
Submitted:
26 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
In this study, the concentrations of metals (As, Cd, Co, Cr, Cu, Hg, Ni, Pb, and Zn) were deter-mined in two freshwater fish species. The same fish were also used to determine the content of fatty acids (FA), including saturated fatty acids (SFA), monounsaturated fatty acids (MUFA), and polyunsaturated fatty acids (PUFA). We investigated the species-specific relationship between bioaccumulation of metals and the content of FA. Roach and white bream were caught at two different sites in the Serbian part of the Danube. The measured metal concentrations were below the standard concentrations recommended in the guidelines of the EU and the Republic of Ser-bia. Among the non-essential metals, Ni had the highest concentration, ranging from 0.158 to 0.591 µg g-1. The muscle of the two fish species contained high concentrations of SFA, ranging from 36.31 to 51.79%, and MUFA, ranging from 33.56 to 43.70%. Polyunsaturated fatty acids were represented with the lowest percentage (5.32 - 27.18). Four of the nine metals studied (As, Hg, Ni and Pb) in one fish species and five of the nine metals studied (A, Cu, Ni, Pb and Zn) in the other fish species showed correlations with fatty acid content. These results help to reveal potential biomarkers of metal stress in lipid composition.
Keywords:
roach
; white bream
; lipids
; inorganic chemicals
; correlation matrix
1. Introduction
The rapid development of industry and agriculture has led to increased levels of inorganic (e.g., metals and trace elements) and organic chemicals in the aquatic environment. Discharge of wastewater enriched with these chemicals can lead to pollution of the aquatic environment and thus affect biological communities [1]. To protect freshwater biota, it is critical to identify the major pollution activities, their sources, and their impacts on the aquatic environment [2]. Understanding the effects of different types of anthropogenic pollution on these processes can help develop conservation programmes and strategies to prevent the disruption of freshwater environments [3].
Metals pose a particular threat to the environment due to their high potential for bioaccumulation and biomagnification in living organisms [4]. The accumulation and toxicity of heavy metals in fish is multidirectional and leads to physiological and chemical changes in the body of fish. Metals can stimulate excessive formation of reactive oxygen species (ROS) and disrupt the balance of oxidation-reduction reactions, resulting in structural damage to lipids, proteins, and DNA. The reactive oxygen species, whose main target is lipids, primarily PUFAs in membrane phospholipids, trigger lipid peroxidation (LPO) reactions. Xenobiotics attack cells and damage their structure and function, which can lead to apoptosis and necrosis. [5]. The presence of contaminants such as metals can greatly affect fatty acid composition [6,7,8,9]. Fat content and composition can vary greatly between different fish species and even within the same species when reared under different conditions [10]. The fatty acid composition of fish can be influenced by many environmental and biological factors, such as fish species, nutritional status, size, age, reproductive cycle, salinity, temperature, geographic location, and season. Reportedly, the amount of lipids (total fatty acids) in fish decreases during the cold season. The studies have also shown that the unsaturated fatty acid content varies significantly depending on the season, which may affect the nutritional properties and storage conditions of fish. In general, a decrease in temperature leads to an increase in the degree of unsaturation [11,12]. In particular, FA composition, degree of unsaturation, and changes in the relative abundance of individual fatty acids have been shown to respond to contamination levels both in experiments and in practise [13,14]. Because fish are at the top of the aquatic food chain and are used in human diets, they are important indicators for environmental biomonitoring [15]. Fish muscle is an important source of fatty acids (FA), especially long-chain polyunsaturated fatty acids (PUFAs) [1,16]. Among PUFAs, omega-3 FA are important because of their involvement in several biological processes and their nutritional significance [17].
The roach (Rutilus rutilus) is omnivorous and feeds on plant and animal matter. They feed opportunistically and eat a variety of small invertebrates such as insects, crustaceans, and mollusks. The diet of the roach is diverse and adaptable, allowing it to survive in a number of different water bodies, making this species very widespread. The white bream (Blicca bjoerkna) is a benthivorous fish that feeds on benthic invertebrates such as insect larvae, mollusks, crustaceans, and small fish. They also feed on aquatic vegetation such as algae, plant debris, and macrophyte seeds, especially during the warmer months of the year. Overall, the diet of white bream is highly variable and influenced by many factors such as season, location, and food availability [18].In Serbia, the lack of wastewater treatment and practices leads to pollution of water resources with heavy metals and other pollutants, which damages the aquatic ecosystem. Belgrade is the most important industrial area in Serbia, and the largest industrial capacities are located mainly on the banks of the Danube. The main problem related to pollution in Belgrade is the discharge of wastewater without prior treatment [19]. Recently, non-traditional biochemical biomarkers, namely fatty acid profiles of organisms, have been tested and proposed as reliable indicators of pollution levels [14, 20–22]. Changes in lipid metabolism and profiles of FA have been used to better understand how pollution affects key organisms in aquatic food webs and as an integrative biochemical response to contaminant exposure and accumulation in aquatic organisms [20] involving several studies with metals [23,24], making them promising biomarkers for contaminant exposure assessment. Lipid metabolism and fatty acid profiles have been studied to better understand how pollution affects key organisms in aquatic food chains. These studies help us understand how pollutants accumulate in aquatic organisms and how they respond to exposure. Previous studies with metals support the use of lipid metabolism and FA profiles as valuable biomarkers for assessing contaminant exposure [20,23,24]. Most of the studies deal exclusively with the accumulation of heavy metals or compositional analysis of FA [25,26,27,28]. In this context, the primary objective of this study was to determine the effects of bioaccumulation of untreated municipal wastewater on the FA content of roach and white bream. We examined the FA profile of fish in relation to metal pollution concentrations in two urban areas with different pollution loads.
2. Materials and Methods
2.1. Study site and sample collection
A total of 16 specimens of roach and bream were collected by commercial fishermen at each locality in April 2021 (8 individuals per species).Veliko Ratno Ostrvo (VRO) is located at the confluence of the Danube and Sava rivers It is separated from the urban environment of the city of Belgrade and is considered as a reference place. The locality of Višnjica (VIS) is exposed to the discharge of the largest sewage collector in the city of Belgrade and additionally suffers from anthropogenic pressure caused by the presence of a port for recreational boats (Figure 1). Fish total body length and total body weight were measured. Length and weight of the sampled fish were in the interval of 22-29 cm and 190-400 g, respectively (See supplementary table S2). For elemental and fatty acid analysis, samples of muscle tissue were collected from each specimen of both species. Samples were washed with distilled water and stored at -20 °C before analysis.
