
Submitted:
29 July 2023
Posted:
31 July 2023
You are already at the latest version
Abstract
Total Parenteral Nutrition (TPN), which uses intravenous administration of nutrients, minerals and vitamins, is essential in sustaining premature infants until they transition to enteral feeds. There is limited information on metabolomic differences between infants on TPN and enteral feeds. We performed untargeted global metabolomics on urine samples collected between 23-30 days of life from 314 infants born <29 weeks gestational age from the TOLSURF and PROP cohorts. Principal component analysis across all metabolites showed a separation of infants solely on TPN as compared to infants who had transitioned to enteral feeds, indicating global metabolomic differences between infants based on feeding status. Among 913 metabolites that passed quality control filters, 609 varied in abundance between infants on TPN vs enteral feeds at p<0.05. Of these, 88% were in the direction of higher abundance in the urine of infants on enteral feeds. In a subset of infants with longitudinal analysis, both concurrent and delayed changes in metabolite levels were observed with initiation of enteral feeds. Infants on enteral feeds had higher concentrations of essential amino acids, lipids, and vitamins, which are necessary for growth and development, suggesting a nutritional benefit of an enteral feeding regimen.
Keywords:
prematurity
; extreme premature birth
; enteral feeding
; total parenteral nutrition
; untargeted metabolomic profiling
1. Introduction
Parenteral Nutrition (PN) is an essential therapy for infants who are born extremely prematurely. These infants have minimal nutrient stores essential for their growth and development plus an immature gastrointestinal system that limits nutrient uptake [1,2]. PN delivers all necessary nutrients, including carbohydrates, proteins, lipids, vitamins, and minerals, directly into the bloodstream [3,4]. Almost all extremely premature infants start their postnatal life on Total Parental Nutrition (TPN), deriving all their nutrition from PN. They are then gradually transitioned to enteral feeds when intestinal perfusion seems sufficient to absorb enteral nutrients safely [5]. While TPN usage is a standard clinical practice to provide essential nutrition for infants, the global impact on circulating metabolites is not well defined.
TPN can also negatively affect extremely premature infants' developing gut and metabolism. Potential adverse effects of TPN usage include hyperglycemia, electrolyte abnormalities, acid-base disturbances, essential fatty acid deficiency, and compromised gut integrity [6,7]. While providing nutrition through TPN is crucial for infants experiencing malnutrition, it is important to carefully manage all aspects of nutrition to avoid excessive intake that may result in negative consequences [8,9]. Understanding the influence that PN and especially TPN has on the metabolome of extremely preterm infants is useful for optimizing the timing and dosage of TPN usage in this patient population [10].
Untargeted global metabolomics can be used to identify and quantify levels of many hundreds of biochemicals in a biological fluid at a time [11,12]. An untargeted approach allows the systematic analysis of biofluid composition across a large number of metabolic pathways, eliminating the need to identify biochemical pathways in advance. Untargeted metabolomic approaches also allows for the identification of potential biomarkers from a large pool of candidates, distinguishes those that vary in response to a clinical intervention and captures a wider range of physiological processes for biological inference [13,14]. Lastly, changes in the biochemical composition of the metabolome reflect the intersection of both environmental and genetic contributions, allowing a more comprehensive understanding of metabolomic changes that occur in response to different nutritional interventions [15,16].
We performed untargeted global metabolomics of urine samples from extremely premature infants on TPN vs enteral feeds from two independent clinical cohorts. We hypothesized that there are changes in the urinary metabolome as infants are weaned off TPN feeds and transition to enteral feeding.
2. Materials and Methods
2.1. Study Description
Our study includes untargeted metabolomic profiling of 314 extremely premature infants from two cohorts; a) The Trial of Late Surfactant for Prevention of Bronchopulmonary Dysplasia (TOLSURF, ClinicalTrials.gov, NCT01022580) and b) The Prematurity and Respiratory Outcomes Program (PROP, NCT01435187).
TOLSURF, was a blinded, randomized, sham-controlled trial performed between 2010-2013 at 25 US centers and designed to assess the effects of late surfactant treatment on respiratory outcome. A total of 511 extremely low gestational age (<28 weeks) infants who required mechanical ventilation at 7-14 days were enrolled and received either surfactant treatment or sham installation every 1-3 days. A detailed description of the cohort, the trial design, infant characteristics, and effects of late surfactant treatment can be found elsewhere[17].
PROP was an observational study performed between 2010-2013 at 8 US centers and designed to collect clinical data and biospecimens for analysis of factors related to respiratory outcomes. A total of 835 infants <29 wks were enrolled at 1-7 days with characteristics as previously described [18]. The research protocol for both studies was approved by the Institutional Review Boards of the participating institutions and a parent of each infant provided written informed consent.
2.2. Metabolomic Profiling
We performed global metabolomic studies on single urine samples collected at day 23-30 from infants of both TOLSURF (n=108 TPN feeds, n=63 enteral feeds) and PROP (n=80 TPN feeds, n=63 enteral feeds) cohorts. TPN was initiated in all infants within the first two days of birth. In our study, the infants included in the enteral feeds group were required to have completely transitioned to enteral feeds at least two days prior of the collection of their urine samples for metabolomic profiling. Detailed feeding data were available for PROP infants; most (86.7%) of those who were on full enteral feeds at week 4-5 received breast milk: (70.2% breast milk only or a mixture of breast milk and formula 16.7%). Additionally, 28 infants in the TOLSURF study had longitudinal metabolomic profiling of their urine (collected at 2-7 days intervals from 7-50 days after birth); all of these infants received breast milk as their enteral feeds.
Untargeted metabolomics was performed by Ultra High Performance Liquid Chromatography-Tandem Mass Spectroscopy (UHPLC-MS/MS) as performed by Metabolon Inc (Morrisville, NC). All methods utilized a Waters ACQUITY ultra-performance liquid chromatography (UPLC) and a Thermo Scientific Q-Exactive high resolution/accurate mass spectrometer interfaced with a heated electrospray ionization (HESI-II) source and Orbitrap mass analyzer operated at 35,000 mass resolution. Each reconstitution solvent contained a series of standards at fixed concentrations to ensure injection and chromatographic consistency. One aliquot was analyzed using acidic positive ion conditions, chromatographically optimized for more hydrophilic compounds. In this method, the extract was gradient eluted from a C18 column (Waters UPLC BEH C18-2.1x100 mm, 1.7 µm) using water and methanol, containing 0.05% perfluoropentanoic acid (PFPA) and 0.1% formic acid. Another aliquot was also analyzed using acidic positive ion conditions optimized for more hydrophobic compounds. In this method, the extract was gradient eluted from the same C18 column using methanol, acetonitrile, water, 0.05% PFPA and 0.01% formic acid and was operated at an overall higher organic content. Another aliquot was analyzed using basic negative ion optimized conditions using a separate dedicated C18 column. The basic extracts were gradient eluted from the column using methanol and water, with 6.5mM ammonium bicarbonate at pH 8. The fourth aliquot was analyzed via negative ionization following elution from a HILIC column (Waters UPLC BEH Amide 2.1x150 mm, 1.7 µm) using a gradient consisting of water and acetonitrile with 10mM ammonium formate, pH 10.8. The MS analysis alternated between MS and data-dependent MSn scans using dynamic exclusion. The scan range varied slightly between methods but covered 70-1000 m/z.
Urine samples were normalized to osmolality and missing values imputed to the minimum detectable level. For quality control, we only included metabolites in our study if they were profiled in at least 70% of infants in each cohort separately (Figure S2). Data are expressed normalized to the median value for each biochemical.
