
Submitted:
27 July 2023
Posted:
28 July 2023
You are already at the latest version
Abstract
Aquaculture assumes a pivotal role in meeting the escalating global food demand, and shrimp farming, in particular, holds indispensable significance for the global economy and food security, providing a rich source of nutrients for human consumption. Nonetheless, the industry faces formidable challenges, primarily attributed to disease outbreaks, and the diminishing efficacy of conventional disease management approaches, such as antibiotic usage. Consequently, there is an urgent imperative to explore alternative strategies to ensure the sustainability of the industry. In this context, the burgeoning field of epigenetics emerges as a promising avenue for combating infectious diseases in shrimp aquaculture. Epigenetic modulations entail chemical alterations in DNA and proteins, orchestrating gene expression patterns without modifying the underlying DNA sequence by utilizing DNA methylation, histone modifications, and non-coding RNA molecules. Exploiting epigenetic mechanisms presents an opportunity to enhance immune gene expression and bolster disease resistance in shrimp, thus revolutionizing disease management strategies and optimizing shrimp health and productivity. Additionally, the concept of epigenetic inheritability in marine animals holds immense potential for the future of the shrimp farming industry. To this end, this comprehensive review thoroughly explores the dynamics of epigenetic modulations in shrimp aquaculture, with a particular emphasis on its pivotal role in disease management. It accentuates the significance of harnessing advantageous epigenetic changes to ensure the long-term viability of shrimp farming while deliberating on the potential consequences of these interventions. Overall, this appraisal highlights the promising trajectory of epigenetic applications, propelling the field toward strengthening sustainability in shrimp aquaculture.
Keywords:
Shrimps
; Disease management
; Epigenetics
; Epigenetic modulation
; Future applications
1. Introduction
In the face of a rapidly expanding global population, ensuring food security has become an increasingly formidable challenge. Climate change and the extensive degradation of natural resources further compound this issue. To address this challenge, the aquaculture sector assumes an indispensable role in meeting the escalating worldwide demand for food. In 2020, the aquaculture industry achieved a momentous feat, attaining a record-breaking aquatic production of 122.6 million tonnes valued at USD 281.5 billion [1]. Global aquaculture primarily centers on the commercial cultivation of a diverse range of marine vertebrates, such as teleost fish, and invertebrates, including mollusks and crustaceans [2]. Among teleost fish, the prevailing species cultivated in the aquaculture sector encompass carp, catfish, salmon, and tilapia [3,4]. However, it is the crustaceans, particularly shrimp, prawns, and crabs, that constitute the two most invaluable crustacean commodities traded on a global scale [3].
Shrimp, as a pivotal source of crustacean-derived seafood, assumes a critical role in the global economic landscape and food security. The shrimp industry serves as a substantial catalyst for the socioeconomic advancement of developing nations, providing a highly prized seafood replete with indispensable nutrients like protein, omega-3 fatty acids, and vitamins [5,6]. The Asian region dominates shrimp production, accounting for over 80% of the total output, which further augments foreign exchange earnings through shrimp exports [4]. According to the Food and Agriculture Organization (FAO), farmed shrimp production reached a staggering 4.1 million metric tons in 2020, with the global shrimp market projected to experience exponential growth in the coming years, fueled by escalating demand from the food industry [4]. However, notwithstanding their paramount importance, shrimp farming confronts formidable challenges primarily stemming from disease outbreaks, environmental fluctuations, and the improper utilization of chemicals in aquaculture [7,8]. Aquaculture enterprises also experience economic losses of over $3 billion every year due to diseases, and their spread is enhanced by the international trade of cultured shrimp, posing significant threats to the long-term sustainability of the industry [9,10]. The initial efficacy of antibiotics in mitigating shrimp infections has been compromised by the emergence and proliferation of antibiotic-resistant bacteria and the dissemination of antibiotic-resistant genes from shrimp farms to the surrounding environment has led to potential risks for the well-being of humans [11,12]. Consequently, the quest for alternative strategies that ensure the safety of humans, aquatic ecosystems, and the environment has become imperative. To address this challenge, various environmentally conscious approaches have been investigated as substitutes for conventional disease management techniques in shrimp aquaculture. There has been growing interest in the utilization of phytochemicals, bacteriophages, immunization protocols (Vaccinations), feed additives (such as probiotics and synbiotics), and cutting-edge genetic methodologies to control shrimp diseases [13,14]. However, an emerging field of scientific research called epigenetics has offered great promise for preventing and managing infectious diseases in shrimp aquaculture [15,16].
Epigenetics focuses on the study of heritable changes in gene expression and gene expression patterns without altering the underlying DNA sequence. Epigenetic modulations encompass chemical alterations that occur in both DNA and proteins, ultimately modulating the accessibility of DNA to the transcription machinery [16,17]. Notably, processes associated with DNA methylation, histone modifications, and non-coding RNA molecules have shown the capacity to orchestrate epigenetic changes in animals [18,19]. Recently, there has been a burgeoning interest in harnessing "targeted epigenetic modulations" to boost the biological capabilities of aquatic animals, including fish and shellfish [15,20]. The strategic deployment of such interventions holds tremendous promise in potentiating immune gene expression, fortifying resistance against diseases, and modulating the composition of the gut microbiome, thereby ultimately enhancing biological functions in shrimps. On the other hand, understanding the intricate interplay between epigenetic modulations and disease susceptibility in shrimp aquaculture has the potential to revolutionize disease management strategies [21,22]. By unraveling the specificities of epigenetic modulations, scientists can further unlock new strategies to optimize the health and productivity of shrimp populations, while minimizing the reliance on antibiotics and other conventional disease prevention methods. Moreover, given the evidence on epigenetic inheritability among marine animals, the proper application of this approach can have a tremendous impact on the future of the shrimp farming industry [15,19,22]. Furthermore, ongoing advancements in epigenetic research, if coupled with technological innovations in the field, present exciting opportunities to develop targeted and tailored interventions for preventing infectious diseases in shrimp aquaculture. Alternatively, it holds significant implications not only for the aquaculture industry but also for global food security and environmental sustainability.
Given this background, the primary objective of this comprehensive review is to elucidate the prospective application of epigenetics in the context of shrimp aquaculture for effective management and prevention of infectious diseases. It further underscores the significance of harnessing epigenetic regulatory mechanisms to ensure the long-term viability of the shrimp farming sector as a sustainable alternative to conventional disease control methodologies, Furthermore, this review will discuss the recent applications of epigenetic research, thereby presenting promising opportunities to develop targeted and precision-based interventions aimed at preventing infectious diseases in relation to the shrimp aquaculture. The elucidation of such scientific developments bears profound implications for the sustainable growth of the aquaculture industry and global food security.
2. Current Status of Global Production and Challenges in Shrimp Aquaculture
Within the realm of crustacean aquaculture, shrimp farming has garnered significant attention due to its high market value and relatively short production cycle. Since the establishment of commercial shrimp farms in 1970, shrimp aquaculture production has experienced consistent growth with accelerated expansion. It's popularity and rapid momentum surged after 1990, and in 2007, shrimp aquaculture production surpassed fisheries production for the first time, accounting for a ratio of 50.2% to 49.8% [4]. Until 2008, farmed shrimp production stood at approximately 3.7 million tons, representing over 50% of the global shrimp market (Figure 1). In 2018, global farmed shrimp production reached 4 million tons, indicating a 3-5% increase from the previous year [23]. The production of shrimp has witnessed a remarkable surge, surging from less than 0.6 million tons in 1980 to over 6 million tons in 2018, signifying nearly a ten-fold upsurge [10]. Projections for global shrimp production indicate a rise to 7.28 million tons by 2025, underscoring the pivotal role of shrimp aquaculture in meeting global food demand and the substantial growth of the industry throughout the years [24]. As the shrimp aquaculture industry has experienced rapid expansion over several decades, it has made significant contributions to the socio-economic advancement of coastal communities in numerous developing nations. Large-scale shrimp farming remains prevalent, with the majority of shrimp production concentrated in East and Southeast Asia and Latin America, while a substantial portion of consumption occurs in developed markets such as the United States of America, the European Union, and Japan [25]. Presently, the leading global shrimp producers include China, Thailand, Vietnam, Indonesia, India, Mexico, Malaysia, Brazil, and the Philippines, underscoring the predominant concentration of the industry in the Asian region, accounting for 80% of global production [26]. Among the various shrimp species cultivated for consumption, the dominant farmed species comprises white-leg shrimp (Litopenaeus vannamei) and the giant tiger prawn (Penaeus monodon), which collectively capture over 90% of the market share (Figure 2). However, L. vannamei has emerged as the preferred choice for cultivation over P. monodon in numerous regions due to factors such as the availability of special pathogen-free broodstock, lower protein requirements, production costs, tolerance to diverse water parameters, distinctive flavor profile, and nutritional value. Additionally, L. vannamei demonstrates adaptability to a broad range of salinity levels and temperatures, enabling convenient cultivation practices in inland waters [27,28,29]. Moreover, L. vannamei reigns as the favored shrimp species for processing owing to its higher meat yield (approximately 66%–68%) in comparison to P. monodon, and this characteristic has propelled them to archive greater popularity among American consumers, who represent the largest shrimp market globally [25]. The continued advancement of technology and the adoption of sustainable practices will be vital in ensuring the sustained prosperity and environmental integrity of shrimp aquaculture in the face of mounting demand and global challenges.
The expansion of shrimp aquaculture has encountered formidable challenges, particularly in the form of emerging diseases that pose substantial threats to production and economic stability. Throughout its history, the industry has grappled with global pandemics and diseases caused by pathogens, but it has demonstrated adaptability and resilience in coexisting with these challenges while expanding production [26,30]. Even in the present day, pathogens remain the most devastating threat to the shrimp aquaculture industry, resulting in significant annual economic losses amounting to approximately $3 billion, which accounts for a considerable amount of the industry's value [31,32]. Until 1990, the restricted transboundary movement of shrimp and their associated materials limited the spread of pathogenic agents such as viruses, parasites, bacteria, and fungi to specific geographic locations. However, the unrestricted international trade of shrimp and related materials, including broodstock, has facilitated the global expansion of shrimp diseases. Among these pathogens, viruses emerge as the most destructive threat to farmed shrimp [33,34]. A multitude of viral organisms capable of causing diseases have been identified, with white spot syndrome virus (WSSV) standing out as the most pervasive pathogen afflicting farmed shrimp on a global scale. The disease, known as white spot disease (WSD), was first reported in cultured kuruma shrimp in China and Taiwan in 1991/1992, rapidly spreading to other shrimp-farming countries in Asia and reaching Latin America by 1995 [35,36]. The cumulative economic losses due to WSD over the past two decades are estimated to be around US$ 15 billion globally, with WSSV causing approximately two-thirds of the annual economic loss, equivalent to approximately US$ 1 billion, and a 15% reduction in global shrimp production [37]. Among other viral diseases, the recently identified infectious myonecrosis disease (IMND) caused by infectious myonecrosis virus (IMNV) in farmed L. vannamei in Brazil in 2002, has rapidly spread to other shrimp farming regions through the importation of broodstock materials and current experimental studies have shown that all other commercial shrimp species, including P. monodon and P. stylirostris, are susceptible to IMNV infection [38]. In Brazil alone, IMNV has resulted in financial losses of approximately U.S. $440 million between 2002 and 2005, and by the end of 2011, estimated losses in Brazil and Indonesia have exceeded $1 billion [39]. Furthermore, there is evidence of co-infection of IMNV with WSSV among L. vannamei shrimp populations which makes the situation even worse [40].
Apart from viral infections, bacterial diseases, Vibriosis, and acute hepatopancreatic necrosis disease (AHPND) particularly mediated by Vibrio parahaemolyticus, have significantly impacted the global aquaculture industry in recent years. AHPND, previously known as early mortality syndrome (EMS), was first reported in China in 2009 and subsequently spread to Southeast Asia (Vietnam, Malaysia, Thailand, Philippines, and Bangladesh) and South America (Mexico and the USA) [30,41,42,43]. The disease is characterized by rapid and widespread mass mortality rates of up to 100% within the first 20-30 days of postlarval stages in commercial shrimp farms cultivating L. vannamei and P. monodon. Since the 2010s, AHPND has caused production declines in major shrimp-producing countries in Asia, resulting in substantial income losses estimated at over $10 billion, with annual losses reaching approximately $1 billion [10,44]. In addition to this significant economic impact, AHPND has also raised concerns regarding food security and the potential for ecological disruption. The disease's ability to cause mass mortalities in shrimp populations has led to disruptions in the global supply for consumption, impacting the global seafood market and potentially leading to higher prices for consumers [45].
Apart from the diseases, shrimps encounter multifaceted challenges in maintaining a balanced gut microbiota, which plays a pivotal role in modulating cellular functions. The gut microbiome exerts a profound influence on the immune response and can enhance resistance to pathogenic infections [46]. Several factors, encompassing water chemistry, feed composition, and antimicrobial usage, can disrupt the delicate equilibrium of the gut microbiome, resulting in recurring encounters with pathogenic invasions [47]. Additionally, fluctuations in environmental conditions, such as temperature and salinity, exert significant impacts on shrimp's immune and physiological functions. For instance, reduced temperatures during winter can attenuate immune responses, rendering shrimps more susceptible to pathogenic infections and salinity variations can impede ion transport and osmoregulation, critical processes for maintaining homeostasis [48,49]. Moreover, the presence of immunosuppressive agents, including pollutants, within their habitats can impair the biological functions of these animals [50]. On the other hand, the phenotypic traits of shrimp, including growth rate, disease resistance, and stress tolerance, are intricately linked to genetic factors. However, the limited genetic diversity, the absence of advanced breeding techniques, and the challenges posed by genetic constraints, such as inbreeding depression in commercial shrimp populations have hindered the development of superior shrimp strains with enhanced desirable traits [51,52].
With the intensification of shrimp aquaculture, the risk of rapid disease dissemination is amplified, impeding the implementation of effective control measures. As a countermeasure, recent scientific advancements have found ways to enhance the biological capabilities of shrimps. There is a burgeoning interest in the utilization of bacteriophages, vaccines, immune stimulants, and feed additives to combat infectious diseases in shrimp as viable solutions to antibiotics in the present industry.
3. Overview of Current Shrimp Disease Management Strategies.
Throughout its historical trajectory, the shrimp aquaculture industry has encountered recurrent global pandemics, with diseases exerting a pivotal influence on its evolution. While diverse pathogenic ailments will persistently present challenges, the industry has demonstrated remarkable resilience in managing and adapting to disease outbreaks during the preceding two decades, coinciding with a substantial amplification in production. This resolute response portends that the industry will persistently acquire knowledge and devise effective strategies to effectively confront and coexist with diseases as its global expansion persists [53]. Consequently, numerous approaches have been embraced to surmount the obstacles imposed by pathogens and are presently under inspection. These methodologies encompass the utilization of antibiotics, bacteriophages, vaccines, immune stimulants (such as algal extracts), and feed additives (including probiotics, prebiotics, and synbiotics) to combat infectious diseases in shrimp. Recent empirical findings substantiate the efficacy and safety of these modalities as viable solutions for disease control in the shrimp aquaculture industry.
