scavenging from a waste dumpsite in Pretoria, South Africa. Journal of environmental and public health, 2018. 2018. [CrossRef]
- Cortés-Avizanda, A., G. Blanco, T. DeVault, A. Markandya, M. Virani, J. Brandt, and J. Donázar, Supplementary feeding and endangered avian scavengers: benefits, caveats, and controversies. Frontiers in Ecology and the Environment, 2016. 14(4): p. 191-199. [CrossRef]
- Kane, A. and C. Kendall, Understanding how mammalian scavengers use information from avian scavengers: cue from above. Journal of Animal Ecology, 2017: p. 837-846. [CrossRef]
- García Alfonso, M., Individual and environmental drivers of resource use in an endangered vulture: Integrating movement, spatial and social ecology. 2020.
- Stattersfield, A., D. Capper, G. Dutson, and T. Morrisey, Threatened birds of the world: the official source for birds on the IUCN Red List. Bird Life International.2000, Barcelona and Cambridge.
- Kasielke, S. and S. Keeper, White-backed Vulture, Gyps africanus. 2015, Berkeley: University of California Press.
- Ferguson-Lees, J. and D. Christie, Raptors of the world2001: Houghton Mifflin Harcourt.
- Duffey, E., Handbook of the birds of the world, vol. 3. Hoatzin to auks: Edited by Josep del Hoyo, Andrew Elliot and Jordi Sargatal. 1996. Lynx Editions, Barcelona. 824 pp. ISBN 84 87334 20 2. Price£ 110.00, 1998, Elsevier.
- Murn, C., Field identification of individual White-headed Vultures Trigonoceps occipitalis using plumage patterns-an information theoretic approach. Bird Study, 2012. 59(4): p. 515-521. [CrossRef]
- Shimelis, A., E. Sande, S. Evans, and P.J. Mundy, International Species Action Plan for the Lappet-faced Vulture, Torgos tracheliotus BirdLife International, 2005, BirdLife International Africa Partnership Secretariat.
- Petrides, G., Competition for food between five species of East African vultures. The Auk, 1959. 76(1): p. 104-106. [CrossRef]
- Kemp, A., G. Kirwan, D. Christie, and C. Sharpe, eds. White-headed Vulture (Trigonoceps occipitalis). In Birds of the World. 2020, Cornell Lab of Ornithology. [CrossRef]
- Portugal, S., C. Murn, and G. Martin, White-headed Vulture Trigonoceps occipitalis shows visual field characteristics of hunting raptors. Ibis, 2017. 159(2): p. 463-466. [CrossRef]
- Hockey, P., W. Dean, and P. Ryan. Roberts birds of southern Africa. 7th edn. Trustees of the John Voelcker Bird Book Fund: Cape Town. 2005.
Figure 1.
Map showing the location of Sinamatella in Hwange National Park, Zimbabwe.
Figure 2.
Pie charts showing the composition of vultures at different decomposition states in different vegetation types. Ele represent Elephant.
Figure 2.
Pie charts showing the composition of vultures at different decomposition states in different vegetation types. Ele represent Elephant.