2.2. Elemental accumulation analysis
Analytical portions of approximately 0.5 g (wet weight) were accurately weighed and then processed in a microwave-digestion system. Samples were mineralized by adding 9 mL of 65% HNO3 and 1 mL of 30% H2O2 (Merck, Darmstadt, Germany). Microwave-assisted digestion was performed in ETHOS EASY Advanced Microwave Digestion System 230 V/50 Hz, Milestone, Italy. After cooling, digested samples were diluted with distilled water to a total volume of 25 mL. To assess the possible presence of trace elements in the reagents or carry-over effect of the digestion vessels, five reagent blanks were prepared during sample preparation, one per each session, according to the described procedure. These samples were analyzed in each analytical batch. All solutions were stored in polyethylene bottles until the trace elements (As, Hg, Cd, Co, Cu, Cr, Ni, Pb, and Zn) were measured using inductively coupled plasma optical emission spectrometry (ICP-OES), the Thermo Scientific™ iCAP™ 7400 ICP-OES. The single element mercury (Hg) calibration standard (1000 mg/ml) and the multi-element standard (100 mg/ml 21 components) purchased from CPAChem, Bulgaria, were used to prepare the calibration standards for the performed ICP-OES analysis. All measurements were performed in triplicate. Thermo Scientific Qtegra Intelligent Scientific Data Solution (ISDS) software was used for data acquisition and processing. The detection limits were 1 ppb for Ni, Cd, Cr, Co and Hg, 2 ppb for As and 30 ppb for Zn.
2.3. Fatty acid profile analysis
Samples of muscle tissue - about 25 g per sample were measured. Before homogenization, an appropriate amount of anhydrous Na2SO4 and PetrolEtra was added to each sample (10 g of anhydrous Na2SO4 and 15 ml of PetrolEtra are added to 10 g of tissue). Then the tissue was homogenized with a Politron, Kinematika (CH -6010 Kriens, Lucerne, Switzerland) for 5 min at speed 5. The supernatant was filtered through filter paper into a flask and evaporated to dryness on a vacuum evaporator from IKA, Germany (IKA RV 10) at 40 °C and 150 rpm.
Analysis of the fatty acid methyl ester profile (FAME) included transesterification by acid methanolysis and gas chromatography-mass spectrometry (GC-MS). The extracted lipids (30 mg) were dissolved in 6 mL of methanol with 2-3 drops of concentrated sulfuric acid. The mixture was refluxed at 80 °C for 2 hours and then the pH was adjusted to 7 with NaHCO3 solution (0.1 g/mL water). The FAMEs were collected using hexane (4 × 6 mL). The hexane layer was collected with a Pasteur pipette and dried with 15 g of anhydrous Na2SO4 for 15 min. The solution was filtered to remove the desiccant, and the solvent was removed in a rotary evaporator from IKA, Germany (IKA RV 10) at 40 °C with 150 rpm. The FAME extract was dissolved in hexane (5 mg/mL) and filtered through a Nachlon syringe filters 0.22 µm. Analysis was performed using a GC-MS KP2010 plus, equipped with an AOC 5000 injector (Shimadzu, Kyoto, Japan) and a FAME column (Phenomenex, L = 30 m, ID = 0.25 mm, df = 0.50 µm ), and using GCMS solution Ver. 2 software (Shimadzu). Samples (1 µL) were injected in split mode (1:30), with the injector temperature set at 250 °C. Mass spectra were obtained in EI mode (± 70 eV) in the m/z range 50–500 amu (SCAN) mode. Helium (99.999%) was used as carrier gas with a flow rate of 1.34 mL/min. The column was heated linearly from 100 °C (hold for 2 min) to 240 °C with a gradient of 3 °C/min and held at 240 °C for 5 min. The ion source temperature was set to 240 °C; interface temperature up to 260 °C. Constituents were identified by comparing their mass spectra with those of the NIST05, Wiley8, and FFNSC3 libraries using different browsers and a set of FAME standards in a Supelco® 37-component mixture FAME dissolved in hexane (1 mg/mL). Quantitative data were determined from the GC peak area using the area normalization method (results obtained are expressed as relative percentage).
2.4. Statistical Analysis
Initial assessment of the differences between groups was performed using the Kruskal-Wallis H test, which indicated differences in metal concentrations between localities (p < 0.05 was used as the threshold, STATISTICA 12.0). Differences in tissue heavy metal concentrations between different sites of the same species were estimated using a Mann-Whitney test (p < 0.05). To test the presence of metal and trace element accumulation with FA profile, the nonparametric Spearman correlation test (p < 0.05) was used to evaluate the relationship between the tissue metal concentrations and FA composition, using the Hmisc package [29] in R-Studio 1.4.1103.
3. Results and Discussion
3.1. Metal concentrations in two fish species
The concentration of nine metals in the muscle tissue of roach and white bream was studied. The average concentrations of the analyzed metals and trace elements (As, Cd, Co, Cr, Cu, Hg, Ni, Pb, and Zn) in the muscle tissue of the two studied species are shown in Table 1. In the total of 32 fish samples, the highest metal values were generally measured for Zn, and the lowest values were found for Co and Cd. Although the concentrations of Cd and Co were below detectable levels, their presence cannot be ruled out. Comparable consequences have been reported by many researchers in studies for different species [30,31,32]. In our study, the detected As, Cu, Hg, Pb and Zn concentrations were below the maximum allowable concentrations set by the EU [33] and the Republic of Serbia [34]. In roach, As, Pb and Zn differed significantly (p < 0.05) between localities (Table 1). Muscle had higher levels of As (p < 0.05), Cr, Cu, Hg, Pb (p < 0.05), and Zn (p < 0.05), except for Ni in roach from VIS compared to VRO. The opposite trend was observed for the levels of As, Cr, Hg, Pb, and Zn, but there were no significant differences between the levels of all metallic trace elements measured in white bream muscle at the two sites (Table 1).