The foremost approach employed for shrimp disease prevention is the utilization of antimicrobials and antibiotics. Nevertheless, toward the conclusion of the 20th century, practical impediments emerged in the application of antibiotics due to the acquisition of tolerance by microorganisms to these chemical agents [11,53]. Consequently, the adoption of antibiotics and chemical agents to combat bacterial infections in shrimps has facilitated the dissemination of antibiotic-resistant bacteria, thereby engendering environmental hazards and the deposition of drug residues in their tissues [54,55,56]. Recent investigations also showcased a higher prevalence of antibiotic residues in farmed shrimp as compared to their wild-caught counterparts, highlighting the pervasive and unregulated usage of antibiotics in shrimp farming to augment production [56,57,58,59]. Furthermore, the genetic elements conferring drug resistance in numerous antibiotic-resistant bacteria can be transmitted to pathogenic microorganisms via contaminated aquaculture feed, thus posing a considerable health concern for human populations [54]. Shrimp pathogens, particularly those belonging to the Vibrio genus, have demonstrated such resistance to antibiotics commonly employed in aquaculture settings. The ingress of antibiotic-resistant strains into shrimp farms can transpire via seawater, as effluents laden with antibiotics and chemicals are discharged into marine environments. On the other hand, excessive employment of antibiotics and chemicals in shrimp aquaculture ponds contributes to the degradation of water quality, culminating in immune suppression induced by stress events [57]. Considering the mounting peril of climate change, the prevalence of antibiotic resistance in pathogens is anticipated to escalate, imposing repercussions on both aquatic organisms and human well-being.
Vaccination serves as a vital prophylactic measure in aquaculture, particularly in marine vertebrates (e.g., fish), to augment the host's immunological defenses against infectious agents. Given the existence of an adaptive immune system and the potential for immune memory, vaccines exert markedly greater effectiveness in these organisms [60,61,62]. Nonetheless, invertebrates such as shrimp lack an adaptive immune system, thereby raising the pertinent query: "Can vaccination be efficacious in shrimp?". However, diligent research endeavors have been directed toward the development of DNA vaccines for shrimp employing biodegradable carriers such as chitosan, and encouraging outcomes from these investigations demonstrate that oral administration of chitosan in P. monodon confers immune enhancement and safeguards against the WSSV [63,64]. Liposome-based vaccines, structurally resembling cellular membranes, have also been explored In shrimp aquaculture, and notably, the administration of a liposome-based recombinant VP28 vaccine has yielded impressive survival rates against WSSV in Marsupenaeus japonicas [65]. Additionally, the exploitable potential of formalin-inactivated Vibrio, administered orally to Fenneropenaeus merguensis postlarvae, has elicited noteworthy protection against subsequent Vibrio harveyi infections [66]. However, the development of a commercial shrimp vaccine represents a multifaceted process that necessitates the requirement of intricate comprehension of the presence of shrimp's unique "adaptive-like immunity."
Bacteriophages (phages) have emerged as promising therapeutic agents in the field of aquaculture, owing to their remarkable efficacy and safety in combating pathogenic bacteria. These viral predators offer an environmentally friendly approach, capitalizing on their innate ability to selectively target and eliminate specific bacteria while sparing non-target microbiota due to their restricted host range and “Phage therapy”, involving the deliberate administration of phages, holds great potential for the management of bacterial diseases in aquaculture [67,68]. The delivery of phages can be achieved through various means, including injection, dietary administration, or their incorporation into rearing systems [69]. Among these, dietary administration is particularly favored as it facilitates the treatment of large populations and offers protection against intestinal infections. In recent times noteworthy successes have been documented in the application of phage therapy for the treatment of AHPND caused by V. parahaemolyticus in shrimp aquaculture [70]. Additionally, the utilization of phage cocktails, consisting of combinations of multiple phages, has demonstrated synergistic effects and superior efficacy compared to individual phages against V. parahaemolyticus and V. harveyi in artemia shrimp models [71]. Furthermore, phages have displayed promising anti-biofilm activities against shrimp pathogens, with specific phages disrupting bacterial protein machinery and thereby implying the existence of encoded antimicrobial compounds [72]. However, it is imperative to acknowledge the possibility of bacterial resistance to phages, arising from receptor loss on the cell membrane as well as the presence of restriction enzymes and CRISPR systems within bacteria [73,74,75]. Horizontal gene transfer has been observed in some lytic phages, enabling the dissemination of antibiotic-resistant genes and the occurrence of virulent mutants among temperate phages may complicate treatment regimens as they possess the capacity to revert to lysogeny [75]. Thus it is essential to carefully consider the development of phages or tailored phage cocktails that are capable of overcoming above obstacles to ensure both efficacy and safety.
In recent years, there has been a growing interest in the utilization of Immunostimulants as a viable alternative to antibiotics for boosting the immune system of animals. Immunostimulants encompass naturally occurring and/or synthetic substances derived from diverse sources, including bacteria, algae, animals, and plants [76]. Within the shrimp aquaculture industry, the efficacy and multifunctionality of immunostimulants, such as polysaccharides derived from seaweeds, have been extensively investigated. Notably, brown seaweeds have been identified as a valuable source of bioactive compounds, including fucoidan and alginate, which possess immunostimulant properties and confer resistance against shrimp pathogens [77]. Moreover, plant extracts from various herbs (such as Eclipta alba, Aegle marmelos, and Cyanodon Dactylon) have demonstrated antiviral and immunostimulant activities against viral organisms like WSSV [78]. These scientific findings underscore the perspectives of immunostimulants derived from diverse sources as effective alternatives for disease management in the shrimp farming industry. Furthermore, the utilization of feed additives (including probiotics, prebiotics, and synbiotics) has caught the eye in recent times as a promising strategy for enhancing shrimp biological functions. Probiotics, which encompass bacteria-based products, have showcased abilities to promote health, prevent diseases, and control the proliferation of pathogenic organisms [79]. Notably, several bacterial species, such as Bacillus, Vibrio, and lactic acid bacteria, have been introduced as probiotics in shrimp farming, demonstrating their potential in enhancing the overall health of shrimp populations [80,81]. On the other hand, prebiotics are indigestible fibers capable of fostering the growth and activity of beneficial bacteria within the gut, thereby promoting shrimp health. Several prebiotics have been introduced to shrimp farming including inulin, fructooligosaccharides (FOS), and mannan oligosaccharides (MOS), which are derived from plant sources and fibers [82,83]. Synbiotics, (representing a combination of prebiotics and probiotics) exhibit a synergistic relationship by facilitating the gut microbiome growth while enhancing the immune resistance in shrimps against pathogens as demonstrated by the supplementation with Lactiplantibacillus. plantarum and galactooligosaccharide [84]. The selection of specific prebiotics and probiotics in synbiotic formulations is based on their ability to enhance the proliferation of beneficial bacteria and confer overall benefits to the host. However, overall, it is important to acknowledge that the efficacy of these control methods may exhibit their positives, negatives, variabilities, and inconsistencies. Also, the possibility of these archived traits may pose challenges in terms of their stability and heritability over multiple generations, thereby raising significant concerns regarding the economic sustainability of the shrimp aquaculture industry.
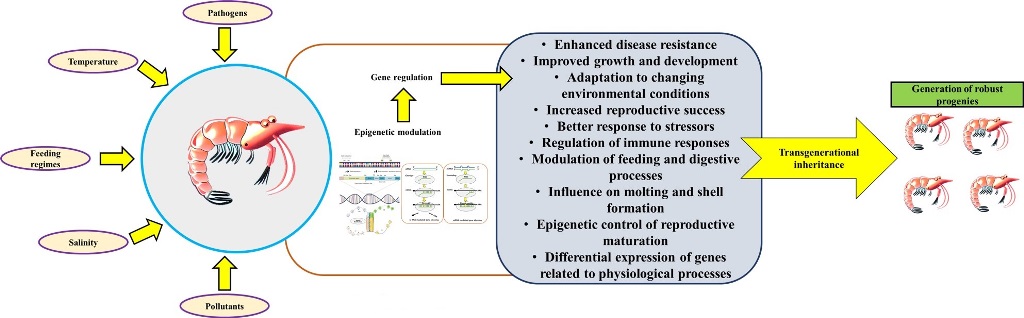
4. The Concept of Epigenetics, Epigenetic Inheritance, and Its Potential Effect on Disease Management in Shrimp
Shrimp aquaculture, as a pivotal component of global food production, confronts a multitude of challenges, including the emergence of diseases that exert a substantial impact on shrimp health and production. Conventional disease management strategies, such as the implementation of antibiotics and vaccines, possess inherent limitations and potential drawbacks. Consequently, researchers and industry professionals have redirected their focus toward alternative approaches, notably the potential of utilizing epigenetic modulation, to enhance disease resistance and bolster aquatic animal health [16,85,86]. Epigenetic modulation, a noteworthy method, encompasses the modification of gene expression patterns devoid of alterations to the underlying DNA sequence [87]. Within aquatic organisms, these epigenetics-associated regulatory mechanisms play a pivotal role in governing a diverse array of biological processes, encompassing development, adaptation, and response to environmental stimuli [16,88]. Epigenetic modulations are governed by three fundamental mechanisms, namely DNA methylation, histone modifications, and non-coding RNA regulation [89,90]. In the context of shrimp disease management, epigenetic modulation offers numerous advantages.
Firstly, it furnishes a mechanism for swift and reversible adaptation to dynamic environmental conditions and pathogenic challenges. It has been demonstrated that shrimp like other animals, possess the capacity to potentially modify their epigenetic marks, thereby activating immune-related genes, has the potential to augment disease resistance, which enables shrimp to mount efficacious defenses against pathogens [15,22,91]. Secondly, epigenetic modulations can influence the expression density of genes implicated in immune response pathways, thereby enabling shrimp to fine-tune their immune response and counteract diseases [91]. Hence, it may serve as a potential avenue for enhancing the shrimp's immune system without reliance on external agents thus mitigating the risks associated with side effects and environmental contamination that are frequently linked with conventional disease management strategies.
An enthralling facet of epigenetic modulation lies in the prospect of “transgenerational inheritance”, whereby these epigenetic marks are transmitted from one generation to the next. This phenomenon has garnered extensive attention in investigations of other organisms, inciting interest in its applicability to shrimp disease management. Epigenetic inheritance furnishes the potential for "priming" future generations of shrimp, rendering them more resistant to diseases [91,92]. Once established, epigenetic marks can be perpetuated and transmitted to future generations, potentially endowing lasting resistance to diseases. This facet of epigenetic modulation aligns harmoniously with the principles of sustainable aquaculture and also mitigates reliance on external interventions. Through exposure to specific environmental cues or epigenetic modifiers, it may be plausible to induce heritable epigenetic changes that enhance the biological functions of these animals. However, the mechanisms and extent of epigenetic inheritance in shrimp remain incompletely understood and necessitate further exploration. Studies aimed at identifying specific epigenetic marks associated with disease resistance and assessing their stability across generations will yield valuable insights into its applicability.
5. Types of Epigenetic Modifications
5.1. DNA Methylation
In the field of epigenetics, DNA methylation emerges as the most extensively investigated phenomenon in the current research landscape. DNA methylation encompasses the addition of a methyl group to nucleotides within a DNA strand, facilitated by DNA methyltransferase (DNMTs) enzymes. These DNMTs associated with DNA methylation are divided into maintenance DNMTs (DNMT1, and DNMT2) and De novo DNMTs (DNMT3) where DNMT1 is the most abundant and responsible for the management of existing DNA methylation patterns (They are capable of recognizing hemimethylated DNA and the addition of methyl groups to the newly synthesized DNA strand to preserve the methylation pattern) while DNMT2 is associated with the methylation of RNA [93,94,95]. De novo DNMTs (including DNMT3 A or B) are involved in the establishment of new DNA methylation patterns and are capable of methylating previously unmethylated DNA regions, hence showcasing pivotal roles in biological processes such as gametogenesis [96]. Many vertebrates possess DNMTs and in shrimp, the presence of all 3 DNMTs has been identified suggesting the possible presence of DNA methylation in this organism [97].
Based on previous scientific research, it has been proven that DNA methylation occurs across diverse genomic regions, including promoters and enhancers regions, ultimately influencing various gene expression outcomes dependent on the level of methylation [98,99]. Notably, methylation of promoter regions is often associated with gene silencing, leading to the inhibition of further gene expression where the impact of DNA methylation on enhancer regions is context-dependent, resulting in either gene expression inhibition or enhancement [100,101]. There is a growing body of evidence highlighting significant differences in DNA methylation patterns between invertebrates (such as shrimps) and vertebrates. Also, DNA methylation among different species might be evolution dependent, showcased by the high variability of methylation dynamics between vertebrate taxonomic groups such as mammals and reptiles [102]. However, in comparison to vertebrate animals, invertebrates have lower levels of DNA methylation in their genomes. Though a significant proportion of invertebrates exhibit DNA methylation levels ranging from 20% to 40%, shrimps, in contrast, display a comparatively low DNA methylation level of approximately 2% relative to their genome size [102]. On the other hand, vertebrates predominantly exhibit prominent CpG site methylation (characterized by regions containing a cystine nucleotide followed by a guanine nucleotide) across their genomes. However, the CpG islands, that contain a high frequency of CpG dinucleotides (which has shown to coincide with gene promoter regions) are typically unmethylated and correlate with active gene expression, allowing the transcription machinery to access the DNA and initiate gene transcription and this process has been shown to play a critical role in the development, cellular differentiation, and maintenance of cellular integrity in vertebrates demonstrated by previous studies in humans and mice [103,104]. Conversely, invertebrates, (including insects, mollusks, and crustaceans) exhibit variable frequencies of CpG sites or lack CpG islands altogether, showcasing that CpG methylation is not the only factor explaining DNA methylation across these species, suggesting the utilization of alternative forms of methylation (Named as non CpG methylation) such as CpA or CpT sites [102,105]. Furthermore, DNA methylation in vertebrates such as fish, birds, and mammals, has shown more pronounced tissue and cell type specificity and inter-individual differences, sculpting gene expression patterns and ensuring proper cellular functions [102]. Vertebrates finely regulate gene activities by selectively methylating and/or demethylating specific genome regions, contributing to this tissue specialization or cell differentiation, and in invertebrates, this has not been the case even though they have versatile ranges of potential methylation regions, and has not shown any significant tissue specificity or inter-individual differences in DNA methylation patterns, where they are often correlated to the developmental stages and/or environmental stimuli, signifying a dynamic and adaptive nature that allows them covey rapid adjustments in respective gene expression patterns [102]. Given this degree of diversity, it has been suggested that invertebrates may be attributable to the utilization of diverse mechanisms beyond DNA methylation for gene expression and cellular processes.
In addition to variations in methylation types, noteworthy distinctions exist in the genome-wide distribution of DNA methylation between vertebrates and invertebrates. Vertebrates typically exhibit a global pattern of DNA methylation throughout the genome, while invertebrates display a mosaic pattern characterized by regions of heavy methylation interspersed with nonmethylated regions, which is more variable and species-specific [102,106]. These mosaic methylation patterns likely play a role in the target-specific regulation of genomic regions or functions, suggesting potential divergence in the regulatory mechanisms associated with DNA methylation in these organisms. Another notable difference between invertebrates and vertebrates lies in gene body methylation, which is prevalent in vertebrates and plays a significant role in alternative splicing and conversely, the majority of invertebrates exhibit lower levels of gene body methylation (Figure 3) [102,107,108]. The absence or reduced presence of gene body methylation in invertebrates suggests the employment of alternative molecular strategies, such as histone modifications, chromatin remodeling, or other epigenetic mechanisms, to achieve similar regulatory outcomes as gene body methylation in vertebrates. Additionally, DNA methylation in vertebrates serves a crucial role in controlling the activities of transposable elements (TEs) within the genome. By adding a methyl group to the DNA sequence of TEs, DNA methylation can manipulate defense mechanisms to reduce their mobility and transcription, thereby preserving genomic integrity [109]. In contrast, invertebrates like shrimp typically rely on mechanisms such as small RNA-based silencing pathways, involving small interfering RNAs (siRNAs) and piwi-interacting RNAs (piRNAs), to control post-transcriptional or transcriptional gene silencing highlighting the diverse strategies employed by these organisms [110,111].