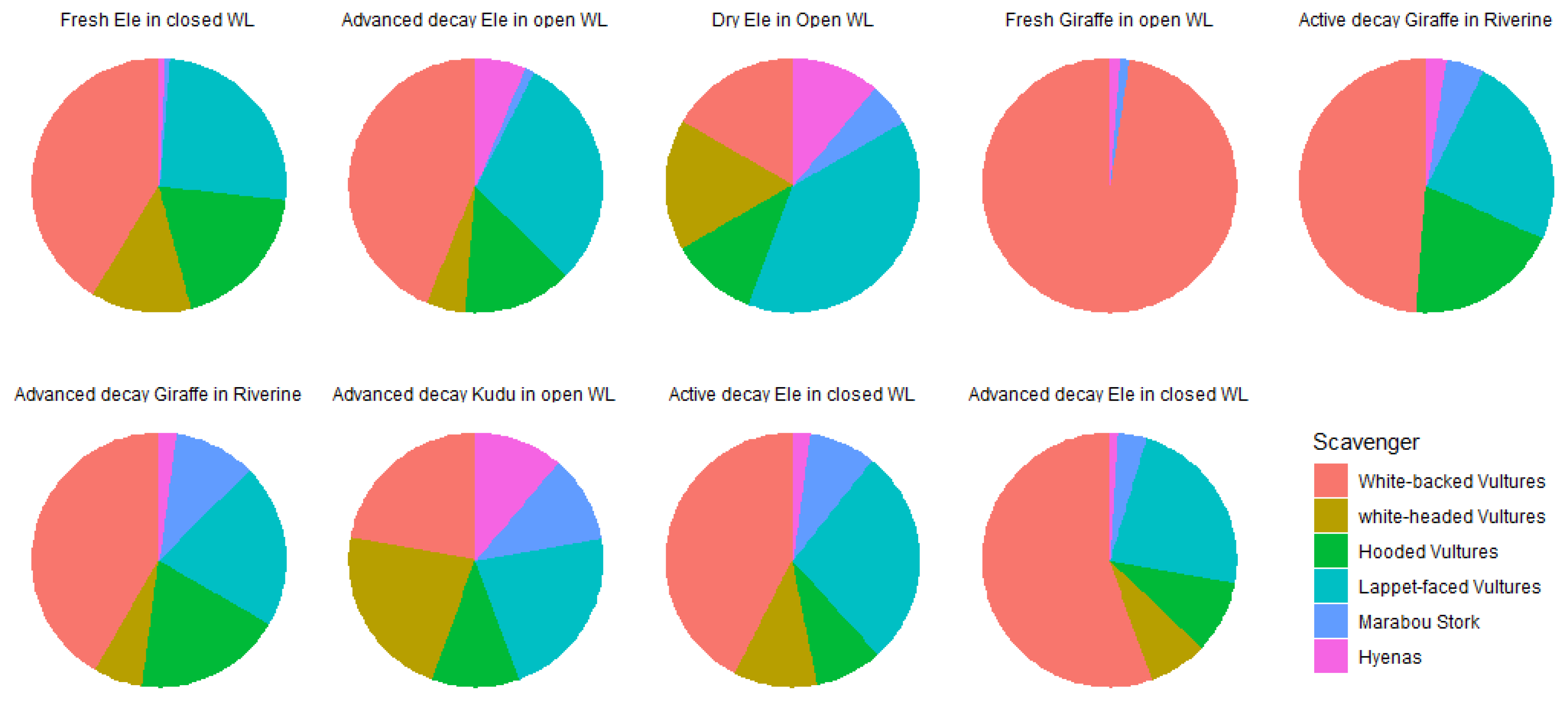
Figure 3.
Graph showing the influence of vegetation types and carcass decomposition state on vulture abundance.
Figure 3.
Graph showing the influence of vegetation types and carcass decomposition state on vulture abundance.
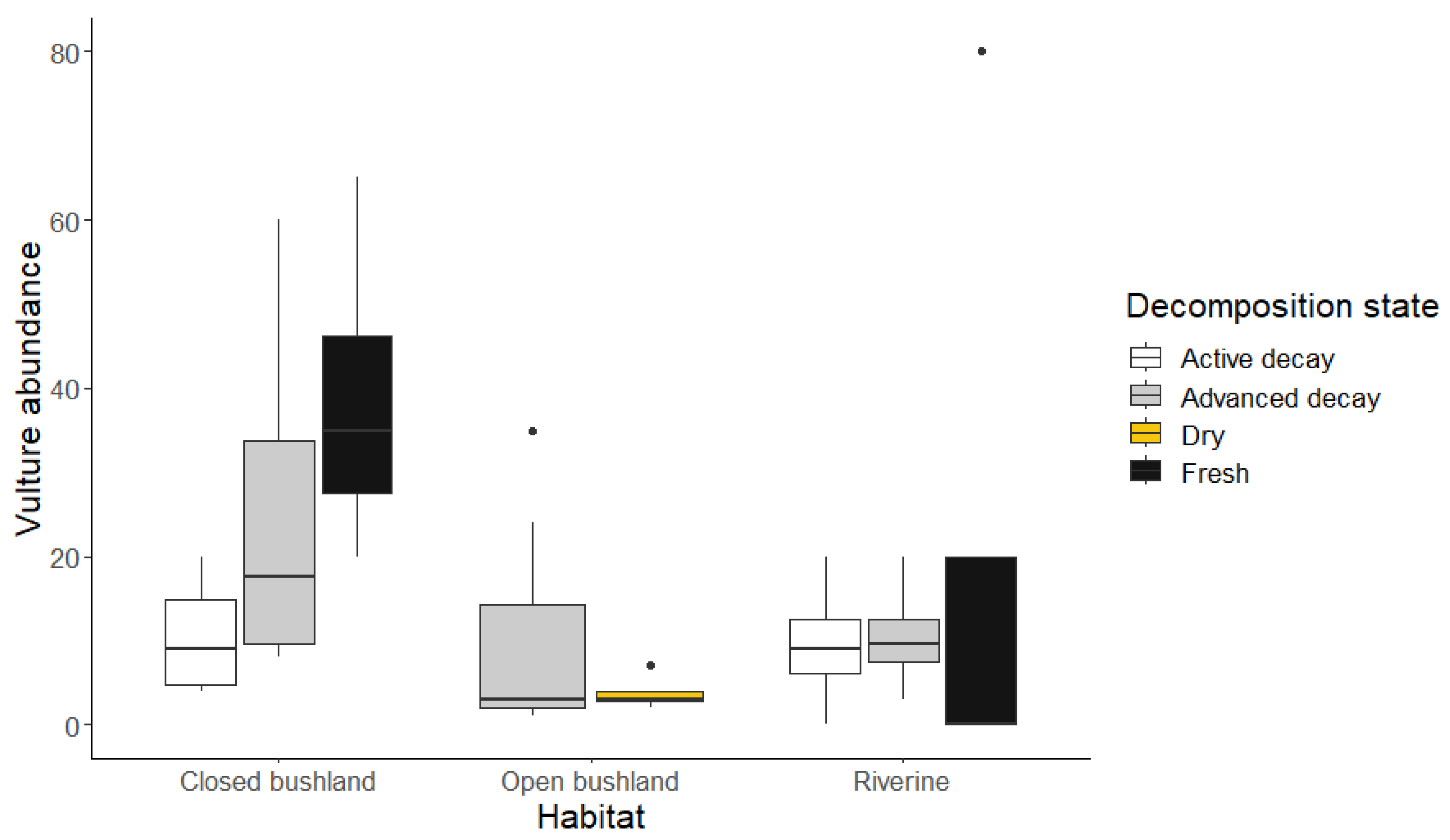