Our results differ from previous analyses of metal concentrations in fish muscles at the same sampling sites as in this study [35˗37]. As for the VRO site, the Cr concentrations in fish muscles determined in this study are consistent with the results of Subotić et al. [35,36] (Table 1). The same authors reported much higher concentrations of As, Cu, and Hg in the muscle of common nase (Chondrostoma nasus), Vimba bream (Vimba vimba), sichel (Pelecus cultratus), and ruffe (Gymnocephalus cernua) compared to our results. Lead concentrations in all of the above fish species were below the detection limit, in contrast to Pb concentrations in roach and white bream, as also shown in Table 1. In addition, we detected Pb in the range of 0.023-0.029 µg g-1 (Table 1), while the Pb concentration in fish from VIS was below the detection limit in the study by Kostić et al. [37]. The Zn concentration in this study has similar mean values to those of Subotić et al. [36]. Nickel enrichment in this study is similar to that measured in sichel, but much higher than that in ruffe [35]. On the other hand, the concentrations of AS, Cr and Cu in the muscles of roach and white bream at the polluted site VIS were notably lower than in white bream sampled at the same site in 2014 [37].
3.2. Fatty acid composition
Twenty-six FAs consisting of 9 SFAs, 7 MUFAs, and 10 PUFAs were identified (Table 2). The species studied contained higher levels of SFAs than MUFAs and PUFAs at both sites, with the exception of roach from VRO, where the levels of SFAs and MUFAs were almost equal. Saturated fatty acids varied greatly among species, and only roach showed large differences among sites. Among SFAs, myristic acid (C14:0), palmitic acid (C16:0), and stearic acid (C18:0) dominated, accounting for an average of 90.4% and 94.8% of SFAs in roach and white bream, respectively. Monounsaturated fatty acid content varied spatially within each fish species (Table 2). The predominant MUFAs, which accounted for approximately 90.1% overall, were palmitoleic acid (C16:1) and oleic acid (C18:1) in roach and 98% in white bream (Table 2).
The percentage of PUFA was higher in both fishes in the reference locality. Among PUFA, n-6 acids dominated compared to n-3 acids, especially C18:2. It was found that alpha-linoleic acid (ALA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA) were present in significantly higher amounts in roach compared to white bream, regardless of the sampling location. In addition, the ratio of n-3 and n-6 fatty acids was calculated (Table 2).
Freshwater fish have higher levels of C18 PUFA and lower levels of n-3 EPA and DHA compared to marine fish. They are also known for their high content of n-6-PUFA, especially linoleic acid and arachidonic acid [38,39]. The content of fatty acids in muscle tissue of the two fishes studied was not significantly different using a Mann-Whitney test (p < 0.05), but differences cannot be excluded. It is already known that differences in FA profiles between species follow taxonomic differences, and these differences are thought to be due not only to dietary differences but also to metabolic differences [40].
Total SFA content in roach was 36.311% and 45.116% at the VRO and VIS sites, respectively, which is comparable to the values obtained in roach from Lake Gusinoe and the Olsztyn Lake District [24,41]. With respect to ƩMUFA, our results were 2.5 to 3 times higher, while for ƩPUFA an opposite trend was found than in the above studies. In bream from both localities studied, we found a much higher content of ƩSFA (Table 2), which contradicts the results of [24,41]. ƩMUFA and ƩPUFA in were not comparable with the other studies, because the value variations for these fatty acids are large.
3.3. Correlation of metals concentrations with fatty acids content
Spearman correlation coefficients between FA and metal concentrations for both species are shown in Figure 2.
Significant negative correlations were found between C13:0 content and As concentration in roach from the VRO area, indicating that an increase in the concentration of this metal led to a decrease in C13:0 content. A positive correlation was found between C15:1, C17:1, C20:2, and C22:6 levels and Hg concentration. A significant positive correlation was found between Ni concentration and C16:1 content, and a negative correlation was found with C20:0 content. Negative correlations were found between Hg concentration and C17:0, C17:1 and C18:0 content in the same types of VIS. Pb concentration showed a positive correlation with C18:1 (n-3) and negative correlations with C14:0, C15:0, C17:0, C18:1 (n-6) and C20:2 (Figure 2a). For the white bream from the VRO locality, Cu concentration showed negative correlations with C14:1, C15:0, C16:0, and C16:1 content and positive correlations with C18:2, C18:3, C20:2, C20:3, C20:4 (n-3), C20:4 (n-6), C20:5, and C22:6. Nickel enrichment correlated negatively with C20:1, while Zn concentration correlated negatively with C18:1. At the VIS site, Ni concentration correlated negatively with C13:0, C14:0, and C14:1. Lead concentration correlated positively with C18:1 and negatively with C20:1, while Zn concentration had a positive correlation with C16:1 (Figure 2b). All correlations reported in this section are statistically significant at p < 0.05.
In the work of Fonseca et al. [21], C14:0 and C14:1 in common goby (Pomatoschistus microps) showed no correlation with metal concentrations, which is in contrast to our results in roach and white bream. In the same study, most FA had negative correlations with the measured metals, while the correlation data between metals and specific FA in this study had an almost equal number of positive and negative correlations. The long-chain FA, i.e., omega-3 and omega-6 PUFA, are the two most important due to their multiple biological functions, such as reducing oxidative stress and protecting the cardiovascular system. The beneficial effects of consuming long-chain PUFA have been associated with positive effects on human health [42]
In the reviewed studies with fish, a change in the content of PUFAs, MUFAs and SFAs was observed after exposure to organic and inorganic pollutants [5,43,44]. A notable observation from the results obtained was that the higher ƩSFA values were recorded in both fish species caught at VIS. Higher ƩMUFA and Ʃ PUFA values were recorded in both fish species caught at the VRO site. The main reason for the changes in PUFAs is that they could be oxidized under oxidative stress conditions induced by metals or other compounds [45,46]. Polyunsaturated fatty acids are susceptible to oxidation because oxygen acts on double bonds to form lipoperoxides, which generate ROS and eventually induce oxidative stress [47]. Metal ions are directly or indirectly involved in lipid peroxidation reactions and enhanced free radical formation [20,48], metals such as Cd, Ni and Hg can induce the formation of ROS due to their oxidative potential, which can lead to lipid peroxidation damage [49,50]. Nickel, which has the highest bioaccumulation in roach and white bream (Table 1), can induce the formation of ROS by binding to macromolecules and inactivating protective antioxidant enzymes [51]. Fish are potentially vulnerable to peroxidative attack due to the large amounts of PUFAs in their tissues [52].