5.2. Histone Modifications
Histone modifications are fundamental epigenetic mechanisms that intricately regulate gene expression patterns. Histones, crucial protein molecules involved in DNA packaging within chromatin, undergo chemical alterations that dynamically modulate DNA accessibility to regulatory proteins. Among animals, a diverse repertoire of histone modification programming has been unraveled, encompassing methylation, acetylation, ubiquitination, phosphorylation, sumoylation, and ADP-ribosylation [112]. While the role of histone modifications in vertebrates (including mammals, birds, reptiles, amphibians, and fish) has been extensively studied providing a comprehensive understanding of their impact on gene regulation, and in invertebrates, such as crustaceans, this epigenetic mechanism has garnered less attention, consequently leaving it’s significance unexplored.
Compared to invertebrates, vertebrates generally exhibit a more complex chromatin structure where they are organized into distinct domains, including euchromatin and heterochromatin, as well as higher-order structures like nucleosomes and chromatin loops. In contrast, invertebrate chromatin possesses a relatively simpler structure with fewer defined domains with some higher-order structures [113,114,115]. It is thought that these structural disparities between vertebrates and invertebrates might be attributed to the differences in evolutionary history, genetic mechanisms, and the specific necessity of the regulatory mechanisms associated with histone modifications [116]. Among different mechanisms used, histone acetylation is the most extensively studied histone modification relies on the enzymatic actions of histone acetyltransferases (HATs) and histone deacetylases (HDACs), that exert pivotal roles in regulating a wide range of physiological phenomena including cell differentiation, proliferation, metabolism, senescence, immune response, and programmed cell death [117,118]. Recent research on the organisms in vertebrates and invertebrates has shown that histone acetylation primarily occurs on four histone proteins (H2A, H2B, H3, and H4), with H3 and H4 exhibiting the highest levels of acetylation [119,120]. Notably, acetylation of histone H3 can modulate ATP-dependent chromatin remodeling, thereby contributing to the inflammatory response, while histone H4 acetylation influences chromatin transcription and replication [121,122,123]. Concerning shrimps, all four histone proteins capable of acetylation have been identified, revealing their involvement in possible regulatory mechanisms [124]. Moreover, a recent study, directed using the artemia model indicated the presence of a significantly higher level of H3 acetylation in the treatment group (parents and their progeny cysts) compared to its control group following phloroglucinol treatment, where H4 acetylation levels were significantly lower in the treatment group compared to the respective controls [15].
Histone methylation and/or demethylation represent another fundamental process implicated in the modulation of epigenetic alterations. The methylation of histones is catalyzed by histone methyltransferases (HMTs), while demethylation is executed by histone demethylases (HDMs), and their collective effort, along with the recruitment of various cofactors and chromatin modifiers, governs the precise patterns of this process, thus contributing to the regulation of diverse cellular functions. Although histone methylation/demethylation is a conserved histone modification method found in both vertebrates and invertebrates, distinct variations and functional outcomes can be expected depending on the context of this process. In contrast, it has been proven histone methylation in vertebrates (such as mammals) can lead to gene activation or repression, depending on the specific site and degree of methylation [125]. Notably, the methylation of histone H3 (at the H3K4 location) has demonstrated the exclusive presence of three types of methylation, namely H3K4me1, H3K4me2, and H3K4me3 in genes and promoter regions. The H3K4me3 and H3K4me2 methylations primarily occurred in promoters, particularly in the 5' end of transcribed regions, while H3K4me1 is depleted in promoters but enriched within transcribed regions [126,127,128]. With respect to shrimps, recent scientific evidence on the artemia model has shown conserved histone H3K4 methylation patterns with increased levels of H3K4me3, H3K4me1, and H3K4me2, indicating enhanced trimethylation at H3K4 sites similar to that of vertebrates. However, the probability of inheriting the H3K4 methylation has not been constant leading to a progressive decline in the respective succeeding generations. The same research study demonstrated the methylation occurrence in K9 and K27 positions in the H3 protein as well (Figure 4) [15]. Another study on Marsupenaeus japonicus has shown the potential utility of histone H3 methylation in enhancing trained immunity, suggesting its involvement in possible epigenetic inheritance [129].
Phosphorylation, ubiquitination, and sumoylation are additional histone modifications that play important roles in gene expression regulation by modulating chromatin structure and recruiting regulatory proteins. These modifications have been extensively characterized in vertebrates, such as mouse, and invertebrates such as Common fruit fly (Drosophila melanogaster) and Oysters (Crassostrea virginica), while our understanding of their significance in crustaceans, remains limited [130,131,132,133]. In the model organism D. melanogaster, phosphorylation of histone H3 at specific serine residues (H3S28 and H3S10) is linked to both gene activation and repression, respectively, at enhancers and promoters [134]. Additional investigations into the role of the chromatin-remodeling factor SMARCA1 in Drosophila neural stem cell differentiation also revealed the involvement of histone H3 phosphorylation (at H3S10) is linked to gene activation and repression [135]. A similar type of serine phosphorylation in the histone molecule H2A has been observed in oysters (C. virginica) following exposure to brevetoxin [132]. Moreover, histone ubiquitination in Drosophila has been implicated in transcriptional activation and repression. For example, histone H2B ubiquitination has been linked to the transcriptional activation of certain genes (such as heat shock proteins), while ubiquitination of histone H2A is associated with gene silencing [136,137,138]. Among the crustaceans, unique H2A H4 phosphorylations have been detected in the mitten crab (Eriochier sinensis) and, in mantis shrimps (Oratosquilla oratoria) in H3 and H2A in the nuclei of spermatozoa [139,140]. Furthermore, a recent study on L. vannamei infected with AHPND-causing pirAB toxins revealed the occurrence of changes in H3 phosphorylation (at H3S10), which led to hemocyte apoptosis events [141]. Additionally, in the Chinese white shrimp (Fenneropenaeus chinensis) increased levels of H3 phosphorylation have been highlighted following WSSV infections [142]. However, It is imperative to acknowledge that the exploration of histone modifications in crustaceans remains a burgeoning area of investigation, necessitating further scientific inquiry to elucidate the intricate mechanisms that govern gene expression and cellular processes in these organisms.
5.3. Elements of RNA
Non-coding RNAs (ncRNAs) have emerged as pivotal players in the realm of epigenetic modifications, encompassing both vertebrates and invertebrates. These ncRNAs, which do not code for proteins, are classified into two categories as housekeeping and regulatory ncRNAs, where based on their size regulatory ncRNAs are further categorized as short-chain ncRNAs (including siRNAs and miRNAs) and long non-coding RNAs (lncRNAs) (Figure 5) [89,143,144]. These ncRNAs have demonstrated crucial roles in various processes, including stress responses and immune functions particularly involving the mechanism called “RNAi (RNA interference)”, which is a conserved biological process that involves the silencing of gene expression by targeting specific mRNA molecules for degradation and/or translational repression [145,146,147]. While there are shared regulatory principles of ncRNAs in vertebrates and invertebrates, the evolutionary and functional divergence among these groups has given rise to some peculiar differences [148,149]. Furthermore, the repertoire and functional significance of these ncRNAs can vary even across the invertebrate taxa, reflecting their adaptations to a wide range of lifestyles.
Among short-chain ncRNAs, the endogenous small RNA molecules that are transcribed from specific genomic regions or commonly known as microRNAs (miRNAs) have garnered significant attention in animals due to their ability on post-translational regulation of genes through partial binding to the 3' untranslated region (UTR) of messenger RNA (mRNA) [150]. This binding can either promote, inhibit or change the translation, leading to changes in various biological processes such as embryonic development, immune responses, neuronal development, and the pathogenesis of diseases (including cancer and neurodegenerative disorders), as demonstrated by various studies from vertebrates and invertebrates [151,152,153,154]. In the context of shrimps, the first set of miRNAs (35 miRNAs) was identified from M. japonicus hemocytes in 2011, 11 of which showed homology to miRNAs found in other arthropods [155]. Subsequent research has focused on miRNAs associated with defense response regulation in P. monodon against WSSV pathogen, revealing specific miRNAs (including miR-7, miR-965, and miR-12) that play crucial roles in virus-host interactions, affecting viral gene expression, replication, immune-related gene targeting, apoptosis, and immune recognition [156]. Similarly, in the case of V. alginolyticus infection in M. japonicus, differentially expressed miRNAs (including miR-100, miR-275, and miR-279) have been linked to innate immune responses involving phenoloxidase enzyme activation, apoptosis, and phagocytosis. Notably, miR-100 has directed a pivotal role in regulating shrimp hemocyte apoptosis, promoting the anti-Vibrio immune response by influencing factors such as the extent of hemocyte count [157]. Furthermore, differentially expressed miRNAs, such as miR-210, miR-10b, and miR-193, have been associated with immune-related processes in AHPND caused by V. parahemolyticus, offering potential insights into immune responses [158]. Additionally, environmental factors such as copper exposure and heat stress have also been instrumental in the identification of novel miRNAs in shrimp [159,160].
Small interfering RNAs (siRNAs) are another type of ncRNA that have been identified as key regulators of potent antiviral defense mechanisms in both vertebrates and invertebrates. The siRNAs are double-stranded RNA molecules that are typically generated from exogenous sources, such as viral genomes or transposable elements. They guide the RNA-induced silencing complex (RISC) to respective complementary target mRNAs, leading to their degradation and translation prevention [161,162]. This process, known as post-transcriptional gene silencing, serves as a defense mechanism against foreign genetic elements, and in shrimp, it has been shown to play a crucial role in the immune defense against viral infections by effectively degrading viral RNAs and restricting their replication. In shrimp, such as M. japonicus, the ability to generate an antiviral siRNA (vp28-siRNA) in response to WSSV infection has been detected and its production and function have been shown to coincide with the availability of RISC [163]. Considering the association of miRNAs and siRNAs with epigenetic regulation, these newly discovered ncRNAs hold great promise for disease management strategies in shrimp aquaculture.
Piwi-interacting RNAs (piRNAs) are another distinct class of non-coding RNAs (ncRNAs) that plays pivotal roles in gene regulation and genome defense mechanisms. These small RNA molecules interact with piwi proteins, which are a subgroup of the Argonaute protein family. In comparison to other classes of small RNAs, such as miRNAs and siRNAs, piRNAs are primarily expressed in the germline and are involved in maintaining genome integrity [164,165]. Moreover, in both vertebrates and invertebrates, piRNAs are capable of epigenetic modulations through the silencing of transposable elements (TEs), which are mobile genetic elements capable of disrupting genome stability. While interacting with piwi proteins, piRNAs guide the recognition and targeting of complementary sequences within TEs, resulting in their transcriptional and post-transcriptional repression and this mechanism is vital for preventing the harmful effects of TEs, such as genomic instability and germ cell dysfunction [166,167]. In invertebrates, the roles of piRNAs have been extensively studied, as they are more abundant and highly diverse in these taxa than in vertebrates and it is hypothesized that the expansion and diversification of transposons in invertebrate genomes have driven the evolution of robust piRNA pathways as a self-defense mechanism [168,169,170]. Regarding shrimps, several piwi proteins and piRNA-like molecules have been identified, however, their role in epigenetic modulation remains to be explored [171,172]. The long non-coding RNAs (lncRNAs) are also a regulatory type of ncRNA, with essential regulatory functions particularly in gene expression regulation at the transcriptional and/or post-transcriptional levels through interactions with DNA, RNA, and proteins. In shrimps a large number of lncRNAs have been identified, revealing species-specific and tissue-specific expression among L. vannamei, P. monodon, and Macrobrachium rosenbergii [173,174,175].
Overall, the diverse repertoire of ncRNAs showcases the intricate mechanisms underlying shrimp biology and the possible utilization of these mechanisms for future applications in epigenetic modulation. Furthermore, harnessing the potential of these ncRNAs for disease management and selective breeding programs in shrimp aquaculture holds great promise for the sustainable and efficient production of these economically important crustaceans.
6. Applications of Epigenetics for Immunity Enhancement and Disease Management in Shrimp Aquaculture
Epigenetic modulation through targeted modification of epigenetic marks holds considerable promise for augmenting various molecular pathways in shrimps. Such modifications can be achieved through multifarious strategies, encompassing the administration of pharmacological agents and/or dietary supplements, environmental interventions, and gene-centric methodologies tailored to specifically target and modify epigenetic signatures. In the context of shrimps, recent investigations have demonstrated the possibility of epigenetic mechanism manipulation by utilizing above mentioned key strategies. For example, the efficacy of employing specific methyl donors with DNA methylation capacity, such as S-adenosylmethionine (SAM) and L-methionine or methionine hydroxy analogs, as dietary supplements have shown to modulate antioxidant capacity, cell proliferation ability, protein synthesis alternatively leading to better growth and defense performance in L. vannamei [176,177]. Conversely, dietary supplements enriched with folic acid, an indispensable nutrient for DNA synthesis, have exhibited ameliorative effects on the immunity-associated functions of M. rosenbergii [178]. Furthermore, in the artemia model, the application of plant-based phenolic compounds as a biological control treatment has evinced transgenerational effects, instigating an inherited enhancement in resistance against Vibrio that persists across subsequent generations, indicating a transgenerational response to the treatment. This heightened resistance has been postulated to be associated with augmented expression of innate immune genes, with DNA methylation serving as a regulatory factor governing the expression of these genes [15,91]. In addition, environmental interventions have been observed to yield promising outcomes in terms of inducing epigenetic modifications. Various factors, including temperature fluctuations, salinity, water quality, air availability, and feeding regimes, have been found to impact epigenetic changes and the functionality of immune-associated processes in animals, including shrimps. The extant evidence suggests that shrimps are capable of undergoing epigenetic changes and exhibiting transgenerational resilience in response to heat stress. For instance, the association between epigenetic mechanisms and temperature-induced tolerance to lethal heat shock and resistance against the pathogenic bacteria V. campbellii has been demonstrated in the artemia model. This study revealed altered levels of DNA methylation and increased acetylation of histones H3 and H4, accompanied by changes in heat shock protein 70 (HSP70) levels [22]. Building upon these intriguing findings subsequent studies validated that exposure to other environmental stimuli, such as bioactive food components, could induce similar epigenomic modulation in artemia, resulting in the generation of robust phenotypes with resistance against V. campbellii over three successive generations [91]. Furthermore, alterations in water quality parameters, such as ammonia/nitrate levels, and exposure to environmental pollutants can instigate oxidative stress and potentially inflict damage to DNA in crustaceans, thereby potentially modulating epigenetic changes [179,180,181]. Additionally, air availability has been shown to influence DNA methylation patterns in kuruma shrimps, coinciding with changes in apoptosis and physiological responses [182]. Moreover, feeding regimes, entailing carefully devised plans encompassing diet composition, nutrient intake, and schedule, can exert significant impacts on the epigenetic changes associated with shrimp immunity and defense functions.