2.3. Global Metabolomic Analysis
Using the 913 metabolites in our study, we performed principal component analysis (PCA) [19] to visualize global differences in the urinary metabolome of infants on TPN vs enteral feeds. Our metabolomic profiling was done in two batches for TOLSURF and PROP separately. For the PROP cohort, we included additional anchor samples from the TOLSURF cohort to merge the two batches into a joint metabolomic matrix, allowing us to compare metabolite abundances between the two cohorts. We performed PCA on a matrix of 314 Infants x 913 Metabolites using Sklearn in Python 3.8. We then performed Unifold Manifold Approximation and Projection (UMAP) for dimension reduction using the UMAP package in Python 3.8 using the raw metabolomic matrices. PCA and UMAP analyses were implemented and performed in Python; the source code is available on GitHub. A Mann Whitney U-test was used to test for spatial separation between infants on TPN vs. enteral feeds for the first two PCs and UMAP coordinates in Python.
2.4. Metabolome Wide Association Analysis (MWAS)
We performed a metabolome-wide association study using logistic regression to identify individual biochemicals that vary in abundance by mode of feeding (enteral vs. TPN), adjusting for potential confounders of clinical importance: birthweight, sex, corticosteroid treatment, and maternal self-identified race/ethnicity. Analyses were performed within each study separately, and results combined in a meta-analysis to account for study and experimental batch effects between the two cohorts (TOLSURF and PROP) (Tables S1, S2 and S3). MWAS was implemented using a custom Python 3.8 script using the statsmodel package for logistic regression analysis. Meta-analyses using a weighted inverse-variance were performed using the metafor package in R using summary statistics from MWAS analyses across TOLSURF and PROP. All source code is available on GitHub.
2.5. Metabolite Set Enrichment Analysis
We performed a metabolite set enrichment analysis (MSEA, or pathway analysis) to identify metabolic pathways enriched for metabolites that vary between infants on TPN vs. enteral feeds following Bonferroni correction for 913 tests (p-value < 5.47e-5). Identified (named) metabolites from Metabolon were annotated to nine super-pathways and 92 sub-pathways. For each pathway, we tested for enrichment of metabolites from our MWAS using a Fisher Exact Test, adjusting for multiple comparisons by inferring the false discovery rate (FDR) using the qvalue package in R.
2.6. Time Course Comparison Analysis
For 28 infants in the TOLSURF study, we conducted longitudinal metabolomic profiling of their urine between 7-50 days after birth. For each infant, the age at full enteral feeds was recorded, with the weaning process beginning approximately a week before. We compared two-time points, one before and one after stopping TPN, for each infant to quantitate the impact of transitioning to enteral feeds [20]. The first time point was at least five days before stopping TPN (TP1), and the second time point was 0-2 days after starting full enteral feeds (TP2). Five infants were excluded due to not having timepoints in the inclusion criteria, leaving 23 infants for analysis. We used TP2/TP1 to generate a fold change of the effect for each metabolite, and abundance of each metabolite was compared between time points using a paired t-test (Table S4).
3. Results
Overall, we identified 1437 biochemicals, of which 913 passed our quality control filters, including detection in at least 70% of all infant samples (Table S2). Of the 913 biochemicals, 689 were named metabolites (by Metabolon) as of July 2022. Clinical characteristics of infants included in our study are shown in Table 1, including maternal self-reported racial-ethnic groups that were balanced between the comparison groups. Infants on hydrocortisone treatment were more likely to be on TPN at the time of metabolomic profiling than enteral feeds (p=0.0029), and thus we adjusted for hydrocortisone treatment in addition to birthweight, sex, and maternal self-identified race/ethnicity in our analyses.
Global analyses over all metabolites using dimension reduction showed both a qualitative separation of infants by feeding status (Figure 1a), and a statistically significant separation for both PC1 (p = 1.51e-15) and PC2 (p = 4.36e-9) (Figure 1b). UMAP showed a similar separation of infants by feeding status across both latent dimensions (Figure S2). These results highlight the global metabolomic shift between infants who are on TPN vs enteral feeds (primarily breast milk).
In our analyses of individual metabolites, we found that 609 of 913 (67%) metabolites were associated with infants feeding status at p<0.05 (Figure 2), consistent with the separation of infants by feeding status in our global metabolomic comparisons. Of the 609 metabolites that varied by feeding status, 309 (51%) were still significant when applying a conservative Bonferroni threshold to adjust for multiple comparisons across 913 tests (p<5.5e-5). Among metabolites that varied by feeding status with p<0.05, the majority (88%) of metabolites were enriched in infants on enteral feeds as compared to TPN (Figure 2) (Table S3).
The top 20 most significant associations with mode of feeding are shown in Table 2. Dexpanthenol (provitamin B5) and gluconate, which belong to the Xenobiotic Super-Pathway, had the strongest signals of association with feeding status and were enriched in infants on TPN. Both chemicals are components of PN as vitamins (dexpanthenol) and mineral (calcium gluconate) supplements, and their concentrations serve as positive controls for the level of TPN administration. We additionally identified Pantothenate, a natural form of vitamin B5, as a top hit in our MWAS study and found higher levels for infants on enteral feeds. Eight of the top 20 hits were cofactors and vitamins; levels were higher on enteral feeds for 6 of these chemicals. Other significant metabolites were members of lipid, amino acid, carbohydrate, and energy super-pathways.
In the amino acid super-pathway, twelve of twenty amino acids were significantly associated with infant feeding status, with concentrations of 11 amino acids being higher in infants on enteral feeds (avg FC = 1.59) (Figure 3a). Four of these were essential amino acids (not synthesized in vivo): lysine, leucine, threonine, and tryptophan. The only amino acid with significantly higher levels in the urine of infants on TPN feeds was cysteine (8.2-fold), a non-essential sulfur-containing amino acid important for protein synthesis and redox status [21].
Additionally, we identified differences in lipid metabolites involved in acylcarnitine pathways, which play a crucial role in energy metabolism and may contribute to necrotizing enterocolitis (NEC) [22]. We examined the impact of feeding status on acylcarnitines using the metabolites profiled by Sylvester et al 2017 in their analysis assessing the impact of acylcarnitine on NEC [22]. C5 was profiled across three metabolites (isovalerylcarnitine, 2-methlbututyrylcarnitine, and pivalerylcarnitine), and two were profiled for C4 (isobutyrylcarnitine and butyrylcarnitine) (Figure 3b). Of the 21 metabolites used in their analysis, 14 were profiled in our analysis. We find 7 acylcarnitines significant in our analysis, with three of them belonging to the C5 subclass (Pivaloylcarnitine, 2-methylbutryrlcarnitine, and isovalerylcarnitine).
Using super- and sub-pathway annotations provided by Metabolon, we performed pathway enrichment analysis to identify metabolomic pathways enriched for metabolites that vary by feeding status. We found two super-pathways (Energy and Nucleotide) and eight sub-pathways significantly enriched in infants on enteral feeds (Figure 4). Of the eight sub-pathways, three were acylcarnitines: long-chain saturated, dicarboxylate, and dihydroxy fatty acids. Additionally, we find the ATP sub-pathway enriched as a component of the Energy super-pathway.