4. Conclusions
From the obtained results, it appears that the content of FA is influenced by various environmental stresses in the Belgrade urban area. The non-essential heavy metals in the muscles of the fish samples could be attributed to the anthropogenic metal sources affecting the aquatic habitats in the Belgrade urban area. The analysis of the concentration of 9 elements of the roach and white bream showed species-specific differences in their bioaccumulation. Although the results show that both fish species are safe from a nutritional and health point of view, and the metal concentrations are below the maximum levels set by the national legislation [34], it is recommended to conduct monitoring studies at regular intervals to investigate the metal concentrations, even if the levels are not above the acceptable norms. Although the measured metals are currently within safe limits, there is a possibility that they could increase and become toxic in the near future. Although our study did not reveal significant changes in FA levels in the fish, the correlations indicate the effects of bioaccumulation of metals on FA levels and affect homeostasis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.J. and V.Đ.; methodology and data collection I.S., M.D. and S.Ž.; data analysis K.J. and J.S.V.; writing—original draft preparation, J.S.V., K.J., and V.Đ.; writing—review and editing, J.S.V.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia, Contract Nos. 451-03-47/2023-01/200007, 451-03-47-/2023-01/200053 and 451-03-47/2023-01/200017.
Data Availability Statement
All data generated or analyzed during this study are included in this article [and its supplementary information file].
Conflicts of Interest
Conflicts of Interest: The authors declare no conflict of interest.
References
- Khoshnoud, M.J.; Mobini, K.; Javidnia, K.; Hosseinkhezri, P.; Aeen Jamshid, K. Heavy metals (Zn, Cu, Pb, Cd and Hg) contents and fatty acids ratios in two fish species (Scomberomorus commerson and Otolithes ruber) of the Persian Gulf. Iran. J. Pharm. Sci. 2011, 7, 191–196. [Google Scholar]
- Zhao, X.M.; Yao, L.A.; Ma, Q.L.; Zhou, G.J.; Wang, L.; Fang, Q.L.; Xu, Z.C. Distribution and ecological risk assessment of cadmium in water and sediment in Longjiang River, China: Implication on water quality management after pollution accident. Chemosphere 2018, 194, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Schmeller, D.S.; Loyau, A.; Bao, K.; Brack, W.; Chatzinotas, A.; De Vleeschouwer, F.; Friesen, J.; Gandois, L.; Hansson, S.V.; Haver, M.; Le Roux, G.; Shen, J.; Teisserenc, R.; Vredenburg, V.T. People, pollution and pathogens - Global change impacts in mountain freshwater ecosystems. Sci. Total Environ. 2018, 622-623, 756–763. [Google Scholar] [CrossRef]
- Erdoğrul, Ö.; Erbilir, F. Heavy metal and trace elements in various fish samples from Sir Dam Lake, Kahramanmaraş, Turkey. Environ. Monit. Assess. 2007, 130, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, A.; Fozia, F.; Ahmad, I.; Ullah, H. , Iqbal, A.; Ullah, R.; Moharram, B.A., Kowalczyk, A. Ecotoxicological assessment of heavy metal and its biochemical effect in fishes. Biomed. Res. Int. 2022, 3787838. [Google Scholar]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Senthamilselvan, D.; Chezhian, A.; Suresh, E. Synergistic effect of nickel and mercury on fatty acid composition in the muscle of fish Lates calcarifer. J. Fish Aquat. Sci. 2016, 11, 77–84. [Google Scholar]
- Strandberg, U.; Palviainen, M.; Eronen, A.; Piirainen, S.; Laurén, A.; Akkanen, J.; Kankaala, P. Spatial variability of mercury and polyunsaturated fatty acids in the European perch (Perca fluviatilis)–Implications for risk-benefit analyses of fish consumption. Environ. Pollut. 2016, 219, 305–314. [Google Scholar] [CrossRef]
- Li, D.; Hu, X. Fish and its multiple human health effects in times of threat to sustainability and affordability: are there alternatives? Asia Pac. J. Clin. Nutr. 2009, 218, 553–563. [Google Scholar]
- Ahmed, I.; Jan, K.; Fatma, S.; Dawood, M.A.O. Muscle proximate composition of various food fish species and their nutritional significance: A review. J. Anim. Physiol. Anim. Nutr. 2022, 106, 690–719. [Google Scholar] [CrossRef]
- Kaur, N.; Brraich, O.S. Impact of industrial effluents on physico-chemical parameters of water and fatty acid profile of fish, Labeo rohita (Hamilton), collected from the Ramsar sites of Punjab, India. Environ. Sci. Pollut. Res. 2022, 29, 11534–11552. [Google Scholar] [CrossRef] [PubMed]
- Kheiri, A.; Aliakbarlu, J.; Tahmasebi, R. Antioxidant potential and fatty acid profile of fish fillet: effects of season and fish species. Vet. Res. Forum. 2022, 13, 91–99. [Google Scholar]
- Duarte, B.; Carreiras, J.; Pérez-Romero, J.A.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Matos, A.R.; Marques, J.C.; Caçador, I. Halophyte fatty acids as biomarkers of anthropogenic-driven contamination in Mediterranean marshes: sentinel species survey and development of an integrated biomarker response (IBR) index. Ecol. Indic. 2018, 87, 86–96. [Google Scholar] [CrossRef]
- Silva, C.O.; Simões, T.; Novais, S.C.; Pimparel, I.; Granada, L.; Soares, A.M.V.M.; Barata, C.; Lemos, M.F.L. Fatty acid profile of the sea snail Gibbula umbilicalis as a biomarker for coastal metal pollution. Sci. Total Environ. 2017, 586, 542–550. [Google Scholar] [CrossRef]
- Meng, H.; Lin, Y.; Zhong, W.; Zhao, Z.; Shen, L.; Ling, Z.; Zhao, K.; Xu, S. Fish biomonitoring and ecological assessment in the Dianchi Lake basin based on environmental DNA. Water 2023, 15, 399. [Google Scholar] [CrossRef]
- Nordov, A.; Macholi, R.; Arnesen, H.; Videbaek, J. N-3 polyunsaturated fatty acids and cardiovascular diseases. Lipids 2001, 36, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Van Dael, P. Role of n-3 long-chain polyunsaturated fatty acids in human nutrition and health: review of recent studies and recommendations. Nutr. Res. Pract. 2021, 15, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Kottelat, M.; Freyhof, J. Handbook of European freshwater fishes. Publications Kottelat, Cornol and Freyhof, Berlin, 2007, pp 646.