In addition to the aforementioned approaches, gene-centric methodologies have recently emerged as effective means to elicit epigenetic alterations in animals. Among these methodologies, RNA interference (RNAi) stands out as the most commonly employed technique, entailing the attenuation or downregulation of specific target genes through the utilization of synthetic small-interfering RNA (siRNA) molecules [183]. Due to its versatility, RNAi has proven to be a valuable tool for inducing epigenetic modifications in shrimps. Notably, recent investigations have unveiled the potential of RNAi-mediated modifications in enhancing growth and development factors critical for mounting effective immune responses in shrimps. For instance, a study employed RNAi to silence the expression of m6A methyltransferase genes in L. vannamei, leading to diverse variations in DNA methylation patterns and subsequent alterations in molting-associated gene networks [184]. Moreover, applications of RNAi targeting immunity-related genes such as the Kruppel-like factor 2 (KLF2) gene, are implicated in the immune responses of L. vannamei [185]. Also, the silencing of Toll and IMD genes has been shown to impact immune response pathways and influence transcription levels of antimicrobial peptides (AMPs) in the same species [186]. Similarly, RNAi-mediated knockdown of the "LGBP" gene in shrimps has induced expression changes in the prophenoloxidase (proPO) system and AMP production [187]. While these studies do not explicitly convey the occurrence of heritable epigenetic changes, they highlight the potential of RNAi as a mediator for delivering immune gene-associated alterations, thus holding significance for future investigations of epigenetic modulation to develop disease management strategies.
The revolutionary CRISPR/Cas9 gene editing approach has emerged as a powerful method for genome manipulation in animals [188]. While not directly considered an epigenetic modulation method, CRISPR/Cas9 can indirectly introduce epigenetic changes by altering the DNA configuration. The CRISPR/Cas9 process entails the precise targeting of specific genes and the introduction of mutations and/or insertions/deletions that can induce epigenetic changes. Majority of the previous reports on shrimps focused on the CRISPR/Cas9 systems related to growth and reproduction. In contrast, the role of CRISPR/Cas9 targeting genes involved in the regulation of molting in the oriental prawn (Exopalaemon carinicauda), resulting in mutations in the chitinase gene (named as EcChl4) has led to alterations in chitin expression, which plays a pivotal role in the primary immune response of shrimp [189]. Conversely, CRISPR/Cas9 system-mediated mutation of the insulin-like peptide-encoding gene has impacted growth and development in various life stages, resulting in high mortality rates within the "gene knockout" populations in the same species [190]. Although the application of CRISPR/Cas9 technology is still in its nascent stages, these studies indicate the potential of employing it as a valuable tool for inducing subsequent epigenetic changes, particularly in the context of growth and development alterations in shrimps.
Withstanding these research findings, it is reasonable to posit that exploring the nature of epigenetics holds enormous potential for disease management in shrimp aquaculture and the augmentation of various facets of shrimp productivity. Manipulating certain epigenetic marks, such as DNA methylation, histone modifications, and non-coding RNAs, allows the alteration of gene expression patterns, thereby enhancing disease resistance and overall performance in shrimps. Furthermore, environmental factors, dietary interventions, and gene-centric methodologies can induce epigenetic changes that enhance shrimp health and optimize their physiological responses. Hence, unraveling the underlying mechanisms governing epigenetic regulation presents novel avenues for developing innovative strategies to revolutionize shrimp farming practices.
7. Challenges and Future Perspectives of Epigenetics in the Disease Management of Shrimp Aquaculture
The integration of epigenetic modulations in shrimp holds substantial promise, yet its practical implementation faces an array of intricate challenges and constraints. One primary hurdle lies in the demand for target-specific, finely-tuned, and precise epigenetic interventions, devoid of inadvertent off-target effects on other biological processes. Given the multi-genomic nature of shrimp genomes, similar to other organisms, implementing epigenetic modulations at a single unique site becomes intricate, as it may inadvertently influence neighboring regions, potentially yielding irreversible consequences for cellular functions [85,86,191]. Thus, the development of high-precision tools and methodologies capable of eliciting specific and localized epigenetic phenomena becomes paramount. Incorporating epigenetic modulations in shrimp aquaculture necessitates careful consideration of potential off-target effects, encompassing the emergence of drug resistance and perturbations in the gut microbiota composition, potentially instigating unforeseen consequences. Prolonged or recurrent interventions may be required to sustain the desired epigenetic changes in animals, engendering logistical challenges regarding practicality and implementation feasibility as one-time interventions. Moreover, the intricate interplay between shrimp physiology, metabolism, and gene expression patterns demands cautious evaluation, as epigenetic alterations could potentially perturb critical metabolic pathways essential for the animals' health and well-being [86,182,192]. Hence, achieving a comprehensive comprehension of the complex interactions between epigenetic modifications and gene expression is essential, as it will pave the way for delineating the implications of such changes and enable the development of targeted interventions that optimize shrimp health and performance. n addition to scientific considerations, ethical obligations take center stage when conducting research involving live animals. Ensuring the welfare of the animals involved is paramount, and researchers must adhere strictly to ethical guidelines and regulations to minimize any harm or distress inflicted upon the animals during epigenetic studies. A responsible and ethical approach underpins the pursuit of epigenetic advancements in shrimp aquaculture, signifying a commitment to the well-being of the animal subjects involved.
Despite the complex challenges encountered in implementing epigenetic modulations in shrimp aquaculture, the immense potential benefits warrant substantial attention. The profound understanding and manipulation of epigenetic changes offer viable avenues to augment diverse biological functions, including immune regulation, growth, and reproduction. Notably, epigenetic modulation emerges as a promising strategy to bolster disease resistance in shrimp by orchestrating immune pathways and augmenting immune responses against pathogens [22,91,92,182]. Targeted manipulation of epigenetic marks can enhance the expression of pivotal genes in immune pathways, fortifying the shrimp's capacity to combat infections and reducing disease incidence, thus reducing the dependency on antibiotics and chemical treatments. The identification of epigenetic markers associated with specific attributes and regulatory pathways assumes utmost significance, as it facilitates precise control of epigenetic changes to yield shrimp progenies characterized by heightened disease resistance and enhanced immunity [15,22,90]. Concurrently, the malleability of epigenetic changes in response to dietary components offers alternative prospects for nutritional programming in commercial shrimp aquaculture. Deliberate manipulation of the shrimp's diet to target nutrients involved in epigenetic regulations presents an opportunity to induce favorable changes in gene expression patterns related to growth and reproduction. In the long term, this approach holds the potential to foster sustainable and cost-effective shrimp performance while mitigating the demand for high-cost feeding materials, thereby contributing to the development of an economically viable shrimp industry. The fisheries sector has already witnessed such advantages of epigenetics, exemplified by studies in rainbow trouts that demonstrated higher feed efficiencies and growth rates resulting from epigenetic interventions [193,194]. Furthermore, the integration of epigenetics into selective breeding programs holds immense promise for genetic enhancement in shrimp populations. Through comprehension of the epigenetic regulation of desirable traits, the incorporation of epigenetic information into breeding strategies accelerates genetic improvement in shrimp populations. This avenue culminates in the production of superior strains endowed with enhanced disease resistance, growth rates, and other economically crucial traits. The potential transfer of epigenetic marks observed in a mass population study of Pacific oysters (C. gigas) may also offer further insights into the interplay of epigenetics and genetics in enhancing desirable traits [195]. Additionally, by discerning epigenetic changes instigated by environmental factors, the development of strategies to fortify the shrimp's resilience to adverse conditions and optimize their growth and survival becomes viable. Finally, comprehensive consideration of the ecological implications of epigenetic modulations in shrimp aquaculture is imperative to identify sustainable and environmentally friendly approaches that promote shrimp health and minimize negative impacts on surrounding ecosystems. The integration of epigenetics in shrimp aquaculture necessitates an interdisciplinary approach, fostering synergistic collaboration among researchers, biotechnologists, and ecologists to usher in a new era of innovative, sustainable, and eco-conscious shrimp farming practices.
8. Conclusion
In conclusion, the domain of epigenetics in shrimp aquaculture holds prodigious promise for disease management, optimized productivity, and the evolution of robust populations. The full realization of epigenetic modulation's potential necessitates the surmounting of multifaceted challenges through ingenious research, precision-driven tools, comprehensive monitoring, and seamless integration of epigenetic insights into breeding programs and aquaculture methodologies. Recent advancements in tailored feed formulations, nutraceuticals, pharmaceutical agents, and state-of-the-art genome-centric technologies such as RNA interference (RNAi) and CRISPR-Cas9 have unveiled captivating avenues for augmenting shrimp capabilities via targeted epigenetic interventions. Nonetheless, the field of epigenetics remains in its infancy, unveiling a plethora of unanswered inquiries into the mechanistic intricacies that call for exploration in the foreseeable future. The profound exploration of epigenetic frontiers bears the potential to catalyze the establishment of an immensely efficient and resilient shrimp farming industry, engendering a symbiotic alliance between industry prosperity and ecological stewardship.
Author Contributions
Conceptualization, C.W.R. and C.H.K; Writing—original draft preparation, C.W.R., W.S.P, and J.L; Writing—review and editing, C.W.R and C.H.K; Supervision, C.H.K.
Funding
This work was supported by a Research Grant from Pukyong National University (2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
All authors have read and agreed to the present version of the manuscript and declared no conflict of interest.
Ethical Statement
The present manuscript does not involve any experimental interventions and/or data collection that involves human subjects or animals, thus ethical approval is not necessary.
References
- The state of world fisheries and aquaculture 2022. State World Fish. Aquac. 2022 2022. [CrossRef]
- Araujo, G.S.; Silva, J.W.A. da; Cotas, J.; Pereira, L. Fish farming techniques: current situation and trends. J. Mar. Sci. Eng. 2022, 10. [Google Scholar] [CrossRef]
- Ahmed, N.; Azra, M.N. Aquaculture production and value chains in the COVID-19 pandemic. Curr. Environ. Heal. Reports 2022, 9, 423–435. [Google Scholar] [CrossRef]
- FAO The state of world fisheries and aquaculture 2020. Available from Http://Www.Fao.Org/3/Ca9229en/Ca9229en.Pdf. Accessed 25 Nov 2020. 2020, 1–244.
- Gunalan B, Nina Tabitha S. , S.P. and T.A. Nutritive value of cultured white leg shrimp Litopenaeus vannamei. 2013, 5, 166–171. [CrossRef]
- Shrimps – A nutritional perspective on JSTOR . Available online: https://www.jstor.org/stable/24092471 (accessed on 21 July 2023).
- Kulkarni, A.; Krishnan, S.; Anand, D.; Kokkattunivarthil Uthaman, S.; Otta, S.K.; Karunasagar, I.; Kooloth Valappil, R. Immune responses and immunoprotection in crustaceans with special reference to shrimp. Rev. Aquac. 2021, 13, 431–459. [Google Scholar] [CrossRef]
- Millard, R.S.; Ellis, R.P.; Bateman, K.S.; Bickley, L.K.; Tyler, C.R.; van Aerle, R.; Santos, E.M. How do abiotic environmental conditions influence shrimp susceptibility to disease? a critical analysis focussed on white spot disease. J. Invertebr. Pathol. 2021, 186, 107369. [Google Scholar] [CrossRef]
- Bank, T.W. Reducing disease risk in aquaculture. World Bank. Agric. Environ. Serv. 2014, 119. [Google Scholar]
- Yu, Y. Bin; Choi, J.H.; Kang, J.C.; Kim, H.J.; Kim, J.H. Shrimp bacterial and parasitic disease listed in the OIE: A review. Microb. Pathog. 2022, 166, 105545. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Steele, J.C.; Meng, X.Z. Usage, residue, and human health risk of antibiotics in chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef]
- Jiang, Y. The use of chemicals in aquaculture in the people’s republic of china. Use Chem. Aquac. Asia 1996, 141–153. [Google Scholar]
- Li, C.; Lin, N.; Feng, Z.; Lin, M.; Guan, B.; Chen, K.; Liang, W.; Wang, Q.; Li, M.; You, Y.; et al. CRISPR/Cas12a based rapid molecular detection of acute hepatopancreatic necrosis disease in shrimp. Front. Vet. Sci. 2022, 8, 1–10. [Google Scholar] [CrossRef]
- Sivakamavalli, J.; Park, K.; Kwak, I.; Baskaralingam, V. Bacterial disease control methods in shrimp ( penaeus, 1798) farming sector in asian countries. Arthropods - Are They Benef. Mankind? 2021. [CrossRef]
- Roy, S.; Kumar, V.; Bossier, P.; Norouzitallab, P.; Vanrompay, D. Phloroglucinol treatment induces transgenerational epigenetic inherited resistance against vibrio infections and thermal stress in a brine shrimp (Artemia franciscana) Model. Front. Immunol. 2019, 10, 1–25. [Google Scholar] [CrossRef]
- Granada, L.; Lemos, M.F.L.; Cabral, H.N.; Bossier, P.; Novais, S.C. Epigenetics in aquaculture – the last frontier. Rev. Aquac. 2018, 10, 994–1013. [Google Scholar] [CrossRef]
- Mazzone, R.; Zwergel, C.; Artico, M.; Taurone, S.; Ralli, M.; Greco, A.; Mai, A. The emerging role of epigenetics in human autoimmune disorders. Clin. Epigenetics 2019, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xue, Y. Emerging roles of non-coding RNAs in epigenetic regulation. Sci. China Life Sci. 2016, 59, 227–235. [Google Scholar] [CrossRef]
- Daxinger, L.; Whitelaw, E. Transgenerational epigenetic inheritance: more questions than answers. Genome Res. 2010, 20, 1623–1628. [Google Scholar] [CrossRef]
- Lacal, I.; Ventura, R. Epigenetic inheritance: concepts, mechanisms and perspectives. Front. Mol. Neurosci. 2018, 11, 1–22. [Google Scholar] [CrossRef]
- Aguilera, O.; Fernández, A.F.; Muñoz, A.; Fraga, M.F. Epigenetics and environment: a complex relationship. J. Appl. Physiol. 2010, 109, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Norouzitallab, P.; Baruah, K.; Vandegehuchte, M.; Van Stappen, G.; Catania, F.; Vanden Bussche, J.; Vanhaecke, L.; Sorgeloos, P.; Bossier, P. Environmental heat stress induces epigenetic transgenerational inheritance of robustness in parthenogenetic artemia model. FASEB J. 2014, 28, 3552–3563. [Google Scholar] [CrossRef]
- Manan, H.; Ikhwanuddin, M. Triploid induction in penaeid shrimps aquaculture: A Review. Rev. Aquac. 2021, 13, 619–631. [Google Scholar] [CrossRef]
- IMARC Group. (2020). Shrimp market: global industry trends, share, size, growth, opportunity and forecast 2020-2025 Available online: https://scholar.google.com/scholar_lookup?title=Shrimp market%3A Global industry trends%2C share%2C size%2C growth%2C opportunity and forecast 2020-2025&publication_year=2020&author=IMARC.
- Tabarestani, M.; Keithly, W.R.; Marzoughi-Ardakani, H. An analysis of the us shrimp market: a mixed demand approach. Mar. Resour. Econ. 2017, 32, 411–429. [Google Scholar] [CrossRef]
- Jory; Darryl, E. Current production, challenges and the future of shrimp farming. Glob. Aquac. Alliance 2018, 1.
- El-Saadony, M.T.; Swelum, A.A.; Abo Ghanima, M.M.; Shukry, M.; Omar, A.A.; Taha, A.E.; Salem, H.M.; El-Tahan, A.M.; El-Tarabily, K.A.; Abd El-Hack, M.E. Shrimp production, the most important diseases that threaten it, and the role of probiotics in confronting these diseases: A Review. Res. Vet. Sci. 2022, 144, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Wang, X.; Chen, K.; Xu, C.; Qin, J.G.; Chen, L. Physiological change and nutritional requirement of pacific white shrimp Litopenaeus vannamei at low salinity. Rev. Aquac. 2017, 9, 57–75. [Google Scholar] [CrossRef]
- Li, E.; Xu, C.; Wang, X.; Wang, S.; Zhao, Q.; Zhang, M.; Qin, J.G.; Chen, L. Gut microbiota and its modulation for healthy farming of pacific white shrimp Litopenaeus vannamei. Rev. Fish. Sci. Aquac. 2018, 26, 381–399. [Google Scholar] [CrossRef]
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Seibert, C.H.; Pinto, A.R. Challenges in shrimp aquaculture due to viral diseases: distribution and biology of the five major penaeid viruses and interventions to avoid viral incidence and dispersion. Brazilian J. Microbiol. 2012, 43, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Yu, Y. Bin; Choi, J.H.; Jo, A.H.; Hong, S.M.; Kang, J.C.; Kim, J.H. Viral shrimp diseases listed by the OIE: A Review. Viruses 2022, 14. [Google Scholar] [CrossRef]
- APEC Trans-Boundary Aquatic Animal Pathogen Transfer and the Development of Harmonized Standards on Aquaculture Health; 2001; ISBN 9747313278.