Finally, we examined the kinetics of changes in metabolite levels with weaning of TPN, which occurs over approximately 1 week. For 28 infants in the TOLSURF study, we conducted longitudinal metabolomic profiling of their urine (7-50 days after birth) (Table S4). We first qualitatively determined the time course of change with transition to enteral feeds for the top metabolites in each of 8 super-pathways identified in our MWAS analysis and compared the temporal pattern with that of Dexpanthenol as a marker of an exogenous chemical in TPN. Each of these metabolites, except for dexpanthenol, was enriched in infants on enteral feeds. For the infant shown in Figure 5, dexpanthenol abundance increased over the first 20 days and then decreased dramatically (74-fold) as TPN feeds decreased over 3 days and full milk feeds began on day 23. Conversely, there are temporally related increases in concentrations of N-acetyl asparagine (3.3-fold, amino acid super-pathway) and 3-methyladipate (2.2-fold increase, lipid super-pathway) over the same 3-day transition to enteral feeds (Figure 5a). For this infant, 4 metabolites demonstrated a rapid increase in concentration concurrent with the decrease in dexpanthenol, 3 metabolites had a delayed increase, and 1 metabolite (prolylglycine) demonstrated a steady increase in concentration over time independent of the transition in feeding status (Figure 5b-d).
We used two time point values, one before and one after stopping TPN, to quantitate the impact of the change in feeding status to complement our single timepoint/cross-sectional analysis. We found 461 metabolites across both approaches with a change in concentration in the same direction by each analysis (p<0.05). Using linear regression, there was a strong correlation between the fold changes of metabolites (R2=0.37, p=2.53e-83) and p values (R2=0.40, p=9.02e-93) (Figure S3) comparing cross-sectional and longitudinal results.
4. Discussion
Our study utilized untargeted metabolomics to identify urinary metabolites that change in response to TPN vs. enteral feeds. Our data indicate a global and dramatic shift of multiple metabolomic pathways when comparing infants on TPN versus enteral feeds. We identified 609 out of 913 urinary chemicals with altered concentrations. Of these metabolites, 88% have higher concentrations for infants on enteral feeds. Metabolites were enriched on enteral feeds across all nine metabolomic super-pathways, indicating the wide scope of responses, and levels of many biochemicals changed concurrently with the gradual reduction of TPN dose.
Prior studies with lower numbers of infants have assessed metabolomic differences by modes of feeding, allowing comparisons with our results. A study by Esturau-Escofet identified eight metabolites in the urine of 34 preterm infants that varied by enteral vs parenteral nutrition [23]. Of the eight, we observed similar differences in five metabolites in the same direction of being enriched in PN vs enteral feeds (gluconate, N-acetyltyrosine, succinate, citrate, and lactose). However, this study included infants of predominantly higher gestational age (26-36 weeks, with only three infants being born extremely premature at < 29 weeks). Nilsson et al. examined levels of 31 blood biochemicals over time in extremely premature infants [24]. Of the eight amino acids also detected in our study (urine), only tyrosine showed a similar pattern of increased abundance on enteral feeds. However, an additional eight biochemicals were in agreement and were more abundant in infants on enteral feeds in both studies, which queried different biological fluids (3-hydroxyisobutyrate, creatinine, glycerol, leucine, lysine, proline, threonine, and uridine). Finally, a study by Falaina et al observed global shifts in the urinary metabolome of term infants exclusively fed breast milk vs formula across the first 3 months of life [15], similar to the dramatic shift we observe between PN vs enteral feeds. Thus, although prior studies varied substantially from our own, a number of similarities were observed in terms of individual metabolites and observations of a global metabolic impact of feeding status. Highlights of our study include two cohorts of extremely premature infants with diverse maternal racial/ethnic backgrounds recruited from 35 different medical centers across the US. From a methodological perspective, our untargeted approach identified close to 1000 different urinary chemicals for univariate and longitudinal comparisons, allowing us to identify individual metabolites that change by type of nutrition while also adjusting for confounding factors.
This study illustrates the benefit of global metabolomics approaches in identifying and quantifying systemic effects of environmental factors such as diet, medications, and contaminants. The change in the human metabolome as infants transition to enteral feeds demonstrates the wide scope of both direct and indirect effects of a change in diet. The large percentage of chemicals that change in both detection and concentration with initiation of enteral feeds is not unexpected given the limited and defined composition of PN compared to both formula, which contains bovine skim milk and whey concentrate (with any bound chemicals), and even more so to breast milk that contains chemicals derived from maternal diet, medications, and exposures. The change in metabolite concentration secondary to altered intake is illustrated by amino acid metabolism, where 11 out of the 12 amino acids with altered levels were enriched for infants on milk feeds; the only metabolite found enriched in infants on TPN feeds was cysteine [25].
Two lipid metabolites affected by feeding status in our study, C5 and C12 acylcarnitine, are associated with the risk of developing NEC, which is an inflammatory condition of the immature gastrointestinal mucosa in premature infants that is often related to enteral feeds. Extremely premature infants often have an insufficient supply of carnitine due to immature liver and kidneys, which can lead to toxic fatty acid build-ups. Recent work has shown TPN usage to increase the risk of developing NEC, suggesting a potential role in regulating fatty acid metabolism [22]. Interestingly, two metabolites were found in consensus directions with developing risk for NEC and TPN usage. C5 was enriched in infants on TPN feed (ORmeta = 2.00), with higher levels associated with an increased risk of developing NEC (ORNEC = 1.75). C12 was enriched in infants on Enteral Feeds (ORmeta = 0.73) with higher levels found protective against developing NEC (ORNEC = 0.74). This finding highlights the potential that the duration of TPN may impact levels of C5, which could contribute to the risk of developing NEC. However, it is unclear whether increased C5 with TPN is contributing to development of NEC or is rather a surrogate for sicker premature infants. Further work will be needed to look at the relationship between acylcarnitine levels and NEC and to establish the optimal dose to use in PN [26].
We report a preliminary survey of the time course of metabolite concentrations related to weaning the dose of TPN and initiating enteral feeds, including examples of both rapid and delayed changes in concentrations of biochemicals. The different temporal patterns likely reflect a direct result of change in intake amount (eg., isocitric lactone, rapid response) versus changes in concentration resulting indirectly from altered levels of a biochemical in the same pathway (eg., lyxonate, delayed response). Additionally, delayed responses in circulating metabolites may arise from altered composition of the gut microbiome secondary to intake of milk, an area for future study.
The dramatic changes we observe in the metabolome by mode of feeding could have implications for Infant Newborn Blood Screening (NBS). NBS is used to detect newborns with severe but treatable rare disorders to help avoid adverse outcomes [27]. NBS screening has evolved over the years but still involves using mass spectrometry on blood samples to identify inborn errors of metabolism or other inherited disorders [27,28]. Previously, strong associations between measured NBS metabolites and persistent pulmonary hypertension, hyperbilirubinemia, and NEC have been reported [26,27,28,29]. Recently, Oltman et al described a metabolomic vulnerability profile that combines clinical characteristics and selected metabolites from NBS to predict the occurrence of common complications of preterm birth as well as overall morbidity and mortality [30]. Their model included 16 metabolites that were measured as part of NBS: 7 amino acids and 9 acylcarnitine. In our study we found that concentrations of 8 of these metabolites were influenced by feeding status (glycine, leucine/isoleucine, proline, tyrosine, and acylcarnitines C-3, C-4, C-5, and C-12). Because NBS screening in premature infants can occur between 1 and 7 days after birth, these infants may be receiving full TPN, full enteral feeds or a mixture of parenteral and enteral feeding, which will influence metabolite levels. Thus, the predictive value of vulnerability profiles using NBS data will be impacted by feeding status, and ideally this information is collected and used for adjustments of predictive profiling.