- Milanović, A. , Kovačević-Majkić, J.; Milivojević, M. Water quality analysis of Danube River in Serbia: pollution and protection problems. Bull. Serb. Geogr. Soc. 2010, 90, 47–68. [Google Scholar]
- Filimonova, V.; Goncalves, F.; Marques, J.C.; De Troch, M.; Goncalves, A.M. Fatty acid profiling as bioindicator of chemical stress in marine organisms: a review. Ecol. Indic. 2016, 67, 657–672. [Google Scholar] [CrossRef]
- Fonseca, V.F.; Duarte, I.A.; Feijão, E.; Matos, A.R.; Duarte, B. Fatty acid-based index development in estuarine organisms to pinpoint environmental contamination. Mar. Pollut. Bull. 2022, 180, 113805. [Google Scholar] [CrossRef]
- Signa, G.; Di Leonardo, R.; Vaccaro, A.; Tramati, C.D.; Mazzola, A.; Vizzini, S. Lipid and fatty acid biomarkers as proxies for environmental contamination in caged mussels Mytilus galloprovincialis. Ecol. Indic. 2015, 57, 384–394. [Google Scholar] [CrossRef]
- Fokina, N.N.; Ruokolainen, T.R.; Nemova, N.N.; Bakhmet, I.N. Changes of blue mussels Mytilus edulis L. lipid composition under cadmium and copper toxic effect. Biol. Trace Elem. Res. 2013, 154, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Łuczyńska, J.; Paszczyk, B. Health risk assessment of heavy metals and lipid quality indexes in freshwater fish from lakes of Warmia and Mazury region, Poland. Int. J. Environ. Res. Public Health. 2019, 16, 3780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ning, X.; He, X.; Sun, X.; Yu, X.; Cheng, Y.; Yu, R.Q; Wu, Y. Fatty acid composition analyses of commercially important fish species from the Pearl River Estuary, China. PLoS One. 2020, 15, e0228276. [Google Scholar] [CrossRef]
- Linhartová, Z.; Krejsa, J.; Zajíc, T.; Másílko, J.; Sampels, S.; Mráz, J. Proximate and fatty acid composition of 13 important freshwater fish species in central Europe. Aquacult. Int. 2018, 26, 695–711. [Google Scholar] [CrossRef]
- Jovičić, K.; Janković, S.; Nikolić, D.; Đikanović, V.; Skorić, S.; Krpo-Ćetković, J.; Jarić, I. Prospects of fish scale and fin samples usage for nonlethal monitoring of metal contamination: a study on five fish species from the Danube River. Knowl. Manag. Aquat. Ecosyst. 2023, 424, 4. [Google Scholar] [CrossRef]
- Nędzarek, A.; Formicki, K.; Kowalska-Góralska, M.; Dobrzański, Z. Concentration and risk of contamination with trace elements in acipenserid and salmonid roe. J. Food Compos. Anal. 2022, 110, 104525. [Google Scholar] [CrossRef]
- Harrell, F.E. Hmisc: a package of miscellaneous R functions. 2020, http://biostat.mc.vanderbilt.
- Tepe,Y. Metal concentrations in eight fish species from Aegean and Mediterranean Seas, Environ. Monit. Assess. 2009, 159, 501–509.
- Ali, M.M.; Ali, M.L.; Proshad, R. ; Islam,S; Rahman, Z; Kormoker, T. Assessment of trace elements in the demersal fishes of a Coastal River in Bangladesh: a public health concern Thalassas. Int. J. Mar. Sci 2020, 36, 641–655. [Google Scholar]
- Töre, Y.; Ustaoğlu, F. ; Tepe,Y., Kalipci,E. Levels of toxic metals in edible fish species of the Tigris River (Turkey); Threat to public health. Ecol. Indic. 2021, 123, 107361. [Google Scholar] [CrossRef]
- EU, 2006. Commission Regulation (EC) No. 1881/2006 of 19 december 2006 setting maximum levels for certain contaminants in foodstuffs (text with EEA relevance). Off. J. Eur. Union 364, 5 - 24. No. 1881/2006.
- Official Gazette of the Republic of Serbia Nos 22/2018 & 90/2018, 2018. Regulation on the Maximum Permitted Residue Levels of Pesticides in Food and Animal Feed and Feed and Animal Feed for Which Maximum Quantities of Residues of Pesticides Are Permitted.
- Subotić, S.; Višnjić-Jeftić, Ž.; Spasić, S.; Hegediš, A. ; Krpo-Ćetković, J; Lenhardt, M. Concentrations of 18 Elements in Muscle, Liver, Gills, and Gonads of Sichel (Pelecus cultratus), Ruffe (Gymnocephalus cernua), and European Perch (Perca fluviatilis) in the Danube River near Belgrade (Serbia). Water Air Soil Pollut. 2015, 226, 287. [Google Scholar] [CrossRef]
- Subotić, S.; Višnjić-Jeftić, Ž.; Đikanović, V.; Spasić, S.; Krpo-Ćetković, J; Lenhardt, M. Metal Accumulation in Muscle and Liver of the Common Nase (Chondrostoma nasus) and Vimba Bream (Vimbavimba) from the Danube River, Serbia: Bioindicative Aspects. Bull. Environ. Contam. Toxicol. 2019, 103, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Kostić-Vuković, J.; Kolarević, S.; Kračun-Kolarević, M.; Višnjić-Jeftić, Ž.; Rašković, B.; Poleksić, V.; Gačić, Z.; Lenhardt, M.; Vuković-Gačić, B. Temporal variation of biomarkers in common bream Abramis brama (L., 1758) exposed to untreated municipal wastewater in the Danube River in Belgrade, Serbia. Environ. Monit. Assess. 2021, 193, 465. [Google Scholar] [CrossRef] [PubMed]