- Jones, B. Transboundary movement of shrimp viruses in crustaceans and their products: A special risk? J. Invertebr. Pathol. 2012, 110, 196–200. [Google Scholar] [CrossRef]
- Karunasagar, I.; Ababouch, L. Shrimp viral diseases, import risk assessment and international trade. Indian J. Virol. 2012, 23, 141–148. [Google Scholar] [CrossRef]
- Flegel, T.W.; Alday-Sanz, V. The crisis in asian shrimp aquaculture: Current status and future needs. J. Appl. Ichthyol. 1998, 14, 269–273. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Pantoja, C.R.; Poulos, B.T.; Redman, R.M.; Lightnere, D. V. In situ hybridization demonstrates that Litopenaeus vannamei, L. stylirostris and Penaeus monodon are susceptible to experimental infection with infectious myonecrosis virus (IMNV). Dis. Aquat. Organ. 2005, 63, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.P.D.; Cruz-Flores, R.; Mai, H.N.; Dhar, A.K. Novel infectious myonecrosis virus (IMNV) variant is associated with recent disease outbreaks in Penaeus vannamei shrimp in Brazil. Aquaculture 2022, 554, 738159. [Google Scholar] [CrossRef]
- Feijó, R.G.; Kamimura, M.T.; Oliveira-Neto, J.M.; Vila-Nova, C.M.V.M.; Gomes, A.C.S.; Coelho, M. das G.L.; Vasconcelos, R.F.; Gesteira, T.C. V.; Marins, L.F.; Maggioni, R. Infectious myonecrosis virus and white spot syndrome virus co-infection in pacific white shrimp (Litopenaeus vannamei) farmed in Brazil. Aquaculture 2013, 380–383, 1–5. [Google Scholar] [CrossRef]
- Gomez-Gil, B.; Soto-Rodríguez, S.; Lozano, R.; Betancourt-Lozano, M. Draft genome sequence of Vibrio parahaemolyticus strain M0605, which causes severe mortalities of shrimps in Mexico. Genome Announc. 2014, 2, 2104. [Google Scholar] [CrossRef]
- Lightner, D. V.; Ph, D. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Advocate 2012, 40, 2012. [Google Scholar]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D. V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Organ. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- FAO Report of the FAO/MARD Technical Workshop on Early Mortality Syndrome (EMS) or Acute Hepatopancreatic Necrosis Syndrome (AHPNS) of Cultured Shrimp (under TCP/VIE/3304); 2013; Vol. 1053; ISBN 9789251079041.
- Aranguren Caro, L.F.; Mai, H.N.; Noble, B.; Dhar, A.K. Acute hepatopancreatic necrosis disease (VPAHPND), a chronic disease in shrimp (Penaeus vannamei) population raised in Latin America. J. Invertebr. Pathol. 2020, 174. [Google Scholar] [CrossRef]
- Hossain, M.S.; Dai, J.; Qiu, D. Dysbiosis of the shrimp (Penaeus monodon) gut microbiome with AHPND outbreaks revealed by 16S rRNA metagenomics analysis. Aquac. Res. 2021, 52, 3336–3349. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut microbiota metagenomics in aquaculture: factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Rahi, M.L.; Azad, K.N.; Tabassum, M.; Irin, H.H.; Hossain, K.S.; Aziz, D.; Moshtaghi, A.; Hurwood, D.A. Effects of salinity on physiological, biochemical and gene expression parameters of black tiger shrimp (Penaeus monodon): potential for farming in low-salinity environments. Biology (Basel). 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, Q.; Shao, H.; Xu, Y.; Liu, P.; Li, J. Effects of low temperature on shrimp and crab physiology, behavior, and growth: A Review. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Kataoka, C.; Kashiwada, S. Ecological risks due to immunotoxicological effects on aquatic organisms. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Argue, B.B.J.; Tolentino, G.; Brock, J.A. Inbreeding cuts growth, reproduction in shrimp. 2018, 7–11.
- Doyle, R.W. Inbreeding and disease in tropical shrimp aquaculture: A reappraisal and caution. Aquac. Res. 2016, 47, 21–35. [Google Scholar] [CrossRef]
- Asche, F.; Anderson, J.L.; Botta, R.; Kumar, G.; Abrahamsen, E.B.; Nguyen, L.T.; Valderrama, D. The economics of shrimp disease. J. Invertebr. Pathol. 2021, 186, 107397. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Davies, J. Origins and evolution of antibiotic resistance. Microbiologia 1996, 12, 9–16. [Google Scholar] [CrossRef]
- Davis, R.P.; Davis, D.A.; Boyd, C.E. A Preliminary survey of antibiotic residues in frozen shrimp from retail stores in the United States. Curr. Res. Food Sci. 2021, 4, 679–683. [Google Scholar] [CrossRef]
- Le, T.X.; Munekage, Y. Residues of selected antibiotics in water and mud from shrimp ponds in mangrove areas in Vietnam. Mar. Pollut. Bull. 2004, 49, 922–929. [Google Scholar] [CrossRef]
- Sharma, L.; Nagpal, R.; Jackson, C.R.; Patel, D.; Singh, P. Antibiotic-resistant bacteria and gut microbiome communities associated with wild-caught shrimp from the United States versus imported farm-raised retail shrimp. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Thornber, K.; Verner-Jeffreys, D.; Hinchliffe, S.; Rahman, M.M.; Bass, D.; Tyler, C.R. Evaluating antimicrobial resistance in the global shrimp industry. Rev. Aquac. 2020, 12, 966–986. [Google Scholar] [CrossRef]
- Sudheesh, P.S.; Cain, K.D. Prospects and challenges of developing and commercializing immersion vaccines for aquaculture. Int. Biol. Rev. 2017, 1, 1–20. [Google Scholar]
- Gudding, R.; Van Muiswinkel, W.B. A History of fish vaccination: Science-based disease prevention in aquaculture. Fish Shellfish Immunol. 2013, 35, 1683–1688. [Google Scholar] [CrossRef]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7. [Google Scholar] [CrossRef]
- Rout, N.; Kumar, S.; Jaganmohan, S.; Murugan, V. DNA vaccines encoding viral envelope proteins confer protective immunity against WSSV in black tiger shrimp. Vaccine 2007, 25, 2778–2786. [Google Scholar] [CrossRef]
- Rajeshkumar, S.; Venkatesan, C.; Sarathi, M.; Sarathbabu, V.; Thomas, J.; Anver Basha, K.; Sahul Hameed, A.S. Oral delivery of DNA construct using chitosan nanoparticles to protect the shrimp from white spot syndrome virus (WSSV). Fish Shellfish Immunol. 2009, 26, 429–437. [Google Scholar] [CrossRef]
- Mavichak, R.; Takano, T.; Kondo, H.; Hirono, I.; Wada, S.; Hatai, K.; Inagawa, H.; Takahashi, Y.; Yoshimura, T.; Kiyono, H.; et al. The effect of liposome-coated recombinant protein VP28 against white spot syndrome virus in kuruma shrimp, Marsupenaeus Japonicus. J. Fish Dis. 2010, 33, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.K.; Gopal, C.; Panigrahi, A.; Rajababu, D.; Pillai, S.M. Oral administration of formalin killed Vibrio anguillarum cells improves growth and protection against challenge with Vibrio harveyi in banana shrimp. Lett. Appl. Microbiol. 2014, 58, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Vivas, J.; Superio, J.; Galindo-Villegas, J.; Acosta, F. Phage therapy as a focused management strategy in aquaculture. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Sillankorva, S.M.; Oliveira, H.; Azeredo, J. Bacteriophages and their role in food safety. Int. J. Microbiol. 2012, 2012. [Google Scholar] [CrossRef]
- Jun, J.W.; Han, J.E.; Giri, S.S.; Tang, K.F.J.; Zhou, X.; Aranguren, L.F.; Kim, H.J.; Yun, S.; Chi, C.; Kim, S.G.; et al. Phage application for the protection from acute hepatopancreatic necrosis disease (AHPND) in Penaeus vannamei. Indian J. Microbiol. 2018, 58, 114–117. [Google Scholar] [CrossRef]
- Quiroz-Guzmán, E.; Peña-Rodriguez, A.; Vázquez-Juárez, R.; Barajas-Sandoval, D.R.; Balcázar, J.L.; Martínez-Díaz, S.F. Bacteriophage cocktails as an environmentally-friendly approach to prevent Vibrio parahaemolyticus and Vibrio harveyi infections in brine shrimp (Artemia franciscana) production. Aquaculture 2018, 492, 273–279. [Google Scholar] [CrossRef]
- Yin, Y.; Ni, P.; Liu, D.; Yang, S.; Almeida, A.; Guo, Q.; Zhang, Z.; Deng, L.; Wang, D. Bacteriophage potential against Vibrio parahaemolyticus biofilms. Food Control 2019, 98, 156–163. [Google Scholar] [CrossRef]
- Borodovich, T.; Shkoporov, A.N.; Ross, R.P.; Hill, C. Phage-mediated horizontal gene transfer and its implications for the human gut microbiome. Gastroenterol. Rep. 2022, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, F. Resistance development to bacteriophages occurring during bacteriophage therapy. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Skurnik, M.; Strauch, E. Phage therapy: Facts and fiction. Int. J. Med. Microbiol. 2006, 296, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- Declarador, R.S.; Serrano, A.E.; Corre, V.L. Ulvan extract acts as immunostimulant against white spot syndrome virus (WSSV) in juvenile black tiger shrimp Penaeus monodon. AACL Bioflux 2014, 7, 153–161. [Google Scholar]
- Citarasu, T. Herbal biomedicines: A new opportunity for aquaculture industry. Aquac. Int. 2010, 18, 403–414. [Google Scholar] [CrossRef]
- Anirudhan, A.; Tosin, O.V.; Wahid, M.; Effendy; Sung, Y. Y. Alternative disease control methods in shrimp aquaculture: A Review. J. Aquac. Res. Dev. 2021, 12, 12–15. [Google Scholar]
- Yu, P.; Wang, T.; Ye, H.; Shan, H.; Ma, S. Isolation and identification of pathogenic vibrio spp. retrieved from diseased Litopenaeus vannamei and beneficial role of some functional probiotic bacteria for control. Aquac. Int. 2020, 28, 1403–1420. [Google Scholar] [CrossRef]
- Khademzade, O.; Zakeri, M.; Haghi, M.; Mousavi, S.M. The effects of water additive Bacillus cereus and Pediococcus acidilactici on water quality, growth performances, economic benefits, immunohematology and bacterial flora of whiteleg shrimp (Penaeus vannamei boone, 1931) reared in earthen ponds. Aquac. Res. 2020, 51, 1759–1770. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Dai, X.; Li, J.; Ding, F. Effects of dietary inulin and mannan oligosaccharide on immune related genes expression and disease resistance of pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 76, 78–92. [Google Scholar] [CrossRef]
- Rungrassamee, W.; Kingcha, Y.; Srimarut, Y.; Maibunkaew, S.; Karoonuthaisiri, N.; Visessanguan, W. Mannooligosaccharides from copra meal improves survival of the pacific white shrimp (Litopenaeus vannamei) after exposure to Vibrio harveyi. Aquaculture 2014, 434, 403–410. [Google Scholar] [CrossRef]
- Huynh, T.G.; Cheng, A.C.; Chi, C.C.; Chiu, K.H.; Liu, C.H. A synbiotic improves the immunity of white shrimp, Litopenaeus vannamei: metabolomic analysis reveal compelling evidence. Fish Shellfish Immunol. 2018, 79, 284–293. [Google Scholar] [CrossRef]
- Roy, S.; Kumar, V.; Behera, B.K.; Das, B.K. Epigenetics: perspectives and potential in aquaculture. Adv. Fish. Biotechnol. 2022, 133–150. [Google Scholar]
- Liu, Z.; Zhou, T.; Gao, D. Genetic and epigenetic regulation of growth, reproduction, disease resistance and stress responses in aquaculture. Front. Genet. 2022, 13, 1–22. [Google Scholar] [CrossRef]
- Jablonka, E.; Lamb, M.J. The changing concept of epigenetics. Ann. N. Y. Acad. Sci. 2002, 981, 82–96. [Google Scholar] [CrossRef]
- Brander, S.M.; Biales, A.D.; Connon, R.E. The role of epigenomics in aquatic toxicology. Environ. Toxicol. Chem. 2017, 36, 2565–2573. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Yang, J.J.; Shi, K.H. Non-coding RNAs as direct and indirect modulators of epigenetic mechanism regulation of cardiac fibrosis. Expert Opin. Ther. Targets 2015, 19, 707–716. [Google Scholar] [CrossRef]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef]
- Norouzitallab, P.; Biswas, P.; Baruah, K.; Bossier, P. Multigenerational immune priming in an invertebrate parthenogenetic artemia to a pathogenic Vibrio campbellii. Fish Shellfish Immunol. 2015, 42, 426–429. [Google Scholar] [CrossRef]