There are some limitations to our study. The TOLSURF and PROP clinical studies were conducted between 2010 and 2013. Thus, the urine samples have been stored (at -70 C) for up to ten years, and it is possible that oxidation or degradation of some biochemicals occurred. Even though we used two clinical cohorts spanning multiple medical centers, larger and inclusive cohorts should replicate this analysis and highlight population-specific effects of infant metabolism. Additionally, for all the metabolites associated with our analysis that may have clinical importance, targeted analyses should be performed to validate the effect of feeding status on metabolite abundances. Another limitation of this analysis was the inability, due to lack of clinical data, to compare the effect of breast milk vs formula milk on the urinary metabolome; future studies can address this interesting topic [31]. It's also important to note that infants on TPN might be sicker than infants who transitioned to enteral feeds, which might impact metabolism. Although we adjusted for BPD, future studies to inform a causal relationship is warranted. Lastly, our study has a limitation regarding the exclusion of rare biochemicals. Our method of imputation results in left-censored data, whereby undetected metabolites are assigned a value equal to the minimum level of detection. However, it is important to note that we identified an additional 204 metabolites of significance below our imputation quality threshold. These biochemicals were profiled in less than 70% of the population (Table S1). It is crucial for future research to address this issue by developing methodologies that incorporate rare biochemicals into metabolomic association studies.
5. Conclusions
In conclusion, we observed a large difference in the urinary metabolome of extremely premature infants on TPN versus enteral feeds, with primarily increased levels of biochemicals in infants on enteral feeds. The results of this study support the common clinical practice to transition neonates as soon as clinically able from TPN to enteral feeds. The metabolomic data presented in this study can be used to further understand the complex interplay between nutrition and infant growth and development.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Concept; Project, All Authors; Methodology, M.G.; Software, M.G.; Validation, M.G.; Formal Analysis, M.G.; Investigation, M.G.; Resources, (All Authors); Data Curation, (All Authors); Writing, M.G.; Review & Editing, (All Authors ); Visualization, M.G., C.C.; Supervision, R.D.H., D.T, P.L.B; Project Administration, D.T, P.L.B.; Funding Acquisition, D.T, P.L.B., R.D.H.
Funding
Supported by NIH grants R01HL153429 (P.L.B, D.T.) and R01GM142112 (R.D.H.). M.G. is a recipient of a Howard Hughes Medical Institute Gilliam Fellowship, Achievement Award for College Scientists Foundation Scholarship, and an UCSF Discovery Fellows Program Award.
Institutional Review Board Statement
The research protocols for both clinical trials were approved by the Institutional Review Boards of the participating institutions, and a parent of each infant provided written informed consent.
Informed Consent Statement
A parent of each infant provided written informed consent.
Data Availability Statement
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors thank the investigators and parents for their contributions to the TOLSURF and PROP trials and gratefully acknowledge the TOLSURF PI leadership (Roberta Ballard). The authors thank Tom Mariani, director of the PROP Biospecimen Repository, for providing samples for this study. The authors also thank Mallika John for her assistance in reviewing the manuscript.
Conflicts of Interest
The authors declare no competing interests.
References
- Demerath, E.W.; Johnson, W.; Davern, B.A.; Anderson, C.G.; Shenberger, J.S.; Misra, S.; Ramel, S.E. New Body Composition Reference Charts for Preterm Infants. Am. J. Clin. Nutr. 2017, 105, 70–77. [Google Scholar] [CrossRef] [PubMed]
- ElHassan, N.O.; Kaiser, J.R. Parenteral Nutrition in the Neonatal Intensive Care Unit. Neoreviews 2011, 12, e130–e140. [Google Scholar] [CrossRef]
- Carnielli, V.P.; Correani, A.; Giretti, I.; D’Ascenzo, R.; Bellagamba, M.P.; Burattini, I.; Biagetti, C. Practice of Parenteral Nutrition in Preterm Infants. Nutr. Care Preterm Infants 2021, 122, 198–211. [Google Scholar] [CrossRef]
- Hamdan, M.; Puckett, Y. Total Parenteral Nutrition; StatPearls Publishing, 2022. [Google Scholar]
- Darmaun, D.; Lapillonne, A.; Simeoni, U.; Picaud, J.-C.; Rozé, J.-C.; Saliba, E.; Bocquet, A.; Chouraqui, J.-P.; Dupont, C.; Feillet, F.; Frelut, M.-L.; Girardet, J.-P.; Turck, D.; Briend, A.; Committee on Nutrition of the French Society of Pediatrics (CNSFP), and French Society of Neonatology (SFN). Parenteral Nutrition for Preterm Infants: Issues and Strategy. Arch. Pediatr. 2018, 25, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Jeejeebhoy, K.N. Total Parenteral Nutrition: Potion or Poison? Am. J. Clin. Nutr. 2001, 74, 160–163. [Google Scholar] [CrossRef]
- Lavoie, J.-C.; Chessex, P. Parenteral Nutrition and Oxidant Stress in the Newborn: A Narrative Review. Free Radic. Biol. Med. 2019, 142, 155–167. [Google Scholar] [CrossRef]
- Chowdary, K.V.R.; Reddy, P.N. Parenteral Nutrition: Revisited. Indian J. Anaesth. 2010, 54, 95–103. [Google Scholar] [CrossRef]
- Kumar, R.K.; Singhal, A.; Vaidya, U.; Banerjee, S.; Anwar, F.; Rao, S. Optimizing Nutrition in Preterm Low Birth Weight Infants-Consensus Summary. Front Nutr 2017, 4, 20. [Google Scholar] [CrossRef]
- Patel, P.; Bhatia, J. Total Parenteral Nutrition for the Very Low Birth Weight Infant. Semin. Fetal Neonatal Med. 2017, 22, 2–7. [Google Scholar] [CrossRef]
- Board on Life Sciences; Division on Earth and Life Studies; National Academies of Sciences, Engineering, and Medicine. Use of Metabolomics to Advance Research on Environmental Exposures and the Human Exposome; National Academies Press (US), 2016. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]
- Kindschuh, W.F.; Baldini, F.; Liu, M.C.; Liao, J.; Meydan, Y.; Lee, H.H.; Heinken, A.; Thiele, I.; Thaiss, C.A.; Levy, M.; Korem, T. Preterm Birth Is Associated with Xenobiotics and Predicted by the Vaginal Metabolome. Nat Microbiol 2023, 8, 246–259. [Google Scholar] [CrossRef]