Figure 4.
Graph showing scenarios when the Hooded Vulture was dominating in the interactions with other vultures.
Figure 4.
Graph showing scenarios when the Hooded Vulture was dominating in the interactions with other vultures.
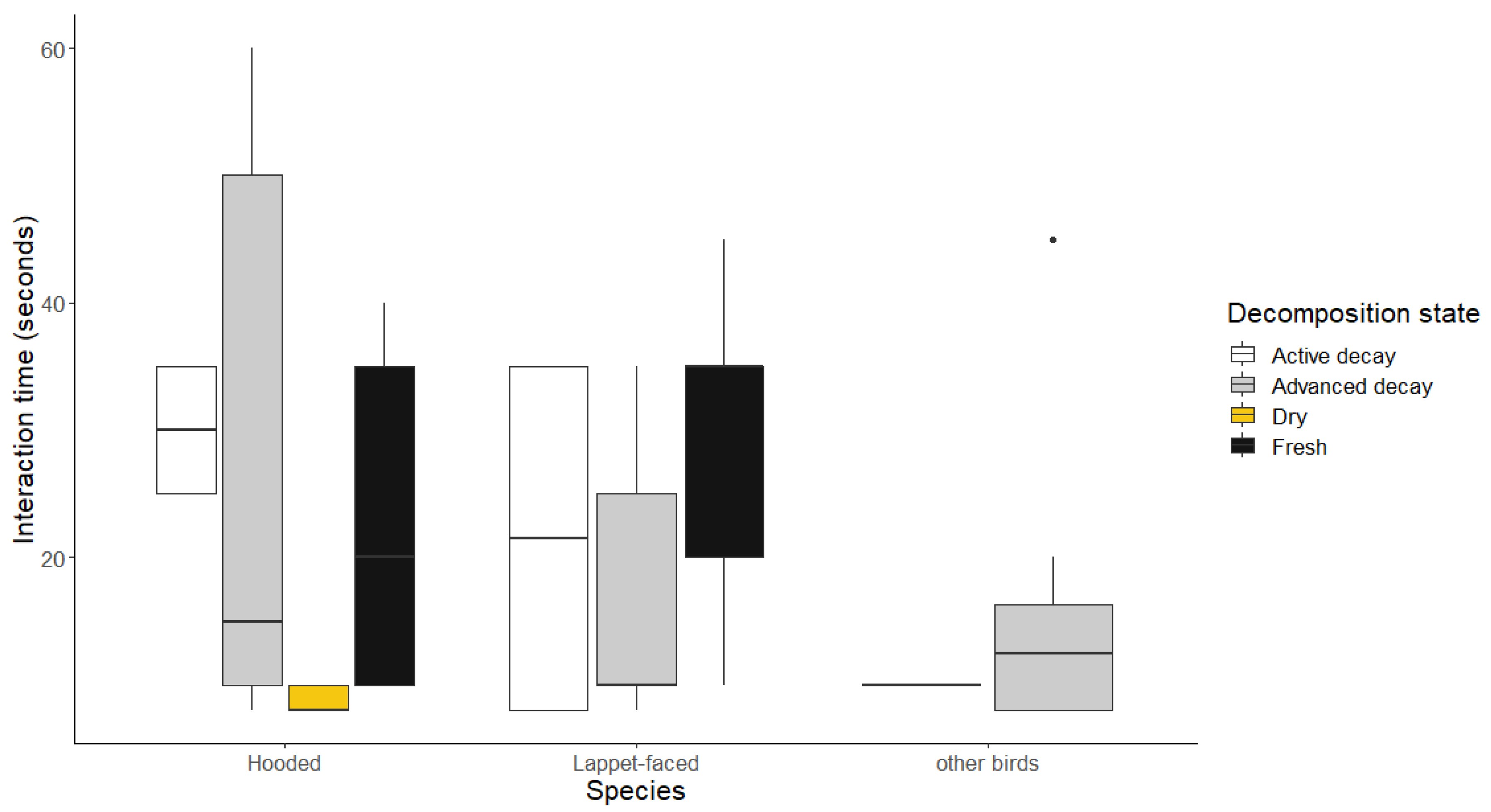
Figure 5.