- Norouzitallab, P.; Baruah, K.; Biswas, P.; Vanrompay, D.; Bossier, P. Probing the phenomenon of trained immunity in invertebrates during a transgenerational study, using brine shrimp artemia as a model system. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Jin, B.; Li, Y.; Robertson, K.D. DNA methylation: superior or subordinate in the epigenetic hierarchy? Genes and Cancer 2011, 2, 607–617. [Google Scholar] [CrossRef]
- Miremadi, A.; Oestergaard, M.Z.; Pharoah, P.D.P.; Caldas, C. Cancer genetics of epigenetic genes. Hum. Mol. Genet. 2007, 16. [Google Scholar] [CrossRef]
- Okano, M.; Xie, S.; Li, E. Cloning and characterization of a family of novel mammalian DNA ( Cytosine-5 ) methyltransferases non-invasive sexing of preimplantation stage mammalian embryos. Nat. Am. Inc. 1998, 19, 219–220. [Google Scholar]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, J.; Scheer, O.; Stadler, P.F.; Prohaska, S.J. Evolution of DNA methylation across ecdysozoa. J. Mol. Evol. 2022, 90, 56–72. [Google Scholar] [CrossRef]
- Lee, H.J.; Lowdon, R.F.; Maricque, B.; Zhang, B.; Stevens, M.; Li, D.; Johnson, S.L.; Wang, T. Developmental enhancers revealed by extensive DNA methylome maps of zebrafish early embryos. Nat. Commun. 2015 61 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Rauscher, G.H.; Kresovich, J.K.; Poulin, M.; Yan, L.; Macias, V.; Mahmoud, A.M.; Al-Alem, U.; Kajdacsy-Balla, A.; Wiley, E.L.; Tonetti, D.; et al. Exploring DNA methylation changes in promoter, intragenic, and intergenic regions as early and late events in breast cancer formation. BMC Cancer 2015, 15, 1–15. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA methylation: A historical perspective. Trends Genet. 2022, 38, 676–707. [Google Scholar] [CrossRef]
- Klughammer, J.; Romanovskaia, D.; Nemc, A.; Posautz, A.; Seid, C.A.; Schuster, L.C.; Keinath, M.C.; Lugo Ramos, J.S.; Kosack, L.; Evankow, A.; et al. Comparative analysis of genome-scale, base-resolution DNA methylation profiles across 580 animal species. Nat. Commun. 2023, 14. [Google Scholar] [CrossRef]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Nery, J.R.; Lee, L.; Ye, Z.; Ngo, Q.M.; et al. Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef]
- Hon, G.C.; Rajagopal, N.; Shen, Y.; McCleary, D.F.; Yue, F.; Dang, M.D.; Ren, B. Epigenetic memory at embryonic enhancers identified in DNA methylation maps from adult mouse tissues. Nat. Genet. 2013, 45, 1198–1206. [Google Scholar] [CrossRef]
- Marhold, J.; Kramer, K.; Kremmer, E.; Lyko, F. The drosophila MBD2/3 protein mediates interactions between the MI-2 chromatin complex and CpT/A-methylated DNA. Development 2004, 131, 6033–6039. [Google Scholar] [CrossRef]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef]
- Yang, X.; Han, H.; DeCarvalho, D.D.; Lay, F.D.; Jones, P.A.; Liang, G. Gene body methylation can alter gene expression and is a therapeutic target in cancer. Cancer Cell 2014, 26, 577–590. [Google Scholar] [CrossRef]
- Shayevitch, R.; Askayo, D.; Keydar, I.; Ast, G. The importance of DNA methylation of exons on alternative splicing. Rna 2018, 24, 1351–1362. [Google Scholar] [CrossRef]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef]
- Sukthaworn, S.; Panyim, S.; Udomkit, A. Homologues of piwi control transposable elements and development of male germline in Penaeus monodon. Comp. Biochem. Physiol. -Part A Mol. Integr. Physiol. 2020, 250, 110807. [Google Scholar] [CrossRef]
- Li, B. Bin; Fan, J.Q.; Lu, K.C.; Chen, G.L.; Chen, Y.H. Identification and functional characterization of a systemic RNA interference defective 1 gene in Litopenaeus vannamei. Fish Shellfish Immunol. Reports 2021, 2, 100033. [Google Scholar] [CrossRef]
- Shilatifard, A. Chromatin modifications by methylation and ubiquitination: Implications in the regulation of gene expression. Annu. Rev. Biochem. 2006, 75, 243–269. [Google Scholar] [CrossRef]
- Marshall, O.J.; Brand, A.H. Chromatin state changes during neural development revealed by in vivo cell-type specific profiling. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Henikoff, S.; Shilatifard, A. Histone modification: Cause or cog? Trends Genet. 2011, 27, 389–396. [Google Scholar] [CrossRef]
- Turner, B.M.; Birley, A.J.; Lavender, J. Histone H4 isoforms acetylated at specific lysine residues define individual chromosomes and chromatin domains in drosophila polytene nuclei. Cell 1992, 69, 375–384. [Google Scholar] [CrossRef]
- Felsenfeld, G.; Groudine, M. Controlling the double helix. Nat. 2003 4216921 2003, 421, 448–453. [Google Scholar] [CrossRef]
- Duman, M.; Martinez-Moreno, M.; Jacob, C.; Tapinos, N. Functions of histone modifications and histone modifiers in schwann cells. Glia 2020, 68, 1584–1595. [Google Scholar] [CrossRef]
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Xiong, L.; Wang, Y. Mapping post-translational modifications of histones H2A, H2B and H4 in Schizosaccharomyces pombe. Int. J. Mass Spectrom. 2011, 301, 159. [Google Scholar] [CrossRef]
- Karmodiya, K.; Anamika, K.; Muley, V.; Pradhan, S.J.; Bhide, Y.; Galande, S. Camello, a novel family of histone acetyltransferases that acetylate histone H4 and is essential for zebrafish development. Sci. Rep. 2014, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, S.; Liao, Y.; Deng, Y.; Jiao, Y. Increased histone H3 acetylation inhibit the inflammatory response and activate the serum immunity of pearl oyster Pinctada fucata martensii. Front. Mar. Sci. 2023, 10, 1–11. [Google Scholar] [CrossRef]
- Chatterjee, N.; Sinha, D.; Lemma-Dechassa, M.; Tan, S.; Shogren-Knaak, M.A.; Bartholomew, B. Histone H3 Tail acetylation modulates ATP-dependent remodeling through multiple mechanisms. Nucleic Acids Res. 2011, 39, 8378–8391. [Google Scholar] [CrossRef] [PubMed]
- Wakamori, M.; Okabe, K.; Ura, K.; Funatsu, T.; Takinoue, M.; Umehara, T. Quantification of the effect of site-specific histone acetylation on chromatin transcription rate. Nucleic Acids Res. 2020, 48, 12648–12659. [Google Scholar] [CrossRef]
- Patat, S.A.; Carnegie, R.B.; Kingsbury, C.; Gross, P.S.; Chapman, R.; Schey, K.L. Antimicrobial activity of histones from hemocytes of the pacific white shrimp. Eur. J. Biochem. 2004, 271, 4825–4833. [Google Scholar] [CrossRef]
- Jambhekar, A.; Dhall, A.; Shi, Y. Roles and regulation of histone methylation in animal development. Nat. Rev. Mol. Cell Biol. 2019 2010 2019, 20, 625–641. [Google Scholar] [CrossRef]
- Lauberth, S.M.; Nakayama, T.; Wu, X.; Ferris, A.L.; Tang, Z.; Hughes, S.H.; Roeder, R.G. H3K4me3 interactions with TAF3 regulate preinitiation complex assembly and selective gene activation. Cell 2013, 152, 1021–1036. [Google Scholar] [CrossRef]
- Gillies, S.D.; Morrison, S.L.; Oi, V.T.; Tonegawa, S. A tissue-specific transcription enhancer element is located in the major intron of a rearranged immunoglobulin heavy chain gene. Cell 1983, 33, 717–728. [Google Scholar] [CrossRef]
- D’Urso, A.; Brickner, J.H. Epigenetic transcriptional memory. Curr. Genet. 2017, 63, 435. [Google Scholar] [CrossRef]
- Zang, S.; Lv, L.X.; Liu, C.F.; Zhang, P.; Li, C.; Wang, J.X. Metabolomic investigation of ultraviolet ray-inactivated white spot syndrome virus-induced trained immunity in Marsupenaeus japonicus. Front. Immunol. 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Baarends, W.M.; Hoogerbrugge, J.W.; Roest, H.P.; Ooms, M.; Vreeburg, J.; Hoeijmakers, J.H.J.; Grootegoed, J.A. Histone ubiquitination and chromatin remodeling in mouse spermatogenesis. Dev. Biol. 1999, 207, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Boros, I.M. Histone modification in drosophila. Brief. Funct. Genomics 2012, 11, 319–331. [Google Scholar] [CrossRef]
- Gonzalez-Romero, R.; Suarez-Ulloa, V.; Rodriguez-Casariego, J.; Garcia-Souto, D.; Diaz, G.; Smith, A.; Pasantes, J.J.; Rand, G.; Eirin-Lopez, J.M. Effects of florida red tides on histone variant expression and DNA methylation in the eastern oyster Crassostrea virginica. Aquat. Toxicol. 2017, 186, 196–204. [Google Scholar] [CrossRef]
- Teperek-Tkacz, M.; Meglicki, M.; Pasternak, M.; Kubiak, J.Z.; Borsuk, E. Phosphorylation of histone H3 serine 10 in early mouse embryos: active phosphorylation at late s phase and differential effects of ZM447439 on first two embryonic mitoses. Cell Cycle 2010, 9, 4674–4687. [Google Scholar] [CrossRef]
- Kellner, W.A.; Ramos, E.; Van Bortle, K.; Takenaka, N.; Corces, V.G. Genome-wide phosphoacetylation of histone H3 at drosophila enhancers and promoters. Genome Res. 2012, 22, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.; Jin, Y.; Johansen, J.; Johansen, K.M. The JIL-1 tandem kinase mediates histone H3 phosphorylation and is required for maintenance of chromatin structure in drosophila. Cell 2001, 105, 433–443. [Google Scholar] [CrossRef]
- Xu, Z.; Song, Z.; Li, G.; Tu, H.; Liu, W.; Liu, Y.; Wang, P.; Wang, Y.; Cui, X.; Liu, C.; et al. H2B ubiquitination regulates meiotic recombination by promoting chromatin relaxation. Nucleic Acids Res. 2016, 44, 9681–9697. [Google Scholar] [CrossRef]
- Wang, H.; Wang, L.; Erdjument-Bromage, H.; Vidal, M.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of histone H2A ubiquitination in polycomb silencing. Nat. 2004 4317010 2004, 431, 873–878. [Google Scholar] [CrossRef]
- Buszczak, M.; Paterno, S.; Spradling, A.C. Drosophila stem cells share a common requirement for the histone H2B ubiquitin protease scrawny. Science 2009, 323, 248. [Google Scholar] [CrossRef]
- Chen, T.; Sun, Z.; Mu, S.; Jiang, L.; Li, C.; Li, L.; Guo, M.; Zhang, Z.; Kang, X. Ultrastructure of spermiogenesis and the distribution of spermatozoal nuclear histones in the japanese mantis shrimp, Oratosquilla oratoria (crustacea: stomatopoda). J. Morphol. 2019, 280, 1170–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Mu, S.M.; Guo, M.S.; Wu, J.L.; Li, Y.Q.; Zhang, H.; Wang, Y.; Kang, X.J. Dynamics of histone H2A, H4 and HS1ph during spermatogenesis with a focus on chromatin condensation and maturity of spermatozoa. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Li, R.; Aweya, J.J.; Yao, D.; Wang, F.; Li, S.; Tuan, T.N.; Zhang, Y. The PirB toxin protein from Vibrio parahaemolyticus induces apoptosis in hemocytes of Penaeus vannamei : PirB induces shrimp hemocytes apoptosis. Virulence 2021, 12, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, T.; Li, X.; Sheng, X.; Xing, J.; Chi, H.; Zhan, W. Protein phosphorylation in hemocytes of fenneropenaeus chinensis in response to white spot syndrome virus infection. Fish Shellfish Immunol. 2022, 122, 106–114. [Google Scholar] [CrossRef]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution - trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Ponjavic, J.; Ponting, C.P.; Lunter, G. Functionality or transcriptional noise? evidence for selection within long noncoding RNAs. Genome Res. 2007, 17, 556–565. [Google Scholar] [CrossRef]
- Zhou, Z.; Leng, C.; Wang, Z.; Long, L.; Lv, Y.; Gao, Z.; Wang, Y.; Wang, S.; Li, P. The potential regulatory role of the LncRNA-MiRNA-MRNA axis in teleost fish. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Cecere, G. Small RNAs in epigenetic inheritance: From mechanisms to trait transmission. FEBS Lett. 2021, 595, 2953–2977. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Ulitsky, I.; Bartel, D.P. LincRNAs: genomics, evolution, and mechanisms. Cell, 2013; 154, 26–46. [Google Scholar] [CrossRef]