- Steurer, M.A.; Oltman, S.; Baer, R.J.; Feuer, S.; Liang, L.; Paynter, R.A.; Rand, L.; Ryckman, K.K.; Keller, R.L.; Jelliffe-Pawlowski, L.L. Altered Metabolites in Newborns with Persistent Pulmonary Hypertension. Pediatr. Res. 2018, 84, 272–278. [Google Scholar] [CrossRef]
- Falaina, V.; Fotakis, C.; Boutsikou, T.; Tsiaka, T.; Moros, G.; Ouzounis, S.; Andreou, V.; Iliodromiti, Z.; Xanthos, T.; Vandenplas, Y.; Iacovidou, N.; Zoumpoulakis, P. Urine Metabolomic Profile of Breast- versus Formula-Fed Neonates Using a Synbiotic-Enriched Formula. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- López-Hernández, Y.; Oropeza-Valdez, J.J.; Blanco-Sandate, J.O.; Herrera-Van Oostdam, A.S.; Zheng, J.; Chi Guo, A.; Lima-Rogel, V.; Rajabzadeh, R.; Salgado-Bustamante, M.; Adrian-Lopez, J.; Castillo, C.G.; Robles Arguelles, E.; Monárrez-Espino, J.; Mandal, R.; Wishart, D.S. The Urinary Metabolome of Healthy Newborns. Metabolites 2020, 10, 165. [Google Scholar] [CrossRef]
- Ballard, R.A.; Keller, R.L.; Black, D.M.; Ballard, P.L.; Merrill, J.D.; Eichenwald, E.C.; Truog, W.E.; Mammel, M.C.; Steinhorn, R.H.; Rogers, E.E.; Ryan, R.M.; Durand, D.J.; Asselin, J.M.; Bendel, C.M.; Bendel-Stenzel, E.M.; Courtney, S.E.; Dhanireddy, R.; Hudak, M.L.; Koch, F.R.; Mayock, D.E.; McKay, V.J.; O’Shea, T.M.; Porta, N.F.; Wadhawan, R.; Palermo, L.; TOLSURF Study Group. Randomized Trial of Late Surfactant Treatment in Ventilated Preterm Infants Receiving Inhaled Nitric Oxide. J. Pediatr. 2016, 168, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, B.B.; Feng, R.; Schmidt, B.; Aschner, J.L.; Ballard, R.A.; Hamvas, A.; Reynolds, A.M.; Shaw, P.A.; Jobe, A.H.; Prematurity and Respiratory Outcomes Program. Comparisons and Limitations of Current Definitions of Bronchopulmonary Dysplasia for the Prematurity and Respiratory Outcomes Program. Ann. Am. Thorac. Soc. 2015, 12, 1822–1830. [Google Scholar] [CrossRef]
- Eicher, T.; Kinnebrew, G.; Patt, A.; Spencer, K.; Ying, K.; Ma, Q.; Machiraju, R.; Mathé, A.E.A. Metabolomics and Multi-Omics Integration: A Survey of Computational Methods and Resources. Metabolites 2020, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, D.; Guardado, M.; Steurer, M.; Chapin, C.; Hernandez, R.D.; Ballard, P.L. The Hydrocortisone-Responsive Urinary Metabolome of Premature Infants. Pediatr. Res. 2023. [Google Scholar] [CrossRef]
- Zhang, Z.; Adelman, A.S.; Rai, D.; Boettcher, J.; Lőnnerdal, B. Amino Acid Profiles in Term and Preterm Human Milk through Lactation: A Systematic Review. Nutrients 2013, 5, 4800–4821. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, K.G.; Kastenberg, Z.J.; Moss, R.L.; Enns, G.M.; Cowan, T.M.; Shaw, G.M.; Stevenson, D.K.; Sinclair, T.J.; Scharfe, C.; Ryckman, K.K.; Jelliffe-Pawlowski, L.L. Acylcarnitine Profiles Reflect Metabolic Vulnerability for Necrotizing Enterocolitis in Newborns Born Premature. J. Pediatr. 2017, 181, 80–85.e1. [Google Scholar] [CrossRef]
- Esturau-Escofet, N.; Rodríguez de San Miguel, E.; Vela-Amieva, M.; García-Aguilera, M.E.; Hernández-Espino, C.C.; Macias-Kauffer, L.; López-Candiani, C.; Naveja, J.J.; Ibarra-González, I. A Longitudinal 1H NMR-Based Metabolic Profile Analysis of Urine from Hospitalized Premature Newborns Receiving Enteral and Parenteral Nutrition. Metabolites 2022, 12, 255. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.K.; Tebani, A.; Malmodin, D.; Pedersen, A.; Hellgren, G.; Löfqvist, C.; Hansen-Pupp, I.; Uhlén, M.; Hellström, A. Longitudinal Serum Metabolomics in Extremely Premature Infants: Relationships With Gestational Age, Nutrition, and Morbidities. Front. Neurosci. 2022, 16, 830884. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.E.; Coody, T.K.; Jeong, M.-Y.; Berg, J.A.; Winge, D.R.; Hughes, A.L. Cysteine Toxicity Drives Age-Related Mitochondrial Decline by Altering Iron Homeostasis. Cell 2020, 180, 296–310.e18. [Google Scholar] [CrossRef] [PubMed]
- Borum, P.R. Carnitine in Parenteral Nutrition. Gastroenterology 2009, 137 (Suppl. 5), S129–S134. [Google Scholar] [CrossRef]
- Wilcken, B.; Wiley, V. Newborn Screening. Pathology 2008, 40, 104–115. [Google Scholar] [CrossRef]
- Therrell, B.L.; Padilla, C.D.; Loeber, J.G.; Kneisser, I.; Saadallah, A.; Borrajo, G.J.C.; Adams, J. Current Status of Newborn Screening Worldwide: 2015. Semin. Perinatol. 2015, 39, 171–187. [Google Scholar] [CrossRef]
- Remec, Z.I.; Trebusak Podkrajsek, K.; Repic Lampret, B.; Kovac, J.; Groselj, U.; Tesovnik, T.; Battelino, T.; Debeljak, M. Next-Generation Sequencing in Newborn Screening: A Review of Current State. Front. Genet. 2021, 12, 662254. [Google Scholar] [CrossRef]
- Oltman, S.P.; Rogers, E.E.; Baer, R.J.; Jasper, E.A.; Anderson, J.G.; Steurer, M.A.; Pantell, M.S.; Petersen, M.A.; Partridge, J.C.; Karasek, D.; Ross, K.M.; Feuer, S.K.; Franck, L.S.; Rand, L.; Dagle, J.M.; Ryckman, K.K.; Jelliffe-Pawlowski, L.L. Newborn Metabolic Vulnerability Profile Identifies Preterm Infants at Risk for Mortality and Morbidity. Pediatr. Res. 2021, 89, 1405–1413. [Google Scholar] [CrossRef]
- Santoro, K.L.; Yakah, W.; Singh, P.; Ramiro-Cortijo, D.; Medina-Morales, E.; Freedman, S.D.; Martin, C.R. Acetaminophen and Xenobiotic Metabolites in Human Milk and the Development of Bronchopulmonary Dysplasia and Retinopathy of Prematurity in a Cohort of Extremely Preterm Infants. J. Pediatr. 2022, 244, 224–229.e3. [Google Scholar] [CrossRef]
Figure 1.
Results of Principal Component Analysis (PCA) of quantitative levels of 913 biochemicals detected in the urine of infants from the TOLSURF and PROP cohorts between 23-30 days of life. Infants who were on TPN at urine collection are shown in red, infants who were on enteral feeds at urine collection are shown in blue. (a) X-Y plot of the first two Principal Components (PC1 and PC2) for each infant, (b) Boxplot of PC1 and PC2 by infant feeding status showing significant differences between TPN vs. enteral feeds for both PC1 (p=1.51e-16) and PC2 (p=4.36e-9).
Figure 1.
Results of Principal Component Analysis (PCA) of quantitative levels of 913 biochemicals detected in the urine of infants from the TOLSURF and PROP cohorts between 23-30 days of life. Infants who were on TPN at urine collection are shown in red, infants who were on enteral feeds at urine collection are shown in blue. (a) X-Y plot of the first two Principal Components (PC1 and PC2) for each infant, (b) Boxplot of PC1 and PC2 by infant feeding status showing significant differences between TPN vs. enteral feeds for both PC1 (p=1.51e-16) and PC2 (p=4.36e-9).