Graph showing cases where the White-headed Vultures were recorded dominating other species.
Figure 5.
Graph showing cases where the White-headed Vultures were recorded dominating other species.
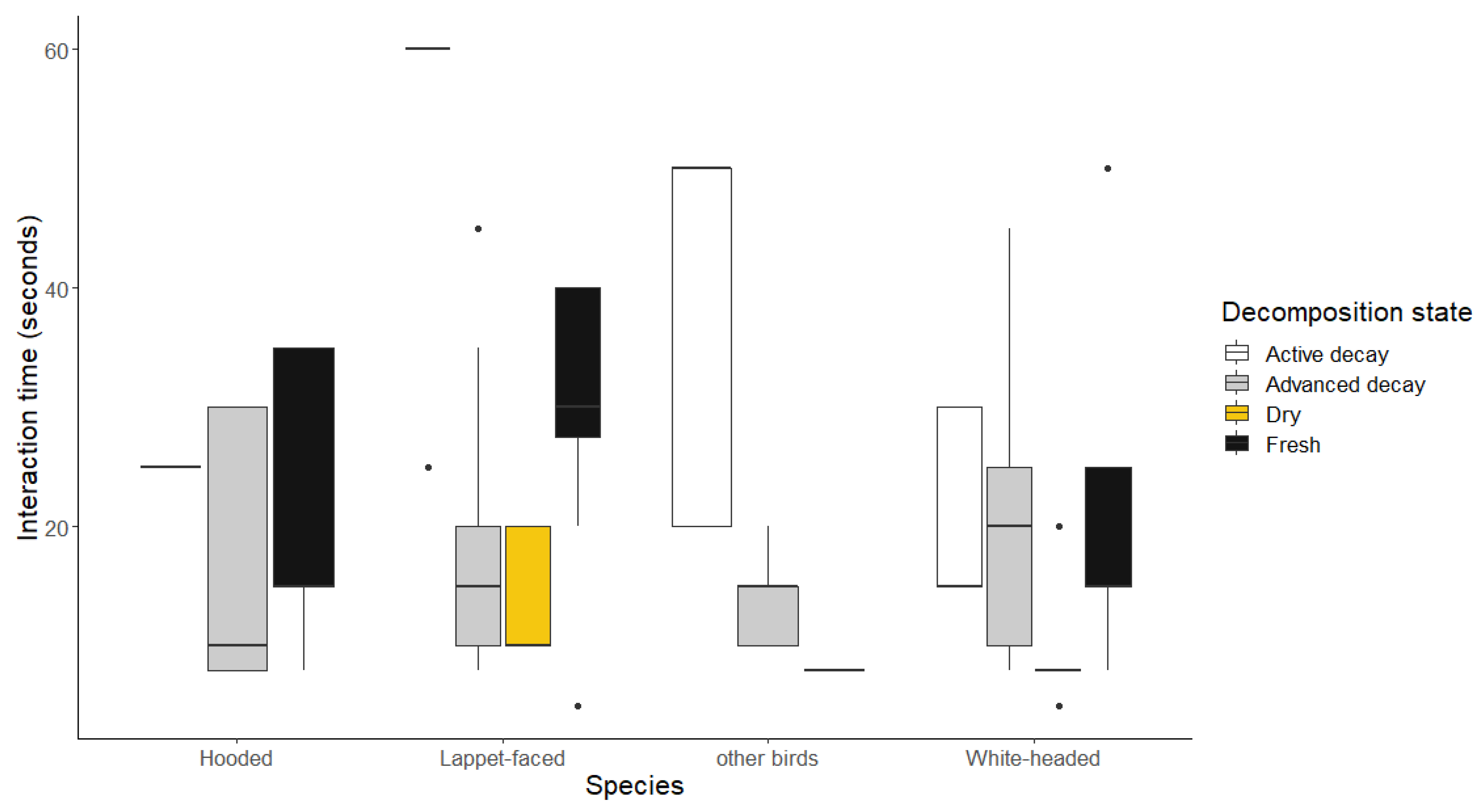
Figure 6.