- Ying, S.Y.; Chang, D.C.; Lin, S.L. The microRNA (MiRNA): Overview of the RNA genes that modulate gene function. Mol. Biotechnol. 2008, 38, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.; Bardhan, A.; Darrah, K.; Tsang, M.; Deiters, A. Optical control of microRNA function in zebrafish embryos. J. Am. Chem. Soc. 2022, 144, 16819–16826. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, Y.; Guo, M.; Guo, M.; Yue, D.; Yue, D.; Chen, C.; Chen, C.; Liang, G.; Liang, G.; et al. MicroRNA-7: Expression and function in brain physiological and pathological processes. Cell Biosci. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Paladini, L.; Fabris, L.; Bottai, G.; Raschioni, C.; Calin, G.A.; Santarpia, L. Targeting microRNAs as key modulators of tumor immune response. J. Exp. Clin. Cancer Res. 2016, 35. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, Z.; Wang, H.; Wang, L.; Wang, W.; Liu, R.; Qiu, L.; Song, L. An invertebrate-specific and immune-responsive microRNA augments oyster haemocyte phagocytosis by targeting CgIκ B2. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Ruan, L.; Bian, X.; Ji, Y.; Li, M.; Li, F.; Yan, X. Isolation and identification of novel microRNAs from Marsupenaeus japonicus. Fish Shellfish Immunol. 2011, 31, 334–340. [Google Scholar] [CrossRef]
- Kaewkascholkul, N.; Somboonviwat, K.; Asakawa, S.; Hirono, I.; Tassanakajon, A.; Somboonwiwat, K. Shrimp MiRNAs regulate innate immune response against white spot syndrome virus infection. Dev. Comp. Immunol. 2016, 60, 191–201. [Google Scholar] [CrossRef]
- Zhu, F.; Wang, Z.; Sun, B.Z. Differential expression of microRNAs in shrimp Marsupenaeus japonicus in response to Vibrio alginolyticus infection. Dev. Comp. Immunol. 2016, 55, 76–79. [Google Scholar] [CrossRef]
- Zheng, Z.; Aweya, J.J.; Wang, F.; Yao, D.; Lun, J.; Li, S.; Ma, H.; Zhang, Y. Acute hepatopancreatic necrosis disease (AHPND) related microRNAs in Litopenaeus vannamei infected with AHPND-causing strain of Vibrio parahemolyticus. BMC Genomics 2018, 19, 1–11. [Google Scholar] [CrossRef]
- Zheng, J.; Cao, J.; Mao, Y.; Su, Y.; Wang, J. Identification of microRNAs with heat stress responsive and immune properties in Marsupenaeus japonicus based on next-generation sequencing and bioinformatics analysis: Essential regulators in the heat stress-host interactions. Fish Shellfish Immunol. 2018, 81, 390–398. [Google Scholar] [CrossRef]
- Guo, H.; Lu, Z. cheng; Zhu, X. wen; Zhu, C. hua; Wang, C. gui; Shen, Y. chun; Wang, W. Differential expression of microRNAs in hemocytes from white shrimp Litopenaeus vannamei under copper stress. Fish Shellfish Immunol. 2018, 74, 152–161. [Google Scholar] [CrossRef]
- Iwakawa, H. oki; Tomari, Y. Life of RISC: Formation, action, and degradation of RNA-induced silencing complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N. RNA interference in functional genomics and medicine. J. Korean Med. Sci. 2003, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Han, F.; Zhang, X. Silencing shrimp white spot syndrome virus (WSSV) genes by SiRNA. Antiviral Res. 2007, 73, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Story, B.; Ma, X.; Ishihara, K.; Li, H.; Hall, K.; Peak, A.; Anoja, P.; Park, J.; Haug, J.; Blanchette, M.; et al. Defining the expression of PiRNA and transposable elements in drosophila ovarian germline stem cells and somatic support cells. Life Sci. Alliance 2019, 2, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.S.; Chuma, S. PiRNAs and their involvement in male germline development in mice. Dev. Growth Differ. 2012, 54, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Kofler, R. Dynamics of transposable element invasions with PiRNA clusters. Mol. Biol. Evol. 2019, 36, 1457–1472. [Google Scholar] [CrossRef]
- Le Thomas, A.; Rogers, A.K.; Webster, A.; Marinov, G.K.; Liao, S.E.; Perkins, E.M.; Hur, J.K.; Aravin, A.A.; Tóth, K.F. Piwi induces PiRNA-guided transcriptional silencing and establishment of a repressive chromatin state. Genes Dev. 2013, 27, 390–399. [Google Scholar] [CrossRef]
- Lewis, S.H.; Quarles, K.A.; Yang, Y.; Tanguy, M.; Frézal, L.; Smith, S.A.; Sharma, P.P.; Cordaux, R.; Gilbert, C.; Giraud, I.; et al. Pan-arthropod analysis reveals somatic PiRNAs as an ancestral defence against transposable elements. Nat. Ecol. Evol. 2017 21 2017, 2, 174–181. [Google Scholar] [CrossRef]
- Waiho, K.; Fazhan, H.; Zhang, Y.; Li, S.; Zhang, Y.; Zheng, H.; Ikhwanuddin, M.; Ma, H. Comparative profiling of ovarian and testicular PiRNAs in the mud crab Scylla paramamosain. Genomics 2020, 112, 323–331. [Google Scholar] [CrossRef]
- Jehn, J.; Gebert, D.; Pipilescu, F.; Stern, S.; Kiefer, J.S.T.; Hewel, C.; Rosenkranz, D. PIWI genes and PiRNAs are ubiquitously expressed in mollusks and show patterns of lineage-specific adaptation. Commun. Biol. 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- ter Horst, A.M.; Nigg, J.C.; Dekker, F.M.; Falk, B.W. Endogenous viral elements are widespread in arthropod genomes and commonly give rise to piwi-interacting RNAs. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Huang, S.; Yoshitake, K.; Asakawa, S. A Review of discovery profiling of piwi-interacting RNAs and their diverse functions in metazoans. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Huerlimann, R.; Wade, N.M.; Gordon, L.; Montenegro, J.D.; Goodall, J.; McWilliam, S.; Tinning, M.; Siemering, K.; Giardina, E.; Donovan, D.; et al. De novo assembly, characterization, functional annotation and expression patterns of the black tiger shrimp (Penaeus monodon) transcriptome. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Liu, D.; Hong, Z.; Gui, L.; Zhao, L.; Wang, Y.; Sun, S.; Li, M. Full-length transcriptomes and sex-based differentially expressed genes in the brain and ganglia of giant river prawn Macrobrachium rosenbergii. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Taneerat, C.; Olasard, P.; Suksri, P.; Whankaew, S.; Sathapondecha, P. Identification and profiling of long non-coding RNAs during molt cycle: An involvement of Lnc1182 in the molt of white shrimp, Litopenaeus vannamei. Aquac. Reports 2023, 30, 101611. [Google Scholar] [CrossRef]
- MacHado, M.; Fernández-Boo, S.; Teixeira, C.; Viegas, M.; Serradeiro, R.; Dias, J.; Costas, B.; Masagounder, K. DL-methionyl-DL-methionine as an efficient methionine source for promoting zootechnical performance and methionine-related pathways in the whiteleg shrimp (Penaeus vannamei). Br. J. Nutr. 2023. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, Y.; Zhang, Y.; Xu, B.; Sagada, G.; Wang, Z.; Chen, C.; Lang, X.; Zhang, J.; Shao, Q. Comparative study on the effects of crystalline L-methionine and methionine hydroxy analogue calcium supplementations in the diet of juvenile pacific white shrimp (Litopenaeus vannamei). Front. Physiol. 2023, 14, 1–13. [Google Scholar] [CrossRef]
- Asaikkutti, A.; Bhavan, P.S.; Vimala, K. Effects of different levels of dietary folic acid on the growth performance, muscle composition, immune response and antioxidant capacity of freshwater prawn, Macrobrachium rosenbergii. Aquaculture 2016, 464, 136–144. [Google Scholar] [CrossRef]
- Marçal, R.; Llorente, L.; Herrero, O.; Planelló, R.; Guilherme, S.; Pacheco, M. Intergenerational patterns of DNA methylation in Procambarus clarkii following exposure to genotoxicants: a conjugation in past simple or past continuous? Toxics 2021, 9. [Google Scholar] [CrossRef]
- Mirbahai, L.; Chipman, J.K. Epigenetic memory of environmental organisms: A reflection of lifetime stressor exposures. Mutat. Res. - Genet. Toxicol. Environ. Mutagen. 2014, 764–765, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Šrut, M. Ecotoxicological epigenetics in invertebrates: Emerging tool for the evaluation of present and past pollution burden. Chemosphere 2021, 282. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, J.; Su, Y.; Liu, Z.; Mao, Y. Air exposure affects physiological responses, innate immunity, apoptosis and DNA methylation of kuruma shrimp, Marsupenaeus japonicus. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, N.; Dasaradhi, P.V.N.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA interference: Biology, mechanism, and applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657–685. [Google Scholar] [CrossRef]
- Lei, Y.; Zeng, Q.; Tan, G.; Yuan, Z.; Zhang, N.; Liu, J.; Wang, W. A first glimpse into the M6A modification machinery of shrimp: Genomic features, expression patterns and potential roles in molting regulation. Aquac. Reports 2023, 29, 101493. [Google Scholar] [CrossRef]
- Yang, P.; Aweya, J.J.; Yao, D.; Wang, F.; Lun, J.; Hong, Y.; Sun, K.; Zhang, Y. The Krüppel-like factor of Penaeus vannamei negatively regulates transcription of the small subunit hemocyanin gene as part of shrimp immune response. Fish Shellfish Immunol. 2020, 100, 397–406. [Google Scholar] [CrossRef]
- Hou, F.; He, S.; Liu, Y.; Zhu, X.; Sun, C.; Liu, X. RNAi knock-down of shrimp Litopenaeus vannamei toll gene and immune deficiency gene reveals their difference in regulating antimicrobial peptides transcription. Dev. Comp. Immunol. 2014, 44, 255–260. [Google Scholar] [CrossRef]
- Amparyup, P.; Sutthangkul, J.; Charoensapsri, W.; Tassanakajon, A. Pattern recognition protein binds to lipopolysaccharide and β-1,3-glucan and activates shrimp prophenoloxidase system. J. Biol. Chem. 2012, 287, 10060–10069. [Google Scholar] [CrossRef]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-Guided Cas9 nuclease in zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.; Xiang, J. A CRISPR/Cas9-mediated mutation in chitinase changes immune response to bacteria in Exopalaemon carinicauda. Fish Shellfish Immunol. 2017, 71, 43–49. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, X.; Yuan, J.; Zhang, C.; Li, S.; Li, F. CRISPR/Cas9-mediated mutation on an insulin-like peptide encoding gene affects the growth of the ridgetail white prawn Exopalaemon carinicauda. Front. Endocrinol. (Lausanne). 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Gavery, M.R.; Roberts, S.B. Epigenetic considerations in aquaculture. PeerJ 2017, 2017. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, H.; Han, D.; Ren, L.; Gong, X.; Jiang, F.; Cui, Y.; Liu, X.; Ren, C.; Xue, J.; et al. Metabolomic alterations in the digestive system of the mantis shrimp Oratosquilla oratoria following short-term exposure to cadmium. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- Balasubramanian, M.N.; Panserat, S.; Dupont-Nivet, M.; Quillet, E.; Montfort, J.; Le Cam, A.; Medale, F.; Kaushik, S.J.; Geurden, I. Molecular pathways associated with the nutritional programming of plant-based diet acceptance in rainbow trout following an early feeding exposure. BMC Genomics 2016, 17, 1–20. [Google Scholar] [CrossRef]
- Panserat, S.; Marandel, L.; Geurden, I.; Veron, V.; Dias, K.; Plagnes-Juan, E.; Pegourié, G.; Arbenoits, E.; Santigosa, E.; Weber, G.; et al. Muscle catabolic capacities and global hepatic epigenome are modified in juvenile rainbow trout fed different vitamin levels at first feeding. Aquaculture 2017, 468, 515–523. [Google Scholar] [CrossRef]
- Jiang, Q.; Li, Q.; Yu, H.; Kong, L.F. Genetic and epigenetic variation in mass selection populations of pacific oyster Crassostrea gigas. Genes and Genomics 2013, 35, 641–647. [Google Scholar] [CrossRef]
Figure 1.
Global shrimp production (in metric tons) trend during 1990-2022 [1].
Figure 1.
Global shrimp production (in metric tons) trend during 1990-2022 [1].
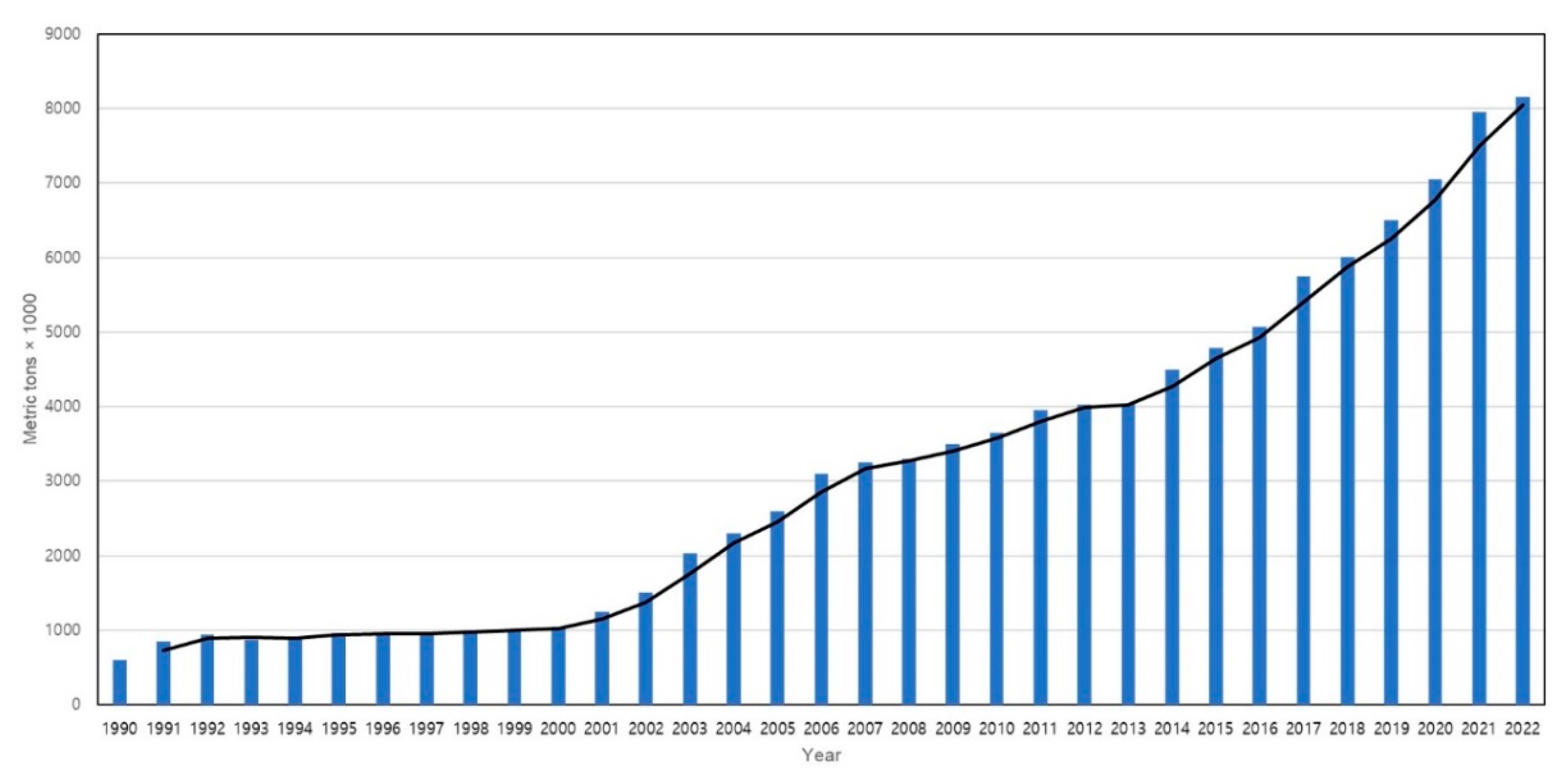
Figure 2.
Global shift in production (in percentage) from black tiger shrimp to white-leg shrimp [1].
Figure 2.
Global shift in production (in percentage) from black tiger shrimp to white-leg shrimp [1].
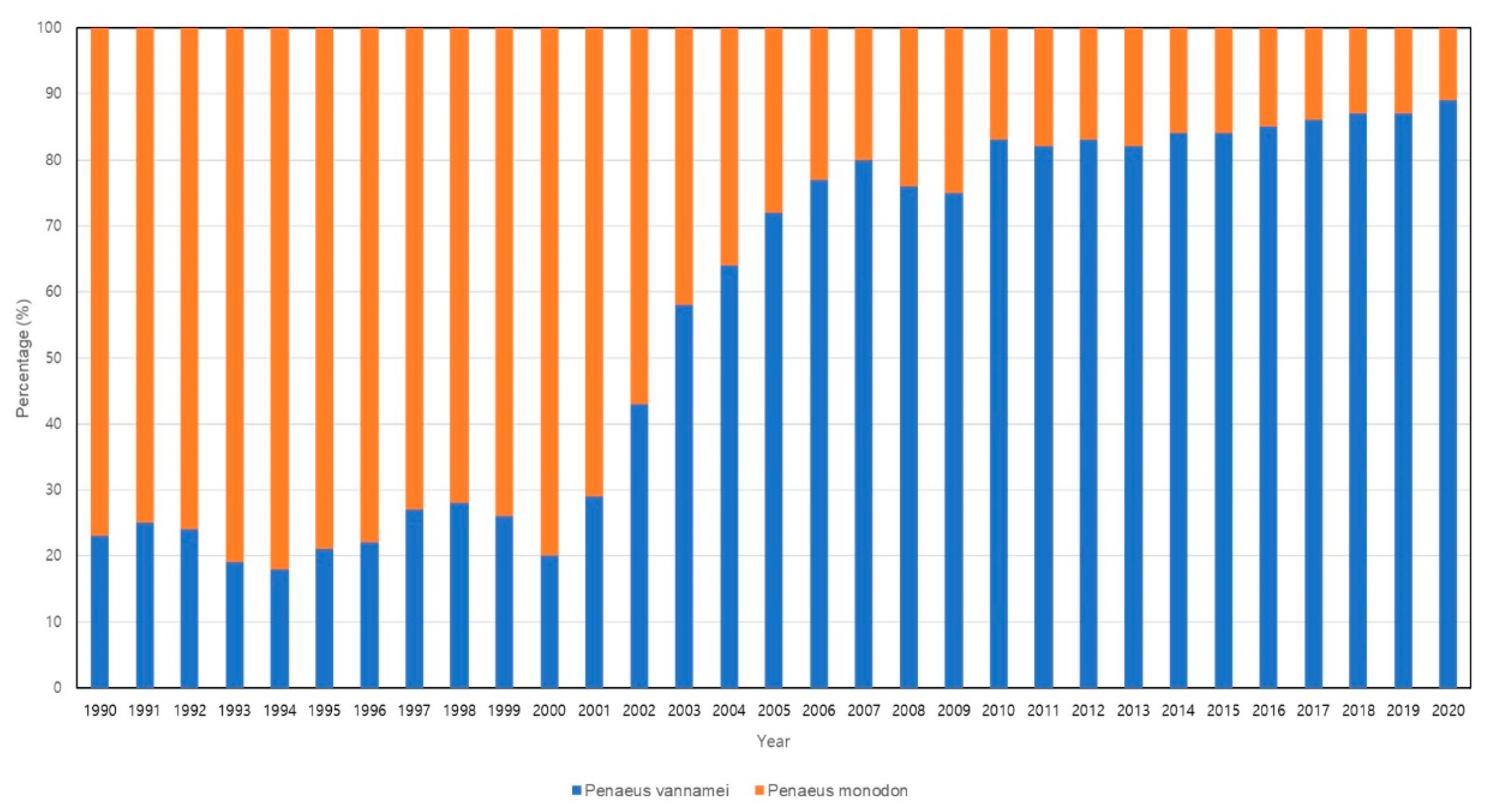
Figure 3.