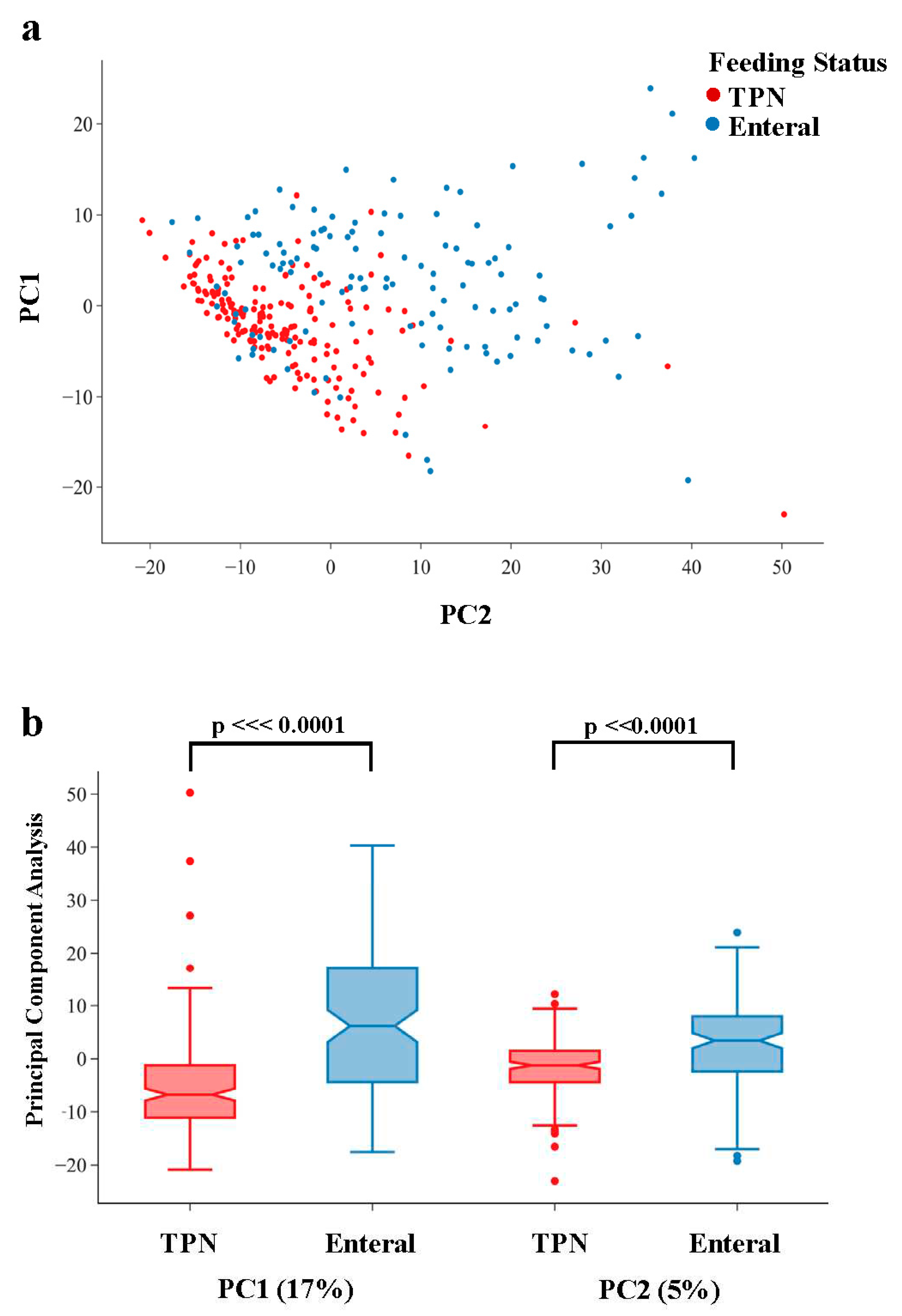
Figure 2.
Volcano plot from MWAS using logistic regression to compare urinary profiles of infants on enteral feeds vs. TPN. The dashed red line represents p=0.05; metabolites with OR >0 are higher in infants on TPN and metabolites with OR <0 are higher in infants on enteral feeds (dashed blue line). Metabolites are significantly enriched in infants who transitioned to enteral feeds for all super-pathways.
Figure 2.
Volcano plot from MWAS using logistic regression to compare urinary profiles of infants on enteral feeds vs. TPN. The dashed red line represents p=0.05; metabolites with OR >0 are higher in infants on TPN and metabolites with OR <0 are higher in infants on enteral feeds (dashed blue line). Metabolites are significantly enriched in infants who transitioned to enteral feeds for all super-pathways.
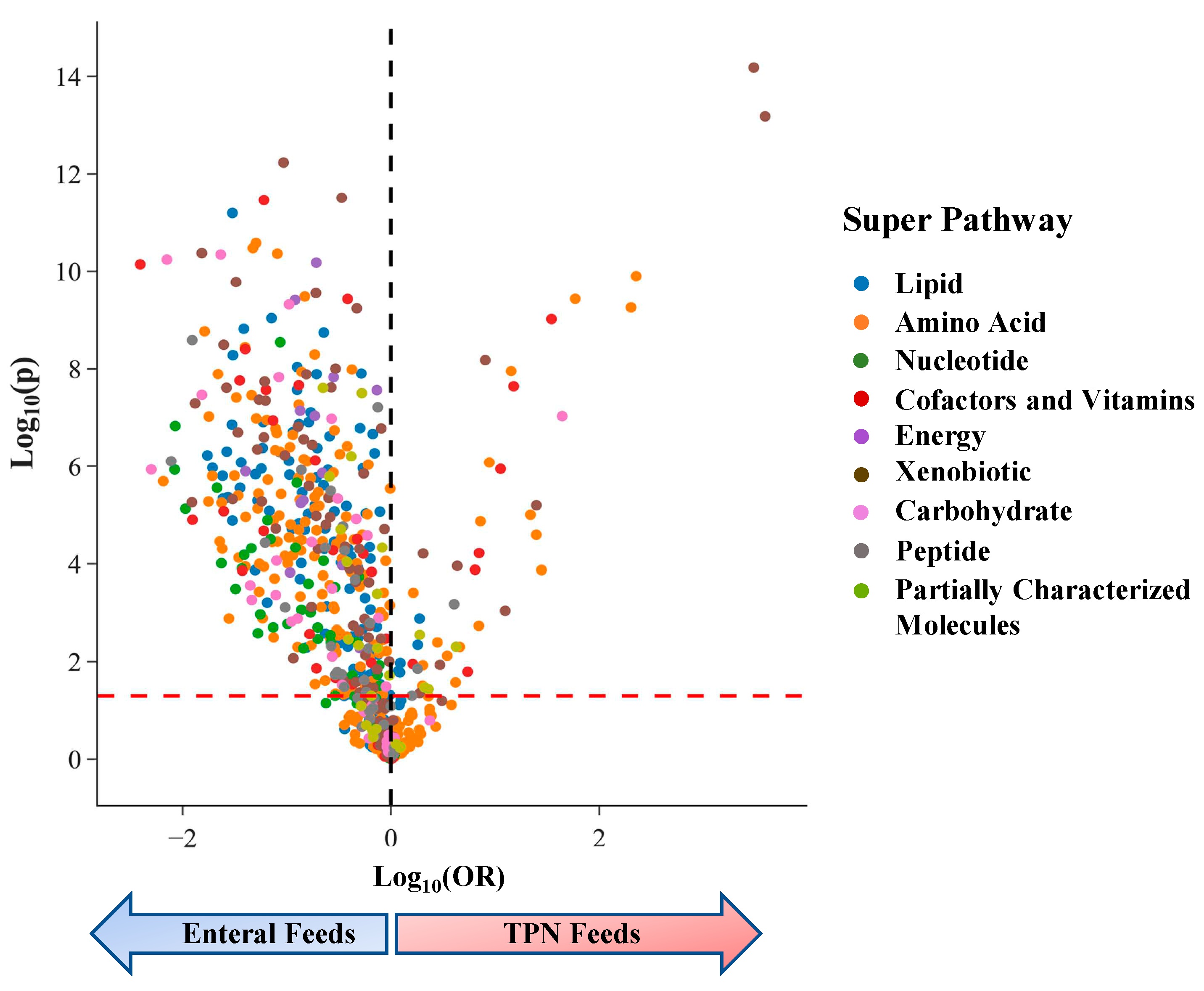
Figure 3.