- Özogul, Y.; Özogul, F.; Alagoz, S. Fatty acid profiles and fat contents of commercially important seawater and freshwater fish species of Turkey: A comparative study. Food Chem. 2007, 103, 217–223. [Google Scholar] [CrossRef]
- Parzanini, C.; Colombo, S.M.; Kainz, M.J.; Wacker, A.; Parrish, C.C.; Arts, M.T. Discrimination between freshwater and marine fish using fatty acids: ecological implications and future perspectives. Environmental Reviews 2020, 28, 546–546. [Google Scholar] [CrossRef]
- Jardine, T.D.; Galloway, A.W.E.; Kainz, M.J. Unlocking the power of fatty acids as dietary tracers and metabolic signals in fishes and aquatic invertebrates. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 2020, 375, 20190639. [Google Scholar] [CrossRef]
- Bazarsadueva, S.V.; Radnaeva, L.D.; Shiretorova, V.G.; Dylenova, E.P. The comparison of fatty acid composition and lipid quality indices of roach, perch, and pike of Lake Gusinoe (Western Transbaikalia). Int. J. Environ. Res. Public Health. 2021, 18, 9032. [Google Scholar] [CrossRef]
- Zheng, S.; Qiu, M.; Wu, J.H.Y.; Pan, X.F.; Liu, X.; Sun, L.; Zhu, H.; Wu, J.; Huang, Y. Long-chain omega-3 polyunsaturated fatty acids and the risk of heart failure. Ther. Adv. Chronic Dis. 2022, 13, 20406223221081616. [Google Scholar] [CrossRef]
- Liu, F.; Xie, Q.; Yu, R.Q.; Xie, Z.; Wu, J.; Zhang, X.; Wu, Y. Fatty acids as bioindicators of organohalogen exposure in marine fish from a highly polluted estuary: First insight into small-scale regional differences, J. Hazard. Mater. 2023, 452, 131337. [Google Scholar] [CrossRef]
- Ferain, A.; Delbecque, E.; Neefs, I.; Dailly, H.; Saeyer, N.D.; Larebeke, M.V.; Cornet, V.; Larondelle, Y.; Rees, J.F.; Kestemont, P.; De Schamphelaere, K.A.C.; Debier, C. Interplay between dietary lipids and cadmium exposure in rainbow trout liver: Influence on fatty acid metabolism, metal accumulation and stress response, Aquat. Toxicol. 2021, 231, 105676. [Google Scholar] [CrossRef]
- Gabryelak, T.; Filipiak, A.; Brichon, G. Effects of zinc on lipids of erythrocytes from carp (Cyprinus carpio L.) acclimated to different temperatures. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2000, 127, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Imran, A.; Nosheen, F.; Fatima, M.; Arshad, M.U.; Afzaal, M.; Ijaz, N.; Noreen, R.; Mehta, S.; Biswas, S.; Rasool, I.F.U.; Aslam, M.A.; Usman, I.; Zahra, S.M.; Segueni, N.; Amer Ali, Y. Functional roles and novel tools for improving-oxidative stability of polyunsaturated fatty acids: A comprehensive review. Food Sci Nutr. 2023, 11, 2471–2482. [Google Scholar] [CrossRef] [PubMed]
- Carminato, A.; Pascoli, F.; Trocino, A.; Locatello, L.; Maccatrozzo, L.; Palazzi, R.; Radaelli, G.; Ballarin, C.; Bortoletti, M.; Bertotto, D. Productive results, oxidative stress and contaminant markers in european sea bass: Conventional vs. organic feeding. Animals 2020, 10, 1226. [Google Scholar] [CrossRef] [PubMed]
- Vlahogianni, T.H.; Valavanidis, A. Heavy-metal effects on lipid peroxidation and antioxidant defence enzymes in mussels Mytilus galloprovincialis. Chemistry and Ecology 2007, 23, 361–371. [Google Scholar] [CrossRef]
- Maria, B.; Maria, M.C.; Antonio, B.; Simona, M.; Rosaria, A.; Andrea, S.; Giulia, M.; Mario, S. Chemical and biochemical responses to sub−lethal doses of mercury and cadmium in gilthead seabream (Sparus aurata). Chemosphere 2022, 307, 135822. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogiannia, T.; Dassenakisb, M.; Scoullosb, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Fokina, N. Copper and Nickel Induce Changes in the Lipid and Fatty Acid Composition of Anodonta cygnea. J. Xenobiot. 2023, 13, 132–147. [Google Scholar] [CrossRef]
- Oliva-Teles, A. Nutrition and health of aquaculture fish. J. Fish Dis. 2012, 35, 83–108. [Google Scholar] [CrossRef]
Figure 1.
Map of sampling localities: Veliko Ratno Ostrvo (VRO) and Višnjica (VIS).
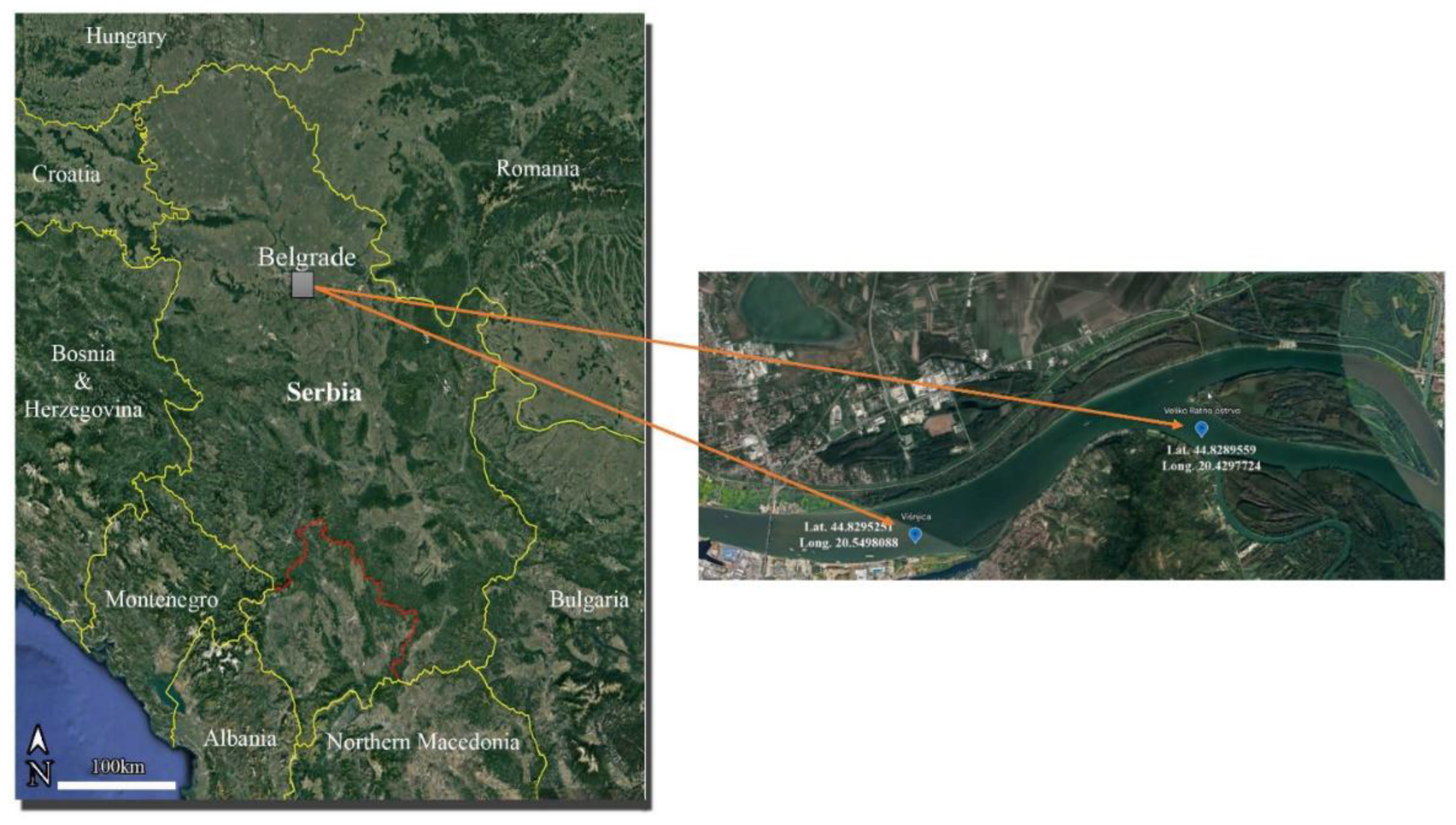
Figure 2.