Graph showing cases where the White-backed Vultures were recorded dominating over other species.
Figure 6.
Graph showing cases where the White-backed Vultures were recorded dominating over other species.
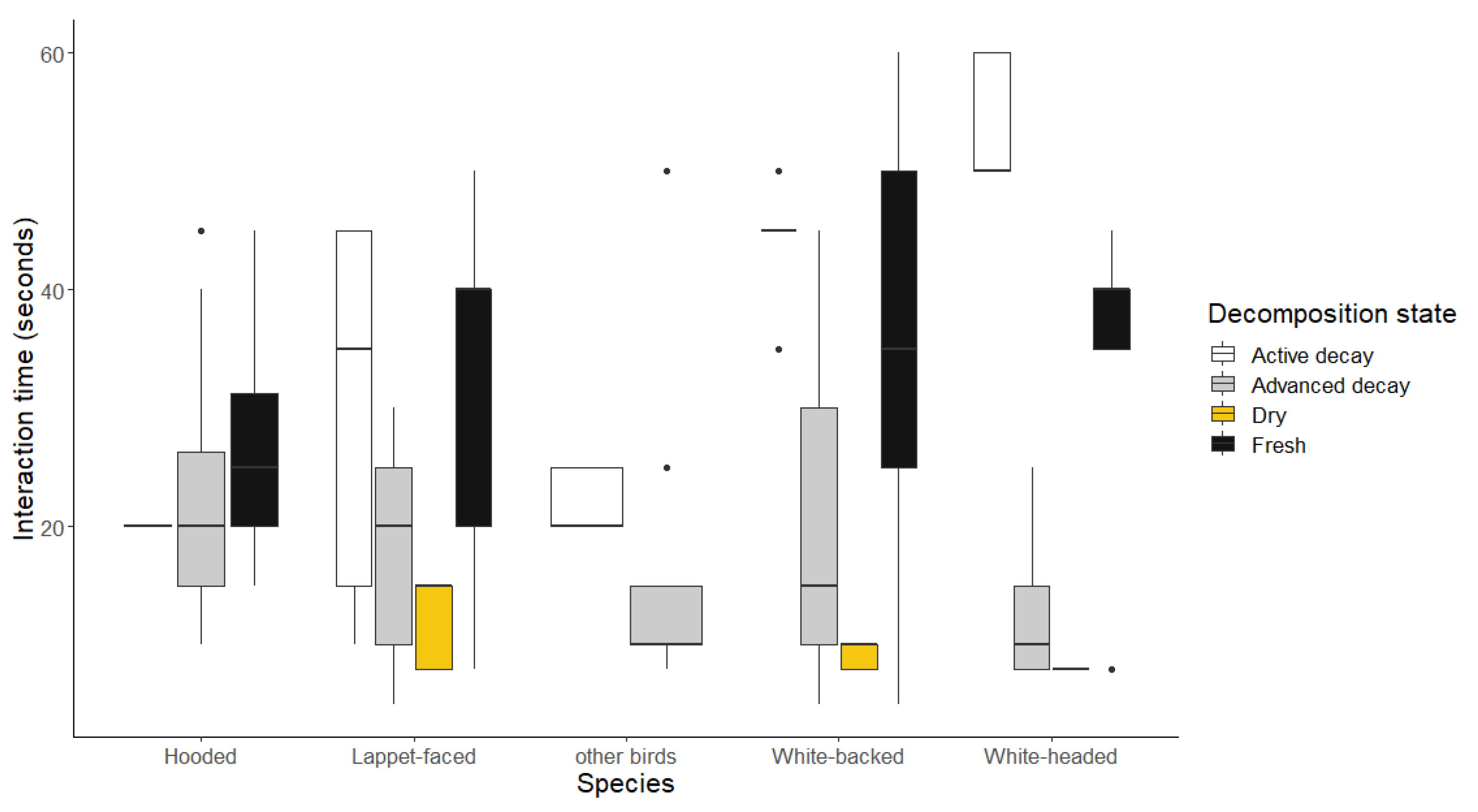
Figure 7.
Graphs showing the intraspecific dominance of Lappet-faced Vultures on (a) in