A comprehensive depiction of diverse categories and consequences of DNA methylation implicated in the epigenetic modulation of shrimps. DNA methylation encompasses two distinct regions: (1) CpG-rich domains, designated as CpG methylation, and regions lacking CpG enrichment, recognized as Non-CpG methylation. (2) Promoter region methylation exerts regulatory control over gene expression by inducing transcriptional inhibition. (3) Gene body methylation transpires within the intron-exon regions, contributing to alterations in gene expression levels and the emergence of "alternative splicing" phenomena.
Figure 3.
A comprehensive depiction of diverse categories and consequences of DNA methylation implicated in the epigenetic modulation of shrimps. DNA methylation encompasses two distinct regions: (1) CpG-rich domains, designated as CpG methylation, and regions lacking CpG enrichment, recognized as Non-CpG methylation. (2) Promoter region methylation exerts regulatory control over gene expression by inducing transcriptional inhibition. (3) Gene body methylation transpires within the intron-exon regions, contributing to alterations in gene expression levels and the emergence of "alternative splicing" phenomena.
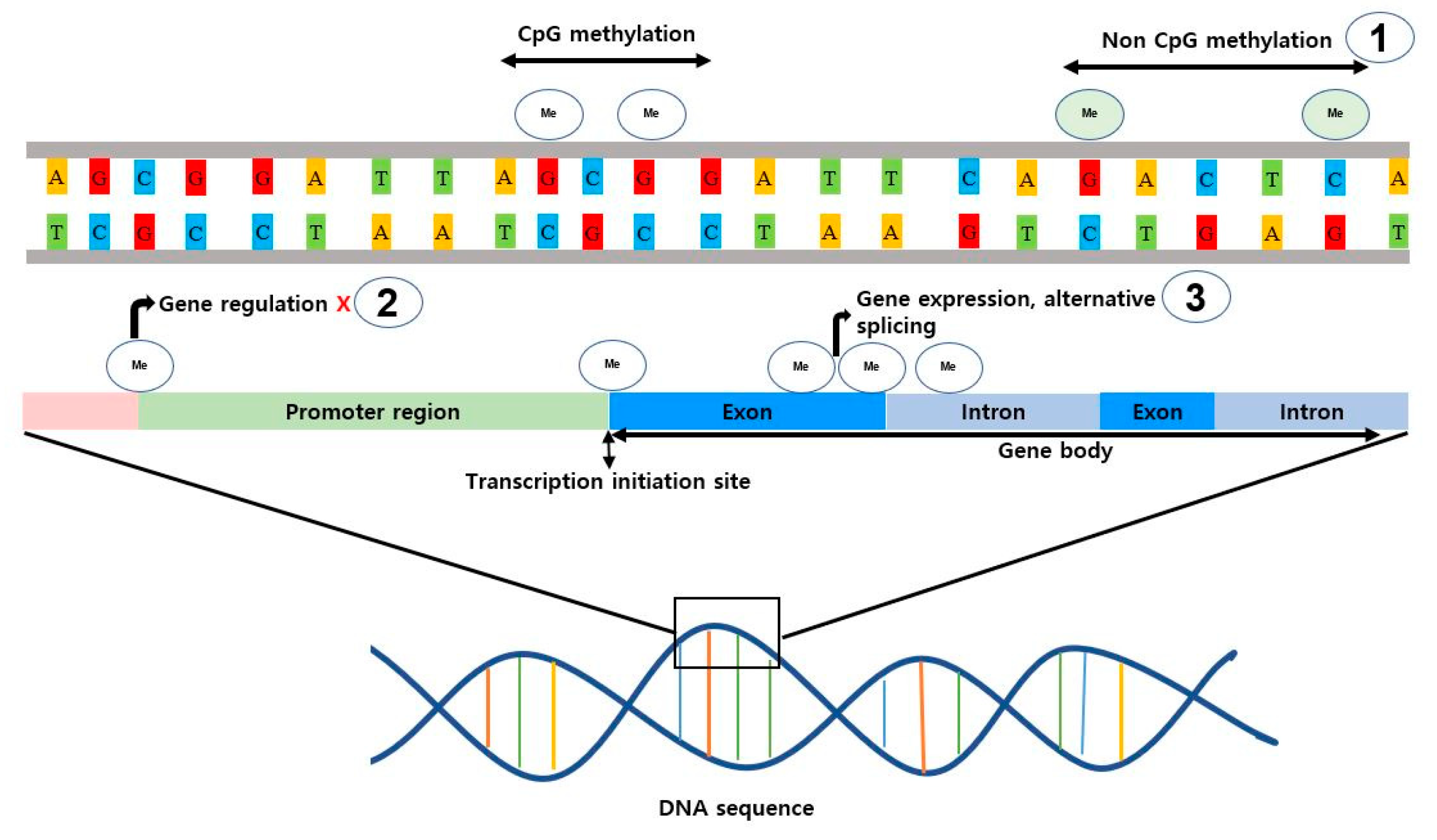
Figure 4.
Schematic representation of post-translational events associated with H2A, H2B, H3, and H4 proteins, highlighting their modification types in epigenetic regulation in animals. Notably, histone methylation (Me), histone phosphorylation (Ph), histone acetylation (Ac), and histone ubiquitination (Ub) are the common mechanisms involved in histone modification. In the context of shrimps, previous investigations have identified specific histone modification patterns, including H3K4, H3K9, and H3K27 methylation, H3K14 and H4 acetylation, and H3S10, H3S28, H2A, and H4 phosphorylation [15,140,141,142]. These modifications play critical roles in regulating gene expression and contribute to various biological processes in shrimps.
Figure 4.
Schematic representation of post-translational events associated with H2A, H2B, H3, and H4 proteins, highlighting their modification types in epigenetic regulation in animals. Notably, histone methylation (Me), histone phosphorylation (Ph), histone acetylation (Ac), and histone ubiquitination (Ub) are the common mechanisms involved in histone modification. In the context of shrimps, previous investigations have identified specific histone modification patterns, including H3K4, H3K9, and H3K27 methylation, H3K14 and H4 acetylation, and H3S10, H3S28, H2A, and H4 phosphorylation [15,140,141,142]. These modifications play critical roles in regulating gene expression and contribute to various biological processes in shrimps.
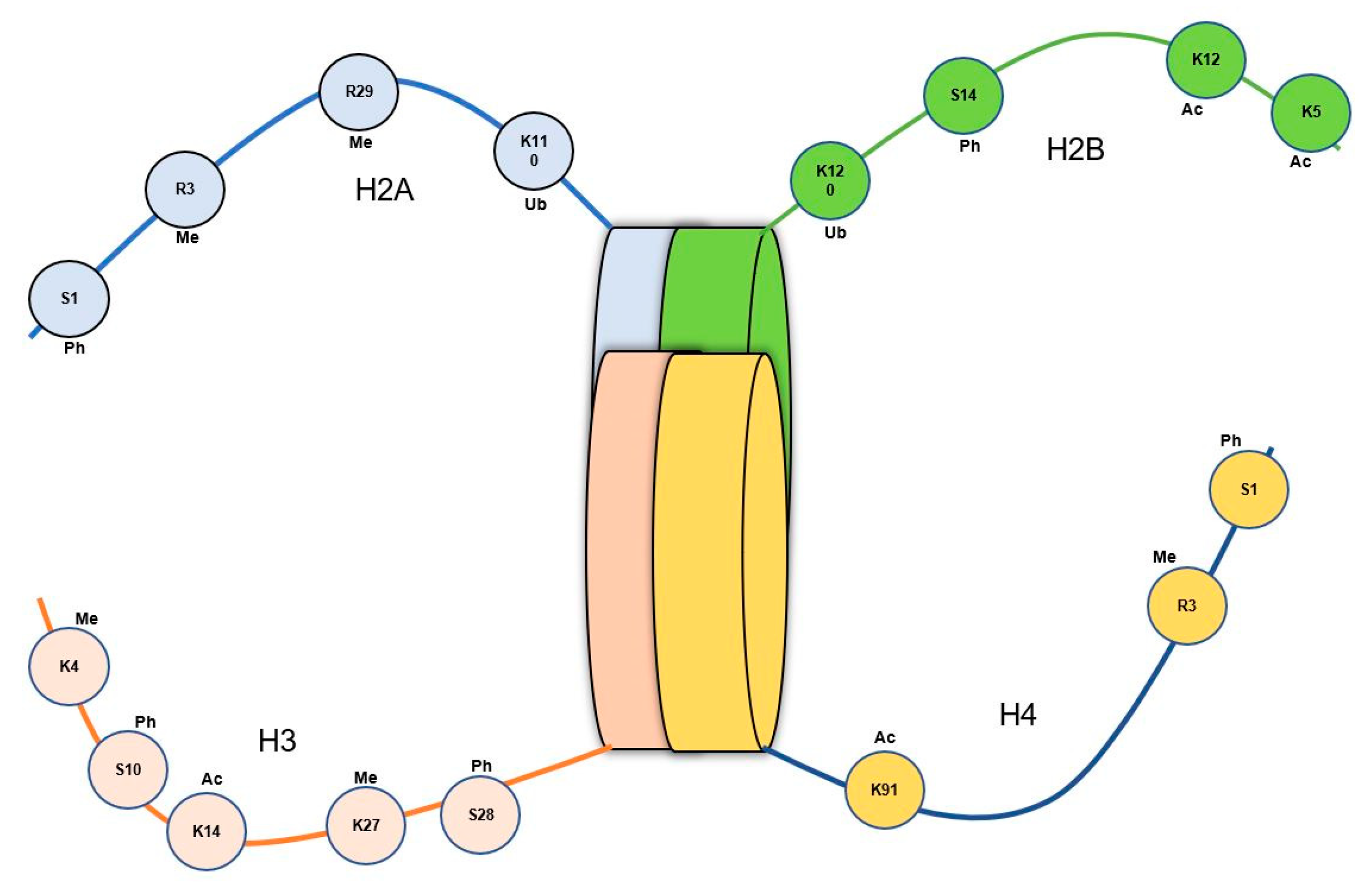
Figure 5.
Schematic diagram depicting the elements of RNA implicated in epigenetic regulation across animal species. Notable examples encompass (1) small interfering RNA (siRNA), (2) micro RNA (miRNA), (3) piwi-interacting RNA (piRNA), and (4) long noncoding RNA (lncRNA). Each type of mechanism involves different types of regulatory molecules. In the context of shrimps, previous investigations have elucidated the existence of specific siRNA (such as vp28-siRNA) and miRNA (such as miR-100, miR-275, miR-7, miR-965), that combat viral and bacterial pathogens including White spot syndrome virus (WSSV) and V. alginolyticus [156,157,163]. Overall, the participation of above noncoding RNAs (ncRNAs) in regulatory processes assumes a pivotal role in the modulation of gene expression and contributes to diverse biological processes in shrimps.
Figure 5.
Schematic diagram depicting the elements of RNA implicated in epigenetic regulation across animal species. Notable examples encompass (1) small interfering RNA (siRNA), (2) micro RNA (miRNA), (3) piwi-interacting RNA (piRNA), and (4) long noncoding RNA (lncRNA). Each type of mechanism involves different types of regulatory molecules. In the context of shrimps, previous investigations have elucidated the existence of specific siRNA (such as vp28-siRNA) and miRNA (such as miR-100, miR-275, miR-7, miR-965), that combat viral and bacterial pathogens including White spot syndrome virus (WSSV) and V. alginolyticus [156,157,163]. Overall, the participation of above noncoding RNAs (ncRNAs) in regulatory processes assumes a pivotal role in the modulation of gene expression and contributes to diverse biological processes in shrimps.
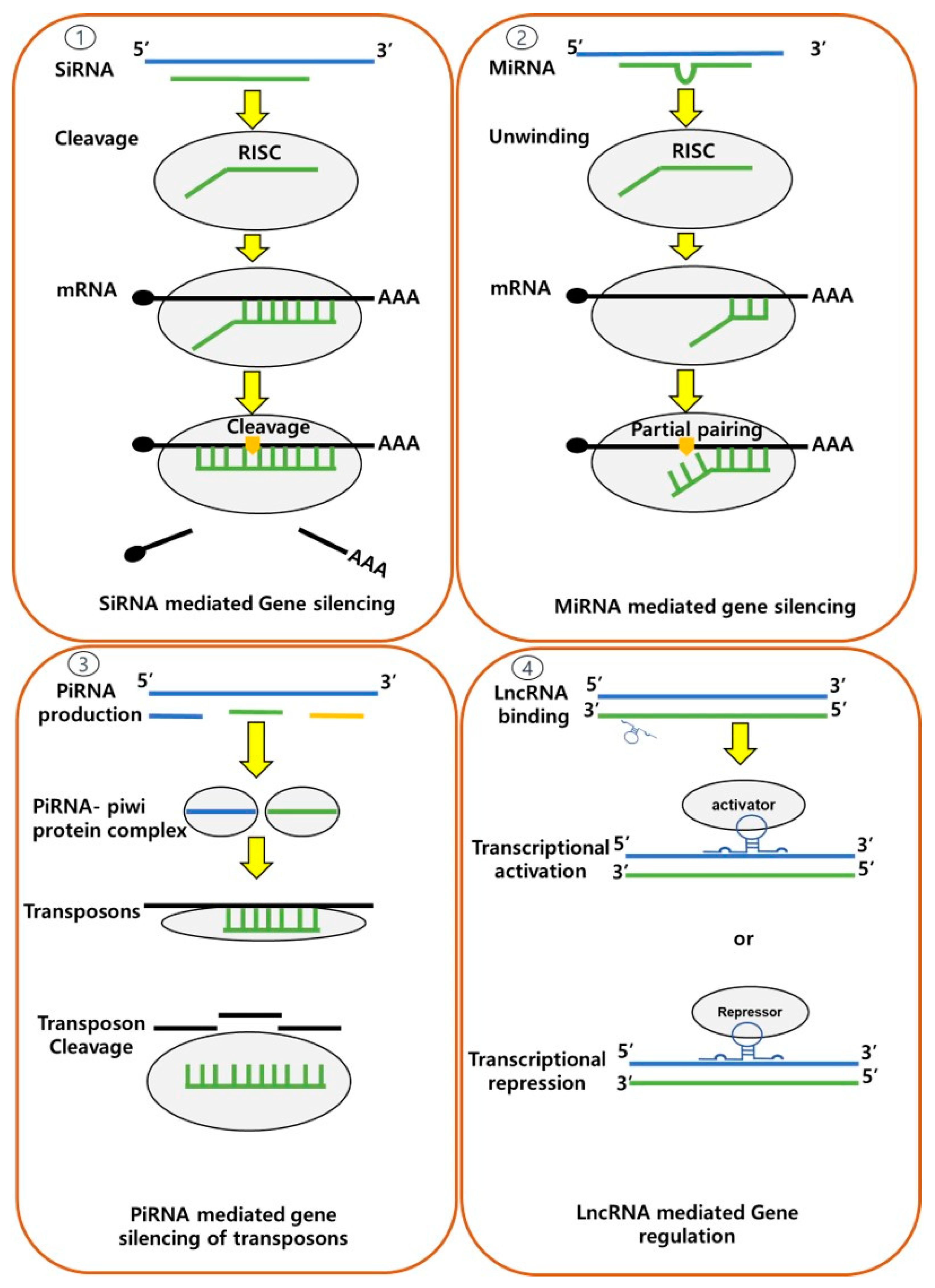
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