Spearman correlation coefficients (scale on the right side of each figure) between all considered variables (metal concentration and individual fatty acids) for (a) roach (Rutilus rutilus) and (b) white bream (Blicca bjoerkna) collected at Veliko Ratno ostrvo (VRO) and Višnjica (VIS) sites in the Serbian part of the Danube.
Figure 2.
Spearman correlation coefficients (scale on the right side of each figure) between all considered variables (metal concentration and individual fatty acids) for (a) roach (Rutilus rutilus) and (b) white bream (Blicca bjoerkna) collected at Veliko Ratno ostrvo (VRO) and Višnjica (VIS) sites in the Serbian part of the Danube.
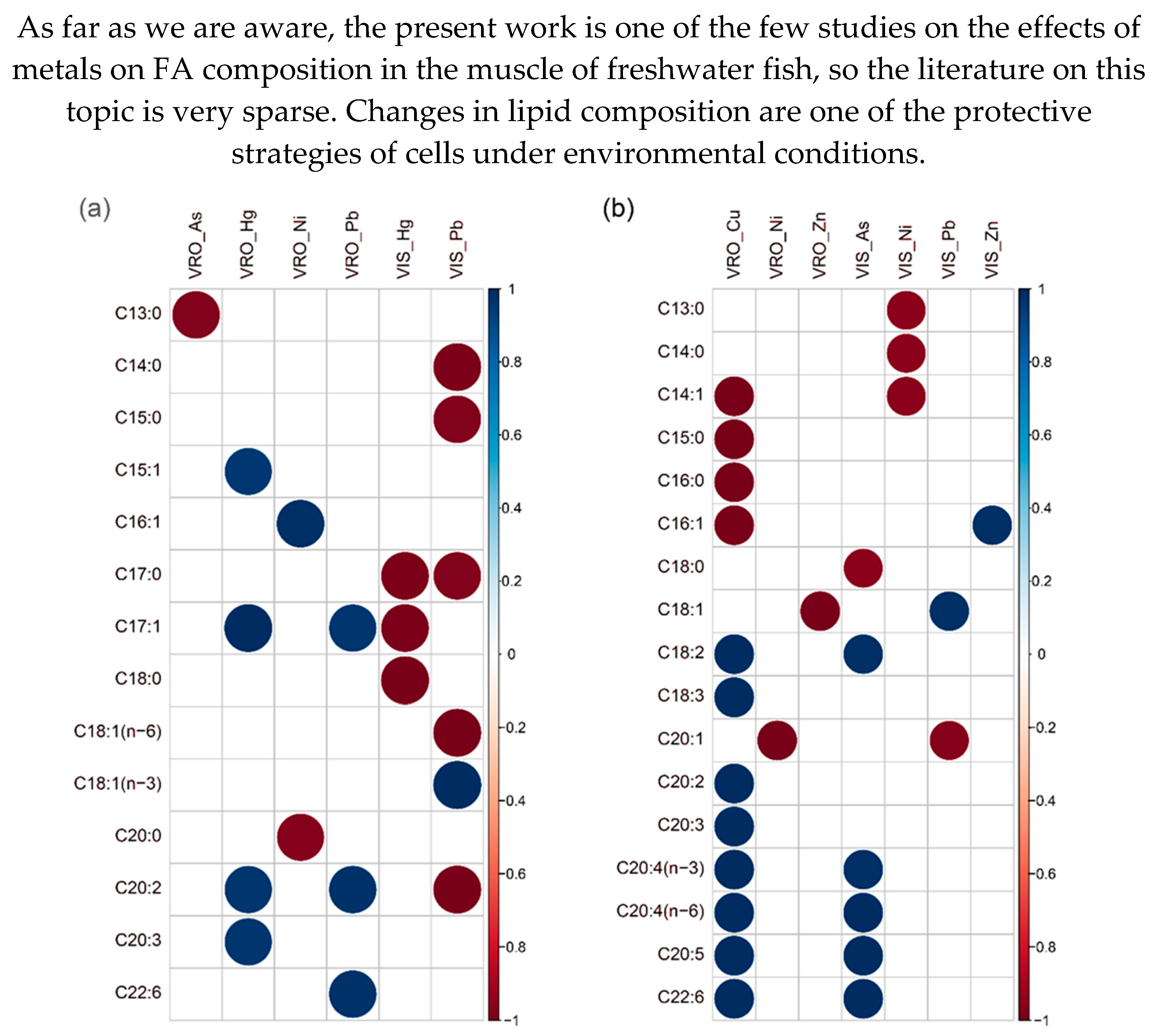

Table 1.
Metal concentrations (As, Cd, Co, Cr, Cu, Hg, Pb, Zn, and Ni) in the muscle of roach (Rutilus rutilus, n = 16) and white bream (Blicca bjoerkna, n = 16) (means ± standard deviation) at studied localities, Veliko Ratno ostrvo (VRO) and Višnjica (VIS). Concentrations are expressed as µg g -1 of wet weight. (Mann-Whitney U test, *—significant results in comparison with reference site – VRO, *p < 0.05).