different vegetation types and (b) across the carcass decomposition states.
Figure 7.
Graphs showing the intraspecific dominance of Lappet-faced Vultures on (a) in different vegetation types and (b) across the carcass decomposition states.
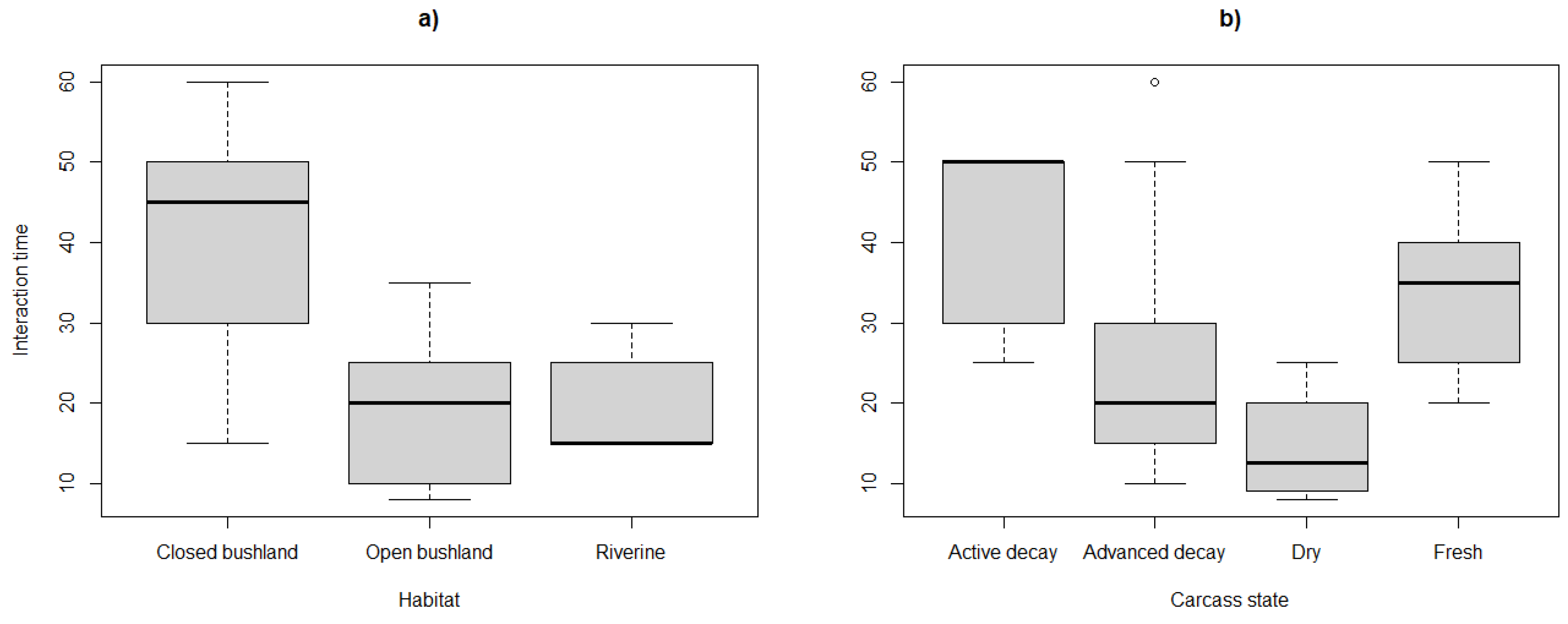

Table 1.