Odds Ratio (OR) of amino acids and acyl-carnitines from metabolomic association analysis of TPN vs enteral feeding. (a) OR estimates for amino acids, with essential amino acids colored in red. (b) OR estimates for acylcarnitines. Metabolites significantly enriched in enteral feeds are colored in blue and those enriched in TPN feeds are colored in red.
Figure 3.
Odds Ratio (OR) of amino acids and acyl-carnitines from metabolomic association analysis of TPN vs enteral feeding. (a) OR estimates for amino acids, with essential amino acids colored in red. (b) OR estimates for acylcarnitines. Metabolites significantly enriched in enteral feeds are colored in blue and those enriched in TPN feeds are colored in red.
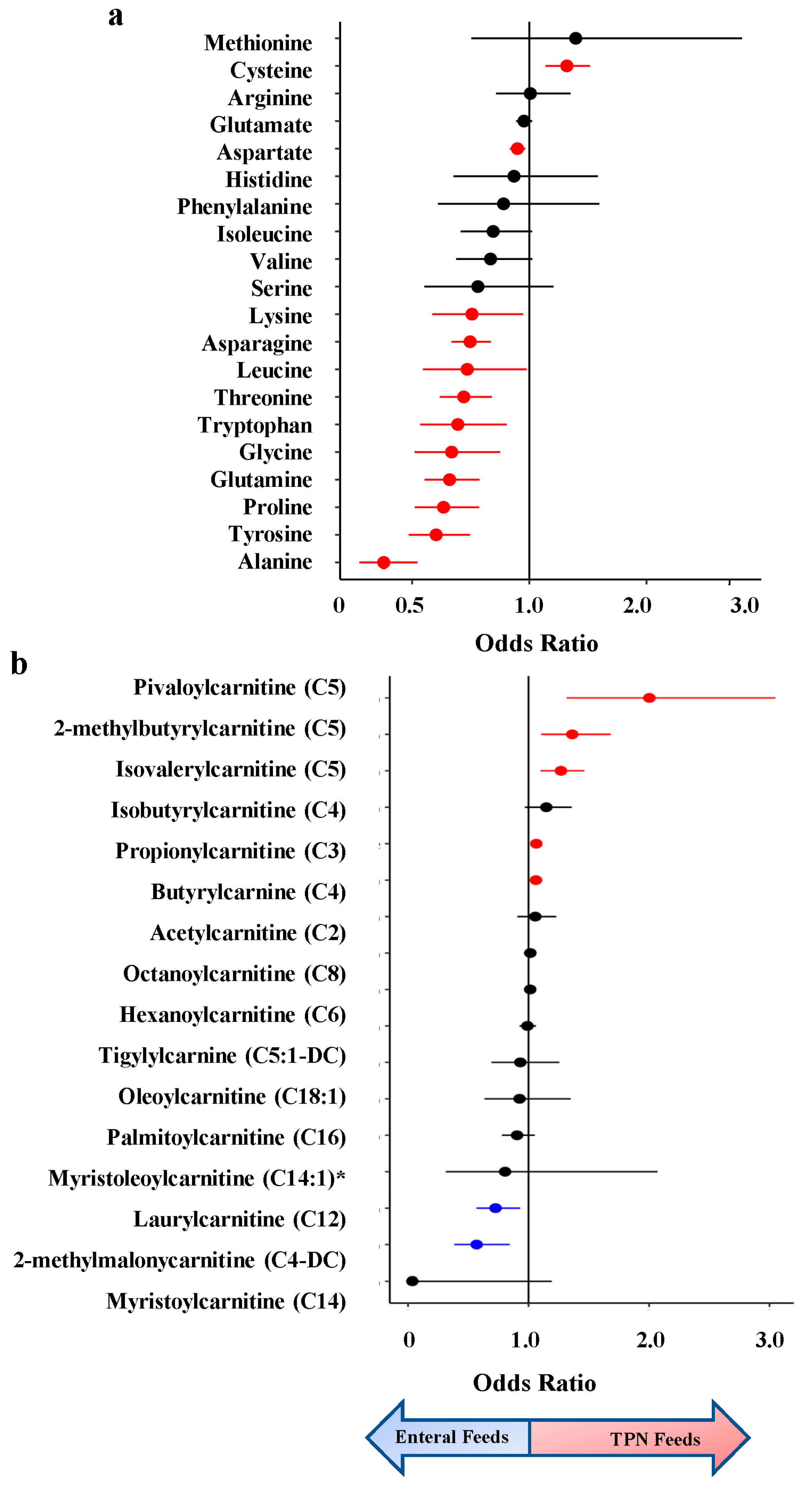
Figure 4.
Bar plot of pathway enrichment analysis of metabolomic differences between infants on TPN vs. enteral feeds. (a) super-pathway; (b) sub-pathway. Metabolite set enrichment analysis was performed using a Fisher Exact Test on sub-pathways with at least 2 named metabolites. 8 sub-pathways were found to be enriched at p<0.05 (dashed red line).
Figure 4.
Bar plot of pathway enrichment analysis of metabolomic differences between infants on TPN vs. enteral feeds. (a) super-pathway; (b) sub-pathway. Metabolite set enrichment analysis was performed using a Fisher Exact Test on sub-pathways with at least 2 named metabolites. 8 sub-pathways were found to be enriched at p<0.05 (dashed red line).
Figure 5.
Trajectories of individual metabolite concentrations from 9 super-pathways for one infant transitioning off TPN. Dexpanthenol (pro-vitamin B5) was used as a marker of TPN treatment. Dashed red line indicates when the infant was fully off TPN.
Figure 5.
Trajectories of individual metabolite concentrations from 9 super-pathways for one infant transitioning off TPN. Dexpanthenol (pro-vitamin B5) was used as a marker of TPN treatment. Dashed red line indicates when the infant was fully off TPN.