Table 1.
Metal concentrations (As, Cd, Co, Cr, Cu, Hg, Pb, Zn, and Ni) in the muscle of roach (Rutilus rutilus, n = 16) and white bream (Blicca bjoerkna, n = 16) (means ± standard deviation) at studied localities, Veliko Ratno ostrvo (VRO) and Višnjica (VIS). Concentrations are expressed as µg g -1 of wet weight. (Mann-Whitney U test, *—significant results in comparison with reference site – VRO, *p < 0.05).
| Roach(Rutilus rutilus) | White bream(Blicca bjoerkna) | |||
|---|---|---|---|---|
| Metal (µg g -1) | VRO | VIS | VRO | VIS |
| As | 0.019 ± 0.003 | 0.072 ± 0.056* | 0.115 ± 0.067 | 0.048 ± 0.059 |
| Cd | bdl | bdl | bdl | bdl |
| Co | bdl | bdl | bdl | bdl |
| Cr | 0.019 ± 0.033 | 0.020 ± 0.034 | 0.110 ± 0.217 | 0.051 ± 0.033 |
| Cu | 0.264 ± 0.255 | 0.566 ± 0.107 | 0.202 ± 0.235 | 0.269 ± 0.186 |
| Hg | 0.080 ± 0.067 | 0.081 ± 0.031 | 0.109 ± 0.043 | 0.050 ± 0.029 |
| Ni | 0.480 ± 0.224 | 0.158 ± 0.085 | 0.317 ± 0.510 | 0.591 ± 0.542 |
| Pb | 0.023 ± 0.003 | 0.029 ± 0.002* | 0.028 ± 0.007 | 0.024 ± 0.007 |
| Zn | 9.002 ± 8.040 | 27.641 ± 7.890* | 12.935 ± 7.606 | 12.228 ± 7.730 |
*bdl – below the detection limit.
Table 2.
Fatty acids profiles (presented as percentage of total fatty acids) in muscle tissues of roach (Rutilus rutilus) and white bream (Blicca bjoerkna) catch from Veliko Ratno ostrvo (VRO) and Višnjica (VIS) localities. Data are expressed as mean ± SD.
Table 2.
Fatty acids profiles (presented as percentage of total fatty acids) in muscle tissues of roach (Rutilus rutilus) and white bream (Blicca bjoerkna) catch from Veliko Ratno ostrvo (VRO) and Višnjica (VIS) localities. Data are expressed as mean ± SD.
| Fatty acid | Roach (Rutilus rutilus) | White bream (Blicca bjoerkna) | ||
|---|---|---|---|---|
| VRO | VIS | VRO | VIS | |
| C11:0 | 0.009±0.008 | 0.008±0.006 | / | / |
| C12:0 | 0.592±0.145 | 0.707±0.327 | 0.400±0.060 | 0.653±0.271 |
| C13:0 | 0.099±0.027 | 0.156±0.133 | 0.082±0.029 | 0.189±0.047 |
| C14:0 | 6.517±0.963 | 7.545±2.456 | 5.035±0.530 | 6.477±1.471 |
| C14:1 | 0.099±0.049 | 0.144±0.093 | 0.076±0.034 | 0.133±0.036 |
| C15:0 | 1.381±0.242 | 1.767±0.841 | 0.825±0.153 | 1.320±0.328 |
| C15:1 | 0.876±0.172 | 1.328±0.928 | / | / |
| C16:0 | 19.255±13.279 | 25.456±10.740 | 38.399±2.296 | 35.152±3.605 |
| C16:1 | 14.210±2.222 | 10.063±7.105 | 11.379±0.648 | 11.973±1.705 |
| C17:0 | 0.988±0.435 | 1.346±0.699 | 0.528±0.185 | 0.949±0.325 |
| C17:1 | 0.692±0.208 | 0.928±0.469 | / | / |
| C18:0 | 7.123±2.120 | 7.711±3.395 | 4.975±0.461 | 6.887±1.292 |
| C18:1 | 0.157±0.051 | 0.185±0.090 | 27.772±1.551 | 24.905±1.381 |
| C18:1 | 19.360±10.579 | 19.512±12.102 | 4.056±0.123 | 5.090±0.832 |
| C18:2 | 11.770±2.241 | 10.551±1.807 | 3.556±2.464 | 3.256±2.010 |
| C18:2 | / | / | 0.090±0.057 | 0.076±0.012 |
| C18:3 | 0.289±0.054 | 0.299±0.109 | 0.231±0.033 | 0.378±0.360 |
| C18:3 | 1.635±0.469 | 1.284±0.512 | / | / |
| C20:0 | 0.347±0.144 | 0.419±0.271 | 0.181±0.039 | 0.167±0.038 |
| C20:1 | 1.106±0.459 | 1.402±0.795 | 0.419±0.063 | 0.788±0.264 |
| C20:2 | 2.095±0.602 | 1.446±0.474 | 0.299±0.219 | 0.394±0.196 |
| C20:3 | 0.593±0.117 | 0.547±0.148 | 0.097±0.054 | 0.057±0.046 |
| C20:4 | 2.543±0.885 | 1.857±0.558 | 0.313±0.057 | 0.241±0.041 |
| C20:4 | / | / | 0.048±0.040 | 0.051±0.019 |
| C20:5 | 3.719±2.439 | 2.563±1.871 | 0.603±0.030 | 0.508±0.297 |
| C22:6 | 4.546±2.799 | 2.774±1.713 | 0.635±0.087 | 0.354±0.169 |
| ƩSFA | 36.311 | 45.116 | 50.424 | 51.794 |
| ƩMUFA | 36.499 | 33.563 | 43.703 | 42.890 |
| Ʃn-3 PUFA | 12.443 | 8.478 | 1.782 | 1.481 |
| Ʃn-6 PUFA | 14.746 | 12.843 | 4.091 | 3.836 |
| n-3/n-6 | 0.844 | 0.660 | 0.436 | 0.386 |
Σ SFA (saturated fatty acid), Σ MUFA (monounsaturated fatty acid), Σ n-3 PUFA (polyunsaturated fatty acid). Σ n-6 PUFA (polyunsaturated fatty acid).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