The Carcass species, allocated identities and vegetation types in which they were found. The decomposition states codes were defined as (i) = fresh, (ii) active decay, (iii) = advanced decay, and (iv) = dry.
Table 1.
The Carcass species, allocated identities and vegetation types in which they were found. The decomposition states codes were defined as (i) = fresh, (ii) active decay, (iii) = advanced decay, and (iv) = dry.
| Carcass ID | Carcass Species | Closed woodland | Open woodland | Riverine | Decomposition states monitored |
|---|---|---|---|---|---|
| 1 | Elephant | √ | i, iii | ||
| 2 | Elephant | √ | iii, iv | ||
| 3 | Giraffe | √ | i, ii, iii | ||
| 4 | Elephant | √ | Iii | ||
| 5 | Kudu | √ | Iii | ||
| 6 | Elephant | √ | Ii |
Table 2.
ble showing the outcome of the model of vulture abundances across vegetation types and decomposition states in Sinamatella, HNP.
Table 2.
ble showing the outcome of the model of vulture abundances across vegetation types and decomposition states in Sinamatella, HNP.
| Variable | Estimate | Std. Error | z value | p value |
|---|---|---|---|---|
| Intercept | 2.351 | 0.154 | 15.239 | < 0.0001 |
| Open woodland | -0.933 | 0.149 | -6.285 | < 0.0001 |
| Riverine | -0.1 | 0.224 | -0.447 | 0.65486 |
| Advanced decay | 0.897 | 0.183 | 4.9 | < 0.0001 |
| Dry state | -0.096 | 0.335 | -0.287 | 0.7743 |
| Fresh state | 1.306 | 0.174 | 7.506 | < 0.0001 |
| Open woodland:Advanced decay | - | - | - | - |
| Riverine:Advanced decay | -0.797 | 0.289 | -2.756 | 0.00586 |
| Open woodland:Dry | - | - | - | - |
| Riverine : Dry state | - | - | - | - |
| Open woodland : Fresh state | - | - | - | - |
| Riverine : Fresh state | -0.561 | 0.263 | -2.136 | 0.0327 |
Table 3.
Table showing the outcome of the interaction between different vulture species in Sinamatela camp, HNP, Zimbabwe.
Table 3.
Table showing the outcome of the interaction between different vulture species in Sinamatela camp, HNP, Zimbabwe.
| Vulture species | ||||||
|---|---|---|---|---|---|---|
| Hooded | Lappet-faced | White-backed | White-headed | Other birds | ||
| Vulture interaction | Hooded loosing | 16 | 16 | 0 | 0 | 0 |
| Hooded winning | 280 | 244 | 0 | 0 | 56 | |
| Lappet-faced loosing | 0 | 24 | 0 | 0 | 0 | |
| Lappet-faced winning | 0 | 404 | 0 | 0 | 0 | |
| White-backed loosing | 0 | 4 | 0 | 16 | 0 | |
| White-backed winning | 272 | 380 | 404 | 272 | 80 | |
| White-Headed winning | 176 | 260 | 0 | 372 | 88 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