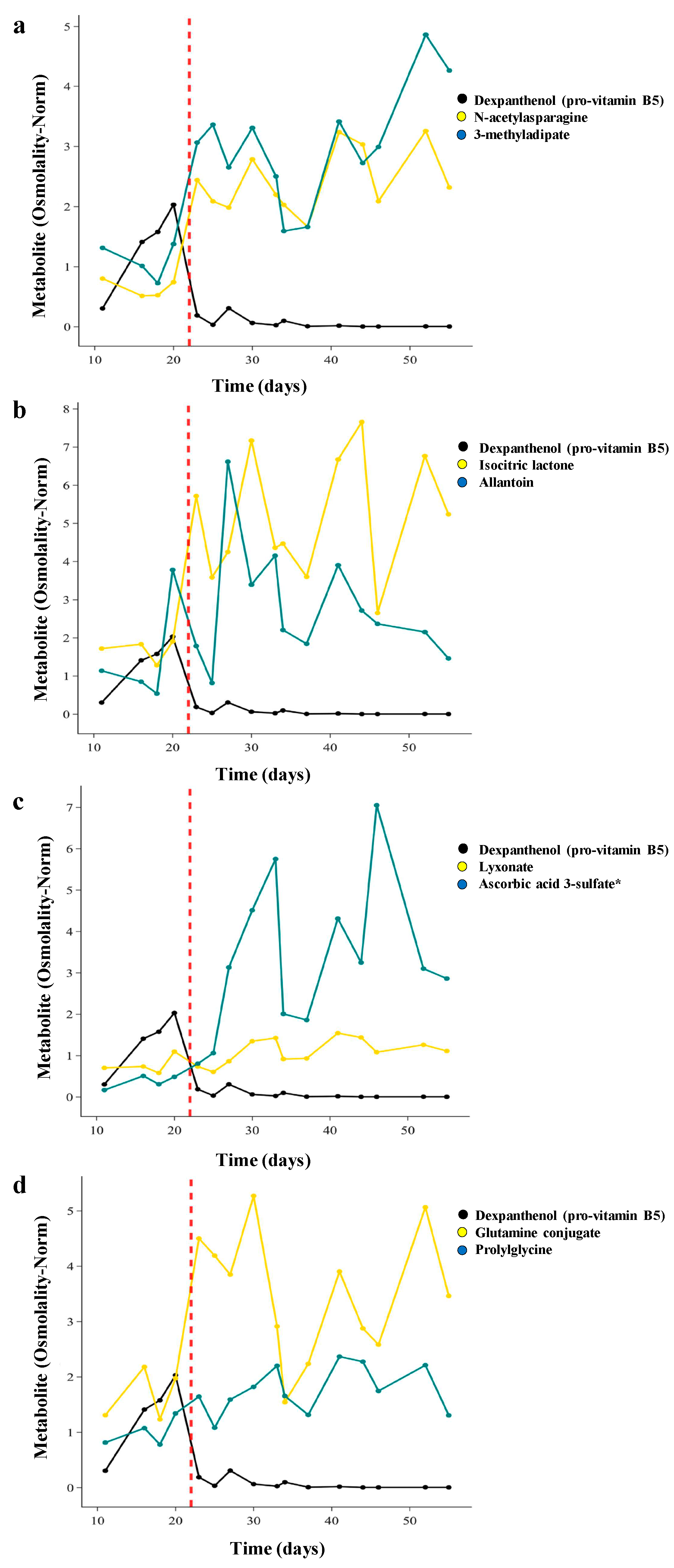

Table 1.
Characteristics of infants included in metabolomic studies from TOLSURF and PROP by feeding status at 28 days. Quantitative measurements were compared using a student’s t-test and categorical data using a Fisher’s Exact Test.
Table 1.
Characteristics of infants included in metabolomic studies from TOLSURF and PROP by feeding status at 28 days. Quantitative measurements were compared using a student’s t-test and categorical data using a Fisher’s Exact Test.
| TOLSURF | PROP | |||||
|---|---|---|---|---|---|---|
| TPN | Enteral | p-value | TPN | Enteral | p-value | |
| Number of Infants | 108 | 63 | - | 80 | 63 | - |
| Age at Sample in days, mean (SD) | 27.3 (2.2) | 28.1 (2.0) | 0.013 | 28.3 (0.9) | 28.0 (0.9) | 0.14 |
| Number of Infants by Maternal Race (NHW|AA|HL)* | 50|40|18 | 35|21|7 | 0.43 | 35|30|15 | 30|25|8 | 0.62 |
| Gestational Age in weeks, mean (SD) | 25.2 (1.2) | 25.6 (1.2) | 0.034 | 25.7 (1.0) | 26.0 (1.3) | 0.11 |
| Male sex (%) | 58.3 | 58.7 | 1.00 | 51.3 | 54.0 | 0.87 |
| Birthweight (g), mean (SD) | 698 (164) | 778 (181) | 0.008 | 770 (161) | 804 (131) | 0.18 |
| On corticosteroids at day 28 (%) | 22.2 | 14.3 | 0.23 | 28.7 | 15.9 | 0.0029 |
*NHW = Non-Hispanic White; AA = African American; HL = Hispanic Latino.
Table 2.
Top 20 most significant metabolites that varied by feeding status at 28 days. Metabolites with OR<1 are enriched in urine of infants who had transitioned to enteral feeds while those with OR>1 are enriched for infants on TPN feeds. AVG FC is the average fold change for Enteral vs TPN feeds from the TOLSURF and PROP cohort.
Table 2.
Top 20 most significant metabolites that varied by feeding status at 28 days. Metabolites with OR<1 are enriched in urine of infants who had transitioned to enteral feeds while those with OR>1 are enriched for infants on TPN feeds. AVG FC is the average fold change for Enteral vs TPN feeds from the TOLSURF and PROP cohort.
| Biochemical Name | Super-Pathway | Meta p | Meta OR | AVG FC |
|---|---|---|---|---|
| Dexpanthenol | Xenobiotics | 6.6e-15 | 11.19 | 6.30 |
| Gluconate | Xenobiotics | 6.7e-14 | 12.06 | 4.83 |
| Ferulic acid 4-sulfate | Xenobiotics | 5.9e-13 | 0.49 | 0.23 |
| 2,6-dihydroxybenzoic acid | Xenobiotics | 3.1e-12 | 0.72 | 0.18 |
| Ascorbic acid 3-sulfate | Cofactors and Vitamins | 3.5e-12 | 0.43 | 0.36 |
| 3-methyladipate | Lipid | 6.4e-12 | 0.35 | 0.50 |
| N-acetylasparagine | Amino Acid | 2.6e-11 | 0.41 | 0.43 |
| Methylsuccinate | Amino Acid | 3.3e-11 | 0.40 | 0.35 |
| Acetylhydroquinone sulfate | Xenobiotics | 4.2e-11 | 0.28 | 0.29 |
| N-succinyl-phenylalanine | Amino Acid | 4.3e-11 | 0.47 | 0.28 |
| Lyxonate | Carbohydrate | 4.5e-11 | 0.32 | 0.43 |
| Arabonate/xylonate | Carbohydrate | 5.7e-11 | 0.22 | 0.50 |
| Isocitric lactone | Energy | 6.6e-11 | 0.61 | 0.36 |
| Threonate | Cofactors and Vitamins | 7.2e-11 | 0.18 | 0.61 |
| N-acetyltyrosine | Amino Acid | 1.3e-10 | 5.11 | 7.08 |
| Tartronate (hydroxymalonate) | Xenobiotics | 1.7e-10 | 0.36 | 0.40 |
| 3-hydroxy-2-methylpyridine sulfate | Xenobiotics | 2.8e-10 | 0.61 | 0.25 |
| N-delta-acetylornithine | Amino Acid | 3.3e-10 | 0.56 | 0.26 |
| N-acetylphenylalanine | Amino Acid | 3.7e-10 | 3.41 | 4.02 |
| Pantothenate | Cofactors and Vitamins | 3.7e-10 | 0.75 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



